
Em meio à caatinga e às onças, uma barraginha faz a diferença!
Upload
truongthienCategory
view
223download
0
Pagamentos por Serviços Ambientais no Corredor das Onças - Remuneração pela
conservação de Puma concolor e conservação de água mediante adequação ambiental em
propriedades rurais na Região Metropolitana de Campinas (RMC).
Resultados parciais da análise da dieta de felinos nos fragmentos florestais selecionados da Região Metropolitana de Campinas.
Claudia Zukaran Kanda*1
Gabriela de Lima Fregonezi*
Helena Alves do Prado*
Mitra Katherina Ferreira*
Marcelo Magioli2
Milton Cezar Ribeiro*
Márcia Rodrigues3
Outubro de 2012
1* Laboratório de Ecologia Espacial e Conservação (LEEC), Depto de Ecologia, IB, UNESP, Rio Claro. 2 Mestrando do Depto de Ciências Florestais, Escola Superior de Agricultura Luiz de Queiroz, USP. 3 Analista ambiental, ARIE Matão de Cosmópolis, ICMBio/MMA.

COMO CITAR ESTE RELATÓRIO:
Kanda, C.Z; Fregonezi, G. de L.; Prado, H.A. do; Ferreira, M.K.; Magioli, M.;
Ribeiro, M.C.; Rodrigues, M.G. 2012. Resultados parciais da análise da dieta de felinos
nos fragmentos florestais selecionados da Região Metropolitana de Campinas.
Relatório técnico, Campinas, Outubro de 2012.
SOBRE O PROJETO:
O projeto PSA – Corredor das Onças tem coordenação geral do Prof. Dr.
Ademar Romeiro (IE/UNICAMP) e coordenação executiva da Profa. Dra. Márcia
Gonçalves Rodrigues (ICMBio/MMA).

I. INTRODUÇÃO
A região Metropolitana de Campinas (RMC) está inserida no domínio da Mata
Atlântica. Esse bioma é considerado como uma área prioritária para conservação já
que se encontra altamente fragmentado, restando menos de 12% de sua vegetação
original e em sua maioria remanescentes com menos de 50 ha (RIBEIRO et al., 2009).
Além disso, ela é classificada como hotspot de biodiversidade (MYERS et al., 2000)
Após o acelerado processo de urbanização e expansão da agricultura nos anos
de 1970 a 2000 (BAENINGER, 2001 e MIRANDA, 2012), a RMC sofreu uma drástica
redução de sua vegetação nativa, o que levou consequentemente a um elevado grau
de fragmentação e isolamento de fragmentos florestais (CANDIDO NUNES, 2010).
Além da redução do habitat, a caça é uma das principais causas de extinções
locais em áreas fragmentadas (CULLEN JR., BODMER e VALLADARES-PÁDUA, 2001),
sendo especialmente prejudicial a espécies com ciclo de vida longo, baixas taxas de
crescimento e prole reduzida, como os mamíferos de maior porte (BODMER,
EISENBERG e REDFORD, 1997).
Embora a cobertura vegetal de Campinas esteja numa situação crítica, o
município ainda é tradicionalmente reconhecido em função das áreas verdes que
possui, constituídas tanto por remanescentes naturais de Mata Atlântica como por
parques e bosques. Portanto, essa região ainda possui grande relevância biológica por
abrigar centenas de espécies, dentre elas carnívoros (MIRANDA, 2012).
A estabilidade de comunidades animais relaciona-se a como diferentes espécies
repartem os recursos disponíveis (MACARTHUR e LEVINS 1967; GORDON, 2000).
Segundo Schoener (1974), a teoria de partição de nicho indica que populações não são
capazes de coexistir no meio por um tempo significativo, se os recursos
compartilhados forem os mesmos. Dessa forma, mamíferos carnívoros simpátricos
geralmente exibem discrepâncias em tamanho corporal ou de caracteres, capacitando-
os a explorar diferentes nichos (ARJO, PLETSCHER e REAM, 2002; DAVIES et al., 2007;
FRISCIA, VAN VALKENBURGH e BIKNEVICIUS, 2007). Além disso, a adoção de
estratégias diferentes de caça e a formação de grupos cooperativos também são

comportamentos visualizados nesse grupo de mamíferos (PIERCE, BLEICH e BOWYER,
2000).
Os carnívoros desempenham na cadeia alimentar o papel de predadores de
topo de cadeia, atuando como controladores das populações de animais de níveis
tróficos intermediários (DONATTI, GUIMARÃES JR. e GALETTI, 2009). Devido ao uso de
extensas áreas de mata contínua para manutenção de suas populações (WILSON e
WILLIS, 1975 apud CHIARELLO, 1999) e baixa densidade demográfica (BRASIL, 2003),
são mais suscetíveis ao desaparecimento, sendo substituídos por mesopredadores de
hábito generalista (LAURANCE, 1993; CROOKS e SOULÉ, 1999), com maior capacidade
de sobreviver em ambientes alterados (GEHRING e SWIHART, 2003).
Apesar da variação corporal, sua arquitetura corporal é fortemente conservada
na história evolutiva, o que pode ser considerado um reflexo de seu comportamento
alimentar (NOWAK, 2005; BARYCKA, 2007). Os dentes caninos são fortes e
pronunciados, os molares especializados e os incisivos reduzidos em tamanho, cada
um servindo a funções específicas (SUNQUIST e SUNQUIST, 2002).
No Brasil, a Ordem Carnivora está representada por 29 espécies (CHEIDA,
2006). Na área de estudo são registradas cinco espécies de felinos e duas espécie de
canídeo, sendo descritos abaixo:
Puma concolor
Conhecida popularmente como onça parda ou suçuarana, é o felídeo de maior
área de distribuição no continente americano, ocorrendo do oeste do Canadá ao
extremo sul do continente sul-americano e por todo o Brasil. Está presente em todos
os biomas brasileiros e possui adaptação a diversos tipos de ambientes e clima
(CIOCHETE, 2007). A espécie é classificada como Vulnerável no Brasil (BRASIL, 2003). A
suçuarana é a segunda maior espécie de felídeo no Brasil, com comprimento total
variando de 155,4 a 169,9 cm e peso de 22,0 a 70,0 kg. Possui hábitos solitários e
terrestres, com atividade predominantemente noturna. Em seu trabalho, Oliveira
(2002), concluiu que as suçuaranas possuem papel de predador de topo e
mesopredador, pois consumem, na maioria das vezes, mamíferos de médio porte (1-

15kg), mas muitas presas pequenas também são consumidas, como roedores, répteis e
aves (CHEIDA, 2006).
Puma yagouaroundi
O gato-mourisco apresenta massa corpórea entre 3,8 e 6,5 kg. Sua coloração é
uniforme e polimórfica e varia do marrom ao cinza-escuro e avermelhado. De ampla
distribuição geográfica, alcança desde os Estados Unidos até o norte da Argentina e é
registrado em todos os biomas brasileiros (SUNQUIST e SUNQUIST, 2002; CHEIDA et al.
2006).
Estudos a respeito de sua dieta indicam o consumo frequente de pequenos
mamíferos, em menor frequência de aves, répteis e artrópodes (KONECNY, 1989;
(SUNQUIST e SUNQUIST, 2002). A atividade do gato-mourisco foi descrita como
essencialmente diurna (CHEIDA et al., 2006).
Leopardus tigrinus
O gato-do-mato é a menor espécie do gênero, com massa corporal variando
entre 1,8 e 3,4 kg (SUNQUIST e SUNQUIST, 2002). Sua coloração varia em tonalidades
de amarelo e castanho, com rosetas ao longo do corpo (CHEIDA et al., 2006). Pode
apresentar alto teor de melanismo, tornando o corpo enegrecido (OLIVEIRA e
CASSARO, 2005).
Distribui-se desde a América Central até o norte da Argentina. No Brasil, é
registrada na Amazônia, Caatinga, Cerrado, Pantanal, Mata Atlântica e Campos Sulinos
(SUNQUIST e SUNQUIST, 2002; CHEIDA et al., 2006). Existem poucos estudos a respeito
da dieta desta espécie, sendo reportada maior frequência de pequenos roedores
cricetídeos, seguido de outros pequenos mamíferos e aves (GIARETTA, 2002; WANG,
2002). Essa espécie é ativa predominantemente no período noturno, com registros
crepusculares e diurnos (EMMONS e FEER, 1997).
Leopardus pardalis
A jaguatirica é a maior espécie do gênero. Sua massa corporal varia entre 8 e 16
kg (OLIVEIRA e CASSARO, 2005). Sua pelagem possui tonalidades variadas de amarelo-

ocre, cinza-avermelhado e acinzentado, coberto por manchas enegrecidas que formam
rosetas e bandas (CHEIDA et al., 2006). Sua distribuição abrange desde o sul do Texas
nos Estados Unidos até o norte da Argentina (SUNQUIST e SUNQUIST, 2002). No Brasil
distribui-se amplamente, com exceção do Rio Grande do Sul (CHEIDA et al., 2006). Essa
espécie é predominantemente de hábito noturno (DI BITETTI, PAVIOLO e DE ANGELO,
2006), com área utilizada variando entre 3 e 50 km2 (KONECNY, 1989; DI BITETTI,
PAVIOLO e DE ANGELO, 2006). A dieta da jaguatirica baseia-se em pequenos
mamíferos, primatas, artiodáctilos, roedores e xenartros (SUNQUIST e SUNQUIST,
2002; MIRANDA et al., 2005; MORENO, KAYS e SAMÚDIA, 2006; BIANCHI e MENDES,
2007; ABREU et al. 2008), além de ser comum o registro de aves e répteis.
Leopardus wiedii
O gato-maracajá apresenta peso corporal de aproximadamente 3,3 kg. Sua
coloração pode variar do amarelo-acinzentado ao castanho, e possuem manchas com
padrão variado (OLIVEIRA e CASSARO, 2005). Além do território brasileiro, também
ocorre do México até o norte do Uruguai e Argentina (EMMONS e FEER, 1997).
Geralmente é registrado em locais relativamente preservados (FONSECA et al., 1996;
MARGARIDO e BRAGA, 2004). Possui hábito predominantemente noturno e é
extremamente adaptado à vida arbórea e é ágil no solo, sendo considerado um animal
escansorial (KONECNY, 1989; FONSECA et al., 1996). Como reflexo desses hábitos,
alimenta-se especialmente de aves e pequenos roedores arborícolas, além de outros
vertebrados terrestres (KONECNY, 1989; OLIVEIRA e CASSARO, 2005).
Cerdocyon thous
O cachorro-do-mato é um canídeo de médio porte, pesando de 5 a 7 kg
(TROVATI et al., 2001). Este geralmente possui pelagem que vai do cinza ao castanho,
sendo a região dorsal, as patas e a cauda preta (EMMONS e FEER, 1997; NOWAK,
1999). Possui ampla distribuição geográfica, sendo encontrado desde o Uruguai e
norte da Argentina até as terras baixas da Bolívia e Venezuela, ocorrendo também na
Colômbia, Guianas, Suriname e Brasil, no qual a distribuição se dá por todo território,
com exceção das planícies da Bacia Amazônica (BERTA, 1982; JUAREZ e MARINHO-

FILHO, 2002; WILSON e REEDER, 2005; CHEIDA et al., 2006). A espécie possui alto grau
de plasticidade, podendo ser encontrado em ambientes antropizados (BERTA, 1982;
JUAREZ e MARINHO-FILHO, 2002).
Este canídeo é uma espécie onívora, generalista e oportunista, cuja dieta varia
sazonalmente, podendo também se diferenciar dentro da sua área de distribuição.
Geralmente a dieta é composta por frutos, pequenos vertebrados terrestres,
invertebrados, aves, moluscos e peixes (GATTI et al., 2006; CHEIDA et al., 2006).
Chrysocyon brachyurus
O lobo-guará possui pernas longas e finas, pelos longos vermelho alaranjado e
orelha grandes (RODDEN et al., 2004). No Brasil é encontrado a partir da foz do Rio
Paranaíba até o Rio Grande do Sul (RODDEN et al., 2004). É um canídeo onívoro,
consumindo pequenos mamíferos, aves, répteis, artrópodes e vários itens vegetais
(BUENO e MOTTA-JUNIOR, 2004; CIOCHETI, 2007).
II. OBJETIVO
• Descrever a dieta de felinos na área enfocada pelo Pagamento por Serviços
Ambientais – Corredor das Onças

III. METODOLOGIA
A área de estudo compreende onze fragmentos de Mata Atlântica Semidecidual
inseridos na Região Metropolitana de Campinas (Tabela 1 e Figura 1). Porém, somente
em 8 (oito) foram registrados amostras de bolos fecais de pequenos felinos. São eles:
Pirapitingui, Mata da Meia Lua, Bom Retiro, Jaguari, International Paper, Tubarão, ARIE
Matão e ARIE Mata de Santa Genebra.
Tabela 1: Descrição dos fragmentos em estudo na Região Metropolitana de Campinas.
Fragmento Município Área (ha)
Pirapitingui (F1) Cosmópolis 44,90
Mata da Meia Lua (F2) Paulínia 204,65
Bom Retiro (F3) Artur Nogueira -
Jaguari (F4) Paulínia 45,56
International Paper (F5) Artur Nogueira 73,07
Tubarão (F6) Holambra 64,68
ARIE Matão (F7) Cosmópolis 164,25
ARIE Mata Santa Genebra (F8) Campinas 234,13
Mata Santa Genebrinha Campinas 40,65
São Bento - 59,50
Mata Quilombo - 34,41
Para a localização das fezes foram realizados transectos no entorno dos
fragmentos e observados possíveis vestígios de felinos e/ou fezes. As amostras
coletadas foram colocadas em sacos zip, sendo anotadas as respectivas coordenadas
geográficas, o nome do fragmento, a data de coleta e quem coletou. Os pontos de
coleta serão utilizados para mapeamento, a fim de desenvolver modelos de possíveis
corredores ecológicos.
Durante a coleta, as fezes foram associadas a um provável carnívoro por meio de
local de deposição, pegadas, odor, formato, tamanho e diâmetro (medido em campo
com um paquímetro de precisão de 0,01mm).

Figura 1: Mapa da Região Metropolitana de Campinas, com destaque para os fragmentos analisados.
Fonte: Google Earth.
Logo após a coleta as amostras foram colocadas em estufas em torno de 50 °C
para conservação. Antes das triagens foram medidos novamente os diâmetros das
amostras não degradadas, e dispostas durante mais duas horas na estufa, sendo
posteriormente pesadas em balança semianalítica.
Após a pesagem e medições, as amostras foram fragmentadas e deixadas de
molho em água e detergente por um dia para facilitar a lavagem e consequentemente
a triagem. Realizou a lavagem em água corrente e com o auxílio de uma peneira de
malha 1x1 mm.
O material triado foi seco em estufa em torno de 50 °C e acondicionado em sacos
plásticos zip para posterior identificação dos itens alimentares e predadores.
Coletou-se pelos de espécimes (Puma concolor, Panthera onca, Chrysocyon
brachyurus, Cerdocyon thous, Puma yagouaroundi, Leopardus pardalis, Leopardus
wiedii e Leopardus tigrinus) dos zoológicos de Americana - SP e Ilha Solteira – SP para
comparar a impressão cuticular destes com os pelos encontrados nas amostras fecais.
Algumas presas serão identificadas com a ajuda de especialistas considerando as
características de dentes para os pequenos mamíferos, escamas para répteis e

também serão identificados os ectoparasitas (carrapatos e pulgas) encontrados nas
amostras.
Para a identificação dos autores das amostras, preparou lâminas de impressão
cuticular adaptando a técnica descrita por Quadros (2002). Foram encapadas lâminas
com durex grosso de forma a não permanecerem bolhas de ar entre a lâmina e o
durex. Posteriormente, foram separados pelos-guarda do possível carnívoro durante a
triagem, que foram limpos com álcool a 70%. As lâminas foram então parcialmente
cobertas com uma camada homogênea de esmalte incolor da marca Colorama e após
um período de 15 a 20 minutos colocou os pelos sobre o esmalte e as lâminas
encapadas foram então depositadas acima da lâmina com esmalte, simulando um
“sanduiche” de lamina com esmalte, pelos e lâmina encapada com durex. Este
conjunto de lâminas foi fixo envolvendo fita crepe. O conjunto foi pressionado com
uma prensa de exsicata e deixado em descanso por um dia.
As lâminas foram separadas e os pelos retirados suavemente com a ponta dos
dedos. O contramolde marcado foi estudado em microscópio com aumento de 150
vezes.
Após as triagens, pesou em uma balança semianalítica todos os itens alimentares
encontrados.
Para caracterizar a dietas foram utilizadas três abordagens: (i) porcentagem de
fezes com o determinado item alimentar. É representada pela porcentagem de
amostras (fezes) com determinado item alimentar no total de amostras; (ii) avaliação
da biomassa. O cálculo da biomassa será realizado como o produto da massa corpórea
estimada de determinado táxon com o número mínimo de espécimes consumidos
deste táxon. Todos os indivíduos consumidos serão considerados com massa corpórea
em estágio adulto; (iii) avaliação do peso de cada item alimentar nas amostras
identificadas.

IV. RESULTADOS E DISCUSSÃO
Foram coletadas 496 amostras de fezes, sendo 485 encontradas nos oito
fragmentos analisados e 11 amostras em outros fragmentos. A quantidade de
amostras coletadas em cada fragmento, em ordem crescente, é: Mata Santa Genebra
(28), Jaguari (30), Tubarão (40), International Paper (43), Pirapitingui (49), Bom Retiro
(62), ARIE Matão (82) e Mata da Meia Lua (151 amostras) (Figura 2). Nos fragmentos
Pirapitingui, Jaguari e ARIE Matão coletaram-se mais amostras no mês de setembro; já
nos fragmentos da Mata da Meia Lua e Mata Santa Genebra coletou mais amostras em
fevereiro e os fragmentos Bom Retiro, International Paper e Tubarão foram em
dezembro (Figura 3). Pelos dados da Figura 4 percebemos que a quantidade de
amostras coletadas não está proporcionalmente relacionada com a quantidade de dias
de coleta.
Figura 2: Quantidade total de amostras coletadas em cada fragmento da área enfocada pelo PSA – Corredor das Onças.
0
20
40
60
80
100
120
140
160
Quantidade de amostras coletadas em cada fragmento
Amostras
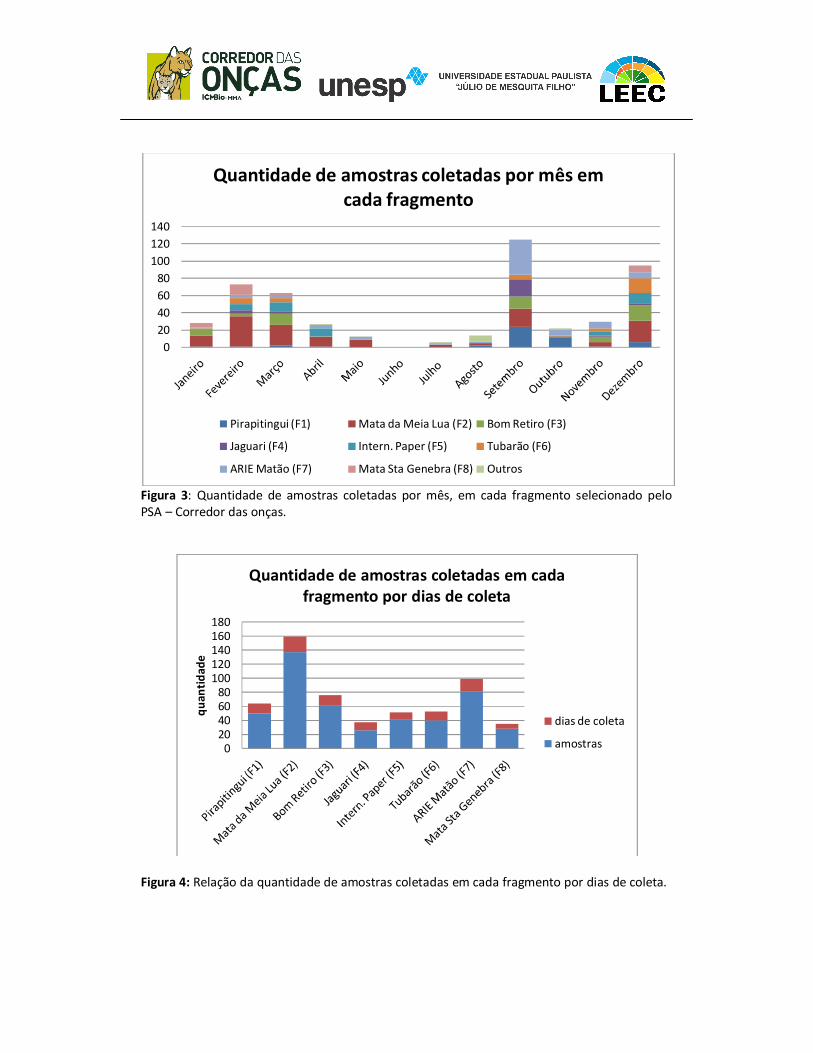
Figura 3: Quantidade de amostras coletadas por mês, em cada fragmento selecionado pelo PSA – Corredor das onças.
Figura 4: Relação da quantidade de amostras coletadas em cada fragmento por dias de coleta.
0
20
40
60
80
100
120
140
Quantidade de amostras coletadas por mês em cada fragmento
Pirapitingui (F1) Mata da Meia Lua (F2) Bom Retiro (F3)
Jaguari (F4) Intern. Paper (F5) Tubarão (F6)
ARIE Matão (F7) Mata Sta Genebra (F8) Outros
020406080
100120140160180
qu
anti
dad
e
Quantidade de amostras coletadas em cada fragmento por dias de coleta
dias de coleta
amostras

Foram triados 251 bolos fecais, sendo 115 identificadas (24 amostras de
Leopardus pardalis; 28 amostras de L. wiedii; 8 amostras de L. tigrinus ; 25 amostras de
Puma yagouarundi; 5 amostras de P. concolor; 17 amostras de Cerdocyon thous e 8
amostras de Chrysocyon bracyurus).
Nas triagens foram identificados os seguintes itens alimentares: pelos, penas e
cartuchos de penas, escamas, molares, incisivos, mandíbulas, garras/unhas, falange,
grama, gravetos, insetos, ossos, sementes e cascas, indeterminados e outros. A
categoria considerada como “Outros” são materiais não frequentes em todas as
amostras como: rochas, folhas, bico, pequenos fragmentos, plásticos, linha, dentre
outros. Os plásticos foram encontrados em duas amostras, uma coletada no
fragmento Pirapitingui e outra no fragmento Tubarão, já a linha foi encontrada em
amostras coletadas no fragmento Pirapitingui (Figura 5).
Figura 5: Alguns materiais encontrados nas amostras fecais. A linha (foto à esquerda) e o plástico (a direita) foram encontradas em amostras coletadas no fragmento Pirapitingui.
Nenhum dos itens alimentares foi registrado em todos os bolos fecais, sendo
que os pelos e os ossos foram encontrados num maior número de amostras (ambos
com 96% de ocorrência). A Figura 6 apresenta os dados referentes a ocorrência de
cada item alimentar, em porcentagem, em comparação ao número total de fezes, e da
figura 7 ao 13, os itens específicos de cada espécie, de acordo com a identificação das
lâminas de pelo.
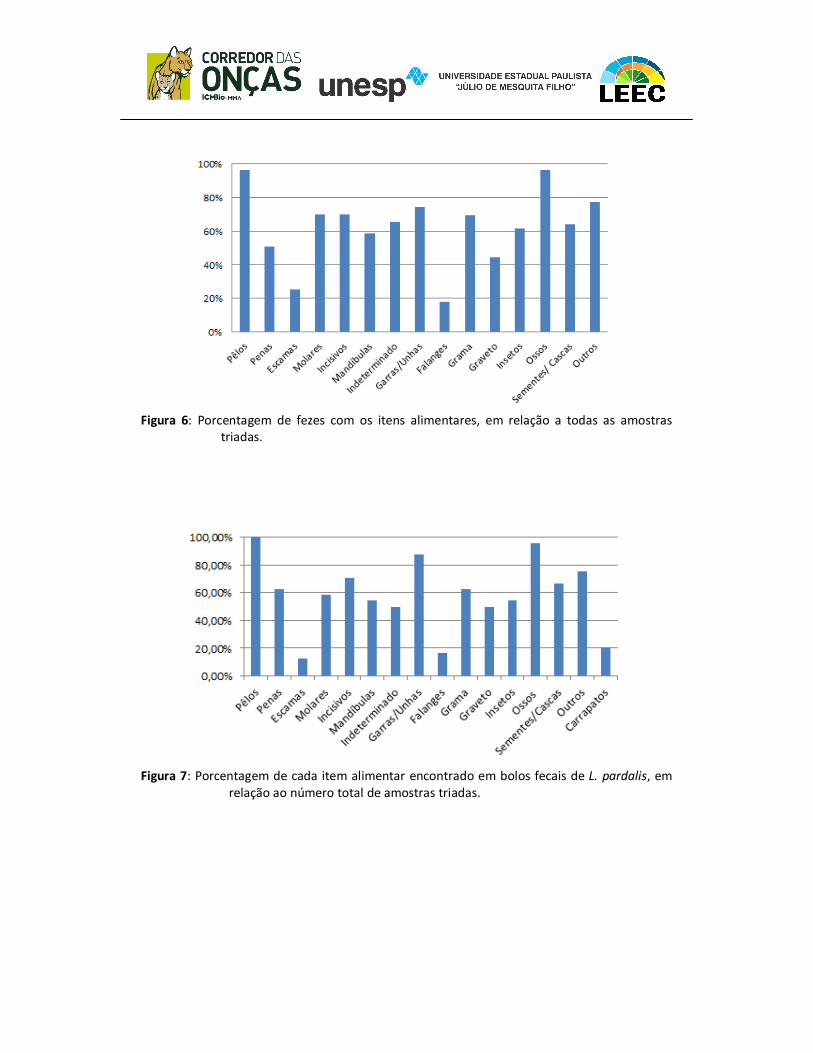
Figura 6: Porcentagem de fezes com os itens alimentares, em relação a todas as amostras
triadas.
Figura 7: Porcentagem de cada item alimentar encontrado em bolos fecais de L. pardalis, em
relação ao número total de amostras triadas.

Figura 8: Porcentagem de cada item alimentar encontrado em bolos fecais de L. wiedii, em relação ao número total de amostras triadas.
Figura 9: Porcentagem de cada item alimentar encontrado em bolos fecais de L. tigrinus, em relação ao número total de amostras triadas.

Figura 10: Porcentagem de cada item alimentar encontrado em bolos fecais de P.
yagouaroundi, em relação ao número total de amostras triadas.
Figura 11: Porcentagem de cada item alimentar encontrado em bolos fecais de P. concolor, em relação ao número total de amostras triadas.
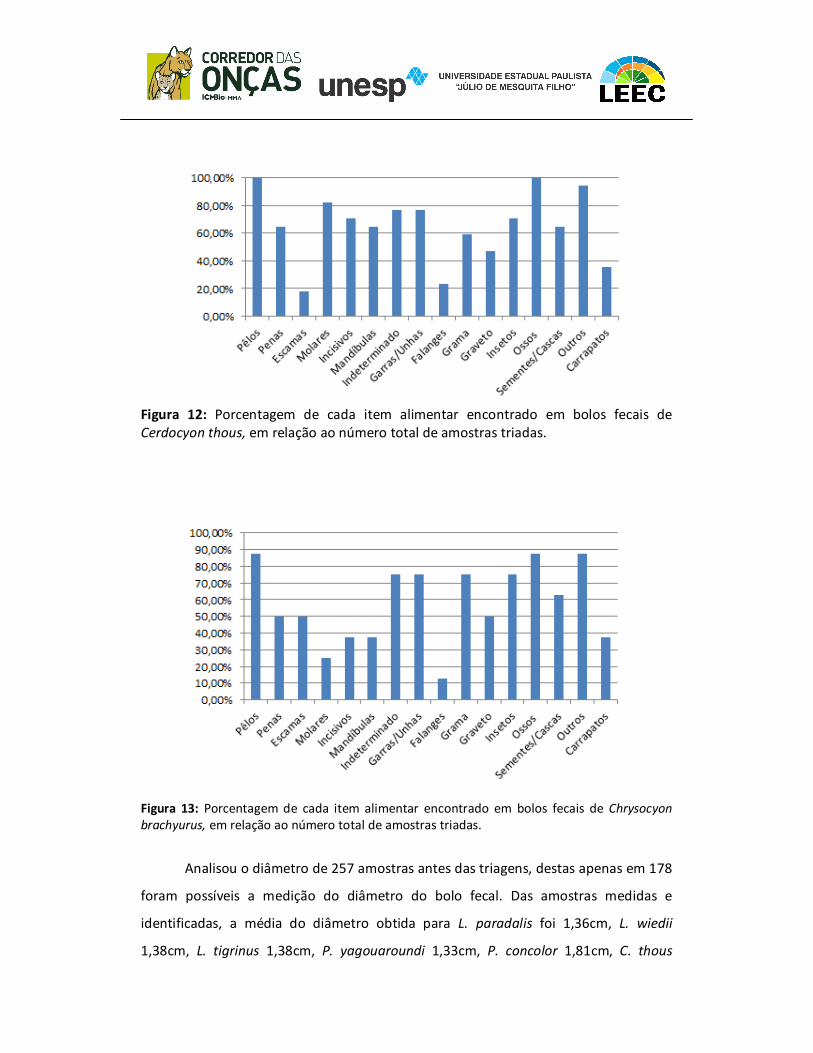
Figura 12: Porcentagem de cada item alimentar encontrado em bolos fecais de Cerdocyon thous, em relação ao número total de amostras triadas.
Figura 13: Porcentagem de cada item alimentar encontrado em bolos fecais de Chrysocyon
brachyurus, em relação ao número total de amostras triadas.
Analisou o diâmetro de 257 amostras antes das triagens, destas apenas em 178
foram possíveis a medição do diâmetro do bolo fecal. Das amostras medidas e
identificadas, a média do diâmetro obtida para L. paradalis foi 1,36cm, L. wiedii
1,38cm, L. tigrinus 1,38cm, P. yagouaroundi 1,33cm, P. concolor 1,81cm, C. thous

1,22cm e C. brachyurus 1,91cm. De acordo com CHAME (2003), no nordeste brasileiro,
um diâmetro menor do que 2,1cm é considerado como pertencente ao bolo fecal de
um pequeno felino e, maior do que este valor é considerado como de grandes felinos.
A média dos diâmetros encontrados em nosso estudo para os pequenos felinos estão
dentro do valor proposto por Chame (2003). Porém, para os grandes felinos, este valor
não coincidiu.
As amostras que continham presença de areia provinda da contaminação na
hora da coleta não foram consideradas para comparar a média de pesos antes e após
as triagens. Desta forma, consideramos apenas 3 amostras para o gato do mato, 10
para o gato mourisco, 9 para o gato maracajá, 12 para a jaguatirica, 2 para onça parda,
9 para o cachorro do mato e 5 para o lobo-guará. Percebemos uma diferença em torno
de 3g para todas as espécies, exceto para a onça parda e para o lobo-guará.
Figura 14: Média da diferença de peso das amostras antes e após serem triadas.
Analisando o peso de cada item alimentar em relação ao peso total dos itens de
todas as amostras identificadas, percebe-se que em todas as espécies que os pelos são
responsáveis pela maior parte do peso dos itens, seguido dos ossos (Figura 15).
02468
1012141618
Pe
so (
g)
Média da diferença de peso antes e após as triagens

Figura 15: Média de pesos de cada item alimentar encontrado em todas as amostras identificadas.
Em geral, a dieta de jaguatirica é composta por pequenos mamíferos, com peso
inferior a 1 kg (BISBAL, 1986; LUDLOW e SUNQUIST, 1987, KONECNY, 1989; SUNQUIST,
SUNQUIST e DANEKE, 1989; CHINCHILLA, 1997; OLIVEIRA et al., 2010), sendo os
pequenos roedores a principal presa em todos os estudos, com exceção de KONECNY
(1989). Rocha-Mendes et al. (2010) registraram o consumo de animais de maior porte
como Quati Nasua nasua e Tapiti Sylvilagus brasiliensis. Em nossos dados apenas
encontramos o consumo de pequenos mamíferos em quase todas as amostras
(95,83%). Assim como Bisbal (1986), Emmons (1987), Konecny (1989) e Sunquist,
Sunquist e Daneke (1989), também encontramos o consumo de aves e folhas, além de
répteis, grama, insetos e sementes.
As principais presas de Leopardus tigrinus são pequenos mamíferos, aves e
repteis (WANG, 2002; OLMOS, 1993, ROCHA-MENDES et al., 2010). Porém, Rocha-
Mendes et al. (2010) também registraram presas maiores, como Nasua nasua,
Sylvilagus brasiliensis e Cuniculus paca. Olmos (1993) também reportou a ingestão de
grama e de invertebrados.
0
0.5
1
1.5
2
2.5
pêl
os
pe
nas
esc
amas
den
tes
ind
eter
min
ado
garr
as/u
nh
asfa
lan
ges
gram
agr
ave
to
inse
tos
oss
inh
os
sem
en
tes
casc
as
roch
asfo
lhas
pel
e/ca
rtila
gem
bic
o
veg.
Div
erso
sfr
agm
en
tos
Pe
so (
g)
Média de pesos dos itens alimentares
L.pardalis
L. tigrunus
L. wiedii
P. yagouaroundi

A dieta do gato-maracajá é composta especialmente por mamíferos arborícolas
(KONECNY, 1989) e aves (LEYHAUSEN 1990), o que pode ser um reflexo de seus
hábitos.
De acordo com a literatura existente, a dieta do jaguarundi é composta
principalmente por pequenos roedores, sendo também citada a presença de
invertebrados (KONECNY 1989; OLMOS 1993). A ocorrência de folhas também foi
citada (BISBAL, 1986).
Ciocheti (2007) encontrou uma dieta estritamente carnívora para onça-parda e
ao contrário do nosso trabalho, encontramos itens vegetais. Martins et al (2008)
obteve na dieta de onça-pintada 80% de ocorrência de mamíferos 12% de répteis e
apenas 8% de aves. Oliveira (2002) encontrou na análise de 23 estudos que as onças
pardas consumiram mais mamíferos de médio porte (1 a 15kg).
Não foram obtidos dados finais a respeito da dieta de cada espécie, pois ainda
não realizou a identificação dos itens pelos especialistas. Portanto, em um momento
posterior, espera-se poder comparar com mais precisão os dados obtidos no trabalho
com as referências existentes.
A biomassa ingerida também será calculada após as identificações das presas.
Notou-se durante as triagens que algumas sementes pequenas e folíolos eram
comuns em quase todas as amostras. Provavelmente estes itens podem ter sido
incorporados nas fezes após a defecação pelos animais já que se encontravam
intactos.

V. CONCLUSÃO
Com o presente trabalho pretende-se diferenciar as dietas entre as espécies de
felinos que se encontram na área de estudo, com o intuito de subsidiar o
desenvolvimento de modelos de possíveis corredores ecológicos que está sendo
realizado por outros pesquisadores, no entanto, para poder concluir a descrição da
dieta dos felinos na área enfocada pelo PSA – Corredor das Onças, necessitamos das
identificações das presas enviadas para analise aos especialistas.
O objetivo final do projeto completo é auxiliar na elaboração de políticas
públicas e buscar a conservação dessas espécies em um ambiente tão perturbado que
é a Região Metropolita de Campinas.
O encontro de materiais de origem antrópica, como plásticos e linha, ingeridos
pelas espécies silvestres ressalta a importância de viabilizar medidas de conservação
nestes locais.

VI. REFERÊNCIAS BIBLIOGRÁFICAS
ABREU, K.C., et al. Feeding habits of ocelot (Leopardus pardalis) in Southern Brazil. Mammalian Biology, 73, 407-411, 2008.
ARJO, W. M.; PLETSCHER, D. H.; REAM, R. R. Dietary overlap between wolves and coyotes in Northwestern Montana. Journal of Mammalogy, 83 (3), 754-766, 2002. BARYCKA, E. Evolution and systematics of feliform Carnivora. Mammalian Biology, 72
(5), 257-282, 2007. BERTA A. Cerdocyon thous. Mammalian Species 186:1-4, 1982. BIANCHI, R.C.; MENDES, S.L. Ocelot (Leopardus pardalis) predation on primates in
caratinga biological station, Southeast Brazil. American Journal of Primatology, 69, 1–6, 2007.
BISBAL, F.J. Food habits of some neotropical carnivores in Venezuela (Mammalia:
Carnivora). Mammalia, 50 (3), 330-339, 1986. BODMER, R. E., EISENBERG, J. F., REDFORD, K. H. Hunting and the likelihood of
extinction of amazonian mammals. Conserv. Biol., 11, 460-466, 1997. BUENO, A. de A.; MOTTA-JUNIOR, J. C. Food habitats of two syntopic canids, the
maned Wolf (Chrysocyon brachyurus) and the crab-eating fox (Cerdocyon thous), in southeastern Brazil. Rev. Chil. Hist. Nat., 77:5-14, 2004.
BRASIL. Instrução Normativa Nº 3, de 26 de maio de 2003. Reconhece como espécies
da fauna brasileira ameaçadas de extinção aquelas constantes da lista anexa à presente Instrução Normativa, considerando apenas anfíbios, aves, invertebrados terrestres, mamíferos e répteis. Diário Oficial da União, 26 mai. 2003.
CHAME. M. Terrestrial mammal feces: amorphometric summary and description.
Memórias do Instituto Oswaldo Cruz, Rio de Janeiro, 98 (suppl. I): 71-94. 2003. CHEIDA, C. C., et al. Ordem Carnivora. In Mamíferos do Brasil, 231-275. Reis, N.R.;
Peracchi, A.L.; Pedro, W.A. & Lima. I.P. (Eds). Nélio R. dos Reis, Londrina, 2006. CHIARELLO, A. G. Effects of fragmentation of the Atlantic Forest on mammal
communities in south-eastern Brazil. Biol. Conserv., 89, 71-82, 1999. CHINCHILLA, F.A. La dieta del jaguar (Panthera onca); el puma (Felis puma) y el
manigordo (Felis pardalis) (Carnivora: Felidae) en al Parque Nacional Corcovado, Costa Rica. Rev. Biol. Trop., 45 (3), 1223-1229, 1997.

CIOCHETI, G., 2007. Uso de habitat e padrão de atividade de médios e grandes mamíferos e nicho trófico de Lobo-Guará (Chrysocyon brachyurus), Onça-Parda (Puma concolor) e Jaguatirica (Leopardus pardalis) numa paisagem agroflorestal, no estado de São Paulo. Dissertação de Mestrado em Ecologia de Sistemas Aquáticos e Terrestres, UFSCAR, 86 p.
CROOKS, K. R., SOULÉ, M. E. Mesopredator release and avifaunal extinctions in a
fragmented system. Nature, 400, 563-566, 1999. CULLEN JR., L., BODMER, E. R., VALLADARES-PÁDUA, C. Ecological consequences of
hunting in Atlantic forest patches. Oryx, 35, 137-144, 2001. DAVIES, T.J. et al. Species coexistence and character divergence across carnivores.
Ecology Letters, 10, 146-152, 2007. DI BITETTI, M.S.; PAVIOLO, A.; DE ANGELO, C. Density, habitat use and activity patterns
of oceltos (Leopardus paralis) in Atlantic Forest of Misiones, Argentina. Journal of Zoology, 270, 153-163, 2006.
DONATTI, C. I., GUIMARÃES JR., P. R., GALETTI, M. Seed dispersal and predation in the
endemic Atlantic Rainforest palm Astrocaryum aculeatissimum across a gradient of seed disperser abundance. Ecol. Res., 2009.
EMMONS, L. H.; FEER, F. Neotropical Rainforest Mammals: A field Guide. 2º ed.
Chicago: University of Chicago Press. 1997. EMMONS, L. H. Comparative feeding ecology of felids in a neotropical rainforest.
Behav Ecol Sociobiol, 20: 271–283, 1987. FONSECA, G. A. B. da, et al. Lista anotada dos mamíferos do Brasil. n. 4. Belo
Horizonte: Conservation International & Fundação Biodiversitas, 1996, 38 p. FRISCIA, A.R.; VAN VALKENBURGH, B.; BIKNEVICIUS, A.R. An ecomorphological analysis
of extant small carnivorans. Journal of Zoology¸ 272, 82-100, 2007. GATTI, A.; BIANCHI, R.; ROSA, C. R. X.; MENDES, S. L. Diet of the crab-eating fox,
Cerdocyon thous (Carnivora, Canidae) in Paulo Cesar Vinha State Park, Espírito Santo State, Brazil. Mammalia. 153-155, 2006.
GEHRING, T. M., SWIHART, R. K. Body size, niche breadth, and ecologically scaled
responses to habitat fragmentation: mammalian predators in an agricultural
landscape. Biol. Conserv., 109, 283-295, 2003.

GIARETTA, K. G. F., 2002. Ecologia alimentar de duas espécies de felino do gênero Leopardus em uma floresta secundária no Sudeste do Brasil. Dissertação de Mestrado em Ecologia, UNICAMP, 86 p.
GORDON, C.E. The coexixtence of species. Rev. Chil. Hist. Nat., 73, 175-198, 2000. JUAREZ K.M. e J MARINHO-FILHO. Diet, habitat use, and home ranges of sympatric
canids in Central Brazil. Jour. of Mamm. 83:925-933, 2002. KONECNY, M. J. Movement Patterns and food habitats of four sympatric carnivore
species in Belize, Central America. Advance in Neotropical Mammalogy, 243-264, 1989.
LAURANCE, W. F. Rainforest fragmentation and the structure of small mammal
communities in Tropical Queensland. Biol. Conserv., 69, 23-32, 1993. LEYHAUSEN, P. Cats. In Grzimek’s encyclopedia mammals. Vol. 3. Van Nostrand
Reinhold, New York, NY.576-632. 1990. LUDLOW, M.E.; SUNQUIST, M.E. Ecology and behavior of Ocelots in Venezuela. Natl.
Geogr. Res., 3 (4), 447-461, 1987. MACARTHUR, R.H.; LEVINS, R. The limiting similarity, convergence and divergence of
coexisting species. Am. Natur., 101, 377- 385, 1967. MARGARIDO, T. C. C.; BRAGA, F. G. Mamíferos. In: MIKICH, S. B.; BÉRNILS, R. S. (Eds.).
Livro Vermelho da Fauna Ameaçada no Estado do Paraná. Curitiba: Instituto Ambiental do Paraná e Mater Natura, 25-142. 2004.
MIRANDA, J. S. Subsídios para Elaboração e Implantação da Agenda 21 do Município
de Campinas – SP. Diagnóstico Ambiental de Campinas – Fauna. Abr/2003. Disponível em <http://www.agenda21cps.cnpm.embrapa.br/>. Acesso em: 01 ago 2012.
MIRANDA, J.M.D., et al. Predation of Alouatta guariba clamitans Cabrera (Primates,
Atelidae) by Leopardus pardalis (Linnaeus) (Carnivora, Felidae). Revista Brasileira de Zoologia, 22, 793-795, 2005.
MORENO, R. S.; KAYS, R. W.; SAMÚDIA JR., R. Competitive release in diets of ocelot
(Leopardus pardalis) and puma (Puma concolor) after jaguar (Panthera onca) decline. Journal of Mammalogy, 87 (4), 808-816, 2006.
MYERS, N., et al. Biodiversity hotspots for conservation priorities. Nature, 403, 853-
858, 2000.

NOWAK, R.M. Walker’s mammals of the world. Baltimore, The John Hopkins University Press, 1921p, 1999.
NOWAK, R.M. Walker’s Carnivores of the World. The Johns Hopkins University Press,
Maryland. 2005. OLIVEIRA, T.G.; CASSARO, K. Guia de campo dos felinos do Brasil. Instituto Pró-
Carnívoros, Fundação Parque Zoológico, Sociedade de Zoológicos do Brasil, Pró-Vida Brasil. 2005.
OLIVEIRA, T.G. et al. Ocelot ecology and its effect on the smallfelid guild in the lowland
neotropics. In: MACDONALD, D. W.; LOVERIDGE, A. (Org.). Biology and Conservation of Wild Felids. Oxford: Oxford University Press, 563-584, 2010.
OLMOS, F. Notes on the food habits of brazilian “caatinga” carnivores. Mammalia, v.
57 (1), 126-130, 1993. PIERCE, B.M.; BLEICH, V.C. & BOWYER, T. Selection of mule deer by mountain lions and
coyotes: effects of hunting style, body size, and reproductive status. Journal of Mammalogy, 81 (2), 462-472, 2000.
QUADROS, J. Identificação microscópica de pêlos de mamíferos e suas aplicações no estudo da dieta de carnívoros. Tese de Doutorado em Zoologia, Universidade
Federal do Paraná. 2002. RIBEIRO, M. C., et al. The brazilian Atlantic Forest: how much is left, and how is the
remaining Forest distributed? Implications for conservation. Biol. Conserv., 142, 1141-1153, 2009.
ROCHA-MENDES, F., et al. Feeding ecology of carnivores (Mammalia, Carnivora)in
Atlantic Forest remnants, Southern Brazil. Biota Neotropical, 10 (4). 2010. RODDEN, M.; RODRIGUES, F. H. G.; BESTELMEYER, S. 2004. Maned wolf Chrysocyon
brachyurus (Illiger, 1815). p. 38-43. In: SILLEROZUBIRI, C.; HOFFMANN, M.; MACDONALD, D. W. (Eds.). Canids: Foxes, Wolves, Jackals and Dogs. Status Surveyand Conservation Action Plan. Gland, Switzerland e Cambridg: Canid Specialist Group, 430 p. 2004.
SCHOENER, T.W. Resource portioning in ecological communities. Science, 185, 27-39,
1974. SUNQUIST, M.E., SUNQUIST, F.; DANEKE, F.H. Ecological separation in a venezuelan
Llanos carnivore community. In Advances in neotropical mammalogy (K.H. Redford & J.F. Eisenberg, ed.). The Sanshill Crane Press, Inc., Gainesville, 197-232, 1989.

SUNQUIST, M.; SUNQUIST F. Wild Cats of the World. University of Chicago Press, Ltd., London. 2002.
TROVATI, R.G.; BRITO, B.; PRADA, M. Captura anestesia e biometria de Cerdocyon
thous (Raposa-cinzenta) na região central do Tocantins. Humanitas, 3:85-91, 2001.
WANG, E. Diets of Ocelots (Leopardus pardalis), Margays (L. wiedii), and Oncillas (L.
tigrinus) in the Atlantic Rainforest in Southeast Brazil. Studies on Neotropical Fauna and Environment, 37 (3), 207-212, 2002.
WILSON, D.E.; REEDER, D.M. Mammal species of the world: a taxonomic and
geographic reference. Baltimore, Johns Hopkins University Press, 2142 p. 2005.


















