
NATÁLIA REGINA SOUZA DA SILVA - repositorio.ufpe.br · Rejane Pereira Neves Ricardo Arraes de...
100
UNIVERSIDADE FEDERAL DE PERNAMBUCO CENTRO DE CIÊNCIAS DA SAÚDE PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA TROPICAL ANÁLISE DA OCORRÊNCIA DO GENE DA BETALACTAMASE EM ISOLADOS CLÍNICOS DE Staphylococcus spp. RESISTENTES À METICILINA PROCEDENTES DE HOSPITAL UNIVERSITÁRIO DA CIDADE DO RECIFE-PE. NATÁLIA REGINA SOUZA DA SILVA RECIFE/PE 2015
Transcript of NATÁLIA REGINA SOUZA DA SILVA - repositorio.ufpe.br · Rejane Pereira Neves Ricardo Arraes de...

UNIVERSIDADE FEDERAL DE PERNAMBUCO
CENTRO DE CIÊNCIAS DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA TROPICAL
ANÁLISE DA OCORRÊNCIA DO GENE DA BETALACTAMASE EM
ISOLADOS CLÍNICOS DE Staphylococcus spp. RESISTENTES À
METICILINA PROCEDENTES DE HOSPITAL UNIVERSITÁRIO DA
CIDADE DO RECIFE-PE.
NATÁLIA REGINA SOUZA DA SILVA
RECIFE/PE
2015

Dissertação apresentada ao Programa de
PósGraduação em Medicina Tropical do
Centro de Ciências da Saúde da
Universidade Federal de Pernambuco, como
parte dos requisitos para a obtenção do Título
de Mestre em Medicina Tropical
NATÁLIA REGINA SOUZA DA SILVA
ANÁLISE DA OCORRÊNCIA DO GENE DA BETALACTAMASE EM
ISOLADOS CLÍNICOS DE Staphylococcus spp. RESISTENTES À
METICILINA PROCEDENTES DE HOSPITAL UNIVERSITÁRIO DA
CIDADE DO RECIFE-PE.
Orientadora: Drª. Maria Amélia Vieira Maciel
Co-orientadora: Drª. Nilma Cintra Leal
RECIFE/PE
2015


UNIVERSIDADE FEDERAL DE PERNAMBUCO (UFPE)
PRÓ-REITORIA PARA ASSUNTOS DE PESQUISA E PÓS-GRADUAÇÃO (PROPESQ)
CENTRO DE CIÊNCIAS DA SAÚDE (CCS)
PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA TROPICAL (PPGMEDTROP)
RELATÓRIO DA BANCA EXAMINADORA DA DISSERTAÇÃO DA MESTRANDA
N A T Á L I A R E G I N A S O U Z A D A S I L V A
No dia 24 de fevereiro de 2015, às 09h00, na Sala de Aula do PPGMEDTROP - no Bl. A, Térreo do
Hospital das Clínicas da UFPE -, os Membros Doutores: a Profa. Dra. Ana Catarina de Souza Lopes
(Presidente da Banca – UFPE), a Profa. Dra. Eulália Camelo Pessoa de Azevedo Ximenes (UFPE) e o
Prof. Dr. Wagner Luís Mendes de Oliveira (LINAT/UFPE), componentes da Banca Examinadora, em
sessão pública, arguiram a mestranda Natália Regina Souza da Silva sobre a sua Dissertação intitulada
“ANÁLISE DA OCORRÊNCIA DO GENE DA BETALACTAMASE EM ISOLADOS CLÍNICOS
DE Staphylococcus spp. RESISTENTES À METICILINA PROCEDENTES DE HOSPITAL UNIVERSITÁRIO DA
CIDADE DO RECIFE-PE”, a qual foi orientada pela Profa. Dra. Maria Amélia Vieira Maciel (UFPE) e co-
orientada pela Profa. Dra. Nilma Cintra Leal (CPqAM/FIOCRUZ). Ao final da arguição de cada membro
da Banca Examinadora e resposta da mestranda, as seguintes menções foram publicamente
fornecidas.
Profa. Dra. Ana Catarina de Souza Lopes
Profa. Dra. Eulália Camelo Pessoa de Azevedo Ximenes
Prof. Dr. Wagner Luís Mendes de Oliveira

UNIVERSIDADE FEDERAL DE PERNAMBUCO
CENTRO DE CIÊNCIAS DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA TROPICAL
REITOR
Anísio Brasileiro de Freitas Dourado
VICE-REITOR
Sílvio Romero de Barros Marques
PRÓ-RETOR PARA ASSUNTOS DE PESQUISA E PÓS-GRADUAÇÃO
Francisco de Sousa Ramos
DIRETOR DO CENTRO DE CIÊNCIAS DA SAÚDE
Nicodemos Teles de Pontes Filho
COORDENADORA DO PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA
TROPICAL
Valdênia Maria Oliveira de Souza
VICE-COORDENADORA DO PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA
TROPICAL
Vera Magalhães da Silveira
CORPO DOCENTE
Ana Catarina de Souza Lopes
Ana Lúcia Coutinho Domingues
Célia Maria Machado Barbosa de Castro
Edmundo Pessoa de Almeida Lopes Neto
Fábio André Brayner dos Santos
Heloísa Ramos Lacerda de Melo
Maria Amélia Vieira Maciel
Maria de Fátima Pessoa Militão de Albuquerque
Maria Rosângela Cunha Duarte Coelho
Marli Tenório Cordeiro
Rejane Pereira Neves
Ricardo Arraes de Alencar Ximenes
Valdênia Maria Oliveira de Souza
Vera Magalhães da Silveira
Vlaudia Maria Assis Costa

DEDICO,
A meus pais, Marcelo e Marli e ao meu
irmão, Marcelinho, pelo apoio constante e
amor incondicional. A Luiz Felipe, que
tornou essa caminhada mais leve e especial.

AGRADECIMENTOS
A Deus, pela força, sabedoria e paciência dadas a mim para o seguimento dessa jornada,
permitindo que eu não desanimasse diante dos novos desafios que surgiam.
A meus pais, Marcelo e Marli, pelo amor, carinho e dedicação e por estarem junto a
mim, ajudando e incentivando cada nova conquista, mesmo quando os problemas da vida eram
tão presentes.
Ao meu irmão, Marcelinho, pelo apoio e amizade, essenciais em minha vida e pelas
risadas e brincadeiras, que tornaram a caminhada menos árdua.
A Luiz Felipe, que sempre confiou e acreditou no meu potencial, mesmo quando eu
desanimava. Pelo amor e paciência sempre presentes, mesmo com a distância existente.
À Professora Amélia Maciel, pela confiança, orientação, ajuda, amizade, paciência e
incentivo dedicados a mim durante o desenvolvimento deste trabalho.
À Dra. Nilma Leal, pela confiança, paciência e orientação que sempre dedicou a mim.
Aos meus amigos, tão importantes nesta caminhada, pelo companheirismo e por alegrar
essa jornada. Quero agradecer de modo especial a Amanda, Camila e Marcelle, por se fazerem
sempre presentes, principalmente nas horas mais difíceis e por me dedicarem tanto amor todos
os dias.
A Armando, Cynthia, Jailton, Jussyêgles, Laura, Lilian, Paula, Stephanie, Valdemir e
todos que fazem o laboratório de Microbiologia, pelo apoio e por compartilharem comigo cada
nova descoberta.
Ao Centro de Pesquisas Aggeu Magalhães, que possibilitou orientação e infraestrutura
para o desenvolvimento de algumas partes desse trabalho.
A Wagner, Fabiana e Silvana, do Departamento de Microbiologia do Centro de
Pesquisas Aggeu Magalhães, por sua disponibilidade em ajudar sempre que necessitei.
À Dona Jussara e Dona Djanira, que sempre me receberam muito bem no laboratório de
microbiologia e que sempre cuidaram de tudo e todos no departamento.
Aos professores e funcionários da Disciplina de Microbiologia e Imunologia, que de
alguma maneira contribuíram para o desenvolvimento da minha pesquisa.
Aos professores da Pós-Graduação em Medicina Tropical da Universidade Federal de
Pernambuco pelo empenho na condução do curso e por todos os ensinamentos.
A Professora Yêda e Árina Leite, que sempre me compreenderam e me incentivaram
em todos os momentos desta caminhada.

A Cassiana, Christophe, Diêgo, Flávia, Gabriel, Geazi, Carlos, Davi, Arquimedes,
Lívia, Monique, Maysa, Roseana, Thaís e todos os que fazem os Niates da UFPE, sem seu
incentivo, compreensão e disponibilidade nada seria possível.
A todos que direta ou indiretamente estiveram ao meu lado e me ajudaram a alcançar
mais este objetivo. Meu muito obrigada!

“Nunca deixe que lhe digam que não vale
a pena
Acreditar no sonho que se tem
Ou que seus planos nunca vão dar certo
Ou que você nunca vai ser alguém
Tem gente que machuca os outros
Tem gente que não sabe amar
Mas eu sei que um dia a gente aprende
Se você quiser alguém em quem confiar
Confie em si mesmo
Quem acredita sempre alcança...”
(Renato Russo)

RESUMO
SILVA, N.R.S Análise da ocorrência do gene da betalactamase em isolados clínicos de
Staphylococcus spp. resistentes à meticilina procedentes de Hospital Universitário da cidade do
Recife-PE. 99f. Dissertação de Mestrado Universidade Federal de Pernambuco- Centro de
Ciências da Saúde. Programa de Pós-Graduação em Medicina Tropical. Recife. Pernambuco
Staphlylococcus desenvolveu a resistência aos betalactâmicos através de dois principais
mecanismos, a expressão das betalactamases e a produção de PBP2a, codificadas pelos genes
blaZ e mecA, respectivamente. Apesar da detecção do gene mecA por técnicas moleculares ser
considerado o padrão ouro para a identificação das cepas de Staphlylococcus resistentes à
meticilina (MRS), alguns estudos têm verificado discrepância entre a apresentação fenotípica
da resistência à meticilina nessas cepas e a presença do gene mecA.O presente estudo teve como
objetivo verificar se existe diferença de frequência do gene blaZ entre isolados clínicos de
Staphylococcus spp. resistentes à meticilina portadores do gene mecA e os não portadores do
gene. Foram estudados 47 isolados provenientes de amostras clínicas de hospital universitário
da cidade do Recife. Essas amostras foram submetidas aos testes de susceptibilidade
antimicrobiana, detecção do gene mecA para separação dos grupos de comparação e posterior
detecção da presença de betalactamase através dos testes de cefinase em disco, Clover-Leaf e
detecção do gene blaZ por PCR. Desta forma, foram observadas diferenças na frequência
quando utilizados os testes fenotípicos para detecção. No teste de cefinase em disco, foram
positivos 33,3% dos isolados portadores do gene mecA e 56,25% dos não portadores. No teste
de Clover-Leaf, foi detectada a produção de betalactamase em 33,3% dos isolados portadores
do gene mecA e 65,62% dos não portadores. Na pesquisa do gene blaZ, no entanto, não foi
verificada diferença significativa nas frequências de detecção entre os grupos. Desta forma,
indica-se o uso dos testes fenotípicos de detecção de produção de betalactamase, por
apresentarem baixo custo e fácil operação, além de demonstrarem boa sensibilidade.
Palavras-chave: Beta-Lactamases. Staphylococcus. Resistência Microbiana a medicamentos

ABSTRACT
SILVA, N.R.S. Analysis of the occurrence of the beta-lactamase gene in clinical isolates of
Staphylococcus spp. methicillin-resistant coming from University Hospital in Recife-PE 99f.
Dissertation, Federal University of Pernambuco Center of Medical Sciences Saude.
Postgraduate Program in Tropical Medicine. Recife. Pernambuco.
Staphlylococcus developed resistance to beta-lactam via two major mechanisms of beta-
lactamase expression and production of PBP2a encoded by gene mecA and blaZ, respectively.
Although the detection of the mecA gene by molecular techniques be considered the gold
standard for identifying Staphlylococcus of methicillin-resistant strains (MRS), some studies
have found discrepancies between the phenotypic presentation of methicillin resistance in these
strains and the presence of the mecA gene. This study aimed to verify if there is frequency
difference of blaZ gene among clinical isolates of Staphylococcus spp. methicillin-resistant
carriers of mecA gene and non-carriers of the gene.47 isolates from clinical samples of
University Hospital in Recife were studied.These samples were subjected to antimicrobial
susceptibility tests and subsequent detection of mecA gene by PCR for separation of the
comparison groups. After this, was performed to detect the production of beta-lactamase using
the disk Cefinase tests, Clover Leaf and detection of blaZ gene by PCR. Thus, diferences in
frequency were observed when phenotypics tests were used for detection. In Cefinase test
disk,were positive 33.3% of the isolates carrying the mecA gene and 56.25% of non-carriers. In
Clover-Leaf assay, the production of beta-lactamase was detected in 33.3% of isolates carrying
the mecA gene and 65.62% of non-carriers.In search of blaZ gene, however, there was no
significant difference in the frequency detection between groups. In this way, it indicates the
use of phenotypic tests for detection of beta-lactamase production for having low cost and easy
operation as well as demonstrating good sensitivity.
Keywords: Beta-Lactamases. Staphylococcus. Drug Resistance, Microbial

LISTA DE ILUSTRAÇÕES
Figura 1: Alexander Ogston.....................................................................................................20
Figura 2: Aspecto macroscópico das colônias de Staphylococcus aureus em Ágar Sangue de
Carneiro 5%...............................................................................................................................22
Figura 3: Demonstração da prova da Coagulase em Tubo........................................................23
Figura 4: Demonstração da prova da DNAse............................................................................24
Figura 5: Demonstração da prova de fermentação do Ágar Manitol.........................................25
Figura 6: Estrutura dos onze tipos de SCCmec.........................................................................31
Figura 7: Arranjo dos domínios da proteína BlaR1...................................................................33
Figura 8: Sistema de regulação de controle da expressão de PBP2a e betalactamase................34
Figura 9: Lesões do impetigo...................................................................................................36
Figura 10: Lesões da Síndrome da pele escaldada....................................................................37
Figura 11: Cronologia da resistência aantibióticos dos S.aureus..............................................40
Figura 1: Teste da Cefinase em disco........................................................................................62
Figura 2: Teste de Clover-Leaf.................................................................................................63
Figura 3: Gel de agarose 1% demostrando produto de amplificação do gene blaZ...................63

LISTA DE TABELAS
Tabela 1: Distribuição da frequência da detecção da betalactamase de acordo com a PCR do
gene blaZ, teste da Cefinase em disco e teste de Clover-Leaf em isolado sdeStaphylococcus
spp. resistentes à meticilina com e sem a presença do gene mecA..............................................64
Tabela 2: Sensibilidade e especificidade dos métodos analíticos para detecção de
betalactamase............................................................................................................................64

LISTA DE ABREVIATURAS E SIGLAS
Ágar Casoy: Ágar Triptona de Soja
ATCC: American Type Culture Collection
BHI: Infusão de cérebro e coração
BlaI: Repressor das betalactamases
BlaR1: Proteína Transdutora de sinal
BlaZ: Betalactamase
CA-MRSA: Staphylococcus aureus Resistente à Meticilina de origem Comunitária
CCIH: Comissão de Controle de Infecção Hospitalar
CCS: Centro de Ciências da Saúde
CDC: Centers of Disease Control and Prevention
CEP: Comitê de Ética em Pesquisa
CLSI: Clinical and Laboratory Standards Insitute
DNA: Ácido Desoxirribonucléico
DNAse: Desoxirribonuclease
dNTP: Desoxirribonucleotídeos Fosfatados
ECN: Estafilococos coagulase negativos
EDTA: Ácido Etileno diamino Tetra-Acético
HC: Hospital das Clínicas de Pernambuco
HCl: Ácido Clorídrico
HIV-AIDS: Vírus da Imunodeficiência Humana – Síndrome da Imunodeficência Aguda
K+: Cátion Potássio
kb: kilobase
MgCl2: Cloreto de Magnésio
MRS: Staphylococcus spp. Resistente à Meticilina
MRSA: Staphylococcus aureus Resistente à Meticilina
MSSA: Staphylococcusaureus Sensível à Meticilina
NaCl: cloreto de Sódio
pb: Pares de base
PBP: Proteína ligadora de Penicilina
PCR: Reação em cadeia da Polimerase
rRNA: RNA ribossomal

RT-PCR: Reação em cadeia da Polimerase em tempo real
qRT-PCR: Reação em cadeia da Polimerase em tempo real quantitava
SCCmec: Staphylococcal Cassete Chromossome mec
TBE: Tampão Tris-borato
TCLE: Termo de Consentimento Livre e Esclarecido
TE: Tris-HCl+EDTA
UFPE: Universidade Federal de Pernambuco
UV: Ultravioleta
VISA/GISA: Staphylococcus aureus Sensível à Vancomicina
VRSA/GRSA: Staphylococcus aureus Resistente à Vancomicina

SUMÁRIO
1. INTRODUÇÃO..................................................................................................................17
2. REVISÃO DE LITERATURA..........................................................................................19
2.1. Características Gerais dos Staphylococus.....................................................................20
2.1.1. Histórico...............................................................................................................20
2.1.2. Classificação e Identificação Bioquímica.............................................................21
2.2. Mecanismo de Resistência aos antibióticos em estafilococos.......................................25
2.2.1. Mecanismo de Resistência aos antibióticos betalactâmicos.................................27
2.3. Betalactamase estafilocócica........................................................................................31
2.4. Staphylococcus spp. como causador de infecções........................................................35
2.5. Epidemiologia das infecções causadas por Staphylococcus spp. resistentes a
meticilina.....................................................................................................................37
2.6. Importância dos Staphylococcus resistentes à meticilina..............................................40
3. OBJETIVOS.......................................................................................................................42
3.1. Objetivo Geral..............................................................................................................43
3.2. Objetivos Específicos...................................................................................................43
4. MATERIAIS E MÉTODOS................................................................................................44
4.1. Desenho do estudo........................................................................................................45
4.2. Local do estudo............................................................................................................45
4.3. Categorização das variáveis.........................................................................................46
4.3.1. Variável Dependente............................................................................................46
4.3.2. Variáveis Independentes......................................................................................46
4.3.2.1. Variáveis Fenotípicas.............................................................................46
4.3.2.2. Variável Genética...................................................................................46
4.4. Operacionalização da pesquisa.....................................................................................46
4.4.1. Obtenção das amostras.........................................................................................46
4.4.2. Seleção dos isolados.............................................................................................47
4.4.2.1. Testes fenotípicos...................................................................................47
4.4.2.2. Identificação do gene mecA através de PCR...........................................47
4.4.3. Identificação fenotípica da produção de betalactamase........................................48
4.4.3.1. Teste da Cefinase em disco.....................................................................48
4.4.3.2. Teste de Clover-Leaf..............................................................................48
4.4.4. Extração de DNA total.........................................................................................49

4.4.5. Identificação do gene da betalactamase por PCR.................................................49
4.5. Considerações éticas....................................................................................................50
4.6. Análise Estatística........................................................................................................50
5. RESULTADOS...................................................................................................................51
Detecção de betalactamase em Staphylococcus spp. Isolados de amostras clínicas
provenientes de Hospital Universitário de Recife-Pernambuco-Brasil................................52
6. CONCLUSÃO....................................................................................................................65
REFERÊNCIAS........................................................................................................................67
APÊNDICE...............................................................................................................................82
ANEXOS..................................................................................................................................94

17
1. INTRODUÇÃO
A detecção do gene mecA por técnicas moleculares vem sendo considerado o padrão
ouro para a identificação das cepas de Staphlylococcus resistentes à meticilina (MRS)
(SWENSON et al., 2007; SHARIATI et al., 2010), porém em alguns estudos verifica-se
discrepância entre a apresentação fenotípica dessas cepas e a presença do gene mecA (ARAJ et
al., 1999; YOSHIDA et al., 2003; CUNY et al., 2011). Em um estudo realizado em hospital
universitário da cidade do Recife-PE foi verificado que várias cepas fenotipicamente resistentes
à meticilina não apresentavam o gene mecA quando submetidas à PCR (RABELO, 2012),
sugerindo que outro mecanismo possa estar envolvido nessa resistência, como por exemplo a
produção de betalactamases.
A betalactamase é codificada pelo gene estrutural blaZ, este por sua vez é regulado pela
proteína DNA ligante BlaI e pelo transdutor de sinal BlaR1 (HAO et al., 2012). Os reguladores
blaI e blaR1 possuem em seus transcritos homologia com os dos reguladores do mecA (mecI e
mecR1), o que permite a eles modular a transcrição do mecA (HACBARTH; CHAMBERS,
1993; McKINNEY et al., 2001; MILHEIRIÇO et al., 2011).
Hackbarth et al. (1994) demonstraram em seus estudos que, a inativação de genes
regulatórios conduz a expressão fenotípica da resistência à meticilina e que quando a produção
de PBP2a é reprimida, outros fatores não-mecA estão associados a este padrão de resistência.
Katayama et al. (2003) verificaram ainda, que os genes reguladores do mec e bla têm capacidade
de estabilizar o mecA, o que facilitaria sua disseminação. Constataram também que os
reguladores do blaZ são mais prováveis de realizar essa função, pois os reguladores do mecA
podem reprimir fortemente a expressão do mecA, sendo uma desvantagem para a sobrevivência
em meios com antibióticos betalactâmicos.
Alguns estudos também demonstram que a regulação exercida pelo blaI e blaR1 sobre
o mecA é mais eficiente que a realizada pelos próprios reguladores do gene (mecI e mecR1),
inferindo, inclusive, que essa regulação seja mais vantajosa para a expansão clonal das cepas
MRS (BLACK et al., 2011; MILHEIRIÇO et al., 2011). Rosato et al. (2003) ainda observaram
que a expressão da resistência à meticilina tem pouca relação com a transcrição do gene mecA,
mas também com a retenção de seus repressores, modulando a quantidade de produtos
produzidos pelo gene na presença de antibióticos betalactâmicos. Também observaram que
90% dos estafilococos retêm o gene blaZ e seus reguladores, apesar da baixa especificidade
para o substrato da betalactamase, não conferindo proteção contra a maioria dos antibióticos
betalactâmicos que são prevalentes em hospitais.

18
Apesar da importância que a betalactamase desempenha nas infecções por
Staphylococcus spp., poucos trabalhos no Brasil fazem referência a ocorrência desta enzima
relacionados ao padrão de resistência bacteriana, tampouco a sua caracterização genética. Com
o aumento dos casos de cepas fenotipicamente resistentes à meticilina, porém sem a presença
do gene mecA, observados nos pacientes hospitalizados em hospital universitário da cidade de
Recife – PE, busca-se, através deste trabalho, a identificação da ocorrência da betalactamase
como responsável pelo padrão de resistência à meticilina encontrados nestas cepas provenientes
de isolados clínicos.

19
2. Revisão de Literatura

20
2.1 Características gerais dos Staphylococcus
2.1.1 Histórico
Staphylococcus foram primeiramente observados e cultivados por Pasteur e Koch,
porém os estudos detalhados iniciais foram feitos por Ogston e por Rosenbach.
Alexander Ogston (1844-1929) (Figura 1) era um cirurgião escocês que em 1880
descobriu a principal causa de pus. Ogston possuía grande preocupação com a taxa de
mortalidade pós-operatória existente na época, por isso, foi um dos primeiros a aceitar a
antissepsia defendida por Joseph Lister, dado que os pacientes pós-operatórios de Lister não
apresentavam sinais de inflamação.
Figura 1: Alexander Ogston
Disponível em: http://www.bbc.co.uk/arts/yourpaintings/paintings/professor-sir-alexander-ogston-18441929-
105196, acesso em 11/11/2014
No entanto, Lister acreditava ser o ar a fonte da putrefação nas feridas. Através da
vaporização de ácido carbólico (fenol) sobre a região que seria realizado o ato cirúrgico,
desinfetando o campo cirúrgico, o que impedia a supuração. Ogston, entretanto, achava essa
explicação insuficiente e suspeitava que um germe pudesse ser o responsável pelo processo
supurativo. Então, ele abriu o abscesso de um de seus pacientes para fazer um esfregaço do pus
e observá-lo ao microscópio (ELEK, 1959).
Ogston então, lançou a hipótese que os abscessos agudos eram causados por pequenos
cocos. Para defender sua hipótese, ele injetou pus de abscessos agudos em cobaias de
camundongos e demonstrou que novos abscessos eram formados, seguidos por sinais de

21
septicemia. Ao observar o sangue dos animais sépticos, foram observados pequenos cocos. Se,
no entanto, o pus fosse pré-tratado com calor ou ácido carbólico antes da inoculação, a formação
do abscesso era evitada. Ogston também desenvolveu um método para cultivar esses cocos
inoculando ovos de galinha com pus e depois os encubando. No conteúdo dos ovos infectados
foi encontrada atividade piogênica semelhante à do pus inicial (OGSTON, 1880; OGSTON,
1881; ORENSTEIN, 2008).
Ogston, porém, não foi o primeiro a examinar pus microscopicamente e descrever cocos
(do grego Kokkos, que significa grão). Em 1874, Billroth observou cadeias de pequenos cocos
e as designou estreptococos. Em 1882, Ogston então nomeou o arranjo de cocos de
estafilococos, do grego staphyle, que significa cacho de uvas (OGSTON, 1882– 1883;
ORENSTEIN, 2008).
Três anos depois, em 1884, Anton J. Rosenbach (1842-1923), um cirurgião alemão,
isolou duas cepas de estafilococos, que ele nomeou a partir da pigmentação aparente das
colônias: Staphylococcus aureus, do latim aurum, que significa ouro e Staphylococcus albus
(hoje chamado epidermidis), do latim albus, que significa branco (ROSENBACH,1884;
ORENSTEIN, 2008).
Rosenbach mostrou que Staphylococcus aureus era responsável por infecções e
formação de furúnculos nas feridas e também que era microbiota normalmente encontrada na
pele.
Sempre que eram descritos por Rosenbach o crescimento destes cocos de coloração
amarela, os cirurgiões passavam a temer cada vez mais as infecções estafilocócicas em feridas
cirúrgicas. Desta forma, na era pré-antibiótico, Staphylococcus aureus ficou reconhecido como
um patógeno de difícil tratamento (VERTONI, 1991), sendo até hoje um patógeno importante
devido ao desenvolvimento de resistência aos antimicrobianos.
2.1.2 Classificação e Identificação Bioquímica
Os estafilococos pertencem ao reino Bacteria, filo Fimicutes, classe Bacilli, ordem
Bacillales, família Staphylococcaceae e gênero Staphylococcus.
O gênero Staphylococcus possui 32 espécies e 14 subespécies, muitas das quais podem
estar associadas a doenças tanto em humanos quanto em animais (KONEMAN et al., 2008).
Os estafilococos se apresentam como cocos gram-positivos ao serem corados pelo
método de Gram, com diâmetro que varia de 0,5 a 1,5µm, podendo ocorrer na forma de células
isoladas, em pares, tétrades e cadeias curtas, porém aparecem predominantemente em grupos

22
semelhantes a cachos de uvas. São imóveis, não formadores de esporos, anaeróbios facultativos
e produtores da enzima catalase. São ainda capazes de crescer em concentrações de 10% a 20%
de cloreto de sódio (NaCl) e em temperaturas que variam de 18ºC a 40ºC, no entanto apresentam
temperatura de crescimento ótimo entre 36ºC e 37ºC e pH ótimos de crescimento 6,0 e 7,0,
(FRANCO et al., 2000; KONEMAN et al., 2008; MURRAY et al., 2010).
Quando semeadas em Ágar Sangue de Carneiro, as colônias apresentam-se
arredondadas, lisas e brilhantes, com coloração variando do acinzentado ao amerelo-ouro,
podendo apresentar β-hemólise característica (Figura 2) (SANTOS et al., 2007).
Figura 2:Aspecto macroscópico das colônias de Staphylococcus aureus em Ágar Sangue de
Carneiro 5%
Disponível em: http://protomag.com/assets/a-brief-history-of-staph, acesso 22/11/2013.
Os isolados de Staphylococcus spp. são ainda classificados quanto à produção ou não
da enzima coagulase. Esta enzima é um importante fator de virulência que coagula o sangue ao
transformar o fibrinogênio em fibrina, da mesma forma que a trombina humana. Esta fibrina
recobre as células bacterianas permitindo sua rápida aglutinação o que dificulta os processos de
opsonização e fagocitose (KONEMANet al., 2008).
Staphylococcus aureus é a única espécie do gênero Staphylococcus encontrada
normalmente em humanos que secreta a enzima coagulase. Desta forma, a detecção desta
enzima vem sendo utilizada como identificação presuntiva do S. aureus em isolados
provenientes de humanos. As cepas pertencentes ao gênero Staphylococcusque não produzem
tal enzima, foram denominadas “estafilococos coagulase negativos” (ECN) (KONEMANet al.,
2008; MURRAYet al., 2010).
Existem 31 espécies de Staphylococcus coagulase negativa conhecidas, das quais os
mais frequentes são:
Staphylococcus epidermidis - causador de infecções provocadas por cateteres e
próteses e o mais frequente microrganismo encontrado em hemoculturas.

23
Staphylococcus saprophyticus - causador de infecção urinária em mulheres jovens.
Staphylococcus haemolyticus - importante devido à resistência aumentada aos
antimicrobianos, e por ser comumente confundido com S. aureus, pois apresenta β-
hemólise na placa de ágar sangue de carneiro (ANVISA, 2013).
A coagulase pode ser encontrada de duas formas: a coagulase conjugada e a coagulase
livre. A coagulase conjugada (prova em lâmina), também conhecida como fator de aglutinação
ou “clumping factor”, está presente na superfície da parede celular bacteriana. Quando as
células são suspensas em plasma, elas reagem com o fibrinogênio e formam-se cordões de
fibrina entre elas, o que causa o agrupamento sob a forma de grumos visíveis.
A coagulase livre (prova em tubo), no entanto, é secretada extracelularmente e reage
com um fator plasmático (Fator de Reação com a Coagulase – CRF) formando um complexo
que atua sobre o fibrinogênio, formando a fibrina (Figura 3)(ANVISA, 2013).
Figura 3: Demonstração da prova da Coagulase em Tubo
Disponível em:http://infectionnet.org,(modificado) acesso em 22/11/2013.
Geralmente, os estafilococos fazem parte da microbiota da pele humana normal e de
outros sítios anatômicos, porém S.aureus está frequentemente relacionado com diversas
infecções em seres humanos, principalmente em ambiente hospitalar, sendo considerada a
espécie de maior interesse médico (CASSETARI et al.,2005; KONEMAN et al.,2008;
SOARES et al.,2008; KLUYTMANS; STRUELENS,2009).
Durante vários anos, os ECN foram considerados saprofíticos da pele e mucosas, e, por
este motivo, eram desconsiderados quando isolados de materiais biológicos humanos, sendo
considerados contaminantes de cultura. A partir da década de 1980, no entanto, eles passaram
a ter importância, sendo S. epidermidis, S. haemolyticus, S. lugdunensis, S. warneri e S.

24
saprophyticus as espécies mais comumente associadas a infecções humanas, dentre esses, S.
epidermidis é o isolado mais comum (SLOOT et al., 1992).
Além da prova de coagulase, S.aureus pode também ser diagnosticado laboratorialmente
através das provas de:
Teste de aglutinação em Látex – é um teste sorológico que detecta a proteínas na
superfície do S. aureus, como proteína A, antígenos específicos de grupo do fator
de aglutinação, polissacarídeos capsulares. Para os testes a amostra deve ser
misturada com esferas de látex revestidas com anticorpo ou antígeno específico,
resultando em agregação das esferas se a substância suspeita estiver presente
(ZURITA, 2010; ANVISA, 2013).
Teste da DNAse – consiste na adição de ácido clorídrico (HCl) sobre as colônias
crescidas em meio Ágar DNAse, e posterior visualização de halo em torno das
mesmas (KONEMAN et al., 2008).
Figura 4: Demonstração da prova da DNAse
Disponível em:http://www.anvisa.gov.br/servicosaude/controle/rede_rm/cursos/
boas_praticas/modulo4/id_sta4.htm, acessado 11/11/2014.
Teste de crescimento em Ágar Manitol – S. aureus cresce bem em ambiente com
alto teor de sal, sendo capaz de fermentar o manitol, produzindo ácido, que pode
ser detectado com o uso de um indicador de pH. O Ágar Manitol Salgado ou Meio
de Chapman possui uma concentração de cloreto de sódio de 7,5%, além disso,
utiliza o Vermelho de Fenol como indicador de pH, que indica uma reação
positiva quando o meio passa de vermelho para amarelo (KONEMAN et al., 2008;
ZURITA, 2010; ANVISA, 2013).

25
Figura 5: Demonstração da prova de Fermentação do Ágar Manitol
Disponível em:http://staff.um.edu.mt/amce1/Stains%20and%20Tests%20PAT2322/
Mannitol%20Salt%20agar.htm, acesso 11/11/2014.
Em relação ao S.epidermidis, esta espécie normalmente é diferenciado das demais
espécies de ECN presentes em amostras clínicas humanas, através da prova da novobiocina, na
qual apresenta sensibilidade à droga (KONEMAN et al., 2008).
2.2 Mecanismo de resistência dos estafilococos aos antimicrobianos
O crescente aumento do número de bactérias cada vez mais resistentes aos
antimicrobianos decorre de múltiplos fatores que incluem; o uso generalizado e por muitas
vezes inadequado de antimicrobianos, uso extensivo como agente promotor de crescimento
animal, uma vez que estes antimicrobianos agem na microbiota intestinal do animal,
diminuindo a competição por nutrientes e reduzindo a produção de metabólitos que deprimem
o crescimento desses animais. Além disso, o crescimento do número de viagens regionais e
internacionais, a aumenta a facilidade com a qual as bactérias resistentes aos antimicrobianos
atravessam as barreiras geográficas (LOWY, 2003; GONZALES et al., 2012).
A resistência aos antimicrobianos pode ocorrer através de dois grandes mecanismos:
mutação num loci cromossomal ou por transferência horizontal de genes, ou seja, por aquisição
de genes de resistência presentes anteriormente em outros microrganismos através de
mecanismos de transdução, transformação ou conjugação (ITO et al., 2003; DZIDIC et al.,
2008).
A resistência de Staphylococcus a quase todas as novas classes de antibióticos é mediada
quase exclusivamente por determinantes adquiridos através de transferência horizontal de

26
DNA. A aquisição de material genético de outro microrganismo oferece várias vantagens ao
Staphylococcus, sendo a principal delas, a possibilidade de ganhar um molde completo de
pacotes de codificação de resistência a múltiplos antibióticos. (PANTOSTI et al., 2007).
Existem diversos mecanismos de resistência encontrados em Staphylococcus, sendo os
mais comuns, a resistência aos macrolídeos, aminoglicosídeos, betalactâmicos e
glicopeptídicos.
A resistência aos macrolídeos e lincosamidas pode ocorrer por dois mecanismos:
1. Modificações no alvo de ligação no ribossomo, que confere resistência cruzada
aos macrolídeos, lincosamidas e estreptogramina B. Esse tipo de resistência é
codificado pelos genes ermA ou ermC.
2. Efluxo ativo, que confere resistência aos macrolídeos e estreptogramina B. É
codificada pelo gene mrsA. (LIVERMORE et al., 2001).
No que se refere aos aminoglicosídeos, o principal mecanismo de resistência encontrado
nos estafilococos é a inativação das drogas por enzimas celulares modificadoras de
aminoglicosídeos. Essas enzimas modificadoras são codificadas por genes presente em diversos
loci distintos e previamente caracterizadas para Staphylococcus spp. As enzimas de maior
importância clínica codificadas por essas regiões são a acetiltransferase (AAC),
adeniltransferase (ANT) e fosfotransferase (APH). Os aminoglicosídeos, modificados em
grupos aminos pela enzima AAC ou em grupos hidroxilas pela enzima ANT ou APH, perdem
a habilidade de se ligar aos ribossomos e assim não inibem mais a síntese protéica das células
bacterianas (NEVES et al., 2007).
Os estafilococos resistentes à meticilina são de difícil tratamento, geralmente por serem
resistentes também a maioria dos outros antibióticos disponíveis (CHAMBERS, 1997). Para
estas infecções, os glicopeptídeos, vancomicina e mais recentemente teicoplanina, são as drogas
de escolha.
Devido ao amplo emprego da vancomicina na prática clínica, surgiu S. aureus com
sensibilidade reduzida tanto à vancomicina, quanto aos demais glicopeptídeos, esses isolados
são denominados S. aureus com sensibilidade intermediaria à vancomicina/glicopeptideos
(VISA/GISA) e S. aureus resistente à vancomicina/glicopeptideos (VRSA/GRSA) (TIWARI;
SEN, 2006).
Acredita-se que aquisição da resistência à vancomicina em estafilococos tenha se
dado devido à transferência conjugativa de plasmídeo contendo o operon vanA em
Enterococcus faecalis. Este operon integra o Transposon Tn1546 composto de vanR, vanS,
vanX, vanY e vanH, presentes em um plasmídeo de 120 Kb (TENOVER et al., 2004).

27
O mecanismo de resistência em linhagens de estafilococos com padrão
intermediário à vancomicina (VISA) inclui a síntese de quantidades adicionais de
peptideoglicano resultando em um espessamento da parede celular. Este espessamento leva ao
aumento no número de resíduos D-alanil-D-alanina, capazes de ligar-se à vancomicina na parte
externa da parede celular, gerando menor disponibilidade do antimicrobiano na molécula alvo
intracelular (BIERBAUM, 2009). Em contrapartida, a resistência em VRSA é causada pela
alteração na parede celular do peptídeo terminal D-alanil-D-alanina da parede celular bacteriana
para D-alanil-D-lactato, impedindo que a vancomicina possa realizar a inibição da síntese de
parede celular. (SCHITO, 2006).
2.2.1 Mecanismo de resistência aos antibióticos betalactâmicos
Nas décadas de 1940 e 1950, a penicilina era o fármaco de escolha para o tratamento das
infecções graves por S. aureus, porém rapidamente surgiu resistência através da aquisição de
elementos genéticos que são capazes de produzir uma enzima designada penicilinase
extracelular (betalactamase classe A), que hidrolisa a ligação amida do anel betalactâmico de
penicilina e ampicilina, obtendo-se um composto inativo. Os plasmídeos que codificam a
produção de penicilinase também transportam outros genes de resistência, tais como a
resistência aos desinfetantes (compostos de amônio quaternário), aos corantes acriflavina e
brometo de etídio ou de metais pesados (como chumbo, mercúrio e cádmio), bem como a outros
antibióticos (eritromicina, ácido fusídico e aminoglicosídeos) (PANTOSTI et al., 2007;
KONEMAN et al., 2008).
No que concerne à resistência aos antibióticos betalactâmicos de 2ª geração, ou penicilinas
semi-sintéticas, surge de mutações nas PBPs que levam à diminuição da afinidade da ligação
do antibiótico ao local de ação Staphylococcus spp. adquiriu o gene cromossômico mecA, que
codifica a PBP2a, que o torna resistente aos betalactâmicos. Esta é uma nova PBP distinta das
restantes presentes neste gênero e que diminui a afinidade pela maioria dos antibióticos
betalactâmicos. Esta proteína confere um elevado padrão de resistência ao Staphylococcus,
provocando inibição de todos os betalactâmicos. Além disso, mantém ativa a síntese de uma
estrutura de peptidoglicano estável, que permite o crescimento e a divisão desses
microrganismos. (HAWKEYet al., 1998; DZIDICet al., 2007; KONEMAN, 2008).
Staphylococcus que expressam o determinante mecA são denominados MRS
(Staphylococcus resistentes à meticilina). A resistência à meticilina e outras penicilinas
resistentes à penicilinase também podem ser observadas em cepas que não possuem o gene

28
mecA. Nessas situações, a resistência se dá por uma hiperprodução de enzimas betalactamases,
que podem hidrolisar as penicilinas semi-sintéticas, causando a resistência (KONEMAN et al.,
2008).
O gene mecA está incluído em um elemento de 30-60kb que é uma ilha genômica de
resistência denominada Staphylococcal Cassete Chromosome mec (SCCmec), que pode conter
também outros genes de resistência a antimicrobianos (LOWY, 2003; PANTOSTI et al., 2007).
Este gene também está presente em estafilococos coagulase negativos resistentes à meticilina e
não está presente em cepas susceptíveis a este antimicrobiano. Acredita-se que este gene tenha
sido adquirido de espécies que não têm relação próxima (ITO; HIRAMATSU, 1998; ENRIGHT
et al., 2002). Segundo o Clinical and Laboratory Standards Institute (CLSI), S.aureus resistentes
à meticilina podem parecer sensíveis in vitro a outros betalactâmicos, combinações de
betalactâmicos e inibidores de betalactamases, carbapenêmicos e cefalosporinas, porém estes
são ineficazes in vivo (CLSI,2013).
Caracteristicamente, SCCmec tem uma localização sítio específica no cromossomo de
estafilococos, dentro de um gene de função desconhecida designada orfX, e contém dois genes
de recombinase (ccrA e ccrB), que medeiam a sítio-específica integração /excisão do elemento
a partir do cromossoma de estafilococos. (PANTOSTI et al., 2007; SHORE; COLEMAN,
2013).
Até o momento, onze diferentes tipos de SCCmec, que podem ser distinguidas com base
na estrutura do complexo mec, dos genes ccr e de elementos adicionais, foram descritos (ITO
et al., 2001; ITO et al.,2003; TAKANO et al., 2007; CONCEIÇÃO et al., 2010; ZONG et al.,
2011). Os SCCmec tipo I, II, III e VI, estão frequentemente associados às infecções
nosocomiais. Os tipos IV, V e VII, no entanto, são mais amplamente disseminados nas cepas
de origem comunitária (HISATA et al., 2005).
Os SCCmec tipos IV e V são elementos genéticos menores e de maior mobilidade que os
outros, pois carregam menos determinantes de resistência que os tipos I, II e III. Por este motivo,
observa-se que os CA-MRSA tendem a ser menos resistentes que os HA-MRSA, mantendo a
sensibilidade a alguns antimicrobianos como, por exemplo, à clindamicina (ITO et al., 2001;
NAIMI et al., 2003; ITO et al., 2004; FRIDKIN et al., 2005; DAUM, 2007).Os SCCmec tipos
I, II e III, por possuírem maior número de determinantes de resistência, possuem como
característica própria a resistência a múltiplos antimicrobianos além dos betalactâmicos, como
os macrolídeos, aminoglicosídeos, tetraciclinas, rifampicina e quinolonas(HIRAMATSU et al.,
1999).

29
O primeiro SCCmec (tipo I) foi isolado em 1961, no Reino Unido, proveniente da cepa
MRSA NCTC10442, o SCCmec tipo II, foi identificado na cepa denominada N315, uma MRSA
isolada no Japão em 1982, já o SCCmec tipo III foi identificado em 1985 na Nova Zelândia, na
cepa MRSA denominada 82/2082. O SCCmec tipo IV foi identificado já nos anos 90, nos
Estados Unidos, em cepas provenientes de infecções comunitárias (DUARTE;
DELENCASTRE, 2002; OKUMA et al., 2002; TURLEJ et al., 2011).
O SCCmec tipo V foi descrito na cepa de origem comunitária denominada WIS
(WBG8318), isolada na Austrália em 1999. Este elemento é estruturalmente semelhante ao
SCCmectipo IV, possuindo apenas o gene mecA como determinante de resistência aos
antimicrobianos (DUARTE; DELENCASTRE, 2002; OKUMA et al.,2002; ITO et al., 2004).
O SCCmec tipo VI foi descrito na cepa HDE288, isolada em Portugal no ano de 1996. Também
é considerado de origem comunitária e, apesar de apresentar similaridade de tamanho com o
SCCmec tipo IV, difere pelo tipo de ccr apresentadora (OLIVEIRA et al.,2006; CONCEIÇÃO
et al., 2010).
O SCCmec tipo VII foi descrito na cepa JCSC6082, de origem comunitária, isolada na
Suécia em 2002 (BERGLUND et al., 2008). Já o SCCmec tipo VIII foi descrito em uma cepa
proveniente de uma epidemia no Canadá no ano de 2003. Os SCCmec tipos IX e X foram
identificados em um estudo realizado na China em cepas provenientes da Tailândia (tipo IX) e
Canada (tipo X), ambas isoladas no ano de 2006. Além da resistência aos antimicrobianos, esses
SCCmec carregam elementos relacionados a desintoxicação de metais pesados como o cádmio,
o cobre, o zinco e o arsenato (LI et al., 2011; TURLEJ et al., 2011). O SCCmec tipo XI foi
identificado pelo Instituto Sanger na cepa bovina de MRSA LGA251 (SHORE et al., 2011).
Os elementos mais recentemente descritos SCCmec, IX, X e XI compartilham
características que fornecem compreensão sobre a possível origem doSCCmec. Em primeiro
lugar, todos os três elementos têm sido encontrados em cepas de MRSA em seres humanos,
mas são considerados como sendo de origem animal. Em segundo lugar, cada um destes
elementos possui pelo menos um operon que codifica resistência a metais pesados tais como
arsênio e cobre, o que parece ser um atributo dos elementos de SCCmec originários de animais.
Além disso, a significante divergência de sequência evidenciada no SCCmec XI sugere que ele
pode ter evoluído separadamente em clones de MRSA humanos e pode ter se originado em
outras espécies de estafilococos ou outros gêneros. Por exemplo, o complexo mec do SCCmec
XI compartilha a sua organização genética como de um complexo mec identificado em um
transposon integrado numa orfX em Macrococcus caseolyticus, e pode representar uma forma
ancestral do complexo mec em MRSA. Outras características incomuns de SCCmec IX e X

30
incluem o tipo ea localização de seus complexos ccr ea orientação da transcrição do mecA no
SCCmec X (LI et al., 2011; SHORE; COLEMAN, 2013).
O elemento SCC é considerado conferir melhor capacidade de sobrevivência e aumento
da virulência da cepa. Visto isto, parece provável que a SCC serve não só como um veículo
para os genes de resistência a drogas, mas também diversos genes úteis para uma melhor
adaptação ao ambiente hostil. Desse modo, a acumulação de vários elementos SCC
transportando várias funções é alcançada para permitir a diversificação do genoma de S. aureus
(HIRAMATSU et al.,2013).
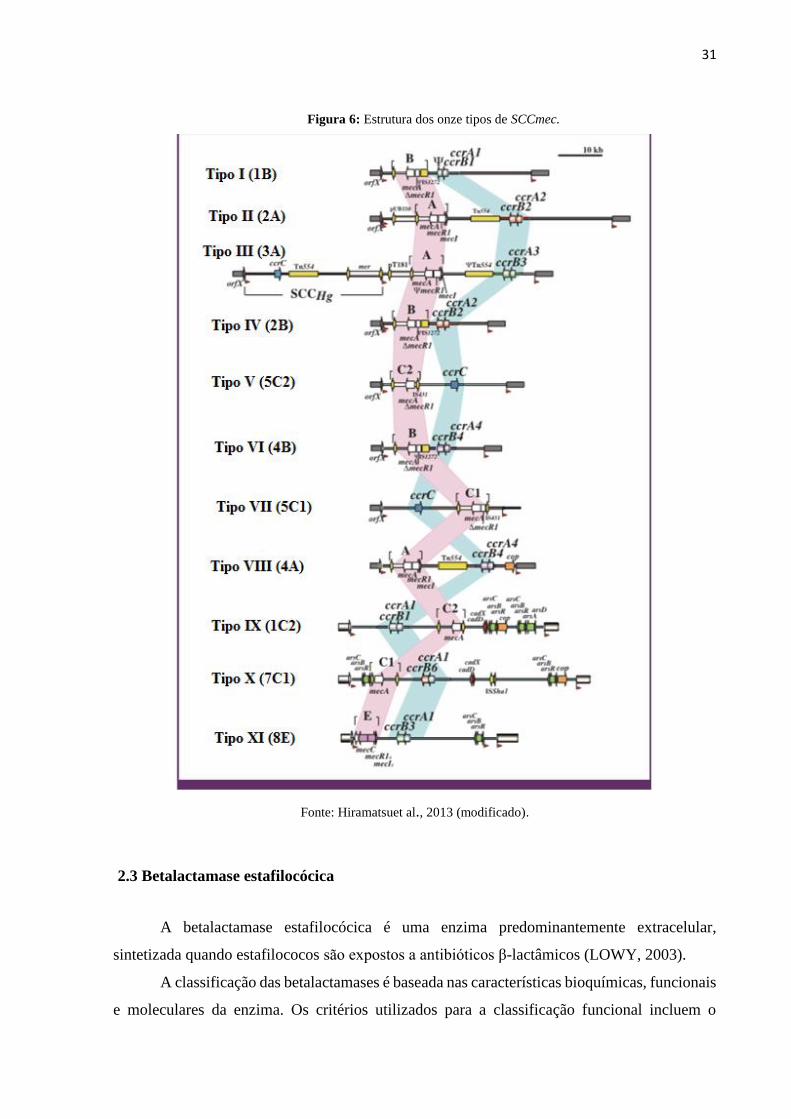
31
Figura 6: Estrutura dos onze tipos de SCCmec.
Fonte: Hiramatsuet al., 2013 (modificado).
2.3 Betalactamase estafilocócica
A betalactamase estafilocócica é uma enzima predominantemente extracelular,
sintetizada quando estafilococos são expostos a antibióticos β-lactâmicos (LOWY, 2003).
A classificação das betalactamases é baseada nas características bioquímicas, funcionais
e moleculares da enzima. Os critérios utilizados para a classificação funcional incluem o

32
substrato antimicrobiano, o perfil de inibição enzimática, a taxa de hidrólise (V max), a
afinidade de ligação (Km), o peso molecular da proteína e a composição de aminoácidos
(BUSH et al., 1995).
Segundo a classificação de Bush, Jacob e Medeiros, as betalactamases estafilocócicas
pertencem ao grupo 2a (classe A de Ambler), porque hidrolisam as penicilinas mais rápido que
cefalosporinas (VOLADRI; KERNODLE, 1998).
A betalactamase estafilocócica é codificada pelo gene estrutural blaZ, que é regulada
por uma proteína de ligação a DNA BlaI e um transdutor de sinal BlaR1 (HAO et al.,2012).
BlaRI é uma proteína de alto peso molecular que se liga à penicilina e é constituída de
dois domínios (GHUYSEN, 1994; BIRCK et al., 2004). O primeiro é um domínio N-terminal
intracelular, com atividade de metaloprotease dependente de zinco, de aproximadamente
38KDa com uma α-hélice atravessando amembrana quatro vezes, via quatro segmentos
transmembrana (TM1, TM2, TM3 e TM4) (HARDT et al., 1997; FREDERICK et al., 2013).
Os segmentos transmembrana são conectados entre si por três alças (L1, L2 e L3), onde L1 e
L3 são direcionadas ao citoplasma e L2 é posicionada fora da célula. O segundo, é um domínio
C-terminal de aproximadamente 27 KDa é constituído por um sensor que se localiza na
superfície da membrana (Figura 7) (KERFF et al., 2003; BIRCK et al., 2004; FREDERICK et
al., 2013).
Quatro tipos de produto blaZ (A, B, C, D) foram distinguidos por sorotipagem e
diferenças na hidrólise em substratos selecionados de betalactâmicos. Os tipos A, C e D são
geralmente localizados em plasmídeos, enquanto que o tipo B é normalmente encontrado no
cromossoma (OLSEN et al., 2006).
As betalactamases agem de duas diferentes formas na destruição do anel β-lactâmico: o
primeiro mecanismo, representado pelas classes A, C e D, baseia-se na ação catalítica de um
resíduo de serina ativo presente nas β-lactamases. O resíduo serina reage irreversivelmente com
o grupo carbonil do anel β-lactâmico, o que resulta em seu rompimento e inativação (KOTRA
et al., 2002; STRATTON, 2000). O segundo mecanismo, representado pela classe B, é
constituído pelas metalo betalactamases, que requerem um íon metálico divalente, geralmente
o zinco, ligado ao resíduo de histidina, cisteína ou ambos, para reagir com o grupo carbonil da
ligação amida de diversas penicilinas, cefalosporinas e carbapenêmicos (PAGE, 2002).

33
Figura7: Arranjo dos domínios da proteína BlaR1
Fonte: Frederick, 2013
Na ausência de betalactâmicos, o homodímero de BlaI se liga à região de operador blaZ
e blaR1-blaI para reprimir a transcrição. Na presença de betalactâmicos, o domínio extracelular
do sensor de BlaR1 pode ser acilado, de modo que o sinal possa ser transduzido pelo domínio
de membrana, atravessando a membrana para ativar o domínio intracelular da metaloprotease
de zinco. O BlaR1 ativado atua como uma protease e perturba ou alteraa formação do dímero
de BlaI. BlaI pode, assim, unir direta ou indiretamente do seu sítio de ligação. Sem BlaI ligado
ao local do operador, a transcrição de ambos blaZ e blaR1-blaI pode iniciar pela síntese de
betalactamase. Um produto de gene adicional, BlaR2, também pode regular a síntese de
betalactamase, apesar de o papel desta proteína ainda não ter sido elucidado (Figura 8)
(STAPLETON; TAYLOR, 2002;HAO et al.,2012).

34
Figura 8: Sistema de regulação de controle da expressão de PBP2a e betalactamase.
Fonte: Hao, 2012 (modificado).
A organização do operon bla é semelhante à encontrada no operon mec, responsável
pela síntese de PBP2a. Em muitas cepas MRSA o gene mecA é induzido pelo sistema regulador
blaR1-blaI. BlaI reprime a transcrição do gene mecA e esta repressão é diminuída na presença
de betalactâmicos. Também tem sido demonstrado que o mecI reprime a síntese de
betalactamases (CLARKE; DYKE, 2001).
Em seus estudos, Hou et al. (2007) demonstraram que o bloqueio da via de
sensores/transdutores de sinal podem representar uma nova abordagem para inverter a
resistência aos antibióticos em MRSA. Este fato foi demonstrado a partir da ideia que quando
um antibiótico betalactâmico se liga ao domínio extracelular do sensor de BlaR1, uma mudança
conformacional dentro BlaR1 conduz à ativação autocatalítica do domínio metaloprotease
integrante-membrana. A protease ativavoltada para o citosol cliva especificamente BlaI, direta
ou indiretamente, que posteriormente inicia expressão blaZ. Este sistema destaca o papel
fundamental da meticilina como o eventual regulador sensor/transdutor transcricional de
resposta MRSA.

35
2.4 Staphylococcus spp. como causador de infecções
S. aureus é frequentemente reconhecido como um patógeno que pode estar presente
intermitente ou permanentemente na microbiota de narinas, pele, garganta e trato intestinal de
alguns indivíduos. Estima-se que 30% da população tenham suas narinas colonizadas de forma
intermitente e cerca de 20% de forma persistente. Esta colonização permite que esses sujeitos
atuem como disseminadores deste microrganismo. A maioria dos portadores de S. aureus é
assintomático e o processo de infecção normalmente se dá devido a algum fator que diminua a
resposta imunológica do indivíduo (GORDON; LOWY,2008; COSTA et al., 2011).
Este patógeno pode ser encontrado colonizando a pele úmida em diferentes regiões
como nariz, períneo e axilas, o que aliado a fatores de virulência determina sua patogenicidade
em infecções (CORBELLA et al., 1997; GORDON; LOWY, 2008).
S. aureus é o agente mais comum de infecções piogênicas. Estas infecções podem se
localizar na pele ou em regiões mais profundas. As infecções cutâneas primárias causadas pelos
estafilococos são chamadas genericamente de piodermites. As piodermites tem contagiosidade
variável e recebem as designações de acordo com a localização e outras características. O
impetigo é uma piodermite superficial que acomete mais crianças que adultos e tem alto grau
de contágio (Figura 9). A foliculite é a inflamação de um folículo piloso e surge em decorrência
de sua obstrução (ocorre apenas na porção superior do folículo). O furúnculo ou abscesso
envolve a inflamação de todo o folículo piloso e de sua glândula sebácea anexa, com
envolvimento do tecido celular subcutâneo. O hordéolo, ou terçol, é a infecção de uma glândula
sebácea marginal das pálpebras (CORBELLA et al., 1997; SANTOS et al., 2007; COSTA et
al., 2011).
36
Figura 9: Lesões do impetigo
Disponível em:http://www.dermis.net/dermisroot/en/27034/image.htm, acesso 17/11/2014.
Em indivíduos debilitados por doenças crônicas, traumas físicos, queimaduras ou
imunossupressão, esse microrganismo pode causar infecções de caráter mais grave. Entre as
infecções profundas destacam-se a osteomielite, a bacteremia (frequentemente associada a
abcessos metastáticos), a endocardite, a pneumonia e, ocasionalmente, a meningite e a artrite
bacteriana (FLOCK et al., 1996; COELLO et al.,1997). A osteomielite pode ser primária, ou
hematogênica, e secundária. No primeiro caso, a doença surge em consequência da
disseminação do S. aureus a partir do foco de infecção. Já a secundária tem decorrência de
traumas penetrantes, processos cirúrgicos ou da presença de um foco de infecção contíguo. A
bacteremia pode ter origem de qualquer foco localizado de infecção por estafilococos
(TORTORA et al., 2012).
Bacteremias prolongadas podem determinar o aparecimento de endocardites, com o
envolvimento das válvulas cardíacas, particularmente a aórtica. As endocardites também
podem surgir em consequência de próteses e processos cirúrgicos. A pneumonia é mais
frequente no primeiro ano de vida, sendo as viroses fatores predisponentes à pneumonia por
esse microrganismo (TORTORA et al., 2012).
Além dessas infecções, o S. aureus pode causar vários tipos de intoxicações, seja na
vigência de um processo infeccioso ou não. A primeira possibilidade pode ser exemplificada
pela síndrome da pele escaldada ou doença de Ritter e é caracterizada pelo deslocamento de
extensas áreas de epiderme (Figura 10), sendo determinado por uma toxina, denominada
esfoliatina, é produzida na área da infecção e levada para áreas distantes pela corrente sanguínea
(TRABULSI et al., 2008; TORTORA et al., 2012).
37
Figura 10:Lesões da Síndrome da pele escaldada
Fonte: Tortora et al., 2012.
Na ausência de processos infecciosos, S. aureus pode causar dois tipos de intoxicações:
a intoxicação alimentar e a Síndrome do Choque Tóxico. A primeira ocorre devido à ingestão
de toxinas previamente formadas no alimento pelo S. aureus. Essas toxinas são denominadas
enterotoxinas. Essas enterotoxinas são superantígenos, e são termoestáveis, desta forma, é
possível a intoxicação alimentar através de alimentos cozidos. Já na síndrome do choque tóxico,
o paciente apresenta febre alta, diminuição da pressão sistólica, eritema com descamação da
pele, insuficiência renal, entre outras manifestações clínicas. As manifestações são atribuídas à
produção da toxina TSST-1. Embora a grande maioria dos doentes seja constituída por mulheres
em período menstrual, sem sinais de infecção estafilocócica, a doença tem sido registrada em
pacientes com infecções de pele, ossos e pulmões (DINGES et al., 2000).
2.5 Epidemiologia das infecções causadas por Staphylococcus spp. resistente à meticilina
Na era pré-antibiótica, a mortalidade de pacientes infectados com S. aureus era superior
a 80%, e, mais de 70% destes desenvolviam infecções metastáticas. A introdução da penicilina
na prática clínica em 1941 revolucionou o tratamento das infecções causadas por bactérias
gram-positivas, principalmente estafilococos e estreptococos, o que melhorou tal prognóstico,
com mais de 94% das cepas exibindo sensibilidade (FUDA et al., 2005; WOODFORD;
LIVERMORE, 2009; MOELLERING, 2012).
Este resultado, no entanto, teve curta duração. O uso da penicilina rapidamente
selecionou cepas que eram resistentes ao antibiótico, como resultado da expressão das
betalactamases. Em 1942 emergiu o primeiro isolado de S. aureus resistente à penicilina, com
proliferação inicial dentro do ambiente hospitalar e após na comunidade. Na década de 50, cerca

38
de 50% dos estafilococos isolados apresentavam resistência à penicilina (DEURENBERG;
STOBBERINGH; 2008; WOODFORD; LIVERMORE, 2009).
Como resposta às altas taxas de resistência à penicilina, surge em 1959 a meticilina,
uma penicilina semi-sintética. Apenas dois anos depois, em 1961, ocorreram os primeiros
relatos de cepas isoladas de S.aureus resistentes à meticilina (MRSA), no Reino Unido, tendo
se espalhado rapidamente para outros países europeus, onde ocorreram surtos de infecções
hospitalares provocados por este microrganismo, ocorrendo também, relatos de MRSA isolados
no Japão, Austrália e Estados Unidos (ENRIGHTetal.,2002; FUDA et al., 2005; WOODFORD;
LIVERMORE, 2009).
Em diversos países, o MRSA tornou-se um patógeno nosocomial endêmico. Estas cepas
MRSA ficaram restritas ao ambiente hospitalar por décadas, causando infecções apenas em
indivíduos que apresentavam os fatores de risco clássicos para infecções associadas aos
cuidados em saúde, tais como, pacientes em pós-operatório, com doenças crônicas, com
histórico de internação hospitalar, ou ainda, portadores de próteses ou dispositivos (MEJIA et
al. 2010).
Desde o final dos anos 1980 e início dos anos 1990 ocorre vigilância regular para MRSA
em hospitais nos EUA e Europa. Mesmo com o intenso programa de vigilância, o CDC estima
que 63% dos isolados de estafilococos isolados nos EUA sejam de MRSA. Em 1974 esses
números mostravam apenas 2% dos isolados, enquanto em 1995, essa taxa subiu para 22%. Em
contrapartida, países com rígidas práticas de controle de infecção, como a Holanda e países
escandinavos, vêm mantendo baixos índices de infecção por MRSA, mesmo em hospitais
(MEJIA et al., 2010).
Entretanto, ao final da década de 90, em 1998, foi documentada na Austrália uma nova
variedade do MRSA, o clone USA300, um MRSA associado à comunidade (CA-MRSA).
Desde então, infecções por CA-MRSA têm sido descritas no mundo inteiro, ocorrendo aumento
significativo no número de casos na última década. (CORREAL et al., 2013).
Infecções por CA-MRSA são definidas como infecções que ocorrem em pacientes não
hospitalizados ou que se inicie dentro das primeiras 48 (quarenta e oito) horas de internação em
pacientes sem nenhum dos seguintes fatores de risco usados para definir infecção por HA-
MRSA: hospitalização ou cirurgia recente, terapia antibiótica prolongada, doença crônica de
base, cateter venoso central ou outros dispositivos, em cuidados de longa permanência ou
institucionalizados (CDC, 2011).
As cepas CA-MRSA diferem em alguns aspectos das amostras HA-MRSA. Cepas CA-
MRSA são sensíveis a uma variedade significativa de antibióticos não betalactâmicos, enquanto

39
as cepas HA-MRSA são tipicamente multirresistentes. Observa-se ainda que cerva de 90% dos
CA-MRSA produzem uma toxina denominada Toxina de Panton Valentine (PVL), capaz de
destruir leucócitos humanos e causar dano tecidual grave (REMONATTO et al., 2007; MEJIA
et al., 2010; CORREAL et al., 2013).
A disseminação das cepas CA-MRSA, bem com seu caráter epidêmico, estão
intimamente relacionados com as mudanças ocorridas na prestação de serviços saúde,
principalmente em programas ambulatoriais para o tratamento de doenças crônicas, tais como,
hemodiálise, diálise peritoneal, programas de hospital dia e asilos de longa permanência, etc.
Esses programas favorecem a disseminação de clones na comunidade. Outro fator relacionado
à disseminação dos clones CA-MRSA é o tipo de SCCmec encontrado. O menor tamanho dos
SCCmec tipos IV a XI, sugerem uma maior facilidade de transmissão entre estes se comparados
com grandes SCCmec tipos I a III (DEURENBERG; STOBBERINGH; 2008; CORREAL et
al., 2013).
No Brasil, registram-se elevadas frequências de isolamento de S. aureus, bem como há
grande relação deste com as infecções hospitalares. Vários estudos têm demonstrado
prevalência de 40 a 80% de isolamento de cepas MRSA em hospitais brasileiros (OLIVEIRA
et al., 2001; TRINDADE et al., 2005; SANTOS et al., 2010). Verifica-se ainda que dados do
Programa de Vigilância a Antimicrobianos apontam que MRSA corresponde a 31% da causa
de infecções nosocomiais e comunitárias, sendo considerado o mais comum dentre os
patógenos mais prevalentes (GALES et al., 2009). O primeiro relato de doença causada por
CA-MRSA no país ocorreu em 2008, por uma amostra de SCCmec tipo IV e PVL positivo
(ROZENBAUM et al, 2009).

40
Figura11: Cronologia da resistência a antibióticos dos S. aureus.
Fonte: Kim, 2009 (modificado)
2.6 Importância dos Staphylococcus resistentes à meticilina
Staphylococcus resistentes à meticilina (MRS) causam infecções de difícil tratamento,
porque estes são resistentes à maioria dos antibióticos utilizados na prática clínica, como os
betalactâmicos, aminoglicosídeos e macrolídeos. (DURANet al., 2012)
S. aureus isolados de unidades de terapia intensiva em todo o país e de culturas de
sangue em todo o mundo estão cada vez mais resistentes a um maior número de agentes
antimicrobianos. Inevitavelmente, isto deixou antibióticos bactericidas menos eficazes para o
tratamento destas infecções, aumentando em muitas vezes o risco de vida dos pacientes. Tão
rapidamente quanto novos antibióticos são introduzidos, estafilococos vêm desenvolvendo
mecanismos eficientes para neutralizá-los (LOWY, 2003).
As infecções por S. aureus resistentes à meticilina (MRSA) atualmente, podem ser
consideradas pandêmicas em todo o mundo. Nos Estados Unidos, por exemplo, a Rede
Nacional de Cuidados em Saúde e Segurança estima que aproximadamente dois milhões de
pacientes sejam internados por adquirirem infecções associadas aos cuidados em saúde a cada
ano, dentre estes as infecções por MRSA são responsáveis por grande porcentagem de
internações. Há aproximadamente 19.000 mortes/ano nos Estados Unidos devido a MRSA,
superando aquelas devido a HIV-AIDS. Com custo estimado em bilhões de dólares, o peso
econômico da infecção é impressionante (HARBARTH et al., 2011; MARLOWE;
BANKOWSKI, 2011).
S. aureus tem a notável capacidade de se adaptar rapidamente às mudanças de ambientes
de hospitais e outras instituições de saúde, superando a atividade antibiótica com uma variedade
de mecanismos de resistência diferentes. Embora a resistência à meticilina ainda seja a principal

41
característica de resistência desta espécie, a resistência à vancomicina está emergindo de forma
preocupante (PANTOSTI et al., 2007).
Recentemente, a comunidade tornou-se um reservatório para novos clones virulentos de
CA-MRSA (S. aureus resistente à meticilina de origem comunitária) que estão sendo
introduzidos em hospitais. Embora as estirpes do CA-MRSA sejam normalmente susceptíveis
aos antibióticos não betalactâmicos, parecem estar adquirindo resistência a outros antibióticos.
MRSA, e CA-MRSA em particular, parecem ser dotados com maior virulência, devido à
presença de um número de diferentes toxinas e outros genes de virulência. É essencial o
monitoramento do desenvolvimento de novos padrões de resistência a antibióticos em S. aureus
para os antigos e novos antibióticos (PANTOSTI et al., 2007).
O diagnóstico rápido e preciso dos genes de resistência a antibióticos no tratamento de
infecções estafilocócicas é importante na prevenção da propagação de infecções. Métodos
moleculares baseados em PCR são muitas vezes preferidos para a determinação dos genes de
resistência a antibióticos por serem mais rápidos e específicos que os métodos tradicionais.
Desta forma, métodos rápidos e confiáveis para a detecção da resistência aos antibióticos são
importantes para instituir a terapêutica adequada nos serviços de saúde (DURAN et al., 2012).

42
3. Objetivos

43
3.1 Objetivo Geral
Verificar se existe diferença na frequência da ocorrência do gene blaZ que codifica as
betalactamase em isolados clínicos de Staphylococcus spp. resistentes à meticilina portadores
do gene mecA, quando comparado aos isolados não portadores do gene mecA, provenientes de
hospital universitário da cidade do Recife-PE no período de abril de 2011 a julho de 2013.
3.2 Objetivos Específicos
Selecionar isolados fenotipicamente resistentes à meticilina;
Detectar a produção fenotípica de betalactamase através dos testes de Cefinase e Clover-
Leaf e relacionar com os achados da PCR do gene blaZ;
Detectar a ocorrência do gene blaZ nos isolados clínicos de Staphylococcus spp.
resistentes à meticilina provenientes de hospital universitário da cidade do Recife-PE
no período de abril de 2011 a julho de 2013;
Comparar a frequência do gene blaZ nos isolados clínicos de Staphylococcus spp.
resistentes à meticilina portadores do gene mecA e dos isolados não portadores do gene
mecA provenientes de hospital universitário da cidade do Recife-PE no período de abril
de 2011 a julho de 2013.

44
4. Materiais e Métodos

45
4.1 Desenho do estudo
Foi realizado um estudo experimental de base laboratorial sobre a ocorrência do gene
da betalactamase em isolados clínicos de Staphylococcus spp. resistentes à meticilina
portadores do gene mecA e dos isolados não portadores do gene mecA provenientes de hospital
universitário da cidade do Recife-PE no período de abril de 2011 a julho de 2013.
Este tipo de estudo permite verificar a presença do gene que codifica a betalactamase
estafilocócica (blaZ), além de comparar as frequências desses genes nos isolados clínicos de
Staphylococcus spp. resistentes à meticilina portadores do gene mecA e dos isolados não
portadores do gene mecA.Além disso, foi possível verificar a produção da betalactamase nos
isolados em estudo através de testes fenotípicos de detecção.
4.2 Local do Estudo
O estudo foi realizado com isolados de Staphylococcus spp. da bacterioteca do
Laboratório de Bacteriologia da Disciplina de Microbiologia e Imunologia do Departamento de
Medicina Tropical-UFPE que foram provenientes de pacientes internados no período de abril
de 2011 a julho de 2013 no Hospital das Clínicas vinculado a UFPE (HC-UFPE).
As amostras foram processadas no setor de Bacteriologia do hospital e transportadas em
ágar Casoy para o Laboratório de Bacteriologia da Disciplina de Microbiologia e Imunologia
do Departamento de Medicina Tropical-UFPE, onde foram feitos testes confirmatórios de
identificação. Também foi utilizado o Laboratório do Departamento de Microbiologia do
Centro de Pesquisas Aggeu Magalhães, para suporte nos procedimentos de biologia molecular.
4.3 Categorização das Variáveis
4.3.1 Variável Dependente

46
Presença do gene mecA(gene que codifica a resistência à meticilina):definida como a
identificação do fragmento de 1.046pbatravés de reação de PCR descrita por PETINAKI et al.
(2001).
Categorizado como 1-Presente ou 2-Ausente.
4.3.2 Variáveis Independentes
4.3.2.1 Variáveis Fenotípicas
Teste da cefinase em disco: definida como um teste fenotípico para detecção de
betalactamase caracterizado pela mudança da coloração nos discos de cefinase (BBL BD) após
esfregaço de cultura nos mesmos.
Categorizado como 1-Presente ou 2-Ausente.
Teste de Clover-Leaf: definida como um teste fenotípico para detecção de betalactamase
caracterizado através da verificação do aparecimento de uma distorção no halo de inibição do
disco de penicilina.
Categorizado como 1-Presente ou 2-Ausente.
4.3.2.2 Variável Genética
Presença do gene blaZ (gene estrutural da betalactamase): definida como a identificação
do fragmento de 533pbatravés de reação de PCR descrita por MILHEIRIÇO et al. (2011).
Categorizado como 1-Presente ou 2-Ausente
4.4 Operacionalização da Pesquisa
4.4.1 Obtenção de Amostras
Foram utilizadas 47 (quarenta e sete) isolados clínicos provenientes do Hospital das
Clínicas da UFPE, coletadas durante o período de abril de 2011 a julho de 2013 de pacientes
internados nesta unidade de saúde. Todos os isolados estão estocados na Bacterioteca do
Laboratório de Bacteriologia do Departamento de Medicina Tropical-UFPE.
Tais isolados foram fornecidos pelos responsáveis dos laboratórios de bacteriologia do
referido hospital, após identificação bacteriana. Desta forma, a coleta da amostra foi de
responsabilidade do hospital.

47
4.4.2 Seleção dos isolados
A resistência à meticilina nos testes fenotípicos foi utilizada como critério de inclusão
no estudo. Todos os isolados já haviam sido previamente identificados e submetidos à PCR do
gene mecA antes de serem armazenados na Bacterioteca do Laboratório de Bacteriologia do
Departamento de Medicina Tropical-UFPE.
4.4.2.1 Testes fenotípicos
Todos os isolados foram previamente identificados como pertencentes ao gênero
Staphylococcus de acordo com os procedimentos padrões. Também foram submetidos aos
testes de difusão em meio sólido utilizando discos de oxacilina e cefoxitina e screening de
oxacilina. Aqueles isolados que se apresentaram resistentes a pelo menos uma das técnicas
utilizada, foram submetidos à PCR para detecção do gene mecA.
4.4.2.2 Identificação do gene mecA através de PCR
Para a amplificação foram utilizados os primers P1, 5'-GGTCCCATTAACTCTGAAG-
3’ e P3, 5’-AGTTCTGCAGTACCGGATTTGC-3’ descritos por PETINAKI, et al. (2001) que
resultam em fragmentos de 1.046pb. As reações de PCR foram preparadas individualmente em
um volume final de 25μL em cada tubo contendo: 20ng de DNA genômico (extraído de acordo
com a técnica de FREITAS et al., 2008), 20 pmol de cada primer, MgCl21,5 mM, dNTP 200
μM, 1U de Go Taq DNA polimerase (Promega, Brasil) e 5μL do tampão Green Go Taq DNA
polimerase (Promega, Brasil). As reações foram realizadas em termociclador (Biometra),
programado inicialmente para 35 ciclos térmicos, com desnaturação de um minuto a 94°C,
anelamento de um minuto a 55°C e uma extensão de dois minutos a 72°C seguido de uma etapa
de alongamento final de 15 minutos a 72°C. Foi utilizado como controle negativo, um tubo
contendo todos os componentes da mistura e sem a presença do DNA molde. Para controle
positivo foi utilizado a cepa Staphylococcus aureus ATCC 33591, a qual é resistente.
O produto da amplificação foi submetido à eletroforese em gel de agarose 1% e corado
com brometo de etídio (2μg/mL), visualizado em transluminador UV e digitalizado através do
software Kodak 1D versão 3.5.2 (Scientific Imaging Systems, USA). O marcador de peso
molecular é de 100pb (Invitrogen).
4.4.3 Identificação fenotípica da produção de betalactamase

48
4.4.3.1 Teste da Cefinase em disco
O teste de cefinase em disco foi realizado utilizando o BBL Cefinase Discs (BD) de
acordo com as especificações do fabricante. Os discos foram distribuídos em uma placa de Petri
esterilizada. Em seguida, cada disco foi umedecido com uma gota (aprox. 30µL) de água miliQ
esterilizada e, com uma alça de platina, foi realizado um esfregaço das colônias em teste no
disco de cefinase. O resultado positivo foi detectado pela mudança de cor, de amarelada para
rosa, apresentada pelo disco. Esta mudança de cor deve-se ao composto Nitrocefin, uma
cefalosporina cromógena, que muda de cor à medida que a ligação amido no anel beta-lactâmico
é hidrolisada por uma β-lactamase. Para Staphylococcus spp. esperou-se até uma hora para que
ocorresse a mudança de coloração.
Como controle positivo foi utilizada a cepa MRSA ATCC 33591 e como controle
negativo foi utilizada a cepa de MSSA ATCC 6538.
4.4.3.2 Teste de Clover-Leaf
O teste de Clover-Leaf consiste num teste de Hodge modificado e foi realizado de
acordo com a técnica de Bergan et al. (1997). Primeiramente preparou-se um inóculo de um
isolado betalactamase negativo (cepa ATCC 25923), de acordo com o 0,5 da escala de
McFarland através de suspensão direta. Em seguida foi realizado o semeio de forma a obter um
crescimento confluente deste isolado em Ágar Mueller-Hinton e colocado um disco de
penicilina 10U (Laborclin) no centro da placa. Com uma alça de platina, foram realizadas estrias
do centro até a periferia da placa de Petri, sem tocar o disco de penicilina. A placa foi incubada
de 18h a 24h a 35ºC e após foi observado o aparecimento da zona de distorção no halo de
inibição dos isolados considerados positivos para a produção da betalactamase.
Como controle positivo do teste, foi utilizado o isolado de Staphylococcus aureus 1171,
produtor de betalactamase, cedido pelo Dr. Wagner Oliveira. Como controle negativo foi
utilizada a cepa MSSA ATCC 25923.
4.4.4 Extração de DNA total
Para a extração do DNA das amostras de Staphylococcus foi utilizada a técnica de
extração por fervura, na qual o DNA é extraído a partir de 1mL de cultura bacteriana
previamente crescida em caldo BHI a 37ºC por 24h e centrifugada a 14.000rpm/4ºC durante

49
cinco minutos. Em seguida, desprezou-se o sobrenadante e o sedimento foi ressuspenso em
200µL de água MiliQ estéril. A suspensão foi incubada a 100ºC por 10 min seguido de
centrifugação a 14.000rpm/4ºC durante cinco minutos. Após centrifugação, 100µL do
sobrenadante foi coletado em novo tubo e estocado a -20ºC.
Uma alíquota de 1 ou 2µL do DNA extraído foi submetida à eletroforese em gel de
agarose 1% em tampão TBE (Tris-borato 0,089M; ácido bórico 0,089M; EDTA 0,002) à
temperatura ambiente sob corrente constante. Após migração, o DNA foi corado e observado
em transiluminador de luz ultravioleta (UV) e registrado em um sistema de foto documentação
Kodak 1D versão3.5.2 (Scientific Imaging Systems, USA).
4.4.5 Identificação do gene da betalactamase por PCR
Para a amplificação do gene blaZ foram utilizados os primers F1, 5'-
GATAAGAGATTTGCCTATGC-3' e R1, 5'-GCATATGTTATTGCTTGACC-3'
(MILHEIRIÇO et al., 2011) que resultam em fragmentos de 533pb. As reações de PCR foram
preparadas individualmente em um volume final de 25µL em cada tubo contendo: 5µl de DNA
genômico, 20pmol de cada primer, MgCl2 1,5mM, dNTP 200µM, 1U de Go Taq DNA
polimerase (Promega, Brasil) e 5µL do tampão Green Go Taq DNA polimerase (Promega,
Brasil). As reações foram realizadas em termociclador (Biometra), programado inicialmente
para desnaturação inicial de quatro minutos a 92ºC, em seguida 30 ciclos térmicos, com
desnaturação de um minuto a 92°C, anelamento de um minuto a 55°C e uma extensão de um
minuto a 72°C seguido de uma etapa de alongamento final de sete minutos a 72°C. Foi utilizado
como controle negativo, um tubo contendo todos os componentes da mistura e ausência de
DNA molde. Para controle positivo foi utilizada uma cepa de S. aureus resistente à meticilina
(código do isolado: 1171) cedida pelo Dr. Wagner Oliveira.
O produto da amplificação foi submetido à eletroforese em gel de agarose 1% e corado
com brometo de etídio (2µg/mL), visualizado em transluminador UV e digitalizado através do
software Kodak 1D versão 3.5.2 (Scientific Imaging Systems, USA). O marcador de peso
molecular é de 100bp (Invitrogen).
4.5 Considerações éticas
Este estudo fez parte do projeto CAAE – 0490.0.172.000-11 Registro CEP/CCS/UFPE
nº 009/11 (Anexo III) submetido à análise pelo Comitê de Ética em Pesquisa (CEP) do Centro

50
de Ciências da Saúde (CCS), da UFPE, com a permissão e colaboração da Comissão de
Controle de Infecção Hospitalar (CCIH) do hospital envolvido.
Não houve Termo de Consentimento Livre e Esclarecido (TCLE) para os pacientes por
se tratar de amostras secundárias.
Foi obtida Carta de Anuência no hospital em estudo, autorizando o acesso aos
prontuários clínicos. Para o fornecimento dos isolados de Staphylococcus spp. Identificados no
período da pesquisa, também foi obtida carta de anuência do laboratório de bacteriologia do
Hospital das Clínicas da UFPE.
4.6 Análise Estatística
Os dados microbiológicos foram analisados estatisticamente através da distribuição de
frequências e Teste χ², utilizando o software EpiInfo versão 6.04.

51
5. Resultados

52
DETECÇÃO DE BETALACTAMASE EM Staphylococcus spp. ISOLADOS DE
AMOSTRAS CLÍNICAS PROVENIENTES DE HOSPITAL UNIVERSITÁRIO DE
RECIFE-PERNAMBUCO-BRASIL
Introdução
A resistência à meticilina, que teve os primeiros relatos descritos na década de 1960,
conferiu maior importância ao gênero Staphlylococcus como um dos principais patógenos
humanos [15, 27]. Staphylococcus resistente à meticilina (MRS) causa infecções de difícil
tratamento, porque estes são resistentes à maioria dos antibióticos utilizados na prática clínica,
como os betalactâmicos, aminoglicosídeos e macrolídeos [4].
Isolados clínicos de S. aureus provenientes de pacientes de unidades de terapia intensiva
em todo o país e de hemoculturas isoladas em todo o mundo descrevem que estas linhagens
bacterianas estão cada vez mais resistentes a um maior número de agentes antimicrobianos.
Inevitavelmente, isto deixou antibióticos bactericidas menos eficazes para o tratamento destas
infecções, aumentando em muitas vezes o risco de vida dos pacientes. Tão rapidamente quanto
novos antibióticos são introduzidos, estafilococos vêm desenvolvendo mecanismos eficientes
para neutralizá-los [12].
Os estafilococos desenvolveram a resistência aos β-lactâmicos através de dois
mecanismos, a expressão das β-lactamases e a produção de PBP2a, codificadas pelos genes
blaZ e mecA, respectivamente [7, 14]. A detecção do gene mecA por técnicas moleculares vem
sendo considerado o padrão ouro para a identificação das cepas de Staphlylococcus resistentes
à meticilina (MRS) [24, 26], porém em alguns estudos verifica-se discrepância entre a
apresentação fenotípica dessas cepas e a presença do gene mecA [1, 3, 28].
A betalactamase estafilocócica é codificada pelo gene estrutural blaZ, que é regulada
por uma proteína de ligação ao DNA, BlaI, e um transdutor de sinal BlaR1 [7]. Na presença de
antibióticos betalactâmicos, o domínio extracelular do sensor de BlaR1 pode ser acilado, de
modo que o sinal possa ser transduzido pelo domínio de membrana, atravessando a mesma para
ativar o domínio intracelular. O BlaR1 ativado atua como uma protease e perturba ou altera a
formação do dímero de BlaI. Sem BlaI ligado ao local do operador, a transcrição de ambos blaZ
e blaR1-blaI pode iniciar a síntese de betalactamase [7, 25].
Diferentes testes podem ser utilizados para avaliar produção de betalactamase em
estafilococos. Os procedimentos qualitativos, como a detecção da produção de betalactamase

53
usando discos de Cefinase, onde a reação é baseada na produção de um composto colorido
quando o disco entra em contato com uma bactéria produtora de betalactamase. Outro
procedimento utilizado é o teste de Clover-Leaf, baseado na formação de uma zona de distorção
no halo de inibição do disco de penicilina [2, 21].
Apesar da importância que a betalactamase desempenha nas infecções por
Staphylococcus spp., poucos trabalhos no Brasil fazem referência a ocorrência desta enzima
relacionados ao padrão de resistência bacteriana, tampouco a sua caracterização genética. Com
o aumento dos casos de cepas fenotipicamente resistentes à meticilina, porém sem a presença
do gene mecA, observados nos pacientes hospitalizados em hospital universitário da cidade de
Recife – PE, busca-se, através deste trabalho, a identificação da ocorrência da betalactamase
como responsável pelo padrão de resistência à meticilina encontrados nestas cepas provenientes
de isolados clínicos.
Materiais e Métodos
Obtenção de Amostras
Foram utilizadas quarenta e sete isolados clínicos provenientes do Hospital das Clínicas
da UFPE, coletadas durante o período de abril de 2011 a julho de 2013.Todos os isolados foram
estocados na Bacterioteca do Laboratório de Bacteriologia do Departamento de Medicina
Tropical-UFPE. Tais isolados foram fornecidos pelos responsáveis dos laboratórios de
bacteriologia do referido hospital, após a identificação bacteriana. Desta forma, a coleta da
amostra foi de responsabilidade do hospital. Essas amostras foram originadas de ponta de
cateter, drenos, sangue, abscesso, ferida operatória e secreções diversas.
Seleção dos isolados
A resistência à meticilina nos testes fenotípicos foi utilizada como critério de inclusão
no estudo. Todos os isolados já haviam sido previamente identificados e submetidos à PCR do
gene mecA antes de serem armazenados na Bacterioteca do Laboratório de Bacteriologia do
Departamento de Medicina Tropical-UFPE.
Para essa identificação, os isolados foram submetidos aos testes de disco-difusão de
oxacilina, disco-difusão de cefoxitina e screening de oxacilina. Aqueles isolados que se
apresentaram resistentes a pelo menos uma das técnicas utilizada, foram submetidos à PCR para

54
detecção do gene mecA onde foram utilizados para amplificação utilizados os primers P1, 5'-
GGTCCCATTAACTCTGAAG-3’ e P3, 5’-AGTTCTGCAGTACCGGATTTGC-3’ descritos
por PETINAKI, et al. (2001) que resultam em fragmentos de 1.046pb.
Teste da Cefinase em disco
O teste de cefinase em disco foi realizado utilizando o BBL Cefinase Discs (BD) de
acordo com as especificações do fabricante. Os discos foram distribuídos em uma placa de Petri
esterilizada. Em seguida, cada disco foi umedecido com uma gota (aprox. 30µL) de água miliQ
esterilizada e, com uma alça de platina, foi realizado um esfregaço das colônias em teste no
disco de cefinase. O resultado positivo foi detectado pela mudança de cor, de amarelada para
rosa, apresentada pelo disco. Esta mudança de cor deve-se ao composto Nitrocefin, uma
cefalosporina cromógena, que muda de cor à medida que a ligação amido no anel beta-
lactâmico é hidrolisada por uma β-lactamase. Para Staphylococcus spp. esperou-se até uma hora
para que ocorresse a mudança de coloração. Como controle positivo foi utilizada a cepa MRSA
ATCC 33591 e como controle negativo foi utilizada a cepa de MSSA ATCC 6538.
Teste de Clover-Leaf
O teste de Clover-Leaf consiste num teste de Hodge modificado e foi realizado de
acordo com a técnica de Bergan et al. (1997). Primeiramente preparou-se um inóculo de um
isolado betalactamase negativo (cepa ATCC 25923), de acordo com o 0,5 da escala de
McFarland através de suspensão direta. Em seguida foi realizado o semeio em tapete deste
isolado em Ágar Mueller-Hinton e colocado um disco de penicilina 10U (Laborclin) no centro
da placa. Com uma alça de platina, foram realizadas estrias do centro até a periferia da placa de
Petri, sem tocar o disco de penicilina. A placa foi incubada de 18h a 24h a 35ºC e após foi
observado o aparecimento da zona de distorção no halo de inibição dos isolados considerados
positivos. Como controle positivo do teste, foi utilizado o isolado 1171, produtor de
betalactamase, da bacterioteca do CPqAM/FIOCRUZ. Como controle negativo foi utilizada a
cepa MSSA ATCC 25923.
Extração de DNA total

55
Para a extração do DNA das amostras de Staphylococcus foi utilizada a técnica de
extração por fervura, na qual o DNA é extraído a partir de 1mL de cultura bacteriana
previamente crescida em caldo BHI a 37ºC por 24h e centrifugada a 14,000rpm em 4ºC durante
cinco minutos. Em seguida, desprezou-se o sobrenadante e o sedimento foi ressuspenso em
200µL de água MiliQ esterilizada. A suspensão foi incubada a 100ºC pór 10 min seguido de
centrifugação a 14,000rpm/4ºC durante cinco minutos. Após centrifugação, 100µL do
sobrenadante foi coletado em novo tubo e estocado a -20ºC.
Identificação do gene da betalactamase por PCR
Para a amplificação do gene blaZ foram utilizados os primers F1, 5'-
GATAAGAGATTTGCCTATGC-3' e R1, 5'-GCATATGTTATTGCTTGACC-3'
(MILHEIRIÇO et al., 2011) que resultam em fragmentos de 533pb. As reações de PCR foram
preparadas individualmente em um volume final de 25µL em cada tubo contendo: 5µL do DNA
genômico, 20pmol de cada primer, MgCl2 1,5mM, dNTP 200µM, 1U de Go Taq DNA
polimerase (Promega, Brasil) e 5µL do tampão Green Go Taq DNA polimerase (Promega,
Brasil). As reações foram realizadas em termociclador (Biometra), programado inicialmente
para desnaturação inicial de quatro minutos a 92ºC, em seguida, 30 ciclos térmicos de um
minuto a 92ºC, um minuto a 55ºC, um minuto a 72ºC e etapa final de sete minutos a 72ºC. Foi
utilizado como controle negativo, um tubo contendo todos os componentes da mistura e
ausência de DNA molde. Para controle positivo foi utilizada uma cepa de S. aureus resistente
à meticilina (código do isolado: 1171 da bacterioteca do CPqAM/Fiocruz-PE).
O produto da amplificação foi submetido à eletroforese em gel de agarose 1% e corado
com brometo de etídio (2µg/mL), visualizado em transluminador UV e digitalizado através do
software Kodak 1D versão 3.5.2 (Scientific Imaging Systems, USA). O marcador de peso
molecular é de 100bp (Invitrogen).
Considerações éticas
Este estudo fez parte do projeto CAAE – 0490.0.172.000-11 Registro CEP/CCS/UFPE
nº 009/11 (Anexo III) submetido à análise pelo Comitê de Ética em Pesquisa (CEP) do Centro
de Ciências da Saúde (CCS), da UFPE, com a permissão e colaboração da Comissão de
Controle de Infecção Hospitalar (CCIH) do hospital envolvido. Não houve Termo de

56
Consentimento Livre e Esclarecido (TCLE) para os pacientes por se tratar de amostras
secundárias.
Análise Estatística
Os dados microbiológicos foram analisados estatisticamente através da distribuição de
frequências e Teste χ², utilizandoosoftwareEpiInfoversão6.04.
Resultados
Foram analisados 47 (quarenta e sete) isolados de Staphylococcus spp., sendo 13 (treze)
isolados identificados como Staphylococcus coagulase negativo e 34 (trinta e quatro)
identificados como Staphylococcus aureus. Todos os isolados selecionados se mostraram
resistentes a pelo menos um dos testes fenotípicos de resistência à meticilina (disco-difusão de
oxacilina, disco-difusão de cefoxitina e screening de oxacilina) e foram submetidos à PCR para
detecção do gene mecA e separados nos grupos para estudo de acordo com o resultado da PCR.
De acordo com os resultados da PCR, os isolados foram selecionados em dois grupos:
Os isolados fenotipicamente resistentes e portadores do gene mecA (15 isolados) e os isolados
fenotipicamente resistentes e não portadores do gene mecA (32 isolados).
Nos isolados pertencentes ao grupo portador do gene mecA, apenas um isolado se
mostrou resistente apenas ao teste de Disco-difusão de cefoxitina, os demais foram resistentes
aos três testes realizados. No que se refere aos isolados pertencentes ao grupo não portador do
gene mecA, ocorreu uma distribuição heterogênea quanto aos resultados apresentados nos testes
fenotípicos de resistência à meticilina, 37% foram resistentes a apenas um teste, 22% foram
resistentes a dois dos testes e 37% foram resistentes aos três testes realizados.
Dos isolados que apresentaram resistência a dois testes fenotípicos diferentes, cinco
apresentaram-se resistentes aos testes de disco-difusão de cefoxitina e ao screening de
meticilina e três isolados demonstraram resistência aos testes de disco-difusão de oxacilina e
cefoxitina.
Dentre os isolados que apresentaram resistência a apenas um teste fenotípico, quatro
foram resistentes apenas ao teste de disco-difusão de oxacilina, cinco apresentaram resistência
apenas ao teste de disco-difusão de oxacilina, enquanto dois se mostraram resistentes apenas
no teste de screening de meticilina.

57
No teste da Cefinase em disco para a detecção fenotípica de betalactamase, foi
observada positividade em apenas cinco dos 15 isolados (33,33%) pertencentes ao grupo
portador do gene mecA. No grupo não portador do gene mecA, foi observada a positividade no
teste para 18 dos 32 isolados (56,25%). O teste é considerado positivo quando o disco de
cefinase muda de coloração, adquirindo uma tonalidade avermelhada (figura 1).
No teste de Clover-Leaf, para detecção fenotípica da produção de betalactamase,
também foi observada a positividade do teste em cinco dos 15 isolados analisados (33,33%)
pertencentes ao grupo portador do gene mecA. Observa-se no entanto, que o isolado 33STVd,
positivo para o teste da Cefinase em disco, foi negativo para o teste de Clover-Leaf enquanto o
isolado 40, que mostrou-se negativo para o teste da Cefinase em disco, foi positivo para o teste
de Clover-Leaf.
No grupo de isolados não portador do gene mecA, foi observado que 21 das 32 amostras
(65,62%) foram positivas para o teste de Clover-Leaf. Observou-se ainda que os isolados 24,
105 e 117, que se mostraram negativos para o teste da Cefinase em disco, mostraram-se
positivos no teste de Clover-Leaf. O teste foi considerado positivo quando ocorreu uma da zona
de distorção no halo de inibição do disco de penicilina (figura 2).
A reação de PCR para o gene blaZ, confirmou a presença do gene de produção de
betalactamase em oito dos 15 isolados do grupo portador do gene mecA, perfazendo um
percentual de 53,33% dos isolados estudados. No grupo não portador do gene mecA, foi
observada a presença do gene em 12 dos 32 isolados estudados, perfazendo um percentual de
37,5%. Foi observada a amplificação do fragmento de tamanho esperado de aproximadamente
533pb (figura 3).
Vários isolados foram positivos para os métodos fenotípicos estudados porém
apresentaram-se negativos na pesquisa do gene blaZ através de PCR (Tabela 1).
Utilizando a reação de PCR como padrão ouro, calculou-se a sensibilidade e
especificidade dos diferentes métodos analíticos utilizados para identificação da produção de
betalactamase (Tabela 2). Deste modo verifica-se que o teste de Clover-Leaf possui maior
sensibilidade para detecção da produção de betalactamase neste estudo, e que ambos os testes
têm especificidade de detecção idênticos. Observa-se ainda que essa especificidade é baixa para
detecção.
Na comparação dos grupos, “Portadores do gene mecA” versus “Não portadores do gene
mecA”, não houve diferença significativa na distribuição de frequências, de acordo com o teste
χ² (p >0,05).

58
Discussão
Desde o surgimento dos primeiros antibióticos, Staphylococcus spp. vem adquirindo
mecanismos de resistência a estes fármacos, sendo dessa forma capazes de persistir no ambiente
hospitalar, tornando-se um dos principais agentes de bacteremia adquirida nesse ambiente [11,
27]. O desenvolvimento de mecanismos de resistência limita as possibilidades de tratamento
do paciente, sendo desta forma, crucial determinar se os isolados de Staphylococcus spp. são
resistentes à meticilina/oxacilina, tanto para o tratamento quanto para o controle de surtos [1,
10]
Apesar do gene mecA ser considerado o padrão ouro para a detecção de resistência à
meticilina, esse método é mais utilizado em laboratórios de referência e não nos laboratórios de
testes microbiológicos de rotina. Esses laboratórios utilizam frequentemente métodos
fenotípicos para identificação da resistência à meticilina [16]. Em alguns estudos, no entanto,
verifica-se discrepância entre a apresentação fenotípica dessas cepas e a presença do gene mecA
[1, 3, 28].
No presente estudo verificou-se alta taxa de discrepância entre a ocorrência do gene
mecA e os testes fenotípicos de resistência à meticilina (disco-difusão de oxacilina, disco-
difusão de cefoxitina e screening de oxacilina), visto que 68% dos isolados resistentes a pelo
menos um dos testes fenotípicos não apresentavam o gene. Em seus estudos, Ghoshal et al.
(2004) também encontrou incoerências entre resultados de testes fenotípicos e a presença do
gene mecA, chegando a conclusão que a hiperprodução de betalactamase seria a responsável
pela indução da resistência à oxacilina.
Foram realizados então testes fenotípicos para detecção da produção de betalactamase
e pesquisa do gene blaZ nestes isolados. Em diversos estudos a taxa de produção de
betalactamase variam de 55,7% a 92,6% para estafilococos [22]. Neste estudo foram
encontradas frequências de detecção mais baixas, se consideradas a totalidade dos isolados,
para todos os testes realizados, visto que, no teste da cefinase em disco a produção de
betalactamase foi detectada em 48,9% dos isolados, no teste de Clover-Leaf detectou-se em
55,3% dos isolados e na PCR para detecção do gene blaZ em 42,55% dos isolados.
No presente estudo, cinco isolados possuíam o gene blaZ, mas não demonstraram
produção de betalactamase em nenhum dos testes fenotípicos realizados, esse fato é comum,
pois a presença do gene não garante, necessariamente, a expressão do mesmo. A expressão
desse fator deve ser avaliada através de técnicas de RT-PCR ou qRT-PCR.

59
Foi verificado também que doze isolados apresentaram produção de betalactamase nos
dois testes fenotípicos realizados, porém não possuíam o gene blaZ. Este fato pode ser explicado
pela presença de polimorfismos no gene, podendo gerar sequências divergentes como já
descritas por Olsen et al. (2006) e Milheiriço et al. (2011). Em seus estudos, El Feghaly et al.
(2012) utilizou três diferentes sequências que codificavam diferentes regiões do gene blaZ a
fim de contabilizar as divergências encontradas.
O teste da cefinase em disco se mostrou com baixa sensibilidade (55%) em relação ao
padrão-ouro utilizado, ficando abaixo dos resultados encontrados por Pitkälä et al. (2007), que
reportou sensibilidade variando de 62% a 100% para o teste. No entanto, ficou acima da
sensibilidade encontrada por Kaase et al. (2008) em seus estudos, que reportou sensibilidade de
apenas 39% utilizando este método. Apesar da sensibilidade dependente de condições do
operador e do tipo de metodologia utilizada, o teste de cefinase é bastante utilizado por sua
rapidez na detecção e baixo custo de operação [5, 20].
O teste de Clover-Leaf foi o que apresentou maior sensibilidade neste estudo (70%),
resultado este que corrobora com diversos outros estudos, que consideram o teste um dos mais
sensíveis para detecção de betalactamase, chegando em alguns estudos a alcançar taxas entre
94% e 98% de sensibilidade. Assim como o teste da Cefinase, este teste é de fácil realização e
baixo custo [2, 5, 8, 20].
Robles et al. (2014), em seus estudos não recomenda assumir a PCR para o gene blaZ
como padrão-ouro para detecção de produção de betalactamase, uma vez que o fenótipo de
produção da betalactamase podia resultar do produto de mais de um gene e ainda reforça a ideia
que a presença do gene não garante sua expressão fenotípica. Além disso defende o uso dos
testes de Cefinase em disco e Clover-Leaf para detecção da produção de betalactamase por
geralmente apresentarem boa sensibilidade. Esses achados podem corroborar com os resultados
do presente estudo, no qual encontrou baixas taxas de sensibilidade nos testes fenotípicos por
utilizar-se da PCR para detecção do gene blaZ como padrão ouro.
Na comparação de frequências do gene blaZ entre os grupos de estudo utilizados, não
foi encontrada diferença significativa na comparação dos grupos, o que pode sugerir tanto que
o fenótipo de resistência a meticilina possa se dever a um fator alheio, que não a presença de
betalactamase, quanto que a baixa detecção da presença do gene pode estar ligada às
divergências de sequências relatadas por El Feghaly et al. (2012).
Apesar de bastante específica, a reação de PCR para detecção do gene blaZ não deve
ser usada como padrão ouro para identificação de resistência, uma vez que a expressão da
betalactamase pode se dever a outros fatores que não apenas a presença deste gene. Desta forma,

60
indica-se o uso dos testes fenotípicos de detecção de produção de betalactamase, por
apresentarem baixo custo e fácil operação, além de demonstrarem boa sensibilidade.
Referências
[1] ARAJ, G.F, TALHOUK, R.S, SIMAAN, C.J, MAASAD, M.J. Discrepancies between
mecA PCR and conventional tests used for detection of methicilin resistant Staphylococcus
aureus.Int J Antimicrob Agents, v. 11, p. 47-52, 1999.
[2] BERGAN, T.; BRUNN, J.N.; DIGRANES, A.; LINGAAS, E.; MELBY, K.K.; SANDER,
J. Susceptibility testing of bacteria and fungi. Report from: “the Norwegian Working Group on
Antibiotics”. Scand J Infect (Suppl.) v. 103, p. 1-36. 1997.
[3] CUNY, C.; LAYER, F.; STROMMENGER, B.; WITTE, W. Rare occurrence of
methicillin- resistant Staphylococcus aureus CC130 with a novel mecA homologue in humans
in Germany. PLoS ONE, v. 6, n. 9, p. e24360, 2011.
[4] DURAN, N, OZER, B, DURAN, G.G, ONLEY, Y, DEMIR, C. Antibiotic resistance genes
& suscebility patterns in staphylococci. Indian J Med Res. v. 135, p. 389-396, 2012.
[5] EL FEGHALY, R.E.; STAMM, J.E.; FRITZ, S.A.; BURNHAM, C.A.D. Presence of the blaZ
beta-lactamase gene in isolates of Staphylococcus aureus that appear penicillin susceptible by
conventional phenotypic methods. Diag Microbiol Infect Dis, v. 74, p. 388-393, 2012.
[6] GHOSHAL, U.; PRASSAD, K.N.; SINGH, M.; TIWARI, D.P.; AYYAGARI, A. A
Comparative evaluation of phenotypic and molecular methods for the detection of oxacillin
resistancce in coagulase-negative staphylococci. J Infect Chemother, v. 10, p. 86-89, 2004.
[7] HAO, H, DAI, M, WANG, Y, HUANG, L, YUAN, Z. Key genetics elements and regulation
systems in methicillin-resistant Staphylococcus aureus. Future Microb. v. 7, n. 11, p. 1315-
1329, 2012.
[8] JARLØV, J.O.; ROSDAHL, V.T. Quantitative determination of beta-lactamase production
in Staphylococcus aureus strains compared to qualitative testing by a microbiological clover
leaf test, a chromogenic cephalosporin test and a iodometric test. Acta Pathol Microbiol
Immunol Scand B, v. 94, p.415-421, 1986.
[9] KAASE, M.; LENGA, S.; FRIEDERICH, S.; SZABADOS, F.; SAKINC, T.; KLEINE, B.;
GATERMANN, S.G. Comparison of phenotipic methods for penicillinase detection in
Staphylococcus aureus. Clin Microbiol Infect, v. 14, p. 614-616, 2008.

61
[10] KIPP, F.; BECKER, K.; PETERS, G.; VON EIFF, C. Evaluation of differen methods to
detect methicillin resistance in small-colony variants of Staphylococcus aureus. J Clin
Microbiol, v. 42, n. 3, p. 1277-1279, 2004.
[11]
KLUYTMANS,J.;STRUELENS,M.MethicillinresistantStaphylococcusaureusinthehospital.B
MJ.v. 12, n. 1, p.338-364,2009.
[12] LOWY, F.D. Antimicrobial resistance: the example of Staphylococcus aureus. J Clin
Invest. v. 111, p.1265-1273, 2003.
[13] MARLOWE, E.M, BANKOWSKI, M.J. Conventional and molecular methods for the
detection oc methicilin-resistant Staphylococcus aureus. J Clin Microbiol. v. 49, n. 9, p. S53-
S56, 2011.
[14] MILHEIRIÇO, C, PORTELINHA, A, KRIPPAHL, L, DE LENCASTRE, H, OLIVEIRA,
D.C. Evidence for a purifying selection acting on the betalactamase locus in epidemic clones
of methicillin-resistant Staphylococcus aureus. BMC Microbiology, v. 11, n. 1, p. 76, 2011.
[15] MOELLERING JR, R.C. MRSA: the first half century. J Antimicrob Chemother, v. 67, p.
4-11, 2012.
[16] ODONKOR, S.T.; ADDO, K.K. Evaluation of three methods for detection of methicillin
resistant Staphylococcus aureus (MRSA). Int J Biol Med Res, v. 2, n.4, p. 1031-1034, 2011.
[17] OLIVEIRA, A.A.F.; PINHEIRO JR, J.W.; MOTA, R.A.; CUNHA, M.L.R.S; LOPES,
C.A.M; ROCHA, N.S. Phenotype characterization of Staphylococcus species strains isolated
from buffalo (Bubalus bubalis) milk. J Vet Diag Invest, v. 23, n. 6, p. 1208-1211, 2011.
[18] OLSEN, J.E, CHRISTENSEN, H, AARESTRUP, FM. Diversity and evolution of blaZ
from Staphylococcu aureus and coagulase-negative sthaphilococci. J Antimicrob Chemoter. v.
57, p. 450-460, 2006.
[19]PETINAKI, E, ARVANITI, A, DIMITRACOPOULOSG, SPILIOPOULOUI. Detection of
mecA, mecR1 and mecI genes among clinical isolates of methicillin-resistant staphylococci by
combined polymerase chain reactions .J Antimicrob Chemother. v. 47, n. 3, p.297-304,2001.
[20] PITKÄLÄ, A.; SALMIKIVI, L.; BREDBACKA, P.; MYLLYNIEMI, A.L.; KOSKINEN,
M.T. Comparison of tests for detection of beta-lactamse-producing staphylococci. J Clin
Microbiol, v. 45, p. 2031-2033, 2007.
[21] ROBLES, B.F.; NOBREGA, D.B.; GUIMARÃES, F.F.; WANDERLEY, G.G.;
LANGONI, H. Beta-lactamase detection in Staphylococcus aureus and coagulase-negative
Staphylococcus isolates from bovine mastites. Pesq. Vet. Bras, v. 34, n.4, p. 325-328, 2014.

62
[22] SAMANT, S.A.; PAI, C.G. Comparative evaluation of β lactamase detection methods in
staphylococci. IJRPBS, v. 3, n. 4, p. 1580-1588, 2012.
[23] SANTOS, A.L, SANTOS D.O, FREITAS C.C, FERREIRA, B.L.A, AFONSO, I.F,
RODRIGUES C.R, CASTRO, H.C. Staphylococcus aureus: visiting a strain of clinical
importance. J Bras Patol Med Lab. v. 43, n. 6, p. 413-423, 2007.
[24] SHARIATI, L.; VALIDI, M.; TABATABAIEFAR, M.A.; KARIMI, A.; NAFISI, M.R.
Comparison of Real-Time PCR with disk diffusion, agar screen and E-test methods for
detection of methicillin-resistant Staphylococcus aureus. Curr Microbiol, v. 61, p. 520–524,
2010.
[25] STAPLETON, P.D, TAYLOR, P.W. Methicilin resistance in Staphylococcus aureus:
mechanisms and modulation. Sci Prog. v. 85, p. 57-72, 2002.
[26] SWENSON, J.M.; LONSWAY, D.; MCALLISTER, S.; THOMPSON, A.; JEVITT, L.;
ZHU, W.; PATEL, J.B. Detection of mecA-mediated resistance using reference and
commercial testing methods in a collection of Staphylococcus aureus expressing boderline
oxacillin MICs. Diag Microbiol Infect Dis, v. 38, n. 5, p. 1346-1350, 2007.
[27] TAVARES, W. Bactérias gram-positivas problemas: resistência do estafilococo, do
enterococo e do pneumococo aos antimicrobianos. Rev Soc Bras Med Trop, v. 33, n. 3, p. 281-
301, 2000.
[28] YOSHIDA, R.; KUWAHARA-ARAI, K.; BABA, T.; CUI, L.; RICHARDSON, J.F.;
HIRAMATSU, K. Physiological and molecular analysis of a mecA-negative Staphylococcus
aureus clinical strain that expresses heterogeneous methicillin resistance. J Antimicrob
Chemother, v. 51, p. 247-255, 2003.
Figura 1: Teste da Cefinase em disco
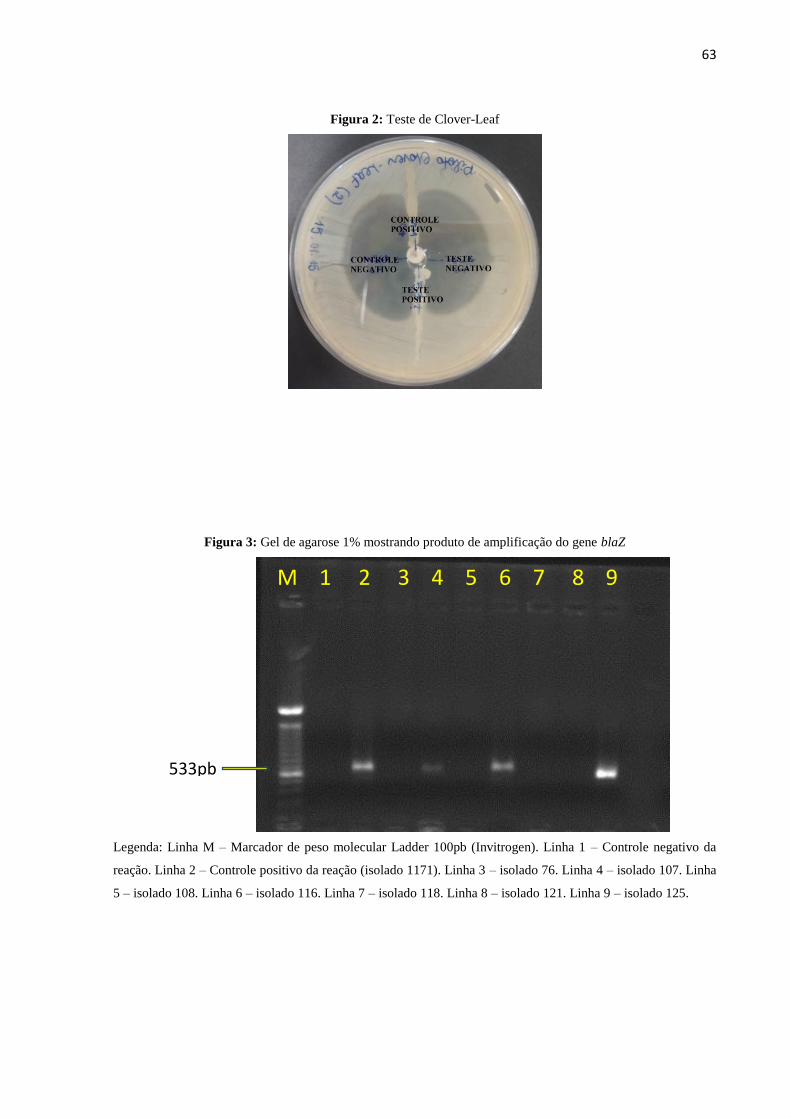
63
Figura 2: Teste de Clover-Leaf
Figura 3: Gel de agarose 1% mostrando produto de amplificação do gene blaZ
Legenda: Linha M – Marcador de peso molecular Ladder 100pb (Invitrogen). Linha 1 – Controle negativo da
reação. Linha 2 – Controle positivo da reação (isolado 1171). Linha 3 – isolado 76. Linha 4 – isolado 107. Linha
5 – isolado 108. Linha 6 – isolado 116. Linha 7 – isolado 118. Linha 8 – isolado 121. Linha 9 – isolado 125.
M 1 2 3 4 5 6 7 8 9
533pb
64
Tabela 1: Distribuição da frequência da detecção da betalactamase de acordo com a PCR do gene blaZ, teste da
Cefinase em disco e teste de Clover-Leaf em isolado sdeStaphylococcus spp. resistentes à meticilina com e sem a
presença do gene mecA.
Isolados com a presença do gene
mecA
Isolados sem a presença do gene
mecA
N % (do total) n % (do total)
GeneblaZ +
-
8
7
17,0
14,9
12
20
25,5
42,6
Cefinase +
-
5
10
10,6
21,3
18
14
38,3
29,8
Clover-Leaf +
-
5
10
10,6
21,3
21
11
44,7
23,4
Tabela 2: Sensibilidade e especificidade dos métodos analíticos para detecção de betalactamase.
Método Sensibilidade Especificidade
Cefinase
Clover-Leaf
55%
70%
44,4%
44,4%
Padrão-ouro = utilização da PCR para detecção do gene blaz

65
6. Conclusão

66
Foi verificado grande número (trinta e dois) de isolados fenotipicamente resistentes aos
testes de detecção de resistência à meticilina, porém sem a presença do gene mecA.
O teste de Clover-Leaf mostrou-se mais sensível para detecção da produção de
betalactamase que o teste da cefinase em disco.
A PCR para detecção do gene blaZ mostrou correlação negativa com os testes
fenotípicos realizados.
Não houve diferença significativa na detecção do gene blaZ entre os grupos estudados.
PERSPECTIVAS
Sequenciamento dos fragmentos amplificados com o gene da betalactamase para
confirmação da identidade.
Padronização de PCR para os reguladores (blaI e blaR1) do gene blaZ.
Identificação de clones entre os isolados pesquisados.
Fazer estudo de expressão nos isolados que demonstraram presença do gene blaZ.

67
Referências

68
ARAJ, G.F, TALHOUK, R.S, SIMAAN, C.J, MAASAD, M.J. Discrepancies between mecA
PCR and conventional tests used for detection of methicilin resistant Staphylococcus aureus.
Int J Antimicrob Agents, v. 11, p. 47-52, 1999.
BERGAN, T.; BRUNN, J.N.; DIGRANES, A.; LINGAAS, E.; MELBY, K.K.; SANDER, J.
Susceptibility testing of bacteria and fungi. Report from: “the Norwegian Working Group on
Antibiotics”. Scand J Infect (Suppl.) v. 103, p. 1-36. 1997.
BERGLUND, C.; ITO, T.; IKEDA, M.; MA, X.X.; SÖDERQUIST, B.; HIRAMATSU, K.
Novel Type of Staphylococcal Cassette Chromosome mec in a methicillin-resistant
Staphylococcus aureus strain isolated in Swede. Antimicrob Agents Chemother, v. 52, n. 10, p.
3512-3516, 2008.
BIRCK, C.; CHA, J.Y.; CROSS, J.; SCHULZE-BRIESE, C.; MEROUEH, S.O.; SCHLEGEL,
H.B.; MOBASHERY, S.; SAMAMA, J.P. X-ray crystal structure of the acylated β-lactam
sensor domain of BlaR1 from Staphylococcus aureus and the mechanism of receptor activation
for signal transduction. J Am Chem Soc. v. 126, n. 43, p. 13945-13947, 2004.
BLACK, C.C, EBERLEIN, L.C, SOLYMN, S.M, WILKES, RP, HARTMANN, FA,
ROHRBACH, B.W, BEMIS, D.A, KANIA, S.A. The role of mecA and blaZ regulatory
elements in mecA expression by regional clones of methicillin-resistant Staphylococcus
pseudintermedius. Veterinaty Microbiolog, v. 151, p. 345-353, 2011.
BRASIL, Agência Nacional de Vigilância Sanitária. Manual de Microbiologia Clínica para o
Controle de Infecção Relacionada à Assistência à Saúde. Módulo 6: Detecção e identificação
de bactérias de importância médica. Brasília: ANVISA, 2013.
BUSH, K.; JACOBY, G.A.; MEDEIROS, A.A. A functional classification scheme for
βlactamases and its correlation with molecular structure. Antimicrob Agents Chemother, v.
39, p. 12111233, 1995.
CASSETARI,V.C.; STRABELLI,T.; MEDEIROS,E.A.S. Staphylococcus aureus bacteremia:
what is the impact of oxacillin resistance on mortality? Braz J Infect Dis.v. 9, n. 1, p.70-76,
2005.

69
CHAMBERS, H.F. Methicillin resistance in Staphylococci: molecular and biochemical basis
and clinical implications. Clin Microbiol Rev, v. 10, n. 4, p. 781-791, 1997.
CLARKE, S.R, DYKE, K.G.H. Studies of the operator region of the Staphylococcus aureus
betalactamase operon. J Antimicrob Chemoter. v. 47, p. 377-389, 2001.
CLSI (Clinical and Laboratory Standards Institute). Performance standards for antimicrobial
susceptibility testing, twenty-third informational supplement, document M100-S23. Wayne,
PA,USA. CLSI, 2013.
COELLO, R.; GLYNN, J.R.; GASPAR, C.; PICAZO, J.J.; FERERES, J. Risk factors for
developing clinical infection with methicillin-resistant Staphylococcus aureus(MRSA)
amongst hospital patients initially only colonized with MRSA. J Hosp Infect, v. 37, n. 1, p. 39-
46, 1997.
CONCEIÇÃO, T.; TAVARES, A.; MIRAGAIA, M.; HYDE, K.; AIRES-DE-SOUSA, M.; DE
LENCASTRE, M. Prevalence and clonality of methicillin-resistant Staphylococcus aureus
(MRSA) in the Atlantic Azores islands: predominance of SCCmec types IV, V and VI. Eur J
Clin Microbiol Infect Dis, v. 29, n. 5, p. 543–550, 2010.
CORBELLA, X.; DOMINGUEZ, M.A., PUJOL, M., AYATS, J., SENDRA, M., PALLARES,
R., ARIZA, J., GUDIOL, F. Staphylococcus aureus nasal carriage as a marker for subsequent
staphylococcal infections in intensive care unit patients. Eur J Clin Microbiol Infect Dis, v. 16,
n. 5, p. 351-357, 1997.
COSTA, T.M.; PINHEIRO, M.G.; CARDOSO, C.A.; ALVES, M.P.; AGUIAR-ALVES, F.
Características clínicas e esquema de tratamento medicamentoso de infecções por
Staphylococcus aureus: uma possível proposta de identificação para o laboratorista e tratamento
para o clínico. Revista Praxis, ano III, n. 5, p. 15-24, 2011.
CORREAL, J.C.D.; MARQUES, E.A.; GUILHERME, W.L.; LEÃO, R.S.; DAMASCO, P.V.
Infecções por Staphylococcus aureus: mudança do perfil epidemiológico no Hospital
Universitário Pedro Ernesto. Revista HUPE, v. 12, n. 3, p. 31-46, 2013.

70
CUNY, C.; LAYER, F.; STROMMENGER, B.; WITTE, W. Rare occurrence of methicillin-
resistant Staphylococcus aureus CC130 with a novel mecA homologue in humans in Germany.
PLoS ONE, v. 6, n. 9, p. e24360, 2011.
DAUM, R.S. Skin and soft-tissue infections caused by methicillin-resistant Staphylococcus
aureus. N Engl J Med, v. 357, n. 4, p. 380-390, 2007.
DEURENBERG, R.H.; STOBBERINGH, E.E. The evolution of Staphylococcus aureus. Infect
Genet Evol, v. 8, n. 6, p. 747-763, 2008.
DINGES, M.M.; ORWIN, P.M.; SCHLIEVERT, P.M. Exotoxins of Staphylococcus aureus.
Clin Microbiol Rev, v. 13, n. 1, p. 16-34, 2000.
DUARTE, C.O.; DE LENCASTRE, H. Multiplex PCR strategy for rapid identification of
structural types and variants of the mec element in methicillin-resistant Staphylococcus aureus.
Antimicrob Agents Chemother, v. 46, p. 2155-2161, 2002.
DURAN, N, OZER, B, DURAN, G.G, ONLEY, Y, DEMIR, C. Antibiotic resistance genes &
suscebility patterns in staphylococci. Indian J Med Res. v. 135, p. 389-396, 2012.
DZIDIC, S, SUSKOVIC, J, KOS, B. Antibiotic resistance mechanisms in bacteria: Biocheical
and genetic aspects. Food Technol Biotechnol. v. 46, n. 11, p. 11-21, 2008.
EL FEGHALY, R.E.; STAMM, J.E.; FRITZ, S.A.; BURNHAM, C.A.D. Presence of the blaZ
beta-lactamase gene in isolates of Staphylococcus aureus that appear penicillin susceptible by
conventional phenotypic methods. Diag Microbiol Infect Dis, v. 74, p. 388-393, 2012.
ELEK, S.D. Staphylococcus pyogenes and its relation to disease. Edinburgh: E. & S.
Livingstone, p. 3, 1959.
ENRIGHT, M.C.; ROBINSON, D.A.; RANDLE, G.; FEIL, E.J.; GRUNDMANN, H.; SPRATT,
B.G. The evolutionary history of methicilin-resistant Staphylococcus aureus (MRSA). Proc Natl
Acad Sci U S A. v. 99, p. 7687-7692, 2002.

71
FLOCK, J.I.; HIENZ, S.A., HEIMDAHL, A., SCHENNINGST, T. Reconsideration of the role
of fibronectin binding in endocartidis caused by Staphylococcus aureus. Infect Immun, v. 64, n.
5, p. 1876-1878, 1996.
FRANCO, B.G.DM, JANDGRAFF, M. Microbiologia de alimentos. São Paulo. Atheneu,
2000.
FREDERICK, T.E.; WILSON, B.D.; CHA, J.; MOBASHERY, S.; PENG, J.W. Revealing cell-
surface intramolecular interactions in the BlaR1 protein of methicillin-resistant Staphylococcus
aureus by NMR spectroscopy. Biochemistry, v. 53, n. 1, p. 10-12, 2013.
FREITAS, M.F.L.; LUZ, I.S.; SILVEIRA-FILHO , V.M.; JÚNIOR, J.W.P.; STAMFORD,
T.L.M.; MOTA, R.A.; SENA, M.J.; ALMEIDA, A.M.P.; BALBINO, V.Q.; LEALBALBINO,
T.C. Staphylococcal toxin genes in strains isolated from cows with subclinical mastitis. Pesq
Vet Bras, v. 28, n. 12, p. 617-621, 2008.
FRIDKIN, S.K.; HAGEMAN, J.C.; MORRISON, M.; SANZA, L.T.; COMO-SABETTI, K.;
JERNIGAN, J. A.; HARRIMAN, K.; HARRISON, L. H.; LYNFIELD, R.; FARLEY, M. M.
Methicillin-resistant Staphylococcus aureus in three comunities. N Engl J Med, v. 352, n. 14,
p. 1436-1444, 2005.
FUDA, C.C.S; FISCHER, J.F.; MOBASHERY, S. β-Lactam resistance in Staphylococcus
aureus: the adaptative resistance of a plastic genome. Cell Mol Life Sci, v. 62, p. 2617-2633,
2005.
GALES, A.C.; SADER, H.S.; RIBEIRO, J.; ZOCCOLI, C.; BARTH, A.; PIGNATARI, A.C.
Antimicrobial susceptibility of gram-positive bacteria isolated in Brazilian hospitals
participating in the SENTRY program (2005-2008). Braz J Infect Dis, v. 13, n. 2, p. 90-98,
2009.
GHOSHAL, U.; PRASSAD, K.N.; SINGH, M.; TIWARI, D.P.; AYYAGARI, A. A
Comparative evaluation of phenotypic and molecular methods for the detection of oxacillin
resistancce in coagulase-negative staphylococci. J Infect Chemother, v. 10, p. 86-89, 2004.

72
GONZALES, E.; MELLO, H.H.C.; CAFÉ, M.B. Uso de antibióticos promotores de
crescimento na alimentação e produção animal. Revista UFG, ano XIII, n. 13, p. 48-53, 2012.
GORDON, R.; LOWY, F.D. Pathogenesis of methicillin-resistant Staphylococcus aureus
infection. Clin Infect Dis, v. 46, p. 350-359, 2008.
HACKBARTH, C.J, CHAMBERS, H.F. blaI and blaR1 Regulate betalactamase and PBP2a
production in methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemoter, v. 37,
n. 5, p. 1144-1149, 1993.
HACKBARTH, C.J, MIICK, C, CHAMBERS, H. Altered production of Penicilin-Binding
Protein 2a can affect phenotypic expression of methicillin resistance in Staphylococcus aureus.
Antimicrob Agents Chemoter, v. 38, n. 11, p. 2568-2571, 1994.
HAO, H, DAI, M, WANG, Y, HUANG, L, YUAN, Z. Key genetics elements and regulation
systems in methicillin-resistant Staphylococcus aureus. Future Microb. v. 7, n. 11, p. 1315-
1329, 2012.
HARBARTH, S, HAWKEY, P.M, TENOVER, F, STEFANI, S, PANTOSTI, A,
STRUELENS, M.J. Update on screening and clinical diagnosis of methicillin-resistant
Staphylococcus aureus (MRSA). Int J Antimicrob Agents. v. 37, p. 110-117, 2011.
HARDT, K., B. JORIS, S. LEPAGE, R. BRASSEUR, J. O. LAMPEN, J. M. FRERE, A. L.
FINK, AND J. M. GHUYSEN. The penicillin sensory transducer, BlaR, involved in the
inducibility of beta-lactamase synthesis in Bacillus licheniformis is embedded in the plasma
membrane via a four-alpha-helix bundle. Mol Microbiol, v. 23, p. 935-944, 1997.
HAWKEY, P. The origins and molecular basis of antibiotic resistance. BMJ. v. 317, p. 657-
660, 1998.
HIRAMATSU, K.; ITO, T.; HANAKI, H. Mechanisms of methicillin and vancomycin in
Staphylococcus aureus. Ballière Clin Infect Dis, v. 5, p. 221-242, 1999.

73
HIRAMATSU, K.; ITO, T.; TSUBAKISHITA, S.; SASAKI, T.; TAKEUCHI, F.;
MORIMOTO, Y.; KATAYAMA, Y.; MATSUO, M.; KUWAHARA-ARAI, K.;
HISHINUMA, T.; BABA, T. Genomic basis for methicillin resistance in Staphylococcus
aureus. Infect Chemother. v. 45, n. 2, p. 117-136, 2013.
HISATA, K.; KUWAHARA-ARI, K.; YAMANOTO, M.; ITO, T.; NAKATOMI, Y.; CUI, L.;
BABA, T.; TERASAWA, M.; SOTOZONO, C.; KINOSHITA, S.; YAMASHIRO, Y.;
HIRAMATSU, K. Dissemination of methicillin-resistant staphylococci among healthy
Japanese children. J Clin Microbiol, v. 43, n. 7, p. 3364-3372, 2005.
HOU, Z, MENG, J, ZHAO, J, HU, B, LIU, J, YAN, X, JIA, M, LUO, X. Inhibiton of
betalactamase-mediated oxacilin resistance in Staphylococcus aureus by a deoxyribozime. Acta
Pharmacol Sin. v. 28, n. 11, p. 1775-1782, 2007.
ITO, T.; HIRAMATSU, K. Acquisition of methicilin resistance and progression of
multiantibiotic resistance in methicilin-resistant Staphylococcus aureus. Yonsei Med J. v. 39,
p. 526-533, 1998.
ITO, T.; KATAYAMA, Y.; ASADA, K.; MORI, N.; TSUTSUMIMITO, K.; TIENSASITON,
C.; HIRAMATSU, K. Structural comparison of three types of Staphylococcal Cassette
Chromosome mec integrated in the chromosome in methicillin resistant Staphylococcus aureus.
Antimicrob Agents Chemother, v. 45, n. 5, p. 1323-1336, 2001.
ITO,T.; OKUMA,K.; MA,X.X.; YUZAWA,H.; HIRAMATSU,K. Insights on antibiotic
resistance of Staphylococcus aureus from its whole genome: genomic island SCC. Drug Resist
Updat .v. 6, p. 41-52, 2003.
ITO, T.; MA, X.X.; TAKEUCHI, F.; OKUMA, K.; YUZAWA, H.; HIRAMATSU, K. Novel
type V staphylococcal cassette chromosome mec driven by a novel cassette chromosome
recombinase, ccrC. Antimicrob Agents Chemother, v. 48, n. 7, p. 2637-2651, 2004.
JIM HUTCHINSON. Tube Coagulase Test, 2013. Disponível em: http://infectionnet.org,
acesso em 22/11/2013.

74
JARLØV, J.O.; ROSDAHL, V.T. Quantitative determination of beta-lactamase production in
Staphylococcus aureus strains compared to qualitative testing by a microbiological clover leaf
test, a chromogenic cephalosporin test and a iodometric test. Acta Pathol Microbiol Immunol
Scand B, v. 94, p.415-421, 1986.
KAASE, M.; LENGA, S.; FRIEDERICH, S.; SZABADOS, F.; SAKINC, T.; KLEINE, B.;
GATERMANN, S.G. Comparison of phenotipic methods for penicillinase detection in
Staphylococcus aureus. Clin Microbiol Infect, v. 14, p. 614-616, 2008.
KATAYAMA, Y, ZHANG, H, HONG, D, CHAMBERS, F, CHAMBERS, H.F. Jumping the
barrier to β-Lactam resistance in Staphylococcus aureus jumping the barrier to β-lactam
resistance in Staphylococcus aureus. J Bacteriol, v. 185, n. 18, p. 5465–5472, 2003.
KERFF, F., P. CHARLIER, M. L. COLOMBO, E. SAUVAGE, A. BRANS, J. M. FRERE, B.
JORIS, AND E. FONZE. Crystal structure of the sensor domain of the BlaR penicillin receptor
from Bacillus licheniformis. Biochemistry, v. 42, p. 12835-12843, 2003.
KIM, J.Y. Understanding the evolution of methicillin-resistant Staphylococcus aureus. Clin
Microbiol Newsletter, v. 31, n. 3, p. 17-23, 2009.
KIPP, F.; BECKER, K.; PETERS, G.; VON EIFF, C. Evaluation of differen methods to detect
methicillin resistance in small-colony variants of Staphylococcus aureus. J Clin Microbiol, v.
42, n. 3, p. 1277-1279, 2004.
KLUYTMANS,J.; STRUELENS,M. Methicillin resistant Staphylococcus aureus in the
hospital. BMJ. v. 12, n. 1, p. 338-364, 2009.
KONEMAN,E.W.; ALLEN,S.D.; JANDA,W.M.; SCHRECKENBERGER,P.C.; JR,W.C.W.
Diagnóstico microbiológico. Texto e Atlas colorido. 6aed. Guanabara Koogan. Rio de Janeiro
p. 551-579, 2008.
LI, S.; SKOV, R.L.; HAN, X.; LARSEN, A.R.; LARSEN, J.; SØRUM, M.; WULF, M.; VOSS,
A.; HIRAMATSU, K.; ITO, T. Novel types of staphylococcal cassette chromosome mec
elements identified in clonal complex 398 methicillin-resistant Staphylococcusaureus strains.
Antimicrob Agents Chemother, v. 55, n. 6, p. 3046-3050, 2011.

75
LIVERMORE, D.M.; WINSTANLEY, T.G.; SHANNON, K.P. Interpretative reading:
recognizing the unusual and inferring resistance mechanisms from resistance phenotypes. J
Antimicrob Chemother, v. 48, Suppl. 1, p. 87-102, 2001.
LLARRULL, L.I, TOTH, M, CHAMPION, M.M, MOBASHERY, S. Activation of BlaR1
protein of methicillin-resistant Staphylococcus aureus, its proteolytic processing, and recovery
from induction of resistance. J Biol Chem, v. 286, n. 44, p. 38148-38158, 2011.
LOWY, F.D. Antimicrobial resistance: the example of Staphylococcus aureus. J Clin Invest. v.
111, p.1265-1273, 2003.
MARLOWE, E.M, BANKOWSKI, M.J. Conventional and molecular methods for the detection
oc methicilin-resistant Staphylococcus aureus. J Clin Microbiol. v. 49, n. 9, p. S53-S56, 2011.
MCKINNEY, T.K, SHARMA, V.K, CRAIG, W.A, CAMPUS, V. Transcription of the gene
mediating methicillin resistance in Staphylococcus aureus (mecA) is corepressed but not
coinduced by cognate mecA and betalactamase regulators. J Bacteriol, v. 183, n. 23, p. 6862–
6868, 2001.
MEJIA, C.; ZURITA, J.; GUZMAN-BLANCO, M. Epidemiology and surveillance of
methicillinresistant Staphylococcus aureus in Latin America. Braz J Infect Dis, v. 14, Suppl. 2,
p. S79-S86, 2010.
MILHEIRIÇO, C, PORTELINHA, A, KRIPPAHL, L, DE LENCASTRE, H, OLIVEIRA, D.C.
Evidence for a purifying selection acting on the betalactamase locus in epidemic clones of
methicillin-resistant Staphylococcus aureus. BMC Microbiology, v. 11, n. 1, p. 76, 2011.
MOELLERING JR, R.C. MRSA: the first half century. J Antimicrob Chemother, v. 67, p. 4-
11, 2012.
MURRAY, P.R, ROSENTHAL K.S, PFALLER, M.A. Microbiologia Médica. 6ª Edição. São
Paulo, Atheneu, 2010.
NAIMI, T.S.; LEDELL, K. H.; COMO-SABETTI, K.; BORCHARDT, S.M.; BOXRUD, D. J.;
ETIENNE, J.; JOHNSON, S. K.; VANDENESCH, F.; FRIDKIN, S.; O´BOYLE, C.; DANILA,

76
R.N.; LYNFIELD, R. Comparison of community and health care-associated methicillin-
resistant Staphylococcus aureus infection. JAMA, v. 290, p. 2976-2984, 2003.
NEVES, M.C.; ROSSI JUNIOR, O.D.; ALVES, E.C.C.; LEMOS, M.V.F. Detecção de genes
de resistência antimicrobiana em cromossomos e plasmídeos de Staphylococcus spp. Arq Inst
Bio, v. 74, n.3, p. 207-213, 2007.
ODONKOR, S.T.; ADDO, K.K. Evaluation of three methods for detection of methicillin
resistant Staphylococcus aureus (MRSA). Int J Biol Med Res, v. 2, n.4, p. 1031-1034, 2011.
OGSTON, A. Ueber Abscesse. Arch Klin Chir, v. 25, p. 588-600, 1880.
OGSTON, A. Report upon micro-organism in surgical diseases. Brit Med J, v. 1, p. 369-375,
1881.
OGSTON, A. Micrococcus poisoning. J Anat Physiol, v. 16, p. 526-567, 1882 (Parte 1) – v. 17,
p. 24-58, 1883 (Parte 2).
OKUMA, K.; IWAKAWA, K.; TURNIDGE, J.D.; GRUBB, W.B.; BELL, J.M.; O'BRIEN,
F.G.; COOMBS, G.W.; PEARMAN, J.W.; TENOVER, F.C.; KAPI, M.; TIENSASITORN, C.;
ITO, T.; HIRAMATSU, K. Dissemination of new methicillin-resistant Staphylococcus aureus
clones in the community. J Clin Microbiol, v. 40, p. 4289-4294, 2002.
OLIVEIRA, G.A.; FARIA, J.B.; LEVY, C.E.; MAMIZUKA, E.M. Characterization of the
Brazilian endemic clone of methicillin-resistant Staphylococcus aureus (MRSA) from hospitals
throughout Brazil. J Infect Dis. 5:163-70, 2001.
OLIVEIRA, D.C.; MILHEIRIÇO, C.; DE LENCASTRE, H. Redefining a structural variant of
staphylococcal cassette chromosome mec, SCCmec type VI. Antimicrob Agents Chemother, v.
50, n. 10, p. 3457–3459, 2006.
OLIVEIRA, A.A.F.; PINHEIRO JR, J.W.; MOTA, R.A.; CUNHA, M.L.R.S; LOPES, C.A.M;
ROCHA, N.S. Phenotype characterization of Staphylococcus species strains isolated from
buffalo (Bubalus bubalis) milk. J Vet Diag Invest, v. 23, n. 6, p. 1208-1211, 2011.

77
OLSEN, J.E, CHRISTENSEN, H, AARESTRUP, FM. Diversity and evolution of blaZ from
Staphylococcu aureus and coagulase-negative sthaphilococci. J Antimicrob Chemoter. v. 57, p.
450-460, 2006.
ORENSTEIN, A. The discovery and naming of Staphylococcus aureus, 2008. Disponível em:
http://www.antimicrobe.org/h04c.files/history/S-aureus.pdf, acesso em 12/01/2015.
PANTOSTI, A, SANCHINI, A, MONACO, M. Mechanisms of antibiotic resistance in
Staphylococcus aureus. Future Microbiol. v. 2, n. 3, p. 323-334, 2007.
PETINAKI, E; ARVANITI, A; DIMITRACOPOULOS, G; SPILIOPOULOUI. Detection of
mecA, mecR1 and mecI genes among clinical isolates of methicillin-resistant staphylococci by
combined polymerase chain reactions. J Antimicrob Chemother. v. 47, n. 3, p.297-304, 2001.
PITKÄLÄ, A.; SALMIKIVI, L.; BREDBACKA, P.; MYLLYNIEMI, A.L.; KOSKINEN, M.T.
Comparison of tests for detection of beta-lactamse-producing staphylococci. J Clin Microbiol,
v. 45, p. 2031-2033, 2007.
RABELO, M.A. Estudo epidemiológico, genético e de susceptibilidade em Staphylococcus spp.
de amostras de pacientes e profissionais de saúde de Hospital Universitário de Pernambuco.
Dissertação de Mestrado, Recife, 2012.
REMONATTO, G.; CARDOSO, C.M.; MARQUES, C.G.; SILVA, A.E.B.; GELATTI, L.C.;
LEITE, C.F.M. CA-MRSA: um patógeno emergente. News Lab, ed. 80, p. 92-96, 2007.
ROBLES, B.F.; NOBREGA, D.B.; GUIMARÃES, F.F.; WANDERLEY, G.G.; LANGONI,
H. Beta-lactamase detection in Staphylococcus aureus and coagulase-negative Staphylococcus
isolates from bovine mastites. Pesq. Vet. Bras, v. 34, n.4, p. 325-328, 2014.
ROSATO, A.E, KREISWIRTH, B.N, CRAIG, W.A, EISNER, W, CLIMO, M.W, ARCHER,
G.L. mecA-blaZ Corepressors in clinical Staphylococcus aureus isolates. Antimicrob. Agents.
Chemoter, v. 47, n. 4, p. 1460-1463, 2003.
ROSEMBACH, A.J. Mikro-Qrganismen bei den Wund-Infections-Krankheiten des Menschen.
Wiesbaden: J.F. Bergmann, p. 18, 1884.

78
ROZENBAUM, R.; SAMPAIO, M.G.; BATISTA, G.S.; GARIBALDI, A.M.; TERRA,
G.M.F.; SOUZA, M.J.; VIEIRA, E.N.; SILVA-CARVALHO, M.C.; TEIXEIRA, L.A.;
FIGUEIREDO, A.M.S. The first report in Brazil of severe infection caused by community-
acquired methicillin-resistant Staphylococcus aureus (CA-MRSA). Braz J Med Biol Res, v. 42,
n. 8, p. 756-760, 2009.
SAMANT, S.A.; PAI, C.G. Comparative evaluation of β lactamase detection methods in
staphylococci. IJRPBS, v. 3, n. 4, p. 1580-1588, 2012.
SANTOS, A.L, SANTOS D.O, FREITAS C.C, FERREIRA, B.L.A, AFONSO, I.F,
RODRIGUES C.R, CASTRO, H.C. Staphylococcus aureus: visiting a strain of clinical
importance. J Bras Patol Med Lab. v. 43, n. 6, p. 413-423, 2007.
SANTOS, H.B.; MACHADO, D.P.; CAMEY, S.A.; KUCHENBECKER, R.S.; BARTH, A.L.;
WAGNER, M.B. Prevalence and acquisition of MRSA amongst patients admitted to a tertiary-
care hospital in Brazil. BMC Infect Dis, v. 10, p. 328-334, 2010.
SCHITO, G.C. The importance of development of antibiotic resistance in Staphylococcus
aureus. Clin Microbiol Infect, v. 12, n. 1, p. 3-8, 2006.
SHARIATI, L.; VALIDI, M.; TABATABAIEFAR, M.A.; KARIMI, A.; NAFISI, M.R.
Comparison of Real-Time PCR with disk diffusion, agar screen and E-test methods for
detection of methicillin-resistant Staphylococcus aureus. Curr Microbiol, v. 61, p. 520–524,
2010.
SHORE, A.C.; DEASY, E.C.; SLICKERS, P.; BRENNAN, G.; O’CONNEL, B.; MONECKE,
S.; EHRICHT, R.; COLEMAN, D.C. Detection of Staphylococcal Cassette Chromosome mec
type XI carrying highly divergent mecA, mecI, mecR1, blaZ, and ccr genes in human clinical
isolates of clonal complex 130 methicillin-resistant Staphylococcus aureus. Antimicrob Agents
Chemother. v. 55, n.8, p. 3765-3773, 2011.
SHORE, A.C, COLEMAN, D.C. Staphylococcal cassette chromosome mec: Recent advances
and new insights. Int J Med Microbiol. v. 303, n. 6-7, p. 350-359, 2013.

79
SLOOT, N, THOMAS, M, MARRE, R, GATRMANN, S. Purification and characterization of
elastase from S. epidermidis. J Med Microbiol. v. 37, p. 201-205, 1992.
STAPLETON, P.D, TAYLOR, P.W. Methicilin resistance in Staphylococcus aureus:
mechanisms and modulation. Sci Prog. v. 85, p. 57-72, 2002.
SOARES,L.C.; PEREIRA,I.A.; COELHO,S.M.O.; CUNHA,C.M.M.; OLIVEIRA,D.F.B.;
MIRANDA,A.N.; SOUZA,M.M.S. Caracterização fenotípica da resistência a antimicrobianos
e detecção do gene mecA em Staphylococcus spp. coagulase negativos isolados de amostras
animais e humanas. Cien Rural. v. 38, n. 5, p.1346-1350, 2008.
SWENSON, J.M.; LONSWAY, D.; MCALLISTER, S.; THOMPSON, A.; JEVITT, L.; ZHU,
W.; PATEL, J.B. Detection of mecA-mediated resistance using reference and commercial
testing methods in a collection of Staphylococcus aureus expressing boderline oxacillin MICs.
Diag Microbiol Infect Dis, v. 38, n. 5, p. 1346-1350, 2007.
TAKANO, T.; HIGUCHI, W.; OTSUKA, T.; BARANOVICH,T.; ENANY,S.; ISOBE, K.S.H.;
DOHMAE, S.; OZAKI,K.; TAKANO, M.; IWAO, Y.; SHIBUYA, M.; OKUBO, T.; YABE,
S.; SHI, D.; REVA, I.; TENG, L.J.; YAMAMOTO, T. Novel characteristics of community-
acquired methicillin-resistant Staphylococcus aureus belonging to multilocus sequence type 59
in Taiwan. Antimicrob Agents Chemother, v. 52, p. 837-845, 2007.
TAVARES, W. Bactérias gram-positivas problemas: resistência do estafilococo, do enterococo
e do pneumococo aos antimicrobianos. Rev Soc Bras Med Trop, v. 33, n. 3, p. 281-301, 2000.
TENOVER, F. C., WEIGEL, L. M., APPELBAUM, P. C., et al, Vancomycin-resistant
Staphylococcus aureus isolate from a patient in Pennsylvania, Antim Agents and Chemother, v.
48, n. 1, p. 275-280, 2004.
TIWARI, H.K.; SEN, M.R. Emergence of vancomycin resistant Staphylococcus aureus
(VRSA) from a tertiary care hospital from northern part of India. BMC Infect Dis, v. 6, p. 156-
161, 2006.

80
TORTORA, G.J; FUNKE, B.R.; CASE, C.L. Microbiologia. 10ª ed. Artmed, Porto Alegre,
2012.
TRABULSI, L.R.; ALTERTHUM, F. Microbiologia. 5ª ed. Atheneu, São Paulo, 2008.
TRINDADE, P.D.A.; PACHECO, R.L.; COSTA, S.F.; ROSSI, F.; BARONE, A.A.;
MAMIZUKA, E.M.; LEVIN, A.S. Prevalence of SCCmec Type IV in nosocomial bloodstream
isolate of methicillin-resistant Staphylococcus aureus clone. J Clin Microbiol, v. 43, p.
34353437, 2005.
TURLEJ, A.; HRYNIEWICZ, W.; EMPEL, J. Staphylococcal Cassette Chromosome mec
(SCCmec) classification and typing methods: an Overview. Pol J Microbiol, v. 60, n. 2, p. 95-
103, 2011.
VERTONI, P.C. Produção de enterotoxinas por linhagens de Staphylococcus
aureusRosenbach, 1884, isoladas de fossas nasais e de algumas amostras clínicas. Dissertação
de Mestrado, 1991.
VOLADRI, R; KERNODLE, D. Characteristics of a chromosomal gene encoding type
betalactamase in phage group II isolates of Staphylococcus aureus. Antimicrob Agents
Chemoter. v. 42, n. 12, p. 3163-3168, 1998.
WEND ORENT. A Brief History Of Staph, 2006. Disponível em:
http://protomag.com/assets/a-brief-history-of-staph, acesso 22/11/2013.
WOODFORD, N.; LIVERMORE, D.M. Infections caused by Gram-positive bacteria: a review
of the global challenge. J Infect, v. 59, Suppl. 1, p. S4-S16, 2009.
YOSHIDA, R.; KUWAHARA-ARAI, K.; BABA, T.; CUI, L.; RICHARDSON, J.F.;
HIRAMATSU, K. Physiological and molecular analysis of a mecA-negative Staphylococcus
aureus clinical strain that expresses heterogeneous methicillin resistance. J Antimicrob
Chemother, v. 51, p. 247-255, 2003.

81
ZONG, Z.; PENG, C.; LU, X. Diversity of SCCmec elements in methicillin-resistant coagulase-
negative staphylococci clinical isolates. Plos One, v. 6, n. 5, p. e20191, 2011.
ZURITA, J.; MEJIA, C.; GUZMÁN-BLANCO, M. Diagnóstico e teste de sensibilidade para
Staphylococcus aureus resistente a meticilina na América Latina. Braz J Infect Dis, v.14, Suppl.
2, p. S97-S107, 2010.

82
Apêndice

83
Artigo a ser submetido à revista Current Microbiology
Instruções aos autores:http://www.springer.com/life+sciences/microbiology/journal/284
DETECTION OF BETA-LACTAMASE IN Staphylococcus spp ISOLATED FROM
CLINICAL SAMPLES PROVIDED FROM UNIVERSITARY HOSPITAL FROM
RECIFE – PERNAMBUCO – BRAZIL
Natália Regina Souza da Silva ¹, Marcelle Aquino Rabelo ¹, Armando Monteiro Bezerra Neto¹, Wagner Luis
Mendes de Oliveira ², Nilma Cintra Leal ², Maria Amélia Vieira Maciel¹.
1Departamento de Medicina Tropical, Universidade Federal de Pernambuco, Recife, Pernambuco, Brasil.
²Centro de Pesquisas Aggeu Magalhães – CPqAM- FIOCRUZ – Recife, Pernambuco, Brasil.
Endereço para Correspondência:
Natália Regina Souza da Silva
Av. Prof. Moraes Rego, 1235- Cidade Universitária, Recife-PE. CEP: 50670-901
e-mail: [email protected]
Resumo: Apesar da detecção do gene mecA por técnicas moleculares ser considerado o padrão ouro para a
identificação das cepas de Staphlylococcus resistentes à meticilina (MRS), verifica-se discrepância entre a
apresentação fenotípica da resistência à meticilina nessas cepas e a presença do gene mecA. O presente estudo teve
como objetivo verificar se existe diferença na frequência da detecção de betalactamase entre isolados clínicos de
Staphylococcus spp. resistentes à meticilina portadores do gene mecA e os não portadores desse gene. Foram
estudados 47 isolados provenientes de amostras clínicas de hospital universitário da cidade do Recife. Essas
amostras foram submetidas aos testes de susceptibilidade antimicrobiana, detecção do gene mecA para separação
dos grupos de comparação e posterior detecção da presença de betalactamase através dos testes de cefinase em
disco, Clover-Leaf e detecção do gene blaZ por PCR. Foram observadas diferenças na frequência quando utilizados
os testes fenotípicos para detecção. No teste de cefinase em disco, foram positivos 33,3% dos isolados portadores
do gene mecA e 56,25% dos não portadores. No teste de Clover-Leaf, foi detectada a produção de betalactamase
em 33,3% dos isolados portadores do gene mecA e 65,62% dos não portadores. Na pesquisa do gene blaZ, no
entanto, não foi verificada diferença significativa nas frequências de detecção entre os grupos.
Palavras-chave: betalactamase, Staphylococcus, resistência.
Summary: Despite the fact that detection of mecA gene by molecular techniques is considered the gold standard
for identifying Staphylococcus strains methicillin-resistant (MRS), there is discrepancy between the phenotypic
presentation of methicillin resistance in these strains and the presence of mecA gene. This study aimed to determine
whether there are differences in the frequency of beta-lactamese detection of clinical isolates of Staphylococcus
spp. methicillin-resistant carriers of mecA gene and non-carriers of the gene. 47 isolates from clinical samples

84
provided from the university hospital in Recife were studied. These samples were subjected to antimicrobial
susceptibility testing, detection of the mecA gene for separation of comparison sequences and subsequent detection
of beta-lactamase by means of tests on cefinase disc, Clover-Leaf and detetection of blaZ gene by PCR. Differences
were observed in the frequency when phenotypic tests were used for detection. In cefinase test disk, were positive
33.3% of the isolates carrying the mecA gene and 56.25% of non-carriers. In the Clover-Leaf test, the production
of beta-lactamase was detected in 33.3% of isolates carrying the mecA gene and 65.62% of non-carriers. In search
of blaZ gene, however, there was no significant difference in the detection frequency between groups.
Keywords: beta-lactamase, Staphylococcus, resistance.
Introduction
The methicillin resistance, which had the first reports described in the 1960’s, gave more importance to
Staphylococcus gender as a major human pathogen [15, 27]. Methicilin-resistant Staphylococcus (MRS) causes
infections difficult to treat because they are resistant to most antibiotics used in clinical practice, such as beta-
lactams, aminoglycosides and macrolides [4].
Clinical isolates of S. aureus from patients in intensive care units across the country and from isolate
blood cultures around the world report that these bacterial strains are becoming more resistant to a greater number
of antimicrobial agents. Inevitably, this left less effective bactericidal antibiotics to treat these infections,
increasing significantly the risk of death in patients. As soon as new antibiotics are introduced, staphylococci
develop efficient mechanisms to neutralize them. [12]
Staphylococci developed resistance to β-lactams by two mechanisms, the expression of β-lactamase and
the production of PBP2a encoded by mecA gene and blaZ, repectively [7, 14]. The detection of the mecA gene by
molecular techniques has been considered the gold standard for indentifying the strains of methicillin-resistant
Staphylococcus (MRS)[24, 26], but in some studies there is discrepancy between the phenotypic presentation of
these strains and the presence of mecA gene. [1, 3, 28].
The staphylococcal beta-lactamase is coded by the structural gene, blaZ, which is regulated by a DNA
binding protein, Blal, and a signal transduces BlaR1 [7]. In the presence of beta-lactam antibiotics, the extracelular
domain of BlaR1 sensor may be acylated, so that the signal can be transduced by the membrane area, crossing it
to activate the intracellular domain. The activated BlaR1 acts as a protease and disturbs or alters the formation of
dimer Blal. Without the Blal connection to the operator site, the transcription of both blaZ and blaR1-blal may
initiate the betalactamase synthesis.
Different tests may be used to assess the production of betalactamase in staphylococci. Qualitative
procedures, such as detection of beta-lactamase production using Cefinase discs, where the reaction is based on
the production of a colored compound when the disc comes in contact with a beta-lactamase producing bacteria.
Another procedure used is Clover-Leaf test, based on the formation of a distorted area in the inhibition halo
penicillin disk. [2, 21].
Despite the importance that beta-lactamase has in infections caused by Stahylococcus spp., few studies
in Brazil mention the occurrence of this enzyme related to the pattern of bacterial resistance, nor their genetic
characterization. With the increased cases of phenotypically methicillin-resistant strains, both the presence of

85
mecA gene, observed in patients hospitalized in a university hospital in the city of Recife – PE, we seek through
this work, to identify the occurrence of beta-lactamase as responsible for the pattern of methicillin resistance found
in these strains provided from clinical isolates.
Materials and methods
Obtainment of samples
Forty-seven clinical isolates were used, provided from Hospital das Clínicas, UFPE, collected during the
period from April 2011 to July 2013. All the isolates were stored in Bacterioteca from Bacteriology Laboratory of
the Department of Tropical Medicine – UFPE. These isolates were provided by the heads of bacteriology
laboratories of the hospital, after bacterial identification. Thus, the collection of the samples was the hospital’s
responsibility. These samples were originated from catheter tip, drains, blood, abscess, surgical wound and various
secretions.
Isolates selection
The methicillin resistance in phenotypic testing was used as a criterion for inclusion in the study. All
isolates had previously been identified and subjected to PCR of mecA before being stored in Bacterioteca from
the Laboratory of the Department of Tropical Medicine – UFPE.
For this identification, the isolates were submitted to disk diffusion testing of oxacillin, cefoxitin disk
diffusion and oxacillin screening. The isolates that showed resistance to at least one of the techniques used were
subjected to PCR for detection of the mecA gene, in which were used for amplification the primers P1, 5'-
GGTCCCATTAACTCTGAAG-3’and P3, 5’-AGTTCTGCAGTACCGGATTTGC-3’ described by PETINAKI et
al. (2001), that result in 1.046pb fragments.
Cefinase disc test
The cefinase disc test was conducted using the BBL cefinase discs (BD) according to the manufacture’s
specifications. The disks were spread in a sterile petri dish. Then, each disc has been moistened with a drop
(approx. 30μL) of sterile miliQ water and, with a platinum loop, there was a smear of test colonies in cefinase disk.
The positive result was detected by color change from yellow to pink, presented by the disc. This color change is
due to the compound Nitrocefin, a chromogen cephalosporin wich changes color as the amido boung in the beta-
lactam ring is hydrolyzed by a betalactamase. For Staphylococcus spp. we waited up to an hour for the color
change to happen. As a positive control we used the MRSA ATCC 33591 strain and as negative control we used
the MSSA ATCC 6538 strain.
Clover-Leaf test

86
The Clover-Leaf test is a modified Hodge test and was conducted according to the technique from Bergan
et al. (1997). First an inoculum was prepared from an isolated negative beta-lactamase (strain ATCC 25923),
according to 0.5 on the McFarland scale by direct suspension. After, the sowing was conducted in this carpet
isolated on Mueller-Hinton Agar and placed on 10U penicillin disc (Laborclin) in the center of the plate. With a
platinum loop, stretches were made from the center to the periphery of the petri dish without touching the penicillin
disk. The plate was incubated from 18 to 24 hours, at 35ºC and then the appearance of a distortion area was
observed at the inhibition zone of the isolates considered positives. As a positive control of the test, the isolate
1171 was used. It produces betalactamase, from Bacterioteca of CPqAM/FIOCRUZ. As a negative control, the
strain MSSA ATCC 25923 was used.
Total DNA extraction
For the extraction of DNA from Staphylococcus, the technique of boiling extraction was used, in which
the DNA is extracted from 1ml of bacterial culture previously grown in BHI broth at 37ºC for 24 hours and
centrifuged at 14,000 rpm for five minutes at 4ºC. Then, the supernatant was ignored and the pellet was
resuspended in 200μL of sterile MilliQ water. The suspension was incubated at 100C for 10 min, followed by the
centrifugation at 14,000rpm /4ºC for five minutes. After centrifugation, 100μL of supernatant was collected in a
new tube and stored ad -20ºC
Beta-lactamase gene identification by PCR
For amplification of the blaZ gene, were used the primers F1, 5'- GATAAGAGATTTGCCTATGC-3'
and R1, 5'-GCATATGTTATTGCTTGACC-3' (MILHEIRIÇO et al., 2011) resulting in 533pb. Fragments. The
PCR reactions were performed individually in a final volume of 25μL in each tube containing: 5μL of genomic
DNA, 20pmol of each primer, 1.5mM, dNTP 200μM, 1U of Go Taq DNA polymerase (Promega, Brazil) and 5μL
of Go Green Taq polymerase buffer (Promega, Brazil). The reactions were performed in a thermocyler (Biometra),
programmed initially to initial denaturation for four minutes at 92ºC, then 30 thermal cycles of one minute at 92ºC,
one minute at 55, one minute at 72ºC and final step, seven minutes at 72ºC. As a negative control, a tube was used,
containing all components of the mixture and the absence of template DNA. For positive control, we used a strain
of S. aureus resistant to methicillin (isolate code: 1171 from Bacterioteca of CPqAM/Fiocruz-PE).
The amplification product was subjected to electrophoresis on 1% agarose gel and stained with ethidium
bromide (2μG/mL), visualized by UV transilluminator and scanned using Kodak 1D software version 3.5.2
(Scientific Imaging Systems, USA). The molecular weight market is 100bp (Invitrogen).
Ethical considerations
This study is part of CAAE project – 0490.0.172.000-11 registration CEP/CCS/UFPE No 009/11 (Annex
III) submitted for analysis by the Research Ethics Committee (Comitê de Ética em Pesquisa, CEP) of the Health
Sciences Center (Centro de Ciências da Saúde, CCS) from UFPE, with the permission and cooperation of the
Hospital Infection Control Commission (Comissão de Controle de Infecção Hospitalar, CCIH) of the involved
hospital. There was no term of Informed Consent (IC) for patients because it’s secondary samples.

87
Statistical analysis
Microbiological data were statistically analyzed through frequency distribution and X² test, using Epi Info
version 6.04.
Results
We analyzed 47 (forty-seven) Staphylococcus spp., 13 (thirteen) isolates identified as coagulase negative
Staphylococcus and 34 (thirty-four) identified as Staphylococcus aureus. All selected isolates were resistant to at
least one of phenotypic testing of methicillin resistance (disk diffusion oxacillin, disk diffusion cefoxitin and
screening oxacillin) and were subjected to PCR for detection of the mecA gene and separated into groups to study
according to the result of PCR.
According to the results of PCR, the isolates were selected into two groups: the phenotypically resistant
isolates and carrying the mecA gene (15 isolates) and phenotypically resistant isolates and not carrying the mecA
gene (32 isolates).
In isolates belonging to the carrier group mecA gene, only one isolate was resistant only to cefoxitin disk
diffusion test, the others were resistant to all three tests. Referring to the isolates that belong to the non carrier
mecA gene group, there was a heterogeneous distribution for the results presented in phenotypic tests of resistance
to methicillin, 37% were resistant to only one test, 22% were resistant to both tests and 37% were resistant to all
three tests.
From the isolates that showed resistance to two different phenotypic tests, five were resistant to cefoxitin
disk diffusion testing and to methicillin screening and three isolates demonstrated resistance to only one
phenotypic test, four were resistant only to disk diffusion oxacillin test, five were resistant only to disk diffusion
test, while two were resistant only in the methicillin screening test.
On the cefinase disc test for phenotypical detection of beta-lactamase, positivity was found only in five
of the 15 isolates (33.33%) belonging to the mecA gen carrier group. In the group not carrying the mecA gene, the
test positivity was observed for 18 of the 32 isolates (56,25%). The test is considered positive when the cefinase
disc changes color, acquiring a reddish hue (Figure 1).
In the Clover-Leaf test for phenotypic detection of beta-lactamase production, the test positivity was also
observed in five of the 15 isolates analyzed (33.33%) belonging to the mecA gene carrier group. However, it is
noted that the isolate 33STVd, positive for the cefinase disc test, was negative to the Clover-Leaf test, while isolate
40, that was negative for the cefinase disc test, was positive for the Clover-Leaf test.
In the group of isolates not carrying the mecA gene, it was noticed that 21 of the 32 samples (65,62%)
were positive for the Clover-Leaf test. It was also noticed that isolates 24, 105 and 117, that were negative for the
cefinase disc test, were positive for the Clover-Leaf test. The test was considered positive when a distortion zone
occurred in the inhibition halo of the penicillin disc (figure 2).
The PCR reaction for blaZ gene confirmed the presence of beta-lactamase production gene in eight of the
15 isolates from the mecA gene carrier group, compromising a percentage of 53.33% of the isolates studied. In the
group not carrying the mecA gene, the presence of the gene was noticed in 12 of the 32 isolates studied,

88
compromising a percentage of 37.5%. Amplification of the expected fragment size was observed, approximately
533pb (figure 3).
Several isolates were positive for the studied phenotypic methods, however they were negative in the
research of gene blaZ through the PCR (Table 1).
Using the PCR reaction as gold standard, we calculated the sensibility and specificity of the different
analytical methods used for beta-lactamase production identification (Table 2). Thus it appears that Clover-Leaf
test has greater sensitivity for detection of beta-lactamase production in this study, and both tests have identical
specificity detection. It is also noticed that this specificity is low for detection.
Comparing the groups, “MecA gene carriers” versus “Not carrying mecA gene”, there was no significant
difference in the distribution of frequencies, according to the χ² test (p >0,05).
Discussion
Since the emergence of the first antibiotics, Staphylococcus spp. has acquired mechanisms of resistance
to these drugs, and thus able to persist in the hospital, becoming one of the main agents of acquired bacteremia in
this environment [11, 27]. The development of resistance mechanisms limits the possibilities of patient treatment,
and thereby crucial to determine whether the Staphylococcus spp. isolates are resistant to methicillin/oxacilina,
both for treatment and outbreak controls [1, 10].
Although the mecA gene is considered the gold standard for the detection of methicillin resistance, this
method is most commonly used in reference laboratories and not in laboratories of microbiological routine tests.
These laboratories often use phenotypic methods for identification of methicillin resistance [16]. In some studies,
however, there is discrepancy between the phenotypic display of the strains and the presence of gene mecA [1, 3,
28].
In this study there was a high rate of discrepancy between the mecA gene and the phenotypic testing of
methicillin resistance (oxacillin disc-diffusion, cefoxitin discdiffusion and oxacillin screening), whereas 68% of
the isolates resistant to at least one of the phenotypic tests did not have the gene. Ghoshak et al. (2004) also found
inconsistencies between results of phenotypic tests and the presence of mecA gene, reaching the conclusion that
the overproduction of beta-lactamase would be responsible for the induction of resistance to oxacillin.
Therefore, phenotypic tests were performed for detection of beta-lactamase production and to research
the blaZ gene in these isolates. In many studies the betalactamase production rate range from 55.7% to 92.6% for
staphylococci [22].
In this study lower detection frequencies were found, when the isolates totality was considered, for all the
tests performed, since the in the cefinase disc test the production of betalactamase was detected in 48,9% of the
isolates, in the Clover-Leaf test it was detected in 55,3% of the isolates and in the PCR for detection of blaZ gene,
42.55% of the isolates. In this study, five isolates had the blaZ gene, but did not demonstrate betalactamase
production in any of the phenotypic tests, this fact is common because the presence of the gene does not necessarily
guarantee its expression. The expression of this factor should be evaluated by RT-PCR and qRT-PCR techniques.
It was also found that twelve isolates showed beta-lactamase production in both phenotypic tests
performed, but they did not have the blaZ gene. This fact can be explained by the presence of polymorphisms in
the gene, which can generate different sequences as alredy described by Olsen et al (2006) and Milheiriço et al

89
(2011). In her studies, El Feghaly et al (2012) used three different sequencies that encode different areas of the
blaZ gene, in order to account the discrepancies found.
The cefinase disc test showed low sensitivity (55%) compared to the gold standard used, lower than the
results found by Pitkälä et al (2007), reporting sensitivity variation from 62% to 100% for the test. However, it is
higher than the sensitivity found by Kaase et al (2008) that reported sensitivity of only 39% using this method.
Despite the sensitivity dependent on conditions of the operator and the type of methodology used, the cefinase test
is highly used because of speed of detection and low operating costs [5, 20].
The Clover-Leaf test showed the highest sensitivity in this study (70%), a result which agrees with other
studies that consider the test one of the most sensitive to beta- lactamase, reaching in some studies rates between
94% and 98% sensitivity. As well as the cefinase test, which is easy to perform and has a low cost [2, 5, 8, 20].
Robles et al. (2014) in his studies does not recommend taking PCR for blaZ gene as a gold standard for
detection of beta-lactamase production, since the beta-lactamase producing phenotype could result from the
product of more than one gene and reinforces the idea that the gene presence does not guarantee its phenotypic
expression. In addition, he defends the use of cefinase disc tests and Clover-Leaf for the detection of beta-lactamase
production because they usually show good sensitivity. These founds may cooperate with the results of this study,
in which were found low sensitivity rates in the phenotypic tests for using PCR for detection of the blaZ gene as a
gold standard.
Comparing gene blaZ frequencies among the study groups used, a significant difference was not found
comparing the groups, which may suggest both that the methicillin resistance phenotype is due to another factor
other than the presence of betalactamase, or the lower detection of the presence of the gene may be linked to
differences in the sequences reported by El Feghaly et al. (2012).
The PCR reaction for detection of blaZ gene is very specific, although it must not be used as a gold
standard for resistance identification, since beta-lactamase expression may be due to factors other thansimply the
presence of this gene. In this way, it indicates the use of phenotypic tests for detection of beta-lactamase
production, for having low cost and easy operation as well as demonstrating good sensitivity.
References
[1] ARAJ, G.F, TALHOUK, R.S, SIMAAN, C.J, MAASAD, M.J. Discrepancies between mecA PCR and
conventional tests used for detection of methicilin resistant Staphylococcus aureus.Int J Antimicrob Agents, v. 11,
p. 47-52, 1999.
[2] BERGAN, T.; BRUNN, J.N.; DIGRANES, A.; LINGAAS, E.; MELBY, K.K.; SANDER, J. Susceptibility
testing of bacteria and fungi. Report from: “the Norwegian Working Group on Antibiotics”. Scand J Infect (Suppl.)
v. 103, p. 1-36. 1997.
[3] CUNY, C.; LAYER, F.; STROMMENGER, B.; WITTE, W. Rare occurrence of methicillin- resistant
Staphylococcus aureus CC130 with a novel mecA homologue in humans in Germany. PLoS ONE, v. 6, n. 9, p.
e24360, 2011.
[4] DURAN, N, OZER, B, DURAN, G.G, ONLEY, Y, DEMIR, C. Antibiotic resistance genes & suscebility
patterns in staphylococci. Indian J Med Res. v. 135, p. 389-396, 2012.

90
[5] EL FEGHALY, R.E.; STAMM, J.E.; FRITZ, S.A.; BURNHAM, C.A.D. Presence of the blaZ beta-lactamase
gene in isolates of Staphylococcus aureus that appear penicillin susceptible by conventional phenotypic methods.
Diag Microbiol Infect Dis, v. 74, p. 388-393, 2012.
[6] GHOSHAL, U.; PRASSAD, K.N.; SINGH, M.; TIWARI, D.P.; AYYAGARI, A. A Comparative evaluation
of phenotypic and molecular methods for the detection of oxacillin resistancce in coagulase-negative
staphylococci. J Infect Chemother, v. 10, p. 86-89, 2004.
[7] HAO, H, DAI, M, WANG, Y, HUANG, L, YUAN, Z. Key genetics elements and regulation systems in
methicillin-resistant Staphylococcus aureus. Future Microb. v. 7, n. 11, p. 1315-1329, 2012.
[8] JARLØV, J.O.; ROSDAHL, V.T. Quantitative determination of beta-lactamase production in Staphylococcus
aureus strains compared to qualitative testing by a microbiological clover leaf test, a chromogenic cephalosporin
test and a iodometric test. Acta Pathol Microbiol Immunol Scand B, v. 94, p.415-421, 1986.
[9] KAASE, M.; LENGA, S.; FRIEDERICH, S.; SZABADOS, F.; SAKINC, T.; KLEINE, B.; GATERMANN,
S.G. Comparison of phenotipic methods for penicillinase detection in Staphylococcus aureus. Clin Microbiol
Infect, v. 14, p. 614-616, 2008.
[10] KIPP, F.; BECKER, K.; PETERS, G.; VON EIFF, C. Evaluation of differen methods to detect methicillin
resistance in small-colony variants of Staphylococcus aureus. J Clin Microbiol, v. 42, n. 3, p. 1277-1279, 2004.
[11] KLUYTMANS,J.;STRUELENS,M.MethicillinresistantStaphylococcusaureusinthehospital.BMJ.v. 12, n. 1,
p.338-364,2009.
[12] LOWY, F.D. Antimicrobial resistance: the example of Staphylococcus aureus. J Clin Invest. v. 111, p.1265-
1273, 2003.
[13] MARLOWE, E.M, BANKOWSKI, M.J. Conventional and molecular methods for the detection oc methicilin-
resistant Staphylococcus aureus. J Clin Microbiol. v. 49, n. 9, p. S53-S56, 2011.
[14] MILHEIRIÇO, C, PORTELINHA, A, KRIPPAHL, L, DE LENCASTRE, H, OLIVEIRA, D.C. Evidence for
a purifying selection acting on the betalactamase locus in epidemic clones of methicillin-resistant Staphylococcus
aureus. BMC Microbiology, v. 11, n. 1, p. 76, 2011.
[15] MOELLERING JR, R.C. MRSA: the first half century. J Antimicrob Chemother, v. 67, p. 4-11, 2012.
[16] ODONKOR, S.T.; ADDO, K.K. Evaluation of three methods for detection of methicillin resistant
Staphylococcus aureus (MRSA). Int J Biol Med Res, v. 2, n.4, p. 1031-1034, 2011.
[17] OLIVEIRA, A.A.F.; PINHEIRO JR, J.W.; MOTA, R.A.; CUNHA, M.L.R.S; LOPES, C.A.M; ROCHA, N.S.
Phenotype characterization of Staphylococcus species strains isolated from buffalo (Bubalus bubalis) milk. J Vet
Diag Invest, v. 23, n. 6, p. 1208-1211, 2011.
[18] OLSEN, J.E, CHRISTENSEN, H, AARESTRUP, FM. Diversity and evolution of blaZ from Staphylococcu
aureus and coagulase-negative sthaphilococci. J Antimicrob Chemoter. v. 57, p. 450-460, 2006.
[19]PETINAKI, E, ARVANITI, A, DIMITRACOPOULOSG, SPILIOPOULOUI. Detection of mecA, mecR1 and
mecI genes among clinical isolates of methicillin-resistant staphylococci by combined polymerase chain reactions
.J Antimicrob Chemother. v. 47, n. 3, p.297-304,2001.
[20] PITKÄLÄ, A.; SALMIKIVI, L.; BREDBACKA, P.; MYLLYNIEMI, A.L.; KOSKINEN, M.T. Comparison
of tests for detection of beta-lactamse-producing staphylococci. J Clin Microbiol, v. 45, p. 2031-2033, 2007.

91
[21] ROBLES, B.F.; NOBREGA, D.B.; GUIMARÃES, F.F.; WANDERLEY, G.G.; LANGONI, H. Beta-
lactamase detection in Staphylococcus aureus and coagulase-negative Staphylococcus isolates from bovine
mastites. Pesq. Vet. Bras, v. 34, n.4, p. 325-328, 2014.
[22] SAMANT, S.A.; PAI, C.G. Comparative evaluation of β lactamase detection methods in staphylococci.
IJRPBS, v. 3, n. 4, p. 1580-1588, 2012.
[23] SANTOS, A.L, SANTOS D.O, FREITAS C.C, FERREIRA, B.L.A, AFONSO, I.F, RODRIGUES C.R,
CASTRO, H.C. Staphylococcus aureus: visiting a strain of clinical importance. J Bras Patol Med Lab. v. 43, n. 6,
p. 413-423, 2007.
[24] SHARIATI, L.; VALIDI, M.; TABATABAIEFAR, M.A.; KARIMI, A.; NAFISI, M.R. Comparison of Real-
Time PCR with disk diffusion, agar screen and E-test methods for detection of methicillin-resistant Staphylococcus
aureus. Curr Microbiol, v. 61, p. 520–524, 2010.
[25] STAPLETON, P.D, TAYLOR, P.W. Methicilin resistance in Staphylococcus aureus: mechanisms and
modulation. Sci Prog. v. 85, p. 57-72, 2002.
[26] SWENSON, J.M.; LONSWAY, D.; MCALLISTER, S.; THOMPSON, A.; JEVITT, L.; ZHU, W.; PATEL,
J.B. Detection of mecA-mediated resistance using reference and commercial testing methods in a collection of
Staphylococcus aureus expressing boderline oxacillin MICs. Diag Microbiol Infect Dis, v. 38, n. 5, p. 1346-1350,
2007.
[27] TAVARES, W. Bactérias gram-positivas problemas: resistência do estafilococo, do enterococo e do
pneumococo aos antimicrobianos. Rev Soc Bras Med Trop, v. 33, n. 3, p. 281-301, 2000.
[28] YOSHIDA, R.; KUWAHARA-ARAI, K.; BABA, T.; CUI, L.; RICHARDSON, J.F.; HIRAMATSU, K.
Physiological and molecular analysis of a mecA-negative Staphylococcus aureus clinical strain that expresses
heterogeneous methicillin resistance. J Antimicrob Chemother, v. 51, p. 247-255, 2003.
Picture 1: Cefinase disc test
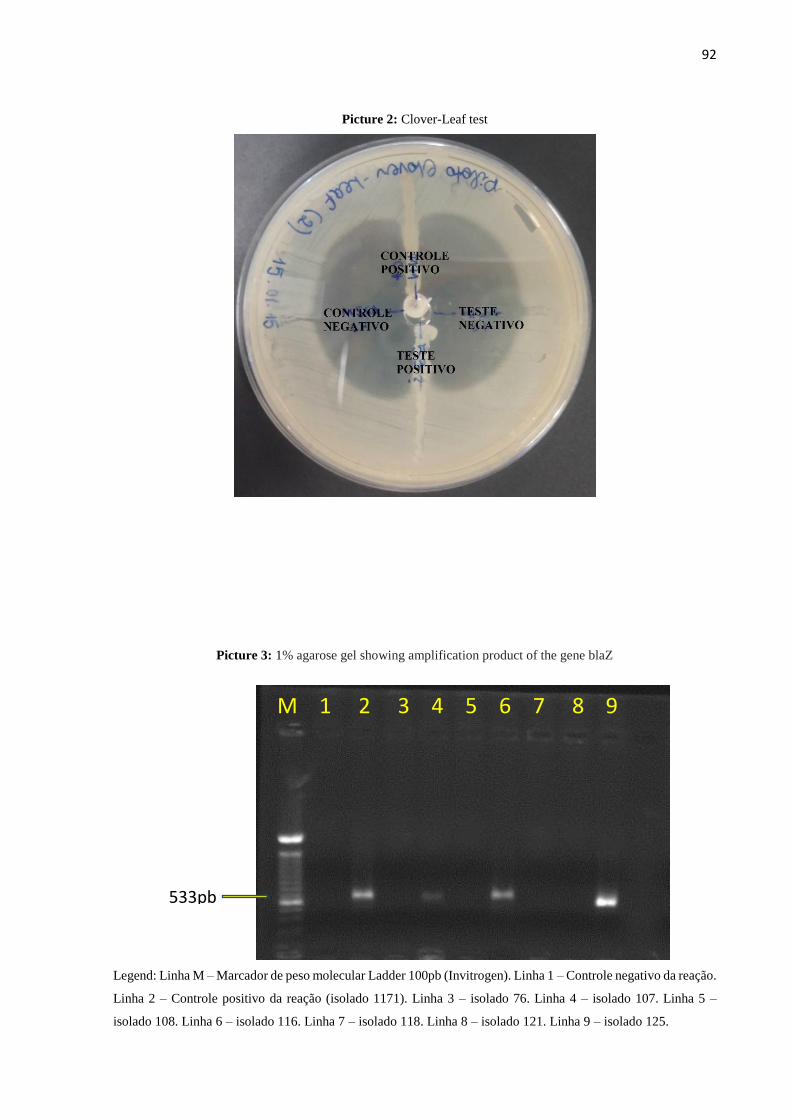
92
Picture 2: Clover-Leaf test
Picture 3: 1% agarose gel showing amplification product of the gene blaZ
Legend: Linha M – Marcador de peso molecular Ladder 100pb (Invitrogen). Linha 1 – Controle negativo da reação.
Linha 2 – Controle positivo da reação (isolado 1171). Linha 3 – isolado 76. Linha 4 – isolado 107. Linha 5 –
isolado 108. Linha 6 – isolado 116. Linha 7 – isolado 118. Linha 8 – isolado 121. Linha 9 – isolado 125.
M 1 2 3 4 5 6 7 8 9
533pb

93
Table 1: Frequency distribution of beta-lactamase detection according to PCR blaZ gene, Cefinase disc test and
Clover-Leaf test in isolated Staphylococcus spp. methicillin-resistant with and without the presence of the mecA
gene.
Isolates with the presence of the
mecA gene
Isolates without the presence of
the mecA gene.
N % (do total) n % (do total)
GeneblaZ +
-
8
7
17,0
14,9
12
20
25,5
42,6
Cefinase +
-
5
10
10,6
21,3
18
14
38,3
29,8
Clover-Leaf +
-
5
10
10,6
21,3
21
11
44,7
23,4
Table 2: Sensibility and specificity of the analytical methods to detect beta-lactamase
Method Sensibility Especificity
Cefinase
Clover-Leaf
55%
70%
44,4%
44,4%
Gold Standar = PCR for detection of blaz gene

94
Anexos

95
ANEXO I
Isolados mecA +
Número do
Isolado
Espécie Qtd de
testes
fenotípicos
de
resitência à
meticilina
positivos
Teste de
Cefinase
em Disco
Teste de
Clover
Leaf
PCR blaZ
23SNLd Staphylococcus
coag neg
3 - - +
33STVd S. aureus 3 + - +
35HAd S. aureus 3 - - -
37FOVd Staphylococcus
coag neg
3 - - -
42HAd S. aureus 3 - - -
49PCLd S. aureus 3 - - -
61PCAd Staphylococcus
coag neg
1 - - +
77PCLd Staphylococcus
coag neg
3 - - -
40 S. aureus 3 - + +
53 S. aureus 2 + + +
59 S. aureus 3 - - -
73 S. aureus 3 + + -
98 S. aureus 3 - - +
102 Staphylococcus
coag neg
3 + + +
125 S. aureus 3 + + +
96
ANEXO II
Isolados mecA –
Número do
Isolado
Espécie Qtd de testes
fenotípicos
de resitência
à meticilina
positivos
Teste de
Cefinase
em Disco
Teste de
Clover
Leaf
PCR blaZ
6PCLd Staphylococcus
coag neg
2 + + +
16SNLd S. aureus 1 - - -
18PCAd Staphylococcus
coag neg
3 - - -
21SNLd S. aureus 1 - - -
22SNLd S. aureus 1 - - +
40STAd Staphylococcus
coag neg
3 - - +
72HAd S. aureus 3 - - -
76PCAd Staphylococcus
coag neg
3 - - -
85PCVd Staphylococcus
coag neg
3 - - -
16 S. aureus 3 - - -
24 S. aureus 3 - + +
36 S. aureus 3 + + -
37 S. aureus 3 + + +
44 S. aureus 2 + + -
46 S. aureus 3 + + -
48 S. aureus 2 - - -
62 S. aureus 1 + + -
70 S. aureus 1 + + -
74 S. aureus 3 + + -
75 S. aureus 3 + + -

97
76 S. aureus 2 + + -
105 S. aureus 1 - + +
107 Staphylococcus
coag neg
1 + + +
108 S. aureus 1 + + -
111 Staphylococcus
coag neg
2 + + +
112 S. aureus 1 + + -
114 S. aureus 2 + + +
115 S. aureus 2 + + +
116 S. aureus 1 + + +
117 Staphylococcus
coag neg
3 - + +
118 S. aureus 1 - - -
121 S. aureus 1 + + -

98
ANEXO III – APROVAÇÃO DO COMITÊ DE ÉTICA

99
ANEXO IV – CARTA DE ANUÊNCIA ULAB-HC


















