New Estudo das alterações morfo-funcionais de espermatozóides … · 2010. 12. 14. · FOLHA DE...
101
UNIVERSIDADE DE SÃO PAULO FACULDADE DE MEDICINA VETERINÁRIA E ZOOTECNIA DEPARTAMENTO DE REPRODUÇÃO ANIMAL Estudo das alterações morfo-funcionais de espermatozóides bovinos submetidos à sexagem por meio da técnica de citometria de fluxo Priscilla Harumi Tanno São Paulo 2009
Transcript of New Estudo das alterações morfo-funcionais de espermatozóides … · 2010. 12. 14. · FOLHA DE...

UNIVERSIDADE DE SÃO PAULO
FACULDADE DE MEDICINA VETERINÁRIA E ZOOTECNIA
DEPARTAMENTO DE REPRODUÇÃO ANIMAL
Estudo das alterações morfo-funcionais de
espermatozóides bovinos submetidos à sexagem por
meio da técnica de citometria de fluxo
Priscilla Harumi Tanno São Paulo 2009

Priscilla Harumi Tanno
Estudo das alterações morfo-funcionais de
espermatozóides bovinos submetidos à sexagem por
meio da técnica de citometria de fluxo São Paulo 2009
Dissertação apresentada ao Programa de Pós-Graduação em Reprodução Animal da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo para a obtenção do título de Mestre em Medicina Veterinária Departamento: Reprodução Animal Área de Concentração: Reprodução Animal Orientador: Prof. Dr. Rubens Paes de Arruda

Autorizo a reprodução parcial ou total desta obra, para fins acadêmicos, desde que citada a fonte.
DADOS INTERNACIONAIS DE CATALOGAÇÃO-NA-PUBLICAÇÃO
(Biblioteca Virginie Buff D’Ápice da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo)
T.2168 Tanno, Priscilla Harumi FMVZ Estudo das alterações morfo-funcionais de espermatozóides bovinos
submetidos à sexagem por meio da técnica de citometria de fluxo / Priscilla Harumi Tanno. – São Paulo : P. H. Tanno, 2009.
100 f. : il.
Dissertação (mestrado) - Universidade de São Paulo. Faculdade de Medicina Veterinária e Zootecnia. Departamento de Reprodução Animal, 2009.
Programa de Pós-Graduação: Reprodução Animal. Área de concentração: Reprodução Animal.
Orientador: Prof. Dr. Rubens Paes de Arruda.
1. Sêmen sexado. 2. Bovino. 3. Fosforilação da tirosina. 4. Membrana plasmática.
5. Reação acrossômica. 6. Citometria de fluxo. I. Título.


FOLHA DE AVALIAÇÃO
Nome: TANNO, Priscilla Harumi Título: Estudo das alterações morfo-funcionais de espermatozóides bovinos submetidos à sexagem por meio da técnica de citometria de fluxo. Dissertação apresentada ao Programa de Pós-Graduação em Reprodução Animal da Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo para a obtenção do título de Mestre em Medicina Veterinária Data: ____/____/_____
Banca Examinadora
Prof. Dr. __________________________ Instituição: _______________________
Assinatura:__________________________ Julgamento: _______________________
Prof. Dr. __________________________ Instituição: ______________________
Assinatura: __________________________ Julgamento: ______________________
Prof. Dr. __________________________ Instituição: _____________________
Assinatura: _________________________ Julgamento: _____________________

“TUDO O QUE TEMOS DE DECIDIR
É O QUE FAZER COM O TEMPO QUE NOS É DADO.”
J. R. R. TOLKIEN

DEDICO ESTA DISSERTAÇÃO
Aos meus Pais, Roberto Toshio Tanno e Amélia Hideco Tanno e à minha irmã,Thais
Hitomi Tanno, por todo o carinho, incentivo e apoio que sempre me deram sem medir
esforços, o meu mais profundo amor e gratidão
À minha família, em especial à minha tia Edna Tomiko Kato, pelo apoio e incentivo
constantes

Ao meu marido, Pablo Eduardo Martins de Paiva,
Que esteve e continua sempre ao meu lado, me apoiando, amparando e norteando os
meus passos, que compartilhou com amor todos os momentos felizes e me confortou, nos mais
difíceis com toda a sua calma e paciência
Digo que sou uma pessoa de sorte por ter você ao meu lado e por fazer parte da sua
vida!
AMO MUITO TODOS VOCÊS!!!

AO MEU ORIENTADOR, PROF. RUBENS,
Gostaria de expressar a minha gratidão, o meu respeito e minha admiração pelo profissional ético,
justo e competente, como toda pessoa formadora de opinião deve ser.
Agradeço por ser parte integrante do seu grupo de pesquisa e espero que se estenda para o
doutorado.
Saiba que o senhor como orientador e professor foi muito importante para o meu crescimento e
amadurecimento profissional e pessoal e que se tornou um amigo dentro do meu círculo de amizades e espero
que assim continue.

AGRADECIMENTOS À DEUS, por me permitir chegar até aqui e colocar pessoas tão especiais no meu caminho. À Sexing Technologies , por permitir a realização de uma das minhas principais metas e por fazer parte desta equipe de trabalho. Aos meus amigos da Sexing Technologies, Juan Moreno, Mr. Maurice Rosenstein, Evanil Pires de Campos Filho, Osvaldo Teobaldo Filho, Cláudia Fernandes Raphael, Thaís Rose dos Santos Hamilton, Esther Furuichi Maia, Flávio Marcelo Chagas, Adriano Ronconi, Fernando Morales Salles Prestes, Rosiára Rosária Dias Maziero, Sthéfano Gaudenzio Pannazzolo, Bruna Cristina de Souza, Janeth Rosendo dos Santos, Eduardo Ramos Bennedetti, Érika Junqueira da Fonseca, pelo apoio e amizade, pois sem vocês isto não seria possível de se concretizar Aos meus amigos da CRV Lagoa, Dra. Lúcia Helena Rodrigues e Carlos Silva pela grande cooperação e incentivo Aos meus amigos pós-graduandos da USP, principalmente o André Furugen César de Andrade e a Fabiane Gilli Zaffalon que me auxiliaram muito com as análises, estatística, gráfico e ao Andres Mejia Escobar Tiquito que me ajudou com os organogramas... Muito obrigada, pois sem vocês seria impossível cumprir no prazo! À Profa. Eneiva Carla Celeghini pela presteza e eficiência em me socorrer sempre nos momentos mais não digo difíceis, mas incisivos!Muito obrigada! Ao Departamento de Reprodução Animal pela oportunidade de fazer parte desta benemérita instituição
Aos Professores do Departamento de Reprodução Animal com quem pude além de conviver, buscar
conhecimentos que foram e serão importantes em toda a minha vida profissional...Prof. Dr. Pietro Sampaio
Baruselli, Prof. Dr. Marcelo Alcindo de Barros Vaz Guimarães, Prof. Dr. José Antonio Visintin, Profa.
Dra. Valquiria Hyppolito Barnabe, Prof. Dr. Renato Campanarut Barnabe, Profa. Dra. Clair Motos de
Oliveira, Profa. Dra. Camila Infantosi Vannucchi, Profa. Dra. Mayra Elena Ortiz D’Avila Assumpção e
ao Prof. Dr. Claudio Alvarenga de Oliveira....muito obrigada
Ao Professor Dr. Paulo Henrique Mazza Rodrigues, por ter tido muita paciência e compreensão em sua disciplina e, principalmente, por me ensinar os conceitos de estatística, fundamental na pesquisa À Harumi D. Shiraishi, pela dedicação e presteza de sempre.

RESUMO
TANNO, P. H. Estudo das alterações morfo-funcionais de espermatozóides bovinos submetidos à sexagem por meio da técnica de citometria de fluxo. [Study of morpho-functions alterations in bovine spermatozoa submitted on sorting through flow cytometry technology]. 2009. 100 f. Dissertação (Mestrado em Medicina Veterinária) – Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2009. O objetivo deste estudo foi avaliar as alterações morfo-funcionais do sêmen bovino submetido
à sexagem através da técnica de citometria de fluxo e comparar com o sêmen convencional e
a diferença entre as subespécies. O sêmen foi obtido de seis touros, sendo três taurinos
(Holandês preto e branco) e três zebuínos (Gir leiteiro), com quatro ejaculados de cada animal
(n = 24). Cinco tratamentos foram realizados: sêmen convencional com diluidor L (Lagoa);
sêmen convencional com diluidor S (Sexing) e sêmen sexado (nos tempos de zero, três e seis
horas após a ejaculação para avaliar se existem alterações de membranas ao longo do tempo).
Foram avaliados pela citometria de fluxo quanto à integridade de membrana plasmática e
reação acrossomal (PI/FITC-PSA); peroxidação lipídica (C11-BODIPY581/591) e capacitação
espermática através de análises da fosforilação da tirosina e aumento da fluidez e
desorganização da membrana plasmática (Merocianina 540 e Yo-Pro1). Os efeitos de
tratamentos foram avaliados por análises de variância (ANOVA), empregando-se o programa
estatístico Stat-View® (SAS Institute, Inc. 1998, Cary, NC, USA). Foram considerados
significativos os valores com P<0,05 e com tendência a significância (P<0,10). O sêmen
proveniente da subespécie taurina teve um efeito negativo mais significante do que a zebuína
na qualidade do sêmen. O sêmen sexado apresentou maior peroxidação lipídica nos
espermatozóides com membrana íntegra e a presença de proteínas fosforiladas na superfície
da membrana plasmática, fatores que são indicativos de capacitação espermática. Porém, ao
contrário do esperado, este aumento não foi acompanhado pela quantidade de células
classificadas como capacitadas pela análise de associação de sondas Yo-Pro-1 e Merocianina
540, pois o sêmen convencional apresentou maior população espermática com membrana
plasmática íntegra desorganizada (P<0,05). Não houve piora na qualidade do sêmen sexado
devido ao tempo de espera do ejaculado (P<0,05).
Palavras-chave: Sêmen sexado. Bovino. Fosforilação da tirosina. Reação acrossômica. Citometria de fluxo.

ABSTRACT
TANNO, P. H. Study of morpho-functions alterations in bovine spermatozoa submitted on sorting through flow cytometry technology. [Estudo das alterações morfo-funcionais de espermatozóides bovinos submetidos à sexagem por meio da técnica de citometria de fluxo]. 2009. 100 f. Dissertação (Mestrado em Medicina Veterinária) – Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2009.
The objective of this study was to evaluate the morpho-functions alterations in bovine semen
submitted on sorting through flow cytometry technology and compare with conventional
semen and the difference between the subspecies. Semen was obtained from six bulls, that
three were taurine (Holstein Black and White) and three were zebuine (Dairy Gir) with four
ejaculates from each animal (n = 24). Five treatments were performed: conventional semen
with L (Lagoa) media; conventional semen with S (Sexing) media and sexed semen (zero,
three and six hours after ejaculation to evaluate if there were membrane alterations over the
time). The treatments were analyzed with flow cytometry as for plasmatic membrane integrity
and acrosome reaction (PI/FITC-PSA); lipid peroxidation (C11-BODIPY581/591) and sperm
capacitation through protein tyrosine phosphorylation and the increase in plasma membrane
fluidity and disorganization (Merocyanine 540 and Yo-Pro-1). The treatments effects were
evaluated by analysis of variance (ANOVA) (Stat-View®, SAS Institute, Inc. 1998, Cary, NC,
USA). Effects were considered significant if the values were P<0.05 and with significant
tendency (P<0.10). Semen from the taurine subspecie had a more significant negative effect
than zebuine on semen quality. Sexed semen showed more lipid peroxidation in sperm with
membrane integrity and the presence of phosphorylated proteins in plasma membrane surface
that seemed to be sperm capacitation. However, in the contrary of expected, this increase did
not accompany with quantity of cells classified as capacitated by the association probes Yo-
Pro-1 e Merocyanine 540 analyse because the conventional semen showed more sperm
population with plasma membrane integrity disorganized (P<0.05). There was not worsening
in sexed semen quality over the time ejaculate waiting (P<0.05).
Keywords: Sexed semen. Bovine. Tyrosine phosphorylation. Acrosome reaction. Flow
cytometry.

LISTA DE FIGURAS
Figura 1 - Esquema resumido do delineamento experimental .................................................50 Figura 2 - Esquema simplificado para a obtenção da concentração espermática.....................52 Figura 3 - Aparelho de Citometria de Fluxo SX MOFLO® .....................................................56 Figura 4 - Gráfico de pontos gerado pela análise de 10.000 células por citometria de fluxo.
Amostra corada com a associação H33342, PI, FITC-PSA permitindo a classificação dos espermatozóides bovinos em quatro categorias – Pirassununga – 2009..........................................................................................................................59
Figura 5 - A. Gráfico de pontos gerado pela análise por citometria de fluxo em amostra corada
com a associação das sondas fluorescentes C11-BODIPY, H33342 e PI. Podem-se observar duas categorias distintas, sendo que o quadrante MPI contém os espermatozóides bovinos com a membrana plasmática íntegra e selecionados para a avaliação da intensidade da peroxidação das membranas. B. Histograma com a população selecionada MPI que está sendo analisada quanto à distribuição da fluorescência captada no fotomultiplicador com band pass de 530 ±15 nm (células com peroxidação lipídica). O valor obtido é indicado no quadro como a média e a mediana em unidades arbitrárias (ua) – Pirassununga – 2009 .................................60
Figura 6 - Gráfico de pontos gerado pela análise por citometria de fluxo, neste caso as células
foram coradas com a associação do H33342, Yo-Pro-1 e da Merocianina 540 permitindo a classificação dos espermatozóides bovinos em três categorias – Pirassununga – 2009 ................................................................................................62
Figura 7 - A. Gráfico de pontos gerado pela análise por citometria de fluxo em amostra corada
com a associação de H33342, com anticorpo antifosfotirosina conjugado ao FITC e a sonda fluorescente PI. Podem-se observar duas categorias distintas, sendo a MPI os espermatozóides bovinos com a membrana plasmática íntegra e selecionados para a avaliação da detecção dos resíduos de tirosina presentes na superfície da célula. B. histograma apresentando a população selecionada MPI que está sendo analisada quanto à distribuição da fluorescência captada no fotomultiplicador com band pass de 530 ±15 nm (células marcadas com o anticorpo antifosfotirosina). O valor obtido é indicado no quadro como a média e a mediana em unidades arbitrárias (ua) – Pirassununga - 2009 .....................................................................63

LISTA DE GRÁFICOS
Gráfico 1 - Médias ± erros padrão da quantidade de células (%) sem reação acrossomal e membrana plasmática íntegra (SRAMI) no sêmen bovino pós-descongelação submetido ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga - 2009..........................................................................................67
Gráfico 2 - Médias ± erros padrão da quantidade de células (%) sem reação acrossomal e
membrana plasmática íntegra (SRAMI) no sêmen de taurinos e zebuínos pós-descongelação submetidos ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga - 2009...............................................................68
Gráfico 3 - Médias ± erros padrão da quantidade de células (%) com reação acrossomal e
membrana plasmática íntegra (CRAMI) no sêmen bovino pós-descongelação submetido ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga - 2009..........................................................................................69
Gráfico 4 - Médias ± erros padrão da quantidade de células (%) com reação acrossomal e
membrana plasmática íntegra (CRAMI) no sêmen de taurinos e zebuínos pós-descongelação submetidos ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga – 2009 ..............................................................70
Gráfico 5 - Médias ± erros padrão da porcentagem de células com membrana plasmática
íntegra (MI) no sêmen bovino pós-descongelação submetido ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga - 2009..................................................................................................................................71
Gráfico 6 - Médias ± erros padrão da quantidade de células (%) com membrana plasmática
íntegra (MI) no sêmen de taurinos e zebuínos pós-descongelação submetidos ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga – 2009 ................................................................................................72
Gráfico 7 - Médias ± erros padrão da quantidade de células (%) sem reação acrossomal (SRA)
no sêmen bovino pós-descongelação nos cinco tratamentos descritos acima – Pirassununga - 2009.................................................................................................73
Gráfico 8 - Médias ± erros padrão da quantidade de células (%) sem reação acrossomal (SRA)
no sêmen de taurinos e zebuinos pós-descongelação submetidos ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga – 2009..................................................................................................................................74
Gráfico 9 - Médias ± erros padrão dos valores lineares médios da intensidade de fluorescência
da população espermática com membrana plasmática íntegra e corada com a sonda fluorescente C11-BODIPY581/591 (PEIFMPI) no sêmen bovino submetido ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h). Os valores estão representados em unidades arbitrárias (u.a.) – Pirassununga – 2009 .............75

Gráfico 10 - Médias ± erros padrão dos valores lineares médios da intensidade de fluorescência da população espermática com membrana plasmática íntegra e corada com a sonda fluorescente C11-BODIPY581/591 (PEIFMPI) no sêmen de taurinos e zebuínos pós-descongelação submetidos ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h). Os valores estão representados em unidades arbitrárias (u.a.) – Pirassununga – 2009...................................................................76
Gráfico 11 - Médias ± erros padrão dos valores lineares médios da intensidade de
fluorescência emitida pela população espermática com membrana plasmática lesada e coradas com a sonda fluorescente C11-BODIPY581/591 (PEIFMPL) do sêmen bovino submetido ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h). Os valores estão representados em unidades arbitrárias (u.a.) – Pirassununga – 2009 ................................................................................................77
Gráfico 12 - Médias ± erros padrão dos valores lineares médios da intensidade de
fluorescência da população espermática com membrana plasmática lesada e corada com a sonda fluorescente C11-BODIPY581/591 (PEIFMPL) no sêmen de taurinos e zebuinos pós-descongelação submetidos ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h). Os valores estão representados em unidades arbitrárias (u.a.) – Pirassununga – 2009...................................................................78
Gráfico 13 - Médias ± erros padrão dos valores lineares médios da intensidade de
fluorescência emitida pela população espermática com a membrana plasmática íntegra e marcada com o anticorpo antifosfotirosina conjugado a uma fluoresceína (FOIFMI) no sêmen bovino pós-descongelação submetido ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h). Os valores estão representados em unidades arbitrárias (u.a.) – Pirassununga – 2009.......................79

LISTA DE ABREVIATURAS E SIGLAS
µL microlitro µm micrômetro µM micromolar A23187 ionóforo de cálcio AIMI membrana plasmática íntegra e acrossomo não reagido ARMI membrana plasmática lesada e acrossomo reagido C11-BODIPY581/591 4,4-difluoro-5-(4-phenyl-1,3-butadienyl)-4-bora-3ª,4ª-diaza-s-indacene-3-undecanoic acid Ca2+ íon calico cAMP adenosina-monofosfato ciclica Cl- íon cloro CRAMI com reação acrossomal e membrana plasmática íntegra Diluidor L diluidor Lagoa Diluidor S diluidor Sexing DNA ácido desoxirribonucléico EROs espécies reativas de oxigênio ERK regulação de sinal extracelular FITC isotiocianato de fluoresceína FOIFMI intensidade da fluorescência emitida pela população espermática com membrana plasmática íntegra e marcada com o anticorpo antifosfotirosina conjugado a uma fluoresceína g grama g gravidade h hora H2O2 peróxido de hidrogênio H33342 Hoechst 33342 HDL lipoproteína de alta densidade HO2 hidroperoxil HOCL ácido hipocloroso HOCl ácido hipocloroso

Hz Hertz IA inseminação artificial IP iodeto de propídio K+ íon potássio LPC lisofosfatidilcolina MAPK proteínas quinases mitógenas-ativadas MEMID quantidade de células com membrana plasmática íntegra desorganizada mg miligrama Mg2+ íon magnésio MI somatória das células com membrana plasmática íntegra min minuto mL mililitro mM milimolar MP espermatozóide com médio potencial de membrana mitocondrial MP motilidade progressiva MPI células com membrana plasmática íntegra (viáveis) MPL espermatozóide com membrana plasmática lesada MT motilidade total Na+ íon sódio NCAP células viáveis não capacitadas NCMPI espermatozóide não capacitado com membrana plasmática íntegra nm nanômetro O2-• ânion superóxido OH- radicais hidroxil p nível de significância PEIFMPI intensidade da fluorescência da população espermática com membrana plasmática íntegra e coradas com a sonda fluorescente C11-BODIPY581/591 pH concentração de íons de hidrogênio PKA proteína quinase A PSA aglutinina de Pisum sativum RA reação acrossômica

RNS espécie reativa de nitrogênio RO• alcoxil ROO- peroxil s segundos SAS Statistical Analysis System (Sistema de Análise Estatística) SNAREs soluble N-ethylmaleimide-sensitive factor attachment protein
receptor proteins SOD superoxido-dismutase SRA somatória das células com membrana acrossomal sem reação SRAMI sem reação acrossomal e membrana plasmática íntegra TALP meio de Tyrode suplementado com albumina, lactato e piruvato
TALPm meio de Tyrode suplementado com lactato e piruvato e sem adição de
CaCl 2H2O e NaHCO3 ua unidades arbitrárias ZP zona pelúcida
ZP3 proteína recombinante não ativa

LISTA DE SÍMBOLOS
°C graus Celsius % percentagem x vezes 106 milhões ® marca registrada < menor que > maior que ± mais ou menos - menos / negativo + mais/positivo = igual ° grau

SUMÁRIO
1. INTRODUÇÃO ..................................................................................................................22
2 REVISÃO DE LITERATURA...........................................................................................27
2.1 HISTÓRICO DA SEXAGEM........................................................................................27
2.2 DIFERENÇAS NO DNA ESPERMÁTICO ..................................................................28
2.3 O ESPERMATOZÓIDE ................................................................................................31
2.3.1 A membrana plasmática .......................................................................................32
2.3.2 Capacitação espermática ......................................................................................35
2.3.3 Fosforilação de proteínas ......................................................................................37
2.3.4 Reação acrossômica ...............................................................................................39
2.4 ESPÉCIES REATIVAS DE OXIGÊNIO ......................................................................41
3 OBJETIVOS ........................................................................................................................45
4 HIPÓTESES ........................................................................................................................47
5 MATERIAL E MÉTODOS ................................................................................................49
5.1 LOCAL...........................................................................................................................49
5.2 ANIMAIS E MANEJO ..................................................................................................49
5.3 DELINEAMENTO EXPERIMENTAL.........................................................................49
5.4 PROCEDIMENTOS EXPERIMENTAIS......................................................................50
5.4.1 Colheita do sêmen..................................................................................................51
5.4.2 Avaliações do sêmen in natura..............................................................................51
5.4.3 Congelação do sêmen pela técnica convencional ................................................54
5.4.4 Sexagem espermática pela técnica de citometria de fluxo .................................54
5.4.5 Criopreservação do sêmen sexado .......................................................................56
5.4.6 Avaliações do sêmen convencional (dois diluidores) e do sexado (três
diferentes tempos)...........................................................................................................57
5.5 ANÁLISE ESTATÍSTICA.............................................................................................63
6 RESULTADOS ....................................................................................................................66
6.1 INTEGRIDADE DA MEMBRANA PLASMÁTICA E REAÇÃO ACROSSOMAL
EM SÊMEN CONVENCIONAL E SEXADO ....................................................................66
6.2 INTEGRIDADE DA MEMBRANA PLASMÁTICA EM SÊMEN CONVENCIONAL
E SEXADO ..........................................................................................................................70
6.3 REAÇÃO ACROSSÔMICA EM SÊMEN CONVENCIONAL E SEXADO...............72

6.4 PEROXIDAÇÃO DA MEMBRANA PLASMÁTICA EM SÊMEN CONVENCIONAL
E SEXADO ..........................................................................................................................74
6.5 FOSFORILAÇÃO DO AMINOÁCIDO TIROSINA EM ESPERMATOZÓIDES COM
MEMBRANA PLASMÁTICA ÍNTEGRA EM SÊMEN CONVENCIONAL E SEXADO
..............................................................................................................................................78
6.6 ESTABILIDADE (FLUIDEZ) DA MEMBRANA PLASMÁTICA ÍNTEGRA
DESORGANIZADA EM SÊMEN CONVENCIONAL E SEXADO.................................80
7 DISCUSSÃO ........................................................................................................................84
8 CONCLUSÕES....................................................................................................................89
REFERÊNCIAS .....................................................................................................................92

1 Introdução

1 Introdução 22
Priscilla Harumi Tanno
1. INTRODUÇÃO
O desenvolvimento da sexagem de sêmen, separando-se o espermatozóide contendo o
cromossomo X do Y baseado na diferença de DNA empregando-se a citometria de fluxo, foi
largamente aceito como o principal avanço na tecnologia da reprodução (BLONDIN et al.,
2009). A pré-seleção do sexo pode acelerar o progresso genético como também, beneficiar o
manejo e a eficiência produtiva (MAXWELL et al., 2004; PARRILLA et al., 2005). Esta
técnica foi desenvolvida por Larry A. Johnson em 1989, no USDA Beltsville Agricultural
Research Center (SEIDEL JR. et al., 1999; JOHNSON, 2000; GARNER; SEIDEL JR., 2003;
WEIGEL, 2004; GARNER, 2006) e é o único método validado até o momento para a pré-
seleção do sexo antes do nascimento (GRAAF et al., 2009). Todavia, somente nos bovinos, a
tecnologia atingiu um nível de desenvolvimento que permitiu a sua aplicação comercial
(RATH; JOHNSON, 2008).
O emprego do sêmen sexado permite aumentar o impacto na eficiência produtiva de
carne e leite, produzindo uma proporção ideal de machos e fêmeas, adquirindo vantagens de
características limitadas e influenciadas pelo sexo, e ainda, obter manejos práticos e flexíveis
economicamente (RATH; JOHNSON, 2008). Adicionalmente permite selecionar entre seus
rebanhos, as matrizes potenciais, produzindo novilhas de reposição apenas de animais
geneticamente superiores (HOHENBOKEN, 1999; WEIGEL, 2004; MOCÉ et al., 2006). É
esperada a elevação do ganho genético de até 15% comparado ao sêmen convencional, além
da redução do custo de programas de teste de progênie e transferência de embrião e, aumento
do valor de marcadores genéticos (HOHENBOKEN, 1999; JOHNSON, 2000; WEIGEL,
2004; DE VRIES et al., 2008).
A precocidade sexual em gado de corte, no âmbito econômico, mostra grande
importância devido à redução de custos de produção de novilhas de reposição e retorno mais
rápido do capital investido. Na maioria dos rebanhos zebuínos, o número de novilhas que
chegam ao desafio para precocidade sexual não é suficiente para realizar toda a reposição de
fêmeas e intensificar a seleção por precocidade sexual. O uso do sêmen sexado para fêmea
poderia aumentar o número de fêmeas candidatas, acelerando tal processo de seleção.
Já na produção in vitro (PIV) de embriões, sabe-se que a proporção de produção de
embriões machos é maior do que de fêmeas e com o uso do sêmen sexado para fêmea,
inverteria esta proporção, já que as fêmeas têm um valor de mercado maior do que de machos
(WHEELER et al., 2006; RATH; JOHNSON, 2008; RATH et al., 2009). No âmbito da saúde

1 Introdução 23
Priscilla Harumi Tanno
humana, uma das aplicações de destaque é na prevenção de doenças genéticas ligadas ao sexo
(SEIDEL JR., 2003; MAXWELL et al., 2004; MOCÉ et al., 2006).
É importante ressaltar que as populações de bezerros resultantes do sêmen sexado não
diferem da população de animais oriundos do sêmen convencional, incluindo o tempo de
gestação, peso ao nascer, taxa de mortalidade e ganho de peso (SEIDEL JR., 2003). Não
foram encontrados até o momento efeitos genotóxicos no sêmen sexado com o uso do
Hoechst 33342 (PARRILLA et al., 2004; GARNER, 2006).
Entretanto, o espermatozóide é sensível a diferentes etapas, incluindo o processo
através do citômetro de fluxo, como a ligação ao corante nuclear Hoechst, a alta diluição,
mudanças na composição dos diluidores, a alta pressão, as forças mecânicas associadas com a
passagem através da máquina, a exposição ao laser UV, a descarga elétrica e a projeção no
tubo coletor em alta velocidade e a centrifugação (MAXWELL; JOHNSON, 1999;
JOHNSON, 2000; GARCIA et al., 2007).
Baseando-se na literatura, o sêmen sexado apresenta algumas desvantagens que estão
relacionadas à sua fertilidade, avaliada através da taxa de prenhez que variou de 70 a 80% em
relação ao sêmen convencional (SEIDEL JR. et al., 1999; GARNER, 2006; MOCÉ et al.,
2006; WHEELER et al., 2006; SEIDEL JR., 2009) em bovinos (SEIDEL et al., 1999),
eqüinos (LINDSEY et al., 2002), ovinos (HOLLINSHEAD et al., 2002) e suínos
(PARRILLA et al., 2004). Dentre eles estão: a baixa concentração (2,1 x 106
espermatozóides) por dose (ANDERSSON et al., 2004; PARRILLA et al., 2004), embora
tenha sido reportado que doses similares tanto do sexado quanto do controle inseminadas
resultaram em taxas de prenhez similares (BODMER et al., 2005); o tempo de sobrevida
menor após a criopreservação (SHENK et al., 1999; SEIDEL JR.; GARNER, 2002;
WHEELER et al., 2006; CERCHIARO et al., 2007; GARCIA et al., 2007; RATH;
JOHNSON, 2008; DE VRIES et al., 2008; SEIDEL, 2009) e aumento de danos nas
membranas (PARRILLA et al., 2005).
A capacitação é definida como uma série de transformações que o espermatozóide
normalmente sofre durante a sua migração através do trato genital feminino, a fim de atingir e
ligar-se à zona pelúcida (ZP) do oócito, sofrendo reação acrossômica e fertilizando o oócito
(LAMIRANDE et al., 1997; URNER; SAKKAS, 2003; BREIBART et al., 2005; GADELLA,
2008). Durante este processo, há extensas alterações intracelulares, ocorrendo aumento
intracelular na concentração de Ca2+, no pH, na adenosina-monofosfato cíclica (cAMP) e na
fosforilação da proteína tirosina (PONS-REJRAJI et al., 2009a; VISCONTI; KOPF, 1998).

1 Introdução 24
Priscilla Harumi Tanno
Todavia, todo o mecanismo que engloba a capacitação ainda não foi completamente
esclarecido.
A fosforilação da tirosina é um pré-requisito necessário para a fertilização do oócito e
tem sido associada com a capacitação em várias espécies, incluindo a bovina (LAMIRANDE
et al., 1997; BRENER et al., 2003; URNER; SAKKAS, 2003; BREIBART et al., 2005),
provocando mudanças de hiperativação da motilidade (requerida para penetrar na ZP), reação
acrossômica, ligação da ZP (URNER; SAKKAS, 2003; BREIBART et al., 2005; GADELLA,
2008) e aquisição na capacidade fertilizante (BUFFONE et al., 2005, PONS-REJRAJI et al.,
2009a).
A regulação da fosforilação da tirosina em espermatozóides é decorrente de uma série
de eventos moleculares envolvendo efluxo de colesterol que aumenta a desordem da porção
dos fosfolipídeos, resultando em um aumento da permeabilidade da bicamada lipídica
(BUFFONE et al., 2005; BREIBART et al., 2005; THOMAS et al., 2006).
A desorganização da membrana pode ser avaliada através da utilização da sonda
Merocianina 540, que é uma sonda hidrofóbica que cora mais intensamente as membranas
que possuem maior desordem de seus componentes (translocação dos fosfolipídeos na
bicamada lipídica), como ocorre no espermatozóide capacitado. Também pode ser combinada
ao Yo-Pro-1, sonda que se liga ao DNA celular, possibilitando uma análise da viabilidade
celular associada ao estado da sua membrana lipídica. Esta associação de corantes pode ser
analisada pela técnica de citometria de fluxo (RATHI et al., 2001; THOMAS et al., 2006,
ANDRADE, 2009).
A reação acrossômica dos espermatozóides com membrana plasmática íntegra pode
ser analisada pela técnica de citometria de fluxo, utilizando uma associação de sondas
fluorescentes, a aglutinina de Pisum sativum (PSA) conjugada ao isotiocianato de fluoresceína
– FITC) e iodeto de propídeo (PI) (GRAHAM et al., 1990; SILVA; GADELLA, 2006;
CELEGHINI et al., 2007; ANDRADE, 2009).
As deteriorações das membranas espermáticas estão possivelmente envolvidas na
subfertilidade e/ou infertilidade dos machos (THOMAS et al., 1998; ƠCONNEL et al., 2002;
BROWERS et al., 2005; GILLAN et al., 2005; AMBROGI et al., 2006). Dentre os fatores que
influenciam estão o processo de criopreservação, pois cerca de 50% dos espermatozóides não
sobrevive e a maioria que resiste, desenvolve várias alterações fisiológicas que afetam sua
capacidade fertilizante (THOMAS et al., 1998; PONS-REJRAJI et al., 2009b) e a produção
excessiva de espécies reativas de oxigênio (EROs) (BURNAUGH et al., 2007).

1 Introdução 25
Priscilla Harumi Tanno
A produção excessiva de EROs causa a peroxidação lipídica da membrana plasmática
celular. O C11-Bodipy581/591, uma sonda fluorescente análoga de ácido graxo insaturado que
possui a capacidade de identificar mudanças nas suas propriedades fluorescentes após a
peroxidação lipídica. A sonda emite uma fluorescência em vermelho quando intercalada
dentro da membrana íntegra, que se altera para a tonalidade de laranja (540nm) após oxidação
pelo peroxinitrito e para o verde (520nm) após ataque com outros radicais oxidativos. É uma
sonda muito apropriada para localizar a peroxidação lipídica nas células espermáticas (usando
um microscópio confocal) e para detectar subpopulações de células espermáticas com
variadas sensibilidades para a peroxidação lipídica empregando a citometria de fluxo,
distinguindo a peroxidação lipídica mediada pelo peroxinitrito das outras EROs (BROWERS;
SILVA; GADELLA, 2005; SILVA; GADELLA, 2006).
De acordo com o exposto, as análises descritas acima são importantes ferramentas
para se determinar a influência da sexagem através da técnica de citometria de fluxo no sêmen
e quais alterações esta tecnologia provoca, pois ainda não foi totalmente elucidado na
literatura qual a real causa da menor fertilidade do sêmen sexado em relação ao sêmen
convencional após a criopreservação.

2 Revisão de Literatura

2 Revisão de Literatura 27
Priscilla Harumi Tanno
2 REVISÃO DE LITERATURA
A sexagem de sêmen através da citometria de fluxo, baseada na diferença de DNA, tem
mostrado ser um método bastante eficaz para diferenciar a determinação sexual entre os
gametas e separá-los para que possam predeterminar o sexo em mamíferos antes da fertilização
com uma acurácia de 90% (GARNER, 2006).
O processo de comercialização de sêmen sexado tem sido acelerado nos últimos anos.
Entretanto, esta tecnologia tem sido caracterizada pelo alto custo, complexidade de
implementação e baixas taxas de prenhez em comparação ao sêmen convencional. E apesar do
sêmen sexado bovino congelado ser produzido comercialmente em vários países, o número de
touros ainda é limitado nesta técnica (SEIDEL, 2007).
2.1 HISTÓRICO DA SEXAGEM
Na primeira metade do século XX, houve grandes avanços científicos na biologia,
especialmente na genética, resultando em numerosas descobertas, incluindo a identificação dos
cromossomos sexuais. A primeira documentação de identificação microscópica dos
cromossomos sexuais encontra-se em 1910 por Guyer (SEIDEL; GARNER, 2002). Entretanto,
somente em 1970, com Barlow e Vossa, foi documentada convincentemente a existência dos
cromossomos sexuais em humanos empregando-se o corante quinacrina (SEIDEL; GARNER,
2002).
Mas foi somente em 1976 que ocorreram os primeiros experimentos relacionados à
análise espermática com o citômetro de fluxo com Gledhill et al. Porém, os resultados
falharam até o desenvolvimento da análise de eritrócitos de galinha em dois feixes de líquido
de sheathfluid para resolver o problema de orientação da célula achatada (FULWYLER,
1977). Para aumentar a resolução da análise de células achatadas foi realizada uma adaptação
de um tubo de injeção para orientar forma e a inclusão de um segundo detector de luz
(GARNER; SEIDEL, 2008). Mas somente Pinkel et al. (1982) modificaram o sistema para
orientar o espermatozóide de frente para o feixe de laser. Estas modificações foram
fundamentais para detectar diferenças no cromossomo X ou Y no DNA espermático,
reportado por Garner et al. (1983); no entanto, estes procedimentos lesavam severamente a

2 Revisão de Literatura 28
Priscilla Harumi Tanno
célula espermática devido à remoção da cauda e das membranas para corar o núcleo com o
corante 4’-6-diamino-2-fenilindol (DAPI), cuja membrana plasmática íntegra é impermeável,
tornando a técnica impraticável até então (GARNER, 2006; SEIDEL 2007) .
Foi somente em 1996, com o emprego do corante de DNA que atravessa a membrana
plasmática íntegra, o bisbenzimidazol, Hoechst 33342, que houve a primeira publicação de
um método (Beltsville Sperm Sexing Technology) com repetibilidade para separar populações
de espermatozóides vivos de mamíferos (cromossomos X do Y), um sistema de sexagem em
alta velocidade e uma orientação através do nozzle (GARNER, 2006; RATH et al., 2009). Em
2001, a tecnologia foi adaptada para a produção comercial através da XY, Inc (Fort Collins,
Colorado e atualmente, Sexing Technologies Navasota, Texas, ambas nos EUA) para sêmen
bovino. O primeiro centro de produção comercial usando o sistema em alta velocidade foi a
Cogent em Chester, Reino Unido (RATH et al., 2009).
2.2 DIFERENÇAS NO DNA ESPERMÁTICO
Em bovinos, cada célula somática possui 60 cromossomos. Os gametas masculinos,
sendo células haplóides, contêm 29 cromossomos autossomos mais o cromossomo Y que
determina o sexo masculino ou o cromossomo X, já que a célula diplóide do macho é
composta dos dois cromossomos (XY ou XX) (GARNER; SEIDEL JR., 2008).
A sexagem do sêmen bovino ocorre devido ao fato do cromossomo X conter em média
3,8% mais DNA do que o cromossomo Y, porém existe uma diferença no conteúdo de DNA
das células espermáticas mesmo entre raças. Touros europeus (Bos taurus taurus) e os
originados na Ásia (Bos taurus indicus) apresentam uma diferença de tamanho no
cromossomo Y entre as duas espécies. O núcleo espermático de touros da raça Brahman, por
exemplo, possui uma diferença menor entre o cromossomo X e o Y, por volta de 3,7%; por
outro lado, a raça Jersey é a que possui a maior diferença entre eles, ao redor de 4,2%
(GARNER, 2006).
A cabeça do espermatozóide quanto mais achatada e oval for, a tendência dele ser
orientado e separado pelo citômetro de fluxo (SX MOFLO®, DakoCytomation Inc., Fort
Collins, CO) é mais rápida do que gametas possuindo cabeças com formato redondo ou
angular. O índice que avalia a qualidade de separação desses espermatozóides é obtido pela

2 Revisão de Literatura 29
Priscilla Harumi Tanno
multiplicação da diferença no DNA do cromossomo X do Y, em porcentagem, pela área da
porção achatada da cabeça do espermatozóide. O espermatozóide bovino é o que possui a
maior área (34,5 µm2) e pelo corte sagital, é um dos mais achatados entre as espécies de
mamíferos domésticos (GARNER, 2006).
A técnica utilizada na atualidade envolve um tratamento das células espermáticas com
o corante fluorescente, Hoechst 33342 (H33342) (trihidrato de trihidrocloreto, invitrogen™
Molecular probes, Eugene, Oregon, USA). É um permeador de membrana que se liga
fortemente e de maneira específica a quatro pares de bases AATT na curvatura menor da
dupla hélice do DNA (GARNER, 2009). Em seguida, as células coradas passam pelo
citômetro de fluxo através do nozzle tip (Cytonozzle™, parte integrante da MOFLO® SX)
que orienta os espermatozóides para serem expostos ao laser UV e forma as gotas na coluna
de fluido devido a vibrações estabelecidas pelo mecanismo pizoelétrico na coluna
(MAXWELL et al., 2004; SUH et al., 2005). Gotas contendo um único espermatozóide
orientado são carregadas eletricamente antes de passarem pelo fluxo eletrostático para a
separação. O feixe de gotas separadas cai em tubos coletores contendo diluidor à base de
gema de ovo com uma velocidade equivalente de 50 km/h. Posteriormente, estes tubos são
centrifugados e o sedimento contendo o sêmen é diluído na concentração desejada para a
preservação espermática (RATH et al., 2009).
As células coradas emitem uma fluorescência de cor azul de 351 e 364 nm de
comprimento de onda quando iluminadas pelo laser VanguardTM, sistema de estado sólido de
laser pulsado (Newport Corporation, Irvine, CA, USA), que evita efeitos lesivos de UV de
baixo comprimento de onda que são absorvidos pelos ácidos nucléicos e proteínas (SEIDEL;
GARNER, 2002), proporcionando uma medida precisa dos espermatozóides que contêm no
DNA o cromossomo X ou o Y, sendo separadas pelo citômetro de fluxo modificado
especificamente para o espermatozóide, ou seja, o espermatozóide corado contendo o
cromossomo X emite proporcionalmente uma maior fluorescência do que o que contém o Y
(JOHNSON; WELCH, 1999; GARNER, 2001; GARNER; SEIDEL JR., 2003; GARNER,
2006).
Quando a fluorescência dos espermatozóides é medida utilizando-se um citômetro de
fluxo, as diferenças entre os cromossomos X e Y são observadas em um histograma com
distribuição dupla Gaussiana (SHARPE; EVANS, 2009).
A eficácia do sistema de separação dos espermatozóides foi aumentada pela
identificação das células mortas ou lesadas identificadas primeiramente pela sonda iodeto de

2 Revisão de Literatura 30
Priscilla Harumi Tanno
propídeo (PI), cuja membrana plasmática íntegra é impermeável ao corante. Os gametas
comprometidos eram identificados e assim descartados junto com as células que não foram
medidas apropriadamente. A substituição do PI, que se intercala entre os pares de bases do
DNA, pelo FD&C #40 (Warner Jenkinson, St. Louis, EUA) proporcionou uma maior
segurança porque este simplesmente bloqueia a fluorescência do Hoechst e não é mutagênico
(JOHNSON; WELCH, 1999; SCHENK et al., 1999; GARNER, 2006).
Entretanto, a eficiência do H33342 nas células espermáticas bovinas é baixa, pois
somente um terço dos espermatozóides que passam pelo sistema pode ser separado e 20% dos
que são separados, são perdidos durante o processo de concentração e envase do sêmen
(SEIDEL; GARNER, 2002; GARNER, 2006; GARCIA et al., 2007).
Outros fatores citados como lesivos a espermatozóides no processo são o uso do
corante de DNA (H33342), reaquecimento e incubação, forças mecânicas durante a passagem
pelo citômetro de fluxo, a exposição intensa ao laser UV, a carga elétrica, a alta gravidade
durante a desaceleração dentro do tubo coletor, a alta diluição, pressão elevada e resistência a
mudanças na composição dos meios diluidores, podendo levar a diminuição na capacidade de
fertilização espermática na inseminação artificial e, conseqüentemente, em menor taxa de
desenvolvimento embrionário, comparado ao sêmen não-sexado (GARNER; SEIDEL JR.,
2003; GARNER, 2006; MOCÉ et al., 2006; GRAAF et al., 2007; GARCIA et al., 2007).
A alta diluição para a sexagem de sêmen por citometria de fluxo é necessária para a
identificação e separação da célula espermática. A esta etapa segue-se a concentração antes da
inseminação artificial. Entretanto, em várias espécies, como por exemplo, ratazana,
camundongo, homem e o cachaço, a centrifugação é um procedimento potencialmente lesivo
durante o processo, levando a uma redução da sobrevida do espermatozóide em conseqüência
de um efeito direto nas membranas (GARCIA et al., 2007).
Testes de viabilidade espermática com SYBR-14, que coram os espermatozóides
vivos, e o PI, células mortas ou com lesão celular, mostraram que o estresse mecânico da
separação e a centrifugação aumentam a mortalidade ou lesões aos espermatozóides (SUH et
al., 2005; GARNER, 2006; AMBROGI et al., 2006, SILVA; GADELLA, 2006).
A diminuição do sistema de pressão de 50 para 40 psi proporcionou uma melhora na
qualidade do sêmen, tais como na motilidade, espermatozóides vivos com membrana intacta,
um aumento na clivagem e desenvolvimento blastocístico sem, no entanto, diminuir a
performance das máquinas (CAMPOS-CHILLON; DE LA TORRE, 2003; SUH et al., 2005).

2 Revisão de Literatura 31
Priscilla Harumi Tanno
Embora a separação das células resulte na eliminação de espermatozóides mortos ou
lesados, aquelas células que sobrevivem ao processo de sexagem tendem a se deteriorar mais
rapidamente do que o controle, em touros e em bodes, mas, aparentemente, isso não ocorre
em sêmen de garanhões e de carneiros (MORRIS et al., 2003; GARNER, 2006).
Outro fator concomitante que pode afetar a diminuição da fertilidade, além do
processo da sexagem em si, é a baixa dose por palheta, porém isso diferiu entre touros
(FRIJTERS et al., 2009).
Parrilla et al. (2005) observaram em sêmen de cachaços que o processo através da
citometria de fluxo é capaz de manter a motilidade, viabilidade e integridade acrossomal em
ótimos níveis durante 10 horas de armazenamento após a separação dos espermatozóides,
porém a capacidade fertilizante foi mantida somente durante 5 horas após a sexagem, já em
eqüinos, a integridade acrossômica manteve-se por 24 h, equivalente ao sêmen a fresco.
Sendo que, após a sexagem, o sêmen armazenado por 12 h a 20ºC resultou em maior
motilidade do que a 4ºC. Foi observado, que o período de coloração e incubação das células
com o H33342 resultou em um decréscimo na proporção de acrossomas intactos (MORRIS et
al., 2003).
2.3 O ESPERMATOZÓIDE
Os espermatozóides são formados dentro dos túbulos seminíferos dos testículos e
armazenados na região do epidídimo, onde adquirem potencial para a motilidade e habilidade
de fertilização, caso receba o estímulo apropriado (PONS-REJRAJI et al., 2009a;
LAMIRANDE, O’FLAHERTY, 2008; SILVA; GADELLA, 2006).
Os espermatozóides completamente desenvolvidos são células alongadas, consistindo
de uma cabeça achatada contendo o núcleo com o genoma masculino e de uma cauda ou
flagelo com o aparelho necessário para a motilidade celular e envolvendo toda a célula, a
membrana plasmática, semi-permeável, que possui um papel muito ativo na capacidade de
fertilização espermática, não sendo, simplesmente, um filme lipídico disposto em bicamada,
passiva, na qual receptores recebem sinais moleculares específicos (LENZI et al., 1996;
LAMIRANDE; O’FLAHERTY, 2008). O acrossomo, ou capa acrossomal, é uma estrutura de
parede dupla situada entre a membrana plasmática e a porção anterior da cabeça. O anel

2 Revisão de Literatura 32
Priscilla Harumi Tanno
posterior separa a cabeça do espermatozóide da peça intermediária que contém mitocôndrias,
envolvidas na produção de energia, sendo esta separada do flagelo, que é subdividido em peça
principal e terminal pelo anel anular (FLESCH; GADELLA, 2000; GARNER; HAFEZ, 2004;
LAMIRANDE; O’FLAHERTY, 2008). O anel posterior e o annulus são estruturas físicas no
interior da membrana plasmática que funcionam como barreiras de difusão para
compartimentalizar a membrana da peça-intermediária e peça principal (JONES et al., 2007).
Até o momento, ainda não está claro como o espermatozóide fertiliza o oócito, embora
seja evidente que somente espermatozóides funcionalmente intactos sejam capazes de
fertilizar e que, de alguma forma, é realizado pela superfície da cabeça espermática
(GADELLA, 2008).
2.3.1 A membrana plasmática
A membrana plasmática é organizada por uma bicamada lipídica formada por
fosfolipídeos e esteróis, na qual as regiões apolares das moléculas dos lipídeos são orientadas
para o centro da bicamada, e os grupos polares, para a região extracelular ou em contato com
o citoplasma, de acordo com o modelo do mosaico fluido, postulado por Singer e Nicolson1
(1972 apud LEHNINGER et al., 2002, p.320); ANDRADE, 2009).
A membrana é formada por significantes níveis de ácidos graxos poliinsaturados
(PUFAs), derivados principalmente do ácido linoléico, importantes na manutenção da fluidez
e da flexibilidade da biomembrana, já que são mais flexíveis do que os saturados (LENZI et
al., 1996). A membrana plasmática tem uma distribuição de fosfolipídeos assimétrica em toda
a sua extensão. Dentre eles, a fosfatidilserina e a fosfatidiletanolamina estão localizadas
principalmente no folheto interno da membrana, e no folheto externo são encontrados a
esfingomielina e a fosfatidilcolina. Essa assimetria é mantida pela ação de várias translocases
ATP-dependente (JANUSKAUSKAS et al., 2003).
A superfície da membrana plasmática é heterogênea e de acordo com estudos
anteriores, sabe-se que as moléculas da superfície espermática exibem propriedades de
1SINGER, S. J.; NICOLSON, G. L. The fluid mosaic model of the structure of cell membranes. Science, v. 175, p. 720-731, 1972.

2 Revisão de Literatura 33
Priscilla Harumi Tanno
difusão lateral, mas permanecem presas a regiões específicas da superfície. Uma
reorganização deste mosaico da superfície espermática sucede-se quando o espermatozóide
reside na proximidade do oócito, simultaneamente quando eles tornam-se competentes para
fertilizar (GADELLA, 2008).
A concentração de fosfolipídeos polinsaturados na membrana indica que os
espermatozóides são extremamente sensíveis ao estímulo externo. A função fertilizante dos
espermatozóides poderia ser explicada por que uma célula espermática tem uma fluidez,
flexibilidade e muita atividade da membrana, que pode ser facilmente desestabilizada e
ativada (LENZI et al., 1996).
2.3.1.1 Função da membrana plasmática
Durante a passagem do testículo para o sítio de fertilização na tuba uterina, os
espermatozóides de mamíferos encontram uma grande variação de fluidos de diferentes
origens e composições, como por exemplo, comparando-se o fluido testicular com o plasma
seminal e estes com as secreções do trato uterino. Estes fluidos têm principal influência nos
processos de desenvolvimento pós-testicular, como a maturação e a capacitação, e como
resultado, os espermatozóides são transformados de um estado imóvel e infértil para células
vigorosamente ativas com capacidade de se ligar especificamente e, por último, fundir-se ao
oócito. O desenvolvimento pós-testicular dos espermatozóides deve ser considerado como
uma hierarquia de sinalizações, iniciados por diferentes agonistas dentro de diversos fluidos,
que levem o espermatozóide a atingir o local da fertilização em um estado totalmente apto.
Este cenário assegura que não resolva apenas o espermatozóide completar todos os estágios
de desenvolvimento na sequência correta, mas que também não respondam a sinais externos
prematuramente (JONES et al., 2007).
O espermatozóide é uma célula altamente diferenciada e como tal, a superfície da
membrana é altamente compartimentalizada o que resulta na presença restrita de certos
lipídeos e proteínas (LENZI et al., 1996; JONES et al., 2007). De acordo com estudos em
outros tipos celulares, sabe-se que a ligação com agonistas e ativação do receptor
correspondente, frequentemente envolve um complexo grupo multi-molecular, que transmite
sinais externos através da membrana plasmática para o interior do citoplasma, onde são

2 Revisão de Literatura 34
Priscilla Harumi Tanno
amplificados para provocar uma resposta (JONES et al., 2007). Como o espermatozóide não
sintetiza novas proteínas de membranas, a formação de complexos de sinalização requer que
moléculas separadas espacialmente, possivelmente de diferentes regiões da célula trabalhem
em conjunto. Estes complexos podem mostrar migrações polarizadas de regiões onde eram
áreas inativas para áreas onde se tornam completamente funcionais (HOWES et al., 2001).
Estes complexos ou em sua forma individual estão sujeitos a forças de difusão que em muitas
situações são geradas por migrações polarizadas controladas por mecanismos ainda
desconhecidos, mas que podem envolver o citoesqueleto (JONES et al., 1990).
2.3.1.2 Alterações da membrana plasmática
As alterações de membrana podem variar na mudança de organização, fluidez,
permeabilidade e composição lipídica da bicamada à total ruptura de membrana
(JANUSKAUSKAS et al., 2003).
Mudanças na estrutura e na integridade da membrana plasmática parecem ser um
importante componente associado com a redução da fertilidade dos espermatozóides pós-
descongelamento (THOMAS; MEYERS; BALL, 2006).
Sabe-se que a membrana plasmática é adversamente afetada pela citometria de fluxo e
pelo processamento do sêmen, o que limita a viabilidade, a capacidade de armazenamento, a
habilidade de fertilização e, além disso, pode acelerar o processo de reação acrossômica após
a criopreservação (MOCÉ et al., 2006; GARCIA et al., 2007).
Uma das funções da membrana plasmática é regular o volume celular, que é crucial
durante a criopreservação (PETRUNKINA et al., 2005).
Foi observada uma correlação entre deterioração de membrana induzida pela
criopreservação e perda da integridade acrossômica e motilidade e a resposta do sêmen a
fresco ao ionóforo de cálcio em ejaculados de cães (PETRUNKINA et al., 2005). O mesmo
foi revelado em uma análise por meio da citometria de fluxo, no qual o sêmen sexado e não-
sexado a fresco possuíram, significativamente, porcentagens menores de reação acrossômica,
lesão de membrana e de atividade mitocondrial, quando comparados com o sêmen congelado
e descongelado sexado e não-sexado (BLONDIN et al., 2009). Januskauskas et al. (2001)
utilizaram H33258 para detectar espermatozóides de touros não-viáveis através da citometria

2 Revisão de Literatura 35
Priscilla Harumi Tanno
de fluxo, observou uma correlação negativa entre a porcentagem de células com membrana
lesada e fertilidade a campo (r= 0.57).
Foi demonstrado que a translocação da fosfatidilserina para a área externa da
membrana, pode ser utilizada como um marcador para a função de membrana deteriorada
pós-descongelação em humanos e bovinos através da anexina-V (cálcio-dependente)
combinada ao iodeto de propídeo (PI), permitindo a detecção simultânea de apoptose celular
e/ou células necróticas, ou células com membrana plasmática comprometida
(JANUSKAUSKAS et al., 2003). Verificou-se que a lesão de membrana causada pelo
processo de criopreservação foi mais pronunciada no sêmen sexado do que no grupo controle
porque exibiu estágios mais avançados de capacitação do que o segundo. Resultados similares
têm sido observados em sêmen de carneiros. Com isso, é razoável que o sêmen sexado
apresente uma fertilidade inferior in vivo comparado ao sêmen convencional (MOCÉ;
GRAHAM; SCHENK, 2006).
2.3.2 Capacitação espermática
Após a saída dos testículos, espermatozóides de mamíferos de muitas espécies são
morfologicamente diferenciados, no entanto, não possuem a capacidade para fertilizar o
oócito em metáfase II. Durante o trânsito epididimário, os espermatozóides adquirem a
habilidade de mover-se progressivamente, entretanto eles ainda são incapazes de fertilizar um
oócito logo após a ejaculação. Essa capacidade é adquirida no trato reprodutivo da fêmea em
um período finito de tempo. As mudanças fisiológicas que conferem a capacidade fertilizante
ao espermatozóide são coletivamente denominadas de capacitação (VISCONTI et al., 1995;
VISCONTI; KOPF, 1998; BREIBART; NAOR, 1999; GADELLA, 2008; WERTHEIMEIR
et al., 2008; PONS-REJRAJI et al., 2009a; PONS-REJRAJ et al., 2009b).
A capacitação foi descrita primeiramente por Chang e Austin, independentemente, em
1951. Eles descobriram que os espermatozóides eram incapazes de fertilizar o oócito a menos
que eles residam no trato genital feminino durante um período específico para sofrer
transformações para tornarem-se férteis; todavia, até o momento a definição de todo o
fenômeno ainda não foi totalmente elucidado (DE LAMIRANDE et al., 1997; VISCONTI;
KOPF, 1998; PONS-REJRAJI et al., 2009b).

2 Revisão de Literatura 36
Priscilla Harumi Tanno
A capacitação envolve aumento na fluidez da membrana plasmática associado a
modificações lipídicas, aumento na permeabilidade de vários íons na cabeça espermática e no
flagelo, geração de espécies reativas de oxigênio (EROs) pelo espermatozóide, assim como a
fosforilação de proteínas (serina, treonina e tirosina) (LAMIRANDE et al., 1997b; RATHI et
al., 2001; LAMIRANDE; O’FLAHERTY, 2008; PONS-REJRAJ et al., 2009a).
Em mamíferos, a capacitação pode ocorrer in vitro espontaneamente em um meio
definido sem a adição de fluidos biológicos, sugerindo que este processo seja intrinsecamente
modulado pelo espermatozóide, desde que seja incubado em um meio apropriado. Embora
não haja um diluidor de capacitação universal, certos componentes são fundamentais,
incluindo a albumina sérica, Ca2+ e o HCO3-, tendo papéis regulatórios importantes para
promover a capacitação, induzindo a fosforilação da tirosina (VISCONTI; KOPF, 1998;
FLESCH; GADELLA, 2000; GADELLA, 2008).
A albumina sérica, geralmente a albumina bovina sérica (BSA), presente no meio de
capacitação in vitro para espermatozóides de mamíferos (camundongo, hamster, bovino,
humano), funciona como um carreador de colesterol, ligando-se e removendo-o da membrana
plasmática, alterando a fluidez da membrana, que ocorre durante a capacitação (VISCONTI;
KOPF, 1998).
O papel do Ca2+ na iniciação e/ou regulação da capacitação é controverso até o
momento. Alguns autores têm descrito um aumento intracelular de Ca2+ durante a
capacitação, enquanto outros têm mostrado que o Ca2+ intracelular não se altera durante este
evento de maturação (VISCONTI; KOPF, 1998). No espermatozóide de camundongos, há a
evidência de que o Ca2+ seja requerido para a capacitação, embora estes autores não tenham
mensurado a concentração de Ca2+ intracelular (DASGUPTA et al., 1993; VISCONTI et al.,
1995). A ação do Ca2+ em nível de enzimas efetoras envolve transdução de sinais no
espermatozóide, como na adenil ciclase, no nucleotídeo de fosfodiesterase cíclico, sugerindo
que este cátion bivalente tem uma importante função na capacitação (VISCONTI; KOPF,
1998).
O bicarbonato (HCO3-) é um dos principais componentes do meio e sua omissão não
somente inibe a fosforilação da tirosina, mas também reduz a capacidade dos espermatozóides
se ligarem às proteínas solubilizadas da zona pelúcida (ZP). O efluxo de colesterol pode
induzir a entrada de bicarbonato, que ativa diretamente uma adenilato ciclase (AC)
espermática específica e, desse modo, induz ao aumento dos níveis de cAMP na célula

2 Revisão de Literatura 37
Priscilla Harumi Tanno
espermática (FLESCH et al., 1999; FLESCH; GADELLA, 2000; HARRISON; GADELLA,
2005).
A subsequente ativação da proteína quinase A (PKA) induz via mecanismo de
sinalização ainda desconhecido, a fosforilação da tirosina de diversas proteínas (FLESCH et
al., 1999).
A célula espermática é submetida a severas condições durante o processo de sexagem,
podendo induzir a uma capacitação parcial (BLONDIN et al., 2009). Em vários trabalhos,
tem-se relatado uma capacitação prematura do sêmen sexado em relação ao controle
semelhante ao que ocorre com o sêmen de caprinos quando congelados (JONHSON, 2000).
Foi observada uma correlação significativa existente entre a porcentagem de reação
acrossômica detectada pela análise através de citometria de fluxo e a porcentagem resultante
de blastocistos produzidos (r = 0,74) (BLONDIN et al., 2009).
Recentemente, foi demonstrado em cachaços que o período de incubação com H33342
induz a uma mudança de pré-capacitação na membrana plasmática, indicado pela
redistribuição da proteína “heat shock” (Hsp70) (SPINACI et al., 2006), enquanto que a
passagem através do citômetro de fluxo e a exposição ao laser foram menos lesivos à
membrana. Entretanto, a carga elétrica e a subseqüente deflecção induziram ainda mais a
relocalização da Hsp70, como também durante o seu armazenamento no fluido de “catch”
(RATH, 2009).
Sinalizadores celulares nos espermatozóides de eqüinos capacitados in vitro parecem
ser dependentes do processo de ativação do ciclo AMPc/PKA, enquanto que as células
espermáticas criopreservadas seguem através de caminhos alternativos, sugerindo que a idéia
de capacitação e criocapacitação não sejam processos equivalentes (THOMAS; MEYERS;
BALL, 2006).
2.3.3 Fosforilação de proteínas
A fosforilação de proteínas é uma modificação pós-translacional de proteínas que
permite à célula controlar vários processos celulares. O estado de fosforilação de
fosfoproteínas é controlado pela atividade de proteínas quinases e fosfatases (URNER;
SAKKAS, 2003).

2 Revisão de Literatura 38
Priscilla Harumi Tanno
Espermatozóides maduros possuem sua própria idiossincrasia particular como células
altamente especializadas. Eles são altamente compartimentalizados, transcricionalmente
inativos e incapazes de sintetizar novas proteínas. Durante a fertilização, a função espermática
é regulada pela ativação intracelular de sistemas de sinalização que controlam a fosforilação
de proteínas. Nos espermatozóides, ocorrem a fosforilação de serina/treonina e tirosina, mas
somente algumas proteínas fosforiladas podem ser identificadas. Embora a via do cAMP
dependente da proteína quinase A represente um papel central na função espermática e tenha
sido estudada em detalhe (VISCONTI; KOPF, 1998), o conhecimento profundo sobre tirosina
quinases e outras serina/treonina quinases permanece limitado (URNER; SAKKAS, 2003).
A fosforilação da proteína tirosina aumenta no espermatozóide durante a capacitação
em várias espécies, incluindo ratos, humanos, bovinos, suínos, hamsters e gatos (VISCONTI;
KOPF, 1998; FLESCH; GADELLA, 2000; URNER; SAKKAS, 2003). A fosforilação da
tirosina parece ser um pré-requisito necessário para um espermatozóide fertilizar um oócito
(URNER; SAKKAS, 2003).
Os processos regulados pela fosforilação da tirosina incluem capacitação,
hiperativação da motilidade, a reação acrossômica e a afinidade à zona pelúcida, sendo que
todas elas são requeridas para que o espermatozóide alcance e fusione-se ao oócito (FLESCH;
GADELLA, 2000; URNER; SAKKAS, 2003). Em geral, a fosforilação de proteínas em
espermatozóides de mamíferos tem sido estudada através do exame de proteínas fosforiladas
em populações espermáticas durante a capacitação e após a exposição aos agentes indutores
de reação acrossômica pela técnica do western blot (FLESCH et al., 1999; URNER;
SAKKAS, 2003; PONS-REJRAJI, 2009a; PONS-REJRAJ, 2009b). Estes estudos levaram ao
acúmulo de informações sobre o peso molecular das proteínas fosforiladas e sua regulação
durante a capacitação e reação acrossômica. A fosforilação no espermatozóide pode ser
avaliada também pela imunocitoquímica, que revela a compartimentalização das
fosfoproteínas no espermatozóide individual e em populações heterogêneas de
espermatozóides (URNER; SAKKAS, 2003).
A fosforilação da tirosina pode induzir a mudanças conformacionais nas proteínas,
desta maneira, leva a uma ativação ou inativação das proteínas (FLESCH et al., 1999). A
fosforilação da tirosina tem sido claramente associada com a aquisição de motilidade e a
indução da capacitação. O papel do monofosfato de adenosina cíclico (cAMP) no controle dos
eventos de fosforilação da tirosina é bem documentada em várias espécies via uma cascata de
transdução de sinais que parece ser única para o gameta masculino. Este mecanismo da

2 Revisão de Literatura 39
Priscilla Harumi Tanno
proteína quinase A-tirosina quinase parece ser regulado através de redução-oxidação (LEWIS;
AITKEN, 2001).
2.3.4 Reação acrossômica (RA)
Espermatozóides capacitados são submetidos à reação acrossômica quando eles
entram em contato com a zona pelúcida que envolve o oócito. A reação acrossômica é
desencadeada naturalmente pela ZP3 que é uma das principais proteínas da zona pelúcida.
Porém, não está disponível comercialmente (a proteína recombinante não é ativa), mas pode
ser reproduzido in vitro utilizando substâncias modelo, tais como ionóforo de cálcio
(A23187), agentes perturbadores de membrana (lisofosfatidilcolina, LPC), ou progesterona
(LAMIRANDE; O’FLAHERTY, 2008).
A reação acrossômica é um evento irreversível, rápido e exocitótico, essencial para o
espermatozóide ligar-se à zona pelúcida e, em seguida, fundir-se com a membrana plasmática
do oócito (RATHI et al., 2001). É um processo complexo, que parece ser controlado por
várias quinases e vias de bloqueio através dos inibidores de PKC, PKA, PTK, P13K, Akt e da
via do ERK, é regulada também pelo processo de regulação redox (DE LAMIRANDE;
O’FLAHERTY, 2008). Está associada com modificações celulares que parecem similares
àquelas da capacitação, como as fosforilações, influxo de Ca2+, geração de EROs, mas os
agentes iniciantes, a cinética, a amplitude das modificações, enzimas, compartimentalização
celular são diferentes (VISCONTI; KOPF, 1998; RATHI et al., 2001; GILLAN et al., 2005;
DE LAMIRANDE; O’FLAHERTY, 2008; BLONDIN et al., 2009).
Ocorrem várias mudanças na superfície da membrana plasmática que são
provavelmente requeridas para permitir ao espermatozóide ligar-se à matriz extracelular do
oócito (ZP), que induz à reação acrossômica (GADELLA, 2008). No momento em que o
espermatozóide encontra o oócito na tuba uterina [ou durante a fertilização in vitro (FIV)
quando se mistura com o oócito], a superfície apical da cabeça do espermatozóide contém
complexas proteínas de membrana funcionais que reconhecem e ligam-se à ZP (GADELLA,
2008). A membrana plasmática apical da cabeça do espermatozóide começa a se fusionar com
a membrana externa do acrossomo subjacente em múltiplos lugares, resultando na dispersão
do conteúdo acrossomal (FLESCH; GADELLA, 2000).

2 Revisão de Literatura 40
Priscilla Harumi Tanno
A maquinaria enzimática requerida para a exocitose e o início da penetração da zona
pelúcida é somente operante funcionalmente na região apical da superfície da cabeça
espermática na proximidade da interação com a zona pelúcida (GADELLA, 2008). Durante a
reação acrossômica enzimas hidrolíticas e proteolíticas são secretadas para liberar, hidrolizar
e dissolver a matriz da ZP localmente na direção imediata da penetração da célula espermática
que, finalmente, assegura a entrada do espermatozóide dentro do espaço perivitelino
(FLESCH; GADELLA, 2000).
A reação acrossômica não deve prosseguir antes da ligação com a ZP porque a parte
do maquinário enzimático desta organela requerida para o sucesso da penetração da ZP será
perdida. Reações acrossômicas preliminares das células espermáticas são consideradas
incompetentes para fertilizar o oócito. Células espermáticas que chegam ao espaço
perivitelino são sempre células com o acrossomo reagido, capazes de se fusionar com o oócito
(FLESCH; GADELLA, 2000).
Antes da fusão para a fertilização, o espermatozóide precisa secretar suas enzimas
acrossomais para penetrar na zona pelúcida. Esta então chamada de reação acrossômica é
desencadeada pela ZP e possuem múltiplos pontos de eventos de fusão entre duas membranas
posicionadas bem próximas. Nas células somáticas, as SNAREs (soluble N-ethylmaleimide-
sensitive factor attachment protein receptor proteins) estão envolvidas na fusão das
membranas, responsável pela secreção das células (GADELLA, 2008). SNAREs requeridas
para a exocitose têm sido identificadas em espermatozóides de mamíferos (GADELLA,
2008). Foram identificadas em espermatozóides de rato, suíno, humano, camundongo. Todas
as SNAREs que foram requeridas para formar complexo de SNARE envolveram fusão de
membrana (GADELLA, 2008). Em espermatozóides de suínos, estas sintaxes (membrana
plasmática SNAREs) mostraram agregação similar para a área do cume apical da superfície
plasmática, como também, foi observado na região para a zona de ligação de proteínas
quando os espermatozóides são capacitados (GADELLA, 2008). Um importante canal de K+
foi identificado no complexo da zona de ligação, que pode estar indiretamente envolvido no
influxo de cálcio para o interior da cabeça do espermatozóide que é, por sua vez, requerido
pela SNARE para que elas se juntem e tenham mudanças conformacionais para que haja a
fusão das membranas do acrossomo (GADELLA, 2008).

2 Revisão de Literatura 41
Priscilla Harumi Tanno
2.4 ESPÉCIES REATIVAS DE OXIGÊNIO (EROs)
Os espermatozóides, como qualquer célula aeróbia, produzem EROs, sendo a maior
parte originada da atividade metabólica normal. As EROs são reconhecidas por seus efeitos
deletérios em quase todos os tecidos e células. Dentre as EROs formadas, podemos citar o
anion superóxido (O2-•), peróxido de hidrogênio (H2O2), os radicais hidroxil (OH•),
hidroperoxil (HO2•), peroxil (ROO-) e alcoxil (RO•), e ainda o ácido hipocloroso (HOCl)
(AITKEN, 1995; DE LAMIRANDE et al, 1997; LEWIS; AITKEN, 2001; SANOCKA;
KURPISZ, 2004; DE LAMIRANDE; O’FLAHERTY, 2008).
Entretanto, foi somente no início dos anos 90, que pesquisadores descobriram que
baixas concentrações de EROs, produzidas no momento da ativação celular, poderiam fazer o
papel de segundo mensageiros e por conseguinte, ter um desempenho positivo nas cascatas de
transdução de sinais (DE LAMIRANDE; O’FLAHERTY, 2008). A geração de EROs pelos
espermatozóides de mamíferos tem um importante papel fisiológico no controle tanto da
capacitação espermática como na reação acrossômica (LEWIS; AITKEN, 2001).
O peróxido de hidrogênio em baixas concentrações induz a capacitação pela promoção
da fosforilação da tirosina, mas altas concentrações (>500 µM) bloqueiam este processo. Por
outro lado, o sêmen tratado com um coquetel oxidante contendo 100 µM de ácido ascórbico,
20 µM de FeSO4 e 500 µM de H2O2 provocou uma peroxidação lipídica da membrana
plasmática, oxidação intracelular e mitocondrial (SILVA et al., 2007).
Um dos principais mecanismos de ação que as EROs agem na transdução de
elementos e vias é pela modulação redox dos pares de sulfidril/dissulfeto nas proteínas.
Condições oxidativas promovem a formação de pontes de dissulfeto, a liberação de Zn2+,
resultando em uma modificação na atividade enzimática. Vários oxidantes de grupos de
sulfidril, como o N-etilmaleimida e a diamida, desencadeiam a capacitação e a geração de O-
•2, inversamente, redutores de dissulfeto, como a glutationa e ditiotreitol, previnem estes
processos (DE LAMIRANDE; O’FLAHERTY, 2008).
As espécies reativas de oxigênio (EROs), tais como o ânion superóxido (•O2-),
peróxido de hidrogênio (H2O2) e o óxido nítrico (NO•), são consideradas elementos
regulatórios. Sabe-se que as EROs, em baixas concentrações, ativam diferentes vias de
transdução de sinal, levando à fosforilação da tirosina (reconhecida como um componente
bioquímico fundamental do processo de capacitação), serina e treonina, mais provavelmente

2 Revisão de Literatura 42
Priscilla Harumi Tanno
via cAMP/proteína quinase (PKA), regulação de sinal extracelular de quinase (ERK), família
de proteínas quinases mitógenas-ativadas (MAPK) e vias PKC (VISCONTI et al., 1995;
LEWIS; AITKEN, 2001; DE LAMIRANDE; O’FLAHERTY, 2008; PONS-REJRAJI et al.,
2009a).
O tratamento dos espermatozóides com antioxidantes [superóxido dismutase (SOD) e
catalase] ou inibidores de NOS (L-NAME, L-NMMA) inibe o aumento de cAMP, necessário
para a capacitação, por outro lado, o aumento de EROs causa um aumento no cAMP,
provavelmente por causa da estimulação da adenil ciclase, pois a atividade das
fosfodiesterases não é afetada (LEWIS; AITKEN, 2001; DE LAMIRANDE; O’FLAHERTY,
2008). Entretanto, os mecanismos através dos quais as EROs influenciam a reação
acrossômica e as fosforilações relacionadas ainda não foram totalmente elucidados. Todavia,
o real conhecimento na duração e na modulação através das quinases, sugere que os
mecanismos regulatórios da capacitação e da reação acrossômica são definitivamente
complexos e diferentes (DE LAMIRANDE; O’FLAHERTY, 2008).
As células espermáticas são particularmente susceptíveis ao estresse oxidativo (EROs)
porque elas contêm uma alta quantidade de ácidos graxos polinsaturados na membrana
plasmática, já as provenientes de manipulação laboratorial são ainda mais sensíveis
(DRUMMEN et al., 2002; BROUWERS et al., 2005; ARRUDA et al., 2006; SILVA et al.,
2007; HENDRICKS; HANSEN, 2009). Brouwers et al. (2005) observaram que a peroxidação
lipídica é mais proeminente na peça intermediária e em menor extensão na parte posterior da
cauda, mas virtualmente ausente na cabeça do espermatozóide.
EROs e espécies reativas de nitrogênio (RNS) têm efeitos lesivos, refletidos pela
peroxidação lipídica, prejuízo da função protéica e dano no DNA, comprometendo a
integridade das membranas dos espermatozóides e a viabilidade espermática (SILVA et al.,
2007).
A peroxidação lipídica foi, particularmente, forte na peça intermediária e na cauda de
espermatozóides congelados/descongelados e, significativamente, menos intenso na cabeça
espermática. Entretanto, a membrana plasmática do espermatozóide está significativamente
correlacionada com a capacidade de desenvolvimento do zigoto pós-fertilização. O estresse
oxidativo não induz um dano significativo no DNA das células espermáticas, a viabilidade
espermática e a membrana acrossomal (BROUWERS et al., 2005; SILVA et al., 2007).
O C11-BODIPY581/591 é uma sonda fluorescente para detectar peroxidação lipídica e
eficácia antioxidante em modelos de sistema de membrana e em células vivas, com excelentes

2 Revisão de Literatura 43
Priscilla Harumi Tanno
características. Possui emissão em um alcance visível, com bom espectro de separação da
forma não-oxidada (595 nm) com a oxidada (520 nm), tem um alto rendimento, necessita de
baixo volume do corante, boa fotoestabilidade, produz poucos artefatos fluorescentes, é
virtualmente insensível a alterações ambientais, como pH e polaridade do solvente, é
lipofílico, entrando facilmente nas membranas, e uma vez oxidado, o C11-BODIPY581/591
permanece lipofílico e não sai espontaneamente da bicamada lipídica, o C11-BODIPY581/591
localiza-se em dois lugares distintos dentro da bicamada lipídica, um é superficial a 18Ǻ e o
outro é profundo a 7,5Ǻ do centro para a bicamada, não é citotóxico, é sensível a vários oxi-
radicais e peroxinitritos, mas não é para o superóxido, óxido nítrico, íons de metais de
transição e hidroperóxidos (DRUMMEN et al., 2002).

3 Objetivos

3 Objetivos 45
Priscilla Harumi Tanno
3 OBJETIVOS
Levando-se em consideração a necessidade de informações mais precisas sobre os
efeitos da sexagem espermática em bovinos pela técnica de citometria de fluxo, os objetivos
desta pesquisa foram:
1. Avaliar os efeitos da sexagem sobre a integridade da membrana plasmática, reação
acrossomal, peroxidação lipídica e capacitação dos espermatozóides bovinos e entre
subespécies taurinas (Bos taurus taurus) e zebuínas (Bos taurus indicus),
2. Comparar possíveis diferenças entre o sêmen convencional e sexado quanto a
integridade da membrana plasmática, reação acrossomal, peroxidação lipídica e
capacitação dos espermatozóides bovinos e entre subespécies taurinas (Bos taurus
taurus) e zebuínas (Bos taurus indicus) e
3. Avaliar se o tempo de espera do ejaculado para a sexagem provoca alterações na
integridade da membrana plasmática, reação acrossomal, peroxidação lipídica e
capacitação dos espermatozóides bovinos e entre subespécies taurinas (Bos taurus
taurus) e zebuínas (Bos taurus indicus).

4 Hipóteses

4 Hipóteses 47
Priscilla Harumi Tanno
4 HIPÓTESES
1 A sexagem de sêmen pela técnica de citometria de fluxo provoca alterações na
integridade da membrana plasmática, reação acrossomal, peroxidação lipídica e
capacitação dos espermatozóides bovinos, e estas alterações são mais severas nas
subespécies taurinas (Bos taurus taurus) do que nas zebuínas (Bos taurus indicus).
2. Os espermatozóides bovinos submetidos à técnica de sexagem por citometria de
fluxo apresentam maior porcentagem de lesão da membrana plasmática, reação
acrossomal, peroxidação lipídica e capacitação do que os espermatozóides
submetidos à técnica convencional, sendo os espermatozóides da subespécie taurina
(Bos taurus taurus) mais sensíveis do que os da zebuína (Bos taurus indicus).
3. O tempo de espera do ejaculado para a sexagem provoca alterações na integridade
da membrana plasmática, reação acrossomal, peroxidação lipídica e capacitação
dos espermatozóides bovinos, sendo os da subespécie taurina (Bos taurus taurus)
mais sensíveis que os da zebuína (Bos taurus indicus).

5 Material e Métodos

5 Material e Métodos 49
Priscilla Harumi Tanno
5 MATERIAL E MÉTODOS
5.1 LOCAL
O presente trabalho foi realizado em duas etapas, sendo que a primeira consistiu na
sexagem e congelamento do sêmen no laboratório da Sexing Technologies do Brasil,
localizada na Central CRV Lagoa, na cidade de Sertãozinho, SP; e a segunda, referente às
análises laboratoriais, realizada no Laboratório de Biotecnologia do Sêmen e Andrologia do
Centro de Biotecnologia em Reprodução Animal do Departamento de Reprodução Animal da
Faculdade de Medicina Veterinária e Zootecnia da Universidade de São Paulo, localizado no
Campus Administrativo de Pirassununga, SP.
5.2 ANIMAIS E MANEJO
Foram utilizados seis touros em idade reprodutiva, sendo três taurinos da raça
Holandesa Preta e Branca (Bos taurus taurus) e três zebuínos da raça Gir Leiteira (Bos taurus
indicus), mantidos sob as mesmas condições ambientais, ou seja, em piquetes individuais com
água e sal mineral ad libitum e alimentação nutricional balanceada. Os animais foram pesados
quinzenalmente e realizados os controles de endo e ectoparasitas regularmente.
5.3 DELINEAMENTO EXPERIMENTAL
Baseado nos objetivos deste trabalho foi delineado um experimento, no qual foram
congeladas 12 palhetas de sêmen de 6 touros (3 taurinos e 3 zebuínos), quatro ejaculados por
touro e cinco tratamentos (12 x 6 x 4 x 5 = 1440 palhetas). Cada ejaculado foi dividido em
cinco alíquotas (cinco tratamentos) para produzir sêmen convencional (não sexado)
utilizando-se dois diluidores diferentes denominados de diluidor (L) Lagoa (T1) e outro com
o diluidor utilizado para a sexagem do sêmen pela técnica de citometria de fluxo, diluidor (S)
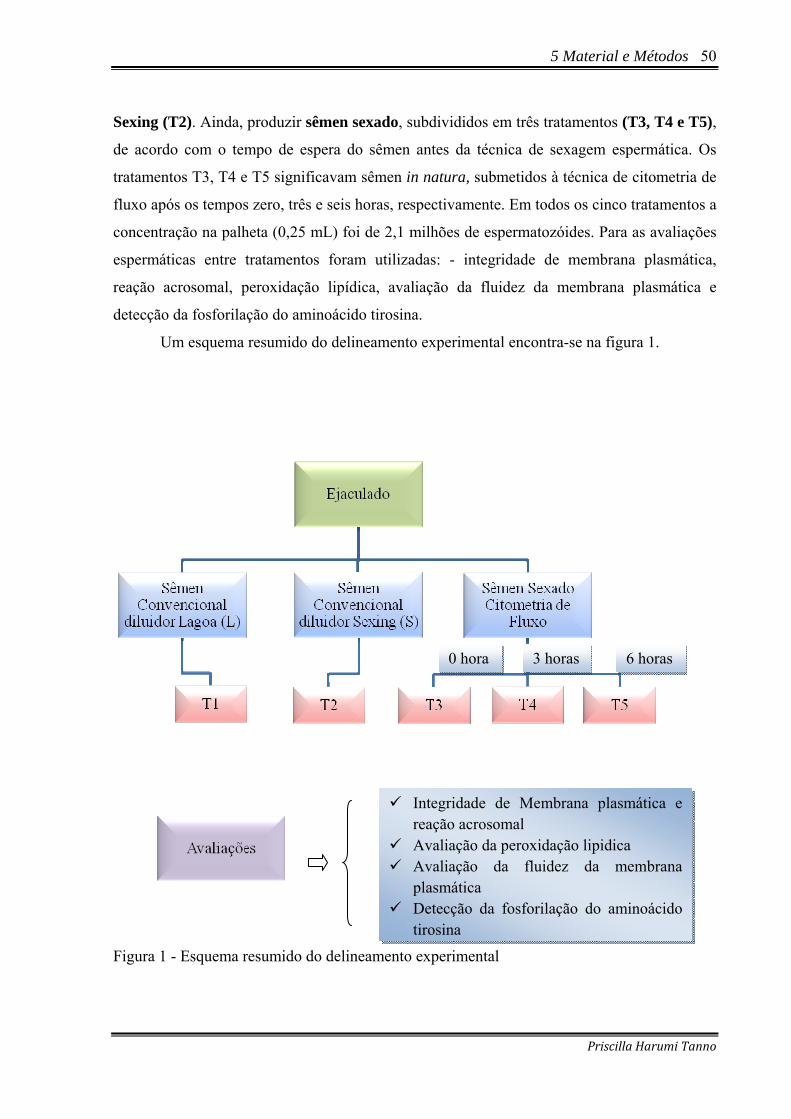
5 Material e Métodos 50
Priscilla Harumi Tanno
Sexing (T2). Ainda, produzir sêmen sexado, subdivididos em três tratamentos (T3, T4 e T5),
de acordo com o tempo de espera do sêmen antes da técnica de sexagem espermática. Os
tratamentos T3, T4 e T5 significavam sêmen in natura, submetidos à técnica de citometria de
fluxo após os tempos zero, três e seis horas, respectivamente. Em todos os cinco tratamentos a
concentração na palheta (0,25 mL) foi de 2,1 milhões de espermatozóides. Para as avaliações
espermáticas entre tratamentos foram utilizadas: - integridade de membrana plasmática,
reação acrosomal, peroxidação lipídica, avaliação da fluidez da membrana plasmática e
detecção da fosforilação do aminoácido tirosina.
Um esquema resumido do delineamento experimental encontra-se na figura 1.
Figura 1 - Esquema resumido do delineamento experimental
0 hora 3 horas 6 horas
Integridade de Membrana plasmática e reação acrosomal
Avaliação da peroxidação lipidica Avaliação da fluidez da membrana
plasmática Detecção da fosforilação do aminoácido
tirosina

5 Material e Métodos 51
Priscilla Harumi Tanno
5.4 PROCEDIMENTOS EXPERIMENTAIS
5.4.1 Colheita do sêmen
As colheitas de sêmen foram realizadas com vagina artificial, com auxílio de uma vaca
como manequim devidamente contida. Foram utilizados seis touros sendo, três taurinos da
raça Holandesa Preta e Branca (Bos taurus taurus) e três zebuínos da raça Gir Leiteira (Bos
taurus indicus). Foram realizadas quatro colheitas de sêmen de cada animal, com intervalos
semanais (6 touros × 4 ejaculados, n = 24).
Após a colheita, o ejaculado foi encaminhado rapidamente ao laboratório para análise.
5.4.2 Avaliações do sêmen in natura
O ejaculado foi analisado quanto ao volume, concentração espermática, motilidade,
vigor e alterações morfológicas. Todo material utilizado na colheita de sêmen e avaliação do
sêmen foi mantido aquecido, na temperatura à 37ºC para evitar o choque térmico e alterações
das características seminais.
5.4.2.1 Volume
O volume foi determinado pela leitura direta do tubo de 15 mL graduado com fração
de 0,1 mL.
5.4.2.2 Concentração espermática
A concentração espermática foi determinada por um contador nuclear denominado
NucleoCounter® SP-100™ System com software versão 1.21 (CM part no. 900-0100) –

5 Material e Métodos 52
Priscilla Harumi Tanno
Chemometec A/S DK – 3450 Allerod, Dinamarca), calibrado para o programa DF 401, para o
qual diluiu-se 25 µL de sêmen em 10 mL de reagente S100 (CM part no. 910-0100),
responsável pela lise da membrana celular, em um tubo de 15 mL.
Em seguida, foi homogeneizado em um vortex durante 12 segundos,
aproximadamente, e uma alíquota foi separada para a leitura em um SP1 - Cassette™ (CM
part no. 941-0006) - Chemometec. O SP1 - Cassete™ contendo iodeto de propídio (PI), que
identifica as células lesadas pelo reagente S100 e realiza a contagem nuclear celular (Figura
2).
Figura 2 - Esquema simplificado para a obtenção da concentração espermática
5.4.2.3 Motilidade, vigor e turbilhonamento
Uma alíquota de 10 µL de sêmen in natura foi diluída em 500 µL de diluidor à base de
Tris e gema de ovo pré-aquecido (37º C) para a avaliação da motilidade. A motilidade foi
avaliada em uma gota de 10 µL da alíquota diluída colocada entre lâmina e lamínula e
25 µLde sêmen 10 mL de S100
VORTEX (12 s)
Cassete
Nucleo Counter
DF 401 Concentração espermática (106 / mL)
Ejaculado

5 Material e Métodos 53
Priscilla Harumi Tanno
visualizada em aumento de 100 x sob microscopia de contraste de interferência diferencial
(DIC).
O vigor, referente à velocidade progressiva uniforme das células em movimento foi
classificado em escores de 1 a 5, sendo o escore 1 o mais lento e aumentando gradativamente
até o escore 5, correspondendo ao mais veloz movimento progressivo e uniforme. Foi
observado em uma gota de 10 µL da alíquota diluída entre lâmina e lamínula em aumento de
100 x em DIC.
Para avaliação do turbilhonamento, uma amostra de 10 µL de sêmen fresco foi
depositada em lâmina pré-aquecida a 37oC e a análise foi efetivada sem lamínula, sob
microscopia de contraste de interferência diferencial (DIC) em aumento de 100 x. O
movimento de massa das células foi classificado em escala que variou de 0 (movimento
ausente) a 5 (máximo).
5.4.2.4 Análise da morfologia espermática
Para a avaliação das alterações morfológicas, foram adicionados 10 µL de formol
salino tamponado no tubo pré-diluído para a avaliação prévia da motilidade. Em seguida, foi
realizada a leitura da morfologia em preparação úmida com aumento de 1000 x sob
microscopia de contraste de interferência diferencial (DIC) (OLYMPUS AX70, Olympus
True Reserch System Microscoper model AX70, Melville, NY), contando 100 células por
amostra. As porcentagens das diversas alterações morfológicas foram agrupadas e
classificadas em defeitos maiores e defeitos menores.
Somente ejaculados com concentração igual ou superior a 1000 x 106 espermatozóides
por mililitros, 60% de motilidade progressiva, vigor maior do que o escore 3 e menos de 20%
de defeitos totais foram utilizados no experimento.

5 Material e Métodos 54
Priscilla Harumi Tanno
5.4.3 Congelação do sêmen pela técnica convencional
Após as avaliações do sêmen in natura, duas alíquotas do ejaculado foram retiradas
para o preparo do sêmen convencional (não sexado) utilizando-se dois tipos de diluidores
denominados de diluidor (L) Lagoa (à base de TRIS-gema de ovo) (T1) e outro com o
mesmo diluidor utilizado para a sexagem do sêmen pela técnica de citometria de fluxo,
diluidor (S) Sexing (à base de TRIS-gema de ovo centrifugado®) (T2).
Logo após a diluição, os tubos contendo sêmen convencional, previamente
identificados, permaneceram em banho-maria a 34°C por uma hora e 30 minutos. Em seguida,
foram encaminhados para a câmara-fria a 5°C por aproximadamente quatro horas, protegidos
da luz para aguardar o envase e congelação.
Foram envasadas 12 palhetas por tratamento, as quais foram cuidadosamente
identificadas com o nome do touro, subespécie, partida do ejaculado e tratamento. A
quantidade de espermatozóides por palheta (0,25 mL) foi de 2,1 milhões.
As palhetas foram congeladas em caixa de isopor contendo 7 cm de altura de
nitrogênio líquido a uma distância de 4 cm das palhetas durante 20 minutos, sob vapor do
nitrogênio líquido. Em seguida, as palhetas foram retiradas e imersas em nitrogênio líquido (-
196ºC). Imediatamente após, as palhetas foram raqueadas e armazenadas em botijões
criogênicos para posterior análise.
5.4.4 Sexagem espermática pela técnica de citometria de fluxo
Após as avaliações do sêmen in natura, três alíquotas do ejaculado foram preparadas
para produzir o sêmen sexado, sendo que cada alíquota foi retirada nos tempos zero, três e
seis horas, respectivamente (T3, T4 e T5) de acordo com o tempo de espera do sêmen.

5 Material e Métodos 55
Priscilla Harumi Tanno
5.4.4.1 Diluição e incorporação do Hoechst 33342
Para a sexagem dos espermatozóides uma solução estoque de Hoechst 33342 foi
preparada a 8,89 mM (5 mg/mL) com água nanopura, corante cuja função foi penetrar na
membrana celular e ligar-se a regiões específicas do DNA, que são os 3 pares de bases ricos
em adenina-timina das menores depressões da hélice de DNA (GARNER, 2009). O sêmen foi
diluído nos tempos zero, três e seis horas na concentração de 200 x 106/mL, em TALP
modificado (meio Tyrode com albumina, lactato e piruvato) em pH de 7,4 visando alcançar
maior eficiência do corante Hoechst 33342. Após a amostra foi incubada a 34ºC durante 45
minutos protegida dos efeitos da luz.
Em seguida, foi adicionado TALP contendo 4% de gema de ovo clarificada e 0,002%
de “food colouring dye” (FD&C#40 Power, Waner Jenkinson Company Inc., St Louis, MO,
USA), que teve como objetivo diminuir o pH, prevenindo dano celular e, também, apagar o
H33342 dos espermatozóides com as membranas celulares comprometidas (SCHENK et al.,
1999; GARNER, 2001; GARNER, 2009), permitindo o descarte destas células no momento
da separação no citômetro de fluxo.
Cada amostra foi filtrada à unidade gravitacional em um filtro de nylon com poros de
50 µm (CellTrics™, Partec GmbH, Germany) para a remoção de debris ou espermatozóides
aglutinados, formando uma solução final de 4 mL com uma concentração de 80 x 106
espermatozóides/mL e mantidas em temperatura à 21ºC durante a separação espermática no
citômetro de fluxo.
O citômetro de fluxo usado no experimento foi o SX MOFLO®, operando a 40 psi
(Figura 3). A fluorescência que foi emitida pelas células quando excitadas pelo laser
Vanguard TM, sistema de estado sólido de diode-pumped (Newport Corporation, Irvine, CA,
USA) passou por dois detectores de fluorescência (PMT), localizados a 0° e a 90°, um com a
função de orientar corretamente as células e, outro para medir o valor da fluorescência.
Dependendo da onda emitida, foi dada uma carga elétrica na célula, ou seja, o espermatozóide
contendo o cromossomo X recebeu uma carga negativa, sendo atraído pelo campo positivo e o
oposto para os espermatozóides contendo o cromossomo Y.
Um diluidor (Sheath Fluid) a base de TRIS (hidroximetil aminometano - TRIS 197,0
Mm), ácido cítrico monohidratado (55,4 mM) e frutose (47,5 mM) foi utilizado para formar o
fluxo hidrodinâmico onde as células espermáticas percorreram.

5 Material e Métodos 56
Priscilla Harumi Tanno
As células separadas pelo citômetro de fluxo (SX MOFLO®) foram coletadas em tubos
cônicos graduados para centrífuga de 50mL, contendo diluidor sem glicerol, denominado de
catch (meio TRIS - 2% gema de ovo – Sexing Technologies, Navasota, USA) que foi diluído
com 16% de água, até atingir o volume de 20 mL. A concentração final de um tubo foi de
aproximadamente 8,0 x 105 espermatozóides/mL, como conseqüência da mistura dos
diluidores durante a separação, catch e o sheath fluid.
Em seguida, foram colocados sob refrigeração a 5ºC por um período de 90 minutos.
Figura 3 - Aparelho de Citometria de Fluxo SX MOFLO®
5.4.5 Criopreservação do sêmen sexado
Após a refrigeração, foi adicionado ao sêmen, diluidor TRIS contendo 12% de glicerol
em duas etapas, com intervalo de 15 minutos (9,5 mL cada). O sêmen foi levado à centrífuga
refrigerada à 5º C, SORVALL® RC-4 (Heareus Sorvall Revco, Asheville, NC), a 850 x g
durante 20 minutos.

5 Material e Métodos 57
Priscilla Harumi Tanno
O sobrenadante foi desprezado e o sedimento formado foi homogeneizado e colocado
em um único tubo, com uma concentração aproximada de 40 x 106 espermatozóides/mL. Em
seguida, foi retirada uma alíquota de 25 µL para a determinação da concentração através do
NucleoCounter® SP-100™ system AN-103 (ChemoMetec A/S Gydevang 43, DK-3450
Allerod, Dinamarca), visando a adição do restante do diluidor TRIS-gema até atingir
concentração final de 9,4 x 106 de espermatozóides/mL (considera-se em torno de um milhão
de espermatozóides perdidos durante o envase).
Logo após a diluição com TRIS-gema de ovo, o sêmen permaneceu protegido da luz à
temperatura de 5ºC durante 3 horas (tempo de equilíbrio) para então, ser envasado em
palhetas de 0,25 mL (IMV®, Aigle, France). As palhetas foram congeladas em caixa de isopor
contendo 7 cm de altura de nitrogênio a uma distância de 4 cm das palhetas durante 20
minutos, sob vapor do nitrogênio liquído. Em seguida, as palhetas foram retiradas e imersas
em nitrogênio líquido (-196ºC). Imediatamente, as palhetas foram raqueadas e armazenadas
em botijões criogênicos para posterior análise.
5.4.6 Avaliações do sêmen convencional (dois diluidores) e do sexado (três diferentes
tempos)
Para a avaliação do sêmen três palhetas do mesmo animal, partida e tratamento foram
descongeladas a 37oC por 30 segundos. Todas as técnicas que serão descritas a seguir foram
realizadas segundo metodologias citadas por Andrade (2009).
5.4.6.1 Avaliação da integridade da membrana plasmática e reação acrossomal por citometria
de fluxo
Uma alíquota foi retirada das amostras e adicionada ao TALP sperm (BAVISTER;
LEIBFRIED; LIEBERMAN, 1983) modificado (TALPm, Anexo A) em tubos de
microcentrifugação (1,5 mL) pré-aquecidos a 37°C, a fim de se obter amostras com
concentração de 5 x 106 espermatozóides por mL em um volume final de 148 µL. A seguir,
adicionou-se 2 µL de Hoechst 33342 (H33342; 40 µg/mL, H-1399, Molecular Probes Inc.,

5 Material e Métodos 58
Priscilla Harumi Tanno
Eugene, Oregon, EUA) com posterior incubação por 10 minutos a 37°C. O uso desta sonda
teve como objetivo, corar o DNA das células espermáticas para que não houvesse a contagem
de forma equivocada de partículas com o mesmo tamanho e granulosidade do
espermatozóide. Passado o período de incubação, foram adicionados a amostra 3 µL de Iodeto
de Propídio (PI-0,5 mg/mL, 28,707-5, Sigma-Aldrich Co., Saint Louis, Missouri, EUA),
conjuntamente com a adição de 10 µL de aglutinina de Pisum sativum conjugada ao
isotiocionato de fluoresceína (FITC-PSA-100 µg/mL, L-0770, Sigma-Aldrich Co., Saint
Louis, Missouri, EUA) estas sondas foram adicionadas com os seguintes objetivos: corar as
células com membrana plasmática lesada (PI positivo) (CELEGHINI et al., 2007; ANDRADE
et al., 2007) e com a membrana acrossomal reagida (FITC-PSA positivo). Após 10 minutos de
incubação a 37°C, os espermatozóides foram diluídos com a adição de 150 µL de TALPm e
transferidos para tubos graduados até 15 mL (37°C). Com isso, as amostras apresentavam
uma concentração de 2,5 x 106 espermatozóides por mL no momento de serem analisadas pela
técnica de citometria de fluxo. Todas as amostras tiveram 10.000 células analisadas. Na figura
4 está apresentado um exemplo de gráfico de pontos gerados pelo software BD FACSDiva 6.0
após a contagem da amostra. A coloração com FITC-PSA diferenciou a população
espermática viável (IP-negativo) em dois grupos distintos (Figura 4). Os espermatozóides que
não se apresentavam corados com FITC-PSA, estes sendo os que possuíam a membrana
acrossomal intacta (AIMPI), ou os positivos para a coloração com FITC-PSA indicando que
estes espermatozóides ou possuíam o acrossomo em processo de exocitose ou já estavam
reagidos (ARMPI).

5 Material e Métodos 59
Priscilla Harumi Tanno
Figura
4 - Gráfico de pontos gerado pela análise de 10.000 células por citometria de fluxo. Amostra corada com a associação H33342, PI, FITC-PSA permitindo a classificação dos espermatozóides bovinos em quatro categorias – Pirassununga – 2009
5.4.6.2 Avaliação da peroxidação das membranas espermáticas por citometria de fluxo
Foram retiradas alíquotas de sêmen durante as etapas de análises a fim de obter
amostras diluídas em TALPm com 5 x 106 espermatozóides/mL e com um volume final de
499,5 µL. Estas foram então coradas com 0,5 µL da sonda C11-BODIPY581/591(1 mg/mL, D-
3861, Molecular Probes Inc., Eugene, Oregon, EUA) por 30 minutos a 37°C. Passado o
período de incubação, foi transferida para outro microtubo 145 µL desta solução, agora,
corada com a sonda C11-BODIPY581/591. A esta amostra foi adicionado 2 µL de Hoechst
33342 (40 µg/mL, H-1399, Molecular Probes Inc., Eugene, Oregon, EUA) com posterior
incubação por 10 minutos a 37°C. O uso desta sonda teve como objetivo, corar o DNA das
células espermáticas para que não houvesse a contagem de forma equivocada de partículas
com o mesmo tamanho e granulosidade do espermatozóide. Então, adicionou-se a amostra
3 µL de Iodeto de Propídio (0,5 mg/mL, 28,707-5, Sigma-Aldrich Co., Saint Louis, Missouri,
EUA) a fim de servir como um marcador das células com membrana plasmática lesada. Após
AIML-membrana plasmática lesada e membrana acrossomal não reagida. ARML-membrana plasmática lesada e membrana acrossomal reagida. AIMI-membrana plasmática íntegra e membrana acrossomal não reagida. ARMI-membrana plasmática íntegra e membrana acrossomal reagida.
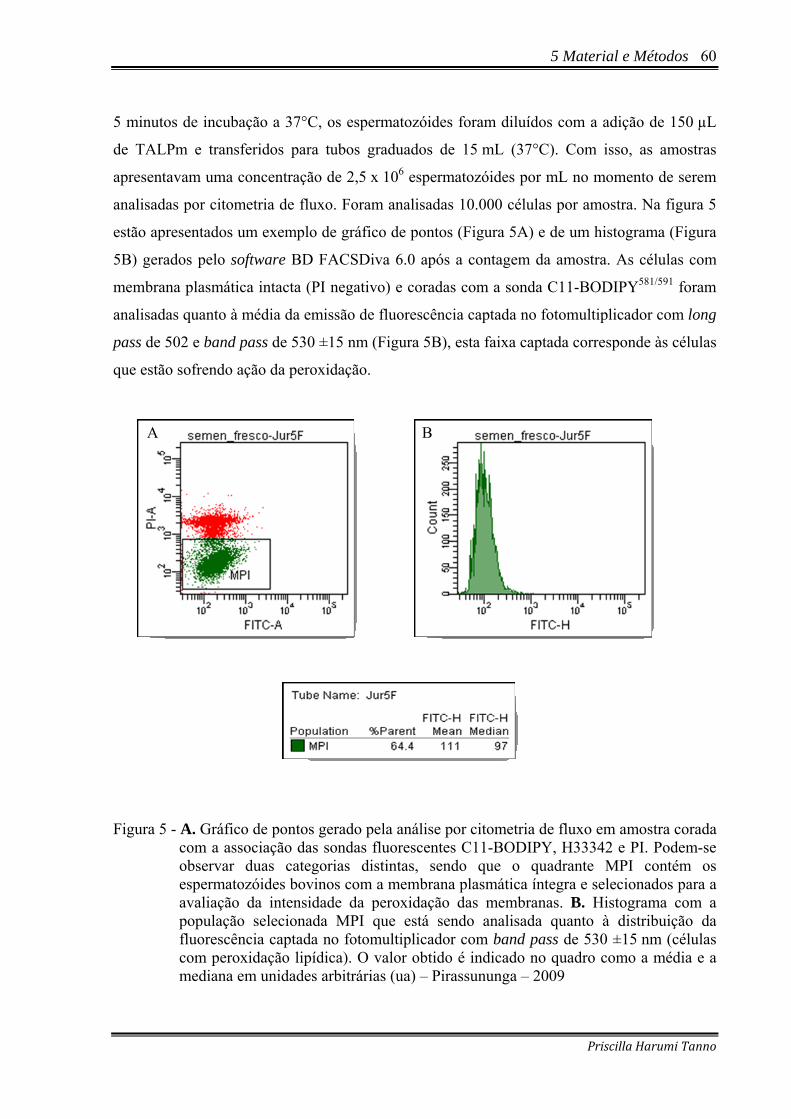
5 Material e Métodos 60
Priscilla Harumi Tanno
5 minutos de incubação a 37°C, os espermatozóides foram diluídos com a adição de 150 µL
de TALPm e transferidos para tubos graduados de 15 mL (37°C). Com isso, as amostras
apresentavam uma concentração de 2,5 x 106 espermatozóides por mL no momento de serem
analisadas por citometria de fluxo. Foram analisadas 10.000 células por amostra. Na figura 5
estão apresentados um exemplo de gráfico de pontos (Figura 5A) e de um histograma (Figura
5B) gerados pelo software BD FACSDiva 6.0 após a contagem da amostra. As células com
membrana plasmática intacta (PI negativo) e coradas com a sonda C11-BODIPY581/591 foram
analisadas quanto à média da emissão de fluorescência captada no fotomultiplicador com long
pass de 502 e band pass de 530 ±15 nm (Figura 5B), esta faixa captada corresponde às células
que estão sofrendo ação da peroxidação.
Figura 5 - A. Gráfico de pontos gerado pela análise por citometria de fluxo em amostra corada com a associação das sondas fluorescentes C11-BODIPY, H33342 e PI. Podem-se observar duas categorias distintas, sendo que o quadrante MPI contém os espermatozóides bovinos com a membrana plasmática íntegra e selecionados para a avaliação da intensidade da peroxidação das membranas. B. Histograma com a população selecionada MPI que está sendo analisada quanto à distribuição da fluorescência captada no fotomultiplicador com band pass de 530 ±15 nm (células com peroxidação lipídica). O valor obtido é indicado no quadro como a média e a mediana em unidades arbitrárias (ua) – Pirassununga – 2009
B A

5 Material e Métodos 61
Priscilla Harumi Tanno
5.4.6.3 Avaliação da estabilidade (fluidez) da membrana plasmática por citometria de fluxo
Para a análise da estabilidade da membrana plasmática pela técnica de citometria de
fluxo as amostras foram incubadas em solução de TALPm na concentração de
5 x 106 espermatozóides/mL em um volume final de 147 µL. Foram então adicionados 2 µL
de Hoechst 33342 (40 µg/mL, H-1399, Molecular Probes Inc., Eugene, Oregon, EUA) com
posterior incubação por 10 minutos a 37°C. O uso desta sonda teve como objetivo, corar o
DNA das células espermáticas para que não houvesse a contagem de forma equivocada de
partículas com o mesmo tamanho e granulosidade do espermatozóide. Em seguida 0,5 µL da
sonda fluorescente Yo-Pro-1 foi adicionado a amostra (7,5 µM-Molecular Probes Inc.,
Eugene, Oregon, EUA) obtendo-se uma concentração final de 25 nM. A membrana
plasmática íntegra é impermeável ao Yo-Pro-1, o qual se liga ao DNA das células com a
membrana plasmática lesada. Então, a amostra foi incubada por 20 minutos para
posteriormente ser adicionada da sonda fluorescente merocianina 540 (810 µM-Molecular
Probes Inc., Eugene, Oregon, EUA) a fim de se obter uma concentração de 2,7 µM em uma
solução final de 150 µL, em seguida estas foram incubadas por 70 segundos. Foram então
diluídas em 150 µL de TALPm e analisadas por citometria de fluxo, no qual foi avaliado as
porcentagens de espermatozóides com membrana plasmática íntegra (Yo-Pro-1 negativo) com
aumento na desordem da bicamada lipídica (Merocianina 540 positivo). Na figura 6 está
apresentado um exemplo de gráfico de pontos gerados pelo software BD FACSDiva 6.0 após
a contagem da amostra. A coloração com Merocianina 540 mostrou duas populações distintas
de células viáveis (Yo-Pro-1 negativa), uma que é caracterizada pelas células com emissão de
baixa fluorescência e outra que é caracterizada por alta (captado no long pass-595 e band-
pass-610/20). Os espermatozóides com baixa emissão de fluorescência são considerados não
capacitados, enquanto que as células que apresentam alta emissão de fluorescência são
caracterizadas como capacitadas (alto grau de desorganização da bicamada lipídica; Figura 6).

5 Material e Métodos 62
Priscilla Harumi Tanno
Figura 6 - Gráfico de pontos gerado pela análise por citometria de fluxo, neste caso as células foram coradas com a associação do H33342, Yo-Pro-1 e da Merocianina 540 permitindo a classificação dos espermatozóides bovinos em três categorias – Pirassununga – 2009
5.4.6.4 Detecção por citometria de fluxo da fosforilação do aminoácido tirosina
As amostras de sêmen foram diluídas em TALPm a fim de se obter uma concentração
final de 1 x 106 espermatozóides/mL em um volume final de 141 µL. Foram então
adicionados 2 µL de Hoechst 33342 (40 µg/mL, H-1399, Molecular Probes Inc., Eugene,
Oregon, EUA) com posterior incubação por 10 minutos a 37°C. Em seguida, foi adicionado
6 µL do anticorpo antifosfotirosina conjugado a uma fluoresceína (100 µg/mL-CLONE PY-
20, F0426, Sigma-Aldrich Co., Saint Louis, Missouri, EUA) a esta suspensão, obtendo-se
uma concentração final de 2 µg/mL de anticorpo. Neste mesmo momento foi adicionado à
amostra 3 µL de Iodeto de Propídio (0,5 mg/mL, 28,707-5, Sigma-Aldrich Co., Saint Louis,
Missouri, EUA) a fim de retirar da análise as células que apresentavam lesão na membrana
plasmática. Em seguida a amostra foi incubada por 5 minutos a 37°C para então ser submetida
à análise por citometria de fluxo (ANDRADE et al., 2008). A figura 7 é constituída por um
exemplo de gráfico de pontos (Figura 7A) e de um histograma (Figura 7B) gerados pelo
software BD FACSDiva 6.0 após a contagem da amostra. As células com membrana
NCMPI- espermatozóide não capacitado com membrana plasmática íntegra. MPL - espermatozóide com membrana plasmática lesada. CAPAC- espermatozóide capacitado e com membrana plamática íntegra.
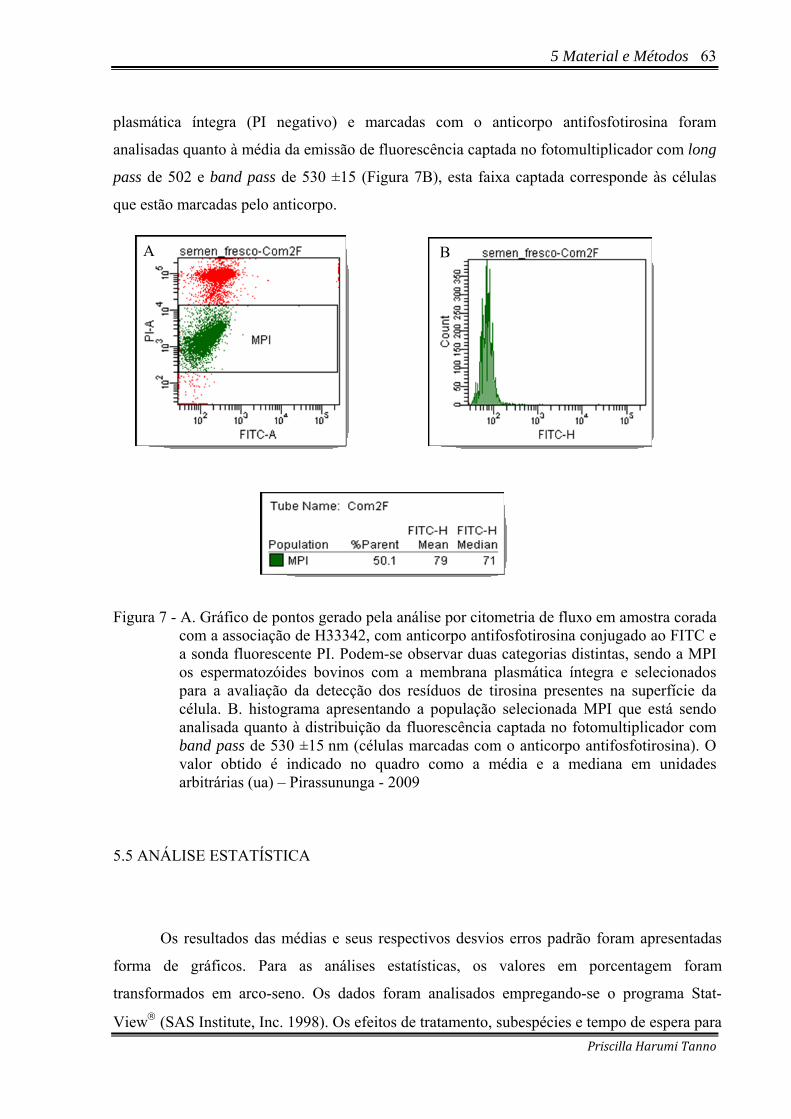
5 Material e Métodos 63
Priscilla Harumi Tanno
plasmática íntegra (PI negativo) e marcadas com o anticorpo antifosfotirosina foram
analisadas quanto à média da emissão de fluorescência captada no fotomultiplicador com long
pass de 502 e band pass de 530 ±15 (Figura 7B), esta faixa captada corresponde às células
que estão marcadas pelo anticorpo.
Figura 7 - A. Gráfico de pontos gerado pela análise por citometria de fluxo em amostra corada com a associação de H33342, com anticorpo antifosfotirosina conjugado ao FITC e a sonda fluorescente PI. Podem-se observar duas categorias distintas, sendo a MPI os espermatozóides bovinos com a membrana plasmática íntegra e selecionados para a avaliação da detecção dos resíduos de tirosina presentes na superfície da célula. B. histograma apresentando a população selecionada MPI que está sendo analisada quanto à distribuição da fluorescência captada no fotomultiplicador com band pass de 530 ±15 nm (células marcadas com o anticorpo antifosfotirosina). O valor obtido é indicado no quadro como a média e a mediana em unidades arbitrárias (ua) – Pirassununga - 2009
5.5 ANÁLISE ESTATÍSTICA
Os resultados das médias e seus respectivos desvios erros padrão foram apresentadas
forma de gráficos. Para as análises estatísticas, os valores em porcentagem foram
transformados em arco-seno. Os dados foram analisados empregando-se o programa Stat-
View® (SAS Institute, Inc. 1998). Os efeitos de tratamento, subespécies e tempo de espera para
A B

5 Material e Métodos 64
Priscilla Harumi Tanno
a sexagem espermática, bem como suas respectivas interações, foram avaliados por meio de
análises de variância (ANOVA). Quando o principal efeito foi significativo, as médias foram
comparadas individualmente pelo teste de Fisher. A hipótese testada foi considerada
significativa quando P<0,05 e com tendência a significância quando P >0,05 e <0,10.

6 Resultados

6 Resultados 66
Priscilla Harumi Tanno
6 RESULTADOS
Nesta seção encontram-se descritos os resultados obtidos no presente experimento. 6.1 INTEGRIDADE DA MEMBRANA PLASMÁTICA E REAÇÃO ACROSSOMAL EM
SÊMEN CONVENCIONAL E SEXADO
Após a descongelação das amostras, foi observado que houve tendência a significância
(P<0,10) da quantidade de espermatozóides sem reação acrossômica e com membrana
plasmática íntegra entre o T1 (sêmen convencional com diluidor L) e T3 (sêmen sexado no
tempo zero), porém não foi encontrada diferença entre os grupos T1, T2 (sêmen convencional
com diluidor S), T4 (sêmen sexado no tempo três horas após) e T5 (sêmen sexado no tempo
seis horas após) e entre os tratamentos T2, T3, T4 e T5, no sêmen bovino pós-descongelação,
submetido ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) (Gráfico
1).
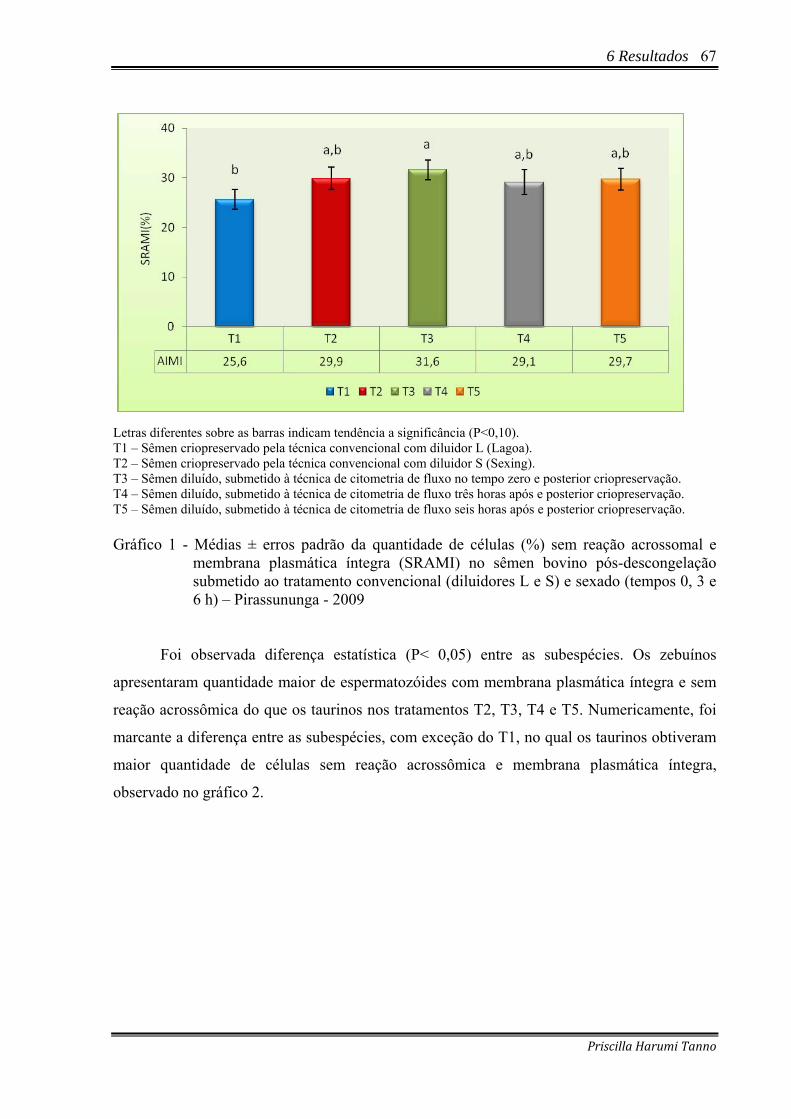
6 Resultados 67
Priscilla Harumi Tanno
Letras diferentes sobre as barras indicam tendência a significância (P<0,10). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído, submetido à técnica de citometria de fluxo no tempo zero e posterior criopreservação. T4 – Sêmen diluído, submetido à técnica de citometria de fluxo três horas após e posterior criopreservação. T5 – Sêmen diluído, submetido à técnica de citometria de fluxo seis horas após e posterior criopreservação. Gráfico 1 - Médias ± erros padrão da quantidade de células (%) sem reação acrossomal e
membrana plasmática íntegra (SRAMI) no sêmen bovino pós-descongelação submetido ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga - 2009
Foi observada diferença estatística (P< 0,05) entre as subespécies. Os zebuínos
apresentaram quantidade maior de espermatozóides com membrana plasmática íntegra e sem
reação acrossômica do que os taurinos nos tratamentos T2, T3, T4 e T5. Numericamente, foi
marcante a diferença entre as subespécies, com exceção do T1, no qual os taurinos obtiveram
maior quantidade de células sem reação acrossômica e membrana plasmática íntegra,
observado no gráfico 2.

6 Resultados 68
Priscilla Harumi Tanno
Letras diferentes sobre barras significam diferenças estatísticas entre as subespécies(P< 0,05). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído, submetido à técnica de citometria de fluxo no tempo zero e posterior criopreservação. T4 – Sêmen diluído, submetido à técnica de citometria de fluxo três horas após e posterior criopreservação. T5 – Sêmen diluído, submetido à técnica de citometria de fluxo seis horas após e posterior criopreservação. Gráfico 2 - Médias ± erros padrão da quantidade de células (%) sem reação acrossomal e
membrana plasmática íntegra (SRAMI) no sêmen de taurinos e zebuínos pós-descongelação submetidos ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga - 2009
Foi observada uma pequena população espermática com membrana plasmática íntegra
e reação acrossômica que variou de 0,3 a 1,0%. Embora os diluidores do sêmen convencional
tenham sido diferentes (T1 e T2) não houve diferença entre eles, no entanto, quando
comparado ao sêmen sexado (T3, T4, T5), houve diferença significativa (P< 0,05). O sêmen
sexado mostrou reação acrossômica e membrana plasmática íntegra menor do que o
convencional. Já entre os tratamentos do sêmen sexado, não houve diferença ao longo do
tempo (zero, três e seis horas após), presente no gráfico 3.

6 Resultados 69
Priscilla Harumi Tanno
Letras diferentes sobre as barras indicam diferença estatística (P< 0,05). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído, submetido à técnica de citometria de fluxo no tempo zero e posterior criopreservação. T4 – Sêmen diluído, submetido à técnica de citometria de fluxo três horas após e posterior criopreservação. T5 – Sêmen diluído, submetido à técnica de citometria de fluxo seis horas após e posterior criopreservação.
Gráfico 3 - Médias ± erros padrão da quantidade de células (%) com reação acrossomal e membrana plasmática íntegra (CRAMI) no sêmen bovino pós-descongelação submetido ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga - 2009
A porcentagem de células com reação acrossomal e membrana plasmática íntegra não
variaram entre taurinos e zebuínos (P>0,05). Provavelmente, a diferença estatística não foi
encontrada, devido à maior variação entre as médias, caracterizado pelo maior erro padrão
(Gráfico 4).
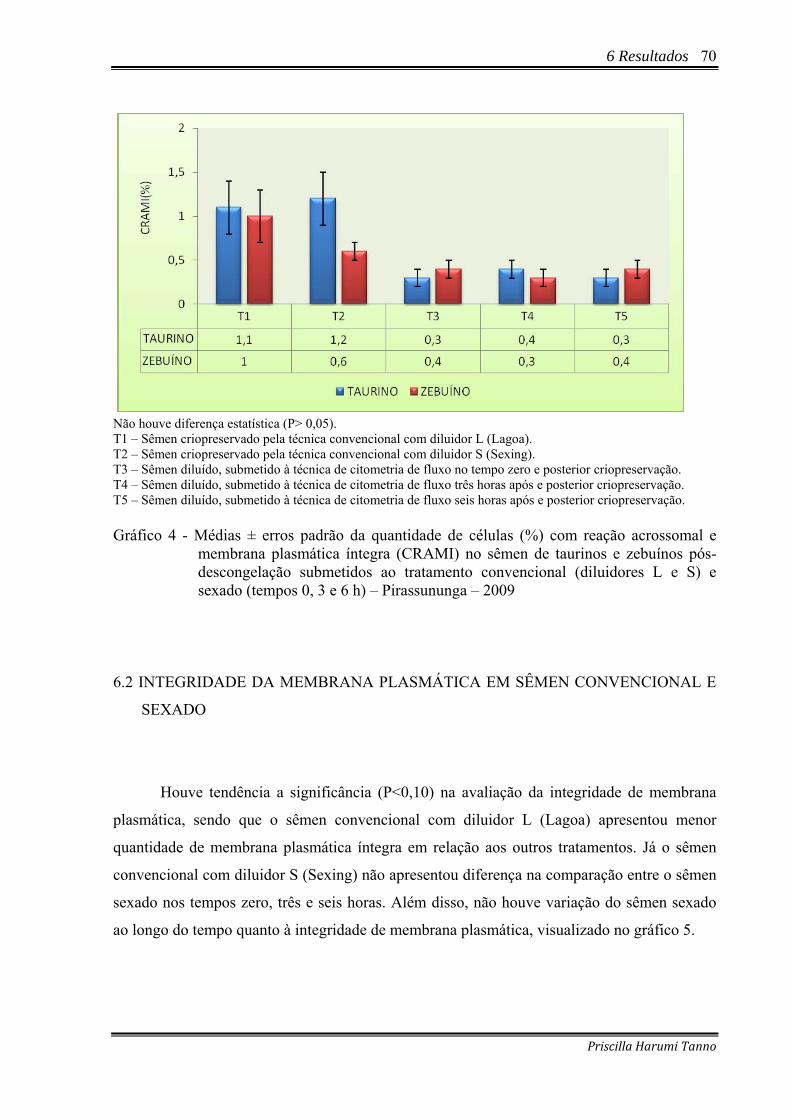
6 Resultados 70
Priscilla Harumi Tanno
Não houve diferença estatística (P> 0,05). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído, submetido à técnica de citometria de fluxo no tempo zero e posterior criopreservação. T4 – Sêmen diluído, submetido à técnica de citometria de fluxo três horas após e posterior criopreservação. T5 – Sêmen diluído, submetido à técnica de citometria de fluxo seis horas após e posterior criopreservação. Gráfico 4 - Médias ± erros padrão da quantidade de células (%) com reação acrossomal e
membrana plasmática íntegra (CRAMI) no sêmen de taurinos e zebuínos pós-descongelação submetidos ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga – 2009
6.2 INTEGRIDADE DA MEMBRANA PLASMÁTICA EM SÊMEN CONVENCIONAL E
SEXADO
Houve tendência a significância (P<0,10) na avaliação da integridade de membrana
plasmática, sendo que o sêmen convencional com diluidor L (Lagoa) apresentou menor
quantidade de membrana plasmática íntegra em relação aos outros tratamentos. Já o sêmen
convencional com diluidor S (Sexing) não apresentou diferença na comparação entre o sêmen
sexado nos tempos zero, três e seis horas. Além disso, não houve variação do sêmen sexado
ao longo do tempo quanto à integridade de membrana plasmática, visualizado no gráfico 5.
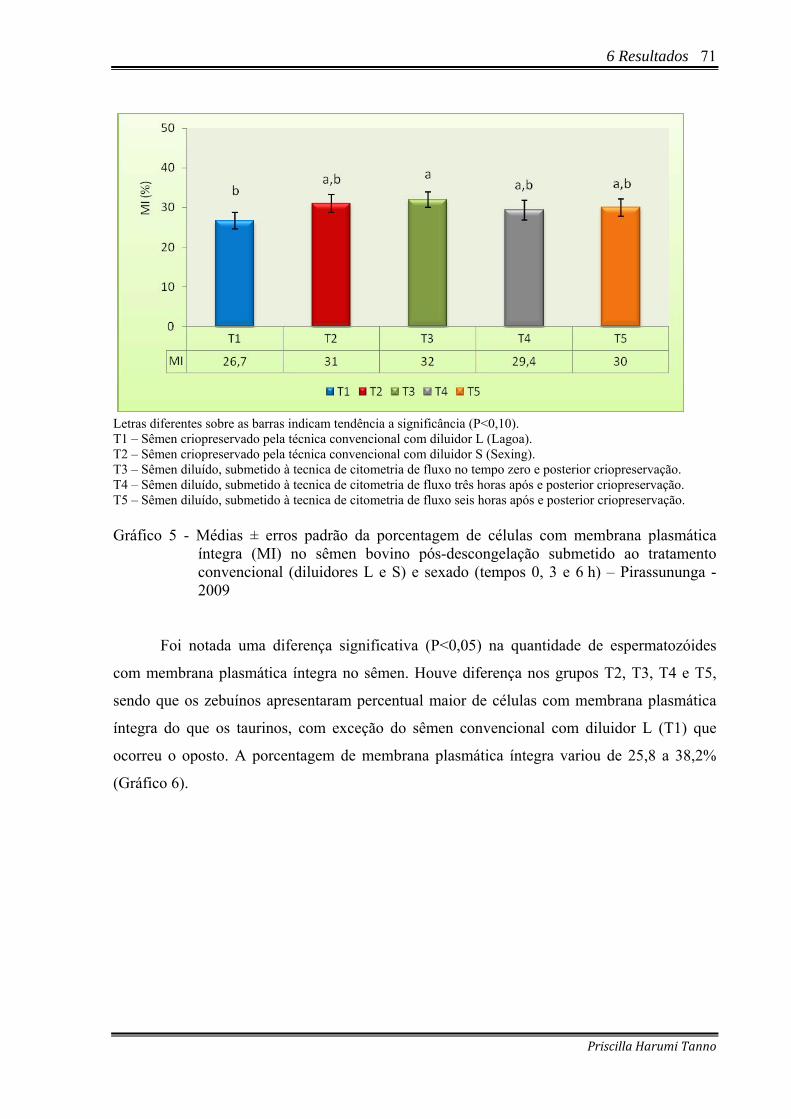
6 Resultados 71
Priscilla Harumi Tanno
Letras diferentes sobre as barras indicam tendência a significância (P<0,10). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído, submetido à tecnica de citometria de fluxo no tempo zero e posterior criopreservação. T4 – Sêmen diluído, submetido à tecnica de citometria de fluxo três horas após e posterior criopreservação. T5 – Sêmen diluído, submetido à tecnica de citometria de fluxo seis horas após e posterior criopreservação. Gráfico 5 - Médias ± erros padrão da porcentagem de células com membrana plasmática
íntegra (MI) no sêmen bovino pós-descongelação submetido ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga - 2009
Foi notada uma diferença significativa (P<0,05) na quantidade de espermatozóides
com membrana plasmática íntegra no sêmen. Houve diferença nos grupos T2, T3, T4 e T5,
sendo que os zebuínos apresentaram percentual maior de células com membrana plasmática
íntegra do que os taurinos, com exceção do sêmen convencional com diluidor L (T1) que
ocorreu o oposto. A porcentagem de membrana plasmática íntegra variou de 25,8 a 38,2%
(Gráfico 6).

6 Resultados 72
Priscilla Harumi Tanno
Letras diferentes sobre barras significam diferenças estatísticas entre as subespécies(P< 0,05). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído e submetido à tecnica de citometria de fluxo no tempo zero. T4 – Sêmen diluído e submetido à tecnica de citometria de fluxo após três horas. T5 – Sêmen diluído e submetido à tecnica de citometria de fluxo após seis horas Gráfico 6 - Médias ± erros padrão da quantidade de células (%) com membrana plasmática
íntegra (MI) no sêmen de taurinos e zebuínos pós-descongelação submetidos ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga – 2009
6.3 REAÇÃO ACROSSÔMICA EM SÊMEN CONVENCIONAL E SEXADO
Houve tendência a significância (P<0,10) na avaliação da reação acrossômica. O
sêmen sexado no tempo zero (T3) e no tempo seis horas após (T5) apresentaram,
numericamente, uma quantidade maior de células sem reação acrossômica (P<0,10) do que o
sêmen convencional com o diluidor S (Sexing – T2). No entanto, não houve diferença entre o
sêmen convencional com diluidor L (Lagoa – T1) e com o sêmen sexado nos tempos zero
(T3), três horas após (T4) e seis horas após (T5) (Gráfico 7).

6 Resultados 73
Priscilla Harumi Tanno
Letras diferentes sobre as barras indicam tendência a significância (P<0,10). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído, submetido à técnica de citometria de fluxo no tempo zero e posterior criopreservação. T4 – Sêmen diluído, submetido à técnica de citometria de fluxo três horas após e posterior criopreservação. T5 – Sêmen diluído, submetido à técnica de citometria de fluxo seis horas após e posterior criopreservação. Gráfico 7 - Médias ± erros padrão da quantidade de células (%) sem reação acrossomal (SRA)
no sêmen bovino pós-descongelação nos cinco tratamentos descritos acima – Pirassununga - 2009
Não houve diferença estatística (P>0,05) entre os tratamentos, quanto à quantidade de
células sem reação acrossomal (SRA) no sêmen tanto de taurinos como de zebuínos. A
variação foi de 69,0 a 79,3 (Gráfico 8).

6 Resultados 74
Priscilla Harumi Tanno
Não houve diferença estatística (P> 0,05). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído, submetido à técnica de citometria de fluxo no tempo zero e posterior criopreservação. T4 – Sêmen diluído, submetido à técnica de citometria de fluxo três horas após e posterior criopreservação. T5 – Sêmen diluído, submetido à técnica de citometria de fluxo seis horas após e posterior criopreservação. Gráfico 8 - Médias ± erros padrão da quantidade de células (%) sem reação acrossomal (SRA)
no sêmen de taurinos e zebuinos pós-descongelação submetidos ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga – 2009
6.4 PEROXIDAÇÃO DA MEMBRANA PLASMÁTICA EM SÊMEN CONVENCIONAL
E SEXADO
Foi observado que houve aumento significativo (P<0,05) na intensidade da
fluorescência captada pelo fotomultiplicador, que corresponde à maior peroxidação em
espermatozóides com membrana plasmática íntegra nos tratamentos T3, T4, T5 (sêmen
sexado), comparado com os tratamentos T1 (sêmen convencional com diluidor L – Lagoa),
T2 (sêmen convencional com diluidor S – Sexing). Entretanto, não houve diferença ao longo
do tempo (0, 3 e 6 horas) na sexagem espermática (Gráfico 9).

6 Resultados 75
Priscilla Harumi Tanno
Letras diferentes sobre as barras significam diferenças estatísticas (P < 0,05). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído, submetido à técnica de citometria de fluxo no tempo zero e posterior criopreservação. T4 – Sêmen diluído, submetido à técnica de citometria de fluxo três horas após e posterior criopreservação. T5 – Sêmen diluído, submetido à técnica de citometria de fluxo seis horas após e posterior criopreservação. Gráfico 9 - Médias ± erros padrão dos valores lineares médios da intensidade de fluorescência
da população espermática com membrana plasmática íntegra e corada com a sonda fluorescente C11-BODIPY581/591 (PEIFMPI) no sêmen bovino submetido ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h). Os valores estão representados em unidades arbitrárias (u.a.) – Pirassununga – 2009
As amostras de sêmen sexado dos taurinos e dos zebuínos após a descongelação
apresentaram, em média, um aumento dos valores lineares médios da intensidade de
fluorescência da população espermática com membrana plasmática íntegra e corada com a
sonda fluorescente C11-BODIPY581/591 (P<0,05) nos tratamentos T3, T4 e T5 (sêmen sexado
nos tempos zero, três e seis horas após, respectivamente) em relação aos grupos T1 e T2
(sêmen convencional com diluidor L e S, respectivamente). Todavia, não apresentou
diferença ao longo do tempo no sêmen sexado. Já entre taurinos e zebuínos, houve maior
peroxidação lipídica (P<0,05) nos taurinos (variou de 101,6 a 240,9 u.a.) do que nos zebuínos
(variou de 91,1 a 225,3 u.a.) (Gráfico 10).
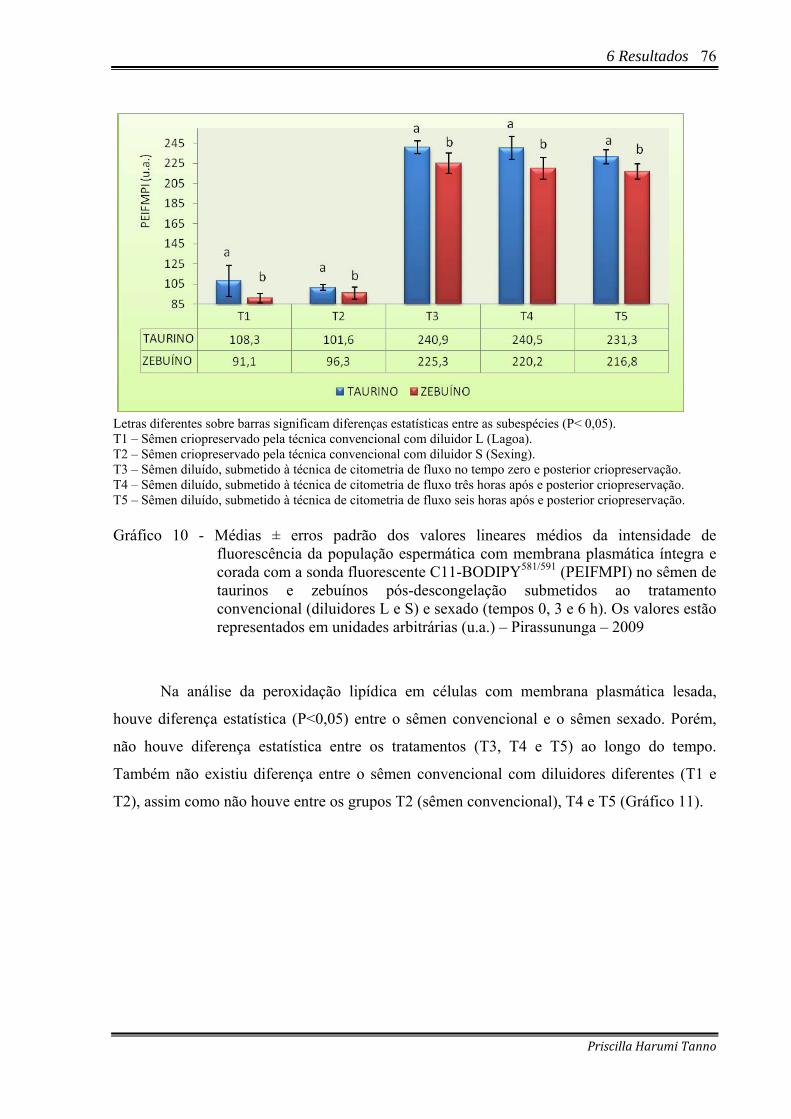
6 Resultados 76
Priscilla Harumi Tanno
Letras diferentes sobre barras significam diferenças estatísticas entre as subespécies (P< 0,05). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído, submetido à técnica de citometria de fluxo no tempo zero e posterior criopreservação. T4 – Sêmen diluído, submetido à técnica de citometria de fluxo três horas após e posterior criopreservação. T5 – Sêmen diluído, submetido à técnica de citometria de fluxo seis horas após e posterior criopreservação. Gráfico 10 - Médias ± erros padrão dos valores lineares médios da intensidade de
fluorescência da população espermática com membrana plasmática íntegra e corada com a sonda fluorescente C11-BODIPY581/591 (PEIFMPI) no sêmen de taurinos e zebuínos pós-descongelação submetidos ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h). Os valores estão representados em unidades arbitrárias (u.a.) – Pirassununga – 2009
Na análise da peroxidação lipídica em células com membrana plasmática lesada,
houve diferença estatística (P<0,05) entre o sêmen convencional e o sêmen sexado. Porém,
não houve diferença estatística entre os tratamentos (T3, T4 e T5) ao longo do tempo.
Também não existiu diferença entre o sêmen convencional com diluidores diferentes (T1 e
T2), assim como não houve entre os grupos T2 (sêmen convencional), T4 e T5 (Gráfico 11).
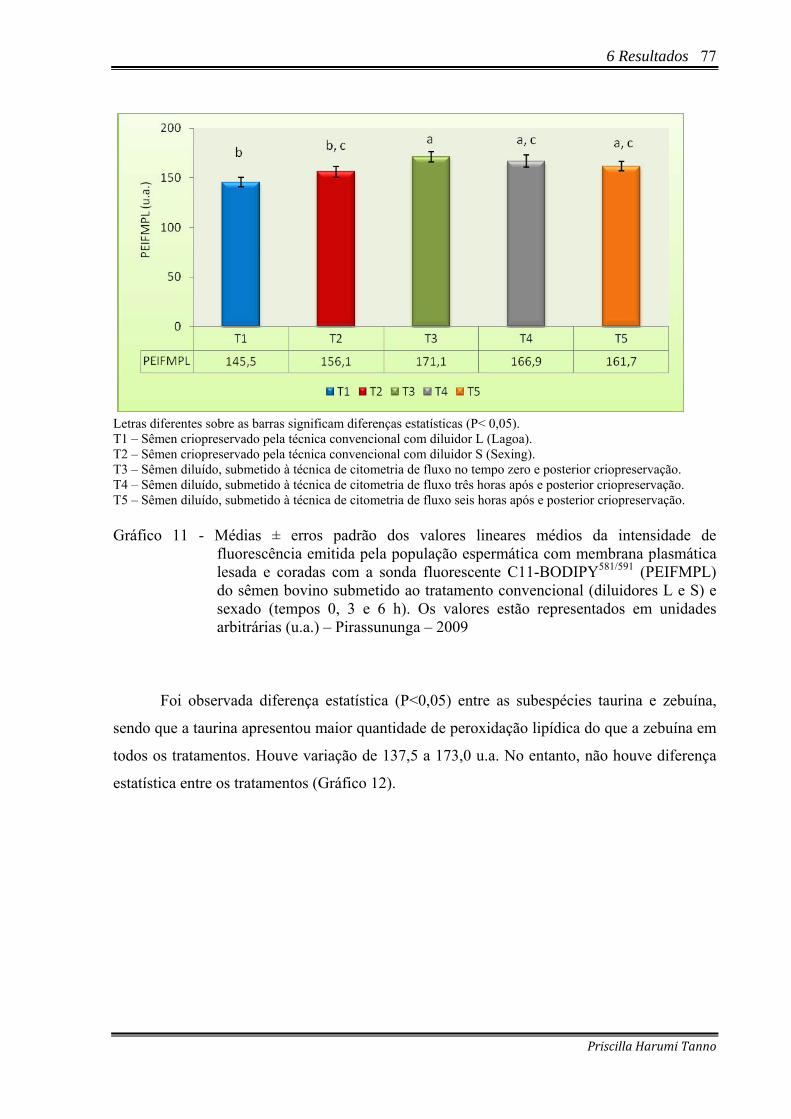
6 Resultados 77
Priscilla Harumi Tanno
Letras diferentes sobre as barras significam diferenças estatísticas (P< 0,05). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído, submetido à técnica de citometria de fluxo no tempo zero e posterior criopreservação. T4 – Sêmen diluído, submetido à técnica de citometria de fluxo três horas após e posterior criopreservação. T5 – Sêmen diluído, submetido à técnica de citometria de fluxo seis horas após e posterior criopreservação. Gráfico 11 - Médias ± erros padrão dos valores lineares médios da intensidade de
fluorescência emitida pela população espermática com membrana plasmática lesada e coradas com a sonda fluorescente C11-BODIPY581/591 (PEIFMPL) do sêmen bovino submetido ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h). Os valores estão representados em unidades arbitrárias (u.a.) – Pirassununga – 2009
Foi observada diferença estatística (P<0,05) entre as subespécies taurina e zebuína,
sendo que a taurina apresentou maior quantidade de peroxidação lipídica do que a zebuína em
todos os tratamentos. Houve variação de 137,5 a 173,0 u.a. No entanto, não houve diferença
estatística entre os tratamentos (Gráfico 12).

6 Resultados 78
Priscilla Harumi Tanno
Letras diferentes sobre barras significam diferenças estatísticas entre as subespécies (P< 0,05). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído, submetido à técnica de citometria de fluxo no tempo zero e posterior criopreservação. T4 – Sêmen diluído, submetido à técnica de citometria de fluxo três horas após e posterior criopreservação. T5 – Sêmen diluído, submetido à técnica de citometria de fluxo seis horas após e posterior criopreservação. Gráfico 12 - Médias ± erros padrão dos valores lineares médios da intensidade de
fluorescência da população espermática com membrana plasmática lesada e corada com a sonda fluorescente C11-BODIPY581/591 (PEIFMPL) no sêmen de taurinos e zebuinos pós-descongelação submetidos ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h). Os valores estão representados em unidades arbitrárias (u.a.) – Pirassununga – 2009
6.5 FOSFORILAÇÃO DO AMINOÁCIDO TIROSINA EM ESPERMATOZÓIDES COM
MEMBRANA PLASMÁTICA ÍNTEGRA EM SÊMEN CONVENCIONAL E SEXADO
As amostras de sêmen sexado apresentaram, em média, um aumento na emissão da
intensidade de fluorescência (FOIFMI - P< 0,05), o que significa que houve uma elevação na
ligação do anticorpo antifosfotirosina na superfície da membrana plasmática íntegra do
espermatozóide nos tempos 0, 3 e 6 horas após em relação ao sêmen convencional (T2). No
entanto, não houve diferença significativa entre o T1 e T2. Foi feita a associação da sonda IP
com a finalidade de excluir os espermatozóides com a membrana plasmática lesada, evitando
assim a contagem destas células durante a análise (Gráfico 13).
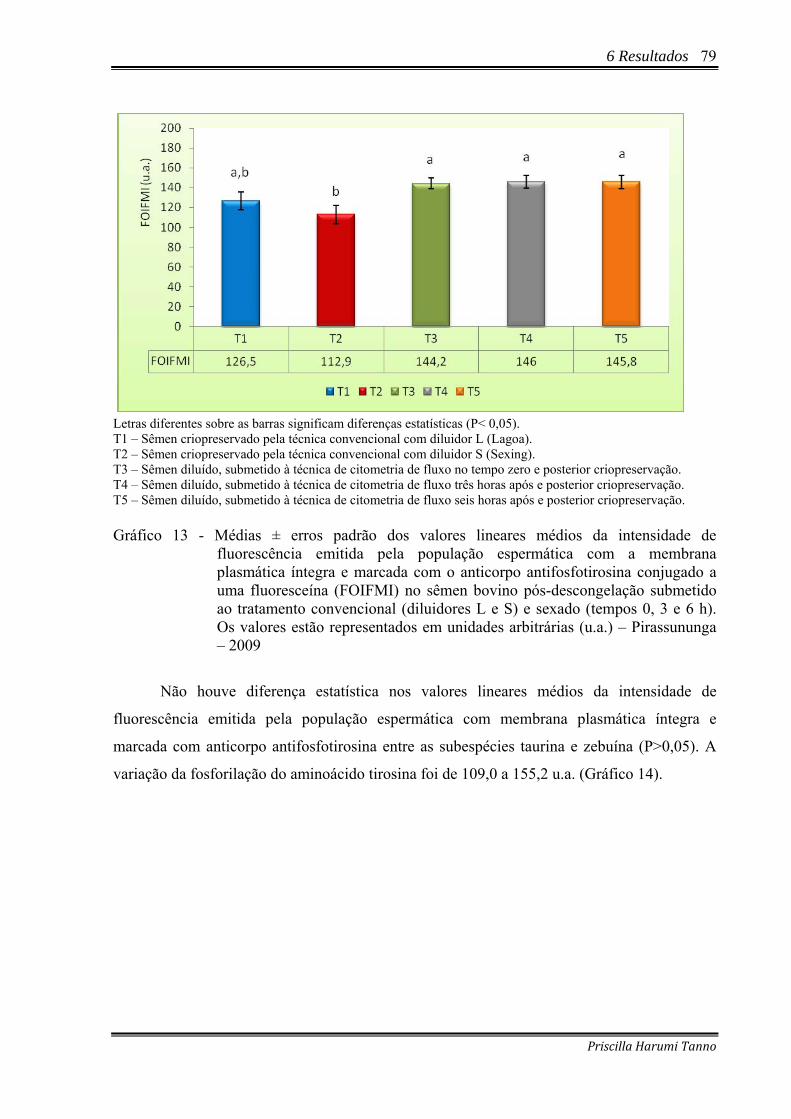
6 Resultados 79
Priscilla Harumi Tanno
Letras diferentes sobre as barras significam diferenças estatísticas (P< 0,05). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído, submetido à técnica de citometria de fluxo no tempo zero e posterior criopreservação. T4 – Sêmen diluído, submetido à técnica de citometria de fluxo três horas após e posterior criopreservação. T5 – Sêmen diluído, submetido à técnica de citometria de fluxo seis horas após e posterior criopreservação. Gráfico 13 - Médias ± erros padrão dos valores lineares médios da intensidade de
fluorescência emitida pela população espermática com a membrana plasmática íntegra e marcada com o anticorpo antifosfotirosina conjugado a uma fluoresceína (FOIFMI) no sêmen bovino pós-descongelação submetido ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h). Os valores estão representados em unidades arbitrárias (u.a.) – Pirassununga – 2009
Não houve diferença estatística nos valores lineares médios da intensidade de
fluorescência emitida pela população espermática com membrana plasmática íntegra e
marcada com anticorpo antifosfotirosina entre as subespécies taurina e zebuína (P>0,05). A
variação da fosforilação do aminoácido tirosina foi de 109,0 a 155,2 u.a. (Gráfico 14).
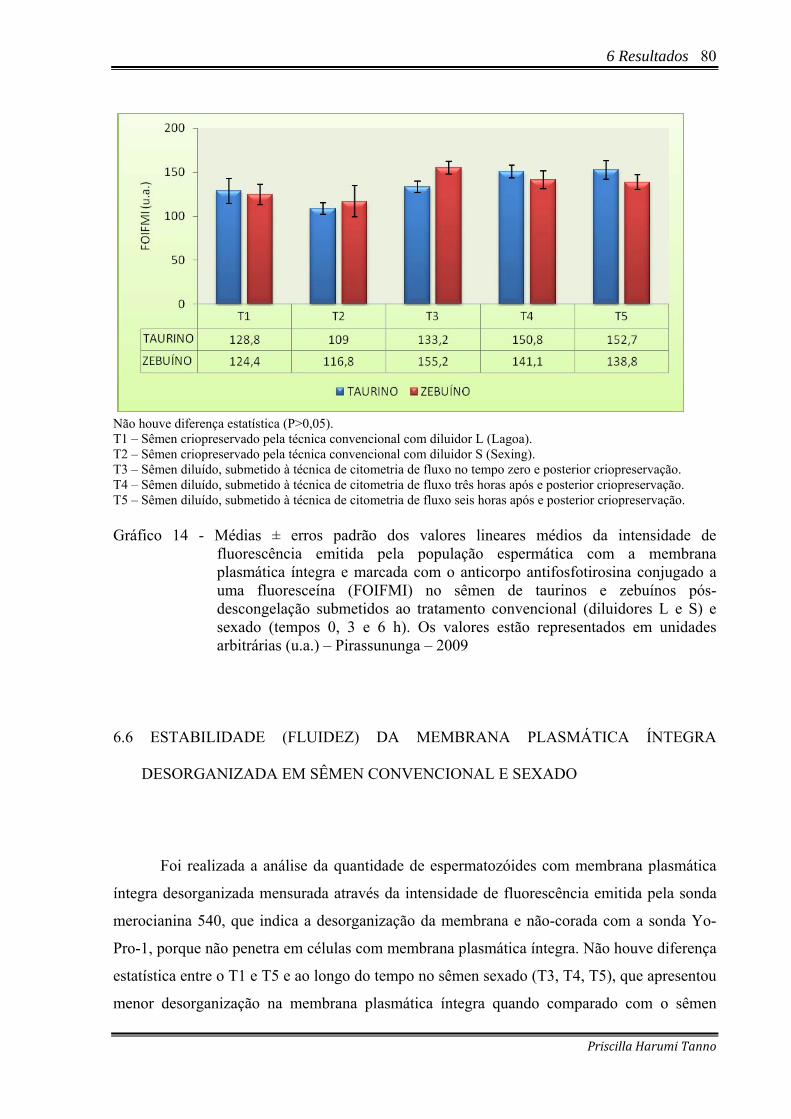
6 Resultados 80
Priscilla Harumi Tanno
Não houve diferença estatística (P>0,05). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído, submetido à técnica de citometria de fluxo no tempo zero e posterior criopreservação. T4 – Sêmen diluído, submetido à técnica de citometria de fluxo três horas após e posterior criopreservação. T5 – Sêmen diluído, submetido à técnica de citometria de fluxo seis horas após e posterior criopreservação. Gráfico 14 - Médias ± erros padrão dos valores lineares médios da intensidade de
fluorescência emitida pela população espermática com a membrana plasmática íntegra e marcada com o anticorpo antifosfotirosina conjugado a uma fluoresceína (FOIFMI) no sêmen de taurinos e zebuínos pós-descongelação submetidos ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h). Os valores estão representados em unidades arbitrárias (u.a.) – Pirassununga – 2009
6.6 ESTABILIDADE (FLUIDEZ) DA MEMBRANA PLASMÁTICA ÍNTEGRA
DESORGANIZADA EM SÊMEN CONVENCIONAL E SEXADO
Foi realizada a análise da quantidade de espermatozóides com membrana plasmática
íntegra desorganizada mensurada através da intensidade de fluorescência emitida pela sonda
merocianina 540, que indica a desorganização da membrana e não-corada com a sonda Yo-
Pro-1, porque não penetra em células com membrana plasmática íntegra. Não houve diferença
estatística entre o T1 e T5 e ao longo do tempo no sêmen sexado (T3, T4, T5), que apresentou
menor desorganização na membrana plasmática íntegra quando comparado com o sêmen

6 Resultados 81
Priscilla Harumi Tanno
convencional (T1 e T2), mas existiu diferença significativa (P<0,05) entre os tratamentos T1 e
T2; T1, T2 e o sêmen sexado (T3, T4 e T5) (Gráfico 15).
Letras diferentes sobre as barras significam diferenças estatísticas (P< 0,05). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído, submetido à técnica de citometria de fluxo no tempo zero e posterior criopreservação. T4 – Sêmen diluído, submetido à técnica de citometria de fluxo três horas após e posterior criopreservação. T5 – Sêmen diluído, submetido à técnica de citometria de fluxo seis horas após e posterior criopreservação. Gráfico 15 - Médias ± erros padrão da quantidade de células com membrana plasmática
íntegra desorganizada (%), corada com a sonda fluorescente merocianina 540 e não-corada com o Yo-Pro-1 (MEMID) no sêmen bovino pós-descongelação submetido ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga – 2009
Na avaliação da quantidade de espermatozóides com membrana plasmática íntegra
desorganizada mensurada através da intensidade de fluorescência emitida pela sonda
merocianina 540 e Yo-Pro-1, foi observado que houve diferença significativa (P<0,05) entre
as subespécies nos tratamentos T2, T3, T4 e T5, com os zebuínos apresentando maior
quantidade de células com membrana plasmática íntegra desorganizada do que os taurinos,
com exceção do T1 que os resultados foram iguais para ambas.

6 Resultados 82
Priscilla Harumi Tanno
Letras diferentes sobre barras significam diferenças estatísticas entre as subespécies(p< 0,05). T1 – Sêmen criopreservado pela técnica convencional com diluidor L (Lagoa). T2 – Sêmen criopreservado pela técnica convencional com diluidor S (Sexing). T3 – Sêmen diluído, submetido à técnica de citometria de fluxo no tempo zero e posterior criopreservação. T4 – Sêmen diluído, submetido à técnica de citometria de fluxo três horas após e posterior criopreservação. T5 – Sêmen diluído, submetido à técnica de citometria de fluxo seis horas após e posterior criopreservação. Gráfico 16 - Médias ± erros padrão da quantidade de células com membrana plasmática
íntegra desorganizada (%), corada com a sonda fluorescente merocianina 540 e não-corada com o Yo-Pro-1 (MEMID) no sêmen de taurinos e zebuinos pós-descongelação submetidos ao tratamento convencional (diluidores L e S) e sexado (tempos 0, 3 e 6 h) – Pirassununga - 2009

7 Discussão

7 Discussão 84
Priscilla Harumi Tanno
7 DISCUSSÃO
O processo de sexagem de sêmen pela técnica de citometria de fluxo envolve várias
etapas que são susceptíveis a alterações morfo-funcionais nos espermatozóides, como o uso do
corante de DNA (Hoechst 33342), reaquecimento e incubação, forças mecânicas de vibração
para a formação da gota durante a passagem da célula espermática pelo nozzle tip, a exposição
ao laser UV, a carga elétrica, a alta gravidade durante a desaceleração do espernatozóide
dentro do tubo coletor, a alta diluição, pressão elevada (40 psi) e resistência a mudanças na
composição dos meios diluidores, a centrifução, a refrigeração e o processo de criopreservação
(GARNER; SEIDEL JR., 2003; GARNER, 2006; MOCÉ et al., 2006; GRAAF et al., 2007;
GARCIA et al., 2007). Desta forma, neste experimento foram testados os efeitos que todas
estas etapas causam aos espermatozóides considerando lesões na membrana plasmática, reação
acrossomal, peroxidação lipídica e capacitação espermática; conjunto de avaliações ainda não
descrito na literatura.
Pela observação dos resultados apresentados no gráfico 1, pôde-se notar uma tendência
do sêmen sexado criopreservado com o diluidor S avaliado logo após a descongelação (T3)
apresentar mais alta porcentagem de espermatozóides com membrana plasmática intacta e
acrossomo sem reação acrossomal do que aquele sêmen convencional criopreservado com
diluidor L (T1); todavia, esta diferença não foi notada quando comparou-se os outros grupos
de tratamento. Esses resultados podem sugerir algum efeito do diluidor, já que numericamente
o sêmen criopreservado com o diluidor L (T1) apresentou os mais baixos percentuais de
células com membrana plasmática intacta e sem reação acrossomal (25,6%) do que os outros
grupos de tratamento (29,9%; 31,6%; 29,1% e 29,7%; respectivamente para os grupos: T2, T3,
T4 e T5). Os efeitos do diluidor sobre as membranas plasmática e acrossomal já vem sendo
observados em vários outros trabalhos (AMIRAT et al., 2004; ARRUDA et al., 2005;
CELEGHINI et al., 2008).
Com as técnicas de análise realizadas no presente experimento, foi possível verificar
que o sêmen convencional (T1 e T2) apresentou maior população de células com reação
acrossômica e membrana plasmática íntegra do que o sêmen sexado (T3, T4 e T5); apesar da
diferença estatística, o percentual desta população foi baixo (Gráfico 3). Estes resultados são
divergentes daqueles encontrados por Oliveira (2009) e Oliveira et al. (2009) que observaram
maior percentual de espermatozóides com membrana plasmática íntegra e lesão de acrossoma

7 Discussão 85
Priscilla Harumi Tanno
após o processo de sexagem (33,8%) do que no sêmen convencional (0,0%). Esta discrepância
se explica pelo fato de que Oliveira et al. (2009) utilizaram a técnica de sexagem por gradiente
de Percoll, a qual poderia induzir a reação acrossomal precocemente. Por estes dados pode-se
dizer que há diferença entre a técnica de sexagem por citometria de fluxo quanto à preservação
da reação acrossômica precoce em relação à outra técnica citada.
Quando foi considerada somente a integridade de membrana plasmática, observou-se
menor percentual (26,7%) para sêmen convencional criopreservado com diluidor L (T1) do
que para o sêmen sexado no tempo zero (T3; 32,0%), nenhuma diferença foi encontrada entre
os outros grupos de tratamento (Gráfico 5). É importante ressaltar que a integridade da
membrana plasmática é fundamental para que ocorra o desencadeamento da capacitação
espermática com a finalidade de fecundar o oócito (FLESCH; GADELLA, 2000).
Quanto à reação acrossômica, tanto o sêmen convencional quanto o sêmen sexado
apresentaram um valor alto na população de células sem reação acrossômica, com variação de
72,8 a 77,9% e tendência a significância (P<0,10) do sêmen sexado (T3 e T5) apresentar mais
alto percentual de células sem reação acrossomal. Com estes resultados, pode-se notar neste
trabalho, que a membrana plasmática é muito mais sensível ao processo de sexagem e
criopreservação do que a membrana acrossomal, tanto no sêmen convencional como no
sexado.
A peroxidação lipídica nos espermatozóides com membrana plasmática íntegra foi mais
alta no sêmen sexado (T3, T4 e T5) com valores de 224,1 a 233,1 u.a., comparado ao sêmen
convencional (T1 e T2) que variou de 99,0 a 99,3 u.a. (Gráfico 9). A peroxidação lipídica é
conseqüência da ação de espécies reativas de oxigênio (EROs) em baixas concentrações, que
funcionam como segundo mensageiros na capacitação espermática e reação acrossômica,
através do aumento dos níveis de cAMP intracelular e a subsequente ativação da proteína
quinase A (PKA), estimulando a fosforilação do aminoácido tirosina de diversas proteínas
(VISCONTI; KOPF, 1998; FLESCH; GADELLA, 2000; LEWIS; AITKEN, 2001;
LAMIRANDE; FLAHERTY, 2008). Provavelmente, isto se deve ao processo de sexagem do
sêmen, pois não houve efeito do diluidor e nem do tempo de espera do ejaculado para a
sexagem. Porém, ainda não foi elucidado até que ponto este aumento interfere na fertilidade,
sabe-se que concentrações excessivamente altas de EROs são associadas a efeitos tóxicos,
levando a lesão de membrana, resultante da depleção de ATP e até morte celular (FLESCH;
GADELLA, 2000; LAMIRANDE, FLAHERTY, 2008).

7 Discussão 86
Priscilla Harumi Tanno
Neste trabalho, pode-se observar no gráfico 11 que o sêmen convencional apresentou
quantidades menores de subpopulação de células com membrana plasmática lesada com
peroxidação lipídica (145,5 – 156,1 u.a.) em relação ao sêmen sexado (161,7 -171,1 u.a.),
porém este valor foi maior comparado com as células com membrana plasmática íntegra com
peroxidação no sêmen convencional (99,0 -99,3 u.a.). No entanto, segundo Gadella
(comunicação pessoal, 2009), a sonda C11-BODIPY581/591 possui uma afinidade menor em
células com membrana plasmática lesada, por isso pode mostrar valores mais baixos de
peroxidação. As células lesadas liberam o seu pequeno, mas existente conteúdo, podendo
alterar o meio em que as células se encontram, desta maneira suprindo os fatores necessários
para que ocorra a capacitação (VADNAIS; ROBERTS, 2007).
Outro fator indicativo de capacitação espermática é a presença de proteínas fosforiladas
na superfície do espermatozóide, que apresentou um aumento significativo (P<0,05) no sêmen
sexado (Gráfico 13), ao contrário do esperado por Rathi et al. (2001), este aumento não foi
acompanhado pela quantidade de células classificadas como capacitadas pela análise de
associação de sondas YO-PRO-1 e Merocianina 540 (Gráfico 15), pois o sêmen convencional
apresentou maior população espermática com membrana plasmática íntegra desorganizada
(P<0,05). De acordo com Flesch et al. (1999), a fosforilação da tirosina pode induzir a
mudanças conformacionais nas proteínas, levando desta maneira, a uma ativação ou inativação
das proteínas. E, segundo Rathi et al. (2001), a sonda Merocianina 540 tem mostrado que cora
células com membrana íntegra mais intensamente se seus componentes lipídicos estiverem em
um alto estado de desordem, como é o caso do espermatozóide capacitado.
Embora o sêmen in natura partisse de um padrão mínimo de características normais, ou
seja, de boa qualidade, tanto para a subespécie taurina quanto para a zebuína, foi possível
observar nestes resultados que os zebuínos parecem ser melhores e mais resistentes do que os
taurinos devido à maior quantidade de espermatozóides com integridade de membrana
plasmática sem reação acrossômica (Gráfico 2), integridade de membrana plasmática (Gráfico
6), à menor população espermática com peroxidação lipídica tanto com membrana plasmática
íntegra (Gráfico 10) quanto lesada (Gráfico 12). Estes resultados diferem daqueles observados
por Lucio et al. (2009) e Oliveira (2009), que não observaram diferença nos percentuais de
lesão de membrana plasmática e acrossomal, nem função mitocondrial de sêmen sexado e
convencional entre taurinos e zebuínos.
No entanto, entre os taurinos e zebuínos, não foram encontradas diferenças estatísticas
(P>0,05) na análise da população espermática com membrana plasmática íntegra e reação

7 Discussão 87
Priscilla Harumi Tanno
acrossômica (Gráfico 4) e nem na quantidade de células sem reação acrossômica (Gráfico 8).
Somente na análise de associação das sondas Merocianina 540 e Yo-Pro-1 (Gráfico 16)
ocorreu o contrário do que seria o esperado, os zebuínos apresentaram maior quantidade de
espermatozóides com membrana plasmática desorganizada do que os taurinos nesta
característica, significando que possuem mais espermatozóides capacitados devido ao aumento
da fluidez da membrana, como descrito por Rathi et al. (2001).
Em relação à fosforilação da tirosina, não houve efeito de subespécie, apenas de
tratamento. No entanto, não foi encontrada nenhuma comparação entre as subespécies
associada a este trabalho na literatura, somente de diferenças de DNA existentes entre as raças
(GARNER, 2006).
Um fato importante observado neste experimento é o de que o sêmen sexado não piora
com o tempo de espera do ejaculado (zero, três e seis horas após) em relação a todas as
análises realizadas, nas quais houve tendência a significância (P<0,10) nas populações de
células sem reação acrossomal e com membrana plasmática íntegra (SRAMI), com membrana
plasmática íntegra (MI) e espermatozóides sem reação acrossomal (SRA). Houve significância
(P<0,05) na quantidade de espermatozóides com reação acrossomal e membrana plasmática
íntegra (CRAMI), peroxidação lipídica com membrana plasmática íntegra (PEIFMI),
peroxidação lipídica com membrana plasmática lesada (PEIFMPL), espermatozóides com
fosforilação da tirosina e membrana plasmática íntegra (FOIFMI) e membrana plasmática
íntegra desorganizada (MEMID). Isto vem a comprovar que o ejaculado mantido à temperatura
ambiente (21°C) por até seis horas para a produção do sêmen sexado vendido comercialmente
aparentemente não altera a taxa de prenhez, visto que não há reclamações de partidas
específicas, ou seja, de acordo com o tempo de espera do ejaculado para o início do processo
de sexagem.
É importante ressaltar que as partidas de sêmen sexado sejam comercializadas com
uma quantidade mínima de células mortas, pois estas produzem altas quantidades de EROs,
que associadas à baixa concentração por palheta interferem na fertilidade a campo. A
vantagem do sêmen sexado é que a técnica de citometria de fluxo reduz a população de células
mortas e com lesão celular através do descarte de gametas comprometidos identificados pelo
corante food colour (FD&C 40).

8 Conclusões

8 Conclusões 89
Priscilla Harumi Tanno
8 CONCLUSÕES
- A sexagem pela técnica de citometria de fluxo não provoca lesões na membrana
plasmática, nem causa reação acrossomal, sendo o percentual de espermatozóides com
membrana plasmática íntegra e sem reação acrossomal semelhante ao sêmen
convencional com tendência a ser melhor.
- O sêmen sexado pela técnica de citometria de fluxo sofre maior peroxidação lipídica
das membranas espermáticas e fosforilação do aminoácido tirosina quando comparado
ao sêmen convencional.
- O sêmen sexado pela técnica de citometria de fluxo apresenta menor quantidade de
espermatozóides com membrana plasmática desorganizada, quando comparado ao
com a técnica convencional.
- Os espermatozóides de zebuínos são mais resistentes a lesão de membrana plasmática,
reação acrossomal e peroxidação lipídica do que espermatozóides de taurinos, quando
submetidos à sexagem pela técnica de citometria de fluxo.
- A fosforilação do aminoácido tirosina de espermatozóides submetidos às técnicas de
sexagem (citometria de fluxo) ou convencional é semelhante para animais de raças
zebuínas e taurinas.
- O sêmen de taurinos quando submetido à sexagem pela técnica de citometria de fluxo
apresenta menor quantidade de espermatozóides com membrana plasmática
desorganizada, quando comparado com o de zebuínos.
- Não existem diferenças na integridade ou desorganização da membrana plasmática,
reação acrossomal, peroxidação lipídica e fosforilação do aminoácido tirosina nos
espermatozóides bovinos, bem como entre zebuínos e taurinos, quando o ejaculado, in
natura, é mantido por zero, três ou seis horas antes de iniciar a sexagem pela técnica
de citometria de fluxo.

8 Conclusões 90
Priscilla Harumi Tanno
- Uma vez que a desorganização da membrana plasmática, reação acrossomal,
peroxidação lipídica e fosforilação do aminoácido tirosina são eventos envolvidos com
a capacitação espermática, os resultados obtidos neste experimento em relação à
sexagem pela técnica de citometria de fluxo merecem maiores estudos.

Referências

Referências 92
Priscilla Harumi Tanno
REFERÊNCIAS
AITKEN, R. J. Free radicals, lipid peroxidation and sperm function. Reproduction, Fertility and Development, v. 7, p. 659-668, 1995.
AMANN, R. P. Issues affecting commercialization of sexed sperm. Theriogenology, v. 52, p. 1441-1457, 1999.
AMBROGI, M. de; SPINACI, M.; GALEATI, G.; TAMANINI, C. Viability and DNA fragmentation in differently sorted boar spermatozoa. Theriogenology, v. 66, p. 1994-2000, 2006.
AMIRAT, L.; TAINTURIER, D.; JEANNAAU, L.; THORIN, C.; G´ERARD, O.; COUNTERS, J. L.; ANTON, M. Bull semen in vitro fertility after cryopreservation using egg yolk LDL: a comparison with Optidyl®, a commercial egg yolk extender. Theriogenology, v. 61, p. 895–907, 2004.
ANDERSSON, M.; TAPONEN, J.; KOSHINEN, E.; DAHLBOM, M. Effect of insemination with doses of 2 or 15 million frozen-thawed spermatozoa and semen deposition site on pregnancy rate in dairy cows. Theriogenology, v. 61, p. 1583-1588, 2004.
ANDRADE, A. F. C. Efeito da adição do plasma seminal nas mudanças semelhantes à capacitação (Criocapacitação) em espermatozóides criopreservados de eqüinos. 2009. 132 f. Tese (Doutorado em Medicina Veterinária) – Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2009. ARRUDA, R. P.; CELEGHINI, E. C. C.; ANDRADE, A. F. C.; RAPHAEL. C. F.; PERES, K. R.; NEVES, L. C. Influência da qualidade do sêmen nos resultados de prenhez em programas de IATF e TETF. In: SIMPÓSIO INTERNACIONAL DE REPRODUÇÃO ANIMAL APLICADA, 2., 2006, Paraná. Anais... 2006, p. 157-164.
ARRUDA, R. P.; FORERO-GONZALEZ, R. A.; CELEGHINI, E. C. C.; RAPHAEL, C. F. Effects of cryopreservation using different freezing techniques and cryoprotectants on plasmatic, acrosomal and mitochondrial membranes of bovine spermatozoa. Acta Scientiae Veterinariae, v. 33, p. 329, 2005.
BLONDIN, P.; BEAULIEU, M.; FOURNIER, V.; MORIN, N.; CRAWFORD, L.; MADAN, P.; KING, W. A. Analysis of bovine sexed sperm for IVF from sorting to the embryo. Theriogenology, v. 71, p. 30-38, 2009.
BODMER, M.; JANETT, F.; HÄSSIG, M.; DASS, N.; REICHERT, P.; THUN, R. fertility in heifers and cows after low dose insemination with sex-sorted and non-sorted sperm under field conditions. Theriogenology, v. 64, p. 1647-1655, 2005.
BREIBART, H.; COHEN, G.; RUBINSTEIN, S. Role of actin cytoskeleton in mammalian sperm capacitation and the acrosome reaction. Reproduction, v. 129, p. 263-268, 2005. Disponível em: <http://www.reproduction-online.org/cgi/reprint/129/3/263>. Acesso em: 24 maio 2009.

Referências 93
Priscilla Harumi Tanno
BREIBART, H.; NAOR, Z. Protein kinases in mammalian sperm capacitation and the acrosome reaction. Reviews of Reproduction, v. 4, p. 151-159, 1999.
BRENER, E.; RUBINSTEIN, S.; COHEN, G.; SHTERNALL, K; RIVLIN, J.; BREIBART, H. Remodeling of the actin cytoskeleton during mammalian sperm capacitation and acrosome reaction. Biology of Reproduction, v. 68, p. 837-845, 2003.
BROWERS, J. F.; SILVA, P. F. N.; GADELLA, B. M. New assays for detection and localization of endogenous lipid peroxidation products in living boar sperm after BTS dilution or after freeze-thawing. Theriogenology, v. 63, p. 458-469, 2005.
BUFFONE, M. G.; CALAMERA, J. C.; VERSTRAETEN, S. V.; DONCEL, G. F. Capacitation-associated protein tyrosine phosphorylation and membrane fluidity changes are impaired in the spermatozoa of asthenozoospermic patients. Reproduction Research, v. 129, p. 697-705, 2005. Disponível em: <http://www.reproduction-online.org/cgi/reprint/129/6/697>. Acesso em: 24 maio 2009.
BURNAUGH, L.; SABEUR, K.; BALL, B. A. Generation of superoxide anion by equine spermatozoa as detected by dihydroethidium. Theriogenology, v. 67, p. 580-589, 2007.
CAMPOS-CHILLON, L. F.; DE LA TORRE, J. F. Effect of concentration of sexed bovine sperm sorted at 40 and 50 psi on developmental capacity of in vitro produced embryos. Theriogenology, v. 59, p. 506, 2003. Abstract.
CATT, S. L.; O'BRIEN, J. K.; MAXWEKK, W. M. C.; EVANS, G. Assessment of ram and boar spermatozoa during cell-sorting by flow cytometry. Reproduction in Domestic Animals, v. 32, p. 251-258, 1997.
CATT, S. L.; SAKKAS, D.; BIZZARO, D.; BIANCHI, P. G.; MAXWELL, W. M. C.; EVANS, G. Hoechst staining and exposure to UV laser during flow cytometric sorting does not affect the frequency of detected endogenous DNA nicks in abnormal and normal human spermatozoa. Molecular Human Reproduction, v. 3, p. 821-5, 1997.
CELEGHINI, E. C. C.; ARRUDA, R. P.; ANDRADE, A. F. C.; NASCIMENTO, J.; RAPHAEL, C. F.; RODRIGUES, P. H. M. Effects that bovine sperm cryopreservation using two different extenders has on sperm membranes and chromatin. Animal Reproduction Science, v. 104, p. 119-131, 2008.
CELEGHINI, E. C. C.; ARRUDA, R. P.; ANDRADE, A. F. C.; NASCIMENTO, J.; RAPHAEL, C. F. Practical techniques for bovine sperm simultaneous fluorimetric assessment of plasma, acrosomal and mitochondrial membranes. Reproduction in Domestic Animals, v. 42, p. 479-488, 2007.
CERCHIARO, I.; CASSANDRO, M.; DAL ZOTTO, R.; CARNIER, P.; GALLO, L. A Field study on fertility and purity of sex-sorted cattle sperm. Journal Dairy Science, v. 90, p. 2538-2542, 2007.
DASGUPTA, S.; MILLS, C. S.; FRASER, L. R. Ca2+ related changes in the capacitation state of human spermatozoa assessed by chlortetracycline fluorescence assay. Journal of Reproduction and Fertility, v. 1, p. 135-143, 1993.

Referências 94
Priscilla Harumi Tanno
DE VRIES, A.; OVERTON, M.; FETROW, J.; LESLIE, K.; EICKER, S.; ROGERS, G. Exploring the impact of sexed semen on the structure of the dairy industry. Journal Dairy Science, v. 91, p. 847-856, 2008.
DRUMMEN, G. P. C.; VAN LIEBERGER, L. C. M.; OP DEN KAMP, J. A. F.; POST, J. A. C-11 BODIPY581/591, an oxidation-sensitive fluorescent lipid peroxidation probe: (micro) spectroscopic characterization and validation of methodology. Free radical Biology & Medicine, v. 33, p. 473-490, 2002.
FLESCH, F. M.; COLENBRANDER, B.; VAN GOLDE, L. M. G.; GADELLA, B. M. Capacitation induces tyrosine phosphorylation of proteins in the boar sperm plasma membrane. Biochemical and Biophysical Research Communications, v. 262, p. 787-792, 1999. Disponível em: < http://www.idealibrary.com>. Acesso em: 25 maio 2009.
FLESCH, F. M.; GADELLA, B. M. Dynamics of the mammalian sperm plasma membrane in the process of fertilization. Biochimica et Biophysica Acta, v. 1469, p. 197-235, 2000.
FRASER, L.; DZIEKÓNSDA, A.; STRZERŹEK, R.; STRZERŹEK, J. Dialysis of boar semen prior to freezing-thawing: its effects on post-thaw sperm characteristics. Theriogenology, v. 67, p. 994-1003, 2007.
FRIJTERS, A. C. J.; MULLAART, E.; ROELOFS, R. M. G.; VAN HOORNE, R. P.; MORENO, J. F.; MORENO, O.; MERTON, J. S. What affects fertility of sexed bull semen more, low sperm dosage or the sorting process? Theriogenology, v. 71, p. 64-67, 2009.
FULWYLER, M. J. Hydrodynamic orientation of cells. The Journal of Histochemistry and Cytochemistry, v. 25, p. 781-783, 1977.
GADELLA, B. M. Sperm membrane physiology and relevance for fertilization. Animal Reproduction Science, v. 107, p. 229-236, 2008.
GARCIA, M. A.; GRAHAM, J. K. Dialysis of bovine semen and its effect on fresh and freeze-thawed spermatozoa. Cryobiology, v. 24, p. 446-454, 1987b.
GARCIA, M. A.; GRAHAM, J. K. Effects of low-molecular proteins (LMWF) from milk, egg yolk and seminal plasma on freezability of bovine spermatozoa. Cryobiology, v. 24, p. 429-436, 1987a.
GARCIA, E. M.; VÁSQUEZ, J. M.; PARRILLA, I.; CALVETE, J. J.; SANZ, L.; CABALLERO, I.; ROCA, J.; VASQUEZ, J. L.; MARTÍNEZ, E. A. Improving the fertilizing ability of sex sorted boar spermatozoa. Theriogenology, v. 68, p. 771-778, 2007.
GARNER, D. L. Flow cytometric sexing of mammalian sperm. Theriogenology, v. 65, p. 943-957, 2006.
GARNER, D. L. Hoechst 33342: the dye that enabled the differentiation of living X- an Y-chrmosome bearing mammalian sperm. Theriogenology, v. 71, p. 11-21, 2009.
GARNER, D. L. Sex-sorting mammalian sperm: concept to application in animals. Journal of Andrology, v. 22. p. 519-526, 2001.

Referências 95
Priscilla Harumi Tanno
GARNER, D. L.; SEIDEL, JR., G. E. History of commercializing sexed semen for cattle. Theriogenology, v. 69, p. 886-895, 2008.
GARNER, D.L.; SEIDEL, JR, G. E. Past, Present and future perspectives on sexing sperm. Canadian Journal of Animal Science, v. 83, p. 375-384, 2003.
GARNER, D. L.; GLEDHILL, B. L.; PINKEL, D.; LAKE, S.; STEPHENSON, D.; VAN DILLA, M. A.; JOHNSON, L. A. Quantification of the X- and Y-chromosome-bearing spermatozoa of domestic animals by flow cytometry. Biology of Reproduction, v. 28, p. 312-321, 1983.
GARNER, D. L.; HAFEZ, E. S. E. Espermatozóides e plasma seminal. In: HAFEZ, E. S. E; HAFEZ, B. Reprodução animal. 7. ed. Barueri: Manole Ltda, 2004. cap. 7, p. 97-110.
GILLAN, L.; EVANS, G.; MAXWELL, W. M. C. Flow Cytometric evaluation of sperm parameters in relation to fertility potential. Theriogenology, v. 63, p. 445-457, 2005.
GRAAF, S. P.; BEILBY, K. H.; UNDERWOOD, S. L.; EVANS, G.; MAXWELL, W. M. C. Sperm sexing in sheep and cattle: the exception and the rule. Theriogenology, v. 71, p. 89-97, 2009.
GRAAF, S. P.; EVANS, G.; GILLAN, L.; GUERRA, M. M. P.; MAXWELL, W. M. C.; O'BRIEN, J. K. The influence of antioxidant, cholesterol and seminal plasma on the in vitro quality of sorted and non-sorted ram spermatozoa. Theriogenology, v. 67, p. 217-227, 2007.
GRAHAM, J. K.; KUNZE, E. HAMMERSTEDT, R. H. Analysis of sperm cell viability, acrosomal integrity, and mitochondrial function using flow cytometry. Biology of Reproduction, v. 43, p. 55-64, 1990.
GRAVANCE, C. G.; GARNER, D. L.; BAUMBER, J.; BALL, B. A. Assessment of equine sperm mitochondrial function using JC-1. Theriogenology, v. 53, p. 1691-1703, 2000.
HARRISON, R. A. P.; GADELLA, B. M. Bicarbonate-induced membrane processing in sperm capacitation. Theriogenology, v. 63, p. 342-351, 2005.
HENDRICKS, K. E. M.; HANSEN, P. J. Can programmed cell death be induced in post-ejaculatory bull and stallion spermatozoa? Theriogenology, v. 71, p. 1138-1146, 2009.
HOHENBOKEN, W. D. Aplication of sexed semen in cattle production. Theriogenology, v. 52, p. 1421-1433, 1999.
HOLLINSHEAD, F. K.; O’BRIEN, J. K.; MAXWELL, W. M. C.; EVANS, G. Production of lambs of predetermined sex after the insemination of ewes with low numbers of frozen-thawed sorted X- or Y-chromosome bearing spermatozoa. Reproduction, Fertility, and Development, v. 14, p. 503-508, 2002.
HOWES, E.; PASCALL, J. C.; ENGEL, W.; JONES, R. Interactions between mouse ZP2 glycoprotein and proacrosin; a mechanism for secondary binding of sperm to the zona pellucid during fertilization. Journal of Cell Science, v. 114, p. 4127-4136, 2001.

Referências 96
Priscilla Harumi Tanno
JANUSKAUSKAS, A.; JOHANNISON, A.; RODRIGUEZ-MARTINEZ, H. Assessment of sperm quality through fluorometry and sperm chromatin structure assay in relation to field fertility of frozen-thawed semen from Swedish AI bulls. Theriogenology, v. 55, p. 947-61, 2001.
JANUSKAUSKAS, A.; JOHANNISON, A.; RODRIGUEZ-MARTINEZ, H. Subtle membrane changes in cryopreserved bull semen in relation with sperm viability, chromatin structure, and field fertility. Theriogenology, v. 60, p. 743-758, 2003.
JOHNSON, L. A. Sexing mammalian sperm for production of offspring: the state-of-the- art. Animal Reproduction, v. 60-61, p. 93-107, 2000.
JOHNSON, L. A.; FLOOK, J. P.; HAWK, H. W. Sex preselection in rabbits: Live births from X and Y bearing sperm separated by DNA and cell sorting. Biology of Reproduction, v. 41, p. 199-203, 1989.
JOHNSON, L. A.; WELCH, G. R. Sex-preselection: high speed flow cytometric sorting of X and Y sperm for maximum efficiency. Theriogenology, v. 52, p. 1323-1341, 1999.
JONES, R.; JAMES, P. S.; HOWES, L.; BRUCKBAUER, J.; KLENERMAN, D. Supramolecular organization of the sperm plasma membrane during maturation and capacitation. Asian Journal of Andrology, v. 9, p. 438-444, 2007.
JONES, R.; SHALGI, R.; HOYLAND, J.; PHILLIPS, D. M. Topographical rearrangement of a plasma membrane antigen during capacitation of rat spermatozoa in vitro. Developmental Biology, v. 139, p. 349-362, 1990.
LAMIRANDE, E.; JIANG, H.; ZINI, A.; KODAMA, H.; GAGNON, C. Reactive oxygen species and sperm physiology. Reviews of Reproduction, v. 2, p. 48–54, 1997.
LAMIRANDE, E.; LECLERC, P.; GAGNON, C. Capacitation as a regulatory event that primes spermatozoa for the acrosoma reaction and fertilization. Molecular Human Reproduction, v. 3, p. 175-194, 1997b. Disponível em: <http://molehr.oxfordjournals.org/cgi/reprint/3/3/175>. Acesso em: 24 maio 2009.
LAMIRANDE, E.; O’FLAHERT, C. Sperm activation: role of reactive oxygen species and kinases. Biochimica et Biophysica Acta, v. 1784, p. 106-115, 2008.
LEHNINGER, A. L.; NELSON, D. L.; COX, M. M. Membranas biológicas e transporte. In: Lehninger, A.L. Princípios da Bioquímica. São Paulo: Sarvier, 2002. cap. 12, p. 301-339.
LENZI, A.; PICARDO, M.; GANDINI, L.; DONDERO, F. Lipids of the sperm plasma membrane: from polyunsaturated fatty acids considered as markers of sperm function to possible scavenger therapy. Human Reproduction Update, v. 2, p. 246-256, 1996.
LEWIS, B.; AITKEN, R. J. A redox-regulated tyrosine phosphorilation cascade in rat spermatozoa. Journal of Andrology, v. 22, p. 611-622, 2001.
LINDSEY, A. C.; MORRIS, L. H.; ALLEN, W. R.; SHENK, J. L.; SQUIRES, E. L.; BRUEMMER, J. E. Hysteroscopic insemination of mares with low numbers of nonsorted or flow sorted spermatozoa. Equine Veterinary Journal, v. 34, p. 128-132, 2002.

Referências 97
Priscilla Harumi Tanno
LU, K. H.; SEIDEL JR., G. E. Effects of heparin and sperm concentration on cleavage rates of bovine oocytes inseminate with flow-citometrically sorted bovine sperm. Theriogenology, v. 62, p. 819-830, 2004.
LUCIO, A. C.; OLIVEIRA, L. Z.; CELEGHINI, E. C. C.; ANDRADE, A. F. C.; ARRUDA, R. P.; HOSSEPIAN DE LIMA, V. F. M. Influence of bovine subspecies in the recovered rate after the separation of X- bearing sperm by centrifugation in discontinuous Percoll density gradient. Animal Reproduction, v. 6, n. 1, p. 336, 2009. Supplement.
MAXWELL,W. M. C.; JOHNSON, L. A. Chlortetracycline analysis of boar spermatozoa after incubation, flow cytometric sorting, cooling, or cryopreservation. Molecular Reproduction and Development, v. 46, p. 408-418, 1997.
MAXWELL, W. M. C.; JOHNSON, L. A. Physiology of spermatozoa at high dilution rates: the influence of seminal plasma. Theriogenology, v. 52, p. 1353-1362, 1999.
MAXWELL, W. M. C.; WELCH, G. R.; JOHNSON, L. A. Viability and membrane integrity of spermatozoa after dilution and flow cytometric sorting in the presence or absence of seminal plasma. Reproduction, Fertility, and Development, v. 8, p. 1165-1168, 1996.
MAXWELL, W. M. C.; EVANS, G.; HOLLINSHEAD, F. K.; BATHGATE, R.; GRAAF, S. P.; ERIKSSON, B. M.; GILLAN, L.; MORTON, K. M.; O’BRIEN, J. K. Integration of sperm sexing technology into the ART toolbox. Animal Reproduction Science, v. 82-83, p. 79-95, 2004.
MOCÉ, E.; GRAHAM, J. K.; SCHENK, J. L. Effect of sex-sorting on the ability of fresh and cryopreserved bull sperm to undergo an acrosome reaction. Theriogenology, v. 66, p. 929-936, 2006.
MOORE, L. H. A.; TATCHER, W. W. Major advances associated with reproduction in dairy cattle. Journal Dairy Science, v. 89, p. 1254-1266, 2006.
MORRIS, L. H. A.; HOLLINSHEAD, F. K.; EVANS, G.; MAXWELL, W. M. C. The longevity and acrosome status of stallion flow-sorted spermatozoa. Theriogenology, v. 59, p. 512, 2003. Aabstract.
NAGY. S.; JANSEN, J.; TOPPER, E. K.; GADELLA, B. M. A triple-stain flow cytometric method to assess plasma- and acrosome-membrane integrity of cryopreserved bovine sperm immediately after thawing in presence of egg-yolk particles. Biology of Reproduction, v. 68, p. 1828-1835, 2003.
O´CONNEL, M.; MCCLURE, N.; LEWIS, S. E. M. The effects of cryopreservation on sperm morphology, motility and mitochondrial function. Human Reproduction, v. 17, n 3, p. 704-709, 2002.
OLIVEIRA, L. Z. Efeitos do processo de seleção do sexo por centrifugação em gradiente de densidade na qualidade de espermatozóides bovinos. 2009. 135 p. Dissertação (mestrado) – Universidade Estadual Paulista, Faculdade de Ciências Agrárias e Veterinárias, Jaboticabal, 2009.

Referências 98
Priscilla Harumi Tanno
OLIVEIRA, L. Z.; ARRUDA, R. P.; CELEGHINI, E. C. C.; ANDRADE, A. F. C.; LUCIO, A. C.; HOSSEPIAN DE LIMA, V. F. M. Sperm demage evaluation caused by separation of X-bearing sperm by centrifugation in discontinuous Percoll density gradient using fluorescence probes association. Animal Reproduction, v. 6, n. 1, p. 335, 2009. Supplement.
PARRILLA, I.; VASQUEZ, J. M.; CUELLO, C.; GILL, M. A.; ROCA, J.; DI BERARDINO, D.; MARTÍNEZ, E. A. Hoechst 33342 Stain and UV exposure do not induce genotoxic effect in flow-sorted boar spermatozoa. Reproduction, v. 128, p. 615-621, 2004.
PARRILLA, I.; VASQUEZ, J. M.; GIL, M. A.; CABALLERO, I.; ALMIÑANA, C.; ROCA, J.; MARTINEZ, E. A. Influence of storage time on functional capacity of flow cytometrically sex-sorted boar spermatozoa. Theriogenology, v. 64, p. 86-98, 2005.
PENFOLD, L. M.; HOLT, C.; HOLT, W. V.; WELCH, G. R.; CRAN, D. G.; JOHNSON, L. A. Comparative motility of X and Y chromosome-bearing bovine sperm separated on the basis of DNA content by flow sorting. Molecular Reproduction and Development, v. 50, p. 323-327, 1998.
PETRUNKINA, A. M.; GRÖPPER, B.; TÖPFER-PETERSEN, E.; GÜNZEL-APEL. A.-R. Volume regulatory function and sperm membrane dynamics as parameters for evaluating cryoprotective efficiency of a freezing extender. Theriogenology, v. 63, p. 1390-1406, 2005.
PINKEL, D.; LAKE, L.; GLEDHILL, B. L.; VAN DILLA, M. A.; STEPHENSON, D.; WATCHMAKER, G. High resolution DNA content measurements of mammalian sperm. Cytometry, v. 3, p. 1-9, 1982.
PONS-REJRAJI, H.; BAILEY, J. L.; LECLERC, P. Cryopreservation affects bovine sperm intracellular parameters associated with capacitation and acrosome exocytosis. Reproduction, Fertility and Development, v. 21, p. 525-537, 2009b.
PONS-REJRAJI, H.; BAILEY, J. L.; LECLERC, P. Modulation of bovine sperm signaling pathways: correlation between intracellular parameters and sperm capacitation and acrosome exocytosis. Reproduction, Fertility and Development, v. 21, p. 511-524, 2009a.
RATH, D.; JOHNSON, L. A. Aplication and commercialization of flow cytometrically sex-sorted semen. Reproduction in Domestic Animals, v. 43, p. 338-346, 2008.
RATH, D.; MOENCH-TEGEDER, G.; TAYLOR, U.; JOHNSON, L. A. Improved quality of sex sorted sperm: a prerequisite for wider commercial application. Theriogenology, v. 71, p. 22-29, 2009.
RATHI, R.; COLENBRANDER, B.; BEVERS, M. M.; GADELLA, B. M. Evaluation of in vitro capacitation of stallion spermatozoa. Biology of Reproduction, v. 65, p. 462-470, 2001.
SANOCKA, D.; KURPISZ, M. Reactive oxygen species and sperm cells. Reproductive Biology and Endocrinology, v. 2, p. 1-7, 2004. Disponível em: <http://www.rbej.com/content/2/1/12>. Acesso em: 24 maio 2009.
SEIDEL JR., G.E. Economics of selecting for sex: the most important genetic trait. Theriogenology, v. 59, p. 585-598, 2003.

Referências 99
Priscilla Harumi Tanno
SEIDEL JR., G. E. Overview of sexing sperm. Theriogenology, v. 68, p. 443-6, 2007.
SEIDEL JR., G. E. Sperm sexing technology – The transition to commercial application – An introduction to the symposium “Update on sexing mammalian sperm”. Theriogenology, v. 71, p. 1-3, 2009.
SEIDEL JR, G.E.; GARNER, D.L. Current status of sexing mammalian spermatozoa. Reproduction, v. 124, p. 733-743, 2002.
SEIDEL JR., G. E.; GARNER, D. L. Sexing mammalian sperm by flow cytometry. Reproduction, v. 124, p. 733-743, 2002.
SEIDEL JR., G. E.; SCHENK, J. L.; HERICKHOFF, L. A.; DOYLE, S. P.; BRINK, Z.; GREEN, R. D. Insemination of heifers with sexed sperm. Theriogenology, v. 52, p. 1407-20, 1999.
SHARPE, J. C.; EVANS, K. M. Advances in flow cytometry for sperm sexing. Theriogenology, v. 71, p. 4-10, 2009.
SCHENK, J. L.; SUH, T. K.; CRAN, D. G.; SEIDEL JR., G. E. Cryopreservation of flow-sorted bovine spermatozoa. Theriogenology, v. 52, p. 1375-1391, 1999.
SILVA, P. F. N.; GADELLA, B. M. Detection of damage in mammalian sperm cells. Theriogenology, v. 65, p. 958-978, 2006.
SILVA, P. F. N.; GADELLA, B. M.; COLENBRANDER, B.; ROELEN, B. A. J. Exposure of bovine sperm to pro-oxidants impairs the developmental competence of the embryo after the first cleavage. Theriogenology, v. 67, p. 609-619, 2007.
SPINNACI, M.; VOLPE, S.; BERNARDINI, C.; DE AMBROGI, M.; TAMANINI, C.; SEREN, E. GALEATI, G. Sperm sorting procedure induces a redistribution of Hsp70 but not Hsp60 and Hsp90 in boar spermatozoa. Journal of Andrology, v. 27, p. 899-907, 2006.
SUH, T. K.; SCHENK, J. L.; SEIDEL JR, G. E. High pressure flow cytometric sorting damage sperm. Theriogenology, v. 64, p. 1035-1048, 2005.
THOMAS, A. D.; MEYERS, S. A.; BALL, B. A. Capacitation-like changes in equine spermatozoa following cryopreservation. Theriogenology, v. 65, p. 1531-1550, 2006.
THOMAS, C. A.; GARNER, D. L.; DEJARNETTE, J. M.; MARSHALL, C. E. Effect of cryopreservation on bovine sperm organelle function and viability as determined by flow cytometer. Biology of Reproduction, v. 58, p. 786-793, 1998.
URNER, F.; SAKKAS, D. Protein phosphorilation in mammalian spermatozoa. Reproduction, v. 125, p. 17-26, 2003. Disponível em: < www.reproduction-online.org/cgi/reprint/125/1/17 >. Acesso em: 24 maio 2009.
VISCONTI, P. E.; BAILEY, J. L.; MOORE, G. D.; DE LAMIRANDE, E.; LECLERC, P.; GAGNON, C. Capacitation as a regulatory event that primes spermatozoa for the acrosoma reaction and fertilization. Molecular Human Reproduction, v. 3, p. 175-194, 1997.

Referências 100
Priscilla Harumi Tanno
Disponível em: < http://molehr.oxfordjournals.org/cgi/reprint/3/3/175 >. Acesso em: 24 maio 2009.
VISCONTI, P. E.; BAILEY, J. L.; MOORE, G. D.; PAN, D.; OLDS-CLARKE, P. Capacitation of mouse spermatozoa I. Correlation between the capacitation state and protein tyrosine phophorylation. Development, v. 121, p. 1129-1137, 1995.
VISCONTI, P. E.; KOPF, G. S. Regulation of protein phosphorilation during sperm capacitation. Biology of Reproduction, v. 59, p. 1-6, 1998.
WEIGEL, K. A. Exploring the role of sexed semen in dairy production systems. Journal Dairy Science, v. 87, p. E120-E130, 2004. Supplement.
WERTHEIMEIR, E. V.; SALICIONI, A. M.; LIU, W.; TREVINO, C. L.; CHAVEZ, J.; HERNÁNDEZ-GONZÁLEZ, E. O.; DARSZON, A.; VISCONTI, P. E. Chloride is essential for capacitation and for the capacitation-associated increase in tyrosine phosphorylation. The Journal of Biological Chemistry, v. 283, p. 35539-35550, 2008.
WHEELER, M. B.; RUTLEDGE, J. J.; FISHER-BROWN, A.; VANETTEN, T.; MALUSKY, S.; BEEBE, D. J. Aplication of sexed semen technology to in vitro embryo production in cattle. Theriogenology, v. 65, p. 219-227, 2006.