
SUBSÍDIOS PARA O MANEJO SUSTENTÁVEL NA FLORESTA … · Aos amigos das comunidades de estudo,...
138
Universidade Federal do Amapá - UNIFAP Pró-Reitoria de Pesquisa e Pós-Graduação Programa de Pós-Graduação em Biodiversidade Tropical - PPGBIO Mestrado e Doutorado UNIFAP / EMBRAPA-AP / IEPA / CI-BRASIL PERSEU DA SILVA APARÍCIO SUBSÍDIOS PARA O MANEJO SUSTENTÁVEL NA FLORESTA ESTADUAL DO AMAPÁ: ESTRUTURA E DINÂMICA MACAPÁ – AP 2013
Transcript of SUBSÍDIOS PARA O MANEJO SUSTENTÁVEL NA FLORESTA … · Aos amigos das comunidades de estudo,...

Universidade Federal do Amapá - UNIFAP
Pró-Reitoria de Pesquisa e Pós-Graduação
Programa de Pós-Graduação em Biodiversidade Tropical - PPGBIO
Mestrado e Doutorado
UNIFAP / EMBRAPA-AP / IEPA / CI-BRASIL
PERSEU DA SILVA APARÍCIO
SUBSÍDIOS PARA O MANEJO SUSTENTÁVEL NA FLORESTA
ESTADUAL DO AMAPÁ: ESTRUTURA E DINÂMICA
MACAPÁ – AP
2013

PERSEU DA SILVA APARÍCIO
SUBSÍDIOS PARA O MANEJO SUSTENTÁVEL NA FLORESTA
ESTADUAL DO AMAPÁ: ESTRUTURA E DINÂMICA
Tese apresentada ao Programa de Pós-
Graduação em Biodiversidade Tropical,
como parte dos requisitos para obtenção do
título de Doutor em Biodiversidade
Tropical.
Orientadora: Dra. Eleneide Doff Sotta
Co-orientador: Dr. Marcelino C. Guedes
MACAPÁ - AP
2013

Dados Internacionais de Catalogação na Publicação (CIP)
Biblioteca Central da Universidade Federal do Amapá
Aparício, Perseu da Silva.
Subsídios para o manejo sustentável na floresta estadual do Amapá:
estrutura e dinâmica / Perseu da Silva Aparício; orientadora Eleneide
Doff Sotta; co-orientador Marcelino C. Guedes. Macapá, 2013.
138 f.
Tese (Doutorado) – Fundação Universidade Federal do Amapá,
Programa de Pós-Graduação em Biodiversidade Tropical.
1. Floresta – Conservação. 2. Desenvolvimento sustentável –
Amapá. 3. Recursos naturais – Conservação. 4. Fitossociologia.
I. Sotta, Eleneide Doff. (orient.). II. Guedes, Marcelino C. (co-orient.).
III. Fundação Universidade Federal do Amapá. IV. Título.
CDD. (22.ed). 634.92098116

PERSEU DA SILVA APARÍCIO
SUBSÍDIOS PARA O MANEJO SUSTENTÁVEL NA FLORESTA
ESTADUAL DO AMAPÁ: ESTRUTURA E DINÂMICA
Banca Examinadora:
_________________________________________________________________
Dr. João da Luz Freitas (IEPA)
_________________________________________________________________
Dr. Luciano Araújo Pereira (UEAP)
__________________________________________________________________
Dr. Silas Mochiutti (EMBRAPA/AP)
__________________________________________________________________
Dr. Niro Higuchi (INPA)
Orientador:
__________________________________________________________________
Dra. Eleneide Doff Sotta (EMBRAPA)
Co-orientador:
__________________________________________________________________
Dr. Marcelino C. Guedes (EMBRAPA)
MACAPÁ-AP
Dezembro/ 2012

DEDICATÓRIA
Dedico este trabalho aos meus filhos e
bênçãos Taiguã e Tauã, fontes de inspiração e
motivação para continuar a trilhar os
desafios.

“O Senhor é a minha luz e a minha salvação. De quem terei medo?
O Senhor é a fortaleza da minha vida. A quem temerei? Ainda que
um exército se acampe contra mim não se atemorizará o meu
coração, e, se estourar contra mim a guerra, ainda assim terei
confiança.”
Salmo 27, v. 1,2 e 3

AGRADECIMENTOS
Agradeço a Deus por ter me ofertado saúde, força e capacidade para enfrentar todos os
obstáculos durante todo o curso de pós-graduação, principalmente na fase final da tese.
Ao Programa de Pós Graduação em Biodiversidade Tropical pela oportunidade de
aperfeiçoamento profissional.
A Universidade do Estado do Amapá (UEAP) e seus dirigentes pela compreensão e
incentivo concedido por meio de licença para coleta de campo e viagem para cumprimento
dos deveres do Doutorado.
A EMBRAPA-AP, pela confiança a mim depositada, pelo apoio financeiro e por todos
os momentos de aprendizado adquiridos no campo cujos ensinamentos são indeletáveis.
A Eleneide Doff Sotta, por todo exemplo demonstrado como pessoa e como
profissional, pela confiança, pelos momentos de descontração, pela simplicidade, paciência,
dedicação, respeito e amizade.
A Marcelino Carneiro Guedes, por aceitar co-orientar-me, por me ingressar no cenário
científico do Amapá, por estar sempre ao meu lado, pelos ensinamentos, apoio total, pela
orientação, amizade, respeito, dedicação e paciência em todos os trabalhos realizados.
A todos os motoristas e funcionários que fazem parte da Instituição EMBRAPA - AP,
que viajaram para o município de Porto Grande por diversas vezes, em especial a Carlão, por
todo companheirismo, paciência, auxílio no levantamento dos dados de campo durante o
trabalho.
A Wegliane Campelo da Silva Aparício, pela coragem, atitude, pelas discussões, pela
revisão de texto, pela luta, por se fazer sempre presente nas dificuldades e conquistas e pela
dedicação incomensurável, a qual jamais desapareceu de minhas lembranças, e, sou
eternamente grato e abençoado por ter participado de sua vida.
Aos amigos das comunidades de estudo, Filismino e família, Inês e família, Paulão
(Finado), Adonias, pela moradia, televisão, peixe, camarão e açaí cedidos, por conceder
autorização para a pesquisa e por toda colaboração, confiança, respeito e apoio.
Aos meus orientandos Diego Armando, Anderson Pedro, João Matos, Robson Borges,
Robson Carmo, Vanessa Santos, Camila Severiano, Rayanny, Harliany, Luandson, Elizabete,
Renam, Samara, Marcos Vinicios, Nicacio, Aislan, Francisco, Aloncio, Erick Santos, por
serem amigos desbravadores, por toda ajuda em campo, pelo companheirismo, união,
consideração, respeito, humildade, pelos memoráveis momentos de descontração e por todo
apoio durante este trabalho.

A minha família, pai, mãe, irmãos e cunhados, por serem meus verdadeiros amigos,
minha referência e alicerce, por estarem ao meu lado nas horas de alegria e de tristeza, pelo
incentivo à conclusão do doutorado.
Ao meu tio Emano Saita Aparicio por ter me ajudado no ingresso da pós-graduação, e
acreditado que poderia concluir meus objetivos.
Ao meu sogro e sogra, por toda confiança, dedicação, paciência, companheirismo,
consideração, incentivo e principalmente pela amizade, parceria e respeito.
Aos meus filhos Taiguã e Tauã por promover os melhores momentos da minha vida,
dedico.

RESUMO
Estudos florísticos e estruturais com espécies arbóreas adultas e em regeneração são
imprescindíveis para garantir a sustentabilidade do manejo de uma comunidade florestal.
Nesses termos, o objetivo deste trabalho foi inferir sobre a potencialidade do manejo florestal
em floresta ombrófila densa, a partir de atributos da estrutura e dinâmica das espécies
arbóreas adultas e em regeneração natural. A área de estudo está localizada na Floresta
Estadual do Amapá (FLOTA/AP), região central do estado, Amazônia, Brasil. As principais
fitofisionomias presentes são Floresta Ombrófila Densa de terras baixas, Floresta Ombrófila
Densa Submontana e Transição Cerrado – Floresta. Inicialmente, foram inventariadas as
árvores caídas presentes em 30 unidades amostrais em cada fitofisionomia da FLOTA, de
maneira aleatória. Todas as árvores caídas com diâmetro à altura do peito a 1,30 m do solo
(DAP) > 10 cm foram submetidas à cubagem rigorosa por meio dos procedimentos descritos
por Smalian para determinação do volume comercial. Para gerar uma equação de predição do
volume, cinco modelos de simples entrada (DAP); e oito de dupla entrada (DAP e altura
comercial do fuste (H), combinadas ou não) foram testados. O volume real das árvores, obtido
pela cubagem rigorosa, foi comparado com os volumes estimados pelas equações são de
simples entrada e de dupla entrada com melhor ajuste dos dados e com o volume cilíndrico
com fator de forma 0,7. O volume gerado por cada método foi comparado com o volume real
da testemunha por meio do teste t pareado. Posteriormente, foram implantadas parcelas
permanentes no segundo módulo da FLOTA/AP, por meio de amostragem em
conglomerados. O universo amostral abrangeu uma população de 500 ha, distribuídos em três
distintas áreas (unidades primárias - UP), equidistantes em, aproximadamente, 2.500 m. Cada
unidade primária foi disposta em forma quadrada com cinco parcelas permanentes de 100
x100 m para inventário de árvores adultas (DAP > 10cm), abrangendo uma área de 5 ha/UP e
15 ha no total. Cada parcela foi subdividida em 100 subparcelas de 10x10 m. Destas foram
sorteadas 20 subparcelas para o estudo da regeneração natural e inventário de arvoretas (5 <
DAP < 10 cm). Em cada subparcela sorteada também foi implantada, aleatoriamente nos
vértices, subunidade de 5x5m para inventário da regeneração natural em vara (2,5 < DAP < 5
cm). Também foram mensuradas as alturas comerciais, presença ou ausência de cipós
competidores, oco e cupim, posição sociológica, qualidade e classe do fuste. O
monitoramento da vegetação ocorreu entre os anos de 2010 e 2012, com remedições anuais.
Foram calculados os parâmetros fitossociológicos e regeneração natural por classe de altura.
Também foram gerados histogramas da estrutura diamétrica e estimado o volume total. A
similaridade florística entre cada um dos dois níveis de regeneração natural e das unidades
primárias, foi realizada por meio de análise de agrupamento. Todas as espécies identificadas
foram separadas em grupos de valor madeireiro. O crescimento das espécies foi avaliado a
partir dos incrementos periódicos anuais percentuais em diâmetro, taxas de recrutamento e
mortalidade. Quando geradas as equações volumétricas por fitofisionomia e geral (n = 216
árvores caídas), foi verificado que os modelos de simples entrada de Husch e dupla entrada de
Schumacher-Hall (Ln) geraram os melhores ajustes para a Floresta Ombrófila Densa, tanto
para as de terras baixas, quanto para as submontanas e de transição cerrado-floresta. Não
foram observadas diferenças significativas entre as estimativas de volume geradas pelas
equações de simples (Husch) e dupla entrada (Schumacher-Hall (Ln)) com o volume real (t =
0,81 e p = 0,42; t = 0,62 e p = 0,53, respectivamente) das árvores da Floresta Ombrófila
Densa do Amapá. Sempre que for possível medir com confiança a altura comercial das
árvores, recomenda-se o uso da equação de dupla entrada (LnV = -8,75 + 1,88*LnDAP +
0,82*LnHc) para estimativa dos volumes. Se apenas medidas do DAP estiverem disponíveis é
possível utilizar a equação (LnV = -7,92 + 2,21*LnDAP) de simples entrada. Foram
inventariados 7557 indivíduos arbóreos vivos com DAP > 10 cm, distribuídos em 38 famílias
botânicas, 115 gêneros, 183 espécies, com 12 indeterminadas. O índice de diversidade de
Shannon encontrado foi de 4,38 nats/ind. Dentre as espécies mais importantes com valor
comercial podem ser citadas: Tabebuia serratifolia, Apeiba tibourbou e Bowdichia nitida. O

número de árvores foi estimado em 504 indivíduos.ha-1
, com volume total de 383,22 m3,
distribuídos em 112,13 m3 nas árvores com DAP’s entre 10 e 40cm, 40,98 m
3 para arvores
com DAP’s entre 40 e 50cm e 201,56 m3 apenas para árvores com DAP’s maior 50cm. A
regeneração natural foi composta por 2.700 indivíduos, pertencentes a 38 famílias botânicas,
93 gêneros e 141 espécies arbóreas. O agrupamento formou três grupos florísticos. Quanto à
dinâmica florestal, ocorreu a perda de aproximadamente 3 % dos dados obtidos em campo,
gerando um total de 10.209 plantas vivas em 2010, com aumento de 0,46 % em 2011 e baixa
de 0,90 % em 2012, em relação ao inicio do monitoramento. A estimativa total por hectare de
varas, arvoretas e árvores foi de 2.303 indivíduos. O número de indivíduos e o crescimento
diamétrico das espécies foram influenciados pelo fator tempo. O inventário demonstrou que
nove espécies contêm taxas de recrutamento contínua nas seis classes diamétricas iniciais,
com variação de 0,02 a 0,87 %: Eschweilera coriacea, Osteophloeum platyspermum,
Pourouma ovata, Eugenia sp., Iryanthera paraensis, Micropholis acutangula, Micropholis
longipedicellata, Pouteria sp.1 e Sclerolobium melanocarpum. Os maiores índices de
mortalidade foram observados nas classes iniciais, com exceção da classe com diâmetros
acima de 110 cm (Mezilaurus itauba - 3,7 %). De maneira geral, as taxas de ingresso
apresentaram valores superiores em relação à mortalidade. Quanto aos incrementos periódicos
médios em DAP, durante os três anos de monitoramento foram encontrados valores para a
floresta variando de 0,6 a 0,8 cm ano-1
. O local de estudo apresentou um satisfatório número
de espécies adultas comercializáveis com relevante número de indivíduos, boas condições
fitossanitárias e volume apto à exploração, além de oferecer indivíduos jovens das espécies
alvo de corte para manter a capacidade produtiva da floresta após a exploração.
Palavras-chave: Fitossociologia; Crescimento; Amazônia.

ABSTRACT
Floristic and structural studies with tree and regeneration species are essential to sustainability
management of forest. So the aim was to know potential of forest management in tropical rain
forest, from attributes of structure and dynamics of tree and regeneration species. The study is
located in Amapá State Forest (FLOTA / AP), Amazon, Brazil. There are three types
vegetation: dense rain forest lowland, lower montane rain forest and savanna-forest transition.
Initially, fallen trees were inventoried present in 30 sampling in types vegetation. All fallen
trees with diameter at breast height to 1.30m (DBH) > 10 cm was mensured by Smalian
procedures to determine volume. To prediction equation of volume, five models with single
entry (DAP), and eight double entry (DBH and commercial height (H), combined or not) were
tested. The actual volume of trees was compared with volumes estimated by simple and
double equations, and with cylindrical volume (form factor 0.7). The volume generated by
each method was compared with the actual volume of the witness by the paired t test. Later,
plots were established three primary units (UP) in the second module FLOTA / AP to
estimated 500 ha. Each primary units was arranged with five plots (100 x100m) for inventory
trees (DBH > 10cm). Each plot was divided into 100 subplots of 10x10 m. These 20 plots
were selected for study regeneration (5 < DBH < 10 cm). In each subplot was also
implemented randomly subunit 5x5m (regeneration in 2.5 < DBH < 5 cm). Also the
commercial heights, presence or absence of competitors vines, hollow termite, sociological
position, quality and class of stem were measured. Vegetation monitoring occurred between
2010 and 2012. Parameters phytosociological were calculated and regeneration by height
class. Histograms diameter structure were generated and estimated total volume. The floristic
similarity between regeneration and primary units was performed by cluster analysis. All
species were separated into groups of timber value. The growth of species was evaluated from
annual percentage increases in diameter, recruitment and mortality rates. With volumetric
equations by vegetation type (n = 216 fallen trees), it was found that of single entry Husch
and double entry Schumacher-Hall (Ln) generated the best for Rain Forest, both for lowland,
submontane and transitional savanna-forest. No significant differences were observed
between volume estimates by Husch and Schumacher-Hall (Ln) with actual volume (t = 0.81
and p = 0.42, t = 0.62 p = 0.53, respectively). Where it is possible to measure with confidence
the commercial height of trees, it is recommended to use of double-entry equation (LNV = -
8.75 + 1.88 * + 0.82 * LnDAP LnHc) to estimate volumes. If only DBH are available it is
possible to use equation (LNV = -7.92 + 2.21 * LnDAP) with single entry. We recorded 7557
individual trees (DBH > 10 cm), distributed in 38 botanical families, 115 genera, 183 species,
with 12 indeterminate. The Shannon diversity index was 4.38 nats / ind. Among the most
important species value can be cited: Tabebuia serratifolia, Apeiba tibourbou and Bowdichia
nitida. The number of trees was estimated at 504 individuals.ha-1
, with a total volume of
383.22 m3, distributed in trees with DBH 10 and 40cm (112.13 m
3), 40.98 m
3 for trees with
DBH 40 and 50cm and 201.56 m3 for trees with DBH larger 50cm. Regeneration was
composed of 2,700 individuals, belonging to 38 botanical families, 93 genera and 141 species.
The cluster formed three floristic groups. As for forest dynamics, the loss occurred in
approximately 3% of data obtained, generating a total of 10,209 plants in 2010, an increase of
0.46% in 2011 and low of 0.90% in 2012, compared to beginning of monitoring. The total
estimate per hectare was 2.303 individuals (tree and regenaration). The number of individuals
and diameter growth of species were influenced by time factor. The survey showed that nine
species contain continuous recruitment rates in six initial diameter classes (0.02 to 0.87%):
Eschweilera coriacea, Osteophloeum platyspermum, Pourouma ovata, Eugenia sp.,
Iryanthera paraensis, Micropholis acutangula, Micropholis longipedicellata, Pouteria sp.1
and Sclerolobium melanocarpum. The highest mortality rates were observed in initial classes
except class with diameters above 110 cm (Mezilaurus itauba - 3.7%). In general, recruitment
rates showed higher values in relation to mortality. As for periodic increments, during the
three years of monitoring values were found ranging from 0.6 to 0.8 cm year-1
. The study site

presented a satisfactory number of marketable species with relevant number of individuals,
good phytosanitary conditions and volume fit to exploitation, as well as offering young
individuals species of target cutting to maintain productive capacity of forest after logging.
Key words: Phytosociology; Growth; Amazon

LISTA DE FIGURAS
Figura 1 Localização da Floresta Estadual do Amapá com seus limites e tipologias
florestais características, com destaque para o segundo módulo ao sul do
estado (Imagem do Satélite LANDSAT7 ETM+
26
Figura 2 Distribuição diamétrica e altura comercial média (Hc) das árvores caídas
na Floresta Estadual do Amapá, Amazônia, Brasil.
37
Figura 3 Distribuição diamétrica de árvores vivas e caídas a cada hectare na
Floresta Estadual do Amapá, Amazônia, Brasil.
38
Figura 4 Análise de distribuição gráfica de resíduos dos modelos de simples
entrada para equação de árvores caídas na Floresta Estadual do Amapá.
Onde: 1 - Koperzky-Gehrhardt; 2 - Hohenald-Krenn; 3 – Husch; 4 –
Brenac; 5 – Husch-Não linear.
45
Figura 5 Análise de distribuição gráfica de resíduos dos modelos de dupla entrada
para equação de árvores caídas na Floresta Estadual do Amapá. Onde: 1 –
Spurr; 2 – Stoate; 3 - Naslund; 4 – Meyer; 5 - Meyer modificada; 6 -
Spurr (Ln); 7 - Schumacher-Hall (Ln); 8 - Schumacher-Hall-não linear.
46
Figura 6 Curva do número de espécie por número de parcelas amostradas (área)
representando a composição florística da floresta ombrófila densa
inventariada no município de Porto Grande, Amapá, Amazônia.
58
Figura 7 Unidade Amostral Secundária (US) permanente, utilizada para o estudo
das espécies arbóreas adultas e em regeneração natural no segundo
módulo da FLOTA/AP
83
Figura 8
Figura 9
Dendrograma representando as sequências de agrupamento em três áreas
de estudo (A1; A2; A3) e dois níveis de regeneração natural (arvoreta -
5.0 < DAP < 10.0 cm; vara - 2.5 <DAP <5.0 cm) em floresta ombrófila
densa, Porto Grande/AP.
Distribuição do numero de indivíduos em distintas classes diamétricas
numa floresta ombrófila densa, Porto Grande, Amapá, Amazônia. Onde: 1
(2.5 < DAP < 5cm); 2 (5 < DAP < 10cm); 3 (10 < DAP < 20cm);4 (20 <
DAP < 30cm); 5 (30 < DAP < 40cm); 6 (40 < DAP < 50cm); 7 (50 <
DAP < 60cm); 8 (60 < DAP < 70cm); 9 (70 < DAP < 80cm); 10 (80 <
DAP < 90cm); 11 (90 < DAP < 100cm); 12 (100 < DAP < 110cm);13
(DAP > 110cm).
92
101

LISTA DE TABELAS
Tabela 1 Modelos volumétricos utilizados para representar a volumetria da Floresta
Estadual do Amapá, Amazônia.
35
Tabela 2 Suficiencia amostral para numero de indivíduos e volume por hectare na
Floresta Estadual do Amapá. FODTB – Floresta ombrofila densa de terras
baixas; FODSm – Floresta ombrofila densa submontana; TCF – Transição
cerrado-floresta. N.U.L – número unidades locadas; N.O.U – numero
ótimo de unidades; IC – intervalo de confiança; E.A. – erro de
amostragem.
36
Tabela 3 Ajuste das equações de volume para as árvores caídas testadas nos
modelos de simples entrada para Floresta Estadual do Amapá
(FLOTA/AP). Onde: β0, β1 – parâmetros estatísticos dos modelos; p –
nível de significância dos parâmetros dos modelos; F – valor calculado da
análise de variância; R²aj – coeficiente de determinação ajustado; Syx –
erro padrão residual absoluto; Syx% - erro padrão residual relativo; IF –
ídice de furnival absoluto; IF% – ídice de furnival relativo.
40
Tabela 4 Ajuste das equações de volume para as árvores caídas testadas nos
modelos de dupla entrada para Floresta Estadual do Amapá (FLOTA/AP).
Onde: β0, β1 – parâmetros estatísticos dos modelos; p – nível de
significância dos parâmetros dos modelos; F – valor calculado da análise
de variância; R²aj – coeficiente de determinação ajustado; Syx – erro
padrão residual absoluto; Syx% - erro padrão residual relativo; IF – ídice
de furnival absoluto; IF% – ídice de furnival relativo.
41
Tabela 5 Relação das espécies arbóreas inventariadas, distribuição espacial pelo
método Payandeh (Pi) e gupo de valor madeireiro (GVM) numa floresta
ombrófila densa, Porto Grande, Amapá, Amazônia.
59
Tabela 6
Tabela 7
Parâmetros Fitossociológicos de espécies arbóreas inventariadas numa
floresta ombrófila densa, Porto Grande, Amapá, Amazônia. Onde: N –
numero total de indivíduos; DR – densidade relativa; FR – freqüência
relativa; DoR – dominância relativa; VI – valor de importância; H – altura
comercial (metros); DAP – diâmetro a altura do peito a 1.30m do solo
(metros).
Estoque volumétrico e potencial das árvores nas unidades primárias (UP)
numa floresta ombrófila densa, Porto Grande, Amapá, Amazônia. Onde:
Qf – qualidade do fuste; N – número total de indivíduos; V – volume em
m³.ha-1
; PC – presença de cupim; C – presença de cipós competidores; O
– presença de oco.
67

72
Tabela 8 Estimativa do percentual da Regeneração Natural Total (RNT) para as
arvoretas* (5.0 < DAP < 10.0 cm) e varas** (2.5 < DAP <5.0 cm) por
classe de altura em Porto Grande/AP, listados em ordem decrescente de
acordo com o maior valor de RNT, onde DR= Densidade Relativa; FR =
Freqüência Relativa e RNC1 = Regeneração Natural na Classe 1 de altura;
RNC2 = Regeneração Natural na Classe 2 de altura e RNC3 =
Regeneração Natural na Classe 3 de altura.
85
Tabela 9
Tabela 10
Taxa de recrutamento (R) e mortalidade (M) por classe diamétrica das
espécies arbóreas inventariadas numa floresta ombrófila densa, Porto
Grande, Amapá, Amazônia. Em que: classe 1(2,5 < DAP < 5cm), classe 2
(5 < DAP < 10cm), classe 3 (10 < DAP < 20cm), classe 4 (20 < DAP <
30cm), classe 5 (30 < DAP < 40cm), classe 6 (40 < DAP < 50cm) classe 7
(50 < DAP < 60cm), classe 8 (60 < DAP < 70cm), classe 9 (70 < DAP <
80cm), classe 10(80 < DAP < 90cm), classe 11(90 < DAP < 100cm),
classe 12 (100 < DAP < 110cm) e classe 13 (≥110cm).
Taxa de recrutamento (t), mortalidade (M), incremento periódico anual
percentual (IPA) e incremento periódico anual médio (IPAm) nas classes
diamétricas numa floresta ombrófila densa, Porto Grande/AP.
103
117
Tabela 11 Teste de hipósete multivariado de Wilks para o efeito tempo (T) e
interações sobre o número de indivíduos (N) e crescimento em diâmetro
(C) entre as Unidades Primárias (UP), Nível de inclusão e Grupo de valor
madeireiro (GVM) em uma floresta ombrófila densa, Porto Grande/AP.
118
Tabela 12 Crescimento em diâmetro (cm), representado pelo desvio padrão, nas
Unidades Primárias (UP) quanto ao Grupo de valor madeireiro (GVM)
em uma floresta ombrófila densa, Porto Grande/AP.
119

SUMÁRIO
1 INTRODUÇÃO GERAL ........................................................................... 17
1.1 OBJETIVOS DE ESTUDO..........................................................................
1.2 ESTRUTURA DA TESE.............................................................................
1.3 REVISÃO DE LITERATURA.....................................................................
19
19
20
1.3.1 Fitossociologia............................................................................... 20
1.3.2 Regeneração Natural no Contexto do Manejo
Sustentável......................................................................................
21
1.3.3 Dinâmica Florestal......................................................................... 22
1.4 DESCRIÇÃO DO LOCAL DE ESTUDO................................................... 25
2 ESQUAÇÕES VOLUMÉTRICAS DE ÁRVORES CAIDAS EM
DIFERENTES FITOFISIONOMIAS DA FLORESTA
AMAZÔNICA, BRASIL............................................................................
27
RESUMO...................................................................................................... 28
ABSTRACT ................................................................................................. 29
2.1 INTRODUÇÃO..................................,......................................................... 30
2.2 MATERIAL E MÉTODOS ......................................................................... 32
2.2.1
2.2.2
Caracterização da Área ..................................................................
Estrutura Florestal...........................................................................
32
33
2.2.3 Equação Volumétrica...................................................................... 34
2.2.4 Validação da Equação Volumétrica................................................ 35
2.3 RESULTADOS............................................................................................. 36
2.3.1 Estrutura Florestal........................................................................... 36
2.3.2 Equação Volumétrica...................................................................... 38
2.3.3 Validação da Equação Volumétrica................................................ 46
2.4 DISCUSSÃO................................................................................................ 47
2.4.1 Estrutura Florestal........................................................................... 47
2.4.2 Equação Volumétrica...................................................................... 48
2.4.3 Validação da Equação Volumétrica................................................ 50
2.5
3
3.1
3.2
3.3
CONCLUSÕES............................................................................................
APTIDÃO DE ESPÉCIES ARBÓREAS O MANEJO EM
FLORESTA OMBRÓFILA DENSA, AMAPÁ, AMAZÔNIA...............
RESUMO......................................................................................................
ABSTRACT..................................................................................................
INTRODUÇÃO............................................................................................
MATERIAL E MÉTODOS..........................................................................
3.2.1 Caracterização da Área..................................................................
3.2.2 Inventário Florestal........................................................................
3.2.3 Análise Estrutural...........................................................................
RESULTADOS.............................................................................................
3.3.1 Florística e Fitossociologia............................................................
51
52
53
54
55
56
56
56
57
57
57

3.4
3.5
3.3.2 Estrutura Arbórea...........................................................................
3.3.3 Potencial Madeireiro......................................................................
DISCUSSÃO................................................................................................
3.4.1 Florística e Fitossociologia............................................................
3.4.2 Estrutura Arbórea...........................................................................
3.4.3 Potencial Madeireiro......................................................................
CONCLUSÕES............................................................................................
71
72
73
73
75
76
77
4 NÍVEIS DE REGENERAÇÃO NATURAL PARA SUBSÍDIO AO
MANEJO EM FLORESTA DE TERRA FIRME NO
AMAPÁ........................................................................................................
78
RESUMO ..................................................................................................... 79
ABSTRACT ................................................................................................. 80
4.1 INTRODUÇÃO ........................................................................................... 81
4.2 MATERIAL E MÉTODOS ......................................................................... 82
4.3 RESULTADOS............................................................................................. 83
4.4 DISCUSSÃO................................................................................................ 92
4.5 CONCLUSÕES............................................................................................ 94
5 DINÂMICA DO CRESCIMENTO DE ESPÉCIES ARBÓREAS EM
FLORESTA OMBRÓFILA DENSA NO AMAPÁ.................................
95
RESUMO...................................................................................................... 96
ABSTRACT.................................................................................................. 97
5.1 INTRODUÇÃO............................................................................................ 98
5.2 MATERIAL E MÉTODOS.......................................................................... 99
5.3 RESULTADOS............................................................................................. 101
5.4 DISCUSSÃO................................................................................................ 120
5.5 CONCLUSÕES............................................................................................ 123
6 CONSIDERAÇÕES FINAIS..................................................................... 124
REFERÊNCIAS.......................................................................................... -

17
1 INTRODUÇÃO GERAL
Os processos evolutivos têm modificado espontaneamente as florestas há milhares de
anos, resultando no equilíbrio dos sistemas estruturais e funcionais da natureza. No entanto,
nas últimas décadas têm sido intensas as mudanças provocadas pelo homem, ocasionando
impactos positivos e negativos à biodiversidade.
O principal responsável pelas expressivas transformações nas florestas é o
desmatamento irracional para expansão urbana e/ou desenvolvimento rural. A atividade
ocorre sem critérios de seleção, comprometendo a estrutura e diversidade florística das áreas
atingidas e adjacentes.
Historicamente, a região amazônica é alvo de interferências, com explorações desde o
descobrimento à atualidade. Segundo Lima Filho et al. (2004), a região amazônica vem
sofrendo interferência humana inadequada, exigindo da sociedade uma perspectiva de
aproveitamento socioeconômico mais elaborado e consistente, incluindo aspecto sobre o
conhecimento da sua biodiversidade. No entanto, é uma região bastante heterogênea no que
diz respeito à sua composição florística, evidenciando a complexidade dos grupos vegetais
que a compõem.
Os impactos decorridos do desmatamento mobilizaram o Governo Brasileiro a
estimular discussões sobre proteção da biodiversidade. Normas e legislações específicas
foram criadas a partir dos conhecimentos empíricos e científico para mitigar as ações de
desenvolvimento.
No ano de 1965, foi criado o código florestal brasileiro (lei 4.771/65) o qual definiu
que as florestas da Amazônia só poderiam ser utilizadas através de plano de manejo. Décadas
mais tarde o ministério do meio ambiente publicou a Instrução Normativa nº4/2006
(Autorização Prévia à Análise Técnica de Plano de Manejo Florestal Sustentável – APAT), e
posteriormente a Instrução Normativa nº 05/2006, que dispõe sobre procedimentos técnicos
para elaboração, apresentação, execução e avaliação técnica de Planos de Manejo Florestal
Sustentável - PMFS nas florestas primitivas e suas formas de sucessão na Amazônia Legal.
O manejo florestal também foi normatizado em áreas de assentamentos (IN
nº65/2010), exigindo o licenciamento ambiental para exploração das florestas com
mecanismos de controle dos cortes, regeneração e do estoque existente. No entanto, são
isentos do PMFS o corte raso através do uso alternativo do solo pela agropecuária (limitado a
3 ha/ano até atingir 20% da área total) e a exploração não comercial realizada pelas pequenas
propriedades e agricultores familiares.

18
Ainda existem inúmeros questionamentos que precisam ser esclarecidos, que variam
desde princípios ecológicos (definição de diâmetro e ciclos de cortes distintos por espécie) aos
econômicos (aumento das áreas destinadas à agropecuária). No ano de 2012, A lei nº 12.651,
que diz respeito ao Novo Código Florestal, estabeleceu o novo regime jurídico de proteção da
vegetação nativa.
No entanto, longe do que se esperava pelos ambientalistas, houve um retrocesso
ambiental em sua reformulação. Uma questão relevante para a conservação é o tamanho da
área, considerando que a diminuição desta resulta em alterações no fluxo de nutrientes,
número de indivíduos e espécies (Mantovani e Santos, 2007). Thomaziello (2007)
complementa que uma floresta aparentemente homogênea apresenta indivíduos de diferentes
características, que podem compor vários conjuntos vegetacionais fisionomicamente distinta.
Nesses termos, o presente trabalho utiliza a forma mais usual de caracterização da
estrutura da floresta, o levantamento fitossociológico. De acordo com Oliveira e Amaral
(2004) a obtenção e padronização dos atributos de diferentes ambientes florísticos e
fisionômicos possibilita a proposição de modelos mais adequados de manejar as florestas.
Estes estudos são essenciais para continuar a embasar a aplicação dos mecanismos
legais de monitoramento e conservação da floresta. Além disso, propicia a atualização de
normas da política ambiental para acompanhar a dinâmica das transformações das paisagens
do meio. Marangon et al. (2003), destacaram que a identidade das espécies e o seu
comportamento em comunidades vegetais é o começo de todo processo para compreensão de
um ecossistema.
Os levantamentos florísticos catalogam as espécies que ocorrem em uma área num
período de tempo, possibilitando acompanhar as mudanças sofridas e tendências ao longo dos
anos (SAWCZUK, 2009).
No Amapá, Aparício (2011) discorre que há poucas informações sobre as florestas,
apesar de apresentar um aspecto notável na sua flora nativa, com alto grau de diversidade,
distribuída em áreas relativamente próximas entre si. Assim, a obtenção dos parâmetros
estruturais (florística, Fitossociologia, regeneração e dinâmica) que permitam a utilização dos
recursos florestais é prerrogativa na execução e aprovação dos projetos ambientais que
orientam as atividades econômicas no estado.

19
1.1 OBJETIVOS DO ESTUDO
O objetivo geral deste trabalho foi inferir sobre a potencialidade do manejo florestal
em floresta ombrófila densa, a partir de atributos da estrutura e dinâmica das espécies
arbóreas adultas e em regeneração natural.
Os objetivos específicos deste trabalho foram:
a) Gerar uma equação volumétrica capaz de estimar o potencial madeireiro da Floresta
Estadual do Amapá (FLOTA/AP);
b) Avaliar os parâmetros fitossociológicos e regeneração natural de espécies arbóreas de
uma floresta ombrofila densa na FLOTA/AP;
c) Conhecer a capacidade produtiva madeireira das espécies arbóreas adultas alvo de
comercialização;
d) Quantificar o crescimento da floresta através da dinâmica de espécies arbóreas adultas
e em regeneração natural.
1.2 ESTRUTURA DA TESE
A fim de se estabelecer critérios técnicos para garantir a sustentabilidade num trecho
da Floresta Estadual do Amapá, foram descritos neste trabalho os principais parâmetros da
vegetação que embasam a legislação florestal vigente para construção do Plano de Manejo
Florestal Sustentável (PMFS).
Para atingir o objetivo, o trabalho foi dividido em quatro linhas de pesquisa, na área de
concentração de uso sustentável, compondo os seguintes capítulos:
a. Equações volumétricas de árvores caídas em diferentes fitofisionomias da floresta
Amazônica, Brasil;
b. Aptidão de espécies arbóreas ao Manejo em Floresta Ombrófila Densa, Amapá, Amazônia;
c. Níveis de regeneração natural para subsidio ao Manejo em Floresta de Terra Firme no
Amapá;
d. Dinâmica do crescimento de espécies arbóreas em Floresta Ombrófila Densa no Amapá.

20
1.3 REVISÃO DE LITERATURA
1.3.1 Fitossociologia
Os distintos ambientes da Amazônia apresentam um contingente florístico rico e
variado, muitas vezes exclusivo de determinado ambiente, o que a torna o maior reservatório
natural da diversidade vegetal do planeta. As múltiplas inter-relações entre seus componentes
bióticos e abióticos formam um conjunto de ecossistemas altamente complexos e de equilíbrio
ecológico extremamente frágil (OLIVEIRA e AMARAL, 2004).
A fragilidade da Amazônia está associada, principalmente, as ações antrópicas de
exploração dos recursos naturais, tendo como conseqüência o desaparecimento das espécies
utilizadas. Durante décadas permaneceu a idéia de que os recursos florestais eram infindáveis,
no entanto a retirada de matéria-prima, em certos momentos, é superior a capacidade da
floresta se recompor (LIMA, 2009), indicando a necessidade prioritária de se conhecer a
composição e distribuição das espécies vegetais como subsidio ao manejo (OLIVEIRA et al.,
2008).
Para gerar indicadores de sustentabilidade sobre os ambientes antrópicamente
descaracterizados, qualificar a estrutura é o primeiro passo para conhecer o ciclo evolutivo da
floresta, apresentando fortes correlações com riqueza florística e a distribuição do número de
indivíduos das diferentes espécies (SILVA et al., 2011), como forma de estudar as inter-
relações dentro de determinada comunidade (SILVA et al., 2008; OLIVEIRA et al., 2009).
Compreender como as espécies florestais vivem em comunidade e se distribuem
espacialmente são essenciais para a elaboração de planos de manejo de uso múltiplo. Esta
prerrogativa permite inferências de maior acurácia sobre o estoque produtivo da floresta,
identificando alguns fatores responsáveis pelas mudanças no padrão da diversidade local,
estimar o grau de distúrbio da área, além de propor medidas conservacionistas (BENTES-
GAMA, 2008; GUIMARÃES e CARIM, 2008; SCHNEIDER, 2008).
Trabalhos realizados em vários estados da Amazônia Legal classificam algumas
espécies potenciais para comercialização a partir do Valor de Importância (VI)
fitossociológico, com destaque para as espécies Eschweilera coriacea, Pouteria sp.,
Sclerolobium sp., Bellucia grossularioides, Eschweilera longipes. (SILVA e BENTES-
GAMA, 2008; GONÇALVES e SANTOS, 2008; SALOMÃO et al., 2012; POSSIMOSER et
al., 2012; PEREIRA et al., 2011).
Destaca-se que a riqueza florística da Amazônia está associada a um elevado número
de indivíduos a cada hectare, variando ente as distintas fitofisionomias. Em alguns estudos

21
fitossociológicos os valores do índice de diversidade de Shannon variam de 2,38 a 4,39
nats.individuo-1
(OLIVEIRA e AMARAL, 2005; FERREIRA JÚNIOR et al., 2008; KUNZ et
al., 2008; ALMEIDA et al., 2012).
Nesse contexto, para a aplicação de projetos com o aproveitamento permanente da
floresta, deve-se compreender o arranjo espacial dos elementos da comunidade, e ainda
conhecer a função das diferentes espécies, como também os habitats preferenciais (QUEIROZ
e MACHADO, 2008). Tais informações caracterizam a região quanto ao potencial econômico
(SANTOS et al., 2012; OLIVEIRA et al., 2008).
Desta forma, para a aplicação do manejo em regime sustentado em florestas na
Amazônia, devem-se analisar as características qualitativas e quantitativas, de modo a
possibilitar que a floresta atinja seu máximo potencial produtivo (FRANCEZ et al., 2009).
1.3.2 Regeneração Natural no contexto do Manejo Sustentável
No que tange o manejo florestal, a regeneração é a base para a sustentabilidade e
garantia de renovação de recursos extraídos da floresta. Isso fica estabelecido na legislação
florestal brasileira através da Lei 11.284 de 2/3/2006, Instrução Normativa 05 MMA de
11/12/2006 e o Decreto 5975 de 30/11/2006 por meio de fundamentos técnicos e científicos
em que trata o artigo 3° do referente Decreto.
Devido à crescente pressão do mercado consumidor de madeira e da opinião pública
quanto à sustentabilidade dos recursos florestais, há um número cada vez maior de iniciativas
de manejo florestal de baixo impacto que visa reduzir os danos causados à natureza pela
interferência humana (FRANCEZ et. al., 2007).
Estudos sobre a dinâmica de regeneração natural são essenciais para a elaboração e
aplicação correta dos planos de manejo e tratamentos silviculturais, permitindo a exploração
racional e permanente dos remanescentes (SILVA et al, 2012).
O termo regeneração natural tem um conceito de avaliação muito amplo, de maneira
que existem diversas metodologias utilizadas pela comunidade científica para sua
caracterização (KERKHOFF, 2011). Para Chami (2008), a regeneração natural é classificada
por meio de classes de tamanho, sendo a padronização dos limites determinada pelo objetivo
do estudo e das características da floresta estudada. Deste modo, ao estudar a vegetação por
meio de classes de tamanho, cada classe é considerada a regeneração natural da classe
subsequente e os tamanhos de mínimo e máximo das plantas consideradas são variáveis
(GARCIA, 2009).

22
Segundo Schorn e Galvão (2006), a regeneração natural é representada pelo estrato
arbóreo inferior da floresta, que geralmente apresentam-se com um comportamento
diferenciado em muitos aspectos da estrutura e dinâmica, tais como a densidade, as taxas de
ingresso e mortalidade, além do crescimento. Também há o uso do termo estrato regenerativo,
referindo-se ao conjunto de indivíduos que sobrevivem ao período crítico da ação seletiva do
ambiente e que representam o verdadeiro potencial regenerativo da estrutura arbórea
(GARCIA, 2009).
A regeneração natural decorre da interação de processos naturais de restabelecimento
do ecossistema e, é, portanto, parte do ciclo de crescimento da floresta. Também se refere às
fases iniciais do estabelecimento e desenvolvimento das plantas. Esse tipo de processo é parte
do complexo biológico ativo das florestas tropicais que formam, desenvolvem e mantém as
fitofisionomias (MARANGON, 1999).
De acordo com Zanetti e Siqueira (2011) a capacidade de regeneração florestal
depende dos elos do processo regenerativo (herbívoros vertebrados, chuva de sementes,
fatores físicos) e/ou vários atributos diretos da regeneração (densidade, riqueza, crescimento,
recrutamento, mortalidade de plantas). Além disso, a regeneração vai ainda depender das
condições alelopáticas encontradas em cada local diferente.
O estudo da regeneração natural permite a realização de previsões sobre o
comportamento e desenvolvimento futuro da floresta, na medida em que fornece a relação e a
quantidade de espécies que constitui o seu estoque, bem como suas dimensões e distribuição
na área (MARAGON et al., 2008). Além de permitir a identificação das espécies que deverão
ter maior importância no futuro, bem como aquelas que tendem a diminuir a participação na
estrutura (SCHORN e GALVÃO, 2006).
O conhecimento da composição da regeneração contribui para definir o estágio, bem
como as direções sucessionais de uma vegetação em desenvolvimento. Sendo a quantificação
da regeneração natural, quando associada à classificação sucessional das espécies, um
indicador extremamente útil das condições de recuperação e de sustentabilidade da floresta
(MARTINS, 2010).
A abordagem do tema de regeneração natural permite avaliar o funcionamento e ações
de manejo, devendo ter cautela no planejamento das operações de exploração para minimizar
a influência direta dos danos causados às plantas emergentes (RIBEIRO e FELFILI, 2009;
FRANCEZ, 2009).
1.3.3 Dinâmica Florestal

23
Na Amazônia, o estudo da dinâmica florestal demonstra um desafio para os
pesquisadores, devido à grande extensão com variabilidade nos ambientes e diversidade
florística. Entender a dinâmica de ocorrência de perturbações periódicas, naturais ou não,
também auxilia na definição dos diferentes estágios de desenvolvimento e o crescimento da
floresta (PANTALEÃO, 2008).
De acordo com Botezelli et al. (2005), estudos de dinâmica têm por objetivo
compreender o processo evolutivo das espécies, na escala ecológica e temporal. Além de
também determinar os fatores abióticos e bióticos que influenciam as alterações nas taxas de
natalidade, mortalidade e recrutamento das espécies, fundamentais em atividades de
sustentabilidade.
Além disso, a dinâmica florestal compreende o balanço entre a mortalidade e o
recrutamento de indivíduos. A taxa de crescimento assume um valor médio dinâmico, em que
a floresta oscila em produção, tanto em número de árvores, espécies e biomassa (HIGUCHI et
al., 2008). O crescimento da floresta é avaliado através do estudo das comunidades e/ou
populações arbóreas no tempo. Assim, a vegetação é mensurada periodicamente, tanto para
fins de caracterização biológica e ecológica, para prever o processo de substituição e propor
estratégias e métodos para manejo da floresta em regime sustentável (HOSOKAWA et al.,
1998; CARVALHO et al.,2010).
Quando se utiliza os termos “crescimento, incremento e mortalidade florestal” estão se
relacionando aos processos temporais da estrutura da floresta. Segundo Moscovich (2006), o
conhecimento das interações desses processos, facilita a interpretação sobre como se
desenvolveu a vegetação através do tempo, transformando-se numa ferramenta de
fundamental importância na tomada de decisões concernentes ao manejo silvicultural.
A determinação dos parâmetros que caracterizam a floresta no tempo deve ser
considerada nos planos de manejo florestal, pois se trata de uma informação básica na
estimativa da produção e ordenamento sustentável. Segundo Berger e Padilha (2007), a
dinâmica determina a viabilidade técnica e econômica de empreendimentos, pois influência
na escolha do regime de manejo, o período de rotação florestal ou o ciclo de corte em
florestas multiâneas.
De acordo com Schneider (2009) o termo crescimento se refere ao aumento das
dimensões de um ou mais indivíduos em um povoamento florestal ao longo de um
determinado período de tempo. Tal fato é verificado pelo avanço de suas dimensões físicas,
ocasionando uma reação à ação das leis naturais, condicionadas ao clima, solo, espécie,
composição florística e idade.

24
O incremento diamétrico, a mortalidade e o recrutamento estão entre os poucos
parâmetros para se fazer predições sobre a produção futura de uma floresta. A estimativa do
crescimento é essencial no ordenamento e credibilidade de um plano de manejo sustentável.
No planejamento florestal, as decisões de manejo são tomadas com base na predição do
crescimento e na produção que os povoamentos podem alcançar de acordo com suas
respectivas taxas (FERREIRA et al., 1998; CARVALHO, 1999).
Quanto à mortalidade, Campos e Leite (2009) definem como o número de indivíduos
que morrem em um dado período no tempo, podendo ser expressa como uma taxa explicita
dada pela razão entre o número de indivíduos que morreram e o total existente anteriormente
ao evento causador da morte.
A mortalidade pode ser um evento aleatório e dinâmico em florestas naturais, e só
pode ser mensurada por meio de parcelas permanentes representativas da floresta (ROSSI et
al., 2007).
Segundo Marra (2010) vários fatores podem causar a mortalidade em povoamentos
florestais, tais como a idade ou senilidade, competição e supressão, doenças e pragas,
alterações climáticas, fogos silvestres, tempestades convectivas, anelamento e
envenenamento, injúrias, corte ou abate de árvore, etc.
Outro fator importante a ser considerado na dinâmica da floresta é o recrutamento ou
ingresso, obtido pelo uso das parcelas permanentes convencionais (LIMA, 2010). Sua
dinâmica esta relacionada à densidade do povoamento, histórico da exploração, tipo de
floresta, presença de árvores porta-semente, dinâmica de clareiras, e outros fatores
empiricamente determináveis na parcela (PINTO, 2008).
O mesmo autor define que o recrutamento, ou ingresso, é o número de árvores que
atinge o diâmetro mínimo considerado no monitoramento da floresta. Conforme Barros
(2009), o recrutamento permite acompanhar e inferir o tempo de crescimento das espécies
florestais em um determinado período.
No sistema de manejo de corte seletivo, a avaliação da regeneração de espécies de
valor comercial, bem como o seu ingresso em classes diamétricas comerciais, são parâmetros
para indício da viabilidade técnica e econômica do regime de manejo empregado
(HOSOKAWA et al., 2008).
Neste sentido, no amplo horizonte do manejo florestal, o estudo da dinâmica
vislumbra-se com um conjunto de interações muito complexas pela multiplicidade dos fatores
nela incluídos. Sendo que estes fatores influem diretamente sobre o crescimento e
desenvolvimento de todas as árvores que formam o povoamento (MOSCOVICH, 2006).

25
1.4 DESCRIÇÃO DO LOCAL DE ESTUDO
O trabalho está localizado na unidade de conservação de uso sustentável denominada
Floresta Estadual do Amapá (FLOTA/AP), região central do estado do Amapá, Amazônia,
Brasil. A unidade compreende uma área de 2.369.400 ha dividida em 4 módulos produtivos,
definidos em função de infraestrutura e vias de acessos para dinamizar os pólos de
desenvolvimento do Estado.
No entanto, o alvo deste estudo são as florestas localizadas no segundo módulo da
FLOTA/AP (Figura 1) no município de Porto Grande/AP, a qual apresenta vegetação
ombrófila densa de terras baixas e submontana (VELOSO et al., 1991; IBGE, 2012).
A vegetação apresenta dossel de alto porte, até 50 m, perenifólia. Estima-se que o
número de espécies arbóreas que compõem esta fitofisionomia gira em torno de 245, com
destaque para Eschweilera coriacea (DC.) S.A. Mori, Pouteria caimito (Ruiz & Pav.) Radlk.,
Protium tenuifolium (Engl.) e Minquartia guianensis Aubl. Do total de espécies, 190 possuem
valor comercial, gerando uma densidade de 27 ind.ha-1
, 133 m3.ha
-1 e 48 % do total de volume
de madeira (DAP > 10cm) da fitofisionomia (IEF, 2009).
Os afloramentos rochosos são encontrados com baixa intensidade. A declividade varia
de ondulada a fortemente ondulada. Possui formação geológica pré-cambriana, com
superfícies pedi planado por canais geralmente curtos, numerosos e pouco aprofundado. O
solo predominante é tipo Latossolo Vermelho-Amarelo distrófico (RADAM, 1974). A
altitude varia de 60 a 100 m.
O clima da região é do tipo Am (equatorial super-úmido), segundo a classificação de
Köppen. A temperatura média é 25 ºC, ocorrendo pouca variação de temperatura durante o
ano. A precipitação anual na região é de 2.300 mm, com índice pluviométrico trimestral mais
seco abaixo de 200 mm. O período mais chuvoso ocorre no trimestre de março a maio (acima
de 1.000 mm). A umidade relativa é alta, com valores entre 80 e 86% (TARDIN et al., 2009).

26
Figura 1. Localização da Floresta Estadual do Amapá com seus limites e tipologias florestais características, com
destaque para o segundo módulo ao sul do Estado (Imagem do Satélite LANDSAT7 ETM).

27
2 EQUAÇÕES VOLUMÉTRICAS DE ÁRVORES CAÍDAS EM DIFERENTES
FITOFISIONOMIAS DA FLORESTA AMAZÔNICA, BRASIL

28
RESUMO
O presente trabalho visa gerar uma equação volumétrica por meio de dados biométricos de
árvores caídas, capazes de estimar o estoque real de madeira em distintas fitofisionomias da
Floresta Estadual do Amapá, Amazônia, Brasil. As principais fitofisionomias presentes são
Floresta Ombrófila Densa de terras baixas, Floresta Ombrófila Densa Submontana e
Transição Cerrado – Floresta. O inventário contemplou as árvores caídas presentes em 30
unidades amostrais locadas aleatoriamente em três distintas fitofisionomias. O número de
unidades amostrais foi proporcional às áreas das fitofisionomias. As unidades amostrais foram
estabelecidas seguindo o sentido Norte-Sul / Leste-Oeste, a partir de faixas contínuas de 20 x
2500 m em forma de cruz. Todas as árvores caídas com diâmetro à altura do peito a 1,30 m do
solo (DAP) > 10 cm foram submetidas à cubagem rigorosa, por meio dos procedimentos
descritos por Smalian para determinação do volume comercial. Para construção das equações,
os volumes outliers máximos e mínimos foram excluídos, bem como árvores com
discrepância nas medidas longitudinais do fuste. Dos modelos volumétricos utilizados, cinco
foram de simples entrada, tendo o DAP como variável independente; e oito de dupla entrada,
tendo o DAP e altura comercial do fuste (H), combinadas ou não, como variáveis
independentes. A melhor equação foi determinada por meio de comparações dos coeficientes
de determinação ajustado (R2
aj), erro padrão da estimativa (SYX) e análise da variância
(ANOVA). Também foi considerada a análise dos resíduos a partir do Desvio Médio
Percentual (DMP), com intuito de verificar a magnitude de subestimativa ou superestimativa
dos volumes estimados pelos modelos. O volume real das árvores, obtido pela cubagem
rigorosa, foi comparado com o volume estimado pela equação de simples e de dupla entrada
com melhor ajuste dos dados e com o volume cilíndrico com fator de forma 0,7, por meio do
teste t pareado. Foram inventariadas 1449 árvores caídas, sendo 71% na Floresta Ombrófila
Densa de terras baixas (FODBP), 15% na Floresta Ombrófila Densa Submontana (FODSm) e
14% na Transição Cerrado-Floresta (TCF) . Baseado na distribuição dos diâmetros das
árvores caídas foram utilizados os volumes reais de 300 indivíduos para a determinação das
equações. Os indivíduos que compõem a amostra apresentaram diâmetros à altura do peito a
1,30 m do solo variando de 10,1 a 112,4 cm. As alturas comerciais mínimas contempladas
foram 4 m, com valores máximos de 30 m. Quando geradas as equações por fitofisionomia,
foi verificado que os modelos de simples entrada de Husch e dupla entrada de Schumacher-
Hall (Ln) geraram os melhores ajustes nas três fitofisionomias, demonstrando independência
desse fator. Para validação das equações volumétricas das árvores caídas foram utilizados os
volumes reais de outras 216 árvores caídas que não foram utilizadas na construção das
equações. Os indivíduos que compõem a amostra apresentaram diâmetros à altura do peito a
1,30 m do solo variando de 10,3 a 123,4 cm. As alturas comerciais mínimas contempladas
foram de 3 m, com valores máximos de 28,9 m. O volume real dos indivíduos contempla de
0,05 a 15,62 m³, com média de 1,7533 m³ e erro padrão de 0,1757 m³. A expressão do volume
cilíndrico com fator de forma, tradicional em inventários e planos de manejos florestais
brasileiros, apresentou 1,4953 m³ (média) e 0,1537 m³ (erro padrão). Os modelos de Husch e
Schumacher-Hall (Ln) foram selecionados para validação de uso e aplicação das equações
volumétricas nas fitofisionomias estudadas.Quando comparado os volumes, não foram
observadas diferenças significativas entre as estimativas geradas pelas equações de simples
(Husch) e dupla entrada (Schumacher-Hall (Ln)) com o volume real (t = 0,81 e p = 0,42; t =
0,62 e p = 0,53, respectivamente). Resultado também encontrado para o volume cilíndrico
com fator de forma (t = 10,52 e p = 0,30). As estruturas diamétrica e hipsométrica das árvores
caídas da Floresta Estadual do Amapá representam a estrutura das árvores vivas em “pé”, nas
distintas fitofisionomias que a compõem. Assim, as equações volumétricas desenvolvidas
neste trabalho estão habilitadas para emprego da estimativa do volume comercial das áreas
destinadas ao manejo florestal sustentável.
Palavras-chave: Estimativa de volume; Cubagem rigorosa; Estrutura.

29
ABSTRACT
The present work aims to generate a volumetric equation by biometric data of fallen trees,
able to estimate real stock of wood in different vegetation types of Amapa State Forest,
Amazon, Brazil. The main vegetation types present are dense rain forest lowland, lower
montane rain forest and savanna-forest transition. The inventory included fallen trees present
in 30 sampling. The number of sample units was proportional to the area of each vegetation
types. The sampling units were established following the north-south / east-west (20 x 2500
m) in cross shape. All fallen trees with diameter at breast height to 1.30m (DBH) > 10 cm
were mensure by procedures Smalian to determine volume. For construction of equations,
maximum and minimum volumes outliers were excluded, as trees discrepancy in
measurements of longitudinal shaft. By volumetric models used, five were simple input and
eight double entry, with DBH and commercial height (H), combined or not. The best
equation was determined by comparing adjusted coefficients of determination (R2aj), standard
error of estimate (SYX) and analysis of variance (ANOVA). It was also considered analysis of
residues from Middle Deviation Percentage (DMP), in order to verify the magnitude of
underestimation or overestimation of volumes estimated by models. The actual volume of
trees was compared with volumes estimated by simple and double equations, and with
cylindrical volume (form factor 0.7). Were surveyed 1449 fallen trees, distributed in Rain
Forest of lowland (71%), Rain Forest submontane (15%) and savanna-forest transition (14%).
Based on distribution of diameters of fallen trees were used real volumes 300 individuals for
determining equations. Individuals in sample had diameters at breast height to 1.30m ranging
from 10.1 to 112.4 cm. Commercial heights were contemplated minimum 4 m, with
maximum values 30 m. With volumetric equations by vegetation type (n = 216 fallen trees), it
was found that of single entry Husch and double entry Schumacher-Hall (Ln) generated the
best for all types vegetation, demonstrating independence of factor. For validation of
volumetric equations of fallen trees were used the actual volume of 216 other fallen trees that
were not used in construction of equations. Individuals in sample had diameters at breast
height to 1.30m ranging from 10.3 to 123.4 cm. Commercial heights were contemplated
minimum of 3 m, with maximum values 28.9 m. The actual volume of individuals comprises
0.05 to 15.62 m³, with average of 1.7533 m³ and standard error of 0.1757 m³. The volume
cylindrical showed 1.4953 m³ (average) and 0.1537 m³ (standard error). Models Husch and
Schumacher-Hall (Ln) were selected for validation of use and application of volumetric
equations in types vegetation. When compared volumes, it was not significant differences
between estimates generated by simple equations (Husch) and double entry (Schumacher-Hall
(Ln)) of real volume (t = 0.81, p = 0.42, t = 0.62, p = 0.53, respectively). Result also found for
volume cylindrical (t = 10.52, p = .30). The diametric structures and hypsometric fallen trees
from the Amapa State Forest represent the structure of living trees alive into different
vegetation types. Thus, volumetric equations developed in this work are enabled to use to
estimate of volume of areas for sustainable forest management.
Key words: Volume estimate; Cubing rigorous; Structure

30
2.1 INTRODUÇÃO
As florestas inequiâneas na Amazônia geram grandes quantidades de matéria orgânica
em diversas classes de tamanho das árvores. Hoje, o bom senso indica o aproveitamento,
comercial ou não, destas árvores, evitando a utilização exagerada do método destrutivo de
seleção. No entanto, é necessário se obter dados sobre a quantidade de resíduos disponíveis e
quão estes podem gerar subsídios para a análise do balanço diamétrico e possíveis
implicações no manejo.
O uso de modelos matemáticos capazes de determinar com precisão a produção
florestal, a partir da estimativa do volume de madeira, é fundamental para implantação do
manejo sustentável (BARROS e SILVA JÚNIOR, 2009). No entanto, ainda existem algumas
dificuldades no gerenciamento da floresta, principalmente quanto à obtenção confiável do
volume de madeira em “pé”, o qual depende de atributos de árvores inventariadas.
As técnicas mais utilizadas possuem um caráter destrutivo de derrubada da árvore para
obtenção do volume de árvores individuais por meio da cubagem rigorosa. O método
geométrico, que utiliza o volume cilíndrico multiplicado por um fator de forma, na tentativa
de corrigir a forma da árvore, é uma alternativa à amostragem destrutiva, mas, normalmente, é
pouco preciso e genérico.
Segundo Thaines et al (2010), a necessidade de quantificação de estoque de matéria-
prima nos leva a buscar métodos de estimativa do volume comercial de madeira das árvores,
que possibilite quantificar o estoque de maneira eficiente e precisa, norteando decisões que
culminem na máxima produção e rentabilidade da floresta.
A atividade de cubagem rigorosa do volume geralmente é feita de maneira separada
das medições das unidades amostrais, o que gera aumento no custo do inventário. Este é o
método direto mais utilizado na rotina de inventários e consiste na medição sucessiva de
diâmetros ao longo do tronco, dividindo-o em seções que variam de comprimento de acordo
com o medidor (LEITE e ANDRADE, 2002).
No entanto, a utilização de dados da cubagem rigorosa pode fornecer informações
individuais das árvores que representam o volume da floresta. A aplicação dessas informações
em modelos de regressão, de acordo com Schneider et al. (2009), tem sido a solução de
grande parte de problemas florestais, especialmente quando se pretende obter estimativas de
parâmetros da floresta, utilizando-se de relações biométricas.
Segundo Silva (2008), os modelos de regressão linear são aplicados para estimar
volumes de árvores individuais. Nesses modelos, simples ou múltiplos, os requisitos de

31
normalidade de erros, de independência nas observações e de variância constante são
usualmente assumidos.
Entretanto, os resultados gerados devem ser interpretados a partir da relação dos
índices e variáveis estatísticas aliadas a fatores ecológicos. No caso da utilização de árvores
caídas da floresta como base do cálculo, ainda possuem a vantagem de redução dos custos
operacionais.
No tocante ao estudo de volumetria, o desenvolvimento de equações torna-se
imprescindível na aplicação de planos de manejo para definir a capacidade produtiva florestal
e garantir o fluxo contínuo dos produtos. No art. 10 da Resolução CONAMA 406/2009, é
descrito que a partir do segundo Plano Operacional Anual-POA, o cálculo do volume de
árvores em “pé” será aceito apenas mediante equação de volume desenvolvida
especificamente para o Plano de Manejo Florestal Sustentável.
Outro ponto importante que deve ser considerado para o desenvolvimento de equações
esta relacionado às variações das unidades florísticas e fisionômicas da região amazônica.
Tendo em vista que a classificação da vegetação pode ser explicada, por meio de alterações
dos parâmetros estruturais como riqueza, densidade, frequência e dominância. Contudo, as
informações sobre a distribuição diamétrica e hipsométrica tendem a ser uniformes em
florestas inequiâneas, independentemente das fitofisionomias.
Diante do exposto, este trabalho questiona se uma equação volumétrica de árvores
caídas pode ser utilizada para estimar com confiança o volume das árvores vivas e se pode ser
empregada em distintas fitofisionomias florestais. Partindo do pressuposto que a partir da
estrutura das árvores caídas é possível entender a estrutura da comunidade florestal matriz,
gerou-se a hipótese que as equações geradas com árvores caídas podem ser usadas para
predição de volumes de árvores em pé em distintas fitofisionomias.
No caso das florestas amapaenses, há necessidade de ampliar o número de
informações, bem como simular a parametrização dos dados de inventários locais para
aumentar a confiança na predição. Assim, o presente trabalho visa gerar uma equação
volumétrica por meio de dados biométricos de árvores caídas, capazes de estimar o estoque
real de madeira em distintas fitofisionomias do Amapá, Amazônia, Brasil.

32
2.2 MATERIAL E MÉTODOS
2.2.1 Caracterização da Área
O trabalho foi realizado na unidade de conservação de uso sustentável denominada
Floresta Estadual do Amapá (FLOTA/AP), localizada na região central do estado do Amapá,
Amazônia, Brasil. A unidade compreende uma área descontínua de 2.369.400 ha dividida em
4 módulos produtivos, definidos em função de infra-estrutura e vias de acessos para dinamizar
os pólos de desenvolvimento do estado.
O clima da região é do tipo Am (equatorial super-úmido), segundo a classificação de
Köppen. A temperatura média é 25 ºC, ocorrendo pouca variação de temperatura durante o
ano. A precipitação anual na região varia de 2.700 a 2.900 mm, com índice pluviométrico
trimestral mais seco abaixo de 200 mm. O período mais chuvoso ocorre no trimestre de março
a maio (acima de 1.000 mm). A umidade relativa é alta, com valores entre 80 e 86% (Souza e
Cunha, 2010).
A vegetação predominantemente é floresta ombrófila densa de baixos platôs e
submontana, bem como transição cerrado-floresta (IBGE, 2012). Considerando toda a área da
FLOTA/AP também podem ser encontrados ambientes de mangue, campos e várzea em
pequenas áreas.
As principais fitofisionomias da Floresta Estadual do Amapá são descritas abaixo
quanto às espécies arbóreas mais importantes, presença de afloramento rochoso, declividade
(IEF, 2009), geologia, solos (RADAM, 1974) e altitudes.
Floresta Ombrófila Densa de terras baixas
Apresenta dossel de alto porte, até 50 m, perenifólia, em planícies costeiras. Estima-se
que o número de espécies arbóreas que compõem esta fitofisionomia gira em torno de 245,
com destaque para Eschweilera coriacea (DC.) S.A. Mori, Pouteria caimito (Ruiz & Pav.)
Radlk., Protium tenuifolium (Engl.) e Minquartia guianensis Aubl. Do total de espécies, 190
possuem valor comercial, gerando uma densidade de 27 ind.ha-1
, 133 m3.ha
-1 e 48 % do total
de volume de madeira (DAP > 10cm) da fitofisionomia.
Os afloramentos rochosos são encontrados com baixa intensidade. A declividade varia
de ondulada a fortemente ondulada. Possui formação geológica pré-cambriana, com
superfícies pedi planado por canais geralmente curtos, numerosos e pouco aprofundado. O
solo predominante é tipo Latossolo Vermelho-Amarelo distrófico. A altitude varia de 60 a
100 m.

33
Floresta Ombrófila Densa Submontana
Situada em encostas, com dossel uniforme e de alto porte (até 50 m), perenifólia. O
número de espécies arbóreas é de aproximadamente 176, com destaque para Eschweilera
coriacea (DC.) S.A. Mori, Pouteria caimito (Ruiz & Pav.) Radlk., Inga auristellae Harms,
Guarea pubescens subsp. pubiflora (A. Juss.) T.D. Penn e Vouacapoua americana Aubl.
Cento e dezessete (117) espécies arbóreas identificadas possuem valor comercial, o que
representa 26 ind.ha-1
, 142 m3.ha
-1 e 49 % do total de volume (DAP > 10cm) por hectare da
fitofisionomia.
Os afloramentos rochosos são encontrados de baixa a moderada intensidade. A
declividade varia de ondulada a fortemente ondulada. Possui formação geológica pré-
cambriana, com relevo montanhoso e medianamente profundo. O solo predominante é tipo
Latossolo Vermelho-Amarelo distrófico. A altitude máxima registrada é 320 m, com média de
195 m.
Transição Cerrado – Floresta
Situada em área mais próxima do litoral, com dossel uniforme e de médio porte (até 30
m), perenifólia. Foram registrados nesta fitofisionomia cerca de 203 espécies arbóreas, com
maior representatividade da Pouteria caimito (Ruiz & Pav.) Radlk., Protium decandrum
(Aubl.) March. e Guarea pubescens subsp. pubiflora (A. Juss.) T.D. Penn, bem como espécies
típicas do cerrado: Ormosia coccínea Jacks., Perebea sp., Cheiloclinium cognatum (Miers)
A.C.Sm. Cento e vinte quatro (124) espécies arbóreas identificadas possuem valor comercial,
o que representa 27 ind.ha-1
, 129 m3.ha
-1 e 48 % do total de volume por hectare da
fitofisionomia.
Os afloramentos rochosos são encontrados de moderada a alta intensidade. A
declividade é inexistente (plana). Possui formação geológica pré-cambriana, com superfícies
pedi planado por drenagem pouco profunda. O solo predominante é tipo Latossolo Amarelo
distrófico e concrecionários lateríticos. A altitude média encontrada gira em torno de 60 m.
2.2.2 Estrutura Florestal
O inventário contemplou as árvores caídas presentes em 30 unidades amostrais locadas
aleatoriamente em três distintas fitofisionomias. O número de unidades amostrais foi
proporcional às áreas das fitofisionomias, distribuídas em vinte e três (23) na Floresta

34
Ombrófila Densa de Baixos Platôs, quatro (4) na Floresta Ombrófila Densa Submontana e três
(3) na Transição Cerrado-Floresta.
As unidades amostrais foram estabelecidas seguindo o sentido Norte-Sul / Leste-
Oeste, a partir de faixas contínuas de 20 x 2500 m em forma de cruz. Todas as árvores caídas
com diâmetro à altura do peito a 1,30 m do solo (DAP) > 10 cm foram submetidas à cubagem
rigorosa por meio dos procedimentos descritos por Smalian para determinação do volume
comercial.
As árvores caídas amostradas foram divididas em dez classes diamétricas com
amplitude de 10 cm. A primeira contemplou indivíduos com 10 < DAP < 20 cm e a última
todos os indivíduos > 100 cm.
A representatividade do número de indivíduos da amostra foi realizada com limite de
20 % de erro, a um nível de significância de α > 0,05.
2.2.3 Equação Volumétrica
Para construção das equações, os volumes outliers máximos e mínimos foram
excluídos, bem como árvores com discrepância nas medidas longitudinais do fuste. Logo,
vinte por cento (20 %) das árvores caídas inventariadas foram selecionadas
proporcionalmente as classes diamétricas para representar as fitofisionomias.
Os dados coletados foram submetidos aos testes de normalidade de Kolmogorov-
Smirnov e homocedasticidade de F-Snedecor. Quando verificado a não normalidade e/ou
variâncias heterogêneas foi realizado a transformação logarítmica.
Estabelecida à preparação dos dados, foram testados modelos volumétricos comuns e
usuais na região amazônica (COUTO e BASTOS, 1987; GOMES e GARCIA, 1993;
MACHADO et al., 2000; ENCINAS e KLEINN, 2001; GIRARDI, 2005; ROLIM et al., 2006;
LEITE, 2008; BARROS e SILVA JUNIOR, 2009; ENCINAS et al., 2009; LEITE e
REZENDE, 2010).
Dos modelos volumétricos utilizados, cinco foram de simples entrada (dois aritméticos
e três não lineares), tendo o DAP como variável independente; e oito de dupla entrada (cinco
aritméticos e três não lineares), tendo o DAP e altura comercial do fuste (H), combinadas ou
não, como variáveis independentes (Tabela 1).

35
Tabela 1. Modelos volumétricos utilizados para representar a volumetria da Floresta Estadual do Amapá,
Amazônia.
Modelo Autor
Simples Entrada
V= β0 + β1 DAP² + ε Koperzky e Gehrhardt
V= β0 + β1 DAP + β2 DAP² + ε Hohenald e kreen
Ln (V)= β0 + ln(DAP) β1+ ε Husch
Ln (V) = β0 + β1 ln DAP + β2 DAP-1
+ ε Brenac
V = β0 * DAP β1
+ ε Husch- Não linear
Dupla Entrada
V= β0 + β1 (DAP² .H) + ε Spurr
V= β0 + β1 .DAP ²+ β2 DAP² H + β3 H + ε Stoate
V= β0 + β1. DAP ²+ β2 DAP² .H + β3 DAP.H² + β4 H² + ε Naslund
V= β0 + β1 .DAP + β2 DAP² + β3 DAP.H + β4 DAP².H + β5 H + ε Meyer
V = β0 + β1.DAP + β2.DAP² + β3 DAP.H + β4 DAP².H + ε Meyer modificada
Ln (V) = β0 + β1 ln (DAP² .H) + ε Spurr (Ln)
Ln (V) = β0 + β1 ln DAP + β2 ln H + ε Schumacher e Hall
V = β0 * DAP β1
* H β2
+ ε Schumacher-Hall-Não linear
Os coeficientes dos modelos foram estimados pelo método dos mínimos quadrados. A
seleção dos coeficientes foi realizada retirando-se aquelas que não apresentaram nível de
significância p < 0,05.
A determinação da melhor equação foi obtida através de comparações dos coeficientes
de determinação ajustado (R2
aj), erro padrão da estimativa (SYX) e análise da variância
(ANOVA). Também foi considerada a análise dos resíduos a partir do Desvio Médio
Percentual (DMP), gerada a partir dos valores observados e estimados do volume, com intuito
de verificar a magnitude de subestimativa ou superestimativa dos volumes estimados pelos
modelos.
Para os modelos logarítmicos, o erro padrão da estimativa foi corrigido na escala
original da variável dependente, para possibilitar a comparação com os modelos aritméticos.
Essa correção foi realizada com o índice de Furnival (FURNIVAL, 1961)
2.2.4 Validação da Equação Volumétrica

36
Para validação das equações volumétricas, cerca de 15 % dos indivíduos (não
utilizados na construção das equações) foram selecionados proporcionalmente ao número de
indivíduos das classes diamétricas e estimado o volume.
As estimativas volumétricas da testemunha (volume real das árvores, obtido pela
cubagem rigorosa) foi comparada com o volume estimado pela equação de simples entrada
com melhor ajuste dos dados, com o volume estimado pela equação de dupla entrada com
melhor ajuste dos dados e com o volume cilíndrico com fator de forma 0,7.
O volume gerado por cada método foi comparado com o volume real da testemunha
por meio do teste t pareado. As análises foram executadas no software STATISTICA TRIAL.
2.3 RESULTADOS
2.3.1 Estrutura Florestal
Foram inventariadas em toda área cerca de 1449 indivíduos (árvores caídas),
distribuídos 1026 na fitofisionomia de Floresta Ombrófila Densa de Baixo Platô (FODBP),
70,81 % do total, seguida da Floresta Ombrófila Densa Submontana (FODSm) com 15,60 % e
a Transição Cerrado-Floresta (TCF) com 13,60%.
O número de árvores caídas para a Floresta Estadual do Amapá foi estimado em 4,95
± 1,00 indivíduos.ha-1
, com um volume médio de 10,45 ± 1,39 m3.ha
-1 e altura comercial
variando de 3,5 a 36,7 m. O erro de amostragem foi de 20,4 % para o número de indivíduos e
13,2 % para o volume (Tabela 2).
Tabela 2. Suficiencia amostral para numero de indivíduos e volume por hectare na Floresta Estadual do Amapá.
FODTB – Floresta ombrofila densa de terras baixas; FODSm – Floresta ombrofila densa submontana; TCF –
Transição cerrado-floresta. N.U.L – número unidades locadas; N.O.U – numero ótimo de unidades; IC –
intervalo de confiança; E.A. – erro de amostragem.
Indivíduos / ha Volume (m³/ ha)
FODTB FODSm TCF Total FODTB FODSm TCF Total
N. U. L. 23 4 3 30 23 4 3 30
N. O. U. 22 4 5 31 10 2 2 14
I. C. 4,53 ± 0,95 5,6 ± 5 6,7 ± 2,24 4,95 ± 1 10,32 ± 1,79 8,52 ± 2,85 13,74 ± 3,07 10,45 ± 1,39
E. A. (%) 20,4 13,2
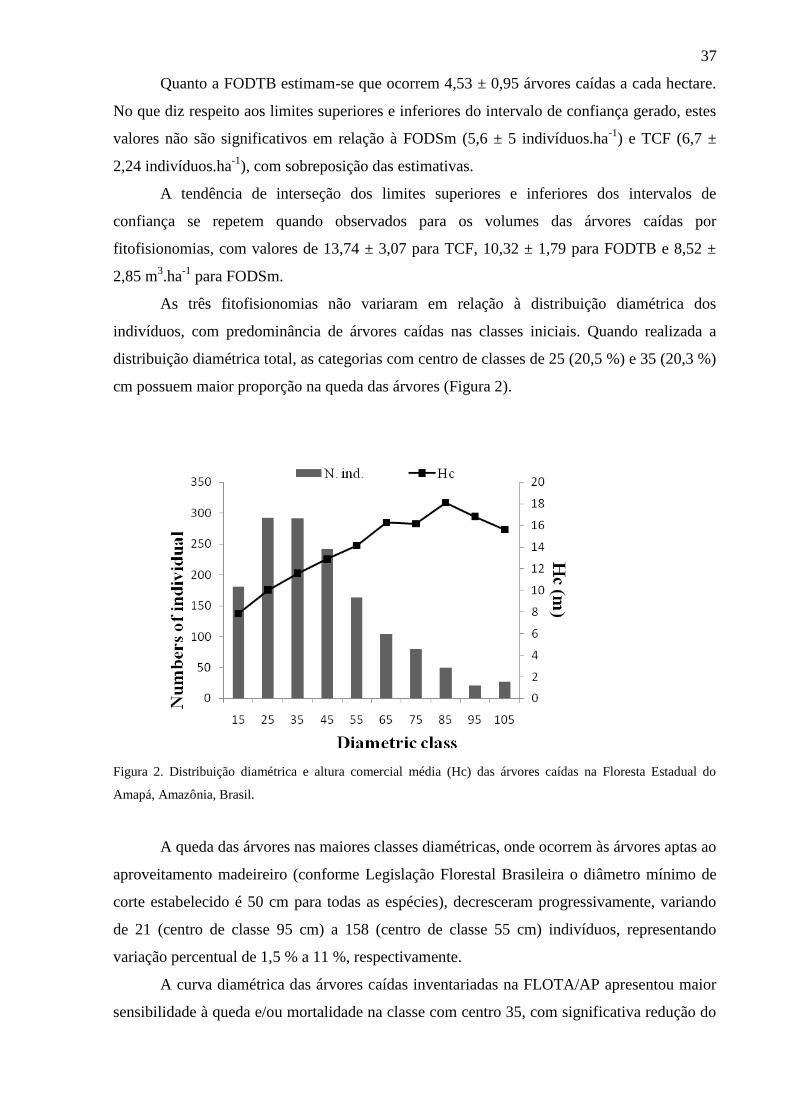
37
Quanto a FODTB estimam-se que ocorrem 4,53 ± 0,95 árvores caídas a cada hectare.
No que diz respeito aos limites superiores e inferiores do intervalo de confiança gerado, estes
valores não são significativos em relação à FODSm (5,6 ± 5 indivíduos.ha-1
) e TCF (6,7 ±
2,24 indivíduos.ha-1
), com sobreposição das estimativas.
A tendência de interseção dos limites superiores e inferiores dos intervalos de
confiança se repetem quando observados para os volumes das árvores caídas por
fitofisionomias, com valores de 13,74 ± 3,07 para TCF, 10,32 ± 1,79 para FODTB e 8,52 ±
2,85 m3.ha
-1 para FODSm.
As três fitofisionomias não variaram em relação à distribuição diamétrica dos
indivíduos, com predominância de árvores caídas nas classes iniciais. Quando realizada a
distribuição diamétrica total, as categorias com centro de classes de 25 (20,5 %) e 35 (20,3 %)
cm possuem maior proporção na queda das árvores (Figura 2).
Figura 2. Distribuição diamétrica e altura comercial média (Hc) das árvores caídas na Floresta Estadual do
Amapá, Amazônia, Brasil.
A queda das árvores nas maiores classes diamétricas, onde ocorrem às árvores aptas ao
aproveitamento madeireiro (conforme Legislação Florestal Brasileira o diâmetro mínimo de
corte estabelecido é 50 cm para todas as espécies), decresceram progressivamente, variando
de 21 (centro de classe 95 cm) a 158 (centro de classe 55 cm) indivíduos, representando
variação percentual de 1,5 % a 11 %, respectivamente.
A curva diamétrica das árvores caídas inventariadas na FLOTA/AP apresentou maior
sensibilidade à queda e/ou mortalidade na classe com centro 35, com significativa redução do
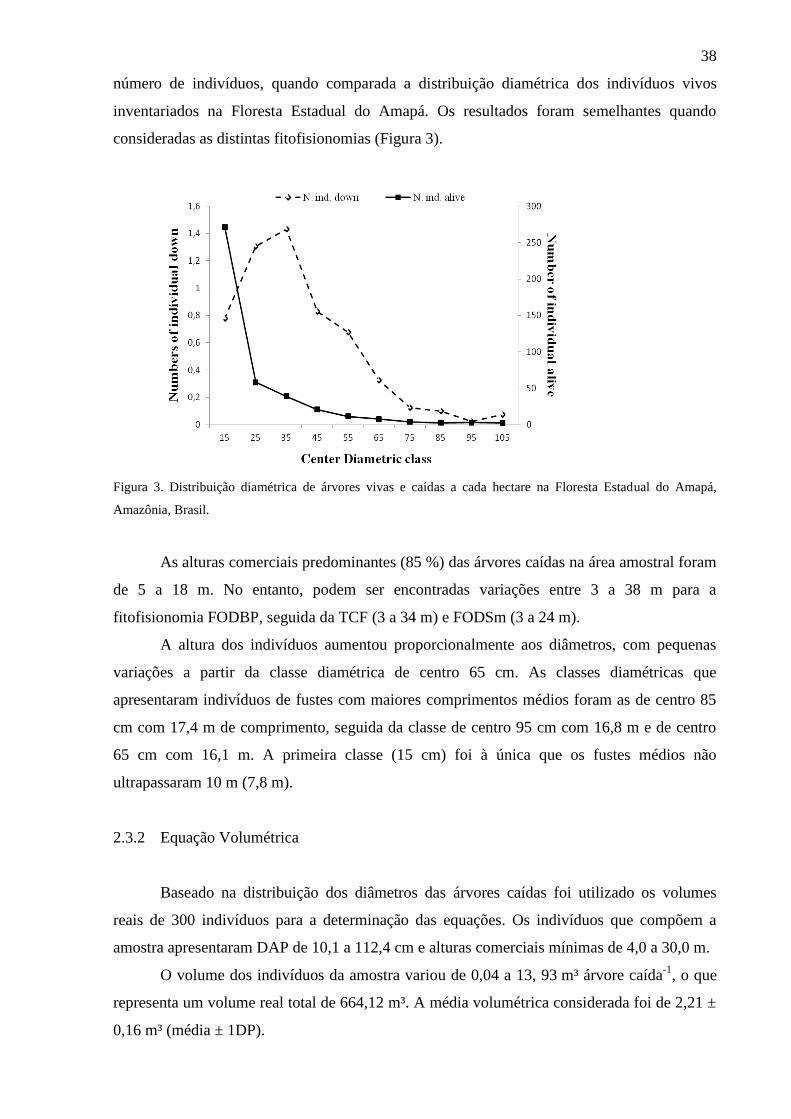
38
número de indivíduos, quando comparada a distribuição diamétrica dos indivíduos vivos
inventariados na Floresta Estadual do Amapá. Os resultados foram semelhantes quando
consideradas as distintas fitofisionomias (Figura 3).
Figura 3. Distribuição diamétrica de árvores vivas e caídas a cada hectare na Floresta Estadual do Amapá,
Amazônia, Brasil.
As alturas comerciais predominantes (85 %) das árvores caídas na área amostral foram
de 5 a 18 m. No entanto, podem ser encontradas variações entre 3 a 38 m para a
fitofisionomia FODBP, seguida da TCF (3 a 34 m) e FODSm (3 a 24 m).
A altura dos indivíduos aumentou proporcionalmente aos diâmetros, com pequenas
variações a partir da classe diamétrica de centro 65 cm. As classes diamétricas que
apresentaram indivíduos de fustes com maiores comprimentos médios foram as de centro 85
cm com 17,4 m de comprimento, seguida da classe de centro 95 cm com 16,8 m e de centro
65 cm com 16,1 m. A primeira classe (15 cm) foi à única que os fustes médios não
ultrapassaram 10 m (7,8 m).
2.3.2 Equação Volumétrica
Baseado na distribuição dos diâmetros das árvores caídas foi utilizado os volumes
reais de 300 indivíduos para a determinação das equações. Os indivíduos que compõem a
amostra apresentaram DAP de 10,1 a 112,4 cm e alturas comerciais mínimas de 4,0 a 30,0 m.
O volume dos indivíduos da amostra variou de 0,04 a 13, 93 m³ árvore caída-1
, o que
representa um volume real total de 664,12 m³. A média volumétrica considerada foi de 2,21 ±
0,16 m³ (média ± 1DP).

39
Todos os modelos testados apresentaram distribuições normais dos resíduos. No
entanto, em modelos de simples entrada (Koperzky-Gehrhardt, Hohenald-kreen e Husch -
Não linear) e de dupla entrada (Meyer modificada) foram constatados a heterocedasticidade
dos resíduos.
A multicolinearidade entre variáveis independentes foi verificada nos modelos de
simples entrada de Hohenald-kreen e Brenac com correlações de 0,9673 e 0,9549,
respectivamente. Fato também constatado em alguns modelos de dupla entrada (Stoate,
Naslund, Meyer e Meyer modificada), com correlação na ordem de 88 a 96 %.
Quando geradas as equações por fitofisionomia, foi verificado que os modelos de
simples entrada de Husch e dupla entrada de Schumacher-Hall (Ln) geraram os melhores
ajustes estatísticos na Floresta ombrófila densa de terras baixas, submontana e transição
cerrado-floresta.
As equações volumétricas totais, geradas para as três fitofisionomias, também foram
significativas nos ajustes das árvores caídas, com valores de Fcal.variando de 345,1782
(p<0,0001; Meyer) a 2550,4323 (p<0,0001; Schumacher-Hall (Ln)).
A estimativa dos coeficientes de determinação ajustados (R²aj) proporcionou distintos
ajustes das variáveis dendrométricas (Tabela 3). As equações de simples entrada de
Koperzky-Gehrhardt, Hohenald-Krenn e Husch - Não linear apresentaram valores em torno
de 70 %.
Para as equações de dupla entrada, foram obtidos coeficientes de determinação
ajustados (R²aj) de 56 ( purr ( n)), 83 ( purr e toate), 84 ( slund, Meyer
modificada e Schumacher-Hall-Não linear) e 85 % (Meyer). Entretanto, quando utilizados
modelos logaritmizados houve ajustes mais representativos (Husch e Brenac - 89 %;
Schumacher-Hall (Ln) - 94 %), e maior eficiência no ajuste dos dados (Tabela 4).
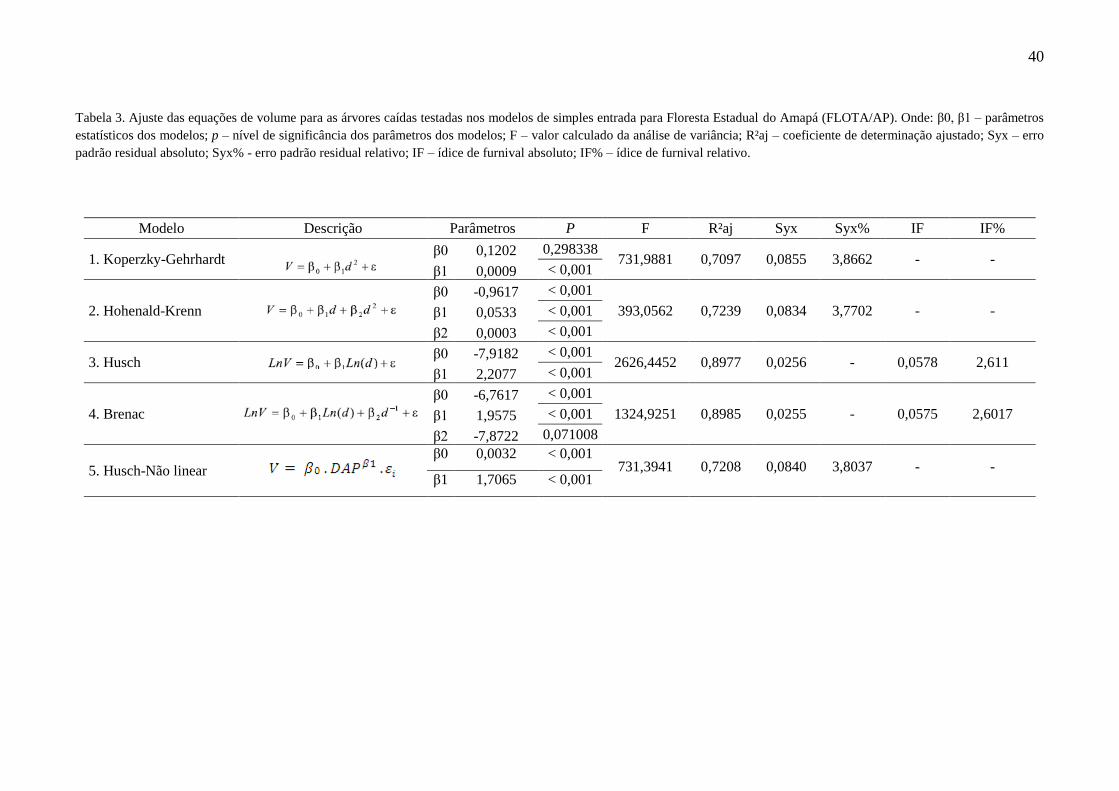
40
Tabela 3. Ajuste das equações de volume para as árvores caídas testadas nos modelos de simples entrada para Floresta Estadual do Amapá (FLOTA/AP). Onde: β0, β1 – parâmetros
estatísticos dos modelos; p – nível de significância dos parâmetros dos modelos; F – valor calculado da análise de variância; R²aj – coeficiente de determinação ajustado; Syx – erro
padrão residual absoluto; Syx% - erro padrão residual relativo; IF – ídice de furnival absoluto; IF% – ídice de furnival relativo.
Modelo Descrição Parâmetros P F R²aj Syx Syx% IF IF%
1. Koperzky-Gehrhardt
β0 0,1202 0,298338 731,9881 0,7097 0,0855 3,8662 - -
β1 0,0009 < 0,001
2. Hohenald-Krenn
β0 -0,9617 < 0,001
393,0562 0,7239 0,0834 3,7702 - - β1 0,0533 < 0,001
β2 0,0003 < 0,001
3. Husch
β0 -7,9182 < 0,001 2626,4452 0,8977 0,0256 - 0,0578 2,611
β1 2,2077 < 0,001
4. Brenac
β0 -6,7617 < 0,001
1324,9251 0,8985 0,0255 - 0,0575 2,6017 β1 1,9575 < 0,001
β2 -7,8722 0,071008
5. Husch-Não linear
β0 0,0032 < 0,001 731,3941 0,7208 0,0840 3,8037 - -
β1 1,7065 < 0,001
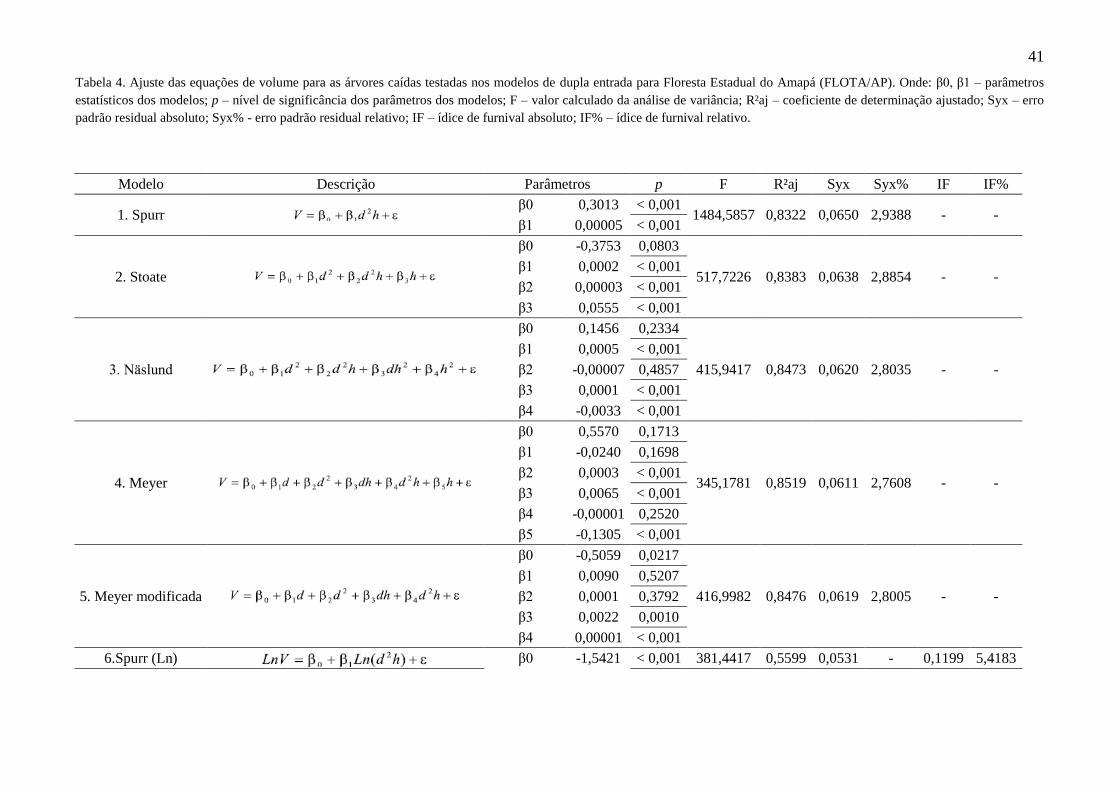
41
Tabela 4. Ajuste das equações de volume para as árvores caídas testadas nos modelos de dupla entrada para Floresta Estadual do Amapá (FLOTA/AP). Onde: β0, β1 – parâmetros
estatísticos dos modelos; p – nível de significância dos parâmetros dos modelos; F – valor calculado da análise de variância; R²aj – coeficiente de determinação ajustado; Syx – erro
padrão residual absoluto; Syx% - erro padrão residual relativo; IF – ídice de furnival absoluto; IF% – ídice de furnival relativo.
Modelo Descrição Parâmetros p F R²aj Syx Syx% IF IF%
1. Spurr
β0 0,3013 < 0,001 1484,5857 0,8322 0,0650 2,9388 - -
β1 0,00005 < 0,001
2. Stoate
β0 -0,3753 0,0803
517,7226 0,8383 0,0638 2,8854 - - β1 0,0002 < 0,001
β2 0,00003 < 0,001
β3 0,0555 < 0,001
3. slund
β0 0,1456 0,2334
415,9417 0,8473 0,0620 2,8035 - -
β1 0,0005 < 0,001
β2 -0,00007 0,4857
β3 0,0001 < 0,001
β4 -0,0033 < 0,001
4. Meyer
β0 0,5570 0,1713
345,1781 0,8519 0,0611 2,7608 - -
β1 -0,0240 0,1698
β2 0,0003 < 0,001
β3 0,0065 < 0,001
β4 -0,00001 0,2520
β5 -0,1305 < 0,001
5. Meyer modificada
β0 -0,5059 0,0217
416,9982 0,8476 0,0619 2,8005 - -
β1 0,0090 0,5207
β2 0,0001 0,3792
β3 0,0022 0,0010
β4 0,00001 < 0,001
6.Spurr (Ln)
β0 -1,5421 < 0,001 381,4417 0,5599 0,0531 - 0,1199 5,4183
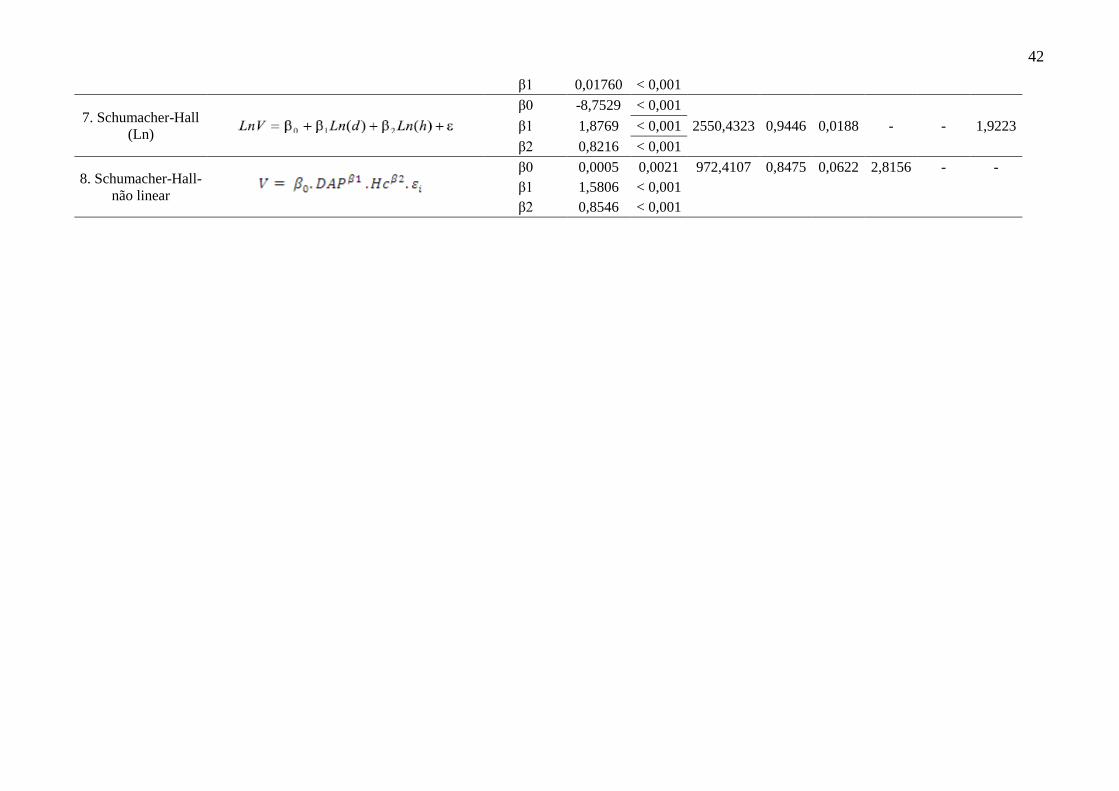
42
β1 0,01760 < 0,001
7. Schumacher-Hall
(Ln)
β0 -8,7529 < 0,001
2550,4323 0,9446 0,0188 - - 1,9223 β1 1,8769 < 0,001
β2 0,8216 < 0,001
8. Schumacher-Hall-
não linear
β0 0,0005 0,0021 972,4107 0,8475 0,0622 2,8156 - -
β1 1,5806 < 0,001
β2 0,8546 < 0,001

43
Nas equações de simples entrada, os modelos aritméticos e Husch-Não linear
obtiveram um erro padrão residual (Sxy) de aproximadamente 3,8 %. Enquanto, os modelos
logarítmicos apresentaram estimativas mais precisas (2,6 %), determinadas pelo índice de
furnival.
erro padrão residual para os modelos de dupla entrada ( purr, toate, slund,
Meyer, Meyer modificada e Schumacher-Hall-Não linear) variaram entre 2,7 e 2,9 %.
Entretanto, dois modelos extrapolaram este intervalo, com amplitude máxima para o modelo
de Spurr (Ln) (5,4 %), e mínima Schumacher-Hall (Ln) (1,9 %).
O desvio médio percentual (DMP), nos modelos de simples entrada (-68,24 a 14,19 %)
teve uma variação quanto à distribuição dos resíduos semelhantemente aos modelos de dupla
entrada (-60,25 a 42,13 %).
Das equações geradas, seis subestimaram o volume das árvores caídas (dois de simples
e quatro de dupla entrada). A superestimação do volume ocorreu nos modelos de Meyer
(5,48 %), Hohenald-Krenn (9,82 %), Brenac (14,19 %), Meyer modificada (17,03 %) e Spurr
(Ln) (42,13 %), em valores positivos.
Nos modelos de simples entrada (Husch-Não linear, Koperzky-Gehrhardt e Hohenald-
Krenn), pode-se observar que a grande limitação é o ajuste dos maiores volumes das árvores
caídas. À medida que há o aumento do volume individual das árvores caídas, a variação dos
resíduos aumenta positiva e negativamente (Figura 4).
Essa limitação também é observada nos modelo de Spurr, Stoate, Naslund, Meyer,
Meyer modificada e Schumacher-Hall-Não linear. Esse comportamento é típico de resíduos
heterocedásticos, com formação gráfica de um cone (Figura 5).

44
Figura 4. Análise de distribuição gráfica de resíduos dos modelos de simples entrada para equação de árvores
caídas na Floresta Estadual do Amapá. Onde: 1 - Koperzky-Gehrhardt; 2 - Hohenald-Krenn; 3 – Husch; 4 –
Brenac; 5 – Husch-Não linear.
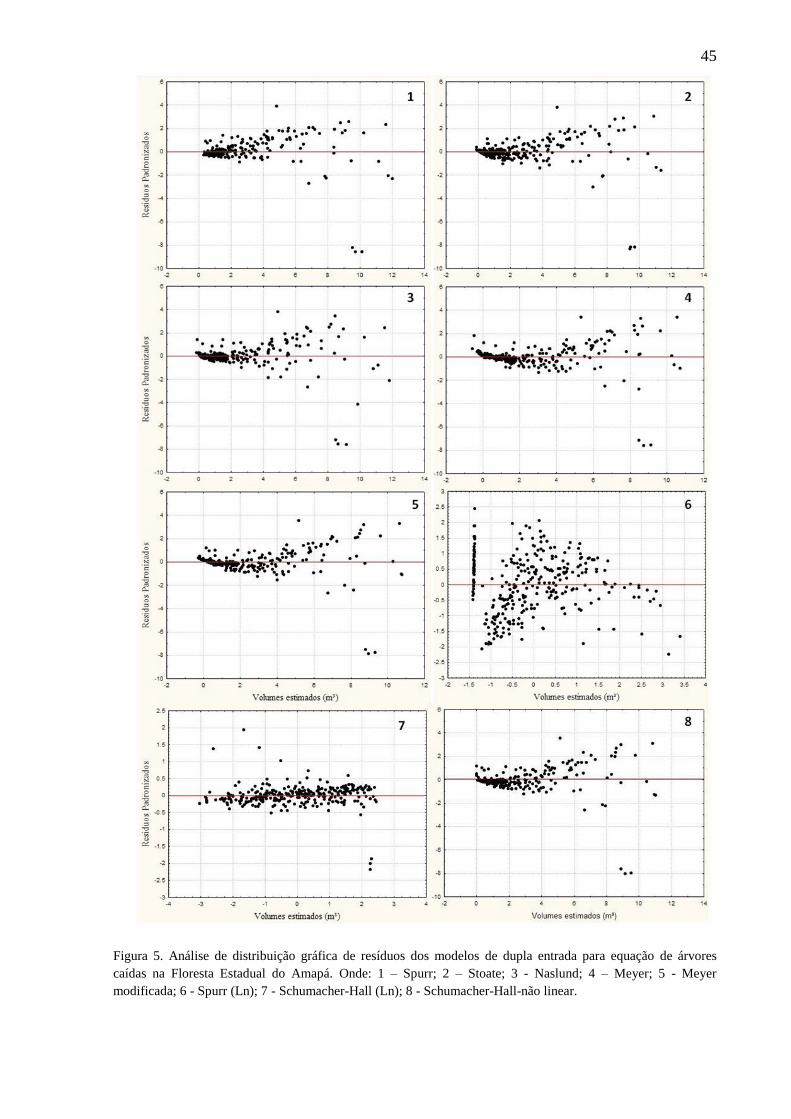
45
Figura 5. Análise de distribuição gráfica de resíduos dos modelos de dupla entrada para equação de árvores
caídas na Floresta Estadual do Amapá. Onde: 1 – Spurr; 2 – Stoate; 3 - Naslund; 4 – Meyer; 5 - Meyer
modificada; 6 - Spurr (Ln); 7 - Schumacher-Hall (Ln); 8 - Schumacher-Hall-não linear.

46
As equações mais significativas foram as formadas pelos modelos de Schumacher-
Hall (Ln) e Husch, com DMP de 2,51 e 2,52 %. Esses valores exprimem o quanto os resíduos
estão distribuídos de forma homogênea na amostra de árvores caídas.
Os modelos de Meyer (5,48 %), Hohenald-Krenn (9,82 %) e Brenac (14,19 %),
também apresentaram bons resultados. Na distribuição dos resíduos verifica-se que os
modelos de Meyer e Hohenald-Krenn estimam com eficiência o volume de pequenas árvores
caídas. No entanto, o modelo de Brenac apesar de possuir maior DMP produz confiabilidade
nas estimativas com maior amplitude de volume de árvores caídas.
2.3.3 Validação da Equação Volumétrica
Para validação das equações volumétricas das árvores caídas foram utilizados os
volumes reais de 216 indivíduos que não fizeram parte da amostragem para ajuste das
equações. Os indivíduos que compõem a amostra apresentaram diâmetros à altura do peito a
1,30 m do solo variando de 10,3 a 123,4 cm. As alturas comerciais mínimas contempladas
foram de 3 m, com valores máximos de 28,9 m.
O volume real médio dos indivíduos foi de 1,7533 ± 0,1757 m³ (média ± erro padrão)
com valores variando de 0,05 a 15,62 m³. A expressão do volume cilíndrico médio com fator
de forma, tradicional em inventários e planos de manejos florestais brasileiros, apresentou
1,4953 ± 0,1537 m³.
Os modelos de Husch e Schumacher-Hall (Ln) foram selecionados para validação de
uso e aplicação das equações volumétricas nas fitofisionomias estudadas. A equação a partir
do modelo de Husch gerou um volume médio de 1,6230 ± 0,1528 m³, enquanto a de
Schumacher-Hall (Ln) de 1,5935 ± 0,1493 m³.
Quando comparado os volumes, não foram observadas diferenças significativas entre
as estimativas geradas pelas equações de simples (Husch; t = 0,81 e p = 0,42) e dupla entrada
(Schumacher-Hall (Ln); t = 0,62 e p = 0,53) com o volume real. Resultado também
encontrado para o volume cilíndrico com fator de forma (t = 10,52 e p = 0,30).
As comparações entre volumes estimados pelas equações (t = 0,04 e p = 0,96) e
expressão do cilindro com fator de forma (Husch; t = 0,26 e p = 0,80 e Schumacher-Hall (Ln);
t = 0,31 e p = 0,76) também não foram diferentes.

47
2.4 DISCUSSÃO
2.4.1 Estrutura florestal
A confiabilidade das estimativas do número de árvores caídas por fitofisionomia
variou em função do número de unidades amostrais estabelecidas. Entretanto, não foram
observadas variações para afetar a representatividade dos volumes.
Quando comparada com a estrutura diamétrica das árvores vivas em “pé”, as
distribuições de indivíduos proporcionaram curvas exponenciais similares às árvores caídas
por fitofisionomia. Estes comportamentos são típicos de florestas inequiâneas, o que também
leva a pensar que as árvores que caem apresentam proporcionalidade com a vegetação
remanescente, portanto capaz de possibilitar quaisquer análises que represente a floresta em
“pé”.
Há evidencias que, em termos médios as florestas inequiâneas na Amazônia têm
percentual de 2 % de árvores mortas a cada ano (SILVA, 2012). Em termos absolutos há
maior mortalidade de árvores nas menores classes de diâmetros, entretanto, em termos
relativos é nas maiores classes de diâmetro que ocorre a maior mortalidade, já que a
distribuição decrescente típica das florestas nativas deixa claro um menor número de árvores
nestas classes (SCOLFORO et al., 1996).
Na Floresta Estadual do Amapá, foram estimados 399 (FODTB), 426 (FODSm) e 465
(TCF) indivíduos vivos.ha-1
, os quais proporcionaram volumes de 276.090, 291.024 e 267.005
m³ ha-1
, respectivamente (INAM, 2009). O pequeno aumento do número de árvores caídas na
TCF está relacionado à mudança de estrutura da floresta, que apresenta indivíduos com
menores dimensões, mais susceptível a intempéries climáticas. Entretanto, o número de
árvores caídas nas fitofisionomias, provavelmente, está relacionado à presença de
afloramentos rochosos e condições edafoclimáticas.
O decréscimo do número de árvores caídas das sucessivas classes diamétricas
configura uma suave curva tendendo a reta com declividade negativa, descendendo para
direita. Este comportamento é típico de comunidades florestais em estágio primário de
sucessão, com aptidão ao manejo florestal. Segundo Sterba (2004), freqüentemente a
estabilidade de distribuições diamétricas de florestas inequiâneas são pensadas com inúmeras
espécies, em distintas idades.
A distribuição diamétrica exponencial das árvores em florestas nativas, com
proporcionalidade constante em classes sucessivas, é o indicio de estabilidade na comunidade
(DE LIOCOURT, 1898). Outra forma de verificar o equilíbrio de uma floresta é assumir que

48
o número de árvores que ingressam numa classe diamétrica é igual à quantidade de árvore que
ingressam na classe imediatamente posterior (SCHUTZ, 1975), ou quando há grande
produção de sementes, germinação satisfatória e com taxa de mortalidade decrescente nas
idades mais avançadas (DAUBERNMIRe, 1968).
A exceção é a primeira classe diamétrica, onde geralmente nos inventários florestais
agrupam o maior número de indivíduos. No entanto, era esperada uma redução do número de
indivíduos na classe inicial, proporcionado pela velocidade de decomposição e mineralização
das árvores menores ao caírem, mesmo compostas por distintas espécies de variadas
densidades.
O efeito de componentes do solo e da vegetação sobre a percentagem de
decomposição de madeira de espécies arbóreas não está associada ao tipo de solo, topografia,
quantidade de liteira fina, biomassa e densidade arbórea (TOLEDO et al., 2012), e sim pelo
material de origem (dimensão da árvore, por exemplo), tempo, microorganismos e condições
microclimáticas do sítio (LEPSCH, 2002).
A estrutura hipsométrica das árvores caídas também foi similar ao das árvores vivas
em “pé” inventariadas em 2009 na F TA. s resultados das alturas comerciais não variaram
sigificativamente entre as fitofisionomias. As árvores vivas em “pé” da F DTB apresentaram
altura média, máxima e mínima de 21,3, 42 e 6 m, respectivamente. Seguida das árvores da
FODSm (média de 21,3 m, máxima de 39 m, mínima de 7m) e TCF (18,8 m, máxima de 42
m, mínima de 6 m).
2.4.2 Equação volumétrica
Os ajustes similares das equações por fitofisionomia é consequência da similaridade da
estrutura diamétrica e hipsométrica das áreas estudadas. As diferenças são observadas apenas
nas composições florísticas, não interferindo na estimativa volumétrica.
Os volumes das árvores caídas nas fitofisionomias apresentaram ajustes similares entre
os modelos de simples e dupla entrada. Esse comportamento dos modelos fornece a vantagem
de uso de equações com menor número de variáveis, tornando a atividade de romaneiro no
campo mais prática e com menor custo.
Estatísticamente, tal fato é explicado pela ausência de efeitos aditivos das variáveis
independentes em alguns modelos. A ocorrência de multicolinearidade proporciona
similaridade entre o poder explicativo adicional e correlação parcial dos diâmetros e/ou
alturas com o volume das árvores caídas, não deixando clara as diferenças de estimativas
quanto ao uso de uma única variável ou combinações.

49
Na medida que as variáveis independentes são correlacionadas, elas compartilham
uma parte do seu poder preditivo, ou seja, as duas variáveis independentes explicam uma
mesma parcela da variável dependente (CORRAR et al., 2007).
Resultados semelhantes foram encontrados por Tonini et al. (2005), trabalhando com
quatro espécies florestais nativas da Amazônia, com equações de volume de simples e dupla
entrada apresentando ajustes semelhantes.
Na Floresta Nacional do Tapajós, no trecho de Santarém a Cuiabá, Brasil, Silva e
Araujo (1984) desenvolveram uma equação de volume para qualquer espécie e para diâmetro
variando de 15 cm a 45 cm, com equações aritméticas apresentando resultados levemente
superiores às equações logarítmicas testadas.
Resultados similares foram encontrados por Ferreira et al (2011) os quais verificaram
que para estimativa de volume de árvores em florestas equianeas, a equação mais
aconselhável e eficiente é de simples entrada com DAP, quando comparada à equação de
Schumacher e Hall, pois a mesma não necessita de obtenção da variável altura, diminuindo
assim o tempo de obtenção de dados, ocasionando a redução de custos totais no trabalho.
Tonini e Melo (2009) verificaram que os modelos de Baden-Wurt, Schumacher-Hall e
Spurr (Ln) apresentaram melhor desempenho nos índices de R²aj, variando de 0,932 a 0,935.
O modelo Baden-Wurt apresentou os valores mais altos para R²aj, porém não foi o
selecionado por apresentar o erro padrão de estimativa elevado.
O menor ajuste de modelos de simples entrada é esperado quando são realizados
trabalhos com variáveis da vegetação, pois assume-se que árvores de mesmo diâmetro têm a
mesma altura, o que não é verdadeiro para florestas heterogêneas (ROLIM et al., 2006). A
pesquisa de Barros e Silva Jr. (2009) acrescenta ainda que se levando em consideração a
praticidade de uso da equação, a diminuição do trabalho de campo, deixando-se de medir a
variável altura, bem como os possíveis erros cometidos durante sua medição, o modelo de
Brenac (simples entrada) é preferido em relação ao modelo de Spurr (dupla entrada).
A precisão de modelos com diâmetro à altura do peito e volume em nível de população
demonstra que os modelos de afilamento são capazes de produzir previsões exatas do
diâmetro do caule e volumes, tanto da população quanto em nível de árvore individual
(YANG et al., 2009).
A hipótese de que o uso da variável independente DAP é suficiente para boas
estimativas em equações volumétricas locais foi rejeitada por Leite e Andrade (2003), os
quais concluíram que o uso da altura no modelo deve ser considerado para obter estimativas
precisas.

50
Santos et al. (2006) com objetivo de testar modelos matemáticos encontradas na
literatura florestal para estimar o volume individual de arvores no estado do Paraná, verificou
que todos os modelos, exceto o de Husch, que utiliza apenas diâmetro, sem considerar altura
do fuste, apresentam boa performance.
Tonini e Melo (2009) trabalhando com espécies nativas, selecionaram as seguintes
equações: Spurr logaritimizada para a castanha-do-brasil; Schumachar-Hall para o angelim-
ferro; Stoate para o Visgueiro.
Segundo Santos (2006), um modelo matemático só pode realmente ser indicado depois
de verificado à existência ou não de um comportamento tendencioso sobre toda a linha de
regressão, com os resíduos o mais uniformemente distribuído sobre a linha.
Os valores do erro padrão residual (Sxy), por expressar a medida de variação dos
volumes estimados das árvores caídas pelas equações, quando comparado ao volume real, são
considerados aceitáveis ao uso.
2.4.3 Validação da Equação Volumétrica
Há praticidade no uso das equações de simples entrada e/ou fator de forma para
obtenção do volume de espécies arbórea, pois fornecem estimativas precisas com menor custo
e praticidade. No entanto, de acordo com a Resolução CONAMA 406/09 é obrigatório, a
partir do segundo ano de exploração florestal, o cálculo do volume de árvores em pé mediante
equação de volume desenvolvida.
Em equações de simples entrada, mesma quando os critérios estatísticos são
obedecidos, podem ocorrer a sub ou superestimação dos volumes de árvores individuais ou
povoamentos. Estes resultados variam em função do tamanho das árvores. Em florestas
nativas, árvores de mesmo diâmetro não possuem necessariamente a mesma altura.
No entento, é cada vez mais comum na Amazônia brasileira a utilização de equações
de simples entrada, com a variável independente DAP, para estimativa do volume de florestas
a serem submetidas ao manejo florestal (FERREIRA et al., 2011; BARROS e SILVA Jr.,
2009; YANG et al., 2009).
As características do conjunto de dados volumétricos faz com que em diferentes sitios,
com características distintas de clima, solo e vegetação, proporcionem ajustes diferenciados
quanto aos modelos de regressão. Brandeis et al. (2006) trabalhado em florestas subtropicais
secas de Porto Rico e outras ilhas do Caribe verificaram que os modelos de Hohenald-Krenn e
Spurr obtiveram melhores ajustes para o volume do tronco das árvores.

51
2.5 CONCLUSÕES
As estruturas diamétrica e hipsométrica das árvores caídas da Floresta Estadual do
Amapá representam a estrutura das árvores vivas em “pé”, nas distintas fitofisionomias que a
compõem.
Assim, as equações volumétricas desenvolvidas neste trabalho estão habilitadas para
emprego da estimativa do volume comercial das áreas destinadas ao manejo florestal
sustentável.
As equações de simples e dupla entrada, bem como o fator de forma apresentaram
resultados confiáveis para estimativa volumétrica. No entanto, deve-se priorizar a utilização
das equações para predição do volume, por atender as indicações legais previstas na
legislação florestal brasileira.
Apesar das vantagens opeacionais observadas para equação volumétrica de Husch,
recomenda-se a utilização da equação volumétrica de Schumacher e Hall (Ln) levando em
consideração que as árvores de mesmo diâmetro da Floresta Estadual do Amapá não possuem
a mesma altura.

52
3 APTIDÃO DE ESPÉCIES ARBÓREAS AO MANEJO EM FLORESTA
OMBRÓFILA DENSA, AMAPÁ, AMAZÔNIA

53
RESUMO
O trabalho pretende verificar a viabilidade de execução de manejo florestal na Floresta
Estadual do Amapá, Amazônia, Brasil. O inventário foi realizado em três unidades primárias.
Cada unidade primária foi disposta em forma quadrada com cinco parcelas de 100 x 100 m. O
nível de inclusão foi de árvores com DAP (Diâmetro a altura do peito a 1,30 m do solo) > 10
cm. Em todos os indivíduos foram coletados nome popular, altura comercial, qualidade do
fuste e presença ou ausência de cipós, oco e cupim. As espécies foram separadas em grupos
de valor madeireiro (GVM). Foram calculados os parâmetros fitossociológicos, agregação,
estrutura vertical e diamétrica. Para cada classe diamétrica foi estimado o volume total. Foram
inventariados 7557 indivíduos, distribuídos em 38 famílias, 115 gêneros, 182 espécies, com
12 indeterminadas. O índice de Shannon foi de 4,38 nats/ind. Dentre as espécies com valor
comercial podem ser citadas: Tabebuia serratifolia (GVM 1), Carapa guianensis (GVM 2),
Dipteryx magnífica (GVM 3) e Aspidosperma excelsum (GVM 4). O número de árvores foi
estimado em 503,80 ± 25,51 ind.ha-1
, com volume médio de 383,22m3.ha
-1. O volume entre as
classes diamétricas evidenciou 67,33 m3.ha
-1 nas árvores com DAP’s entre 10 e 40 cm, 22,37
m3.ha
-1 (DAP’s entre 40 e 50 cm) e 293,53 m
3.ha
-1 (DAP’s maior 50 cm). A distribuição
diamétrica dos indivíduos evidenciou uma curva decrescente. Dessa forma, há indícios de
potencial volumétrico para fins de manejo, com estimativas por hectare superior a permitida
nas bases legais da política florestal brasileira.
Palavras-chave: Diversidade; Volume; Distribuição diamétrica.

54
ABSTRACT
The work aims to verify the feasibility of implementing forest management in Amapa State
Forest, Amazon, Brazil. The inventory was conducted in three primary units. Each primary
units was arranged in a square with five plots of 100 x 100 m. The inclusion level was trees
with DBH (diameter at breast height to 1.30m) > 10 cm. In all individuals were collected
popular name, commercial height, trunk quality and presence or absence of vines, hollow
termite. The species were separated into groups of timber value (GVM). Parameters
phytosociological, aggregation, vertical structure and diameter were calculated. For each
diameter class was estimated total volume. We recorded 7557 individuals belonging to 38
families, 115 genera, 182 species, with 12 indeterminate. The Shannon index was 4.38 nats /
ind. Among the species with commercial value can be cited: Tabebuia serratifolia (GVM 1)
Carapa guianensis (GVM 2), Dipteryx magnificent (GVM 3) and Aspidosperma excelsum
(GVM 4). The number of trees was estimated at 503.80 ± 25.51 ind.ha-1
, with an average
volume of 383.22 m3.ha
-1. The volume between the diameter classes showed 67.33 m
3.ha
-1 in
trees with DBH between 10 and 40 cm, 22.37 m3.ha
-1 (DBH between 40 and 50 cm) and
293.53 m3.ha
-1 (DAP's largest 50 cm). The diameter distribution showed a decreasing curve.
Thus, there is evidence of potential volume for management purposes, with estimates per
hectare higher than permitted in legal bases Brazilian forest policy.
Key words: Diversity, Volume, Distribution diameter.

55
3.1 INTRODUÇÃO
O registro de inúmeras espécies da flora com capacidade de fornecer matéria-prima
para a construção civil, cosméticos, farmacêuticos, etc. tem contribuído para destacar a
Amazônia no cenário internacional. O código florestal brasileiro (Lei 12.651/2012) prevê a
utilização destes recursos a partir da elaboração de “Plano de Manejo Florestal ustentável de
uso múltiplo - PMF ”, com discriminação dos procedimentos técnicos em normas específicas.
Atualmente, os procedimentos e especificações técnicas para o acesso aos recursos
provenientes de florestas primitivas da Amazônia são orientados pelas Instruções Normativas
04 e 05/2006 MMA e Resolução CONAMA 406/2009. Tradicionalmente são consideradas
com aptidão ao manejo florestal, espécies com número de indivíduos superior a três a cada
100 hectares, diâmetro mínimo de corte (50 cm para todas as espécies), fuste cilíndrico e com
ausência de oco.
No entanto, há generalidade da legislação quanto às espécies e intensidade de
exploração, permitindo a máxima retirada de madeira por unidade de área, com danos
irreversíveis à floresta remanescente (ROCHA et al., 2000; PINTO et al., 2002).As diretrizes
estabelecidas nas normativas brasileiras atende apenas as florestas como comunidade, não
atendendo especificidades de populações de espécies arbóreas com distintos comportamentos
ecológicos.
Para dar inicio as discussões sobre procedimentos técnicos considerando a
peculiaridade de cada espécie, é necessário identificar as relações fitossociológicas,
distribuição diamétrica, vertical e espacial para fundamentar a adoção de planejamentos mais
adequados de exploração.
Nos últimos dez anos, diversos autores (JARDIM e SOARES, 2010; SAMPAIO et al.,
2005; PROCÓPIO e SECCO, 2008; HIRAI et al., 2008)têm estudado o comportamento
individual de espécies arbóreas comerciais da Amazônia, como Sterculia pruriens (Aublet)
Schumann, Manilkara huberi Standley, Aniba rosaeodora Ducke e Couratari guianensis
Aublet descrevendo o número de árvores susceptíveis ao manejo com distintas estruturas na
floresta.
Diante do exposto, o trabalho pretende verificar a viabilidade de execução de manejo
florestal em floresta ombrófila densa na Amazônia, avaliando o comportamento individual e
da comunidade. Este trabalho pretende responder se existem espécies com estruturas distintas
capazes de serem submetidas ao manejo florestal madeireiro.

56
3.2 MATERIAL E MÉTODOS
3.2.1 Caracterização da Área
A área de estudo está localizada no módulo II da Floresta Estadual do Amapá
(FLOTA/AP), região central do estado do Amapá, Amazônia, Brasil. O universo amostral
abrangeu uma população de 500 ha, com vegetação descontínua de floresta ombrófila densa
de terras baixas (IBGE, 2012).
O solo predominante é tipo Latossolo Vermelho-Amarelo distrófico (RADAM, 1974).
A altitude varia de 60 a 100 m. O clima da região é do tipo Am (equatorial super-úmido),
segundo a classificação de Köppen.
3.2.2 Inventário Florestal
O inventário foi realizado em unidades amostrais conglomeradas, distribuídas em três
distintas áreas (unidades primárias - UP), equidistantes em aproximadamente 2.500 m. Cada
unidade primária foi disposta em forma quadrada com cinco parcelas permanentes (unidades
secundárias) de 100 x 100 m, abrangendo uma área de 5 ha por UP.
As parcelas foram estabelecidas de acordo com os critérios da “Rede de
Monitoramento da Dinâmica das Florestas da Amazônia”, com nível de inclusão das árvores
abrangendo DAP > 10 cm (SILVA et al., 2005).
Todos os indivíduos foram etiquetados com numeração progressiva e coletados nome
popular, DAP, altura comercial (h) (até a inserção do primeiro galho com auxílio de
hipsômetro TRIPULSE 360º) e presença ou ausência de cipós competidores (C), oco (O) e
cupim (PC).
Os fustes das árvores foram classificados quanto a qualidade do fuste (Qf)
(VACCARO et al., 2003), onde: (1) reto, (2) levemente tortuoso e (3) tortuoso. A classe
"reto" abrangeu os fustes que apresentam qualidade entre 80 e 100 % de seu comprimento; a
classe "levemente tortuoso" entre 50 e 80 % e a classe "tortuoso" menos que 50 %.
A identificação taxonômica das espécies foi realizada com levantamento prévio em
campo e os nomes botânicos seguiram o sistema APG III (2010).Todas as espécies
identificadas foram separadas em grupos de valor madeireiro (GVM), onde: 0 – menor valor
comercial; 1 – alto valor comercial; 2 – reconhecido valor comercial; 3 – comumente
comercializadas; 4 – comercialização local; 5 – não classificadas.

57
3.2.3 Análise Estrutural
A suficiência amostral florística foi determinada a partir da confecção da curva do
coletor. A representatividade do número de indivíduos e volume foi efetuada com limite de
erro em torno da média de até 10 %, para um nível de probabilidade de 95 % (P = 0,95),
precisão recomendada na IN 05/2006 IBAMA.
Com base nas informações coletadas foram calculados os parâmetros fitossociológicos
utilizando o software MATA NATIVA 2.O padrão de agregação das espécies foi definido
com auxílio do índice de Payandeh (Pi).
A estrutura vertical foi estimada a partir de três estratos: estrato inferior -árvores com
altura comercial inferior a média aritmética menos um desvio padrão; estrato médio - árvores
cujas alturas se inserem nos limites inferior e superior da média aritmética e desvio padrão; e
estrato superior - árvores com altura superior a média das alturas mais um desvio
padrão(SCOLFORO, 1998).
Também foram gerados histogramas da estrutura diamétrica por qualidade do fuste. O
número de classes diamétricas foi calculado segundo Higuchi et al. (2008).Para cada classe
diamétrica foi estimado o volume total, utilizando a equação de Schumacher e Hall (Ln):
Onde:
V = volume (m³)
DAP = diâmetro a altura do peito a 1,30m do solo (cm)
Hc = altura comercial (m)
3.3 RESULTADOS
3.3.1 Florística e Fitossociologia
A composição florística foi representada com sucesso a partir da décima quarta parcela,
quando se iniciou a estabilidade linear da curva de suficiência amostral (Figura 6). A
vegetação representada foi composta por 7.557 indivíduos arbóreos vivos, agrupados em 38
famílias botânicas, 115 gêneros, 182 espécies, com 12 indeterminadas.
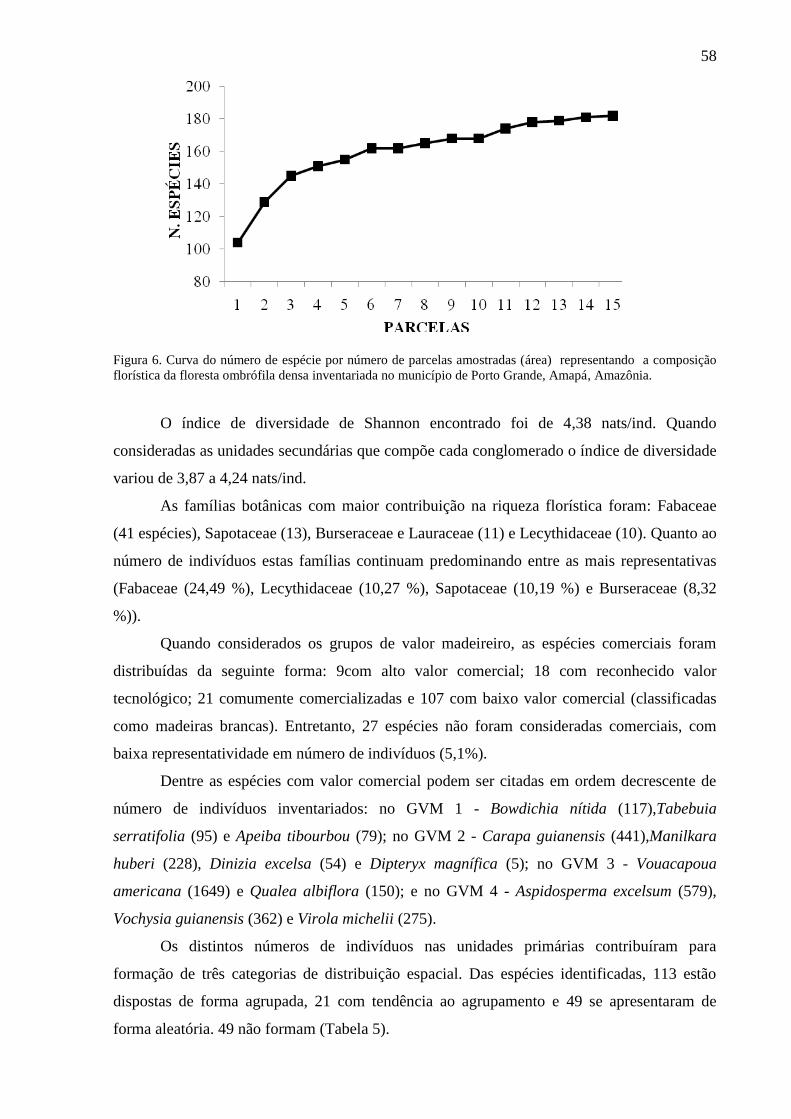
58
Figura 6. Curva do número de espécie por número de parcelas amostradas (área) representando a composição
florística da floresta ombrófila densa inventariada no município de Porto Grande, Amapá, Amazônia.
O índice de diversidade de Shannon encontrado foi de 4,38 nats/ind. Quando
consideradas as unidades secundárias que compõe cada conglomerado o índice de diversidade
variou de 3,87 a 4,24 nats/ind.
As famílias botânicas com maior contribuição na riqueza florística foram: Fabaceae
(41 espécies), Sapotaceae (13), Burseraceae e Lauraceae (11) e Lecythidaceae (10). Quanto ao
número de indivíduos estas famílias continuam predominando entre as mais representativas
(Fabaceae (24,49 %), Lecythidaceae (10,27 %), Sapotaceae (10,19 %) e Burseraceae (8,32
%)).
Quando considerados os grupos de valor madeireiro, as espécies comerciais foram
distribuídas da seguinte forma: 9com alto valor comercial; 18 com reconhecido valor
tecnológico; 21 comumente comercializadas e 107 com baixo valor comercial (classificadas
como madeiras brancas). Entretanto, 27 espécies não foram consideradas comerciais, com
baixa representatividade em número de indivíduos (5,1%).
Dentre as espécies com valor comercial podem ser citadas em ordem decrescente de
número de indivíduos inventariados: no GVM 1 - Bowdichia nítida (117),Tabebuia
serratifolia (95) e Apeiba tibourbou (79); no GVM 2 - Carapa guianensis (441),Manilkara
huberi (228), Dinizia excelsa (54) e Dipteryx magnífica (5); no GVM 3 - Vouacapoua
americana (1649) e Qualea albiflora (150); e no GVM 4 - Aspidosperma excelsum (579),
Vochysia guianensis (362) e Virola michelii (275).
Os distintos números de indivíduos nas unidades primárias contribuíram para
formação de três categorias de distribuição espacial. Das espécies identificadas, 113 estão
dispostas de forma agrupada, 21 com tendência ao agrupamento e 49 se apresentaram de
forma aleatória. 49 não formam (Tabela 5).
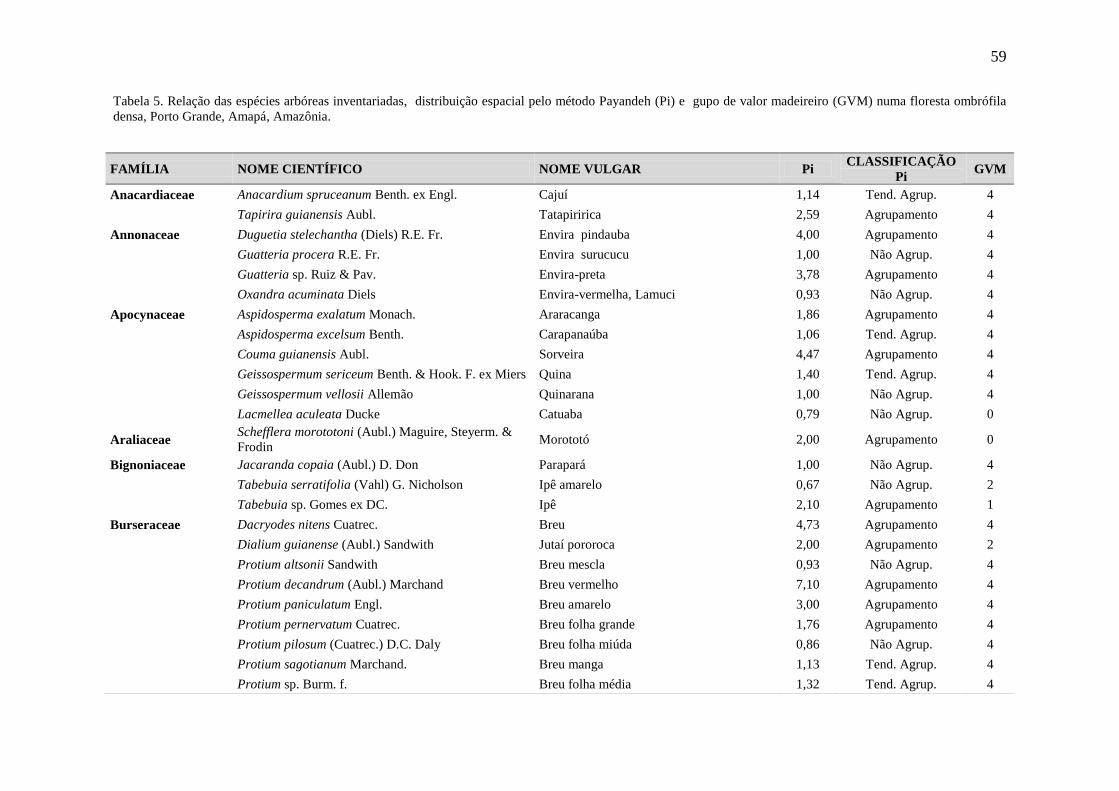
59
Tabela 5. Relação das espécies arbóreas inventariadas, distribuição espacial pelo método Payandeh (Pi) e gupo de valor madeireiro (GVM) numa floresta ombrófila
densa, Porto Grande, Amapá, Amazônia.
FAMÍLIA NOME CIENTÍFICO NOME VULGAR Pi CLASSIFICAÇÃO
Pi GVM
Anacardiaceae Anacardium spruceanum Benth. ex Engl. Cajuí 1,14 Tend. Agrup. 4
Tapirira guianensis Aubl. Tatapiririca 2,59 Agrupamento 4
Annonaceae Duguetia stelechantha (Diels) R.E. Fr. Envira pindauba 4,00 Agrupamento 4
Guatteria procera R.E. Fr. Envira surucucu 1,00 Não Agrup. 4
Guatteria sp. Ruiz & Pav. Envira-preta 3,78 Agrupamento 4
Oxandra acuminata Diels Envira-vermelha, Lamuci 0,93 Não Agrup. 4
Apocynaceae Aspidosperma exalatum Monach. Araracanga 1,86 Agrupamento 4
Aspidosperma excelsum Benth. Carapanaúba 1,06 Tend. Agrup. 4
Couma guianensis Aubl. Sorveira 4,47 Agrupamento 4
Geissospermum sericeum Benth. & Hook. F. ex Miers Quina 1,40 Tend. Agrup. 4
Geissospermum vellosii Allemão Quinarana 1,00 Não Agrup. 4
Lacmellea aculeata Ducke Catuaba 0,79 Não Agrup. 0
Araliaceae Schefflera morototoni (Aubl.) Maguire, Steyerm. &
Frodin Morototó 2,00 Agrupamento 0
Bignoniaceae Jacaranda copaia (Aubl.) D. Don Parapará 1,00 Não Agrup. 4
Tabebuia serratifolia (Vahl) G. Nicholson Ipê amarelo 0,67 Não Agrup. 2
Tabebuia sp. Gomes ex DC. Ipê 2,10 Agrupamento 1
Burseraceae Dacryodes nitens Cuatrec. Breu 4,73 Agrupamento 4
Dialium guianense (Aubl.) Sandwith Jutaí pororoca 2,00 Agrupamento 2
Protium altsonii Sandwith Breu mescla 0,93 Não Agrup. 4
Protium decandrum (Aubl.) Marchand Breu vermelho 7,10 Agrupamento 4
Protium paniculatum Engl. Breu amarelo 3,00 Agrupamento 4
Protium pernervatum Cuatrec. Breu folha grande 1,76 Agrupamento 4
Protium pilosum (Cuatrec.) D.C. Daly Breu folha miúda 0,86 Não Agrup. 4
Protium sagotianum Marchand. Breu manga 1,13 Tend. Agrup. 4
Protium sp. Burm. f. Breu folha média 1,32 Tend. Agrup. 4
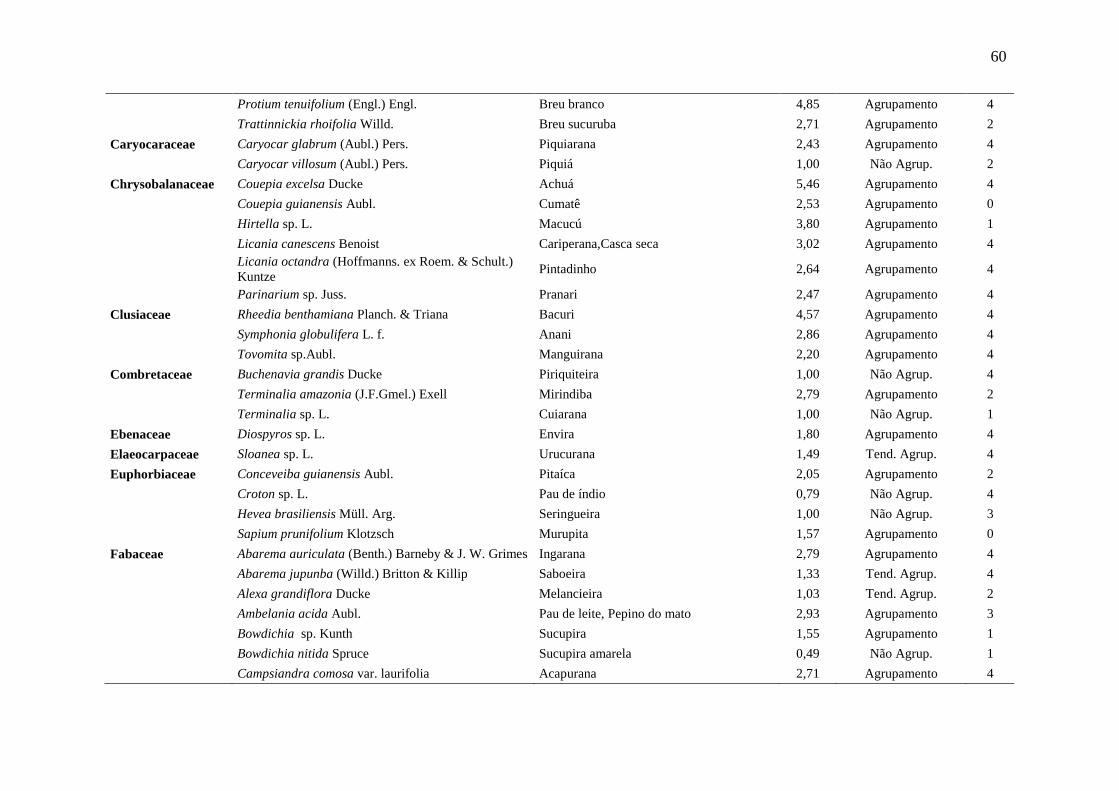
60
Protium tenuifolium (Engl.) Engl. Breu branco 4,85 Agrupamento 4
Trattinnickia rhoifolia Willd. Breu sucuruba 2,71 Agrupamento 2
Caryocaraceae Caryocar glabrum (Aubl.) Pers. Piquiarana 2,43 Agrupamento 4
Caryocar villosum (Aubl.) Pers. Piquiá 1,00 Não Agrup. 2
Chrysobalanaceae Couepia excelsa Ducke Achuá 5,46 Agrupamento 4
Couepia guianensis Aubl. Cumatê 2,53 Agrupamento 0
Hirtella sp. L. Macucú 3,80 Agrupamento 1
Licania canescens Benoist Cariperana,Casca seca 3,02 Agrupamento 4
Licania octandra (Hoffmanns. ex Roem. & Schult.)
Kuntze Pintadinho 2,64 Agrupamento 4
Parinarium sp. Juss. Pranari 2,47 Agrupamento 4
Clusiaceae Rheedia benthamiana Planch. & Triana Bacuri 4,57 Agrupamento 4
Symphonia globulifera L. f. Anani 2,86 Agrupamento 4
Tovomita sp.Aubl. Manguirana 2,20 Agrupamento 4
Combretaceae Buchenavia grandis Ducke Piriquiteira 1,00 Não Agrup. 4
Terminalia amazonia (J.F.Gmel.) Exell Mirindiba 2,79 Agrupamento 2
Terminalia sp. L. Cuiarana 1,00 Não Agrup. 1
Ebenaceae Diospyros sp. L. Envira 1,80 Agrupamento 4
Elaeocarpaceae Sloanea sp. L. Urucurana 1,49 Tend. Agrup. 4
Euphorbiaceae Conceveiba guianensis Aubl. Pitaíca 2,05 Agrupamento 2
Croton sp. L. Pau de índio 0,79 Não Agrup. 4
Hevea brasiliensis Müll. Arg. Seringueira 1,00 Não Agrup. 3
Sapium prunifolium Klotzsch Murupita 1,57 Agrupamento 0
Fabaceae Abarema auriculata (Benth.) Barneby & J. W. Grimes Ingarana 2,79 Agrupamento 4
Abarema jupunba (Willd.) Britton & Killip Saboeira 1,33 Tend. Agrup. 4
Alexa grandiflora Ducke Melancieira 1,03 Tend. Agrup. 2
Ambelania acida Aubl. Pau de leite, Pepino do mato 2,93 Agrupamento 3
Bowdichia sp. Kunth Sucupira 1,55 Agrupamento 1
Bowdichia nitida Spruce Sucupira amarela 0,49 Não Agrup. 1
Campsiandra comosa var. laurifolia Acapurana 2,71 Agrupamento 4

61
Candolleodendron sp. R.S. Cowan Fava 2,49 Agrupamento 4
Copaifera multijuga Hayne Copaíba 0,93 Não Agrup. 4
Dimorphandra multiflora Ducke Fava amarela 1,00 Não Agrup. 3
Dinizia excelsa Ducke Angelim vermelho 1,57 Agrupamento 2
Diplotropis purpurea (Rich.) Amshoff. Sucupira preta 3,77 Agrupamento 1
Dipteryx magnifica Ducke Cumaru rosa 1,00 Não Agrup. 3
Dipteryx odorata (Aubl.) Willd. Cumaru 2,16 Agrupamento 3
Dussia discolor (Benth.) Amshoff Mututirana 2,88 Agrupamento 0
Elizabetha bicolor Ducke Boa macaca 11,49 Agrupamento 4
Eperua schomburgkiana Benth. Apá 5,71 Agrupamento 4
Eperua sp. Aubl. Iperana 1,36 Tend. Agrup. 1
Hymenaea courbaril L. Jatobá 1,14 Tend. Agrup. 3
Hymenaea parvifolia Huber Jutaí 1,49 Tend. Agrup. 4
Hymenolobium petraeum Ducke Angelim pedra 0,86 Não Agrup. 2
Inga auristellae Harms Ingá vermelho 6,64 Agrupamento 4
Inga gracilifolia Ducke Ingá folha miúda 1,57 Agrupamento 4
Inga paraensis Ducke Ingá xixica 0,86 Não Agrup. 4
Inga sp. 1 Mill. Ingá 4,71 Agrupamento 4
Inga sp. 2 Mill. Ingá branco 1,67 Agrupamento 4
Parkia gigantocarpa Ducke Fava esponja, Atanã 1,57 Agrupamento 4
Peltogyne paniculata Benth. Roxinho 0,93 Não Agrup. 3
Peltogyne paradoxa Ducke Coataquiçaua 1,57 Agrupamento 4
Pentaclethra macroloba (Willd.) Kuntze Pracaxi 18,24 Agrupamento 4
Piptadenia gonoacantha (Mart.) J.F.Macbr. Timborana 1,46 Tend. Agrup. 4
Pterocarpus rohrii Vahl Mututi 1,00 Não Agrup. 4
Schizolobium amazonicum Huber ex Ducke Paricá da mata 1,00 Não Agrup. 0
Sclerolobium melanocarpum Ducke Taxi vermelho 4,86 Agrupamento 4
Stryphnodendron pulcherrimum (Willd.) Hochr. Fava folha fina 2,88 Agrupamento 4
Swartzia racemosa Benth. Pacapeuá 0,93 Não Agrup. 0
Swartzia sp. Schreb. Tento 4,00 Agrupamento 3
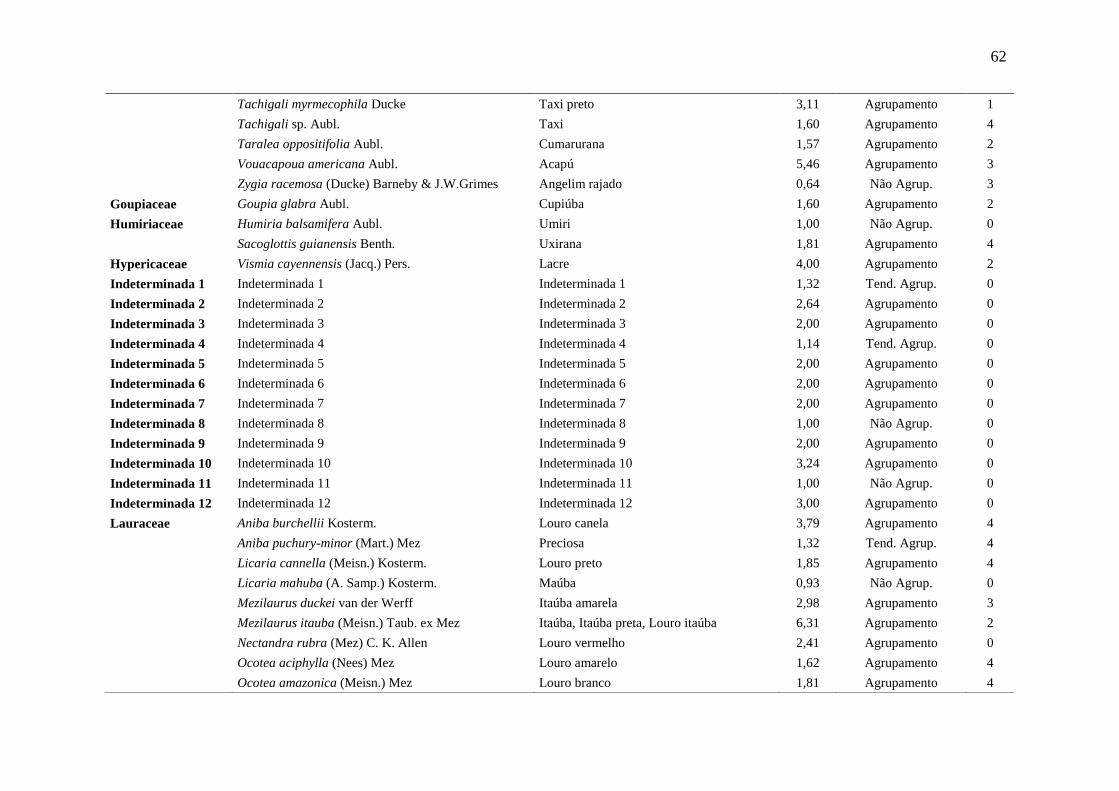
62
Tachigali myrmecophila Ducke Taxi preto 3,11 Agrupamento 1
Tachigali sp. Aubl. Taxi 1,60 Agrupamento 4
Taralea oppositifolia Aubl. Cumarurana 1,57 Agrupamento 2
Vouacapoua americana Aubl. Acapú 5,46 Agrupamento 3
Zygia racemosa (Ducke) Barneby & J.W.Grimes Angelim rajado 0,64 Não Agrup. 3
Goupiaceae Goupia glabra Aubl. Cupiúba 1,60 Agrupamento 2
Humiriaceae Humiria balsamifera Aubl. Umiri 1,00 Não Agrup. 0
Sacoglottis guianensis Benth. Uxirana 1,81 Agrupamento 4
Hypericaceae Vismia cayennensis (Jacq.) Pers. Lacre 4,00 Agrupamento 2
Indeterminada 1 Indeterminada 1 Indeterminada 1 1,32 Tend. Agrup. 0
Indeterminada 2 Indeterminada 2 Indeterminada 2 2,64 Agrupamento 0
Indeterminada 3 Indeterminada 3 Indeterminada 3 2,00 Agrupamento 0
Indeterminada 4 Indeterminada 4 Indeterminada 4 1,14 Tend. Agrup. 0
Indeterminada 5 Indeterminada 5 Indeterminada 5 2,00 Agrupamento 0
Indeterminada 6 Indeterminada 6 Indeterminada 6 2,00 Agrupamento 0
Indeterminada 7 Indeterminada 7 Indeterminada 7 2,00 Agrupamento 0
Indeterminada 8 Indeterminada 8 Indeterminada 8 1,00 Não Agrup. 0
Indeterminada 9 Indeterminada 9 Indeterminada 9 2,00 Agrupamento 0
Indeterminada 10 Indeterminada 10 Indeterminada 10 3,24 Agrupamento 0
Indeterminada 11 Indeterminada 11 Indeterminada 11 1,00 Não Agrup. 0
Indeterminada 12 Indeterminada 12 Indeterminada 12 3,00 Agrupamento 0
Lauraceae Aniba burchellii Kosterm. Louro canela 3,79 Agrupamento 4
Aniba puchury-minor (Mart.) Mez Preciosa 1,32 Tend. Agrup. 4
Licaria cannella (Meisn.) Kosterm. Louro preto 1,85 Agrupamento 4
Licaria mahuba (A. Samp.) Kosterm. Maúba 0,93 Não Agrup. 0
Mezilaurus duckei van der Werff Itaúba amarela 2,98 Agrupamento 3
Mezilaurus itauba (Meisn.) Taub. ex Mez Itaúba, Itaúba preta, Louro itaúba 6,31 Agrupamento 2
Nectandra rubra (Mez) C. K. Allen Louro vermelho 2,41 Agrupamento 0
Ocotea aciphylla (Nees) Mez Louro amarelo 1,62 Agrupamento 4
Ocotea amazonica (Meisn.) Mez Louro branco 1,81 Agrupamento 4
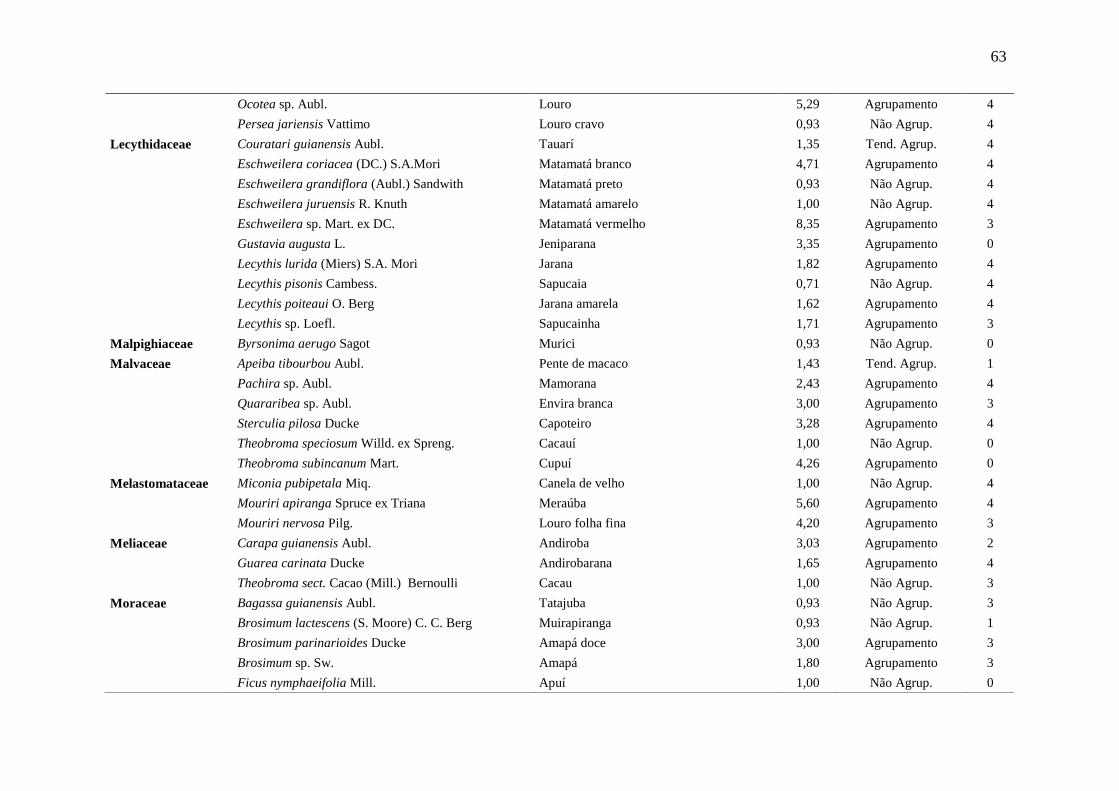
63
Ocotea sp. Aubl. Louro 5,29 Agrupamento 4
Persea jariensis Vattimo Louro cravo 0,93 Não Agrup. 4
Lecythidaceae Couratari guianensis Aubl. Tauarí 1,35 Tend. Agrup. 4
Eschweilera coriacea (DC.) S.A.Mori Matamatá branco 4,71 Agrupamento 4
Eschweilera grandiflora (Aubl.) Sandwith Matamatá preto 0,93 Não Agrup. 4
Eschweilera juruensis R. Knuth Matamatá amarelo 1,00 Não Agrup. 4
Eschweilera sp. Mart. ex DC. Matamatá vermelho 8,35 Agrupamento 3
Gustavia augusta L. Jeniparana 3,35 Agrupamento 0
Lecythis lurida (Miers) S.A. Mori Jarana 1,82 Agrupamento 4
Lecythis pisonis Cambess. Sapucaia 0,71 Não Agrup. 4
Lecythis poiteaui O. Berg Jarana amarela 1,62 Agrupamento 4
Lecythis sp. Loefl. Sapucainha 1,71 Agrupamento 3
Malpighiaceae Byrsonima aerugo Sagot Murici 0,93 Não Agrup. 0
Malvaceae Apeiba tibourbou Aubl. Pente de macaco 1,43 Tend. Agrup. 1
Pachira sp. Aubl. Mamorana 2,43 Agrupamento 4
Quararibea sp. Aubl. Envira branca 3,00 Agrupamento 3
Sterculia pilosa Ducke Capoteiro 3,28 Agrupamento 4
Theobroma speciosum Willd. ex Spreng. Cacauí 1,00 Não Agrup. 0
Theobroma subincanum Mart. Cupuí 4,26 Agrupamento 0
Melastomataceae Miconia pubipetala Miq. Canela de velho 1,00 Não Agrup. 4
Mouriri apiranga Spruce ex Triana Meraúba 5,60 Agrupamento 4
Mouriri nervosa Pilg. Louro folha fina 4,20 Agrupamento 3
Meliaceae Carapa guianensis Aubl. Andiroba 3,03 Agrupamento 2
Guarea carinata Ducke Andirobarana 1,65 Agrupamento 4
Theobroma sect. Cacao (Mill.) Bernoulli Cacau 1,00 Não Agrup. 3
Moraceae Bagassa guianensis Aubl. Tatajuba 0,93 Não Agrup. 3
Brosimum lactescens (S. Moore) C. C. Berg Muirapiranga 0,93 Não Agrup. 1
Brosimum parinarioides Ducke Amapá doce 3,00 Agrupamento 3
Brosimum sp. Sw. Amapá 1,80 Agrupamento 3
Ficus nymphaeifolia Mill. Apuí 1,00 Não Agrup. 0
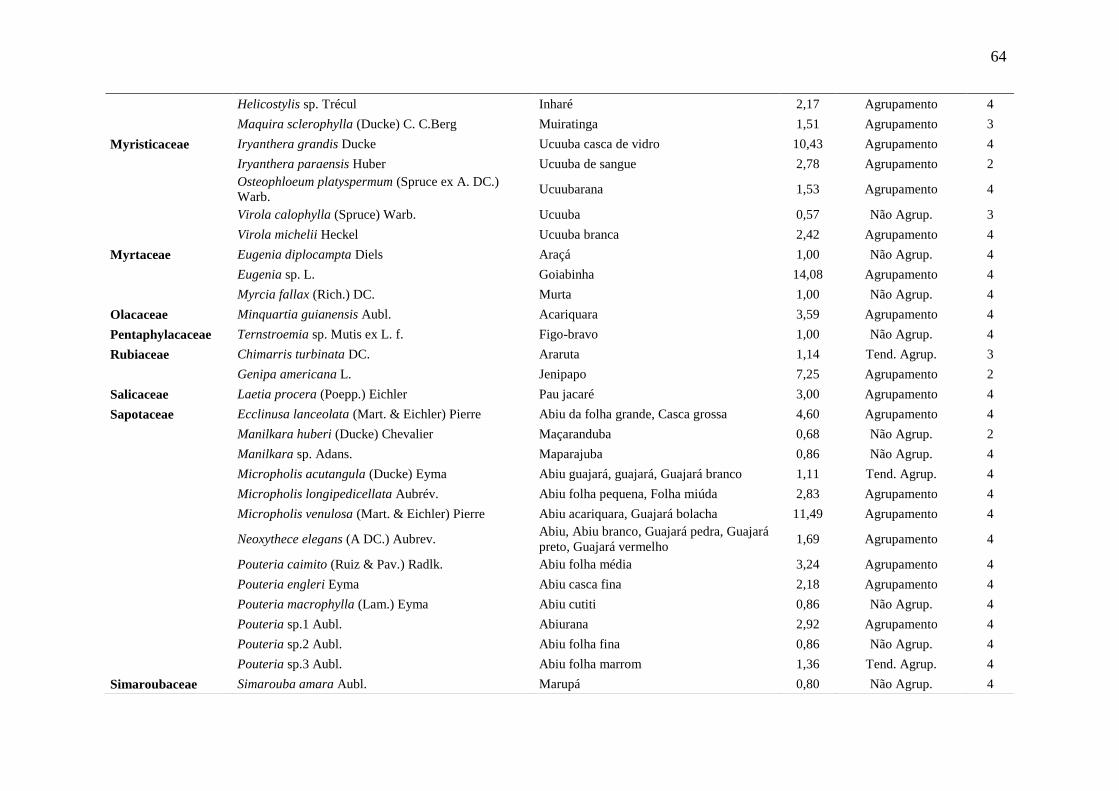
64
Helicostylis sp. Trécul Inharé 2,17 Agrupamento 4
Maquira sclerophylla (Ducke) C. C.Berg Muiratinga 1,51 Agrupamento 3
Myristicaceae Iryanthera grandis Ducke Ucuuba casca de vidro 10,43 Agrupamento 4
Iryanthera paraensis Huber Ucuuba de sangue 2,78 Agrupamento 2
Osteophloeum platyspermum (Spruce ex A. DC.)
Warb. Ucuubarana 1,53 Agrupamento 4
Virola calophylla (Spruce) Warb. Ucuuba 0,57 Não Agrup. 3
Virola michelii Heckel Ucuuba branca 2,42 Agrupamento 4
Myrtaceae Eugenia diplocampta Diels Araçá 1,00 Não Agrup. 4
Eugenia sp. L. Goiabinha 14,08 Agrupamento 4
Myrcia fallax (Rich.) DC. Murta 1,00 Não Agrup. 4
Olacaceae Minquartia guianensis Aubl. Acariquara 3,59 Agrupamento 4
Pentaphylacaceae Ternstroemia sp. Mutis ex L. f. Figo-bravo 1,00 Não Agrup. 4
Rubiaceae Chimarris turbinata DC. Araruta 1,14 Tend. Agrup. 3
Genipa americana L. Jenipapo 7,25 Agrupamento 2
Salicaceae Laetia procera (Poepp.) Eichler Pau jacaré 3,00 Agrupamento 4
Sapotaceae Ecclinusa lanceolata (Mart. & Eichler) Pierre Abiu da folha grande, Casca grossa 4,60 Agrupamento 4
Manilkara huberi (Ducke) Chevalier Maçaranduba 0,68 Não Agrup. 2
Manilkara sp. Adans. Maparajuba 0,86 Não Agrup. 4
Micropholis acutangula (Ducke) Eyma Abiu guajará, guajará, Guajará branco 1,11 Tend. Agrup. 4
Micropholis longipedicellata Aubrév. Abiu folha pequena, Folha miúda 2,83 Agrupamento 4
Micropholis venulosa (Mart. & Eichler) Pierre Abiu acariquara, Guajará bolacha 11,49 Agrupamento 4
Neoxythece elegans (A DC.) Aubrev.
Abiu, Abiu branco, Guajará pedra, Guajará
preto, Guajará vermelho 1,69 Agrupamento 4
Pouteria caimito (Ruiz & Pav.) Radlk. Abiu folha média 3,24 Agrupamento 4
Pouteria engleri Eyma Abiu casca fina 2,18 Agrupamento 4
Pouteria macrophylla (Lam.) Eyma Abiu cutiti 0,86 Não Agrup. 4
Pouteria sp.1 Aubl. Abiurana 2,92 Agrupamento 4
Pouteria sp.2 Aubl. Abiu folha fina 0,86 Não Agrup. 4
Pouteria sp.3 Aubl. Abiu folha marrom 1,36 Tend. Agrup. 4
Simaroubaceae Simarouba amara Aubl. Marupá 0,80 Não Agrup. 4

65
Siparunaceae Siparuna sp. Aubl. Louro folha grande 1,62 Agrupamento 4
Urticaceae Cecropia sciadophylla Mart. Embaúba torem 0,93 Não Agrup. 4
Pourouma heterophylla Mart. ex Miq. Embaúba pata de anta 8,80 Agrupamento 4
Pourouma minor Benoist Embaúba branca 2,55 Agrupamento 4
Pourouma ovata Trécul Embaúba 12,30 Agrupamento 4
Pourouma villosa Trécul Embaúba vick 1,57 Agrupamento 4
Vochysiaceae Qualea albiflora Varm. Mandioqueira 2,67 Agrupamento 3
Vochysia guianensis Aubl. Quaruba branca 21,40 Agrupamento 4
Vochysia maxima Ducke Quaruba vermelha 1,45 Tend. Agrup. 4
Vochysia sp. Aubl. Quaruba 5,29 Agrupamento 4

66
O maior valor de frequência relativa (FR) foi partilhado por 28 espécies
(1,09 %), as quais podemos citar Ocotea amazonica, Bowdichia sp. E Vouacapoua
americana, com número de indivíduos variando de 51 (Bowdichia sp.) a 361
(Eschweilera coriacea). A frequência nas unidades secundárias variou de 107 a 80
espécies, com 29 espécies comuns as unidades primárias, 30 presente apenas na UP1,
seguida de 5 e 14 espécies exclusivas das demais UP’s. Dentre as espécies com menor
frequência relativa (0,07 %), foi observado que estão inseridas oito das doze
indeterminadas.
As espécies com maior densidade relativa (DR) foram: Eschweilera coriacea
(4,78 %), Inga auristellae (4,61 %) e Iryanthera paraensis (4,02 %). Também foi
observado que apenas 32 espécies apresentam percentuais de DR acima de 1 %.
Entretanto, 72 espécies apresentaram percentuais abaixo de 0,1 %, entre elas estão dez
das indeterminadas, bem como Hevea brasiliensis, Hymenaea parvifolia e Terminalia
amazonia.
Maiores percentuais de dominância relativa (DoR) foram obtidas para
Vouacapoua americana com 6,45 %, seguida da Eschweilera coriacea (3,81 %) e Inga
auristellae (3,38 ). Quando consideradas as UP’s, a variação de dominância ocorreu
de 0,0005 a 2,1452 m²ha-1
, com representantes florísticos predominantes as espécies
Inga auristellae Harms eEschweilera coriacea (DC.) S.A.Mori (UP1); Inga auristellae
Harms e Protium tenuifolium (Engl.) Engl. (UP2); Eschweilera coriacea (DC.)
S.A.Mori e Eugeniasp. L. (UP3).
Quando consideradas em conjunto as contribuições dos parâmetros
fitossociológicos, verificou-se que a amplitude do Valor de Importância (VI) apresentou
baixa variabilidade na floresta. Tal fato gerou uma variação de 3,34 a 0,03 % (Tabela
5).

67
Tabela 6. Parâmetros Fitossociológicos de espécies arbóreas inventariadas numa floresta ombrófila densa,
Porto Grande, Amapá, Amazônia. Onde: N – numero total de indivíduos; DR – densidade relativa; FR –
freqüência relativa; DoR – dominância relativa; VI – valor de importância; H – altura comercial (metros);
DAP – diâmetro a altura do peito a 1.30m do solo (metros).
NOME CIENTÍFICO N DR FR DoR VI Hcmax Hcmin DAPmax DAPmin
Vouacapoua americana 193 2,55 1,02 6,45 3,34 30,5 4,3 1,02 0,1
Eschweilera coriacea 361 4,78 1,09 3,81 3,22 26,4 2,2 1,08 0,1
Inga auristellae 348 4,61 1,09 3,38 3,02 23 1,9 0,77 0,1
Eschweilera sp. 275 3,64 1,09 2,75 2,49 26,8 1,5 0,9 0,1
Pouteria sp.1 225 2,98 1,09 3,38 2,48 25,7 2,7 0,95 0,1
Iryanthera paraensis 304 4,02 1,09 1,91 2,34 25 3,2 0,68 0,1
Protium tenuifolium 269 3,56 1,09 2,3 2,31 25,3 2,2 0,95 0,1
Protium decandrum 195 2,58 1,09 1,54 1,74 39,7 2 0,76 0,1
Pourouma ovata 191 2,53 0,94 1,43 1,63 24,1 4 0,62 0,1
Sclerolobium
melanocarpum 148 1,96 1,09 1,71 1,59 32,6 3,8 1,28 0,1
Theobroma subincanum 182 2,41 1,09 0,93 1,48 21,1 1,6 0,42 0,1
Nectandra rubra 57 0,75 0,87 2,8 1,47 26,4 5,1 1,68 0,1
Micropholis venulosa 82 1,09 0,94 2,35 1,46 25,3 5,8 1,02 0,1
Sterculia pilosa 116 1,54 1,09 1,71 1,45 28,2 3,1 0,89 0,1
Micropholis acutangula 120 1,59 1,09 1,64 1,44 23,6 3,3 0,95 0,1
Osteophloeum
platyspermum 89 1,18 1,09 1,69 1,32 24,7 3,2 0,87 0,1
Tachigali myrmecophila 82 1,09 1,09 1,61 1,26 28 4,5 1,09 0,1
Eugenia sp. 160 2,12 0,94 0,7 1,25 19,3 2 0,46 0,1
Maquira sclerophylla 135 1,79 1,09 0,85 1,24 20 2,6 0,48 0,1
Dacryodes nitens 111 1,47 1,02 1,22 1,23 21,8 3 0,83 0,1
Dipteryx odorata 33 0,44 0,87 2,38 1,23 25 7,5 1,27 0,12
Inga sp. 2 141 1,87 1,09 0,72 1,23 22,4 1,4 0,39 0,1
Eperua schomburgkiana 69 0,91 0,94 1,79 1,22 26,5 3,6 0,97 0,11
Mouriri apiranga 121 1,6 1,09 0,93 1,21 26,1 2,6 0,65 0,1
Ocotea sp.1 96 1,27 1,09 1,26 1,21 29,9 4,3 1,27 0,1
Aspidosperma excelsum 80 1,06 1,09 1,43 1,19 26,4 3 0,94 0,1
Iryanthera grandis 115 1,52 1,09 0,81 1,14 25 2,6 0,67 0,1
Mezilaurus itauba 107 1,42 1,02 0,92 1,12 23,8 4,6 1,24 0,1
Couratari guianensis 34 0,45 0,8 2,09 1,11 33,2 6,6 1,59 0,11
Inga sp. 1 104 1,38 1,02 0,95 1,11 22,3 4,3 1,15 0,1
Couepia excelsa 100 1,32 1,02 0,89 1,07 20,2 5,2 0,55 0,1
Goupia glabra 26 0,34 0,87 2,01 1,07 25,1 5,9 1,34 0,11
Conceveiba guianensis 61 0,81 1,02 1,38 1,07 20,6 3 0,95 0,11
Ecclinusa lanceolata 81 1,07 1,09 0,96 1,04 23,8 3,6 0,92 0,1
Vochysia guianensis 75 0,99 0,8 1,19 0,99 32,6 2,9 1,26 0,1
Hirtella sp. 104 1,38 1,09 0,51 0,99 25,4 2,5 0,48 0,1
Licania canescens 73 0,97 0,94 1,05 0,99 21,7 5 0,58 0,1
Alexa grandiflora 86 1,14 1,09 0,7 0,98 25,3 1,5 0,68 0,1
Guatteria sp. 91 1,2 1,09 0,56 0,95 25,9 3,7 0,54 0,1
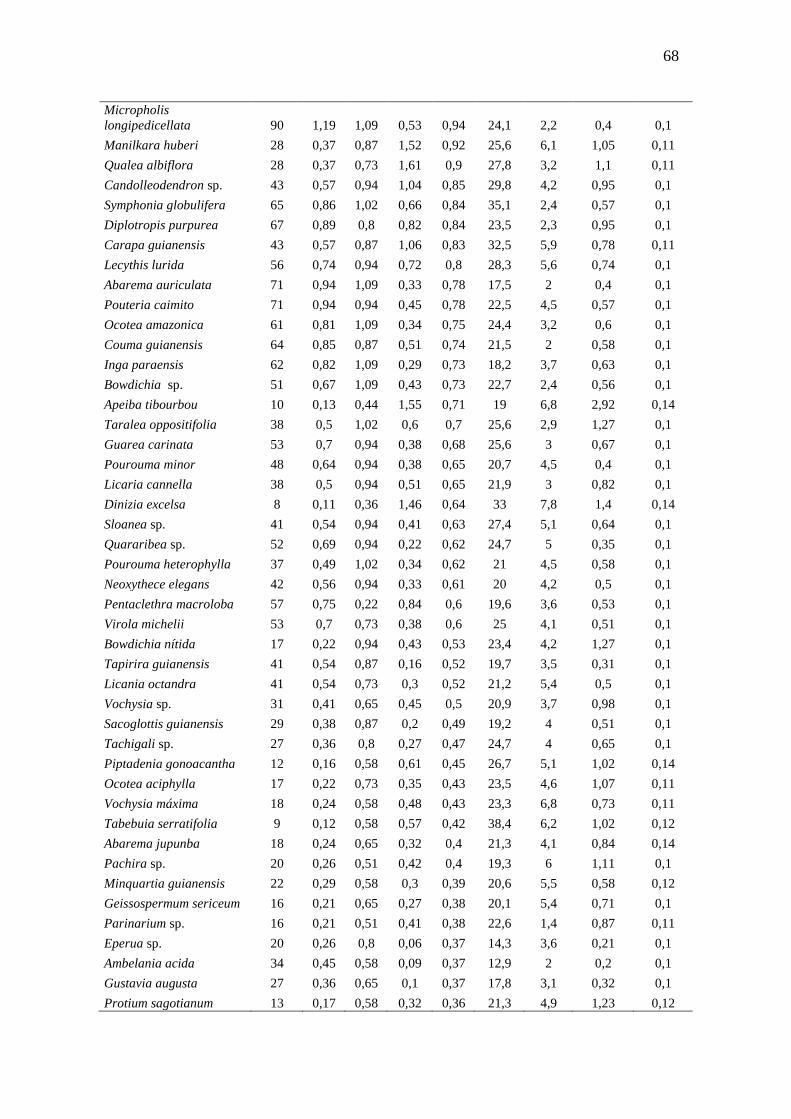
68
Micropholis
longipedicellata 90 1,19 1,09 0,53 0,94 24,1 2,2 0,4 0,1
Manilkara huberi 28 0,37 0,87 1,52 0,92 25,6 6,1 1,05 0,11
Qualea albiflora 28 0,37 0,73 1,61 0,9 27,8 3,2 1,1 0,11
Candolleodendron sp. 43 0,57 0,94 1,04 0,85 29,8 4,2 0,95 0,1
Symphonia globulifera 65 0,86 1,02 0,66 0,84 35,1 2,4 0,57 0,1
Diplotropis purpurea 67 0,89 0,8 0,82 0,84 23,5 2,3 0,95 0,1
Carapa guianensis 43 0,57 0,87 1,06 0,83 32,5 5,9 0,78 0,11
Lecythis lurida 56 0,74 0,94 0,72 0,8 28,3 5,6 0,74 0,1
Abarema auriculata 71 0,94 1,09 0,33 0,78 17,5 2 0,4 0,1
Pouteria caimito 71 0,94 0,94 0,45 0,78 22,5 4,5 0,57 0,1
Ocotea amazonica 61 0,81 1,09 0,34 0,75 24,4 3,2 0,6 0,1
Couma guianensis 64 0,85 0,87 0,51 0,74 21,5 2 0,58 0,1
Inga paraensis 62 0,82 1,09 0,29 0,73 18,2 3,7 0,63 0,1
Bowdichia sp. 51 0,67 1,09 0,43 0,73 22,7 2,4 0,56 0,1
Apeiba tibourbou 10 0,13 0,44 1,55 0,71 19 6,8 2,92 0,14
Taralea oppositifolia 38 0,5 1,02 0,6 0,7 25,6 2,9 1,27 0,1
Guarea carinata 53 0,7 0,94 0,38 0,68 25,6 3 0,67 0,1
Pourouma minor 48 0,64 0,94 0,38 0,65 20,7 4,5 0,4 0,1
Licaria cannella 38 0,5 0,94 0,51 0,65 21,9 3 0,82 0,1
Dinizia excelsa 8 0,11 0,36 1,46 0,64 33 7,8 1,4 0,14
Sloanea sp. 41 0,54 0,94 0,41 0,63 27,4 5,1 0,64 0,1
Quararibea sp. 52 0,69 0,94 0,22 0,62 24,7 5 0,35 0,1
Pourouma heterophylla 37 0,49 1,02 0,34 0,62 21 4,5 0,58 0,1
Neoxythece elegans 42 0,56 0,94 0,33 0,61 20 4,2 0,5 0,1
Pentaclethra macroloba 57 0,75 0,22 0,84 0,6 19,6 3,6 0,53 0,1
Virola michelii 53 0,7 0,73 0,38 0,6 25 4,1 0,51 0,1
Bowdichia nítida 17 0,22 0,94 0,43 0,53 23,4 4,2 1,27 0,1
Tapirira guianensis 41 0,54 0,87 0,16 0,52 19,7 3,5 0,31 0,1
Licania octandra 41 0,54 0,73 0,3 0,52 21,2 5,4 0,5 0,1
Vochysia sp. 31 0,41 0,65 0,45 0,5 20,9 3,7 0,98 0,1
Sacoglottis guianensis 29 0,38 0,87 0,2 0,49 19,2 4 0,51 0,1
Tachigali sp. 27 0,36 0,8 0,27 0,47 24,7 4 0,65 0,1
Piptadenia gonoacantha 12 0,16 0,58 0,61 0,45 26,7 5,1 1,02 0,14
Ocotea aciphylla 17 0,22 0,73 0,35 0,43 23,5 4,6 1,07 0,11
Vochysia máxima 18 0,24 0,58 0,48 0,43 23,3 6,8 0,73 0,11
Tabebuia serratifolia 9 0,12 0,58 0,57 0,42 38,4 6,2 1,02 0,12
Abarema jupunba 18 0,24 0,65 0,32 0,4 21,3 4,1 0,84 0,14
Pachira sp. 20 0,26 0,51 0,42 0,4 19,3 6 1,11 0,1
Minquartia guianensis 22 0,29 0,58 0,3 0,39 20,6 5,5 0,58 0,12
Geissospermum sericeum 16 0,21 0,65 0,27 0,38 20,1 5,4 0,71 0,1
Parinarium sp. 16 0,21 0,51 0,41 0,38 22,6 1,4 0,87 0,11
Eperua sp. 20 0,26 0,8 0,06 0,37 14,3 3,6 0,21 0,1
Ambelania acida 34 0,45 0,58 0,09 0,37 12,9 2 0,2 0,1
Gustavia augusta 27 0,36 0,65 0,1 0,37 17,8 3,1 0,32 0,1
Protium sagotianum 13 0,17 0,58 0,32 0,36 21,3 4,9 1,23 0,12
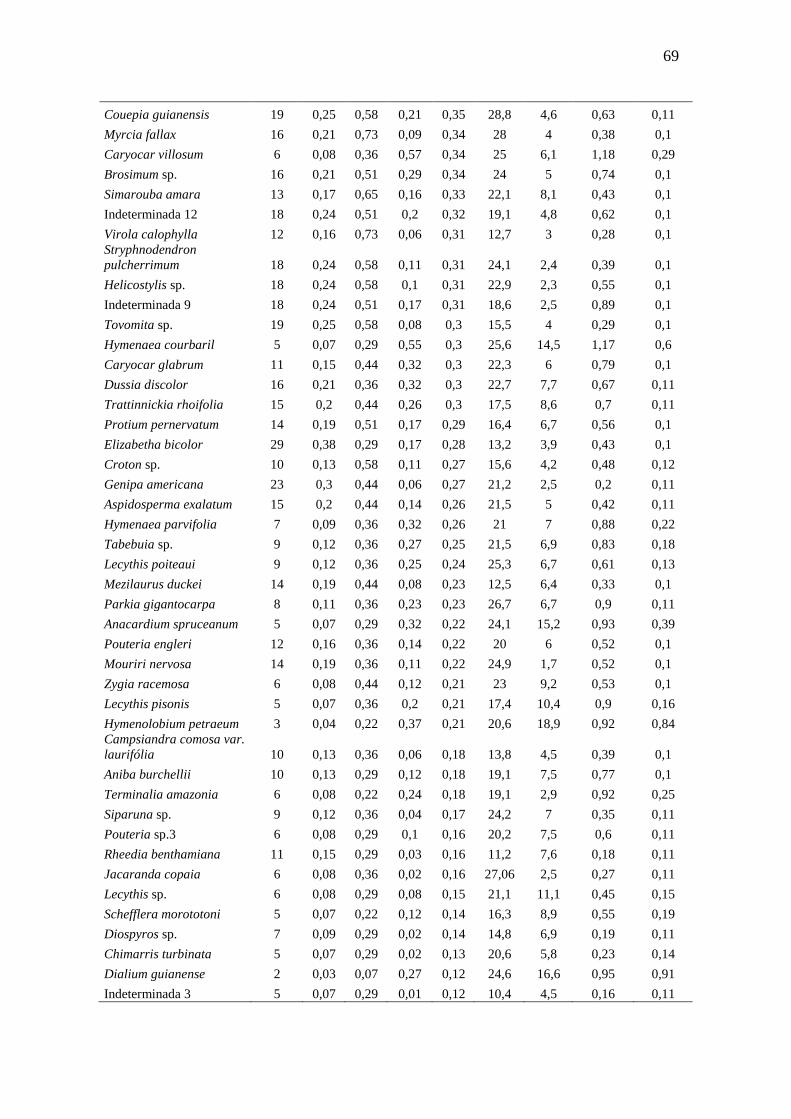
69
Couepia guianensis 19 0,25 0,58 0,21 0,35 28,8 4,6 0,63 0,11
Myrcia fallax 16 0,21 0,73 0,09 0,34 28 4 0,38 0,1
Caryocar villosum 6 0,08 0,36 0,57 0,34 25 6,1 1,18 0,29
Brosimum sp. 16 0,21 0,51 0,29 0,34 24 5 0,74 0,1
Simarouba amara 13 0,17 0,65 0,16 0,33 22,1 8,1 0,43 0,1
Indeterminada 12 18 0,24 0,51 0,2 0,32 19,1 4,8 0,62 0,1
Virola calophylla 12 0,16 0,73 0,06 0,31 12,7 3 0,28 0,1
Stryphnodendron
pulcherrimum 18 0,24 0,58 0,11 0,31 24,1 2,4 0,39 0,1
Helicostylis sp. 18 0,24 0,58 0,1 0,31 22,9 2,3 0,55 0,1
Indeterminada 9 18 0,24 0,51 0,17 0,31 18,6 2,5 0,89 0,1
Tovomita sp. 19 0,25 0,58 0,08 0,3 15,5 4 0,29 0,1
Hymenaea courbaril 5 0,07 0,29 0,55 0,3 25,6 14,5 1,17 0,6
Caryocar glabrum 11 0,15 0,44 0,32 0,3 22,3 6 0,79 0,1
Dussia discolor 16 0,21 0,36 0,32 0,3 22,7 7,7 0,67 0,11
Trattinnickia rhoifolia 15 0,2 0,44 0,26 0,3 17,5 8,6 0,7 0,11
Protium pernervatum 14 0,19 0,51 0,17 0,29 16,4 6,7 0,56 0,1
Elizabetha bicolor 29 0,38 0,29 0,17 0,28 13,2 3,9 0,43 0,1
Croton sp. 10 0,13 0,58 0,11 0,27 15,6 4,2 0,48 0,12
Genipa americana 23 0,3 0,44 0,06 0,27 21,2 2,5 0,2 0,11
Aspidosperma exalatum 15 0,2 0,44 0,14 0,26 21,5 5 0,42 0,11
Hymenaea parvifolia 7 0,09 0,36 0,32 0,26 21 7 0,88 0,22
Tabebuia sp. 9 0,12 0,36 0,27 0,25 21,5 6,9 0,83 0,18
Lecythis poiteaui 9 0,12 0,36 0,25 0,24 25,3 6,7 0,61 0,13
Mezilaurus duckei 14 0,19 0,44 0,08 0,23 12,5 6,4 0,33 0,1
Parkia gigantocarpa 8 0,11 0,36 0,23 0,23 26,7 6,7 0,9 0,11
Anacardium spruceanum 5 0,07 0,29 0,32 0,22 24,1 15,2 0,93 0,39
Pouteria engleri 12 0,16 0,36 0,14 0,22 20 6 0,52 0,1
Mouriri nervosa 14 0,19 0,36 0,11 0,22 24,9 1,7 0,52 0,1
Zygia racemosa 6 0,08 0,44 0,12 0,21 23 9,2 0,53 0,1
Lecythis pisonis 5 0,07 0,36 0,2 0,21 17,4 10,4 0,9 0,16
Hymenolobium petraeum 3 0,04 0,22 0,37 0,21 20,6 18,9 0,92 0,84
Campsiandra comosa var.
laurifólia 10 0,13 0,36 0,06 0,18 13,8 4,5 0,39 0,1
Aniba burchellii 10 0,13 0,29 0,12 0,18 19,1 7,5 0,77 0,1
Terminalia amazonia 6 0,08 0,22 0,24 0,18 19,1 2,9 0,92 0,25
Siparuna sp. 9 0,12 0,36 0,04 0,17 24,2 7 0,35 0,11
Pouteria sp.3 6 0,08 0,29 0,1 0,16 20,2 7,5 0,6 0,11
Rheedia benthamiana 11 0,15 0,29 0,03 0,16 11,2 7,6 0,18 0,11
Jacaranda copaia 6 0,08 0,36 0,02 0,16 27,06 2,5 0,27 0,11
Lecythis sp. 6 0,08 0,29 0,08 0,15 21,1 11,1 0,45 0,15
Schefflera morototoni 5 0,07 0,22 0,12 0,14 16,3 8,9 0,55 0,19
Diospyros sp. 7 0,09 0,29 0,02 0,14 14,8 6,9 0,19 0,11
Chimarris turbinata 5 0,07 0,29 0,02 0,13 20,6 5,8 0,23 0,14
Dialium guianense 2 0,03 0,07 0,27 0,12 24,6 16,6 0,95 0,91
Indeterminada 3 5 0,07 0,29 0,01 0,12 10,4 4,5 0,16 0,11
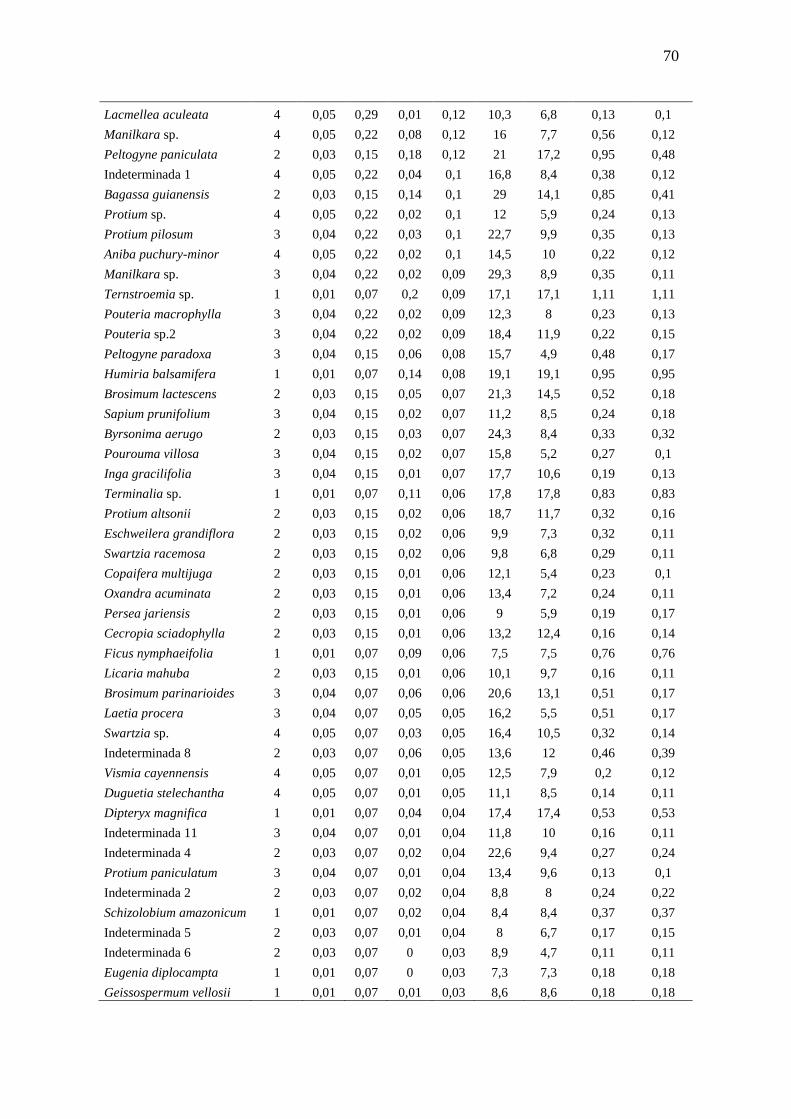
70
Lacmellea aculeata 4 0,05 0,29 0,01 0,12 10,3 6,8 0,13 0,1
Manilkara sp. 4 0,05 0,22 0,08 0,12 16 7,7 0,56 0,12
Peltogyne paniculata 2 0,03 0,15 0,18 0,12 21 17,2 0,95 0,48
Indeterminada 1 4 0,05 0,22 0,04 0,1 16,8 8,4 0,38 0,12
Bagassa guianensis 2 0,03 0,15 0,14 0,1 29 14,1 0,85 0,41
Protium sp. 4 0,05 0,22 0,02 0,1 12 5,9 0,24 0,13
Protium pilosum 3 0,04 0,22 0,03 0,1 22,7 9,9 0,35 0,13
Aniba puchury-minor 4 0,05 0,22 0,02 0,1 14,5 10 0,22 0,12
Manilkara sp. 3 0,04 0,22 0,02 0,09 29,3 8,9 0,35 0,11
Ternstroemia sp. 1 0,01 0,07 0,2 0,09 17,1 17,1 1,11 1,11
Pouteria macrophylla 3 0,04 0,22 0,02 0,09 12,3 8 0,23 0,13
Pouteria sp.2 3 0,04 0,22 0,02 0,09 18,4 11,9 0,22 0,15
Peltogyne paradoxa 3 0,04 0,15 0,06 0,08 15,7 4,9 0,48 0,17
Humiria balsamifera 1 0,01 0,07 0,14 0,08 19,1 19,1 0,95 0,95
Brosimum lactescens 2 0,03 0,15 0,05 0,07 21,3 14,5 0,52 0,18
Sapium prunifolium 3 0,04 0,15 0,02 0,07 11,2 8,5 0,24 0,18
Byrsonima aerugo 2 0,03 0,15 0,03 0,07 24,3 8,4 0,33 0,32
Pourouma villosa 3 0,04 0,15 0,02 0,07 15,8 5,2 0,27 0,1
Inga gracilifolia 3 0,04 0,15 0,01 0,07 17,7 10,6 0,19 0,13
Terminalia sp. 1 0,01 0,07 0,11 0,06 17,8 17,8 0,83 0,83
Protium altsonii 2 0,03 0,15 0,02 0,06 18,7 11,7 0,32 0,16
Eschweilera grandiflora 2 0,03 0,15 0,02 0,06 9,9 7,3 0,32 0,11
Swartzia racemosa 2 0,03 0,15 0,02 0,06 9,8 6,8 0,29 0,11
Copaifera multijuga 2 0,03 0,15 0,01 0,06 12,1 5,4 0,23 0,1
Oxandra acuminata 2 0,03 0,15 0,01 0,06 13,4 7,2 0,24 0,11
Persea jariensis 2 0,03 0,15 0,01 0,06 9 5,9 0,19 0,17
Cecropia sciadophylla 2 0,03 0,15 0,01 0,06 13,2 12,4 0,16 0,14
Ficus nymphaeifolia 1 0,01 0,07 0,09 0,06 7,5 7,5 0,76 0,76
Licaria mahuba 2 0,03 0,15 0,01 0,06 10,1 9,7 0,16 0,11
Brosimum parinarioides 3 0,04 0,07 0,06 0,06 20,6 13,1 0,51 0,17
Laetia procera 3 0,04 0,07 0,05 0,05 16,2 5,5 0,51 0,17
Swartzia sp. 4 0,05 0,07 0,03 0,05 16,4 10,5 0,32 0,14
Indeterminada 8 2 0,03 0,07 0,06 0,05 13,6 12 0,46 0,39
Vismia cayennensis 4 0,05 0,07 0,01 0,05 12,5 7,9 0,2 0,12
Duguetia stelechantha 4 0,05 0,07 0,01 0,05 11,1 8,5 0,14 0,11
Dipteryx magnifica 1 0,01 0,07 0,04 0,04 17,4 17,4 0,53 0,53
Indeterminada 11 3 0,04 0,07 0,01 0,04 11,8 10 0,16 0,11
Indeterminada 4 2 0,03 0,07 0,02 0,04 22,6 9,4 0,27 0,24
Protium paniculatum 3 0,04 0,07 0,01 0,04 13,4 9,6 0,13 0,1
Indeterminada 2 2 0,03 0,07 0,02 0,04 8,8 8 0,24 0,22
Schizolobium amazonicum 1 0,01 0,07 0,02 0,04 8,4 8,4 0,37 0,37
Indeterminada 5 2 0,03 0,07 0,01 0,04 8 6,7 0,17 0,15
Indeterminada 6 2 0,03 0,07 0 0,03 8,9 4,7 0,11 0,11
Eugenia diplocampta 1 0,01 0,07 0 0,03 7,3 7,3 0,18 0,18
Geissospermum vellosii 1 0,01 0,07 0,01 0,03 8,6 8,6 0,18 0,18
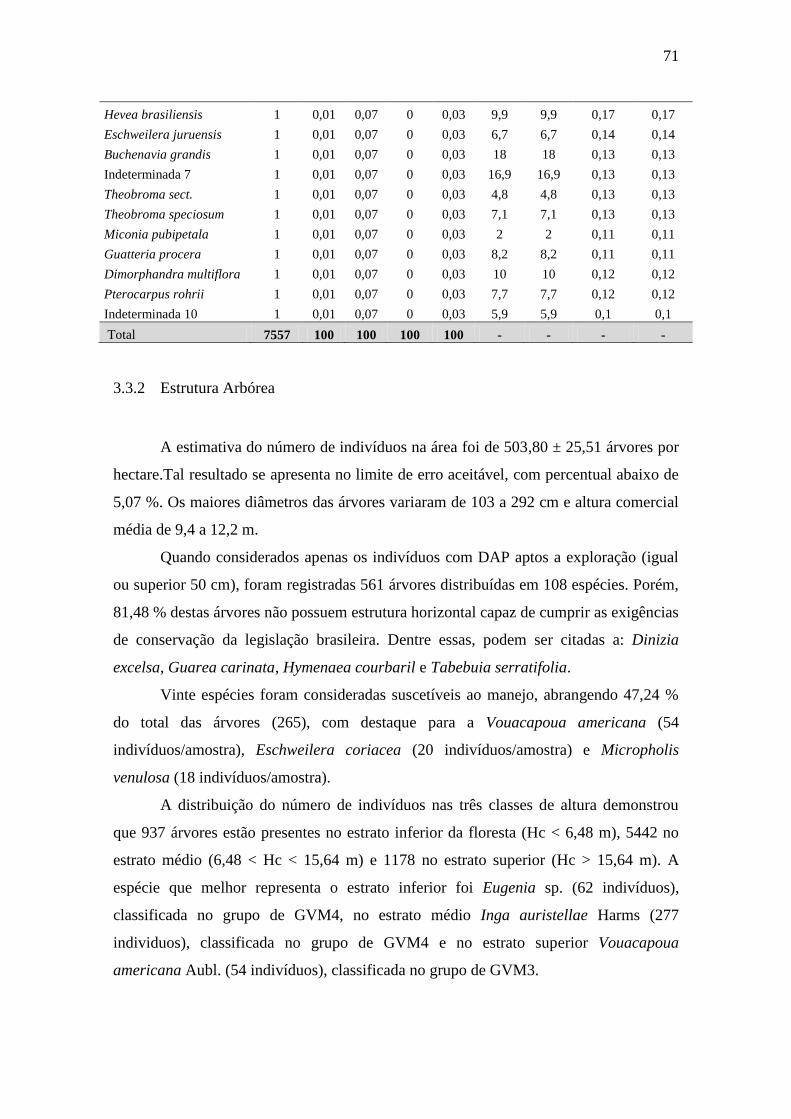
71
Hevea brasiliensis 1 0,01 0,07 0 0,03 9,9 9,9 0,17 0,17
Eschweilera juruensis 1 0,01 0,07 0 0,03 6,7 6,7 0,14 0,14
Buchenavia grandis 1 0,01 0,07 0 0,03 18 18 0,13 0,13
Indeterminada 7 1 0,01 0,07 0 0,03 16,9 16,9 0,13 0,13
Theobroma sect. 1 0,01 0,07 0 0,03 4,8 4,8 0,13 0,13
Theobroma speciosum 1 0,01 0,07 0 0,03 7,1 7,1 0,13 0,13
Miconia pubipetala 1 0,01 0,07 0 0,03 2 2 0,11 0,11
Guatteria procera 1 0,01 0,07 0 0,03 8,2 8,2 0,11 0,11
Dimorphandra multiflora 1 0,01 0,07 0 0,03 10 10 0,12 0,12
Pterocarpus rohrii 1 0,01 0,07 0 0,03 7,7 7,7 0,12 0,12
Indeterminada 10 1 0,01 0,07 0 0,03 5,9 5,9 0,1 0,1
Total 7557 100 100 100 100 - - - -
3.3.2 Estrutura Arbórea
A estimativa do número de indivíduos na área foi de 503,80 ± 25,51 árvores por
hectare.Tal resultado se apresenta no limite de erro aceitável, com percentual abaixo de
5,07 %. Os maiores diâmetros das árvores variaram de 103 a 292 cm e altura comercial
média de 9,4 a 12,2 m.
Quando considerados apenas os indivíduos com DAP aptos a exploração (igual
ou superior 50 cm), foram registradas 561 árvores distribuídas em 108 espécies. Porém,
81,48 % destas árvores não possuem estrutura horizontal capaz de cumprir as exigências
de conservação da legislação brasileira. Dentre essas, podem ser citadas a: Dinizia
excelsa, Guarea carinata, Hymenaea courbaril e Tabebuia serratifolia.
Vinte espécies foram consideradas suscetíveis ao manejo, abrangendo 47,24 %
do total das árvores (265), com destaque para a Vouacapoua americana (54
indivíduos/amostra), Eschweilera coriacea (20 indivíduos/amostra) e Micropholis
venulosa (18 indivíduos/amostra).
A distribuição do número de indivíduos nas três classes de altura demonstrou
que 937 árvores estão presentes no estrato inferior da floresta (Hc < 6,48 m), 5442 no
estrato médio (6,48 < Hc < 15,64 m) e 1178 no estrato superior (Hc > 15,64 m). A
espécie que melhor representa o estrato inferior foi Eugenia sp. (62 indivíduos),
classificada no grupo de GVM4, no estrato médio Inga auristellae Harms (277
individuos), classificada no grupo de GVM4 e no estrato superior Vouacapoua
americana Aubl. (54 indivíduos), classificada no grupo de GVM3.

72
O total de indivíduos inventariados subsidiaram a formação de quatorze classes
diamétricas, com amplitude de 5 cm, formando uma curva exponencial negativa, padrão
característico de florestas inequiâneas mistas. A primeira classe contemplou árvores
com DAP entre 10 e 15 cm, enquanto que na última classe foram agrupadas as árvores
com DAP superior a 75 cm.
As espécies que apresentaram maior número de indivíduos na qualidade do
fuste 1 (Qf1) foram: Eschweilera coriacea, Eschweilera sp. eInga auristellae,
Iryanthera paraensis e Protium tenuifolium, Pourouma ovata, Mezilaurus itauba e
Vouacapoua americana. Entretanto, as espécies com atributos do fuste menos aceitáveis
comercialmente foram: Inga auristellae Harms, e Eugeniasp. L. (Qf2); Tachigali
myrmecophila Ducke, Inga auristellae Harms e Theobroma subincanum Mart. (Qf3).
3.3.3 Potencial Madeireiro
O erro de amostragem para o volume das árvores foi de 8,96 %, com volume
total de 5748,2964 m3 em 15 ha, acarretando em valor médio de 383,22 m
3 ha
-1.O
volume por hectare distribuído entre as classes diamétricas evidenciou 67,33 m3 ha
-1 nas
árvores com DAP’s entre 10 e 40 cm, 22,37 m3 ha
-1 para arvores com DAP’s entre 40 e
50 cm e 293,53 m3 ha
-1 apenas para árvores com DAP’s maior 50 cm.
Quando consideradas as unidades primárias, verificou-se volumes superiores a
365 m³ por hectare, com predominância de indivíduos na qualidade do fuste 1. Foram
registradas poucas árvores tortuosas (Qf3) distribuídas em variações de 17 (UP1), 45
(UP2) e 164 (UP3) (Tabela 6).
Tabela 7. Estoque volumétrico e potencial das árvores nas unidades primárias (UP) numa floresta
ombrófila densa, Porto Grande, Amapá, Amazônia. Onde: Qf – qualidade do fuste; N – número total de
indivíduos; V – volume em m³.ha-1
; PC – presença de cupim; C – presença de cipós competidores; O –
presença de oco.
UP N V PC C O
1
1 2183 380,58 0 362 1
Qf 2 404 30,99 0 71 0
3 17 1,56 0 2 0
Subtotal 2604 413,12 0 435 1
2
1 1967 315,51 1633 466 0
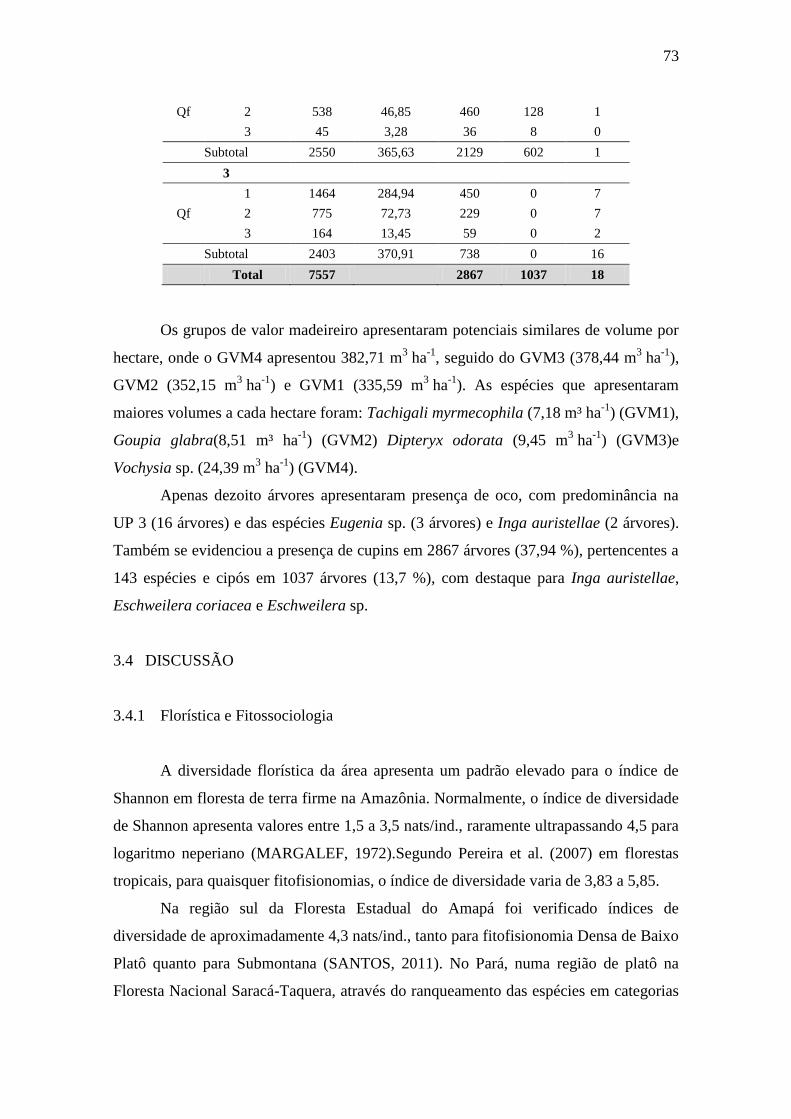
73
Qf 2 538 46,85 460 128 1
3 45 3,28 36 8 0
Subtotal 2550 365,63 2129 602 1
3
1 1464 284,94 450 0 7
Qf 2 775 72,73 229 0 7
3 164 13,45 59 0 2
Subtotal 2403 370,91 738 0 16
Total 7557
2867 1037 18
Os grupos de valor madeireiro apresentaram potenciais similares de volume por
hectare, onde o GVM4 apresentou 382,71 m3 ha
-1, seguido do GVM3 (378,44 m
3 ha
-1),
GVM2 (352,15 m3 ha
-1) e GVM1 (335,59 m
3 ha
-1). As espécies que apresentaram
maiores volumes a cada hectare foram: Tachigali myrmecophila (7,18 m³ ha-1
) (GVM1),
Goupia glabra(8,51 m³ ha-1
) (GVM2) Dipteryx odorata (9,45 m3 ha
-1) (GVM3)e
Vochysia sp. (24,39 m3 ha
-1) (GVM4).
Apenas dezoito árvores apresentaram presença de oco, com predominância na
UP 3 (16 árvores) e das espécies Eugenia sp. (3 árvores) e Inga auristellae (2 árvores).
Também se evidenciou a presença de cupins em 2867 árvores (37,94 %), pertencentes a
143 espécies e cipós em 1037 árvores (13,7 %), com destaque para Inga auristellae,
Eschweilera coriacea e Eschweilera sp.
3.4 DISCUSSÃO
3.4.1 Florística e Fitossociologia
A diversidade florística da área apresenta um padrão elevado para o índice de
Shannon em floresta de terra firme na Amazônia. Normalmente, o índice de diversidade
de Shannon apresenta valores entre 1,5 a 3,5 nats/ind., raramente ultrapassando 4,5 para
logaritmo neperiano (MARGALEF, 1972).Segundo Pereira et al. (2007) em florestas
tropicais, para quaisquer fitofisionomias, o índice de diversidade varia de 3,83 a 5,85.
Na região sul da Floresta Estadual do Amapá foi verificado índices de
diversidade de aproximadamente 4,3 nats/ind., tanto para fitofisionomia Densa de Baixo
Platô quanto para Submontana (SANTOS, 2011). No Pará, numa região de platô na
Floresta Nacional Saracá-Taquera, através do ranqueamento das espécies em categorias

74
ecológicas de prioridade alta, intermediária e baixa, encontrou-se um índice de Shannon
de 5,43 nats/ind. (SALOMÃO et al., 2012).
Quanto à riqueza de espécies, o número encontrado (182) está abaixo dos
resultados de Oliveira et al. (2008) e Alves e Miranda (2008), os quais trabalharam com
o mesmo nível de inclusão, mas com áreas amostrais distintas. Entretanto, Espírito-
Santo et al. (2005) e Pereira et al. (2011), em áreas amostrais similares, apresentaram
riqueza variando de 100 a 200 espécies. Comumente, a riqueza explorada em florestas
submetidas ao manejo sustentável no Amapá varia de 10 a 20 espécies, com volumes
inferiores a 25 m³ ha-1
.
Atualmente, são mais de 1500 espécies comercializadas no Brasil, registradas no
sistema de controle de origem florestal (DOF), sendo 50 % do volume total concentrado
em 33 espécies (NETO et al., 2010).
O elevado número de espécies comuns entre as unidades primárias demonstra
uniformidade no estabelecimento. Além disso, nenhuma das espécies identificadas
consta na lista de espécies da flora brasileira ameaçadas de extinção (IN 06/2008
MMA).
A ausência de espécies ameaçadas e o registro de inúmeras espécies comerciais
proporcionam o planejamento dos processos operacionais da cadeia de produção
florestal. Além da riqueza florística, os padrões espaciais de distribuição das espécies na
área auxiliam na exploração das toras manejadas, com alternativas para construção de
ramais de escoamento distintos para minimizar custos.
A presença do agrupamento da maioria das espécies permite concentrar as
atividades de exploração num número menor de ramais e, consequentemente, na
utilização reduzida de pátios na floresta.
Yared e Souza (1993) afirmam que a distribuição espacial contribui na seleção
das árvores alvo de exploração, bem como daquelas que deverão ser mantidas como
matrizes (porta-sementes), a fim de garantir a regeneração natural. Cabe ressaltar que os
padrões espaciais são próprios de cada espécie, determinados a partir da dispersão de
frutos e sementes e adaptações edáficas do sítio.
Evidenciou-se uma baixa variação da densidade e frequência relativa entre as
espécies ao longo da área, determinando o estabelecimento de distintas espécies nas
variadas condições do sítio, mesmo com número reduzido de indivíduos. Resultados
diferentes foram encontrados por Alves e Miranda (2008), os quais na busca para

75
descrever a estrutura de uma floresta de terra firme sob regime de manejo na Amazônia
Oriental, Pará, constataram que algumas espécies de valor comercial apresentaram
variações importantes na densidade, sugerindo risco de extinção em comunidades onde
as espécies apresentam densidade muito baixa.
Comportamento diferente foi observado quanto à dominância relativa. As
espécies com maior crescimento diamétrico e número de indivíduos se apresentaram
mais adaptadas à região. Estas espécies geralmente estão presentes entre as de maior
valor de importância, com poucos indivíduos, mas com grandes diâmetros, acarretando
numa falsa interpretação de adaptação e estabelecimento na área.
3.4.2 Estrutura Arbórea
O número de indivíduos registrados no inventário está dentro dos padrões
encontrados na Amazônia. Souza et al. (2006) trabalhando com objetivo de analisar a
estrutura horizontal (árvores com DAP≥ 15 cm) numa Unidade de Manejo Florestal em
Paragominas, Pará, estimou uma densidade variando de 309 a 322,4 indivíduos por
hectare.
A quantidade de árvores com DAP aptos a exploração variou em razão dos
critérios selecionados. Atualmente, os critérios de seleção são orientados pela IN
05/2006 MMA, a qual dispõe sobre procedimentos técnicos para elaboração,
apresentação, execução e avaliação técnica de Planos de Manejo Florestal Sustentável
(PMFS) nas florestas da Amazônia legal.
Um ponto relevante trata da obrigação do detentor em manter pelo menos 10 %
do numero de árvores por espécie na área de efetiva exploração, respeitando o limite
mínimo de manutenção de 3 árvores por espécie por 100 ha.
O dossel da floresta foi considerado de alto porte. Esta característica viabiliza a
estimativa de maiores volumes por árvore. Entretanto, as variações das alturas
comerciais por espécies estão mais relacionadas às avaliações da inserção do primeiro
galho. A altura comercial considerada pela indústria madeireira na Amazônia varia de 6
a 33 m (RIVEIRA, 2007; GONÇALVES et al., 2008), em diferentes critérios de
avaliação, com predominância da estimativa através do principio geométrico, sem uso
de aparelhos ópticos.

76
O decréscimo do número de indivíduos nas sucessivas classes diamétricas
configura uma suave curva tendendo a reta com declividade negativa, descendendo para
direita. Segundo Sterba (2004), frequentemente a estabilidade de distribuições
diamétricas de florestas inequiâneas são pensadas em comunidades, compostas por
populações de espécies com distintas idades.
A distribuição diamétrica exponencial das árvores em florestas nativas, com
proporcionalidade constante em classes sucessivas, é o indicio de estabilidade na
comunidade (DE LIOCOURT, 1898).
3.4.3 Potencial Madeireiro
Verificou-se que há potencial volum étrico na área amostral para fins de manejo
florestal, com estimativas por hectare similar a florestas já exploradas. É importante
salientar que as árvores de estoque, situadas entre os DAP’s 40 e 50 cm, possuem um
volume capaz de repor a quantidade permitida para exploração nos ciclos de corte (até
30 m³ ha-1
).
A conicidade das árvores também contribui para agregar valor madeireiro à área.
Em todas as UP’s o número de indivíduos com características de fuste mais cilíndricas
foi superior aos indivíduos mais tortuosos. Tal fato evidencia maior rendimento das
toras que serão submetidas à exploração, com garantia de manutenção das árvores que
possuem menor valor econômico, porém com fundamental função ecológica.
A aptidão ao manejo florestal também foi evidenciada por baixos percentuais de
árvores com oco e cupim. Além disso, foi constatado que a presença de cipós não será
limitante nas operações pré-exploratórias, devido sua baixa incidência. Segundo Vidal e
Gerwing (2003), a abundância de cipós reduz o valor silvicultural da floresta, sendo
necessário o desenvolvimento de uma estratégia eficiente de manejo nas florestas
designadas para a produção madeireira.
Estudos apontam que altas densidades de cipós na floresta amazônica
representam evidencias de distúrbios severo no passado (BALÉE E CAMPBELL,
1990), fato não constatado na área em estudo.

77
3.5 CONCLUSÕES
A diversidade florística da área apresentou um padrão elevado para o ambiente
de terra firme na Amazônia, representada por elevado número de indivíduos distribuídos
em poucas espécies.
A estrutura fitossociológica da área amostral apresenta inúmeras espécies com
capacidade de perpetuação de remanescentes, bem como distribuição espacial e relações
interespecíficas que garantem a exploração de árvores com aptidão ao manejo.
Na área em estudo há potencial volumétrico para fins de manejo, com
estimativas por hectare superior a permitida nas bases legais da política florestal
brasileira. No entanto, em razão dos critérios técnicos previstos na legislação, a
quantidade de árvores com DAP aptos a exploração variou entre as espécies e áreas.
O planejamento de operações de manejo é promissor na região, devido ao grande
número de espécies aptas ao manejo. No entanto, a área de efetivo manejo deve ser
definida numa única Unidade de Produção Anual para viabilizar a utilização dos
recursos.

78
4 NÍVEIS DE REGENERAÇÃO NATURAL PARA SUBSIDIO AO
MANEJO EM FLORESTA DE TERRA FIRME NO AMAPÁ

79
RESUMO
Este trabalho objetivou identificar a regeneração natural na fitofisionomia ombrófila
densa da Floresta Estadual do Amapá, descrevendo as relações entre estratos verticais.
A área de estudo está localizada no município de Porto Grande/AP, Amazônia Oriental.
Foram implantados três conglomerados, equidistantes em 2.500 m, para a estimativa da
regeneração natural. Foram utilizadas cem subparcelas de 10 x 10 m para o estudo das
arvoretas (5,0 < DAP’s (Diâmetro a altura do peito a 1.30 m do solo) < 10,0 cm) e varas
(5 x 5 m) (2,5 < DAP’s < 5,0 cm) por conglomerado. Foram coletadas as alturas e
nomes populares. As alturas foram divididas em três classes de regeneração para
estimativa dos parâmetros fitossociológicos de freqüência e densidade, bem como a
regeneração por classe de tamanho e total. A diversidade foi estimada pelo Índice de
Shannon. Também se analisou a similaridade florística entre as varas e arvoretas por
conglomerado. O inventário contemplou 2.700 indivíduos, pertencentes a 38 famílias
botânicas, 93 gêneros e 141 espécies arbóreas, com 6 indeterminadas. O índice de
Shannon foi de 4,21 (arvoretas) e 4,11 nats.ind.-1
(varas). Foi constatada a ocorrência de
33 espécies comuns nas três classes de regeneração. A regeneração natural total
apresentou variações de 10,3 a 0,1 % (varas) e 5,6 a 0,1 % (arvoretas). As espécies com
menores percentuais foram as Indeterminadas, Vouacapoua americana, Carapa
guianensis, Virola calophylla e Manilkara huberi. Logo, fica evidente que as espécies
estão desenvolvendo seu processo sucessional de forma eficiente, garantindo a
conservação da fitofisionomia da região.
Palavras-chave: Classes de altura; Similaridade florística; Métodos de amostragem.

80
ABSTRACT
This study aimed to identify the natural regeneration in dense rain forest vegetation,
Amapa, describing relations between vertical stract. The study area is located in Porto
Grande / AP, eastern Amazon. Were implanted three conglomerates, equidistant at
2.500 m, for regeneration estimation. We used a hundred plots of 10 x 10 m for the
study of saplings (5.0 < DBH (diameter at breast height to 1.30 m soil) < 10.0 cm) and
sticks (5 x 5 m) (2.5 <DBH <5.0 cm) by conglomerate. We collected the heights and
popular names. The heights were divided into three classes of regeneration to estimate
parameters phytosociological frequency and density, as well as regeneration by size
class and overall. Diversity was estimated by Shannon Index. Also analyzed the floristic
similarity between cluster analisys. The inventory included 2,700 individuals belonging
to 38 botanical families, 93 genera and 141 species of trees, with 6 undetermined. The
Shannon index was 4.21 (saplings) and 4.11 nats.ind.-1
(sticks). It has been found the
occurrence of 33 common species in three classes of regeneration. The total natural
regeneration varied from 10.3 to 0.1% (sticks) and 5.6 to 0.1% (saplings). The species
with the lowest percentages were Indeterminate, Vouacapoua americana, Carapa
guianensis, Virola calophylla and Manilkara huberi. Thus, it is evident that species are
developing their successional process efficiently, ensuring conservation of vegetation
type.
Key words: Height classes; floristic similarity; Sampling

81
4.1 INTRODUÇÃO
A exuberância e fragilidade dos ambientes na Amazônia são alvos de inúmeras
discussões na sociedade, sejam para utilização dos recursos ou preservação dos
ecossistemas. No caso das florestas, a composição e distribuição das árvores,
principalmente dos indivíduos em fase de regeneração natural, orientam o
desenvolvimento dos temas sobre sustentabilidade.
Atualmente, a sustentabilidade das atividades de exploração nas florestas
tropicais é questionada, devido à quantidade retirada de madeira ocorrer sem considerar
a peculiaridade de cada sítio. A legislação brasileira permite nas florestas primitivas da
Amazônia a retirada de até 30m³ a cada hectare, prevista na IN 05/2006 MMA.
Entretanto, o estabelecimento da regeneração natural varia de acordo com as condições
de cada sítio e das operações de manejo.
Diante desta problemática, caracterizar o estado da regeneração natural em
floresta tropicais da Amazônia é fundamental para inferir sobre o estado de conservação
e estoque disponível de espécies para garantir a resiliência do meio. Tal prerrogativa é
fundamentada na teoria de que a regeneração natural representa o conjunto de
indivíduos capazes de serem recrutados para os estádios posteriores (SILVA et al.,
2007; GAMA et al., 2003).
De acordo com Martins e Rodrigues (2002), é preciso caracterizar o momento de
estabelecimento das espécies na floresta para definir estratégias de conservação. A
maneira mais tradicional de caracterizar as espécies regenerantes é por meio de
amostras quali-quantitativas na floresta. Dessa maneira, as informações permitem
reconhecer a função das espécies na comunidade, bem como suas preferências em
habitat e relações ecológicas inter e intraespecíficas.
Nesses termos, uma avaliação momentânea da regeneração natural subsidia a
predição da estrutura da floresta adulta. Assim, a garantia da permanência de
determinada espécie em uma floresta é função direta do número de indivíduos e da
distribuição nas classes de tamanho, baseados no diâmetro ou altura.
Em florestas ombrófila densas da Amazônia, estudos apontam alta diversidade
de espécies arbóreas em fase de regeneração, porém com grande percentual de espécies
representadas apenas por um indivíduo (PRANCE et al., 1976; LIMA FILHO, 2001;
APARÍCIO, 2011). No Amapá, o desenvolvimento de uma economia sustentável

82
baseada nas pretensões de destinar a Floresta Estadual a concessões é limitado pela
carência de informações técnicas sobre o estado das espécies arbóreas nas distintas fases
de crescimento.
Este cenário tem despertado o interesse da sociedade científica para desenvolver
pesquisas que possam garantir a conservação das espécies após inicio das atividades
exploratórias na Floresta Estadual. Logo, para se obter respostas que fortaleçam a
eficiência da implantação do manejo florestal sustentável, este trabalho tentou responder
a seguinte questão: A estrutura das espécies arbóreas regenerantes garante uma
significativa representatividade para compor o futuro dossel da florestal?
Diante do exposto, pretende-se verificar como ocorre a regeneração natural na
fitofisionomia ombrófila densa de terras baixas da Floresta Estadual do Amapá,
descrevendo as relações entre estratos verticais.
4.2 MATERIAL E MÉTODOS
O trabalho foi realizado na Floresta Estadual do Amapá (FLOTA/AP), no
município de Porto Grande, região central do estado do Amapá, Amazônia Oriental,
Brasil. A vegetação é perenifólia, característica da fitofisionomia ombrófila densa de
terras baixas (IBGE, 2012). O solo predominante é do tipo Latossolo Vermelho-
Amarelo distrófico. A declividade característica é ondulada, com altitude variando de
60 a 100 m (RADAM, 1974). O clima da região é Am (equatorial super-úmido),
segundo a classificação de Köppen.
O inventário florestal foi realizado em três conglomerados, equidistantes em
aproximadamente 2.500 m. Cada conglomerado foi constituído por cinco parcelas
permanentes de 100 x 100 m, equidistantes em 250 m. A área total estimada abrangeu
15 ha amostrados.
Para a estimativa da regeneração natural cada parcela foi dividida em cem
subparcelas de 10 x 10 m. Destas, foram selecionadas aleatoriamente vinte
subparcelas/parcela para avaliação das arvoretas. Nos vértices das subparcelas sorteadas
foram locadas, aleatoriamente, subunidades de 5 x 5 m para estimativas das varas.
O critério de inclusão foi adotado foi baseado nos diâmetros dos indivíduos para
distinguir arvoretas (5,0 < DAP’s (Diâmetro a altura do peito a 1.30 m do solo) < 10,0

83
cm) e varas (2,5 < DAP’s < 5,0 cm) seguindo a metodologia de Silva et al. (2005)
(Figura 7).
Figura 7. Unidade Amostral Secundária (US) permanente, utilizada para o estudo das espécies arbóreas
adultas e em regeneração natural no segundo módulo da FLOTA/AP.
Dos indivíduos amostrados foram registrados o nome popular e altura comercial
(Hc). Os nomes científicos foram conferidos no MOBOT segundo o sistema APG III
(2010). Para representar a regeneração natural como estrutura vertical, as alturas foram
divididas em classes de tamanho, onde a classe I contemplou indivíduos de 1,0 < H <
5,0 m; classe II com indivíduos de 5,0 < H < 10,0 m e a classe III com indivíduos > 10,0
m.
Os parâmetros fitossociológicos foram estimados nas classes de tamanho
(FINOL, 1971) das arvoretas e varas com auxílio do Microsoft EXCEL 2010. A
diversidade de espécies foi estimada a partir do Índice de hannon (H’) ( DUM, 1988).
A similaridade florística entre as classes de tamanho de arvoretas e varas nos
conglomerados foi realizada por meio de análise de agrupamento (método de ligação
simples), utilizando como medida a distância euclidiana.
4.3 RESULTADOS
O número total de indivíduos amostrados foi de 2.700 plantas vivas,
pertencentes a 38 famílias botânicas, 93 gêneros e 141 espécies arbóreas, com 6
indeterminadas. Quando consideradas apenas as arvoretas, foram inventariados 1.770
indivíduos, pertencentes a 30 famílias, 126 espécies e 4 indeterminadas. No caso dos

84
indivíduos em nível de vara foram inventariados 919 indivíduos, onde foram
quantificadas 29 famílias, 111 espécies e 5 indeterminadas.
índice de diversidade de hannon (H’) encontrado foi de 4,21 nats/ind. para a
regeneração natural em arvoreta e 4,11 nats/ind. em nível de vara. A diversidade de
espécies foi melhor representada entre as famílias com maior riqueza: Fabaceae (33
espécies), Sapotaceae (11), Lauraceae (10), Burseraceae (8) e Chrysobalanaceae (6).
Estas famílias representaram 50,37 % do total amostrado.
As espécies que se destacaram com maior número de indivíduos foram
respectivamente: Hirtella sp. (139 plantas), Eschweilera coriaceae (117 plantas),
Mouriri apiranga (116), Protium tenuifolium (112) e Guatteria sp. (99).
Os parâmetros fitossociológicos relativos demonstraram que a composição
florística e distribuição das espécies modifica à medida que as plantas vão atingindo o
dossel imediatamente superior. Quanto à freqüência relativa, observou-se que o número
de espécies reduziu com o aumento das alturas das plantas, tanto para arvoretas como
varas. As arvoretas apresentaram 48 (classe I), 44 (classe II) e 35 (classe III) espécies.
Nas varas foi observado uma menor riqueza, com 38 espécies na classe I, 37 na classe II
e apenas 4 na classe III.
As espécies que se destacaram com densidades relativas elevadas nas classes de
regeneração natural foram: Eugenia sp. e Hirtella sp. (5,3 %) e Mouriri apiranga (4,2
%) (arvoretas) e Mouriri apiranga e Hirtella sp. (varas) para classe I; Eschweilera
coriacea e Protium tenuifolium (arvoretas) e Guatteria sp. e Pourouma ovata (varas)
para classe II; e destaque apenas para a Protium tenuifolium tanto arvoreta quanto para
vara na classe III.
A distribuição das arvoretas em regeneração no estrato vertical evidenciou que
665 indivíduos, em 100 espécies, estão presentes na classe de altura I, seguida da classe
de altura II com 1.056 indivíduos pertencentes a 115 espécies e da classe de altura III
com apenas 49 indivíduos em 33 espécies.
As espécies inventariadas em nível de arvoreta (126) apresentaram apenas 33
táxons presentes em todas as classes de altura. Os percentuais de arvoretas presentes nas
classes de regeneração foram principalmente da Hirtella sp. e Eschweilera coriacea
(classe I e II) e Protium tenuifolium (classe III). A distribuição percentual das arvoretas
nas classes de regeneração natural demonstrou que 41,12 % estão concentradas na
classe I, seguida da classe II (40,77 %) e classe III (18,09 %) (Tabela 8).

85
Tabela 8. Estimativa do percentual da Regeneração Natural Total (RNT) para as arvoretas* (5.0 < DAP < 10.0 cm) e varas** (2.5 < DAP <5.0 cm) por classe de altura em
Porto Grande/AP, listados em ordem decrescente de acordo com o maior valor de RNT, onde DR= Densidade Relativa; FR = Freqüência Relativa e RNC1 = Regeneração
Natural na Classe 1 de altura; RNC2 = Regeneração Natural na Classe 2 de altura e RNC3 = Regeneração Natural na Classe 3 de altura.
NOME CIENTÍFICO FR* FR** DR* DR** RN* RN** FR* FR** DR* DR** RN* RN** FR* FR** DR* DR** RN* RN** RNT* RNT**
C1 C1 C1 C1 C1 C1 C2 C2 C2 C2 C2 C2 C3 C3 C3 C3 C3 C3
Abarema auriculata (Benth.) Barneby e J. W. Grimes 2,4 2,3 2,7 1,7 2,6 2,0 2,0 2,3 2,2 2,5 2,1 2,4 2,0 0,0 2,0 0,0 2,0 0,0 2,2 1,5
Alexa grandiflora Ducke 0,7 0,9 0,5 0,5 0,6 0,7 1,1 1,5 0,9 1,3 1,0 1,4 2,0 0,0 2,0 0,0 2,0 0,0 1,2 0,7
Ambelania acida Aubl. 2,9 1,2 3,2 0,7 3,0 0,9 1,3 0,8 0,9 0,6 1,1 0,7 0,0 0,0 0,0 0,0 0,0 0,0 1,4 0,5
Aniba burchellii Kosterm. 0,2 0,2 0,2 0,1 0,2 0,2 0,6 0,0 0,3 0,0 0,4 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,1
Aniba puchury-minor (Mart.) Mez 0,0 0,0 0,0 0,0 0,0 0,0 0,4 0,0 0,2 0,0 0,3 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,0
Apeiba tibourbou Aubl. 0,2 0,0 0,2 0,0 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,0
Aspidosperma exalatum Monach. 0,5 0,7 0,3 0,4 0,4 0,5 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,2
Aspidosperma excelsum Benth. 0,2 0,7 0,2 0,4 0,2 0,5 0,6 0,0 0,4 0,0 0,5 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,2
Bowdichia sp. Kunth 1,5 1,8 1,4 1,8 1,4 1,8 0,6 0,8 0,5 0,6 0,5 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,6 0,8
Bowdichia nitida Spruce 0,7 0,5 0,6 0,4 0,7 0,4 0,4 0,0 0,2 0,0 0,3 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,3 0,1
Brosimum sp. Sw. 0,2 0,7 0,2 0,4 0,2 0,5 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,2
Campsiandra comosa var. laurifólia L. 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Candolleodendron sp. R.S. Cowan 0,2 0,0 0,2 0,0 0,2 0,0 0,4 0,0 0,3 0,0 0,3 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,0
Carapa guianensis Aubl. 0,5 0,2 0,3 0,3 0,4 0,2 0,2 0,8 0,1 0,6 0,1 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,3
Caryocar villosum (Aubl.) Pers. 0,0 0,2 0,0 0,1 0,0 0,2 0,2 0,8 0,1 0,6 0,1 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,3
Caryocar villosum (Aubl.) Pers. 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,8 0,0 0,6 0,0 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2
Cecropia sciadophylla Mart. 0,5 0,0 0,3 0,0 0,4 0,0 0,2 1,5 0,1 1,3 0,1 1,4 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,5
Conceveiba guianensis Aubl. 0,5 0,0 0,3 0,0 0,4 0,0 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,0
Couepia excelsa Ducke 1,2 0,9 0,8 0,5 1,0 0,7 1,3 1,5 1,2 1,3 1,3 1,4 2,0 0,0 2,0 0,0 2,0 0,0 1,4 0,7
Couepia guianensis Aubl. 0,5 0,5 0,3 0,3 0,4 0,4 0,4 0,8 0,4 0,6 0,4 0,7 2,0 0,0 2,0 0,0 2,0 0,0 0,9 0,4
Couma guianensis Aubl. 1,2 0,7 1,2 0,4 1,2 0,5 1,3 0,0 0,7 0,0 1,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,7 0,2
Couratari guianensis Aubl. 1,0 0,7 0,6 0,5 0,8 0,6 0,4 0,0 0,2 0,0 0,3 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,4 0,2

86
Croton sp. L. 0,2 0,0 0,2 0,0 0,2 0,0 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,0
Dacryodes nitens Cuatrec. 0,7 1,6 0,6 1,8 0,7 1,7 1,5 0,0 1,3 0,0 1,4 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,7 0,6
Dimorphandra multiflora Ducke 0,2 0,0 0,2 0,0 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,0
Diospyros sp. L. 0,7 0,5 0,6 0,3 0,7 0,4 0,7 0,0 0,4 0,0 0,6 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,4 0,1
Diplotropis purpurea (Rich.) Amshoff. 1,2 0,9 0,8 0,8 1,0 0,9 0,6 0,0 0,3 0,0 0,4 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,5 0,3
Dipteryx odorata (Aubl.) Willd. 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Duguetia stelechantha (Diels) R.E. Fr. 0,2 0,2 0,2 0,1 0,2 0,2 0,0 0,8 0,0 0,6 0,0 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,3
Ecclinusa lanceolata (Mart. e Eichler) Pierre 1,5 0,9 1,1 0,5 1,3 0,7 1,3 0,8 0,9 0,6 1,1 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,8 0,5
Elizabetha bicolor Ducke 0,7 0,7 0,5 0,5 0,6 0,6 0,4 0,0 0,6 0,0 0,5 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,4 0,2
Eperua schomburgkiana Benth. 0,5 0,2 0,3 0,1 0,4 0,2 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,1
Eperua sp. Aubl. 1,0 0,5 0,8 0,3 0,9 0,4 0,4 0,8 0,3 0,6 0,3 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,4 0,4
Eschweilera coriacea (DC.) S.A.Mori 2,4 3,0 3,9 4,7 3,2 3,8 2,6 1,5 4,8 1,3 3,7 1,4 4,1 0,0 4,1 0,0 4,1 0,0 3,7 1,7
Eschweilera sp. Mart. ex DC. 2,7 2,3 4,1 2,2 3,4 2,3 2,2 3,1 2,2 2,5 2,2 2,8 2,0 0,0 2,0 0,0 2,0 0,0 2,5 1,7
Eugenia diplocampta Diels 0,5 0,5 0,5 0,7 0,5 0,6 0,2 0,8 0,1 0,6 0,1 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,4
Eugenia sp. L. 2,4 2,1 5,3 2,7 3,8 2,4 1,8 1,5 3,2 1,9 2,5 1,7 4,1 0,0 4,1 0,0 4,1 0,0 3,5 1,4
Geissospermum vellosii Allemão 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Genipa americana L. 1,0 0,9 1,2 0,9 1,1 0,9 1,1 0,8 0,9 0,6 1,0 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,7 0,5
Goupia glabra Aubl. 0,5 0,2 0,3 0,1 0,4 0,2 0,2 3,1 0,3 3,8 0,2 3,4 0,0 0,0 0,0 0,0 0,0 0,0 0,2 1,2
Guarea carinata Ducke 1,5 2,1 1,1 1,7 1,3 1,9 2,2 0,0 2,6 0,0 2,4 0,0 0,0 0,0 0,0 0,0 0,0 0,0 1,2 0,6
Guatteria procera R.E. Fr. 0,2 0,2 0,2 0,1 0,2 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,1
Guatteria sp. Ruiz e Pav. 2,2 2,8 3,2 3,8 2,7 3,3 2,8 5,3 3,7 5,7 3,2 5,5 2,0 0,0 2,0 0,0 2,0 0,0 2,6 2,9
Gustavia augusta L. 1,2 0,9 0,9 0,8 1,1 0,9 0,7 0,8 0,5 0,6 0,6 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,6 0,5
Helicostylis sp. Trécul 1,7 1,6 1,5 2,2 1,6 1,9 0,9 2,3 0,9 1,9 0,9 2,1 4,1 0,0 4,1 0,0 4,1 0,0 2,2 1,3
Hevea brasiliensis Müll. Arg. 0,0 0,2 0,0 0,1 0,0 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1
Hirtella sp. L. 3,4 3,2 5,3 6,8 4,3 5,0 2,8 3,1 4,2 5,1 3,5 4,1 0,0 0,0 0,0 0,0 0,0 0,0 2,6 3,0
Hymenaea parvifolia Huber 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 2,0 0,0 2,0 0,0 2,0 0,0 0,7 0,0
Indeterminada 1 0,5 1,4 0,3 0,8 0,4 1,1 0,6 1,5 0,3 1,3 0,4 1,4 0,0 0,0 0,0 0,0 0,0 0,0 0,3 0,8
Indeterminada 10 0,5 0,2 0,3 0,3 0,4 0,2 0,4 0,0 0,2 0,0 0,3 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,1

87
Indeterminada 11 0,0 0,2 0,0 0,1 0,0 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1
Indeterminada 13 0,5 0,2 0,3 0,1 0,4 0,2 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,1
Indeterminada 3 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Indeterminada 5 0,0 0,2 0,0 0,3 0,0 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1
Inga auristellae Harms 1,5 1,4 1,8 1,6 1,6 1,5 2,0 1,5 2,8 1,3 2,4 1,4 2,0 0,0 2,0 0,0 2,0 0,0 2,0 1,0
Inga paraensis Ducke 1,0 1,8 0,6 1,2 0,8 1,5 1,7 0,8 1,4 0,6 1,5 0,7 2,0 0,0 2,0 0,0 2,0 0,0 1,5 0,7
Inga sp. 1 Mill. 1,0 1,2 1,1 1,4 1,0 1,3 1,5 3,1 1,4 2,5 1,4 2,8 2,0 0,0 2,0 0,0 2,0 0,0 1,5 1,4
Inga sp. 2 Mill. 2,4 1,2 2,4 1,4 2,4 1,3 1,8 3,1 2,0 5,1 1,9 4,1 0,0 25,0 0,0 25,0 0,0 25,0 1,4 10,1
Inga velutina Willd. 0,2 0,0 0,2 0,0 0,2 0,0 0,0 0,8 0,0 0,6 0,0 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,2
Iryanthera grandis Ducke 0,2 0,0 0,3 0,0 0,3 0,0 1,1 0,8 1,0 0,6 1,1 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,4 0,2
Iryanthera paraensis Huber 2,4 1,2 2,3 0,9 2,3 1,0 2,6 1,5 2,7 1,3 2,6 1,4 2,0 0,0 2,0 0,0 2,0 0,0 2,3 0,8
Jacaranda copaia (Aubl.) D. Don 0,0 0,2 0,0 0,1 0,0 0,2 0,4 0,0 0,4 0,0 0,4 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,1
Lacmellea aculeata Ducke 0,0 0,2 0,0 0,1 0,0 0,2 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1
Lecythis lurida (Miers) S.A. Mori 0,5 0,0 0,5 0,0 0,5 0,0 0,7 0,0 0,5 0,0 0,6 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,4 0,0
Licania canescens Benoist 0,5 0,5 0,3 0,3 0,4 0,4 1,1 1,5 1,2 1,9 1,2 1,7 2,0 0,0 2,0 0,0 2,0 0,0 1,2 0,7
Licania octandra (Hoffmanns. ex Roem. e Schult.) Kuntze 0,7 0,9 0,5 0,7 0,6 0,8 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,3
Licaria cannella (Meisn.) Kosterm. 0,7 1,2 0,8 1,2 0,7 1,2 1,1 0,0 0,7 0,0 0,9 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,5 0,4
Licaria mahuba (A. Samp.) Kosterm. 0,5 0,5 0,3 0,3 0,4 0,4 0,2 0,8 0,1 0,6 0,1 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,4
Manilkara huberi (Ducke) Chevalier 0,2 0,0 0,2 0,0 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,0
Manilkara sp. Adans. 0,2 0,0 0,2 0,0 0,2 0,0 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,0
Maquira sclerophylla (Ducke) C. C.Berg 1,0 1,8 0,8 1,4 0,9 1,6 2,0 0,0 1,8 0,0 1,9 0,0 2,0 0,0 2,0 0,0 2,0 0,0 1,6 0,5
Mezilaurus itauba (Meisn.) Taub. ex Mez 1,0 0,9 0,8 0,7 0,9 0,8 1,5 0,8 1,8 0,6 1,6 0,7 2,0 0,0 2,0 0,0 2,0 0,0 1,5 0,5
Miconia pubipetala Miq. 1,0 0,0 0,6 0,0 0,8 0,0 0,4 0,0 0,2 0,0 0,3 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,4 0,0
Miconia surinamensis Gleason 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Micropholis acutangula (Ducke) Eyma 2,2 2,5 2,1 3,0 2,1 2,8 2,0 3,1 2,6 3,2 2,3 3,1 4,1 25,0 4,1 25,0 4,1 25,0 2,8 10,3
Micropholis longipedicellata Aubrév. 1,0 0,5 0,8 0,3 0,9 0,4 1,3 0,8 1,3 0,6 1,3 0,7 2,0 0,0 2,0 0,0 2,0 0,0 1,4 0,4
Micropholis venulosa (Mart. e Eichler) Pierre 0,5 0,0 0,3 0,0 0,4 0,0 0,4 0,8 0,2 0,6 0,3 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,2
Minquartia guianensis Aubl. 0,0 0,2 0,0 0,1 0,0 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1

88
Mouriri apiranga Spruce ex Triana 2,9 3,0 4,2 6,5 3,6 4,8 2,4 2,3 2,8 3,8 2,6 3,0 4,1 0,0 4,1 0,0 4,1 0,0 3,4 2,6
Mouriri nervosa Pilg. 0,5 0,2 0,6 0,1 0,5 0,2 0,6 0,0 0,4 0,0 0,5 0,0 2,0 0,0 2,0 0,0 2,0 0,0 1,0 0,1
Myrcia fallax (Rich.) DC. 1,9 3,2 1,8 4,7 1,9 4,0 1,7 2,3 1,1 3,2 1,4 2,7 4,1 0,0 4,1 0,0 4,1 0,0 2,5 2,2
Nectandra rubra (Mez) C. K. Allen 0,7 0,9 0,8 0,5 0,7 0,7 0,4 0,0 0,2 0,0 0,3 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,3 0,2
Neoxythece elegans (A DC.) Aubrev. 0,5 0,7 0,3 0,5 0,4 0,6 0,9 1,5 0,7 1,3 0,8 1,4 0,0 0,0 0,0 0,0 0,0 0,0 0,4 0,7
Ocotea aciphylla (Nees) Mez 0,2 0,5 0,2 0,3 0,2 0,4 0,4 0,0 0,5 0,0 0,4 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,1
Ocotea amazonica (Meisn.) Mez 1,0 1,6 1,2 1,2 1,1 1,4 1,7 2,3 2,0 1,9 1,8 2,1 0,0 0,0 0,0 0,0 0,0 0,0 1,0 1,2
Ocotea sp.1 Aubl. 1,9 2,5 2,0 3,6 2,0 3,1 1,5 1,5 1,5 1,3 1,5 1,4 2,0 0,0 2,0 0,0 2,0 0,0 1,8 1,5
Ocotea sp.2 Aubl. 0,0 0,2 0,0 0,1 0,0 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1
Osteophloeum platyspermum (Spruce ex A. DC.) Warb. 1,0 1,4 0,9 0,8 0,9 1,1 1,3 1,5 1,2 1,9 1,3 1,7 0,0 0,0 0,0 0,0 0,0 0,0 0,7 0,9
Oxandra acuminata Diels 0,0 0,0 0,0 0,0 0,0 0,0 0,4 0,8 0,2 0,6 0,3 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,2
Pachira sp. Aubl. 0,0 0,5 0,0 0,3 0,0 0,4 0,4 0,0 0,2 0,0 0,3 0,0 2,0 0,0 2,0 0,0 2,0 0,0 0,8 0,1
Parinarium Juss. 0,0 0,2 0,0 0,1 0,0 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1
Parkia gigantocarpa Ducke 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,8 0,0 0,6 0,0 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2
Pentaclethra macroloba (Willd.) Kuntze 0,5 0,0 0,3 0,0 0,4 0,0 0,6 0,8 0,6 0,6 0,6 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,3 0,2
Poecilanthe effusa (Huber) Ducke 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Pourouma minor Benoist 1,0 0,5 0,6 0,3 0,8 0,4 0,6 1,5 0,3 1,9 0,4 1,7 4,1 0,0 4,1 0,0 4,1 0,0 1,8 0,7
Pourouma ovata Trécul 1,5 1,6 2,0 2,2 1,7 1,9 1,5 6,1 2,7 6,3 2,1 6,2 4,1 0,0 4,1 0,0 4,1 0,0 2,6 2,7
Pourouma villosa Trécul 0,0 0,2 0,0 0,1 0,0 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1
Pouteria caimito (Ruiz e Pav.) Radlk. 2,2 1,2 1,8 1,2 2,0 1,2 1,3 0,8 0,9 0,6 1,1 0,7 0,0 0,0 0,0 0,0 0,0 0,0 1,0 0,6
Pouteria sp.1Aubl. 1,7 1,6 1,4 1,6 1,5 1,6 1,8 1,5 1,4 1,3 1,6 1,4 6,1 0,0 6,1 0,0 6,1 0,0 3,1 1,0
Pouteria sp.2 Aubl. 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Protium decandrum (Aubl.) Marchand 0,7 1,2 0,6 0,7 0,7 0,9 2,0 0,8 2,0 0,6 2,0 0,7 2,0 0,0 2,0 0,0 2,0 0,0 1,6 0,5
Protium pernervatum Cuatrec. 0,2 0,5 0,2 0,3 0,2 0,4 0,2 0,8 0,1 0,6 0,1 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,4
Protium pilosum (Cuatrec.)D.C. Daly 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,8 0,0 0,6 0,0 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2
Protium sagotianum Marchand. 0,0 0,2 0,0 0,1 0,0 0,2 0,2 0,0 0,1 0,0 0,1 0,0 0,0 25,0 0,0 25,0 0,0 25,0 0,0 8,4
Protium sp. Burm. f. 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Protium tenuifolium (Engl.) Engl. 2,2 2,8 3,0 3,6 2,6 3,2 2,8 1,5 5,2 1,9 4,0 1,7 10,2 25,0 10,2 25,0 10,2 25,0 5,6 10,0

89
Pterocarpus rohrii Vahl 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Qualea albiflora Varm. 0,0 0,2 0,0 0,1 0,0 0,2 0,6 0,8 0,4 0,6 0,5 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,3
Quararibea sp. Aubl. 1,5 0,9 1,1 0,8 1,3 0,9 0,7 1,5 0,9 1,3 0,8 1,4 0,0 0,0 0,0 0,0 0,0 0,0 0,7 0,7
Rheedia benthamiana Planch. e Triana 0,7 0,0 0,5 0,0 0,6 0,0 0,4 0,0 0,2 0,0 0,3 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,3 0,0
Rinorea lindeniana (Tul.) Kuntze 0,5 0,5 0,3 0,5 0,4 0,5 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,2
Rinorea racemosa (Mart.) Kuntze 0,0 0,2 0,0 0,3 0,0 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1
Rinorea sp. Aubl. 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,8 0,0 0,6 0,0 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2
Sacoglottis guianensis Benth. 1,7 1,6 2,6 1,2 2,1 1,4 1,1 2,3 2,0 1,9 1,5 2,1 2,0 0,0 2,0 0,0 2,0 0,0 1,9 1,2
Schizolobium amazonicum Huber ex Ducke 0,0 0,2 0,0 0,1 0,0 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1
Sclerolobium melanocarpum Ducke 1,2 1,2 0,8 0,9 1,0 1,0 1,5 0,8 1,3 0,6 1,4 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,8 0,6
Sclerolobium melinonii Harms 0,2 0,0 0,2 0,0 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,0
Simarouba amara Aubl. 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Siparuna sp. Aubl. 0,0 0,5 0,0 0,3 0,0 0,4 0,4 0,0 0,2 0,0 0,3 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,1
Sloanea sp. L. 0,5 0,0 0,3 0,0 0,4 0,0 0,6 0,0 0,3 0,0 0,4 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,3 0,0
Sterculia pilosa Ducke 0,2 0,9 0,3 0,9 0,3 0,9 0,9 1,5 0,8 1,3 0,8 1,4 0,0 0,0 0,0 0,0 0,0 0,0 0,4 0,8
Stryphnodendron pulcherrimum (Willd.) Hochr. 0,2 0,2 0,2 0,1 0,2 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,1
Swartzia racemosa Benth. 0,0 0,2 0,0 0,1 0,0 0,2 0,4 0,0 0,2 0,0 0,3 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,1
Swartzia sp. Schreb. 0,2 0,0 0,3 0,0 0,3 0,0 0,2 0,8 0,1 0,6 0,1 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,1 0,2
Symphonia globulifera L. f. 0,5 0,5 0,3 0,4 0,4 0,4 0,6 1,5 0,3 1,3 0,4 1,4 0,0 0,0 0,0 0,0 0,0 0,0 0,3 0,6
Tachigali myrmecophila Ducke 1,2 1,4 0,8 1,0 1,0 1,2 1,5 1,5 0,9 1,3 1,2 1,4 6,1 0,0 6,1 0,0 6,1 0,0 2,8 0,9
Tachigali sp. Aubl. 0,0 2,1 0,0 1,4 0,0 1,8 0,9 0,0 0,8 0,0 0,8 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,3 0,6
Tapirira guianensis Aubl. 1,5 0,9 0,9 0,9 1,2 0,9 1,5 1,5 2,9 1,9 2,2 1,7 0,0 0,0 0,0 0,0 0,0 0,0 1,1 0,9
Taralea oppositifolia Aubl. 1,2 2,5 1,5 2,1 1,4 2,3 0,9 0,0 0,9 0,0 0,9 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,7 0,8
Terminalia sp. L. 0,0 0,2 0,0 0,1 0,0 0,2 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1
Theobroma subincanum Mart. 1,9 1,8 2,7 1,7 2,3 1,8 1,7 0,8 1,0 0,6 1,4 0,7 2,0 0,0 2,0 0,0 2,0 0,0 1,9 0,8
Tovomita sp.Aubl. 0,7 0,5 0,5 0,4 0,6 0,4 0,6 0,0 0,3 0,0 0,4 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,3 0,1
Trattinnickia rhoifolia Willd. 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Virola calophylla (Spruce) Warb. 0,5 0,2 0,3 0,1 0,4 0,2 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,1

90
Virola michelii Heckel 1,0 0,9 0,9 0,8 0,9 0,9 1,5 1,5 1,1 1,3 1,3 1,4 2,0 0,0 2,0 0,0 2,0 0,0 1,4 0,7
Vismia cayennensis (Jacq.) Pers. 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Vochysia guianensis Aubl. 0,7 0,2 1,1 0,3 0,9 0,2 0,7 0,8 0,5 0,6 0,6 0,7 0,0 0,0 0,0 0,0 0,0 0,0 0,5 0,3
Vochysia maxima Ducke 0,2 0,0 0,2 0,0 0,2 0,0 0,4 0,0 0,3 0,0 0,3 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,2 0,0
Vochysia sp. Aubl. 0,7 0,7 0,6 0,5 0,7 0,6 0,6 0,0 0,5 0,0 0,5 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,4 0,2
Vouacapoua americana Aubl. 0,0 0,2 0,0 0,1 0,0 0,2 0,2 0,0 0,1 0,0 0,1 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,1
Total 100 100 100 100 100 100 100 100 100 100 100 100 100 100 100 100 100 100 100 100

91
Dentre as espécies inventariadas em nível de vara, apenas 31 estavam presentes
em todas as classes de regeneração natural. Quanto ao número de indivíduos nas
mesmas classes foi observada a seguinte distribuição: 55,97% na classe I, 41,84% na
classe II e 2,17 % na classe III.
As espécies em nível de vara predominantes nas três classes de regeneração
natural foram: Mouriri apiranga e Hirtella sp. (classe I); Guatteria sp. e Pourouma
ovata (classe II); e Protium tenuifolium, Micropholis acutangula e Inga sp. 2 (classe
III).
Quanto à regeneração natural total (RNT) na comunidade, as maiores variações
foram encontradas para as espécies em nível de vara (10,3 a 0,1 %), enquanto que as
arvoretas pouco variaram (5,6 a 0,1 %). O número de espécies com percentuais
inferiores a 1 % é elevado, com 121 e 103 espécies para varas e arvoretas,
respectivamente, podendo-se citar as Indeterminadas, Vouacapoua americana, Carapa
guianensis, Virola calophylla e Manilkara huberi.
Para a similaridade florística entre as arvoretas e varas nos distintos
conglomerados adotou-se como limite de significância a linha de fenon a
aproximadamente 50 % da distância de ligação (6,75). Neste limite houve a
interceptação em três ramos do agrupamento, resultando em três grupos florísticos
distintos (Figura 8).
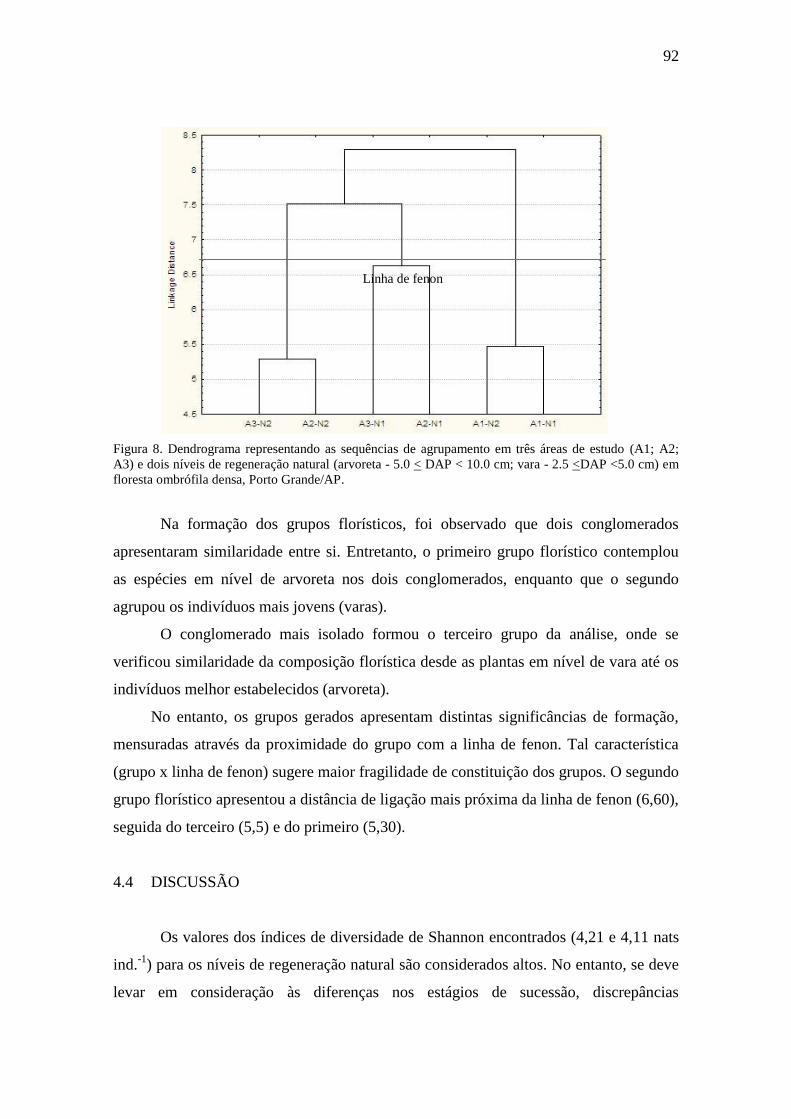
92
Figura 8. Dendrograma representando as sequências de agrupamento em três áreas de estudo (A1; A2;
A3) e dois níveis de regeneração natural (arvoreta - 5.0 < DAP < 10.0 cm; vara - 2.5 <DAP <5.0 cm) em
floresta ombrófila densa, Porto Grande/AP.
Na formação dos grupos florísticos, foi observado que dois conglomerados
apresentaram similaridade entre si. Entretanto, o primeiro grupo florístico contemplou
as espécies em nível de arvoreta nos dois conglomerados, enquanto que o segundo
agrupou os indivíduos mais jovens (varas).
O conglomerado mais isolado formou o terceiro grupo da análise, onde se
verificou similaridade da composição florística desde as plantas em nível de vara até os
indivíduos melhor estabelecidos (arvoreta).
No entanto, os grupos gerados apresentam distintas significâncias de formação,
mensuradas através da proximidade do grupo com a linha de fenon. Tal característica
(grupo x linha de fenon) sugere maior fragilidade de constituição dos grupos. O segundo
grupo florístico apresentou a distância de ligação mais próxima da linha de fenon (6,60),
seguida do terceiro (5,5) e do primeiro (5,30).
4.4 DISCUSSÃO
Os valores dos índices de diversidade de Shannon encontrados (4,21 e 4,11 nats
ind.-1
) para os níveis de regeneração natural são considerados altos. No entanto, se deve
levar em consideração às diferenças nos estágios de sucessão, discrepâncias
Linha de fenon

93
metodológicas e identificações taxonômicas, as quais variam nos levantamentos
florísticos da região. Esta afirmação é baseada na variabilidade das diversidades
usualmente encontradas em florestas tropicais (entre 1,5 e 3,5 nats.ind.-1
), onde
raramente ultrapassam 4,5 (DIAS et al.,2000; SILVA et al., 2007; MARANGON et al.,
2003). Os fatores que normalmente contribuem para o aumento da diversidade são a
existência de elevado número de plântulas, brotação de cepas e banco de sementes de
variadas espécies sob distintas condições edafoclimáticas.
Em florestas ombrófila densa, Aparício et al. (2011) analisando a regeneração
natural de espécies arbóreas tropicais obtiveram diversidade de Shannon de 3,32 nats
ind.-1
. Resultados semelhantes foram expostos em florestas secundárias no Pará, na
avaliação momentânea da composição florística em regeneração (3,5 nats ind.-1
)
(RAYOL et al., 2008).
As espécies inventariadas com maior número de indivíduos são consideradas
mais aptas a se estabelecer às condições impostas pelo meio. Estas mantêm uma
proporção de plantas entre os níveis de vara para arvoreta. No entanto foi observado que
os parâmetros de freqüência e densidade das espécies variaram em função do nível de
regeneração. Acredita-se que a variação pode estar relacionada ao tamanho das classes
de altura que foram pré-estabelecidas para o estudo e ao estágio sucessional em que se
encontra a área.
As espécies que ocorrem nas três classes de altura de regeneração natural são
aquelas que teoricamente possuem um maior potencial de estabelecimento na floresta e
que deverão estar presentes no futuro dossel (SILVA et al., 2007; CITADINI-
ZANETTE, 1995).
Entretanto, algumas espécies que apresentaram maiores índices de regeneração
natural em nível de vara, não foram observadas nas arvoretas, como o Protium
sagotianum, que quando jovem foi classificado entre as espécies mais adaptadas, mas
poucos indivíduos foram registrados na classe de tamanho imediatamente superior.
Além disso, houveram espécies encontradas apenas em uma classe de tamanho,
apresentando uma regeneração natural baixa. Tal fato indica dificuldades no processo de
estabelecimento, possivelmente ocasionado pela seletividade a que estão expostas ou
pela dinâmica de germinação do banco de sementes. Como destaque, pode-se citar a
Vouacapoua americana, que ocorreu apenas na classe I (tanto para varas como
arvoretas) com densidade relativa elevada.

94
A presença de uma espécie apenas na classe de menor altura (RNC1), com
densidade elevada, pode apresentar valores de regeneração natural total (RNT) maior do
que a de outras espécies presentes em todas as classes de altura (SILVA JUNIOR et al.,
2008). Porém, de acordo com Volpato (1994), esse resultado deve ser analisado com
cautela, já que a espécie pode desaparecer ainda nessa fase inicial de desenvolvimento.
Os baixos valores estimados de regeneração para algumas espécies na área de
estudo também podem ser atribuídos a grande diversidade da área, a exposição da
vegetação à declividade do terreno, solo, grupo ecológico e principalmente ao fato da
área ter poucos indícios antrópicos.
Em relação à similaridade florística, foi observada que as condições do estado de
conservação das áreas são semelhantes, fortalecendo a permanência de específicos
exemplares florísticos em cada fase da vegetação. No entanto, merece destaque o
terceiro grupo florístico, formado pelo agrupamento de varas e arvoretas numa mesma
área, demonstrando que o processo de sucessão está ocorrendo com perpetuação das
espécies com maior índice de regeneração.
4.5 CONCLUSÕES
As espécies arbóreas em regeneração demonstraram potencial para a auto-
recuperação, com diversidade elevada nos níveis estudados e número de indivíduos
capazes de compor o futuro dossel da floresta.
Estima-se que a estrutura e as relações entre os estratos verticais das espécies
arbóreas em regeneração natural são capazes de desenvolver o processo sucessional de
forma eficiente, garantindo a conservação da fitofisionomia da região.

95
5 DINÂMICA DO CRESCIMENTO DE ESPÉCIES ARBÓREAS
EM FLORESTA OMBRÓFILA DENSA NO AMAPÁ

96
RESUMO
Este capítulo teve como objetivo realizar uma avaliação inicial da dinâmica das espécies
arbóreas numa floresta ombrófila densa destinada ao manejo florestal no estado do
Amapá.As parcelas foram implantadas no segundo módulo da Floresta Estadual do
Amapá, através da amostragem em conglomerados. Cada unidade primária foi disposta
em forma quadrada com cinco parcelas permanentes de 100x100 m (árvores adultas
com DAP’s (Diâmetro a altura do peito a 1,30 m do solo) > 10cm), abrangendo uma
área de 5 ha/UP. Cada parcela foi subdividida em 100 subparcelas de 10x10 m. Destas
foram sorteadas apenas 20 subparcelas para o estudo da regeneração natural em arvoreta
(5 < DAP < 10cm). Nas subparcelas sorteadas também foram implantadas,
aleatoriamente nos vértices, subunidades de 5x5m para inventário da regeneração
natural em vara (2,5 < DAP < 5cm). A identificação taxonômica foi realizada com um
levantamento prévio em campo por especialistas e os nomes botânicos foram conferidos
no Missouri Botanical Garden. O monitoramento da vegetação ocorreu entre os anos de
2010 e 2012. Foi realizado o teste de esfericidade de Mauchly para justificar a análise
multivariada de medidas repetidas. O crescimento das espécies foi avaliado a partir dos
incrementos periódicos anuais percentuais em diâmetro, taxas de recrutamento e
mortalidade. Para interpretação das mudanças estruturais ocorridas no tempo de
monitoramento na floresta utilizou-secomo medida de dispersão em torno da média o
desvio padrão.Quanto à dinâmica florestal, ocorreu a perda de aproximadamente 3 %
dos dados obtidos em campo, gerando um total de 10.209 plantas vivas em 2010, com
aumento de 0.46 % em 2011 e baixa de 0.90 % em 2012, em relação ao inicio do
monitoramento. A estimativa total por hectare de varas, arvoretas e árvores foi de 2.303
indivíduos. O número de indivíduos e o crescimento diamétrico das espécies foi
influenciado pelo fator tempo. O inventário demonstrou que nove espécies contêm taxas
de recrutamento contínua nas seis classes diamétricas iniciais, com variação de 0,02 a
0,87 %: Eschweilera coriacea, Osteophloeum platyspermum, Pourouma ovata, Eugenia
sp., Iryanthera paraensis, Micropholis acutangula, Micropholis longipedicellata,
Pouteria sp.1 e Sclerolobium melanocarpum. Os maiores índices de mortalidade foram
observados nas classes iniciais, com exceção da classe com diâmetros acima de 110 cm
(Mezilaurus itauba - 3.7 %). Quanto aos incrementos periódicos médios durante os três
anos de monitoramento, os valores foram decrescendo à medida que o tamanho das
árvores aumentava. A superioridade das taxas de ingresso em relação à mortalidade
indica que a floresta se apresenta num estágio sucessional de constante renovação.
Baseado nesta renovação acredita-se que a floresta é capaz de ser submetida ao manejo,
sem comprometer a estrutura da comunidade.
Palavras-chave: Amazônia, Mortalidade, Recrutamento

97
ABSTRACT
This chapter aims to perform an initial assessment of dynamics of tree species in
tropical rain forest allocated to forest management in Amapa. Plots were established in
second module of Amapa State Forest, through cluster sampling. Each primary unit was
arranged with five plots of 100x100 m (trees with DBH (diameter at breast height to
1.30m) > 10cm). Each plot was divided into 100 plots of 10x10 m. These only 20 were
selected for study of regeneration (5 < DBH < 10cm). Subunits were also implanted
(5x5m) for inventory other (2.5 < DBH < 5cm). Taxonomic identification was realized
by Missouri Botanical Garden. Vegetation monitoring occurred between 2010 and 2012.
Mauchly sphericity test was performed to justify the multivariate repeated measures.
The growth of species was evaluated from annual percentage increases in diameter,
recruitment and mortalidade rates. To interpretation of structural changes in the
monitoring time forest used measure of dispersion around the mean. As for forest
dynamics, the loss occurred in approximately 3% of data obtained, generating a total of
10,209 plants in 2010, an increase of 0.46% in 2011 and low of 0.90% in 2012,
compared to beginning of monitoring. Estimate per hectare of sticks, saplings and trees
was 2303 individuals. The number of individuals of species and diameter growth was
influenced by time factor. The survey showed that nine species contain continuous
recruitment rates in six initial diameter classes (0.02 to 0.87%): Eschweilera coriacea,
Osteophloeum platyspermum, Pourouma ovata, Eugenia sp., Iryanthera paraensis
Micropholis acutangula, Micropholis longipedicellata , Pouteria sp.1 and Sclerolobium
melanocarpum. The highest mortality rates were observed in initial classes, except class
with diameters above 110 cm (Mezilaurus itauba - 3.7%). As for periodic increments
average during three years of monitoring, the values were decreasing as the size of the
trees grew. The superiority of recruitment rates in relation to mortality indicates that
forest has a successional stage of constant renewal. Based on this renovation it is
believed that forest can be subjected to handling, without compromising the community
structure.
Key words: Amazon, Mortality, Recruitment

98
5.1 INTRODUÇÃO
O planejamento do manejo sustentável nas florestas tropicais deve ser subsidiado
a partir da utilização da projeção das espécies nos sítios de produção. A projeção do
crescimento da floresta se constitui no estudo da dinâmica, seja de árvores adultas ou
em regeneração. Segundo Mendonça (2003) a partir do crescimento também é possível
mensurar a capacidade da floresta de gerar bens e serviços.
Esforços têm sido realizados em pesquisas que abrangem a dinâmica de espécies
arbóreas de valor comercial madeireiro, visando fundamentar a adoção de práticas
silviculturais de manejo. Tais práticas contribuem para acelerar as taxas de crescimento
e diminuir a competição no povoamento (AZEVEDO et al., 2008), pois o número de
árvores exploradas influência no aumento do crescimento da floresta (FONSECA et al.,
2008).
Atualmente a taxa de crescimento é definida na Resolução CONAMA 406/2009
para espécies comerciais, quando não houver estudos para a área, estimativa da
produtividade anual de 0,86 m³/ha/ano. No entanto, estudos têm apontado taxas de
crescimento diferentes por espécies (OLIVEIRA et al., 2005; TEIXEIRA et al., 2007;
COLPINI et al., 2010), sugerindo a utilização de ciclos específicos por grupos de
espécies, de acordo com suas características ecológicas ou de interesse comercial.
Segundo Teixeira et al. (2007), a partir das taxas de crescimento, recrutamento e
mortalidade são definidos os ciclos de corte.
Do ponto de vista ambiental, nenhuma floresta deveria ser destinada a produção,
antes de sofrer estudos ecológicos, especialmente dinâmicos, para conhecer o
crescimento das espécies e a taxa de rendimento sustentado, a fim de estabelecer o ciclo
de corte e o volume a ser cortado periodicamente. (SCHNEIDER e FINGER, 2000;
VACCARO et al., 2003).
Desta forma, entender como ocorre à dinâmica florestal em sítios da Amazônia é
fundamental para gerar informações que subsidiem manejos distintos por espécie e área.
Nesses termos, para gerar prognósticos temporais da estrutura da floresta devem ser
implantadas parcelas permanentes, as quais, de acordo com Phillips e Baker (2006), são
essenciais para predição e rendimentos no manejo florestal.
No caso do estado do Amapá, pretende-se averiguar se a utilização dos
principais indicadores dinâmicos das florestas pode auxiliar nas práticas de

99
sustentabilidade do manejo. Para responder este questionamento parte-se do pressuposto
que os indivíduos apresentam crescimento similar aos catalogados em outras áreas
tropicais submetidas ou não a intervenções antrópicas.
Logo, o objetivo deste trabalho é realizar uma avaliação inicial da dinâmica das
espécies arbóreas numa floresta ombrófila densa destinada ao manejo florestal no estado
do Amapá.
5.2 MATERIAL E MÉTODOS
O trabalho está localizado num perímetro de floresta ombrófila densa de terras
baixas (IBGE, 2012) numa área de 500 ha ao sul da Floresta Estadual do Amapá
(FLOTA/AP), região central do estado, Amazônia, Brasil.
Os solos são do tipo Latossolo vermelho-amarelo, ligado a variações amplas de
relevo, com destaque para classe ondulada (ZEE/AP, 2008). O clima da região é do tipo
Am (equatorial super-úmido), segundo a classificação de Köppen. A temperatura média
é 25 ºC, ocorrendo pouca variação de temperatura durante o ano. A precipitação anual
na região é de 2.300 mm, com índice pluviométrico trimestral mais seco abaixo de 200
mm. O período mais chuvoso ocorre no trimestre de março a maio (acima de 1.000
mm). A umidade relativa é alta, com valores entre 80 e 86% (TARDIN et al., 2009).
O inventário foi realizado em unidades amostrais conglomeradas, distribuídas
em três distintas áreas (unidades primárias - UP), equidistantes em aproximadamente
2.500 m. Cada unidade primária foi disposta em forma quadrada com cinco parcelas
permanentes de 100 x 100 m, abrangendo uma área de 5 ha / UP com total de 15 ha
amostrados.
As parcelas foram estabelecidas de acordo com os critérios da “Rede de
Monitoramento da Dinâmica das Florestas da Amazônia”, com três níveis de inclusão:
varas (2,5 < DAP (Diâmetro a altura do peito a 1,30 m do solo) < 5,0 cm); arvoretas
(5,0 < DAP < 10,0 cm); e árvores (DAP > 10,0 cm) (SILVA et al., 2005).
Para inventário das árvores foram utilizadas como unidades amostrais as
parcelas de 100 x 100 m. Quanto às arvoretas, cada parcela foi dividida em 100
subparcelas de 10 x 10 m, com posterior seleção aleatória de 20 subparcelas / parcela
para avaliação. Nos vértices das subparcelas sorteadas foram locadas, aleatoriamente, as
subunidades de 5 x 5 m para inventário das varas.

100
Todos os indivíduos foram etiquetados com numeração progressiva,
identificados pelo nome popular e medidos os DAP´s. A identificação taxonômica foi
de acordo com o sistema APG III (2010) Todas as espécies identificadas foram
separadas em grupos de valor madeireiro (GVM), onde: 0 – menor valor comercial; 1 –
alto valor comercial; 2 – reconhecido valor comercial; 3 – comumente comercializadas;
4 – comercialização local; 5 – não classificadas.
Para interpretação das mudanças estruturais ocorridas na floresta utilizou-se
como referência as variações diamétricas das espécies em três ocasiões de coleta (2010
2011 e 2012). Foram consideradas dez classes diamétricas com amplitude de 10 cm,
com exceção das classes 1 e 2 as quais foram mantidas as amplitudes dos níveis de
inclusão de vara e arvoreta.
As espécies presentes em todos os níveis de inclusão e UP’s formaram um
conjunto de dados submetidos ao teste de esfericidade de Mauchly, com intuito de
justificar a análise multivariada de medidas repetidas (NEMEC, 1996). A análise de
medidas repetidas subsequentemente foi empregada para avaliar o quanto o fator tempo
influenciou o número de indivíduos e o crescimento em diâmetro das espécies.
Para facilitar a interpretação do crescimento em diâmetro das espécies nas
classes diamétricas foi utilizada como medida de dispersão em torno da média o desvio
padrão, captando a variabilidade no desenvolvimento dos indivíduos.
O crescimento das espécies foi avaliado a partir dos incrementos periódicos
anuais percentuais em diâmetro, taxas de recrutamento e mortalidade, para todas as
espécies e nas classes de diâmetro.
Para as estimativas de taxas de recrutamento das árvores, arvoreta e varas, foi
considerado como ingresso o indivíduo vivo que inicialmente se localizava na i-ésima
classe diamétrica e ao final do monitoramento migrou para classe imediatamente
posterior ou não.
Quanto à mortalidade, foi considerado como morto o indivíduo vivo no primeiro
ano de monitoramento e que estava morto no ano final de monitoramento. A partir das
estimativas do número de indivíduos mortos estimou-se a taxa de mortalidade. Todas as
análises foram realizadas com auxílio do Microsoft Excel 2007 e STATISTICA TRIAL.
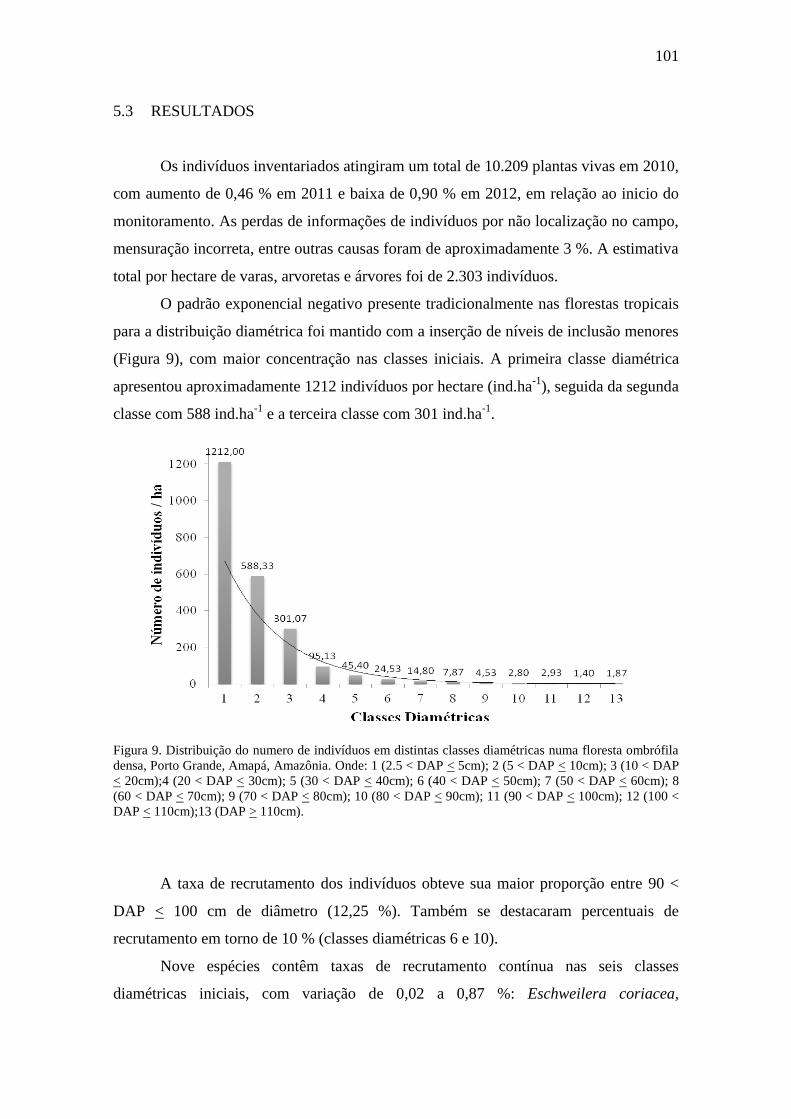
101
5.3 RESULTADOS
Os indivíduos inventariados atingiram um total de 10.209 plantas vivas em 2010,
com aumento de 0,46 % em 2011 e baixa de 0,90 % em 2012, em relação ao inicio do
monitoramento. As perdas de informações de indivíduos por não localização no campo,
mensuração incorreta, entre outras causas foram de aproximadamente 3 %. A estimativa
total por hectare de varas, arvoretas e árvores foi de 2.303 indivíduos.
O padrão exponencial negativo presente tradicionalmente nas florestas tropicais
para a distribuição diamétrica foi mantido com a inserção de níveis de inclusão menores
(Figura 9), com maior concentração nas classes iniciais. A primeira classe diamétrica
apresentou aproximadamente 1212 indivíduos por hectare (ind.ha-1
), seguida da segunda
classe com 588 ind.ha-1
e a terceira classe com 301 ind.ha-1
.
Figura 9. Distribuição do numero de indivíduos em distintas classes diamétricas numa floresta ombrófila
densa, Porto Grande, Amapá, Amazônia. Onde: 1 (2.5 < DAP < 5cm); 2 (5 < DAP < 10cm); 3 (10 < DAP
< 20cm);4 (20 < DAP < 30cm); 5 (30 < DAP < 40cm); 6 (40 < DAP < 50cm); 7 (50 < DAP < 60cm); 8
(60 < DAP < 70cm); 9 (70 < DAP < 80cm); 10 (80 < DAP < 90cm); 11 (90 < DAP < 100cm); 12 (100 <
DAP < 110cm);13 (DAP > 110cm).
A taxa de recrutamento dos indivíduos obteve sua maior proporção entre 90 <
DAP < 100 cm de diâmetro (12,25 %). Também se destacaram percentuais de
recrutamento em torno de 10 % (classes diamétricas 6 e 10).
Nove espécies contêm taxas de recrutamento contínua nas seis classes
diamétricas iniciais, com variação de 0,02 a 0,87 %: Eschweilera coriacea,

102
Osteophloeum platyspermum, Pourouma ovata, Eugenia sp., Iryanthera paraensis,
Micropholis acutangula, Micropholis longipedicellata, Pouteria sp,1 e Sclerolobium
melanocarpum.
Os maiores percentuais de recrutamento foram observados para as espécies
presentes na classe 10 (Conceveiba guianensis, Dipteryx odorata e Tachigali
myrmecophila - 2,5 %), 12 (Pouteria sp,1 - 4,76 %) e 13 (Qualea albiflora - 3,57 %)
(Tabela 9).
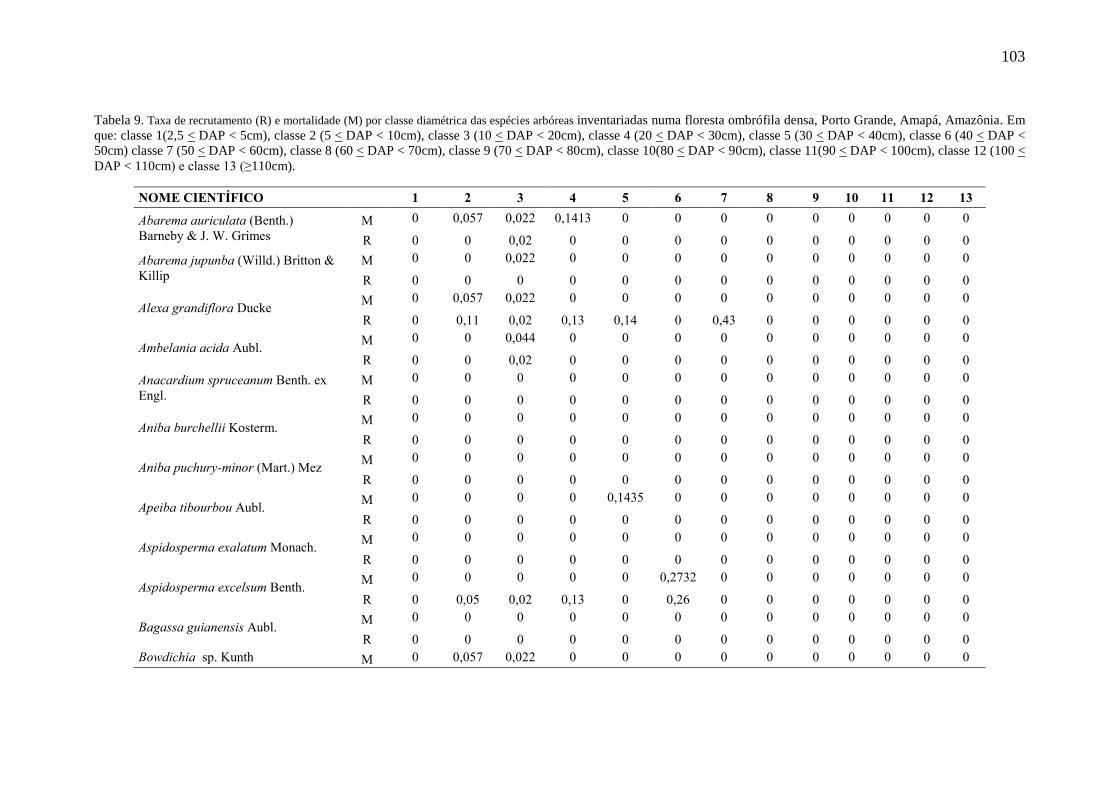
103
Tabela 9. Taxa de recrutamento (R) e mortalidade (M) por classe diamétrica das espécies arbóreas inventariadas numa floresta ombrófila densa, Porto Grande, Amapá, Amazônia. Em
que: classe 1(2,5 < DAP < 5cm), classe 2 (5 < DAP < 10cm), classe 3 (10 < DAP < 20cm), classe 4 (20 < DAP < 30cm), classe 5 (30 < DAP < 40cm), classe 6 (40 < DAP <
50cm) classe 7 (50 < DAP < 60cm), classe 8 (60 < DAP < 70cm), classe 9 (70 < DAP < 80cm), classe 10(80 < DAP < 90cm), classe 11(90 < DAP < 100cm), classe 12 (100 <
DAP < 110cm) e classe 13 (≥110cm).
NOME CIENTÍFICO 1 2 3 4 5 6 7 8 9 10 11 12 13
Abarema auriculata (Benth.)
Barneby & J. W. Grimes
M 0 0,057 0,022 0,1413 0 0 0 0 0 0 0 0 0
R 0 0 0,02 0 0 0 0 0 0 0 0 0 0
Abarema jupunba (Willd.) Britton &
Killip
M 0 0 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Alexa grandiflora Ducke M 0 0,057 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0,11 0,02 0,13 0,14 0 0,43 0 0 0 0 0 0
Ambelania acida Aubl. M 0 0 0,044 0 0 0 0 0 0 0 0 0 0
R 0 0 0,02 0 0 0 0 0 0 0 0 0 0
Anacardium spruceanum Benth. ex
Engl.
M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Aniba burchellii Kosterm. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Aniba puchury-minor (Mart.) Mez M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Apeiba tibourbou Aubl. M 0 0 0 0 0,1435 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Aspidosperma exalatum Monach. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Aspidosperma excelsum Benth. M 0 0 0 0 0 0,2732 0 0 0 0 0 0 0
R 0 0,05 0,02 0,13 0 0,26 0 0 0 0 0 0 0
Bagassa guianensis Aubl. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Bowdichia sp. Kunth M 0 0,057 0,022 0 0 0 0 0 0 0 0 0 0
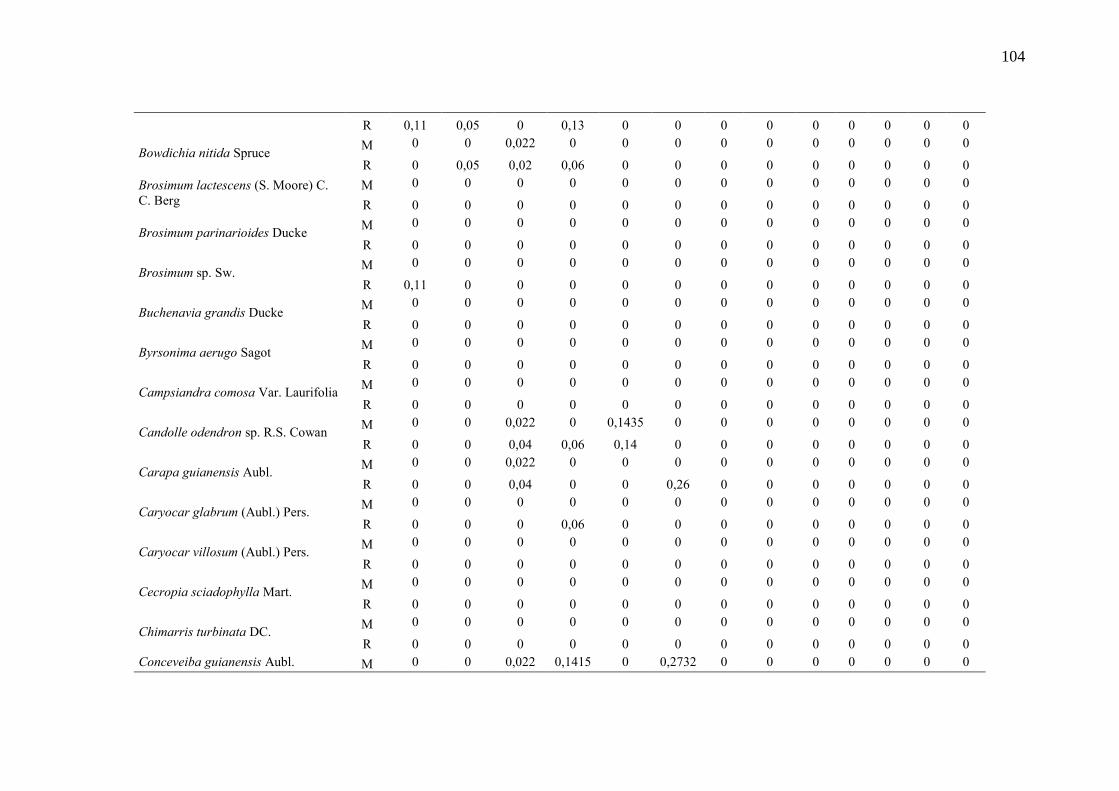
104
R 0,11 0,05 0 0,13 0 0 0 0 0 0 0 0 0
Bowdichia nitida Spruce M 0 0 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0,05 0,02 0,06 0 0 0 0 0 0 0 0 0
Brosimum lactescens (S. Moore) C.
C. Berg
M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Brosimum parinarioides Ducke M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Brosimum sp. Sw. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0,11 0 0 0 0 0 0 0 0 0 0 0 0
Buchenavia grandis Ducke M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Byrsonima aerugo Sagot M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Campsiandra comosa Var. Laurifolia M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Candolle odendron sp. R.S. Cowan M 0 0 0,022 0 0,1435 0 0 0 0 0 0 0 0
R 0 0 0,04 0,06 0,14 0 0 0 0 0 0 0 0
Carapa guianensis Aubl. M 0 0 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0 0,04 0 0 0,26 0 0 0 0 0 0 0
Caryocar glabrum (Aubl.) Pers. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0,06 0 0 0 0 0 0 0 0 0
Caryocar villosum (Aubl.) Pers. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Cecropia sciadophylla Mart. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Chimarris turbinata DC. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Conceveiba guianensis Aubl. M 0 0 0,022 0,1415 0 0,2732 0 0 0 0 0 0 0
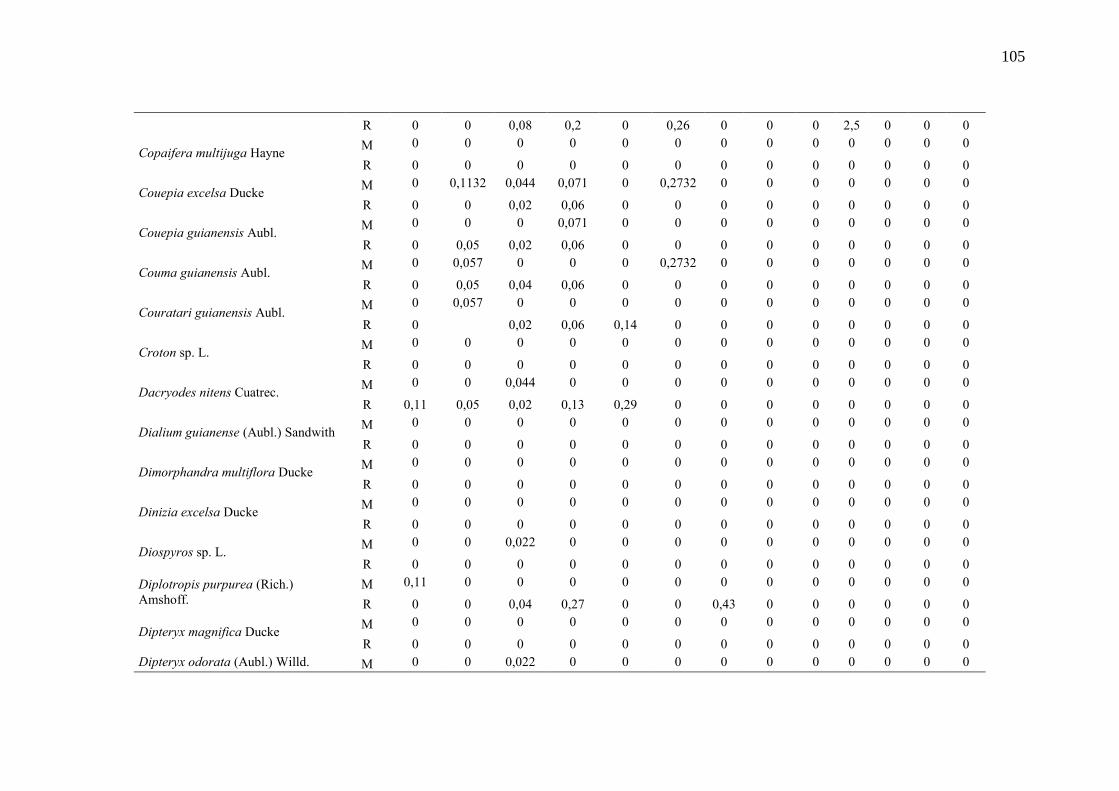
105
R 0 0 0,08 0,2 0 0,26 0 0 0 2,5 0 0 0
Copaifera multijuga Hayne M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Couepia excelsa Ducke M 0 0,1132 0,044 0,071 0 0,2732 0 0 0 0 0 0 0
R 0 0 0,02 0,06 0 0 0 0 0 0 0 0 0
Couepia guianensis Aubl. M 0 0 0 0,071 0 0 0 0 0 0 0 0 0
R 0 0,05 0,02 0,06 0 0 0 0 0 0 0 0 0
Couma guianensis Aubl. M 0 0,057 0 0 0 0,2732 0 0 0 0 0 0 0
R 0 0,05 0,04 0,06 0 0 0 0 0 0 0 0 0
Couratari guianensis Aubl. M 0 0,057 0 0 0 0 0 0 0 0 0 0 0
R 0 0,02 0,06 0,14 0 0 0 0 0 0 0 0
Croton sp. L. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Dacryodes nitens Cuatrec. M 0 0 0,044 0 0 0 0 0 0 0 0 0 0
R 0,11 0,05 0,02 0,13 0,29 0 0 0 0 0 0 0 0
Dialium guianense (Aubl.) Sandwith M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Dimorphandra multiflora Ducke M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Dinizia excelsa Ducke M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Diospyros sp. L. M 0 0 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Diplotropis purpurea (Rich.)
Amshoff.
M 0,11 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0,04 0,27 0 0 0,43 0 0 0 0 0 0
Dipteryx magnifica Ducke M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Dipteryx odorata (Aubl.) Willd. M 0 0 0,022 0 0 0 0 0 0 0 0 0 0
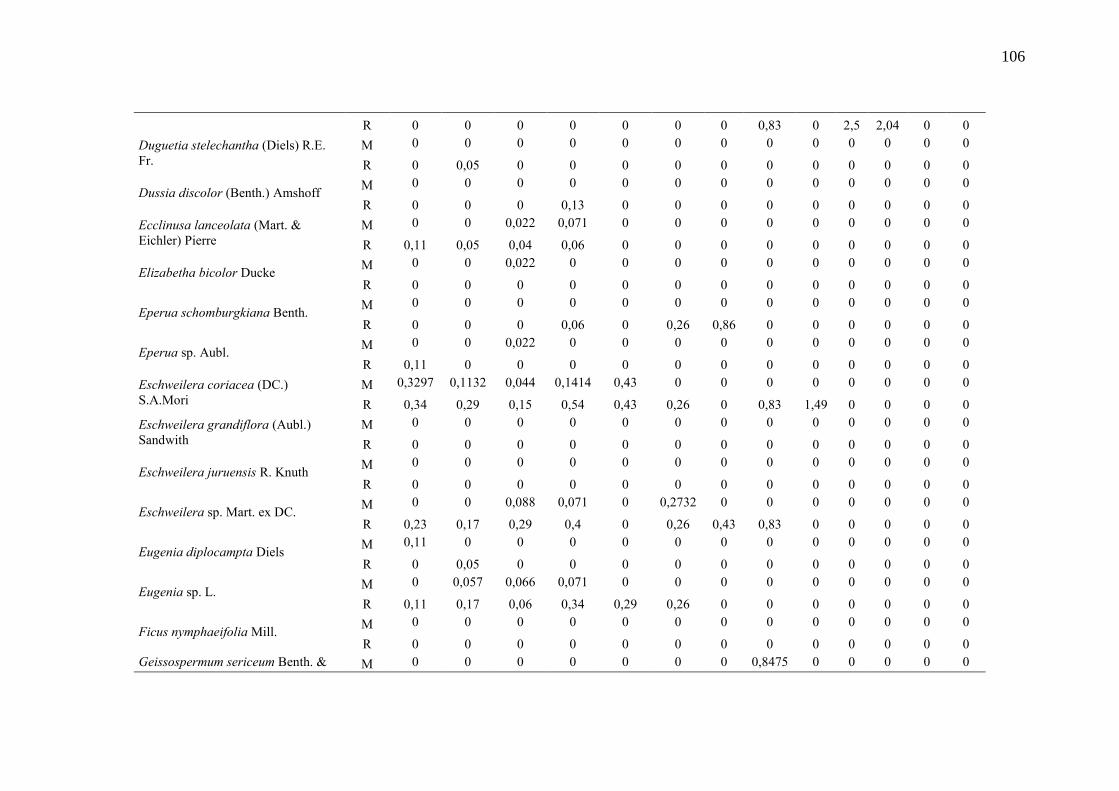
106
R 0 0 0 0 0 0 0 0,83 0 2,5 2,04 0 0
Duguetia stelechantha (Diels) R.E.
Fr.
M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0,05 0 0 0 0 0 0 0 0 0 0 0
Dussia discolor (Benth.) Amshoff M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0,13 0 0 0 0 0 0 0 0 0
Ecclinusa lanceolata (Mart. &
Eichler) Pierre
M 0 0 0,022 0,071 0 0 0 0 0 0 0 0 0
R 0,11 0,05 0,04 0,06 0 0 0 0 0 0 0 0 0
Elizabetha bicolor Ducke M 0 0 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Eperua schomburgkiana Benth. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0,06 0 0,26 0,86 0 0 0 0 0 0
Eperua sp. Aubl. M 0 0 0,022 0 0 0 0 0 0 0 0 0 0
R 0,11 0 0 0 0 0 0 0 0 0 0 0 0
Eschweilera coriacea (DC.)
S.A.Mori
M 0,3297 0,1132 0,044 0,1414 0,43 0 0 0 0 0 0 0 0
R 0,34 0,29 0,15 0,54 0,43 0,26 0 0,83 1,49 0 0 0 0
Eschweilera grandiflora (Aubl.)
Sandwith
M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Eschweilera juruensis R. Knuth M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Eschweilera sp. Mart. ex DC. M 0 0 0,088 0,071 0 0,2732 0 0 0 0 0 0 0
R 0,23 0,17 0,29 0,4 0 0,26 0,43 0,83 0 0 0 0 0
Eugenia diplocampta Diels M 0,11 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0,05 0 0 0 0 0 0 0 0 0 0 0
Eugenia sp. L. M 0 0,057 0,066 0,071 0 0 0 0 0 0 0 0 0
R 0,11 0,17 0,06 0,34 0,29 0,26 0 0 0 0 0 0 0
Ficus nymphaeifolia Mill. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Geissospermum sericeum Benth. & M 0 0 0 0 0 0 0 0,8475 0 0 0 0 0
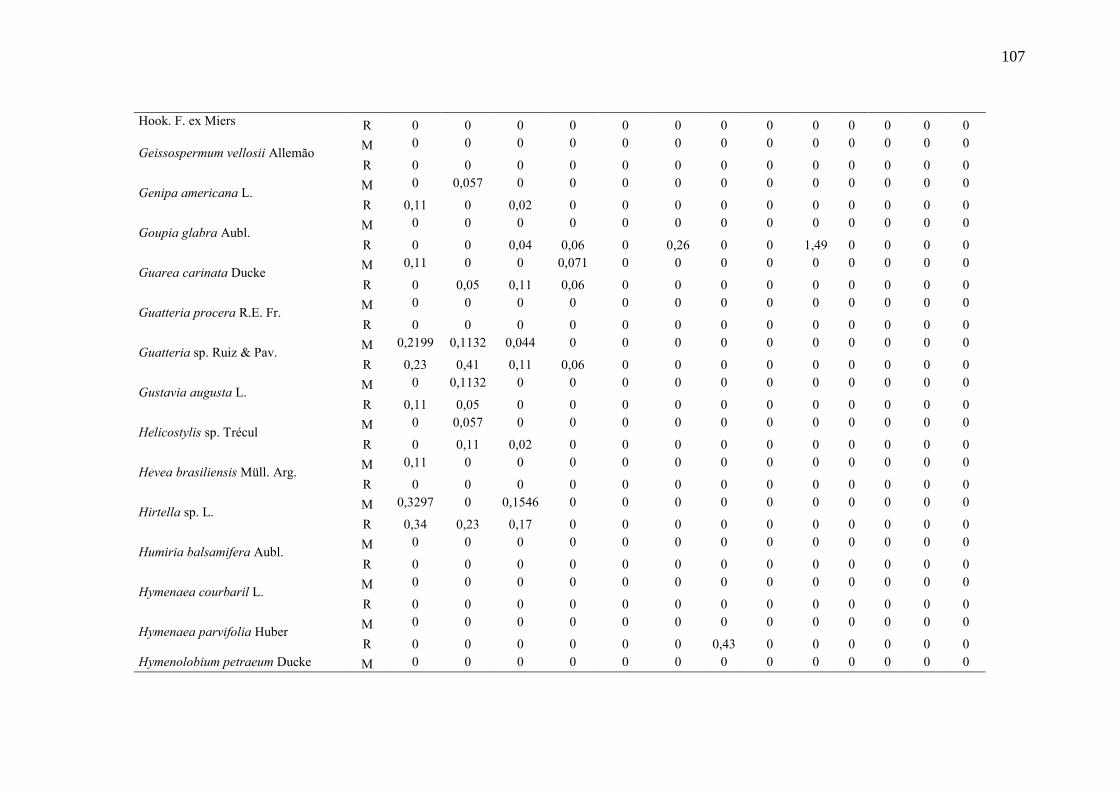
107
Hook. F. ex Miers R 0 0 0 0 0 0 0 0 0 0 0 0 0
Geissospermum vellosii Allemão M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Genipa americana L. M 0 0,057 0 0 0 0 0 0 0 0 0 0 0
R 0,11 0 0,02 0 0 0 0 0 0 0 0 0 0
Goupia glabra Aubl. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0,04 0,06 0 0,26 0 0 1,49 0 0 0 0
Guarea carinata Ducke M 0,11 0 0 0,071 0 0 0 0 0 0 0 0 0
R 0 0,05 0,11 0,06 0 0 0 0 0 0 0 0 0
Guatteria procera R.E. Fr. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Guatteria sp. Ruiz & Pav. M 0,2199 0,1132 0,044 0 0 0 0 0 0 0 0 0 0
R 0,23 0,41 0,11 0,06 0 0 0 0 0 0 0 0 0
Gustavia augusta L. M 0 0,1132 0 0 0 0 0 0 0 0 0 0 0
R 0,11 0,05 0 0 0 0 0 0 0 0 0 0 0
Helicostylis sp. Trécul M 0 0,057 0 0 0 0 0 0 0 0 0 0 0
R 0 0,11 0,02 0 0 0 0 0 0 0 0 0 0
Hevea brasiliensis Müll. Arg. M 0,11 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Hirtella sp. L. M 0,3297 0 0,1546 0 0 0 0 0 0 0 0 0 0
R 0,34 0,23 0,17 0 0 0 0 0 0 0 0 0 0
Humiria balsamifera Aubl. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Hymenaea courbaril L. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Hymenaea parvifolia Huber M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0,43 0 0 0 0 0 0
Hymenolobium petraeum Ducke M 0 0 0 0 0 0 0 0 0 0 0 0 0
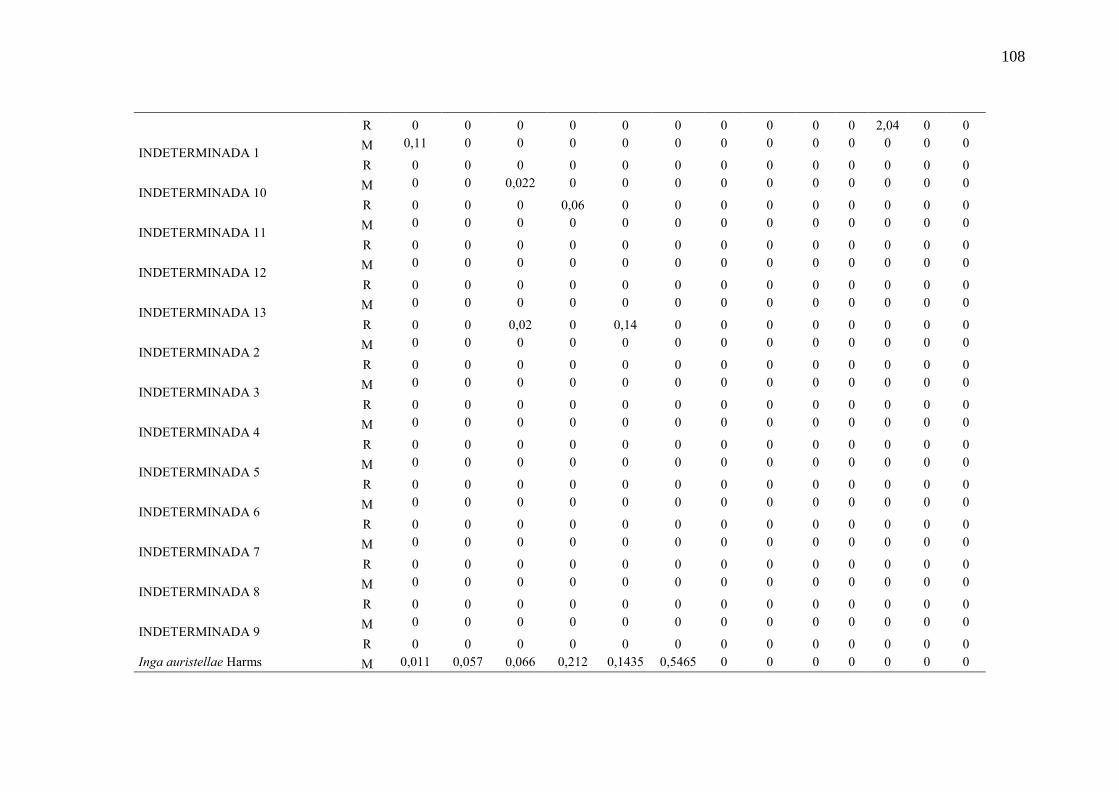
108
R 0 0 0 0 0 0 0 0 0 0 2,04 0 0
INDETERMINADA 1 M 0,11 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
INDETERMINADA 10 M 0 0 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0,06 0 0 0 0 0 0 0 0 0
INDETERMINADA 11 M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
INDETERMINADA 12 M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
INDETERMINADA 13 M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0,02 0 0,14 0 0 0 0 0 0 0 0
INDETERMINADA 2 M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
INDETERMINADA 3 M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
INDETERMINADA 4 M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
INDETERMINADA 5 M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
INDETERMINADA 6 M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
INDETERMINADA 7 M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
INDETERMINADA 8 M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
INDETERMINADA 9 M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Inga auristellae Harms M 0,011 0,057 0,066 0,212 0,1435 0,5465 0 0 0 0 0 0 0
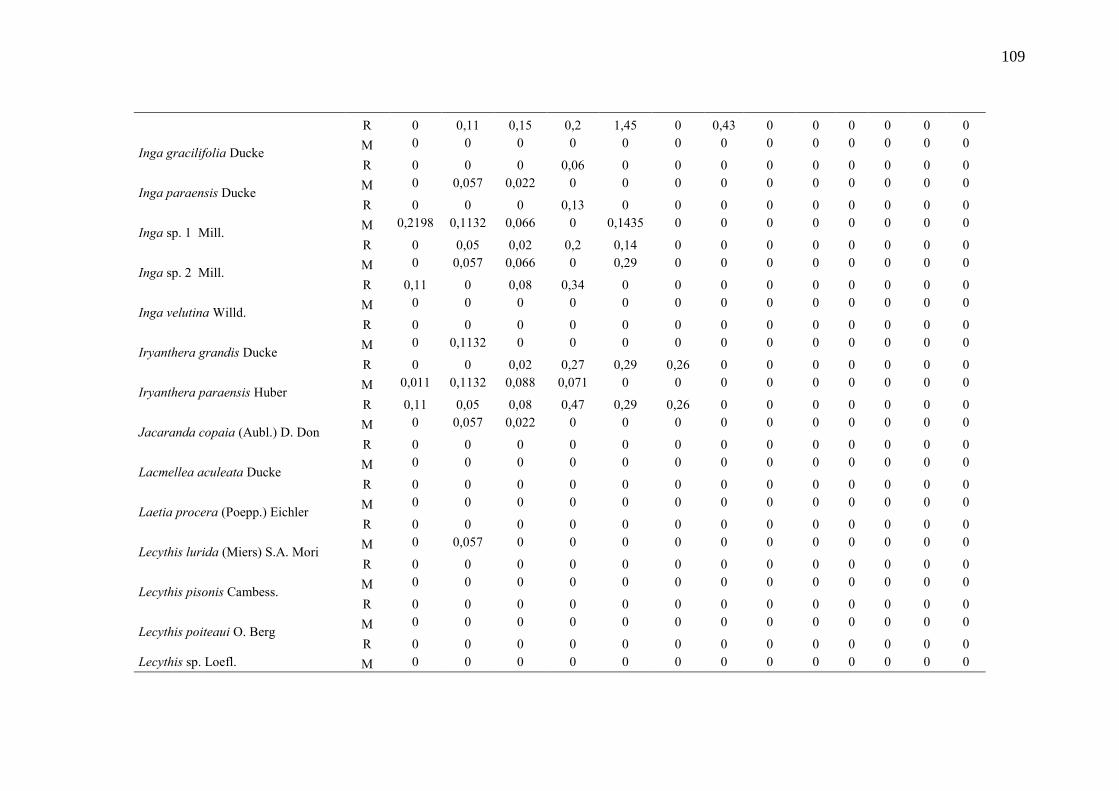
109
R 0 0,11 0,15 0,2 1,45 0 0,43 0 0 0 0 0 0
Inga gracilifolia Ducke M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0,06 0 0 0 0 0 0 0 0 0
Inga paraensis Ducke M 0 0,057 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0,13 0 0 0 0 0 0 0 0 0
Inga sp. 1 Mill. M 0,2198 0,1132 0,066 0 0,1435 0 0 0 0 0 0 0 0
R 0 0,05 0,02 0,2 0,14 0 0 0 0 0 0 0 0
Inga sp. 2 Mill. M 0 0,057 0,066 0 0,29 0 0 0 0 0 0 0 0
R 0,11 0 0,08 0,34 0 0 0 0 0 0 0 0 0
Inga velutina Willd. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Iryanthera grandis Ducke M 0 0,1132 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0,02 0,27 0,29 0,26 0 0 0 0 0 0 0
Iryanthera paraensis Huber M 0,011 0,1132 0,088 0,071 0 0 0 0 0 0 0 0 0
R 0,11 0,05 0,08 0,47 0,29 0,26 0 0 0 0 0 0 0
Jacaranda copaia (Aubl.) D. Don M 0 0,057 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Lacmellea aculeata Ducke M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Laetia procera (Poepp.) Eichler M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Lecythis lurida (Miers) S.A. Mori M 0 0,057 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Lecythis pisonis Cambess. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Lecythis poiteaui O. Berg M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Lecythis sp. Loefl. M 0 0 0 0 0 0 0 0 0 0 0 0 0
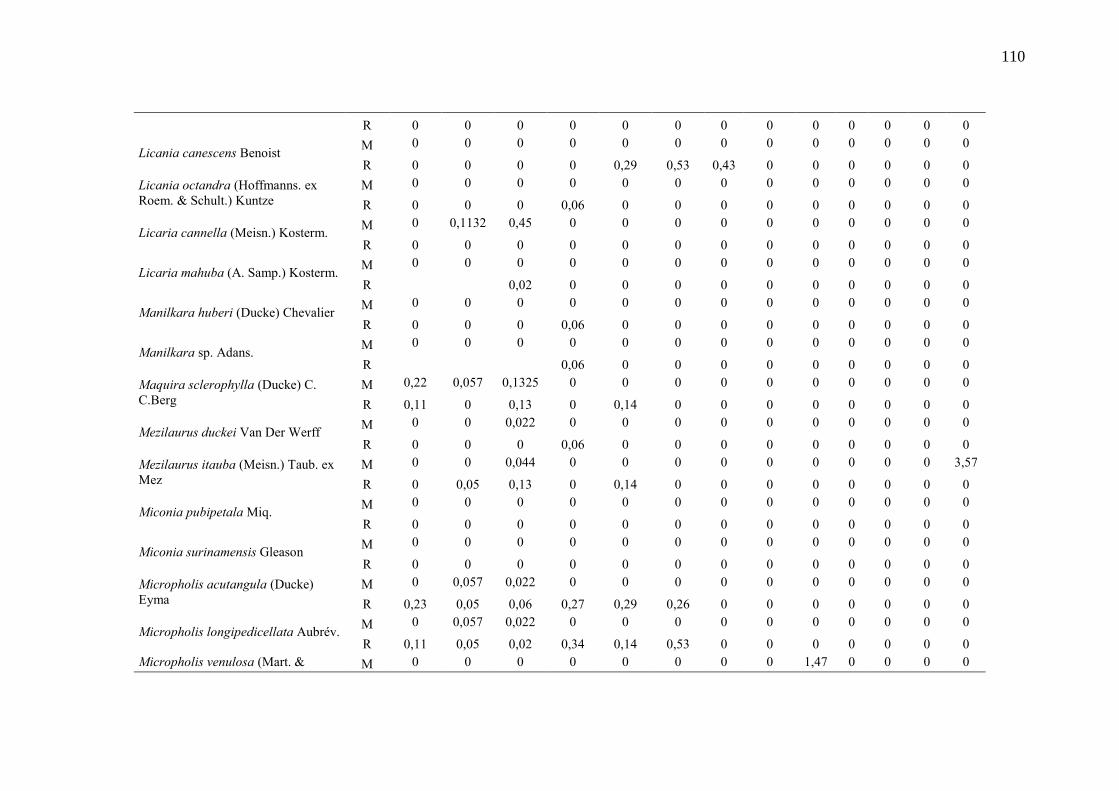
110
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Licania canescens Benoist M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0,29 0,53 0,43 0 0 0 0 0 0
Licania octandra (Hoffmanns. ex
Roem. & Schult.) Kuntze
M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0,06 0 0 0 0 0 0 0 0 0
Licaria cannella (Meisn.) Kosterm. M 0 0,1132 0,45 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Licaria mahuba (A. Samp.) Kosterm. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0,02 0 0 0 0 0 0 0 0 0 0
Manilkara huberi (Ducke) Chevalier M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0,06 0 0 0 0 0 0 0 0 0
Manilkara sp. Adans. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0,06 0 0 0 0 0 0 0 0 0
Maquira sclerophylla (Ducke) C.
C.Berg
M 0,22 0,057 0,1325 0 0 0 0 0 0 0 0 0 0
R 0,11 0 0,13 0 0,14 0 0 0 0 0 0 0 0
Mezilaurus duckei Van Der Werff M 0 0 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0,06 0 0 0 0 0 0 0 0 0
Mezilaurus itauba (Meisn.) Taub. ex
Mez
M 0 0 0,044 0 0 0 0 0 0 0 0 0 3,57
R 0 0,05 0,13 0 0,14 0 0 0 0 0 0 0 0
Miconia pubipetala Miq. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Miconia surinamensis Gleason M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Micropholis acutangula (Ducke)
Eyma
M 0 0,057 0,022 0 0 0 0 0 0 0 0 0 0
R 0,23 0,05 0,06 0,27 0,29 0,26 0 0 0 0 0 0 0
Micropholis longipedicellata Aubrév. M 0 0,057 0,022 0 0 0 0 0 0 0 0 0 0
R 0,11 0,05 0,02 0,34 0,14 0,53 0 0 0 0 0 0 0
Micropholis venulosa (Mart. & M 0 0 0 0 0 0 0 0 1,47 0 0 0 0
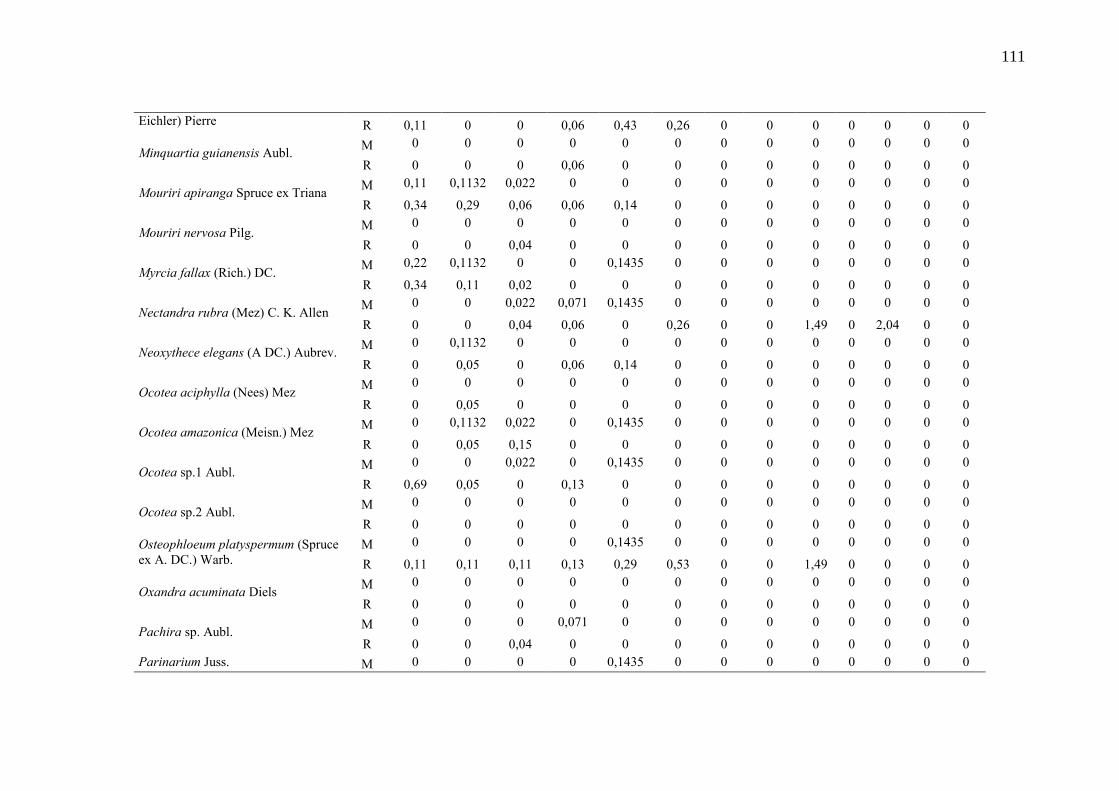
111
Eichler) Pierre R 0,11 0 0 0,06 0,43 0,26 0 0 0 0 0 0 0
Minquartia guianensis Aubl. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0,06 0 0 0 0 0 0 0 0 0
Mouriri apiranga Spruce ex Triana M 0,11 0,1132 0,022 0 0 0 0 0 0 0 0 0 0
R 0,34 0,29 0,06 0,06 0,14 0 0 0 0 0 0 0 0
Mouriri nervosa Pilg. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0,04 0 0 0 0 0 0 0 0 0 0
Myrcia fallax (Rich.) DC. M 0,22 0,1132 0 0 0,1435 0 0 0 0 0 0 0 0
R 0,34 0,11 0,02 0 0 0 0 0 0 0 0 0 0
Nectandra rubra (Mez) C. K. Allen M 0 0 0,022 0,071 0,1435 0 0 0 0 0 0 0 0
R 0 0 0,04 0,06 0 0,26 0 0 1,49 0 2,04 0 0
Neoxythece elegans (A DC.) Aubrev. M 0 0,1132 0 0 0 0 0 0 0 0 0 0 0
R 0 0,05 0 0,06 0,14 0 0 0 0 0 0 0 0
Ocotea aciphylla (Nees) Mez M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0,05 0 0 0 0 0 0 0 0 0 0 0
Ocotea amazonica (Meisn.) Mez M 0 0,1132 0,022 0 0,1435 0 0 0 0 0 0 0 0
R 0 0,05 0,15 0 0 0 0 0 0 0 0 0 0
Ocotea sp.1 Aubl. M 0 0 0,022 0 0,1435 0 0 0 0 0 0 0 0
R 0,69 0,05 0 0,13 0 0 0 0 0 0 0 0 0
Ocotea sp.2 Aubl. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Osteophloeum platyspermum (Spruce
ex A. DC.) Warb.
M 0 0 0 0 0,1435 0 0 0 0 0 0 0 0
R 0,11 0,11 0,11 0,13 0,29 0,53 0 0 1,49 0 0 0 0
Oxandra acuminata Diels M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Pachira sp. Aubl. M 0 0 0 0,071 0 0 0 0 0 0 0 0 0
R 0 0 0,04 0 0 0 0 0 0 0 0 0 0
Parinarium Juss. M 0 0 0 0 0,1435 0 0 0 0 0 0 0 0
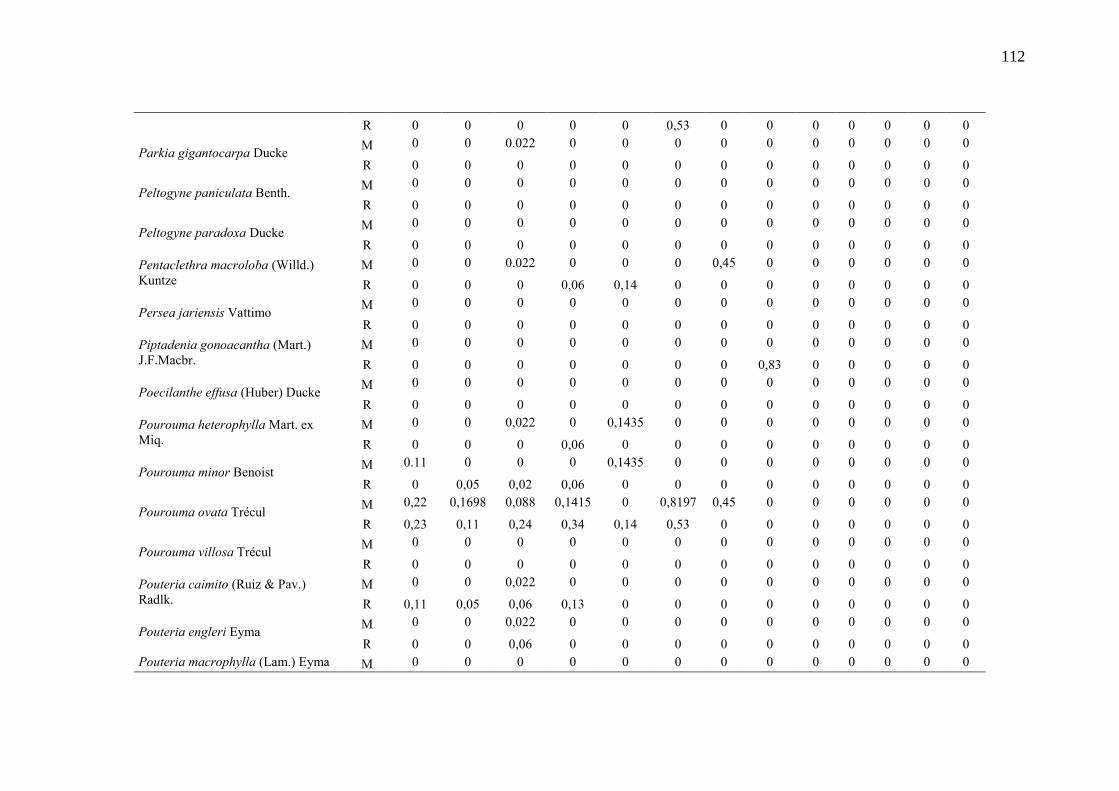
112
R 0 0 0 0 0 0,53 0 0 0 0 0 0 0
Parkia gigantocarpa Ducke M 0 0 0.022 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Peltogyne paniculata Benth. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Peltogyne paradoxa Ducke M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Pentaclethra macroloba (Willd.)
Kuntze
M 0 0 0.022 0 0 0 0,45 0 0 0 0 0 0
R 0 0 0 0,06 0,14 0 0 0 0 0 0 0 0
Persea jariensis Vattimo M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Piptadenia gonoacantha (Mart.)
J.F.Macbr.
M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0,83 0 0 0 0 0
Poecilanthe effusa (Huber) Ducke M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Pourouma heterophylla Mart. ex
Miq.
M 0 0 0,022 0 0,1435 0 0 0 0 0 0 0 0
R 0 0 0 0,06 0 0 0 0 0 0 0 0 0
Pourouma minor Benoist M 0.11 0 0 0 0,1435 0 0 0 0 0 0 0 0
R 0 0,05 0,02 0,06 0 0 0 0 0 0 0 0 0
Pourouma ovata Trécul M 0,22 0,1698 0,088 0,1415 0 0,8197 0,45 0 0 0 0 0 0
R 0,23 0,11 0,24 0,34 0,14 0,53 0 0 0 0 0 0 0
Pourouma villosa Trécul M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Pouteria caimito (Ruiz & Pav.)
Radlk.
M 0 0 0,022 0 0 0 0 0 0 0 0 0 0
R 0,11 0,05 0,06 0,13 0 0 0 0 0 0 0 0 0
Pouteria engleri Eyma M 0 0 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0 0,06 0 0 0 0 0 0 0 0 0 0
Pouteria macrophylla (Lam.) Eyma M 0 0 0 0 0 0 0 0 0 0 0 0 0
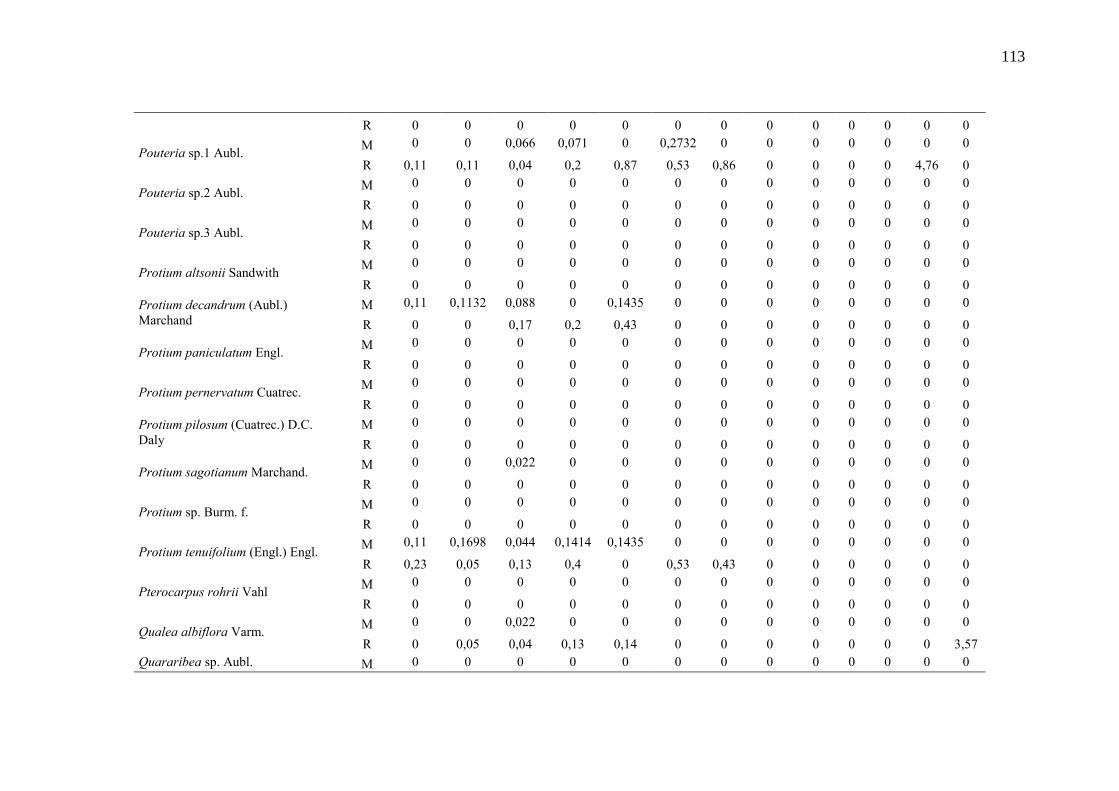
113
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Pouteria sp.1 Aubl. M 0 0 0,066 0,071 0 0,2732 0 0 0 0 0 0 0
R 0,11 0,11 0,04 0,2 0,87 0,53 0,86 0 0 0 0 4,76 0
Pouteria sp.2 Aubl. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Pouteria sp.3 Aubl. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Protium altsonii Sandwith M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Protium decandrum (Aubl.)
Marchand
M 0,11 0,1132 0,088 0 0,1435 0 0 0 0 0 0 0 0
R 0 0 0,17 0,2 0,43 0 0 0 0 0 0 0 0
Protium paniculatum Engl. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Protium pernervatum Cuatrec. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Protium pilosum (Cuatrec.) D.C.
Daly
M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Protium sagotianum Marchand. M 0 0 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Protium sp. Burm. f. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Protium tenuifolium (Engl.) Engl. M 0,11 0,1698 0,044 0,1414 0,1435 0 0 0 0 0 0 0 0
R 0,23 0,05 0,13 0,4 0 0,53 0,43 0 0 0 0 0 0
Pterocarpus rohrii Vahl M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Qualea albiflora Varm. M 0 0 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0,05 0,04 0,13 0,14 0 0 0 0 0 0 0 3,57
Quararibea sp. Aubl. M 0 0 0 0 0 0 0 0 0 0 0 0 0
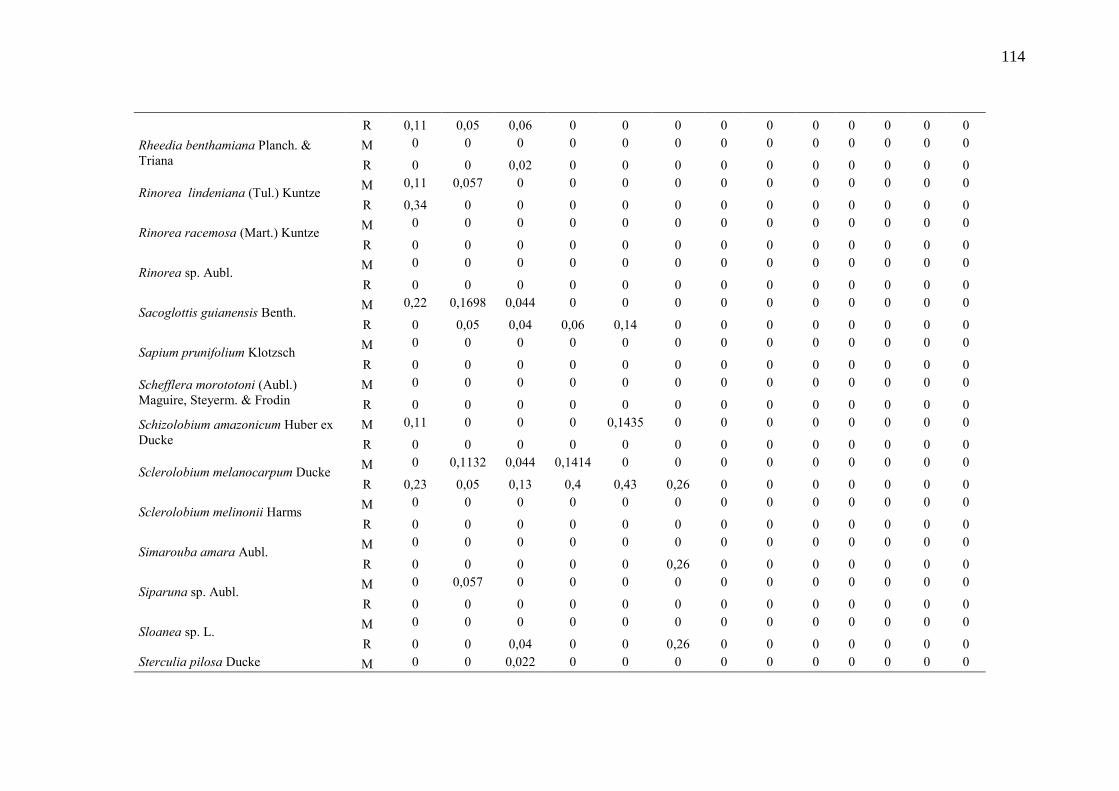
114
R 0,11 0,05 0,06 0 0 0 0 0 0 0 0 0 0
Rheedia benthamiana Planch. &
Triana
M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0,02 0 0 0 0 0 0 0 0 0 0
Rinorea lindeniana (Tul.) Kuntze M 0,11 0,057 0 0 0 0 0 0 0 0 0 0 0
R 0,34 0 0 0 0 0 0 0 0 0 0 0 0
Rinorea racemosa (Mart.) Kuntze M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Rinorea sp. Aubl. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Sacoglottis guianensis Benth. M 0,22 0,1698 0,044 0 0 0 0 0 0 0 0 0 0
R 0 0,05 0,04 0,06 0,14 0 0 0 0 0 0 0 0
Sapium prunifolium Klotzsch M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Schefflera morototoni (Aubl.)
Maguire, Steyerm. & Frodin
M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Schizolobium amazonicum Huber ex
Ducke
M 0,11 0 0 0 0,1435 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Sclerolobium melanocarpum Ducke M 0 0,1132 0,044 0,1414 0 0 0 0 0 0 0 0 0
R 0,23 0,05 0,13 0,4 0,43 0,26 0 0 0 0 0 0 0
Sclerolobium melinonii Harms M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Simarouba amara Aubl. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0,26 0 0 0 0 0 0 0
Siparuna sp. Aubl. M 0 0,057 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Sloanea sp. L. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0,04 0 0 0,26 0 0 0 0 0 0 0
Sterculia pilosa Ducke M 0 0 0,022 0 0 0 0 0 0 0 0 0 0

115
R 0,23 0,05 0,02 0 0 0 1,29 0 0 0 0 0 0
Stryphnodendron pulcherrimum
(Willd.) Hochr.
M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0,13 0 0,26 0 0 0 0 0 0 0
Swartzia racemosa Benth. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0,11 0 0 0 0 0 0 0 0 0 0 0 0
Swartzia sp. Schreb. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Symphonia globulifera L. f. M 0 0 0,022 0 0 0 0,45 0 0 0 0 0 0
R 0 0,05 0 0,2 0 0 0,43 0 0 0 0 0 0
Tabebuia serratifolia (Vahl) G.
Nicholson
M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Tabebuia sp. Gomes ex DC. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Tachigali myrmecophila Ducke M 0 0 0,044 0 0 0 0,45 0 0 0 0 0 0
R 0 0,05 0,06 0,06 0,14 0 0,43 0 0 2,5 0 0 0
Tachigali sp. Aubl. M 0 0 0,044 0,071 0 0 0 0 0 0 0 0 0
R 0,34 0 0,04 0 0,14 0 0 0 0 0 0 0 0
Tapirira guianensis Aubl. M 0,11 0 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0,05 0,04 0,13 0,14 0 0 0 0 0 0 0 0
Taralea oppositifolia Aubl. M 0,11 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0,02 0 0 0 0 0 0 0 0 0 0
Terminalia amazonia (J.F.Gmel.)
Exell
M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Terminalia sp. L. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Ternstroemia sp. Mutis ex L. f. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Theobroma sect. Cacao (Mill.) M 0 0 0 0 0 0 0 0 0 0 0 0 0
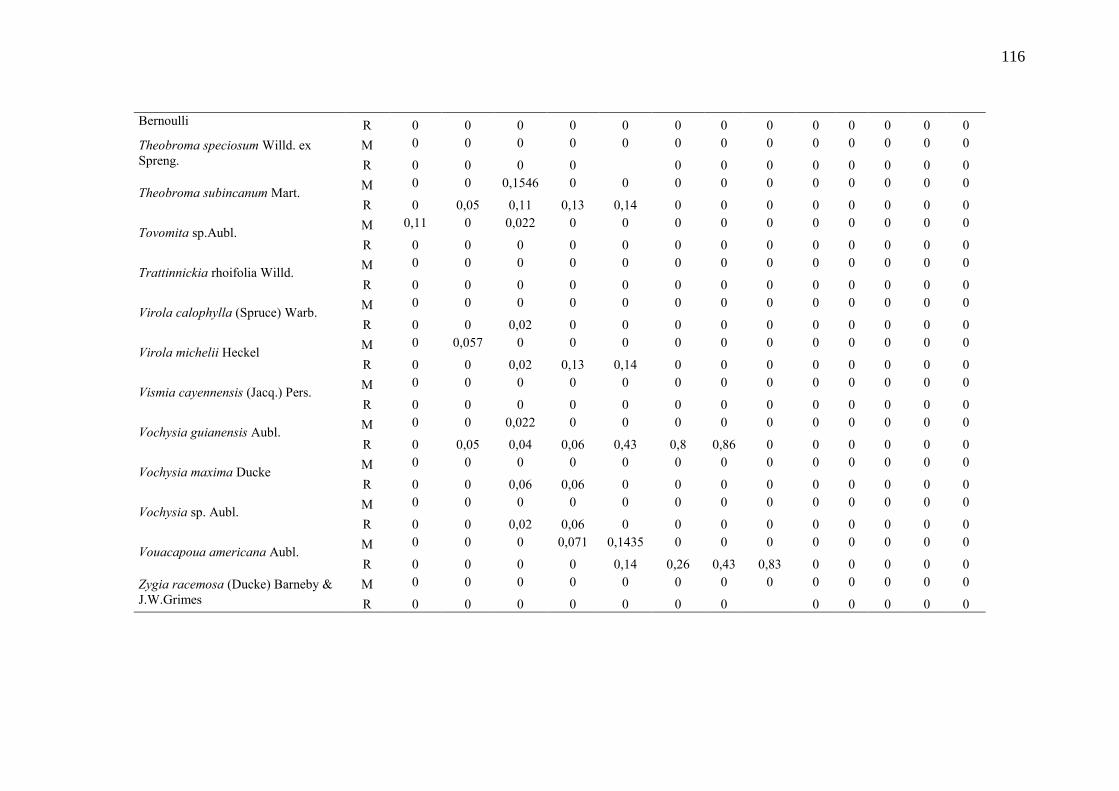
116
Bernoulli R 0 0 0 0 0 0 0 0 0 0 0 0 0
Theobroma speciosum Willd. ex
Spreng.
M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0
Theobroma subincanum Mart. M 0 0 0,1546 0 0 0 0 0 0 0 0 0 0
R 0 0,05 0,11 0,13 0,14 0 0 0 0 0 0 0 0
Tovomita sp.Aubl. M 0,11 0 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Trattinnickia rhoifolia Willd. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Virola calophylla (Spruce) Warb. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0,02 0 0 0 0 0 0 0 0 0 0
Virola michelii Heckel M 0 0,057 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0,02 0,13 0,14 0 0 0 0 0 0 0 0
Vismia cayennensis (Jacq.) Pers. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0 0
Vochysia guianensis Aubl. M 0 0 0,022 0 0 0 0 0 0 0 0 0 0
R 0 0,05 0,04 0,06 0,43 0,8 0,86 0 0 0 0 0 0
Vochysia maxima Ducke M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0,06 0,06 0 0 0 0 0 0 0 0 0
Vochysia sp. Aubl. M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0,02 0,06 0 0 0 0 0 0 0 0 0
Vouacapoua americana Aubl. M 0 0 0 0,071 0,1435 0 0 0 0 0 0 0 0
R 0 0 0 0 0,14 0,26 0,43 0,83 0 0 0 0 0
Zygia racemosa (Ducke) Barneby &
J.W.Grimes
M 0 0 0 0 0 0 0 0 0 0 0 0 0
R 0 0 0 0 0 0 0 0 0 0 0 0

117
Quanto à taxa de mortalidade, observou-se percentuais variando de 3,88 a 3,70
%. As duas classes diamétricas iniciais, varas e arvoretas, apresentam elevados
percentuais de mortalidade. No entanto, proporcionalmente também ocorre elevado
número de árvores mortas (Mezilaurus itauba - 3,70 %) entre os indivíduos com DAP´s
superiores a 110 cm (Tabela 10).
Tabela 10. Taxa de recrutamento (t), mortalidade (M), incremento periódico anual percentual (IPA) e
incremento periódico anual médio (IPAm) nas classes diamétricas numa floresta ombrófila densa, Porto
Grande/AP.
C.D. (%) IPAm
(cm) T M IPA
1 2,5 < DAP < 5 6,64 3,88 9.13 0.30
2 5 < DAP < 10 4,52 3,27 6.13 0.50
3 10 < DAP < 20 4,43 2,47 4.41 0.60
4 20 < DAP < 30 9,88 1,87 3.16 0.80
5 30 < DAP < 40 9,75 3,19 2.36 0.80
6 40 < DAP < 50 10,24 3,10 1.76 0.80
7 50 < DAP < 60 9,48 2,30 1.25 0.70
8 60 < DAP < 70 5,83 0,85 0.98 0.60
9 70 < DAP < 80 5,97 1,49 0.85 0.60
10 80 < DAP < 90 10,00 0,00 0.85 0.70
11 90 < DAP < 100 12,24 0,00 0.82 0.80
12 100 < DAP < 110 4,76 0,00 0.57 0.60
13 DAP > 110 3,57 3,70 0.46 0.60
Na classe 1 os maiores percentuais de mortalidade foram divididos entre duas
espécies, a Eschweilera coriacea e Hirtella sp, (0,33 %). Sequencialmente, para a classe
2 foram: Pourouma ovata, Protium tenuifolium e Sacoglottis guianensis (0,17 %). No
entanto, na terceira classe apenas a Licaria cannella (0,45 %) mereceu destaque.
Quanto aos IPA’s percentuais médios durante os três anos de monitoramento, os
valores de volume foram decrescendo à medida que o tamanho das árvores aumentava,
Foram estimados variações de 0,47 a 9,14 % nas classes diamétricas. O maior
percentual foi verificado na menor classe diamétrica, enquanto que a última classe
obteve estimativas mais discretas.
Os diâmetros médios por classe diamétricas também decresceram
progressivamente, com variação 3,4 a 130,4 cm. Quando estimados os crescimentos
diamétricos em centímetros das classes diamétricas a partir dos percentuais de
incrementos periódicos, constatou-se que os indivíduos mais jovens crescem 0,30 e 0,50

118
cm,ano-1
, para a primeira e segunda classe diamétrica, respectivamente. Além disso, o
crescimento pouco variou nas demais classes, com valores de 0,6 a 0,8 cm,ano-1
.
Apenas três espécie obtiveram incrementos médio acima de 10 %, Duguetia
stelechantha (11,97 %), Indeterminada 11 (12,90 %) e Rinorea sp. (16,81 %).
Entretanto, oito espécies apresentaram percentuais abaixo de 1 %, como a Anacardium
spruceanum, Dialium guianense e Hymenolobium petraeum. Também foi observado
que 57 espécies obtiveram incrementos acima de 5 %.
Espécies comerciais como Carapa guianensis, Dinizia excelsa, Dipteryx odorata
e Tabebuia serratifolia apresentaram incrementos percentuais de 4,64, 2,51, 2,0 e 3,67
%. Entretanto, Dinizia excelsa apresentou uma baixa frequência nas classes diamétrica
(30,76 % total das classes). Se considerada a variação de incremento nas espécies com
alto valor tecnológico e comumentes comercializadas na região amazônica, foram
encontrados valores de 0,86 % (Peltogyne paniculata) a 5,54 % (Quararibea sp,).
Quando analisado o incremento das espécies classificadas como madeira branca,
também comercializadas na Amazônia, mas com preços mais acessíveis aos
empresários locais, se verificou maiores percentuais. Contudo, a variação de
crescimento também aumentou (Ternstroemiasp,) de 0,59 % a (Duguetia
stelechantha)11,97 %.
Quanto ao número de indivíduos e ao crescimento diamétrico das espécies, foi
constatado que o fator tempo influenciou de maneira significativa em ambas variáveis
(número de indivíduos (Gl=2; W=0,4496); crescimento diamétrico (Gl=2; W=0,6386) -
p< 0,0001).
O número de indivíduos apresentou mudanças significativas entre as interações
do tempo com os níveis de inclusão e GVM (Tabela 11), Entretanto, o crescimento em
diâmetro entre os níveis não evidenciou diferenças entre as varas, arvoretas e árvores.
Além disso, verificou-se que o crescimento dos diâmetros entre as UP’s e GVM variou
ao longo do tempo.
Tabela 11. Teste de hipósete multivariado de Wilks para o efeito tempo (T) e interações sobre o número
de indivíduos (N) e crescimento em diâmetro (C) entre as Unidades Primárias (UP), Nível de inclusão e
Grupo de valor madeireiro (GVM) em uma floresta ombrófila densa, Porto Grande/AP.
Efeito GL Wilks F p
N C N C N C N C
T 2 2 0,920 0,977 12,518 3,377 0,000* 0,036*
T*UP 4 4 0,982 0,965 1,279 2,582 0,277 0,036*
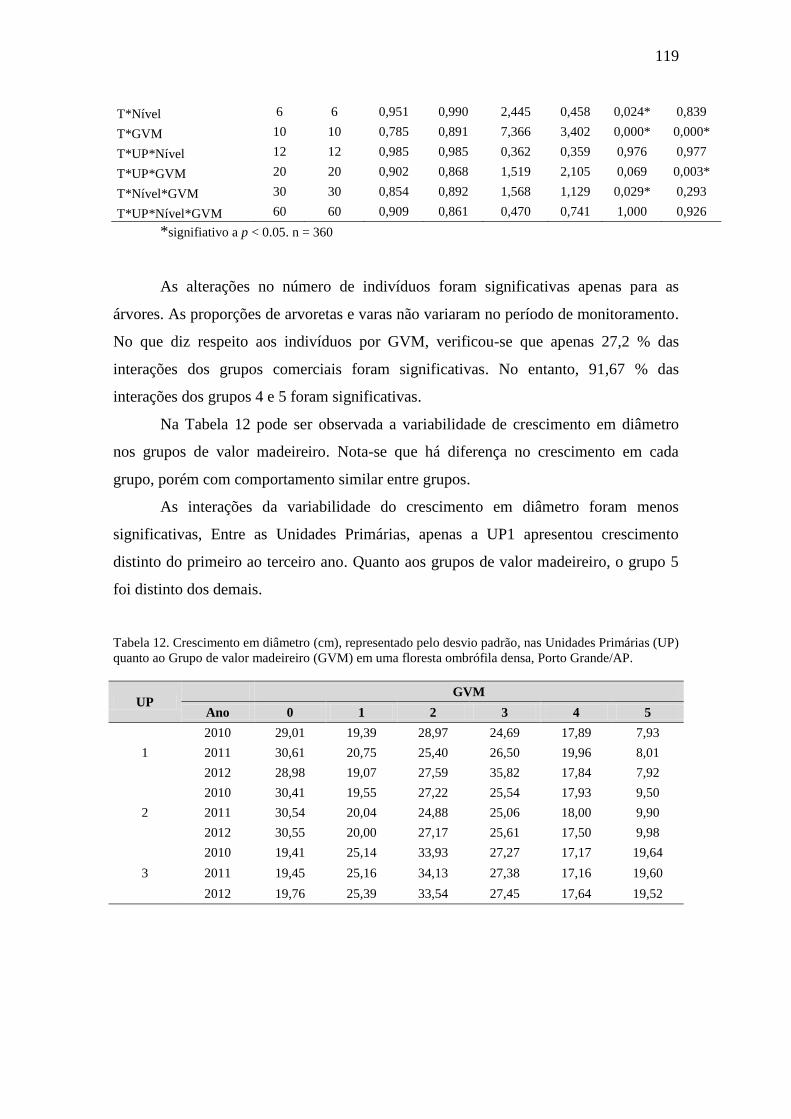
119
T*Nível 6 6 0,951 0,990 2,445 0,458 0,024* 0,839
T*GVM 10 10 0,785 0,891 7,366 3,402 0,000* 0,000*
T*UP*Nível 12 12 0,985 0,985 0,362 0,359 0,976 0,977
T*UP*GVM 20 20 0,902 0,868 1,519 2,105 0,069 0,003*
T*Nível*GVM 30 30 0,854 0,892 1,568 1,129 0,029* 0,293
T*UP*Nível*GVM 60 60 0,909 0,861 0,470 0,741 1,000 0,926
*signifiativo a p < 0.05. n = 360
As alterações no número de indivíduos foram significativas apenas para as
árvores. As proporções de arvoretas e varas não variaram no período de monitoramento.
No que diz respeito aos indivíduos por GVM, verificou-se que apenas 27,2 % das
interações dos grupos comerciais foram significativas. No entanto, 91,67 % das
interações dos grupos 4 e 5 foram significativas.
Na Tabela 12 pode ser observada a variabilidade de crescimento em diâmetro
nos grupos de valor madeireiro. Nota-se que há diferença no crescimento em cada
grupo, porém com comportamento similar entre grupos.
As interações da variabilidade do crescimento em diâmetro foram menos
significativas, Entre as Unidades Primárias, apenas a UP1 apresentou crescimento
distinto do primeiro ao terceiro ano. Quanto aos grupos de valor madeireiro, o grupo 5
foi distinto dos demais.
Tabela 12. Crescimento em diâmetro (cm), representado pelo desvio padrão, nas Unidades Primárias (UP)
quanto ao Grupo de valor madeireiro (GVM) em uma floresta ombrófila densa, Porto Grande/AP.
UP GVM
Ano 0 1 2 3 4 5
1
2010 29,01 19,39 28,97 24,69 17,89 7,93
2011 30,61 20,75 25,40 26,50 19,96 8,01
2012 28,98 19,07 27,59 35,82 17,84 7,92
2
2010 30,41 19,55 27,22 25,54 17,93 9,50
2011 30,54 20,04 24,88 25,06 18,00 9,90
2012 30,55 20,00 27,17 25,61 17,50 9,98
3
2010 19,41 25,14 33,93 27,27 17,17 19,64
2011 19,45 25,16 34,13 27,38 17,16 19,60
2012 19,76 25,39 33,54 27,45 17,64 19,52

120
5.4 DISCUSSÃO
O número de indivíduos por hectare encontrado foi considerado elevado em
relação aos demais inventários florísticos realizados na região Amazônica. O fato é
consequência do nível de inclusão adotado, onde se abrangeu plantas jovens que
geralmente não são inventariados em trabalhos de estoque madeireiro com fins ao
manejo florestal. Pesquisadores da região norte (OLIVEIRA et al., 2008; SALOMÃO et
al., 2012) trabalhando com indivíduos com diâmetros superiores a 10 cm estimaram
entre 450 a 670 árvores/ha em florestas ombrófila densas.
A renovação de indivíduos arbóreos na composição da floresta é garantida pelo
excesso de plantas por unidade de área nas classes iniciais, principalmente por que o
número de árvores aptas à exploração (DAP > 50 cm) apresentou suscetibilidade ao
corte. No entanto, este comportamento é comum em florestas tropicais inequiâneas
mistas, com decréscimo proporcional do número de indivíduos à medida que os
diâmetros aumentam (DE LIOCOURT, 1898), devendo também ser considerado as
relações estruturais e fitossociológicas das populações para definir as espécies, as
classes de tamanho e as taxas de corte que serão alvo do manejo.
A significância da interação verificada entre o tempo e o GVM para o número de
indivíduos e para o crescimento em diâmetro eram esperados, em virtude da ecologia
das espécies inventariadas. Estas características induzem a distintas adaptações no
ambiente, bem como distribuições de densidade e espacial na floresta, formando grupos
ecológicos com maior (madeiras brancas/mole) e menor (madeiras vermelhas/dura)
taxas de crescimento. As mudanças verificadas ao longo do monitoramento também são
consequência das distintas características dos sítios, os quais possuem diferentes
estruturas na composição do solo e histórico de uso.
A não ocorrência de taxas diferenciadas de crescimento em diâmetro para os
níveis deve-se ao curto tempo de monitoramento das parcelas, com tendência a maiores
variações entre as árvores com menores diâmetros, seja pela própria regeneração ou
competição intra e interespecífica.
No que diz respeito às altas taxas de recrutamento em classes diamétricas
superiores a 50 cm de DAP, pode-se inferir duas possibilidades: a) em virtude da
amplitude das classes, as quais o tamanho das árvores encontrava-se sob os limites
superiores das classes, e no intervalo de medição o crescimento proporcionou o

121
recrutamento para a classe posterior; b) devido a erros sistemáticos de medição, com
ponto de medição do diâmetro distinto durante o monitoramento.
Para as espécies que apresentaram taxas de recrutamento em várias classes
diamétricas, existem indícios que estas estão mais adaptadas aos sítios, com
recrutamento dos estratos inferiores aos superiores da floresta. Outro ponto é o
comportamento sucessional das espécies, onde há um rápido crescimento de espécies
pioneiras para posteriormente as secundárias apresentarem aceleração no crescimento.
A exposição das copas das árvores à luz solar influência diretamente no
incremento em diâmetro, com variação de 0,57 cm.ano-1
(árvores expostas à luz solar), a
0,28 cm.ano-1
(árvores sombreadas) (D’ IVEIRA e BRAZ, 2006).
No ano de 1999, Ferreira e colaboradores consideraram que a aplicação correta
da exploração de impacto reduzido em florestas tropicais pode fornecer incrementos
diamétricos médios entre 0,7 e 1,0 cm/ano. Numa floresta primária da Amazônia
Central, Higuchi et al, (2008) estimaram o recrutamento em 0,7% entre 1986 a 2000.
Em estudos sobre incrementos das árvores, Braz et al. (2012) afirmaram que o
potencial de crescimento das florestas naturais são um forte suporte a projeção do
crescimento e estrutura futura das mesmas quando manejadas.
Os percentuais médios encontrados para a mortalidade nas classes diamétricas
encontram-se dentro dos padrões estabelecidos para florestas inequiâneas mistas.
Geralmente, se espera valores entre 1 e 3 % para florestas não perturbadas por
exploração (MANOKARAN e KOCHUMMEN, 1987). No entanto, as duas classes
iniciais, bem como a última (contempla os maiores indivíduos) obtiveram altos
percentuais.
Tais resultados são justificados pela amplitude das classes e nível de inclusão
adotado. Quando jovens, há uma competição mais agressiva, e, consequentemente
aumentam a mortalidade dos indivíduos. Além disso, as maiores árvores inventariadas,
provavelmente, se encontravam em condições fitossanitárias desfavoráveis e/ou estado
senescente no inicio do monitoramento, tendo como consequência o ressecamento e
morte.
Também foi relatado que os valores-padrão das taxas médias de mortalidade de
árvores em florestas tropicais primárias são de 1 a 2 % ao ano, com uma taxa bem mais
elevada para espécies pioneiras (KÖHLER et al., 2002). Em cinco anos de
monitoramento em Floresta de Terra Firme, Rocha (2001) encontrou taxa de

122
mortalidade de 0,86 % ao ano. Similar ao encontrado por Higuchi et al. (2004), os quais
obtiveram 0,7 % anuais de mortalidade em 14 anos de monitoramento numa floresta
primária da Amazônia Central.
Segundo Braz (2010), em povoamentos que apresentam comportamento
diamétrico com decréscimo do número de árvores à medida que os diâmetros
aumentam, as taxas de mortalidade serão mais altas do que povoamentos com pequena
razão numérica entre o número de árvores nas sucessivas classes de diâmetro.
A relação entre as taxas de recrutamento e mortalidade evidenciam que a floresta
apresenta uma estrutura balanceada, mantendo proporcionalidade entre os indivíduos
das espécies nas classes diamétricas. Campos et al. (1983) afirmaram que quando ocorre
um decréscimo da frequência das árvores com menor diâmetro, consequentemente há
condições favoráveis para o aumento do número de indivíduos de maior diâmetro,
tornando a floresta de maior valor comercial.
Os incrementos periódicos anuais demonstraram que as árvores das espécies
inventariadas crescem de forma similar a outras florestas de mesma tipologia. Vidal et
al. (2002), trabalhando na Amazônia Oriental, afirmaram que árvores submetidas ao
manejo apresentaram incremento periódico anual em DAP de 0,63 cm.ano-1
, duas vezes
maior do que florestas sem manejo, sem qualquer tipo de intervenção.
Menores crescimentos foram verificados por Vieira (2003) na região de Rio
Branco (0,39 cm.ano-1
) e Santarém (0,31 cm.ano-1
). Entretanto, foram verificados no
Projeto Jarí, Amapá, incrementos periódicos baixos (0,14 cm.ano-1
) para uma floresta
primária (CV=8 %), e altos (0,60 cm.ano-1
) para uma floresta secundária (CV=132 %),
em indivíduos com diâmetro mínimo acima de 5 cm (GOMIDE, 1997),
Os parâmetros de ingresso, mortalidade e incremento estão em acordo com
outras florestas da Amazônia já submetidas ao processo de exploração, com resultados
de sucesso. Este fato induza uma projeção de restauração da capacidade produtiva da
floresta para o primeiro ciclo de corte, o qual ocorre entre 25 e 35 anos.
A respeito da projeção da distribuição diamétrica da floresta, percebeu-se que as
altas estimativas de mortalidade esperada, provavelmente, serão proporcionadas pela
renovação da floresta, a partir da competição, bem como do rápido crescimento das
espécies. Cunha et al. (2002) constataram que espécies clímax geralmente possuem
crescimento lento, podendo ser aferida com base nos autovalores da matriz de transição.

123
Quando ocorre uma intensidade de exploração mais severa, um grande número
de plantas pioneiras é recrutado, o que pode acarretar um atraso no desenvolvimento da
floresta para o próximo ciclo de corte. Souza et al. (2004) analisando a dinâmica numa
Floresta Ombrófila Densa secundária, após o corte de cipós, indicaram que atividade
está favorecendo a sucessão secundária, aumentando o número de indivíduos na
população, com taxas de recrutamento e crescimento superiores as de mortalidade.
5.5 CONCLUSÕES
Ocorreram poucas variações de incrementos em DAP entre os grupos comerciais
e nas classes diamétricas.
A superioridade das taxas de ingresso em relação à mortalidade indica que a
floresta se apresenta num estágio sucessional de constante renovação.
Baseado na renovação de indivíduos na área acredita-se que a floresta é capaz de
ser submetida ao manejo, sem comprometer a estrutura da comunidade.

124
6 CONSIDERAÇÕES FINAIS
A utilização das toras comerciais das árvores caídas presentes na Floresta
Estadual do Amapá é uma alternativa real para formação de equações volumétricas
locais. O fato baseia-se na semelhança das estruturas diamétrica e hipsométrica das
árvores caídas com as das árvores vivas em “pé” em distintas fitofisionomias.
O manejo madeireiro da Floresta Estadual do Amapá está subsidiado pelo
elevado volume e número de árvores aptas ao corte. Além disso, a estrutura da floresta
atende todos os critérios legais da política brasileira de acesso aos recursos florestais.
Destaca-se a regeneração natural que garante de renovação das espécies arbóreas
comerciais para ciclos de corte subseqüente.
Logo, os parâmetros fitossociológicos e a regeneração natural resultaram em
respostas positivas quando relacionados ao manejo florestal da área em estudo, para o
primeiro ciclo de colheita. O local de estudo apresentou um satisfatório número de
espécies adultas comercializáveis com relevante número de indivíduos, boas condições
fitossanitárias e volume apto à exploração, além de oferecer indivíduos jovens das
espécies alvo de corte que devem ser recrutadas para um estágio superior no futuro.
Teoricamente a floresta em estudo apresenta crescimento suficiente para
restaurar a floresta para um próximo ciclo de corte conforme a legislação brasileira.

REFERÊNCIAS
ALMEIDA; L. S.; GAMA; J. R. V.; OLIVEIRA; F. A.; CARVALHO; J. O. P.;
GONÇALVES, D. C. M.; ARAÚJO, G. C. Fitossociologia e uso múltiplo de espécies
arbóreas em floresta manejada, Comunidade Santo Antônio, município de Santarém,
Estado do Pará. Acta Amazônica. vol. 42(2), 185 – 194, 2012.
ALVES, J. C. Z. O.; MIRANDA , I. S. Análise da estrutura de comunidades arbóreas de
uma floresta amazônica de Terra Firme aplicada ao manejo florestal. Manaus-AM: Acta
Amazonica, vol. 38(4), p. 657- 666, 2008.
APARÍCIO, W. C. S. Estrutura da vegetação em diferentes ambientes na Resex do
Rio Cajari: Interações do solo-floresta e relações com a produção de castanha.
2011, 150 f. Tese (Doutorado em Ciências Florestais) – Universidade Federal Rural de
Pernambuco, Recife, 2011.
APG III. An update of the angiosperm phylogeny group classification for orders and
families of flowering plants. Bot. J. Linn. 2010.
AZEVEDO, C. P.; SANQUETTA, C. R.; SILVA, J. N. M.; MACHADO, S. A.;
SOUZA, C. R.; OLIVEIRA, M. M. Simulação de estratégias de manejo florestal na
Amazônia com o uso do modelo SYMFOR. Acta Amazônica. v. 38, n.1, p. 51 – 70,
2008.
BALÉE, W.; CAMPBELL, D. G. Evidence for the sucessional status of liana forest
(Xingu river Basin, Amazônia, Brasil). Biotropica. v. 22, n. 1:36-47. 1990.
BARROS, P. C.; Dinâmica de carbono de uma floresta primária na Resex – Auatí –
Paraná, Fonte Boa, Amazonas-Manaus, 2009. Dissertação (mestrado). Instituto
Nacional de Pesquisas da Amazônia – INPA/UFAM.
BARROS, P. L. C; SILVA JÚNIOR, A. T. Equação de volume para árvores de uma
floresta Tropical densa no município de anapu, oeste do estado do Pará, Amazônia
Oriental. Revista Ciências Agrárias, Belém, n. 51, p.115-126. 2009.
BENTES-GAMA, M. M. Manejo florestal sustentável. Revista da madeira, Curitiba,
n. 116, p. 15-20, 2008.
BERGER, R. PADILHA JUNIOR, J. B. Economia Florestal. Curitiba, Paraná, Editado
pelos autores, 2007, 143 p.
BOTEZELLI, L.; MACEDO, R. L. G.; YOSHITANI JUNIOR, M.; GOMES, J. E.
Relações entre estudos da dinâmica florestal, conservação da biodiversidade e
ecoturismo. Revista científica eletrônica de engenharia florestal. 2005.
BRAZ, E. M. Subsídios para o planejamento do manejo de florestas tropicais na
Amazônia. 2010. 236 f. Tese (Doutorado em Engenharia Florestal) – Universidade
Federal de Santa Maria, Santa Maria, 2010.

BRAZ, E. M.; SCHNEIDER, P. R.; MATTOS, P. P.; SELLE, G. L.; THAINES, F.;
RIBAS, L. A.; VUADEN, E. Taxa de corte sustentável para manejo das florestas
tropicais. Scientia Forestalis v. 22, n. 1, p. 137-145, 2012.
CAMPOS, J. C. C.; LEITE, H. G. Mensuração florestal: Perguntas e respostas. 3 ed.
Viçosa, MG, Editora UFV, 2009, 548 p.
CAMPOS, J. C. C.; RIBEIRO, J. C.; COUTO, L. Emprego da distribuição diamétrica
na determinação da intensidade de corte em matas naturais submetidas ao sistema de
seleção. Revista Árvore, v. 7, n. 2, p. 110-121, 1983.
Carvalho, A. E. F. B. de. Estrutura fitossociológica e pós-estratificação
multidimensional de uma área na Flona do Tapajós - Pará. Dissertação (Mestrado em
Ciências Florestais) - Faculdade de Ciências Agrárias do Pará, Belém. 173p. 1999.
CARVALHO, F. A; FAGG, C.W; FELFILI, J. M. Dinâmica populacional de Acacia
tenuifolia (L.) Willd. em uma floresta decidual sobre afloramentos calcários no Brasil
Central. Scientia Forestalis, v. 38, n. 86, p. 297-306, jun. 2010.
CITADINI-ZANETTE, V. Fitossociologia e aspectos dinâmicos de um remanescente
da Mata Atlântica na microbacia do Rio Novo, Orleans, SC. 1995. 236 f. Tese
(Doutorado em Ecologia e Recursos Naturais) – Universidade Federal de São Carlos,
São Carlos, 1995.
COLPINI, C.; SILVA, V.S.M.; SOARES, T.S.; HIGUCHI, N.; TRAVAGIN, D.P.;
ASSUMPÇÃO, J.V.L. Incremento, ingresso e mortalidade em uma foresta de contato
ombrófla aberta/estacional em Marcelândia, Estado do Mato Grosso, Acta Amazônica,
VOL. 40(3): 549 – 556, 2010.
CORRAR, L. J.; PAULO, E.; DIAS FILHO, J. M. Análise multivariada. FIPECAFI:
Atlas, 2007.
COUTO, H.T.Z.; BASTOS, N.L.M. Modelos de equações de volume e relações
hipsométricas para plantações de Eucalyptus no Estado de São Paulo. IPEF, n.37, p.33-
34, 1987.
CUNHA, U, S,; MACHADO, S, A,; FILHO A, F,; SANQUETTA, C, R, Predição da
estrutura diamétrica de espécies comerciais de terra firme da amazônia por meio de
matriz de transição, Revista Ciência Florestal, v,12, n, 1, p, 109-122, 2002.
DAUBENMIRE, R. Plant communities. Harper & Row, New York. 1968.
DE LIOCOURT, F. De L’aménagement des sapinieres. Bull. Soc. Franche-Comté et
Belfort. 1898.
D’ IVEIRA, M.V. .; BRAZ, E.M. Forest dynamics study of the managed forest of
the PC Peixoto Communit forest management project in western Amazon. Acta
Amazonica, 36 (2):177-182. 2006.

DIAS, A. C.; CUSTÓDIO FILHO, A.; FRANCO, G. A. D. C. Diversidade do
componente arbóreo, em floresta pluvial Atlântica secundária, São Paulo, Brasil.
Revista do Instituto Florestal de São Paulo, v. 12, n. 2, p.127-153, 2000.
ENCINAS, J. I; KLEINN, C. Determinação do volume de madeira em Florestas de
galeria. Brasil florestal, Nº 70, 2001. p. 10.
ENCINAS, J, I; SANTANA, O, A; DE PAULA, J, E; IMAÑA, C, R. Equações de
volume de madeira para o cerrado de Planaltina de Goiás, Revista Floresta, Curitiba, v.
39, n. 1, p. 107-116, 2009.
ESPÍRITO-SANTO, F. D. B., SHIMABUKURO, Y.E., ARAGÃO, L.E.O.; Machado,
E.L.M.; 2005. Análise da composição florística e fitossociológica da floresta nacional
do Tapajós com o apoio geográfico de imagens de satélites. Acta Amazonica 35(2):
155-173.
FERREIRA, C. A. et al. Manejo florestal na Amazônia brasileira: situação atual e
perspectivas. Colombo: Embrapa Florestas, 1999. 20 p. (Embrapa Florestas.
Documentos, 37).
FERREIRA, C. A.; CARVALHO, J. O. P. de; SANTOS, A. F. dos; AZAVEDO, C. P.
de; LIMA, R. M. B. de; NEVES, E. J. M.; SCHWENGBER, D. R.; ARAUJO, H. J. B.
de. Manejo florestal na Amazônia brasileira: situação atual e perspectivas.
Colombo: Embrapa Florestas. 20 p. (Embrapa Florestas. Documentos, 37), 1999.
FERREIRA JÚNIOR, E. V.; SOARES, T. S.; COSTA, M. F. F.; SILVA, V. S. M.
Composição, diversidade e similaridade florística de uma floresta tropical semidecídua
submontana em Marcelândia – MT. Acta Amazônica. vol. 38(4), 673 – 680, 2008.
FINOL, U. H. Nuevos parametros a considerarse en el analisis estrutural de las selvas
virgenes tropicales. Revista Forestal Venezolana, v.18, n.12, p.29-42. 1971.
FONSECA. M. G.; VIDAL, E.; SANTOS, F. A. M. Efeito da exploração madeireira
sobre o número de indivíduos férteis de três espécies arbóreas comerciais na Amazônia
Oriental. Acta Amazônica, v. 38, n.4, p.681-686, 2008.
FRANCEZ, L.M.B.; CARVALHO, J.O.P.; JARDIM, F.C.S.; PINHEIRO, K.A.O.
Efeito de duas intensidades de colheita de madeira na estrutura de uma floresta natural
na região de Paragominas, Pará. Acta Amazonica, Manaus, v.39, n.4, p.857 -864, 2009.
FURNIVAL, G.M. An index for comparing equations used in constructing volume
tables. Forest science, Madison, 7(4): 337-41, 1961.
GAMA, J. R. V.; BOTELHO, S. A.; BENTES-GAMA, M. M.; SCOLFORO, J. R. S.
Estrutura e potencial futuro de utilização da regeneração natural de Floresta de Várzea
Alta no município de Afuá, estado do Pará. Ciência Florestal, Santa Maria, v. 13, n. 2,
2003.

GARCIA, C. C. Regeneração natural de espécies arbóreas em fragmento florestal
da zona da mata mineira. 2009. 83 p. Dissertação (Mestrado em Ciências Florestais) –
Universidade Federal de Viçosa, Minas Gerais, 2009.
GIRARD, E. A. Volume, biomassa e rendimento de óleos essenciais do Craveiro
(pimenta pseudocaryophyllus (gomes) landrum) em floresta ombrófil a mista. 72f.
Dissertação (Mestrado em Ciências Florestais), Universidade Federal do Paraná, 2005.
GOMES, F. P.; GARCIA, C. H. A Determinação de equações volumétricas na
Engenharia Florestal. Série Técnica IPEF, Piracicaba, 9(26): 1-36, mar. 1993.
GOMIDE, G. L. A. Estrutura ë dinâmica de crescimento De florestas tropicais
primária e secundária no estado do Amapá. 1997. Dissertação (Doutorado em
Ciências Agrárias) – Universidade Federal do Paraná, Curso de Pós-Graduação em
Engenharia. Paraná, 1997.
GONÇALVES, F. G.; SANTOS, J.R.; LOPES, A. L. B. Análise de dados sar para
estimativa volumétrica da floresta tropical em regime de exploração madeireira.
VII seminário em atualização em sensoriamento remoto e sistemas de informações
geográficas aplicados a engenharia florestal. p. 42-48. 2008.
GONÇALVES, F. G.; SANTOS, R. Composição florística e estrutura de uma unidade de
manejo florestal sustentável na Floresta Nacional do Tapajós, Pará. Acta Amazônica.
vol. 38(2), 229 – 244. 2008.
GUIMARÃES, J. R. S.; CARIM, M. J. V. Análise fitossociológica e florística em três
hectares de floresta tropical ombrófila densa na reserva de desenvolvimento sustentável
do Rio Iratapuru. Revista pesquisa de iniciação científica – Amapá. P. 32-34, 2008.
HIGUCHI, N.; SANTOS, J.; SILVA, R. P.; LIMA, A. N.; TEIXEIRA, L. M.;
CARNEIRO, V. M. C.; FELSEMBURGH, C. A; TRIBUZY, E. S. Noções Básicas de
Manejo Florestal. Manaus: INPA/ CPST/ LMF, 2008.
HIGUCHI, N.; CHAMBERS, J.Q.; SANTOS, J.; RIBEIRO, R.J.; PINTO, A.C.M.;
SILVA, R.P.; ROCHA, R.M.; TRIBUZY, E.S. Carbon balance and dynamics of
primary vegetation in the Central Amazon. Floresta, 34 (3): 377-384. 2004.
HIGUCHI, N.; SANTOS, J.; LIMA, A.J.N. Biometria Florestal. INPA, Manuas-AM.
P-14. 2008.
HIRAI, E. H.; CARVALHO, J. O. P.; PINHEIRO, K. A. O. Estrutura da população de
maçaranduba ( Manilkara huberi Standley) em 84 ha de floresta natural na fazenda Rio
Capim, Paragominas, PA. Rev. cienc. agrar., Belem, n. 49, 65-76p. 2008.
HOSOKAWA, R. T.; MOURA, J. B.; CUNHA, U. S.; Introdução ao manejo e
economia de florestas. Curitiba, Paraná: Editora UFPR, 2008, 164 p.
HOSOKAWA, R. T.; MOURA, J. B.de; CUNHA, U. S. da. Introdução ao manejo e
economia de florestas. Curitiba: Ed. da UFPR, 1998.

IBGE. Manual Técnico da Vegetação Brasileira. Instituto Brasileiro de Geografia e
Estatística, Rio de Janeiro, 2012.
IEF. 2009. Relatório de Inventário Florestal Amostral da Floresta Estadual do
Amapá. Macapá.
INAM. Instituto Natureza Amazônica. Relatório técnico de inventario florestal na
Floresta Estadual do Amapá. Instituto Estadual de Florestal (IEF), 2009.
JARDIM, F. C. S.; SOARES, M. S. Comportamento de Sterculia pruriens (Aubl.)
Schum. em floresta tropical manejada em Moju-PA. Acta Amazonica, Manaus, v. 40,
n.3 535 – 542p. 2010.
KERKHOFF, D. T.; RIL, F. L.; ORO, D.; FORTES, F. O. Regeneração natural
estabelecida em um fragmento da floresta estacional decidual. Ciências Florestal,
Santa Maria, v.5, p. 100-107. 2011.
KOHLER, H.-P., F. Billari e J. A. rtega, “The emergence of lowest-low fertility in
Europe during the 1990s”, in Population and Development Review, 28 (4), pp. 641-
680, 2002.
KUNZ, S. H.; IVANAUSKAS, N. M.; MARTINS, S. V.; SILVA, E.; STEFANELLO, D.
Aspectos florísticos e fitossociológicos de um trecho de Floresta Estacional Perenifólia na
Fazenda Trairão, Bacia do rio das Pacas, Querência-MT. Acta Amazônica. vol. 38(2), 245 –
254, 2008.
LEITE, F. S. Estimativa do volume de madeira a partir do Diâmetro da cepa em uma
área explorada de Floresta amazônica de terra firme. 2008, 74f. Dissertação (Mestrado
em Ciências Florestais), Universidade de Brasília, DF.
LEITE F. S; REZENDE, A. V. Estimativa do volume de madeira partindo do diâmetro
da cepa em uma área explorada de floresta Amazônica de terra firme. Revista Ciência
Florestal, Santa Maria, v. 20, n. 1, p. 71-82. 2010.
LEITE, H. G.; ANDRADE, V. C. L. Um método para condução de inventários
florestais sem o uso de equações volumétricas. Revista Árvore, v.26, n.3, p.321-328,
2002.
LEITE, H. G.; ANDRADE, V. C. L. Importância das variáveis altura dominante e altura
total em equações hipsométricas e volumétricas. Revista Árvore, Viçosa, v. 27, n. 3, p.
301-310, 2003
LEPSCH, I. F. Formação e Conservação dos Solos. São Paulo: Oficina de Textos,
2002.
LIMA, A. J. N. Avaliação de um sistema de inventário florestal contínuo em áreas
manejadas e não manejadas do estado do amazonas (AM). 2010. 181 p. Tese
(Doutorado em Ciências de Florestas Tropicais) – Instituto Nacional de Pesquisas da
Amazônia, Manaus.

LIMA FILHO, D.A.; MATOS, F.D.A.; AMARAL, I.L.; REVILLA, J.; COÊLHO, L.S.;
RAMOS, J.F.; SANTOS, J.L. Inventário florístico de floresta ombrófila densa de terra
firme, na região do Rio Urucu-Amazonas, Brasil. Acta Amazonica, v.31, p.565-579.
2001.
LIMA-FILHO, D.A; REVILLA, J., AMARAL, I. L. do. Aspectos florísticos de 13
hectares da área de Cachoeira Porteira-PA. Acta Amazonica, vol.34, n.3, p.415-423,
2004.
MACHADO, S. A. ; MELLO, J. M.; BARROS, D. A. Comparação entre métodos para
avaliação de volume total de madeira por unidade de área, para o pinheiro do paraná, na
região sul do Brasil. Cerne, v. 6, n. 2, p. 55-66, 2000.
MANOKARAN, N.; KOCHUMMEN, K. M. Recruitment, growth and mortality of tree
species in a lowland dipterocarp forest in Peninsular Malaysia. Journal of Tropical
Ecology, Cambridge, v. 3, n. 4, p. 315-330, 1987.
MANTOVANI, W.; SANTOS, R. F. dos. Vegetação, vulnerabilidade e qualidade
ambiental. In: SANTOS, R. F. dos (org.). Vulnerabilidade Ambiental. Brasília: MMA,
p. 144-164, 2007.
MARANGON, L. C. Florística e fitossociologia de área de floresta estacional
semidecidual visando dinâmica de espécies florestais arbóreas no município de
Viçosa – MG. 1999. 139 f. Tese (Doutorado em Ecologia e Recursos Naturais) –
Universidade Federal de São Carlos, São Carlos, 1999.
MARAGON, L. C; SOARES, J. J; FELICIANO, A. L. P; SILVA, C. F. L. Reneração
natural em um fragmento de floresta estacional semidecidual em Viçoça, Minas Gerais.
Revista Árvore, v.32, n.1, p.183-191, 2008.
MARANGON, L. C., SOARES, J. J. e FELICIANO, A. L. P. Florística arbórea da Mata
da Pedreira, município de Viçosa, Minas Gerais. Rev. Árvore, v.27, n. 2, p.207-215,
2003.
MARGALEF, R. Homage to Evelyn Hutchinson or why is there an upper lilit to
diversity. Transactions of the Illinois State Academy of Science, 44, p. 221-235, 1972
MARRA, D. M. Sucessão florestal em área atingida por tempestade convectiva na
Região de Manaus, Amazônia Central. 2010. 106 f. Dissertação (Mestrado em
Ciências de Florestas Tropicais) – Instituto Nacional de Pesquisas da Amazônia,
Manaus.
MARTINS, S. V. Recuperação de áreas degradadas: Ações em Áreas de
Preservação Permanentes, Voçorocas, Tabulares Rodoviários e de Mineração. 2
Ed. Viçosa – MG, Editora Aprenda Fácil, 2010.
MARTINS, S. V.; RODRIGUES, R.R. Gap-phase regeneration in a semideciduous
mesophytic forest, south-eastern Brazil. Plant ecology, London, v. 163, p. 51-62, 2002.

MENDONÇA, A. C. A. Caracterização e Simulação dos Processos Dinâmicos de
uma Área de Floresta Tropical de Terra Firme Utilizando Matrizes de Transição,
79f. Dissertação (mestrado). Universidade Federal do Paraná. 2003.
MANOKARAN, N,; KOCHUMMEN, K, M, Recruitment, growth and mortality of tree
species in a lowland dipterocarp forest in Peninsular Malaysia, Journal of Tropical
Ecology, Cambridge, v, 3, n, 4, p, 315-330, 1987.
MOSCOVICH, F. A. Dinâmica de crescimento de uma floresta ombrófila Mista em
nova prata, RS. 2006. 130 p. Tese (Doutorado em Ciências Florestais) – Universidade
Federal de Santa Maria, Santa Maria, 2006.
NEMEC, A, F, L, Analysis of repeated measures and time series: an introduction
with forestry examples, Victoria, B,C, Work, 90p, 1996.
NETO, A. C. F.; SCÁRDUA, F. P.; JACINTO, J. M. M. Rede de inovação tecnológica
para o setor madeireiro da Amazônia Legal. Brasília, DF: Centro de Gestão e Estudos
Estratégicos. 116 p, 2010.
ODUM, E.P. Fundamentos de Ecologia. Fundação Calouste Gulbenkian. 4 ed. Lisboa,
1988, 927p.
OLIVEIRA, A. N.; AMARAL, I. L.; RAMOS, M. B. P.; NOBRE, A. D.; COUTO, L.
B.; SAHDO, R. M. Composição e diversidade florístico-estrutural de um hectare de
floresta densa de terra firme na Amazônia Central, Amazonas, Brasil. Acta Amazonica,
Manaus, v. 38, n. 4, 627-642 p, 2008.
OLIVEIRA, A. N. & AMARAL, I. L. do. Aspectos florísticos, fitossociológicos e
ecológicos de um sub-bosque de terra firme na Amazônia Central, Amazonas, Brasil.
Acta Botanica, 35(1): 1-16. 2005.
OLIVEIRA, A. N.; AMARAL, I.L. Floristica e fitossociologia de uma floresta de
vertente na Amazonia Central, Amazonas, Brasil. Acta amazonica. v. 34, n.1, 21-34 p,
2004.
OLIVEIRA, E.B. de; MARANGON, L.C.; FELICIANO, A.L.P.; FERREIRA,
R.L.C.; RÊGO, P.L. Estrutura fitossociológica de um fragmento de mata ciliar, Rio
Capibaribe Mirim, Bacia do Rio Goiana, Aliança, Pernambuco. Revista Brasileira de
Ciências Agrárias, v.4, n.2, p.167-172, 2009.
OLIVEIRA, L.C. Efect of logging and thinning intensities on the dynamic of vegetation
of an area of 136 ha in the Tapajos National Forest. Tese de Doutorado, Escola
Superior de Agricultura Luiz de Queiroz, Piracicaba. 183 pp. 2005.
PANTALEÃO, J. C. Avaliação da estrutura, crescimento, mortalidade e recrutamento
em uma floresta semidecidual submontana na Amazônia mato-grossense. 2008. 32 p.
Monografia (Graduação em Engenharia Florestal) – Universidade Federal de Mato
Grosso, Cuiabá, 2008.

PEREIRA, L. A.; SOBRINHO, F. A. P.; COSTA NETO, S. V. Florística e estrutura de
uma mata de terra firme na reserva de desenvolvimento sustentável Rio Iratapuru,
Amapá, Amazônia Oriental, Brasil. Floresta, Curitiba, v. 41, n.1, p.113-122, 2011.
PEREIRA, L. A; SENA, K. S; SANTOS, M. R; COSTA NETO, S. V. Aspectos
florísticos da Flona do Amapá e sua a importância na conservação da biodiversidade.
Revista Brasileira de Biociências – Nota Científica. v. 5, n.2, p.693 – 695, 2007.
PHILLIPS, O.; BAKER, T. Manual de Campo para a Remedição e Estabelecimento
de Parcelas (RAINFOR). 2006.
PINTO, A. C. M. Dinâmica de uma floresta de terra firme manejada experimentalmente
na região de Manaus (AM). 2008. 167 f. Tese (Doutorado em Ciências Florestais
Tropicais) – Instituto Nacional de Pesquisas da Amazônia/Universidade Federal do
Amazonas, Manaus.
PINTO, A. C. M.; SOUZA, A. L.; SOUZA, A. P.; MACHADO, C. C.; MINETTE, L.
J.; VALE, A. B. Análise de dados de colheita de madeira em floresta tropical úmida sob
regime de manejo florestal sustentado na Amazônia Ocidental. R. Árvore, Viçosa-MG,
v. 26, n. 4, p. 459-466, 2002.
POSSIMOSER, D.; JESUS, A. G. de; RIBEIRO, S. B.; CAVELHEIRO, C. S.; ALVES,
W. C. Levantamento fitossocioológico de espécies nativas com potencial produtivo de
fitoterápicos na Zona da Mata rodoniense. Revista Brasileira de Ciências da
Amazônia, vl, n1-2012.
PRANCE, G.T.; RODRIGUES, W.A.; SILVA, M.F. Inventário florestal de um hectare
de mata de terra firme, km 30 da estrada Manaus-Itacoatiara. Acta Amazonica, 6:9-35.
1976.
PROCÓPIO, L. C.; SECCO, R. S. A importância da identificação botânica nos
inventários florestais: o exemplo do “tauari” (Couratari spp. e Cariniana spp. -
Lecythidaceae) em duas áreas manejadas no estado do Pará. Acta Amazonica, Manaus,
v. 38, n. 1: 31 – 44p. 2008.
QUEIROZ, J. A. L; MACHADO, S. A. Fitossociologia em Floresta de Várzea do
Estuário Amazônico no Estado do Amapá. Pesquisa Florestal Brasileira, n 57, p. 05-
20, 2008.
RADAM. Projeto Radam Brasil. Folha SB22 Araguaia e parte da folha SC 22
Tocantins. Instituto Brasileiro de Geografia e Estatística, Rio de Janeiro. 1974.
RAYOL, B.P.; ALVINO, F. O.; SILVA, F.F. Estrutura e composição florística da
regeneração natural de duas florestas secundárias em Capitão Poço, Pará, Brasil.
Amazônia: Ci. & Desenv., Belém, v. 4, n. 7, jul./dez. 2008.
RIBEIRO, G. H. P. M.; FELFILI, J. M. Regeneração natural em diferentes ambientes da
Mata de Galeria do Capetinga, na Fazenda Água Limpa - DF. Revista Cerne, Lavras, v.
15, n.1, p. 1 - 9, 2009.

RIVERA, H. Ordenamento territorial de áreas florestais Utilizando avaliação
multicritério apoiada por geoprocessamento, fitossociologia e análise multivariada.
Dissertação de Mestrado, Pós-Graduação em Engenharia Florestal do Setor de Ciências
Agrárias da Universidade Federal do Paraná, Curitiba, Paraná. 212p. 2007.
ROCHA, K.; MOREIRA, A. R. B.; CARVALHO, L.; REIS, E. J. O valor das
concessões nas florestas nacionais da Amazônia. Texto pra Discussão Nº37. Instituto de
Pesquisa Econômica Aplicada , 2000.
ROCHA, R. M. Taxas de recrutamento e mortalidade da floresta de terra firme da bacia
do Rio Cuieiras na região de Manaus – AM. Master Thesis. INPA/UFAM. Manaus.
2001.
ROLIM, S, G; COUTO, H, T, Z; JESUS, R, M; FRANÇA, J, T. Modelos volumétricos
para a Floresta Nacional do Tapirapé-Aquirí, Serra dos Carajás (PA), Acta
Amazonica, Vol, 36(1)p. 107-114, 2006.
ROSSI, L. M. B.; KOEHLER, H. S.; SANQUETTA, C. R. ARCE, J. E. Modelagem de
mortalidade em florestas naturais. Floresta, Curitiba, v. 37, n. 2, p. 275–291, 2007.
SALOMÃO, R. P.; SANTANA, A. C.; BRIENZA JÚNIOR, S.; GOMES, V. H. F.
Análise fitossociológica de floresta ombrófila densa e determinação de espécies-chave
para recuperação de área degradada através da adequação do índice de valor de
importância. Bol. Mus. Pará. Emílio Goeldi. Cienc. Nat., Belém, v. 7, n. 1, p 57-102.
2012.
SALOMÃO, R. P.; SANTANA, A. C.; COSTA NETO, S. V. Construção de índices de
valor de importância de espécies para análise fitossociológica de floresta Ombrófila
através de análise multivariada. Floresta, Curitiba, PR, v. 42, n. 1, p. 115 - 128,
jan./mar. 2012.
SAMPAIO, P. T. B.; BARBOSA, A. P.; VIEIRA, G.; SPIRONELLO, W.R.; BRUNO,
F. M. S. Biomassa da rebrota de copas de pau-rosa (Aniba rosaeodora Ducke) em
plantios sob sombra parcial em floresta primária. Acta Amazonica, Manaus, v. 35, n.4 :
491 – 494p. 2005.
SANTOS, C. S. dos; CHIOSSI, R. Y.; AVILA, A. L. de; GASPARIN, E. Levantamento
florístico e fitossociológico de um fragmento florestal no município de Faxinal dos
Guedes, SC. Unoesc & Ciência – ACET, Joaçaba, v. 3, n. 1, p. 7-22, jan./jun. 2012.
SANTOS, K; SANQUETA, C.R; EISFIELD, R.; WATZLAWICK, L.F.; ZILIOTTO,
M.A. Equações volumétricas por classe diamétrica para algumas espécies folhosas da
floresta ombrófila mista no Paraná, Brasil. Revista Ciências Exatas e Naturais, v.8, n.1,
p.2-8, 2006.
SANTOS, V. S. Estrutura horizontal de uma área de terra firme na Floresta
Estadual do Amapá – AP. 2011. 36f. Monografia (Graduação em Engenharia
Florestal) - Universidade do Estado do Amapá, Macapá, 2011.

SANTOS, W.C. Análise de características dendrométricas e fatores edáficos no
crescimento e produção de um povoamento de Araucaria angustifólia (Bertol.)
Kuntze. 2006. 124 f. Dissertação (Mestrado em Engenharia Florestal) – Universidade
Federal do Paraná. Curitiba.
SAWCZUK,A. R. Alteração na composição florística de um fragmento de Floresta
Ombrófila mista na região centro-sul do estado do Paraná. 2009. Dissertação
(Mestrado em Ciências Florestais), UNICENTRO, Paraná, 2009.
SCHNEIDER, P. R. Manejo Florestal: Planejamento da Produção Florestal.
Universidade Federal de Santa Maria, Centro de Ciências Rurais Departamento de
Ciências Florestais, 2008.
SCHNEIDER, P. R.; FINGER, C. A. G. Manejo Sustentado de Florestas
Inequiâneas. 2000.
SCHNEIDER, P.R.; SCHNEIDER, P.S.P. & SOUZA, C.A.M. Análise de regressão
aplicada à engenharia florestal. Santa Maria, Universidade Federal de Santa Maria,
2009. 294p.
SCHORN, L. A.; GALVÃO, F. Dinâmica da regeneração natural em três estágios
Sucessionais de uma floresta ombrófila densa em Blumenau, SC. FLORESTA,
Curitiba, PR, v. 36, n. 1, jan./abr. 2006.
CHÜTZ, J. Dynamique et conditions d’équilibre des peuplements jardinées sur les
stations de la hêtraie à sapin. Schweizerische Zeitschrift für Forstwesen v. 126, n. 9,
p. 637–671, 1975.
SCOLFORO, J. R. S. Manejo florestal. Lavras: UFLA/FAEPE, 1998. 438 p.
SCOLFORO, J. R. S.; PULZ, F. A.; MELLO, J. M.; OLIVEIRA FILHO, Ary Teixeira
de. Modelo de produção para floresta nativa como base para manejo sustentado.
CERNE (UFLA), LAVRAS, v. 2, n.1, p. 112-137, 1996.
SILVA, A.P.F.F. da; BENTES-GAMA, M. de M. Fitossociologia de uma Floresta
Ombrófila Aberta em área de assentamento rural no distrito de Jaci-Paraná, Porto
Velho, Rondônia. Ambiência, Santa Cruz, v.4, p. 343-352, 2008.
SILVA JUNIOR, J. F. ; MARANGON, L. C. ; FELICIANO, A. L. P. ; FERREIRA, R.
L. C. ; BRANDÃO, C.F.L.S . Estudo Fitossociológico de Espécies Florestais Arbóreas
em um Fragmento de Floresta Atlântcia no Cabo Santo Agostinho PE. Ciências
Agrárias e Ambientais (UFAM), Piracicaba/SP, v. 1, p. 1, 2008.
SILVA, A. S. ; APARICIO, W. C. S ; APARICIO, P. S ; BATISTA, A. P. B ; MATOS
FILHO, J. R. ; LIMA, R. B. . Estrutura, distribuição espacial e volumetria da
Carapa guianensis Aubl. em floresta de várzea no estado do amapá, brasil. In: 5
simpósio latino-americano de manejo florestal, 2011, santa maria,rs. 5 simpósio latino-
americano de manejo florestal: sustentabilidade ambiental, 2011.

SILVA, J. N. M.; LOPES, J. C. A.; OLIVEIRA, L. C.; SILVA, S. M. A.; CARVALHO,
J. O. P.; COSTA, D. H. M.; MELO, M. C.; TAVARES, M. J. M. Diretrizes para
instalação e medição de parcelas permanentes em florestas naturais da Amazônia
Brasileira. Belém:Pará. ed 1. 2005
SILVA, G. O. Modelos de regressão quando a função de taxa de falha não é
monótona e o modelo probabilística beta Weibull modificado. Tese (Doutorado em
Agronomia) – Universidade de São Paulo, Escola Superior de Agricultura Luiz de
Queiroz, São Paulo. 2008.
SILVA, J, N, M; ARAÚJO, S, M. Equação de volume para árvores de pequeno
diâmetro, na Floresta nacional do tapajós, Boletim de Pesquisa Florestal, Colombo, n,
8/9, p, 16-25. 1984.
SILVA, K. E; MATOS, F. D. A; FERREIRA, M. M. Composição florística e
fitossociologia de espécies arbóreas do Parque Fenológico da Embrapa Amazônia
Ocidental. Acta Amazonica. v. 38. n.2, p. 213-222, 2008.
SILVA R.D. Indicadores de recuperação ambiental em diferentes coberturas
florestais, Alegre-ES [dissertação]. Jerônimo Monteiro: Universidade Federal do
Espírito Santo; 2012.
SILVA, S. O et al. Regeneração natural em um remanescente de caatinga com
Diferentes históricos de uso no agreste pernambucano Revista Árvore, Viçosa-MG,
v.36, n.3, p.441-450, 2012.
SILVA, W. C.; MARANGON, L. C.; FERREIRA, R. L. C.; FELICIANO, A. L. P.;
COSTA JUNIOR, R. F. Estudo da regeneração natural de espécies arbóreas em
fragmento de floresta ombrófila densa, Mata das Galinhas, no município de Catende,
Zona da Mata Sul de Pernambuco. Ciência Florestal, Santa Maria, v. 17, n. 4, p. 321-
331, 2007.
SOUZA, D. R.; SOUZA, A. L.; LEITE, H. G.; YARED, J. A. G. Análise estrutural em
floresta ombrófila densa de terra firme não explorada, Amazônia Oriental. Revista
Árvore, Viçosa, v. 30, n. 1, p 75-87, 2006.
SOUZA, D. R.; SOUZA, A. L.; SILVA, M. L.; RODRIGUES, F. L. Ciclo de corte
econômico ótimo em floresta ombrófila densa de terra firme sob manejo florestal
sustentável, Amazônia Oriental. Revista Árvore, Viçosa, v.28, n.5, p.681-689, 2004.
SOUZA, E. B.; CUNHA, A. C. Climatologia de Precipitação no Amapá e Mecanismos
Climáticos de Grande Escala. In: CUNHA, A. C.;SOUZA, E. B., et al (Ed.). Tempo,
clima e recursos hídricos: resultados do Projeto REMETAP no Estado do Amapá.
Macapá: IEPA, p.216 p, 2010.
STERBA, H. Equilibrium Curves and Growth Models to Deal with Forests in Transition
to Uneven-Aged Structure – Application in Two Sample Stands. Silva Fennica,
Vienna, v. 38, n. 4, p 413-423, 2004.

TARDIN, A. T.; RABELO, B. V.; FACUNDES, F. S.; ALMEIDA, J. C.; ÁVILA, J. E.
S.; LOBATO JUNIOR, J. P. A.; PICANÇO, J. R. A.; SILVA, L. M. S.; GUEDES, L. A.
C.; OLIVEIRA, M. R. M.; DIAS, M. C. S.; MELO, R. M. S.; FERREIRA, T. J. M.
Plano de recuperação do Pa Nova Canaã. Porto Grande. 2009.
TEIXEIRA, L. M.; CHAMBERS, J. Q.; SILVA, A. R.; LIMA, A. J. N.; CARNEIRO,
V. M. C.; SANTOS, J.; HIGUCHI, N. Projeção da dinâmica da floresta natural de
Terra-firme, região de Manaus-AM, com o uso da cadeia de transição probabilística de
Markov. Acta Amazônica, v. 37, n. 3, p 377 – 384. 2007.
THAINES, F.; BRAZ, E. M.; MATTOS, P. P.; THAINES, A. A. R. Equações para
estimativa de volume de madeira para a região da bacia do Rio Ituxi, Lábrea, AM.
Pesquisa Florestal Brasileira. Colombo, v. 30, n. 64, p. 283-289, 2010.
THOMAZIELLO, S. Uso da terra e sua influência sobre a qualidade ambiental. In:
SANTOS,R. F. dos (org.). Vulnerabilidade Ambiental. Brasília: MMA, p. 23-38,
2007.
TOLEDO, J.A.; KAMINSKI, J.; SANTANNA, M.A. & SANTOS, D.R. Tampão Santa
Maria (TSM) como alternativa ao tampão SMP para medição da acidez potencial de
solos ácidos. R. Bras. Ci. Solo, 36:427-435, 2012.
TONINI, H.; ARCO-VERDE, M.F.; SÁ, S.P. Dendrometria de espécies nativas em
plantios homogêneos no Estado de Roraima - andiroba (Carapa guianensis Aubl),
castanha-doBrasil (Bertholletia excelsa Bonpl.), ipê-roxo (Tabebuia avellanedae
Lorentz ex Griseb) e jatobá (Hymenaea courbaril L.). Acta Amazônica, v.35, n.3,
p.2005.
TONINI, H.; MELO, S.R. Equações volumétricas para espécies nativas e exóticas no
município do Cantá (RR). Boletim de Pesquisa e desenvolvimento, 2009.
VACCARO, S.; FINGER, C. A. G.; SCHNEIDER, P. R.; LONGHI, S. J. Incremento
em área basal de árvores de uma floresta estacional decidual, em três fases sucessionais,
no município de Santa Tereza, RS. Ciência Floresta, v.13, n. 2, p 131-142. 2003.
VELOSO, H. P.; RANGEL FILHO, A. L. R.; LIMA, J. C. A. Classificação da
vegetação brasileira adaptada a um sistema universal. Rio de Janeiro: IBGE,
Departamento de Recursos Naturais e Estudos Ambientais. 1991, 124p.
VIDAL, E,; GERWING, J. J Ecologia e manejo de cipós na Amazônia Oriental.
Belém: Imazon. 141p. 2003.
VIDAL, E.; VIANA, V. M.; BATISTA, J. L. F. Crescimento de floresta tropical três
anos após colheita de madeira com e sem manejo florestal na Amazônia oriental.
Scientia Forestalis. n. 61, p. 133-143, 2002.
VIEIRA, S.A. Global changes and tree growth rate in the Amazon forest. Tese de
Doutorado, Escola Superior de Agricultura Luiz de Queiroz, Piracicaba. 133 pp. 2003.

VOLPATO, M. M. L. Regeneração natural em uma floresta secundária no domínio
de Mata Atlântica: uma análise fitossociológica. 1994. 123f. Dissertação (Mestrado
em Ciência Florestal) - Universidade Federal de Viçosa, Viçosa, 1994.
YANG, Y.; HUANG, S.; MENG, S. X. Development of a tree-specific stem profile
model for white spruce: a nonlinear mixed model approach with a generalized
covariance structure. Forestry, Vol. 82, No. 5, 2009.
YARED, J.A.G.; SOUZA, A.L. Análise dos impactos ambientais no manejo de florestas
tropicais. Viçosa: UFV: Sociedade de Investigações Florestais. 38. 1993
ZANETTI, E.; SIQUEIRA, J. Manejo florestal sustentável na amazônia brasileira:
monitoramento de indicadores silviculturais. Ciências Florestal, Santa Maria, v.5,
p.137-147. 2011.
ZEE - Macrodiagnóstico do Estado do Amapá: primeira aproximação do ZEE/Equipe
Técnica do ZEE-AP, Macapá:, 2008.


















