
MIGUEL FERNANDES KOLODIUK DIETA E COMPORTAMENTO DE ...
44
MIGUEL FERNANDES KOLODIUK DIETA E COMPORTAMENTO DE FORRAGEIO DE DUAS ESPÉCIES DE TROPIDURUS (SQUAMATA, TROPIDURIDAE) EM UMA ÁREA DE CAATINGA DO NORDESTE DO BRASIL Dissertação apresentada à Universidade Federal do Rio Grande do Norte, para obtenção do título de Mestre em Psicobiologia. Natal 2008
Transcript of MIGUEL FERNANDES KOLODIUK DIETA E COMPORTAMENTO DE ...

MIGUEL FERNANDES KOLODIUK
DIETA E COMPORTAMENTO DE FORRAGEIO DE DUAS ESPÉCIES DE
TROPIDURUS (SQUAMATA, TROPIDURIDAE) EM UMA ÁREA DE CAATINGA
DO NORDESTE DO BRASIL
Dissertação apresentada à Universidade
Federal do Rio Grande do Norte, para
obtenção do título de Mestre em
Psicobiologia.
Natal
2008

Miguel Fernandes Kolodiuk
DIETA E COMPORTAMENTO DE FORRAGEIO DE DUAS ESPÉCIES DE
TROPIDURUS (SQUAMATA, TROPIDURIDAE) EM UMA ÁREA DE CAATINGA
DO NORDESTE DO BRASIL
Dissertação apresentada à Universidade
Federal do Rio Grande do Norte, para
obtenção do título de Mestre em
Psicobiologia.
Orientadora: Dra Eliza Maria Xavier Freire
Natal
2008

Título: “Dieta e compotamento de forrageio de duas espécies de Tropidurus
(Squamata,Tropiduridae) em uma área de Caatinga do Nordeste do Brasil”
Autor: Miguel Fernandes Kolodiuk
Data: 28 de agosto de 2008

AGRADECIMENTOS
À Universidade Federal do Rio Grande do Norte e ao Programa de Pós-Graduação em
Psicobiologia, onde tive a oportunidade de contar com uma excelente equipe de professores e
funcionários.
Ao PELD/CNPq pelo fomento da pesquisa, e à administração e funcionários da
Estação Ecológica do Seridó – ESEC/IBAMA pela permissão da pesquisa, e pelo alojamento
disponibilizado durante todo o trabalho de campo.
Este trabalho não poderia ser realizado sem a contribuição de inúmeras pessoas que de
alguma forma me ajudaram em seu desenvolvimento. Em especial agradeço a minha
orientadora, Profa. Dra. Eliza Maria Xavier Freire que me orienta desde a minha iniciação
científica e também contribuiu muito em minha formação pessoal, ela será sempre o meu
exemplo de amor à ciência e com certeza serei sempre grato.
A todos os professores do Programa de Pós-Graduação em Psicobiologia pelos
ensinamentos e aos colegas de turma do mestrado.
Ao meu amigo e parceiro de atividades em campo, o doutorando Leonardo Ribeiro,
que foi imprescindível em todas as etapas deste trabalho; agradeço pelo companheirismo,
organização das viagens, e principalmente, pelos ensinamentos.
Ao professor Adalberto Antônio Varela-Freire, do Departamento de Microbiologia e
Parasitologia da UFRN pela ajuda nas identificações dos artrópodes do ambiente e do
conteúdo estomacal.
Aos atuais companheiros do Laboratório de Herpetologia; Carolina Lisboa, Pablo
Souza, Mariana Capistrano, Leonardo Ribeiro e Melissa Gogliath que me proporcionaram
ótimos momentos de trabalho e de descontração.
A Edson Santana, Técnico de laboratório da UFRN, que contribuiu muito nos
conduzindo durante as viagens a campo, sempre com sua alegria contagiante.
Agradeço, em especial, aos meus pais, Ricardo Miguel e Ana Luiza pelo amor e
carinho que sempre me deram e por me incentivarem ao estudo, possibilitando uma formação
profissional e principalmente pessoal.
Aos meus irmãos André e Fernanda pelo amor e amizade com que convivemos.
Aos meus Abuelos Miguel e Maria, e aos meus tios Ana Amélia e Jardelino pela ajuda
prestada com informações e conhecimento.
A Manuela Sales pelo companheirismo e ajuda durante a fase final deste trabalho.
Ao amigo Thiago Marinho pelas fotos e demais ajudas prestadas.

Aos meus amigos André Luis, Sérgio, Thiago Marinho, Murillo e Jefferson que
durante muitos anos me acompanham sempre me incentivando nas minhas escolhas e
atividades e contribuindo com inúmeros momentos de diversão e alegria.
Aos meus amigos herpetólogos Bruno Maggi e Lucio Rodrigues pelas discussões
científicas e momentos de alegria.
Finalizando, queria agradecer a todos que de qualquer maneira tenha me ajudado na
minha formação pessoal e profissional.
Muito Obrigado.

RESUMO
Tropidurus semitaeniatus e Tropidurus hispidus são espécies de lagartos que têm ampla
distribuição no Brasil, sendo que a primeira é reconhecida como endêmica das Caatingas,
onde vive em sintopia com a segunda, que se distribui em outros ecossistemas e biomas.
Neste trabalho, avaliou-se a dieta — incluindo variações sazonais — e o comportamento de
forrageio, para averiguar a importância de tais fatores na estrutura dessas populações e
coexistência dessas espécies em sintopia. Não se constatou diferença significativa na
composição da dieta das duas espécies, embora T. hispidus se alimente de presas maiores do
que T. semitaeniatus; este fato, aliado à diferença no comportamento de forrageio na estação
chuvosa, são fatores ecológicos que contribuem para a coexistência sintópica dessas espécies
na área de Caatinga estudada.
PALAVRAS CHAVE: Lagartos, Dieta, Comportamento de forrageio, Tropidurus, Caatinga

ABSTRACT
Tropidurus semitaeniatus and Tropidurus hispidus are lizard species that are widely
distributed in Brazil. The former is endemic to the Caatingas, where they live in syntopy with
the latter, which is distributed in other ecosystems and biomes. In this work, diet was
evaluated – including seasonal variations – as well as foraging behavior, to determine the
importance of these factors in the structure of these populations and the coexistence of these
species in syntopy. No significant difference was found in diet composition between the two
species, although T. hispidus feed on larger preys than do T. semitaeniatus; this fact, along
with the different foraging behavior in the rainy season, are ecological factors that contribute
to the syntopic coexistence of these species in the Caatinga area studied.
KEYWORDS: Lizards, Diet, Foraging behavior, Tropidurus, Caatinga

LISTA DE FIGURAS
Figura 1 - Tropidurus hispidus, sobre afloramento rochoso na Estação Ecológica do Seridó –
ESEC Seridó. maio 2007. (Foto: Thiago Marinho)....................................................................5
Figura 2 – Tropidurus semitaeniatus, adulto, sobre afloramento rochoso na Estação Ecológica
do Seridó – ESEC Seridó. maio 2007. (Foto: Thiago Marinho)................................................5
Figura 3 – Localização da ESEC-Seridó no estado do Rio Grande do Norte, Brasil. Fonte:
Instituto Brasileiro do Meio Ambiente dos Recursos naturais Renováveis – IBAMA.
www.ibama.gov.br acessado em 05/06/2008.............................................................................7
Figura 4 – Correlação entre o número (NI), a freqüência de ocorrência dos itens (F) na dieta
de Tropidurus semitaeniatus e o IDRA na estação seca (A); a mesma correlação na estação
chuvosa (B); Correlação entre o número (NI), a freqüência de ocorrência dos itens (F) na dieta
de Tropidurus hispidus e o IDRA na estação seca (C); A mesma correlação na estação
chuvosa(D). ESEC Seridó, outubro de 2006 a setembro de 2007 ...........................................20
LISTA DE TABELAS
Tabela I – Freqüência de ocorrência (F), número (#), volume de itens (mm³) e índice de
importância relativa (Ix) por Categorias de itens alimentares na dieta de Tropidurus
semitaeniatus (N=50) e Tropidurus hispidus (N=27) na ESEC Seridó, durante a estação seca,
de outubro de 2006 a fevereiro de 2007 e de junho a setembro de 2007.............................16
Tabela II – Freqüência de ocorrência (F), número (#), volume de itens (mm³) e índice de
importância relativa (Ix) por Categorias de itens alimentares na dieta de Tropidurus
semitaeniatus (N=33) e Tropidurus hispidus (N=14) na ESEC Seridó, durante a estação
chuvosa, de março a maio de 2007 ..........................................................................................18
Tabela III – Correlação entre o comprimento dos cinco maiores itens encontrados nos
estômagos de T. semitaeniatus e as medidas morfométricas. ESEC Seridó, outubro de 2006 a
setembro de 2007......................................................................................................................21
Tabela IV – Correlação entre o comprimento dos cinco maiores itens encontrados nos
estômagos de T. hispidus e as medidas morfométricas. ESEC Seridó, outubro de 2006 a
setembro de 2007......................................................................................................................21

SUMÁRIO
Página
Lista de Figuras ........................................................................................................ vii
Lista de Tabelas......................................................................................................... vii
RESUMO ................................................................................................................. viii
ABSTRACT ............................................................................................................. viii
INTRODUÇÃO......................................................................................................... 1
MATERIAL E MÉTODOS....................................................................................... 6
Área de Estudo.......................................................................................................... 6
Trabalho em Campo.................................................................................................. 7
Análise da dieta ........................................................................................................ 9
Métodos de observação e análise dos comportamentos de forrageio ................. 11
RESULTADOS ........................................................................................................ 14
Dieta .......................................................................................................................... 14
Comportamento de forrageio .................................................................................... 23
DISCUSSÃO ............................................................................................................ 25
Dieta........................................................................................................................... 25
Comportamento de forrageio .................................................................................... 28
CONCLUSÕES......................................................................................................... 30
REFERÊNCIAS BIBLIOGRÁFICAS...................................................................... 31

INTRODUÇÃO
Os lagartos constituem o grupo mais diversificado dos répteis, com cerca de 5.000
espécies (Uetz et al. 2008), possuem distribuição geral ampla, ocupando habitats que variam
de desertos, acima de faixa de florestas, em montanhas e em áreas tropicais e subtropicais
(Pough et al. 2003) O Brasil possui alta diversidade de espécies de lagartos, com 234 espécies
(Uetz et al. 2008) das quais a Caatinga abriga 45 espécies; mas, se incluídas as que ocorrem
nos enclaves de floresta úmida nas Caatingas, este número sobe para 73, comparável ao do
Cerrado e da Mata Atlântica (Rodrigues 2005).
A fauna de lagartos das Caatingas foi considerada até recentemente bem conhecida,
principalmente com base no trabalho clássico de Vanzolini et al. (1980), no qual foram
efetuadas descrições e comentários ecológicos sobre as espécies habitantes desse bioma.
Durante algum tempo, também prevaleceu a idéia de que o bioma Caatinga não possuía fauna
própria (Rodrigues 2003); acreditava-se que as comunidades ali encontradas consistiam de
espécies pertencentes às comunidades do Cerrado e da Mata Atlântica, com as quais as
Caatingas fazem fronteiras (Vanzolini 1974, 1976; Vitt e Vangilder 1983). Segundo
Rodrigues (2003), essas afirmativas resultam de visão precipitada, baseada em coleções
pouco representativas, mas que acabaram contribuindo para que as Caatingas fossem descritas
por muito tempo na literatura como pobre e de pouca importância ecológica. Dois exemplos
apóiam essas observações de Rodrigues (2003): este autor, trabalhando a partir de 1980 nas
Caatingas, descreveu duas espécies novas de lagartos para a região do Cariri paraibano
(Rodrigues 1986, 2000) e 14 novas e endêmicas das dunas paleoquaternárias distribuídas ao
longo do Rio São Francisco (Rodrigues 1991a, b, c, 1996, 2000).
Embora atualmente sejam reconhecidas para a região com feições características das
Caatingas semi-áridas, entre 45 e 47 espécies de lagartos (Rodrigues 2003, 2005), Rodrigues
(2003) considera o nível de conhecimento sobre a herpetofauna das Caatingas muito

insatisfatório e fragmentado, uma vez que poucas localidades têm sido amostradas. Além
disso, quanto a estudos ecológicos sobre estrutura de comunidades de lagartos das Caatingas,
único trabalho publicado (Vitt 1995), resultou de estudo na região de Exú – PE.
A fauna de répteis da região que compreende o Seridó Norte-riograndense e o Cariri
paraibano foi reconhecida como de provável importância, mas insuficientemente conhecida
(Rodrigues et al. 2002, 2003). Por isso, desde 2002, sob fomento do CNPq através do
Programa Ecológico de Longa Duração (PELD/Caatingas), vários estudos ecológicos sobre
herpetofauna das Caatingas dessas duas regiões vêm sendo efetuados, inclusive este. Alguns
outros resultados sobre ecologia de lagartos e serpentes estão sendo submetidos à publicação
(Maggi e Freire, in press; Costa e Freire, in press).
Os lagartos constituem um grupo animal muito adequado ao estudo dos diversos
modelos em Ecologia por serem, em sua maioria, diurnos, abundantes, de fácil observação,
manuseio e captura, além de sua taxonomia ser relativamente bem conhecida (Huey et al.
1983, Rocha 1994).
A maioria dos padrões ecológicos de populações e/ou comunidades de lagartos foram
identificados e estabelecidos durante estudos efetuados a partir da década de 60, com espécies
de lagartos da América do Norte, Austrália e África (Pianka 1966a, b, 1969, 1971, 1973).
Esses estudos clássicos investigaram a estrutura das comunidades, considerando a competição
entre espécies como um fator relevante (Pianka 1973). No entanto, além da competição,
outros fatores devem ser considerados, uma vez que as espécies de lagartos em simpatria
podem diferir em um ou mais aspectos do seu nicho ecológico e, desta forma, partilhar
recursos de maneira a possibilitar a coexistência (Schoener 1977, Dunham 1983). No Brasil,
os estudos sobre a ecologia e história natural de populações de lagartos têm enfocado diversos
aspectos, com destaque para dieta e modos de forrageio (Magnusson et al. 1985, Vitt 1990;
Rocha, 1989, 1992, 1996, 1998, Van Sluys 1993, Bergallo & Rocha 1994, Vrcibradic &

Rocha 1996; Vitt et al. 1997, Vrcibradic & Rocha 1998, Teixeira & Giovanelli 1999, Van
Sluys et al. 2004).
Os estudos sobre dieta de lagartos têm contribuído para o melhor entendimento sobre a
ecologia das espécies, bem como acerca das estratégias de forrageio por elas desenvolvidas. O
modo de forrageio pode ser determinado com base na distância percorrida pelo animal no
ambiente (Pianka 1986, Rocha 1996, 1998) e/ou inferido pelo tipo de presa ingerida pelos
lagartos (Huey & Pianka 1981). Embora exista forte influência filogenética na determinação
do modo de forrageio dos lagartos (Cooper 1995), uma espécie pode modular sua estratégia
em função de alterações na disponibilidade de alimentos (Huey & Pianka 1981).
Huey & Pianka (1981) estabeleceram que lagartos, como muitos outros predadores,
basicamente forrageiam de dois modos: ou são ativos (wild foraging) ou caçam de tocaia
como forrageadores sedentários (sit-and-wait). No entanto, esses dois modos devem ser
considerados como pontos extremos de um gradiente de variações das táticas de forrageio,
podendo existir tipos intermediários na natureza, como os forrageadores errantes (Rocha
1994), ou seja, aqueles que podem modular suas estratégias de caça de acordo com as
oportunidades e disponibilidade de presas, além das pressões impostas pelo ambiente, como
por exemplo, a sazonalidade. Os modos de forrageio podem influenciar quase todos os
aspectos da biologia dos lagartos (Huey et al. 1983), contribuindo para a evolução da forma
do corpo, fisiologia, reprodução e comportamento do animal (Huey & Pianka 1981).
Tropidurus hispidus (Spix 1825) e T. semitaeniatus (Spix 1825) são duas espécies de
lagartos heliófilos que têm ampla distribuição geográfica e ocorrem em sintopia nas Caatingas
do nordeste brasileiro (Vanzolini et al. 1980; Rodrigues 1987, Vitt 1995). No entanto,
Tropidurus hispidus (Figura 1), que é a maior espécie do gênero (maiores espécimes podem
alcançar 35 cm de comprimento total), se distribui no nordeste da América do Sul,
predominantemente nas Caatingas, Serra do Espinhaço e adjacências, com limite sul em Brás

Fortes – MG (Rodrigues 1987). Já T. semitaeniatus (Figura 2) é uma espécie menor (maiores
espécimes podem alcançar 20 cm de comprimento total), e reconhecido originalmente como
habitante de toda a Caatinga nordestina, de onde é considerado endêmico (Vanzolini et al.
1980, Rodrigues 1987) e com distribuição relictual (Rodrigues 2003). Recentemente tem sido
encontrado em afloramentos rochosos em clareira e/ou entorno de remanescentes da Mata
Atlântica nordestina (Freire 2001), e no litoral em regiões próximas da praia ao norte de
Salvador, também em afloramentos rochosos (Freitas & Pavie 2002).
Na Estação Ecológica do Seridó (ESEC Seridó), Estado do Rio Grande do Norte, T.
semitaeniatus e T. hispidus são sintópicas, especialmente nos afloramentos rochosos. Este
estudo, portanto, teve como objetivos identificar a dieta e o comportamento de forrageio
dessas duas espécies, e analisar como esses fatores contribuem para a estrutura dessas
populações e possibilitam sua coexistência em sintopia na área de Caatinga estudada. Foram
analisadas: (i) a composição e as variações sazonais, sexuais e diferenças ontogenéticas das
dietas de ambas as espécies; (ii) a existência de correlação entre a composição das dietas e a
disponibilidade relativa de presas no ambiente; (iii) a diversidade de presas e o grau de
sobreposição nas dietas; e (iv) as possíveis diferenças no comportamento e intensidade de
forrageio entre T. hispidus e T. semitaeniatus.

Figura 1 – Tropidurus hispidus, sobre afloramento rochoso na Estação Ecológica do Seridó – ESEC Seridó. Maio 2007 (Foto: Thiago Marinho).
Figura 2 – Tropidurus semitaeniatus, sobre afloramento rochoso na Estação Ecológica do Seridó – ESEC Seridó. Maio 2007 (Foto: Thiago Marinho).

MATERIAL E MÉTODOS
Área de Estudo
A Estação Ecológica do Seridó (ESEC Seridó, Figura 3), está inserida no Domínio
Morfoclimático das Caatingas (sensu Ab’Sáber 1974), mais precisamente na ecorregião
denominada Depressão Sertaneja Setentrional que divide com a Depressão Sertaneja
Meridional a paisagem mais típica do semi-árido nordestino: uma extensa planície baixa, de
relevo predominantemente suave-ondulado com elevações residuais disseminadas na
paisagem. Os solos são rasos, pedregosos, de origem cristalina e fertilidade média a alta, mas
muito suscetíveis à erosão. A depressão Sertaneja Setentrional contém duas áreas
diferenciadas com extremos climáticos que condicionam uma vegetação de porte mais baixo
que caracteriza o Seridó e o Cariri (Velloso et al. 2002).
O Seridó é bastante semelhante ao resto da Depressão Setentrional, de relevo
suavemente ondulado com elevações residuais, porém os solos são pedregosos e tendem a ser
mais rasos que o resto da ecorregião, com alta fertilidade natural e grande potencial de
minério. Predominam os solos brunos não cálcicos, mas também ocorrem planosolos e solos
podzólicos, além dos solos litólicos das elevações residuais, mas também são comuns
afloramentos de rochas, na forma de grandes lajedos ou blocos desagregados, que formam a
paisagem típica da região (Velloso et al. 2002).
A ESEC Seridó (06º 34’ S, 37º 15’ W), compreende uma área de 1.166,38 hectares e
está situada no município de Serra Negra do Norte (Varela-Freire 2002). O clima, como o de
outras regiões das Caatingas, é do tipo semi-árido quente e seco (Ab’Sáber 1974), com curta
estação chuvosa que predomina nos meses de março a maio, com índices de precipitações
variando entre 500 e 700 mm/ano. As temperaturas médias anuais variam de 28ºC a 30ºC, no
decorrer do ano chegando a mais de 40ºC; as mínimas variam de 17ºC a 20ºC.

A vegetação é do tipo hiperxerófila arbóreo-arbustiva, com estrato herbáceo apenas na
estação chuvosa (Varela-Freire 2002). Em meio a essa vegetação arbóreo-arbustiva são
encontradas várias extrusões rochosas que, durante a estação chuvosa, ficam recobertas pela
vegetação, proporcionando ambiente parcialmente sombreado; durante a estação seca, com o
fenômeno da caducifolia, ficam expostas em forma de grandes afloramentos rochosos.
Figura 3 – Localização da ESEC-Seridó no Estado do Rio Grande do Norte, Brasil. Fonte: Instituto Brasileiro do Meio Ambiente dos Recursos naturais Renováveis – IBAMA. www.ibama.gov.br acessado em 05/06/2008.
Trabalho em Campo
Foram efetuadas excursões mensais de outubro de 2006 a setembro de 2007, sendo
que as observações comportamentais foram efetuadas apenas nas estações seca (outubro a
dezembro/2006) e chuvosa (abril a junho/2007). Em todo o período, foram realizadas coletas
de lagartos para análise da dieta e coleta de artrópodes no ambiente, totalizando um ano de

trabalho em campo, possibilitando a abordagem comparativa sazonal. Para este estudo, a
estação chuvosa correspondeu aos meses de março a junho de 2007, período em que
ocorreram as primeiras chuvas e que se caracterizou pela fisionomia exuberante da vegetação.
Para a estação seca foram considerados os meses de outubro de 2006 a fevereiro de 2007 e de
julho a setembro de 2007, ambos períodos de estiagem, caracterizado pela fisionomia
esbranquiçada da vegetação, acentuando o processo de caducifolia e conseqüentemente,
aumento da insolação.
A coleta dos lagartos para a análise da dieta foi efetuada em área com afloramentos
rochosos, escolhida aleatoriamente, específica para esta finalidade e situada a cerca de 500 m
da área de observação comportamental para que não haja interferência nos lagartos
observados. Os lagartos foram capturados através do uso de carabina de pressão 4,5 mm e de
armadilhas de cola (Glue Traps), sendo imediatamente mortos com éter. Ao final de cada dia
de coleta, os lagartos eram dissecados para remoção dos estômagos, que foram mantidos em
álcool a 70% para análise posterior. Em todos os exemplares coletados para análise da dieta
foram tomadas as seguintes medidas, utilizando-se paquímetro com precisão de 0,1 mm:
comprimento rostro-cloacal (CRC) - distância do focinho à cloaca; rostro-canto do tímpano
(RCT) - distância da ponta do focinho à margem posterior do tímpano (comprimento da
cabeça); rostro-comissura labial (RCL) - distância da ponta do focinho à comissura labial
(comprimento da boca); largura da cabeça (LC) - a maior largura, medida à altura da abertura
auditiva.
Os lagartos coletados foram identificados com um número de campo, fixados em
formol a 10%, preservados em álcool a 70% e tombados na Coleção Herpetológica do
Departamento de Botânica, Ecologia e Zoologia (CHBEZ) da Universidade Federal do Rio
Grande do Norte.

A classificação dos indivíduos coletados nas categorias “adultos” e “jovens” foi feita
conforme Vitt (1995).
A existência de dimorfismo sexual quanto ao tamanho do corpo (CRC) para as duas
espécies de lagartos foi avaliada com o Teste t de Student (Zar, 1999).
Análise da dieta.
Em laboratório, os estômagos mantidos em álcool a 70% foram abertos, o seu
conteúdo colocado em placas de Petri e identificado em nível de ordem. A exceção foi a
ordem Hymenoptera, para a qual foi identificada a família Formicidae, devido a sua
importância na dieta dessas espécies de lagartos. Para as ordens Coleoptera e Lepidoptera, o
estágio de desenvolvimento do indivíduo (larva ou adulto) também foi considerado devido ao
alto número de larvas encontradas. Os fragmentos que não puderam ser identificados foram
categorizados como tais. As partes vegetais encontradas nos estômagos foram identificadas
quanto às seguintes categorias: folhas, flores e sementes.
Para a análise da dieta, os itens foram contados e utilizados os métodos numéricos
(freqüência de ocorrência e freqüência numérica dos itens) e volumétricos (medida do maior
comprimento e largura, com o auxílio de papel milimetrado). O volume das categorias
alimentares foi estimado através da fórmula esferóide, conforme Faria e Araújo (2004):
Volume = ( .comprimento.largura2)/6.
O Índice de Importância Relativa (Ix) foi calculado para cada categoria de item
alimentar, somando-se as percentagens de freqüência de ocorrência, número e volume de
itens, e dividindo-se por três (Menezes et al. 2006, Rocha & Anjos 2007).
Para a coleta de artrópodes (possível oferta de alimento), foram distribuídas
aleatoriamente 30 armadilhas ao longo da área de coleta e mantidas por três dias consecutivos
em cada mês de coleta, durante um ano. Cada armadilha era composta por bandeja plástica de

50x25 cm, contendo água e sabão. Ficavam expostas das 7 às 17 h, quando os artrópodes que
caíam nas armadilhas eram coletados. Também foram utilizadas armadilhas contendo óleo de
sardinha em conserva. Para estimar a disponibilidade de artrópodes, no ambiente, foi utilizado
o Índice de Disponibilidade Relativa de Artrópodes (IDRA; Strüsmann et al. 1984, Van Sluys
1991, Vrcibradic & Rocha, 1995).
Todos os artrópodes coletados foram fixados em álcool a 70% para posterior
identificação.
As dietas das duas espécies de Tropidurus foram comparadas com o índice de
disponibilidade relativa de artrópodes no ambiente (IDRA), por número e freqüência de
presas, utilizando-se o teste de Correlação de Spearman (Zar 1999).
A ocorrência de diferenças sazonal, sexual e ontogenética nas dietas de T. hispidus e
T. semitaeniatus foi avaliada através do teste de Kolmogorov-Smirnov (Zar 1999), também
aplicado para constatar se as dietas dessas duas espécies são diferentes. Em todos os casos a
comparação das dietas foi feita através do volume e do número dos itens alimentares
encontrados nos estômagos.
A Análise de Regressão Simples (Zar 1999) foi utilizada para testar a relação entre as
medidas morfométricas dos lagartos (CRA, RCT, RCL, LC) e o comprimento médio dos itens
ingeridos. Para isso, foram utilizados os cinco maiores itens alimentares encontrados em cada
estômago. As regressões foram realizadas com os valores das medidas morfométricas e do
tamanho dos itens transformados em logaritmo na base 10.
O teste U de Mann-Whitney (Zar 1999) foi utilizado para constatar a existência de
diferenças sexuais e ontogenéticas no tamanho dos itens alimentares ingeridos pelos
indivíduos das duas espécies. O mesmo teste foi aplicado para avaliar se o tamanho dos itens
ingeridos por T. hispidus e T. semitaeniatus diferem em geral e entre as diferentes categorias
de lagartos (machos, fêmeas e jovens) de cada espécie.

Para todos os testes estatísticos e no cálculo das proporções dos itens não foram
consideradas as partes de artrópodes e as partes vegetais não identificadas. Em todos os testes
o nível de significância adotado foi de 0,05 (5%).
Para avaliar a diversidade das presas nas dietas nas diferentes estações foi utilizado o
Índice de Diversidade de Shanonn-Wiener, que se baseia na abundância relativa das espécies,
utilizando-se o programa Bio-Dap (Magurran 1988).
A sobreposição no nicho alimentar (em número e volume de itens) entre T. hispidus e
T. semitaeniatus foi averiguada utilizando-se a fórmula de sobreposição simétrica (Pianka
1973), utilizada também por Vitt (1995), Vitt et al. (2001), Faria & Araújo (2004) e Van
Sluys et al. (2004).
Onde: “pi” = proporção do recurso “i”; “j” e “k” representam as duas espécies de lagartos.
Os valores próximos a zero indicam não haver similaridade nas dietas; valores próximos a um
indicam sobreposição na dieta.
Métodos de observação e de análise dos comportamentos de forrageio
Para as observações comportamentais, foram estabelecidos cinco afloramentos
rochosos, circundados por vegetação e distantes cerca de 500 m da área de coleta. Em todos

estes foram utilizados os métodos de amostragem “Ad Libitum” e “Animal focal” (Altmann
1974, Martin & Bateson, 1973).
O método Ad Libitum foi empregado em estudo piloto para determinar as categorias
comportamentais que foram utilizadas nas observações posteriores, e o método “Animal
focal” foi utilizado para observar os indivíduos em atividade, dentre as categorias
estabelecidas previamente.
As categorias comportamentais, definidas para avaliar o comportamento e a
intensidade de forrageio pelos espécimes de T. hispidus e T. semitaeniatus, definidos a partir
do método Ad Libitum foram: (i) Número de deslocamentos; (ii) Tempo parado (%); (iii)
Distância (cm) percorrida; (iv) Tentativas de captura de presa.
O período de observação foi de 7:00 às 10:00 e de 14:00 às 17:00h, dividido em seis
intervalos de uma hora cada para obter uma amostragem homogênea. Nesses intervalos foram
realizadas sessões focais pelo método de Animal Focal (Altmann 1974, Martin & Bateson,
1993) com duração de 10 minutos, alternando as espécies observadas sempre que possível.
Todas as observações foram realizadas com auxílio de gravadores de voz e cronômetro
regressivo digital. Após a visualização do lagarto, a observação era iniciada depois de 5
minutos, para minimizar a interferência pela presença do observador. Quando ocorria
qualquer reação do lagarto, ou quando este desaparecia do foco, a observação era
desconsiderada, conforme método utilizado por Vitt et al. (1996).
O número de movimentos, a distância percorrida em cm e o número de tentativas de
captura de presa por 10 minutos, foram registrados e calculados para cada lagarto, utilizando-
se a média aritmética por estação do ano e por espécie. Esta metodologia foi adaptada da
descrita em Strüssman et al. (1984), Magnusson et al. (1985) e Gasnier et al. (1994).
O teste U de Mann-Whitney (Zar 1999) foi utilizado para avaliar a existência de
variação sazonal nos índices de forrageio (freqüência de movimentos, distância percorrida e

tentativa de captura de presas) de T. hispidus e T. semitaeniatus, bem como para verificar se
as duas espécies apresentavam diferença quanto ao modo de forrageio, de acordo com os
índices definidos.

RESULTADOS
Foram coletados 83 exemplares de Tropidurus semitaeniatus durante as estações seca
(50) e chuvosa (33), sendo 26 machos adultos, 36 fêmeas adultas e 21 jovens. Os machos têm
o comprimento rostro-cloacal médio (CRC = 76,6 ± 6,4 mm, n = 27) significativamente maior
que as fêmeas (CRC = 65,3 ± 3,0 mm, n = 36; t = 9,251 gl= 61, p < 0,001). Em relação a
Tropidurus hispidus, foram coletados 45 exemplares, durante as estações seca (27) e chuvosa
(18), sendo 13 machos adultos, 21 fêmeas adultas e 11 jovens. Os machos tiveram o CRC
médio (CRC = 102,9 ± 16,2 mm, n = 12) significativamente maior que as fêmeas (CRC =
84,5 ± 8,7 mm, n = 21; t = 4,257, gl = 31, p < 0,001).
Entre os adultos das duas espécies, Tropidurus hispidus teve o CRC médio (CRC =
91,1 ± 14,7 mm, n = 33) significativamente maior do que Tropidurus semitaeniatus (CRC =
70,1 ± 7,3 mm, n = 63; t = 9,309, gl = 94, p = 0,000).
Dieta
Composição, diferenças sazonais, sexuais e ontogenéticas
A composição da dieta, resultante da análise dos estômagos dos lagartos coletados,
está listada nas tabelas I e II.
Na estação seca (Tabela I), para T. semitaeniatus, folhas foi o item alimentar mais
importante (Ix = 43,2). Hymenoptera/Formicidae foi o item com o segundo maior índice de
importância relativa (Ix = 29,0), seguido por Isoptera (Ix = 17, 7). Em relação à freqüência de
ocorrência dos itens, o mais freqüente foi também Hymenoptera/Formicidae (50%), seguido
por folhas (30%) e Coleoptera (24%). Em relação ao número de itens, a maior quantidade
também foi de Hymenoptera/Formicidae (35,5%), seguido por Isoptera (29,3%) e folhas

(22,3%). O item mais importante em termos de volume foi folhas (77,3%), seguido por
Isoptera (5,9%) e Larvas de Lepidoptera (4,1%).
Tropidurus hispidus, teve folhas como o item com o maior índice de importância
relativa (Ix = 47,5), seguido por Hymenoptera/Formicidae (Ix = 42,3) e Isoptera (Ix = 11,1).
Em relação aos itens mais freqüentes, Hymenoptera/Formicidae esteve em 59,3% dos
estômagos, seguido por folhas (48,1%) e larvas de Lepidoptera (18,5%). Em relação ao
número de itens, o que apareceu em maior quantidade foi Hymenoptera/Formicidae (65,5%),
seguido por Isoptera com (20,6%). O item mais importante em termos de volume foi folhas
(84,4%), seguido por Orthoptera (6,5%).
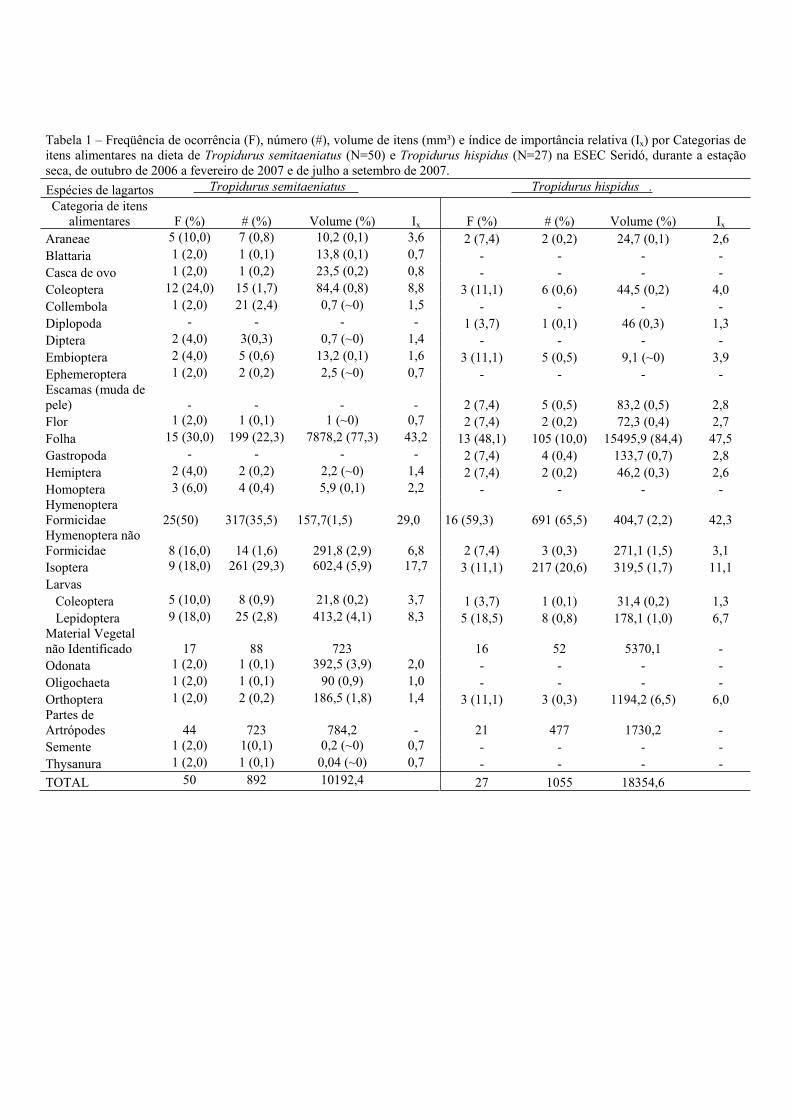
Tabela 1 – Freqüência de ocorrência (F), número (#), volume de itens (mm³) e índice de importância relativa (Ix) por Categorias de itens alimentares na dieta de Tropidurus semitaeniatus (N=50) e Tropidurus hispidus (N=27) na ESEC Seridó, durante a estação seca, de outubro de 2006 a fevereiro de 2007 e de julho a setembro de 2007. Espécies de lagartos Tropidurus semitaeniatus Tropidurus hispidus .Categoria de itens
alimentares F (%) # (%) Volume (%) Ix F (%) # (%) Volume (%) Ix
Araneae 5 (10,0) 7 (0,8) 10,2 (0,1) 3,6 2 (7,4) 2 (0,2) 24,7 (0,1) 2,6 Blattaria 1 (2,0) 1 (0,1) 13,8 (0,1) 0,7 - - - - Casca de ovo 1 (2,0) 1 (0,2) 23,5 (0,2) 0,8 - - - - Coleoptera 12 (24,0) 15 (1,7) 84,4 (0,8) 8,8 3 (11,1) 6 (0,6) 44,5 (0,2) 4,0 Collembola 1 (2,0) 21 (2,4) 0,7 (~0) 1,5 - - - - Diplopoda - - - - 1 (3,7) 1 (0,1) 46 (0,3) 1,3 Diptera 2 (4,0) 3(0,3) 0,7 (~0) 1,4 - - - - Embioptera 2 (4,0) 5 (0,6) 13,2 (0,1) 1,6 3 (11,1) 5 (0,5) 9,1 (~0) 3,9 Ephemeroptera 1 (2,0) 2 (0,2) 2,5 (~0) 0,7 - - - - Escamas (muda de pele) - - - - 2 (7,4) 5 (0,5) 83,2 (0,5) 2,8 Flor 1 (2,0) 1 (0,1) 1 (~0) 0,7 2 (7,4) 2 (0,2) 72,3 (0,4) 2,7 Folha 15 (30,0) 199 (22,3) 7878,2 (77,3) 43,2 13 (48,1) 105 (10,0) 15495,9 (84,4) 47,5 Gastropoda - - - - 2 (7,4) 4 (0,4) 133,7 (0,7) 2,8 Hemiptera 2 (4,0) 2 (0,2) 2,2 (~0) 1,4 2 (7,4) 2 (0,2) 46,2 (0,3) 2,6 Homoptera 3 (6,0) 4 (0,4) 5,9 (0,1) 2,2 - - - - Hymenoptera Formicidae 25(50) 317(35,5) 157,7(1,5) 29,0 16 (59,3) 691 (65,5) 404,7 (2,2) 42,3 Hymenoptera não Formicidae 8 (16,0) 14 (1,6) 291,8 (2,9) 6,8 2 (7,4) 3 (0,3) 271,1 (1,5) 3,1 Isoptera 9 (18,0) 261 (29,3) 602,4 (5,9) 17,7 3 (11,1) 217 (20,6) 319,5 (1,7) 11,1 Larvas Coleoptera 5 (10,0) 8 (0,9) 21,8 (0,2) 3,7 1 (3,7) 1 (0,1) 31,4 (0,2) 1,3 Lepidoptera 9 (18,0) 25 (2,8) 413,2 (4,1) 8,3 5 (18,5) 8 (0,8) 178,1 (1,0) 6,7 Material Vegetal não Identificado 17 88 723 16 52 5370,1 - Odonata 1 (2,0) 1 (0,1) 392,5 (3,9) 2,0 - - - - Oligochaeta 1 (2,0) 1 (0,1) 90 (0,9) 1,0 - - - - Orthoptera 1 (2,0) 2 (0,2) 186,5 (1,8) 1,4 3 (11,1) 3 (0,3) 1194,2 (6,5) 6,0 Partes de Artrópodes 44 723 784,2 - 21 477 1730,2 - Semente 1 (2,0) 1(0,1) 0,2 (~0) 0,7 - - - - Thysanura 1 (2,0) 1 (0,1) 0,04 (~0) 0,7 - - - - TOTAL 50 892 10192,4 27 1055 18354,6

Na estação chuvosa (Tabela II), T. semitaeniatus teve Hymenoptera/Formicidae como
o item com o maior índice de importância relativa (Ix = 42,0), seguido por larvas de
Lepidoptera (Ix = 34,8). Em relação aos itens mais freqüentes, Hymenoptera/Formicidae
esteve em 60,2% dos estômagos, seguido por larvas de Lepidoptera (48,5%). Em relação ao
número de itens, o mais numeroso foi Hymenoptera/Formicidae (62,3%), seguido por larva de
Lepidoptera (16,5%). O item mais importante em termos de volume foi larva de Lepidoptera
(39,4%), seguido por flor (19,5%).
Tropidurus hispidus teve Orthoptera com o maior índice de importância relativa (Ix =
29,2), seguido por Hymenoptera/Formicidae (Ix = 27,3) e larvas de Lepidoptera (Ix = 19,6).
Em relação aos itens mais freqüentes, Hymenoptera/Formicidae estava presente em 50,0%
dos estômagos, seguidos por Orthoptera (44,4%) e Coleoptera (38,9%). O item mais
importante em termos de volume foi Orthoptera (40,4%), seguido por Lepidoptera (16,6%) e
larvas de Lepidoptera (16,5%).

Tabela 2 – Freqüência de ocorrência (F), número (#), volume de itens (mm³) e índice de importância relativa (Ix) por Categorias de itens alimentares na dieta de Tropidurus semitaeniatus (N=33) e Tropidurus hispidus (N=18) na ESEC Seridó, durante a estação chuvosa, de março a maio de 2007. Espécies de lagartos Tropidurus semitaeniatus Tropidurus hispidus . Categorias de itens
alimentares F (%) # (%) Volume (%) Ix F (%) # (%) Volume (%) Ix
Araneae 6 (18,2) 13 (2,2) 28,1 (0,7) 7,0 1 (5,6) 1 (0,3) 461,6 (2,8) 2,9 Casca de ovo 1 (3,0) 1 (0,2) 10,4 (0,2) 1,1 1 (5,6) 1 (0,3) 65,4 (0,4) 2,1 Coleoptera 3 (9,1) 5 (0,9) 128,3 (3,0) 4,3 7 (38,9) 17 (4,5) 784,6 (4,8) 16,1 Diplopoda 4 (12,1) 4 (0,7) 72,6 (1,7) 4,8 5 (27,8) 17 (4,5) 784,6 (4,8) 12,4 Diptera 5 (15,2) 9 (1,5) 424,1 (10,0) 8,9 - - - - Embioptera 2 (6,1) 2 (0,3) 5,4 (0,1) 2,2 - - - - Ephemeroptera 2 (6,1) 3 (0,5) 3,2 (0,1) 2,2 - - - - Escamas (muda de pele) 3 (9,1) 4 (0,7) 119,3 (2,8) 4,2 - - - - Flor 6 (18,2) 34 (5,9) 827,1 (19,5) 14,5 - - - - Folha 5 (15,2) 5 (0,9) 20,5 (0,5) 5,5 4 (22,2) 11 (2,9) 57,5 (0,4) 8,5 Hemiptera 2 (6,1) 9 (1,5) 127,3 (1,8) 3,5 1 (5,6) 1 (0,3) 669,8 (4,1) 3,3 Homoptera 1 (3,0) 1 (0,2) 4,7 (0,1) 1,1 - - - - Hymenoptera Formicidae 20 (60,2) 362 (62,3) 125,8 (3,0) 42,0 9 (50) 116 (31,0) 130 (0,8) 27,3 Hymenoptera não Formicidae 7 (21,2) 12 (2,1) 77,1 (1,8) 8,4 2 (11,1) 4 (1,1) 524 (3,2) 5,1 Isoptera 2 (6,1) 3 (0,5) 3,9 (0,1) 2,2 2 (11,1) 157 (42,0) 245,4 (1,5) 18,2 Larvas Coleoptera 9 (27,3) 14 (2,4) 251,9 (5,9) 11,9 1 (5,6) 1 (0,3) 8,2 (0,1) 2,0 Lepidoptera 16 (48,5) 96 (16,5) 1670 (39,4) 34,8 6 (33,3) 33 (8,8) 2691,9 (16,5) 19,6 Lepidoptera 2 (6,1) 2 (0,3) 321,8 (7,6) 4,7 2 (11,1) 3 (0,8) 2708,2 (16,6) 9,5 Material Vegetal não Identificado 4 8 200,5 - 3 5 1475,8 - Odonata - - - - 2 (11,1) 2 (0,5) 580,2 (3,6) 5,1 Orthoptera - - - - 8 (44,4) 10 (2,7) 6589,6 (40,4) 29,2 Partes de Artrópodes 28 310 1675,8 - 15 303 3012,1 - Phasmida 1 (3,0) 1 (0,2) 13 (0,3) 1,2 - - - - Semente 1 (3,0) 1 (0,2) 1,1 (~0) 1,1 - - - - TOTAL 33 581 4235,6 18 374 16301

Na estação seca, as dietas de T. semitaeniatus e de T. hispidus não diferiram
significativamente em número (Kolmogorov-Sminorv, Dmax = 0,259, p > 0,10), nem em
volume dos itens (Kolmogorov-Smirnov, Dmax = 0,259, p > 0,10). Estes resultados também
não foram significativos na estação chuvosa em número (Kolmogorov-Smirnov, Dmax =
0,222, p > 0,10), e em volume dos itens (Kolmogorov-Smirnov, Dmax = 0,222, p > 0,10).
A dieta de T. semitaeniatus não diferiu significativamente em número de itens entre as
estações seca e chuvosa (Kolmogorov-Smirnov, Dmax = 0,111, p > 0,10), nem em volume
dos itens (Kolmogorov-Smirnov, Dmax = 0,074, p > 0,10). O mesmo ocorreu para T. hispidus
em número de itens entre as estações seca e chuvosa (Kolmogorov-Smirnov, Dmax = 0,111, p
> 0,10), e em volume dos itens (Kolmogorov-Smirnov, Dmax = 0,074, p > 0,10).
Machos e fêmeas de T. semitaeniatus apresentaram dietas semelhantes quanto ao
número e volume dos itens (Kolmogorov-Smirnov, Dmax = 0,076, p > 0,10; Dmax = 0,115, p
> 0,10). Para T. hispidus também não ocorreu diferença significativa em número de itens
entre machos e fêmeas (Kolmogorov-Smirnov, Dmax = 0,055, p > 0,10), nem em volume
(Kolmogorov-Smirnov, Dmax = 0,166, p > 0,10).
Quando a comparação se deu entre a dieta de jovens e adultos de T. semitaeniatus,
constatou-se diferença significativa em número (Kolmogorov-Smirnov, Dmax = 0,500, p <
0,005) e em volume dos itens (Kolmogorov-Smirnov, Dmax = 0,615, p < 0,001). Já a dieta de
T. hispidus não diferiu em número de itens entre adultos e jovens (Kolmogorov-Smirnov,
Dmax = 0,411, p > 0,10), mas diferiu significativamente em relação ao volume dos itens
(Kolmogorov-Smirnov, Dmax = 0,529, p < 0,25).
Relação entre a dieta das espécies e o Índice de Disponibilidade Relativa de Artrópodes no
Ambiente (IDRA)
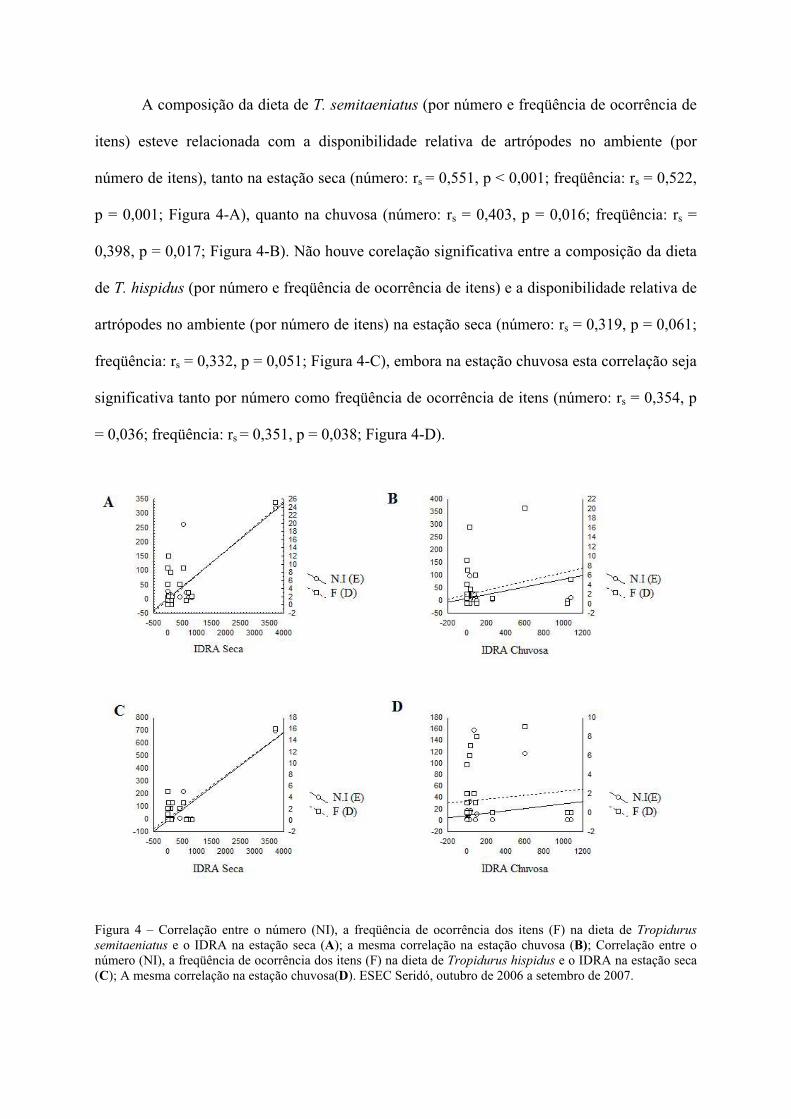
A composição da dieta de T. semitaeniatus (por número e freqüência de ocorrência de
itens) esteve relacionada com a disponibilidade relativa de artrópodes no ambiente (por
número de itens), tanto na estação seca (número: rs = 0,551, p < 0,001; freqüência: rs = 0,522,
p = 0,001; Figura 4-A), quanto na chuvosa (número: rs = 0,403, p = 0,016; freqüência: rs =
0,398, p = 0,017; Figura 4-B). Não houve corelação significativa entre a composição da dieta
de T. hispidus (por número e freqüência de ocorrência de itens) e a disponibilidade relativa de
artrópodes no ambiente (por número de itens) na estação seca (número: rs = 0,319, p = 0,061;
freqüência: rs = 0,332, p = 0,051; Figura 4-C), embora na estação chuvosa esta correlação seja
significativa tanto por número como freqüência de ocorrência de itens (número: rs = 0,354, p
= 0,036; freqüência: rs = 0,351, p = 0,038; Figura 4-D).
Figura 4 – Correlação entre o número (NI), a freqüência de ocorrência dos itens (F) na dieta de Tropidurus semitaeniatus e o IDRA na estação seca (A); a mesma correlação na estação chuvosa (B); Correlação entre o número (NI), a freqüência de ocorrência dos itens (F) na dieta de Tropidurus hispidus e o IDRA na estação seca (C); A mesma correlação na estação chuvosa(D). ESEC Seridó, outubro de 2006 a setembro de 2007.

Tamanho dos itens alimentares: relação com as medidas morfométricas e com variações inter
e intraespecíficas
A regressão entre o comprimento médio dos cinco maiores itens encontrado nos
estômagos e a medida do rosto-comissura labial (RCL) para T. semitaeniatus adultos foi
significativa (Tabela III), enquanto nos T. semitaeniatus jovens, todas as medidas
apresentaram regressão significativas com o comprimento médio dos cinco maiores itens
encontrados nos estômagos (Tabela III). Para Tropidurus hispidus não houve nenhuma
correlação significativa entre o comprimento médio dos cinco maiores itens encontrados nos
estômagos e suas medidas (Tabela IV).
Tabela III – Correlação entre o comprimento dos cinco maiores itens encontrados nos estômagos de T.semitaeniatus e as medidas morfométricas. ESEC Seridó, outubro de 2006 a setembro de 2007. Adultos Jovens
r2 P r2 P CRC 0,036 0,137 0,386 0,002* RCT 0,027 0,199 0,326 0,006* RCL 0,078 0,027* 0,440 0,001* LC 0,019 0,279 0,424 0,001* *indica que a regressão foi significativa, p < 0,05.
Tabela IV – Correlação entre o comprimento dos cinco maiores itens encontrados nos estômagos de T. hispidus e as medidas morfométricas. ESEC Seridó, outubro de 2006 a setembro de 2007. Adultos Jovens
r2 P r2 P CRC 0,046 0,226 0,003 0,860 RCT 0,057 0,177 0,003 0,860 RCL 0,081 0,107 0,062 0,456 LC 0,044 0,237 0,012 0,741 *indica que a regressão foi significativa, p < 0,05.
Em relação ao tamanho médio dos itens da dieta, T. semitaeniatus não apresentou
diferença significativa entre machos (6,6 ± 6,5 mm, n = 40) e fêmeas (6,1 ± 5,0 mm, n = 44;
Teste U de Mann-Whitney, z = -0,367, p = 0,713); no entanto, a diferença foi significativa
entre adultos (7,5 ± 6,2 mm, n = 63) e jovens (2,9 ± 1,6 mm, n = 21; Teste U de Mann-

Whitney, z = 4,008, p < 0,001). Também houve diferença significativa entre as estações seca
(5,3 ± 6,4 mm, n = 50) e chuvosa (7,8 ± 4,3 mm, n = 34; Teste U de Mann-Whitney, z = -
3,822, p < 0,001).
Para T. hispidus, em relação ao tamanho médio dos itens da dieta, o resultado foi
semelhante ao de T. semitaeniatus, pois também não houve diferença significativa entre
machos (14,9 ± 13,1 mm, n = 19) e fêmeas (9,7 ± 5,2 mm, n = 25; Teste U de Mann-Whitney,
z = 0,781, p = 0,434), mas houve entre adultos (14,0 ± 10,3 mm, n = 33) e jovens (6,0 ± 3,9
mm, n = 11; Teste U de Mann-Whitney, z = -3,035, p = 0,002), e entre as estações seca (8,2 ±
4,7 mm, n = 27) e chuvosa (18,0 ± 12,4 mm, n = 17; Teste U de Mann-Whitney, z = -3,278, p
= 0,001).
Comparando-se a dieta geral das duas espécies, ocorreu diferença no tamanho dos
itens da dieta entre T. semitaeniatus (6,3 ± 5,8 mm, n = 84) e T. hispidus (12,0 ± 9,7 mm, n =
44; Teste U de Mann-Whitney, z = -4,301, p < 0,001). Também houve diferença significativa
entre o tamanho dos itens consumidos pelos machos adultos de T. semitaeniatus (8,5 ± 7,2
mm, n = 27) e machos adultos de T. hispidus (19,9 ± 14,1 mm, n = 12; Teste U de Mann-
Whitney, z = 2,936, p = 0,003). O mesmo ocorreu entre as fêmeas adultas de T. semitaeniatus
(6,7 ± 5,3 mm, n = 36) e fêmeas adultas de T. hispidus (10,6 ± 5,1 mm, n = 21; Teste U de
Mann-Whitney, z = -3,068, p = 0,002), e entre os jovens de T. semitaeniatus (2,9 ± 1,6 mm, n
= 21) e jovens de T. hispidus (6,0 ± 3,9 mm, n = 11; Teste U de Mann-Whitney, z = -2,779, p
= 0,004).
Diversidade de presas por espécie de lagarto e sobreposição nas dietas
Em relação à diversidade de presas, foi encontrado para T. semitaeniatus o índice de
H’= 1,63 na estação seca e H’= 1,43 na chuvosa; e para T. hispidus foi encontrado o índice de
H’= 1,05 na estação seca e H’= 1,60 na chuvosa.

A dieta das duas espécies teve sobreposição de jk = 0,886 por número de itens e jk =
0,995 por volume, na estação seca. Na estação chuvosa, as dietas tiveram uma sobreposição
de jk = 0,690 por número de itens e jk = 0,370 por volume.
Comportamento de forrageio
As observações comportamentais totalizaram 122 sessões de 10 minutos cada: 84 para
T. semitaeniatus (43 na estação seca e 41 na chuvosa) e 38 para T. hispidus (22 na estação
seca e 16 na chuvosa); o maior número de sessões para T. semitaeniatus é resultado do maior
número de avistamentos desta espécie.
Durante a estação seca não houve diferenças significativas em nenhuma das
categorias comportamentais analisadas: Número de deslocamentos, T. semitaeniatus (12,5 ±
9,2, n = 43) e T. hispidus (10,1 ± 7,0, n = 22; Teste U de Mann-Whitney, z = -0,887, p =
0,374); Tempo parado (%), T. semitaeniatus (96,6 ± 3,7 %, n = 43) e T. hispidus (97,5 ± 2,3
%, n = 22; Teste U de Mann-Whitney, z = -0,630, p = 0,528); Distância percorrida (cm), T.
semitaeniatus (212,7 ± 151,1, n = 43) e T. hispidus (252,5 ± 225,9, n = 22; Teste U de Mann-
Whitney, z = -0,263, p = 0,792) e Tentativas de capturas de presas, T. semitaeniatus (1,3 ±
1,5, n = 43) e T. hispidus (1,8 ± 2,0, n = 22; Teste U de Mann-Whitney, z = -1,116, p =
0,264).
Na estação chuvosa só não ocorreu diferença significativa na categoria
comportamental, Distância percorrida (cm), T. semitaeniatus (185,0 ± 141,9, n = 41) e T.
hispidus (144,3 ± 182,6, n = 16; Teste U de Mann-Whitney, z = 1,740, p = 0,081). Nas outras
categorias avaliadas ocorreram diferenças significativas: Número de deslocamentos, T.
semitaeniatus (9,8 ± 8,0, n = 41) e T. hispidus (3,0 ± 3,4, n = 16; Teste U de Mann-Whitney, z
= 3,827, p < 0,001); Tempo parado (%), T semitaeniatus (97,8 ± 2,2, n = 41) e T. hispidus
(99,4 ± 0,6, n = 16; Teste U de Mann-Whitney, z = -3,827, p < 0,001) e Tentativa de captura

de presa, T. semitaeniatus (1,7 ± 2,0, n = 41) e T. hispidus (0,3 ± 0,7, n = 16; Teste U de
Mann-Whitney, z = 2,770, p = 0,003).

DISCUSSÃO
Dieta
Composição, diferenças sazonais, sexuais e ontogenéticas
Por terem consumido várias categorias de artrópodes e de material vegetal, Tropidurus
semitaeniatus e T. hispidus podem ser consideradas generalistas, embora predominantemente
insetívoras, com dietas semelhantes às registradas para outras espécies deste gênero (Colli et
al. 1992, Van Sluys 1993, Vitt 1993, Vitt et al. 1996, Teixeira & Giovanelli 1999, Faria &
Araújo 2004, Vans Sluys et al. 2004, Faria 2006, Lima & Rocha 2006).
Quanto à ausência de significância entre os tipos de itens consumidos por Tropidurus
semitaeniatus e por T. hispidus em todas as estações, algumas considerações merecem ser
destacadas. Na estação seca, a categoria alimentar que obteve o maior índice de importância
relativa (Ix) para ambas as espécies foi folhas (Ix = 43,2 e Ix = 47,5 respectivamente, Tabela
I) mostrando um alto consumo de material vegetal durante esta estação, fato que pode estar
relacionado à escassez de outros recursos e à maior disponibilidade de folhas no ambiente,
resultante do processo de caducifolia da vegetação. Resultado semelhante foi obtido para
Tropidurus itambere (Van Sluys 1993) e para Liolaemos lutzae (Rocha 1996), ambos
estudados em formações abertas, embora com sazonalidade menos acentuada que nas
Caatingas. Portanto, o consumo de material vegetal parece ser comum para lagartos
filogeneticamente próximos e habitantes de formações abertas.
Outro aspecto relevante na composição da dieta foi a alta importância de
Hymenoptera/Formicidae na dieta de ambas as espécies durante as duas estações. Apesar da
alta importância de formigas, a dieta não chega a ser praticamente exclusiva de formigas
como acontece em T. hispidus de área florestada da Amazônia (Vitt et al. 1996). A alta
importância de larvas de Lepidoptera na dieta das duas espécies na estação chuvosa também

foi registrada para T. hispidus em afloramentos rochosos na região de Diamantina - MG (Van
Sluys et al. 2004). A alta importância de Orthoptera para T. hispidus na estação chuvosa deve
estar relacionada com a maior disponibilidade dessa presa nesse período, além de constituir
uma presa volumetricamente importante e também por este tipo de presa, assim como as
formigas, serem consideradas ativas, portanto mais consumidas por lagartos que caçam de
tocaia (sit-and-wait lizards; Huey & Pianka 1981, Faria, 2006).
A dieta de T. semitaeniatus não apresentou diferenças significativas sazonais nem
sexuais, mas ontogenética quanto ao número e volume de itens. Já para T. hispidus esta
diferença foi constatada apenas entre jovens e adultos quanto ao volume dos itens. Diferenças
ontogenéticas são comuns a várias espécies de lagartos filogeneticamente próximos (Van
Sluys 1993, Rocha 1998).
Relação entre a dieta das espécies e o Índice de Disponibilidade Relativa de Artrópodes no
Ambiente (IDRA)
A relação significativa (por freqüência de ocorrência e número de itens) entre a dieta
de T. semitaeniatus e o índice de disponibilidade relativa de artrópodes no ambiente (IDRA),
nas estações seca e chuvosa, indica que essa espécie é oportunista, ou seja, se alimenta do que
está mais disponível no ambiente; esta relação positiva também foi encontrada em estudo com
as espécies congenéricas Tropidurus itambere e Tropidurus oreadicus (Faria 2006), fato este
que concorda com o estabelecido pela teoria do forrageamento ótimo (Huey & Pianka 1981),
de que espécies de lagartos que forrageiam de espreita apresentam dietas mais generalizadas,
capturadas de acordo com a disponibilidade (Huey & Pianka 1981).
A dieta de T. hispidus possui relação significativa (por freqüência de ocorrência e
número de itens) com o IDRA apenas na estação chuvosa; na estação seca parece desenvolver
estratégia para capturar, de tocaia, presas que vivem agrupadas, como as formigas, gastando

assim menos energia do que gastaria buscando ativamente presas em ambiente durante
escassez.
Tamanho dos itens alimentares: relação com as medidas morfométricas e com variações inter
e intraespecíficas
Os adultos de Tropidurus semitaeniatus mostraram relação significativa entre o
tamanho médio das presas e do Rostro-comissura labial, que corresponde ao tamanho da
boca. Então, os indivíduos adultos de T. semitaeniatus que possuem a maior boca, se
alimentam de presas maiores. Nos jovens de T. semitaeniatus todas as medidas dos indivíduos
tiveram relação positiva significativa com o tamanho da presa. Essa correlação positiva entre
o tamanho da presa e as medidas dos lagartos parece ser comum para outras espécies de
lagartos (Vitt et al. 1996, Vans Sluys et al. 2004)
Todas as medidas dos indivíduos de T. hispidus (adultos e jovens) tiveram relação
positiva com o tamanho da presa embora nenhuma dessas relações tenha sido significativa.
No que se refere a diferenças intra-específicas quanto ao tamanho das presas ingeridas,
as duas espécies se comportam de maneira semelhante; ou seja, machos e fêmeas não diferem
significativamente quanto ao tamanho das presas ingeridas, embora adultos se alimentem de
presas maiores do que os jovens. Na estação chuvosa, os indivíduos se alimentam de presas
maiores do que na estação seca; essa diferença sazonal no tamanho das presas pode estar
relacionada com o fato de que na estação chuvosa a diversidade de presas é maior e permite
escolha por presas maiores, proporcionando melhor custo-benefício.
Tropidurus hispidus se alimenta de presas significativamente maiores do que T.
semitaeniatus tanto na dieta geral de todos os indivíduos, como entre os machos adultos,
fêmeas adultas e entre os jovens; essa diferença no tamanho da dieta é explicada pelo fato de
T. hispidus ser maior do que T. semitaeniatus, mas difere do que ocorre em duas espécies de

Tropidurus simpátricas no cerrado, as quais ingerem presas do mesmo tamanho (Faria &
Araújo 2004). vale ressaltar, entretanto, que as espécies do Cerrado têm tamanhos
semelhantes.
Diversidade de presas por espécie de lagartos e sobreposição nas dietas
A maior sobreposição nas dietas de T. semitaeniatus e de T. hispidus durante a estação
seca deve estar relacionada com a menor disponibilidade de alimento nesta estação, fato que
leva as duas espécies a se alimentarem de itens semelhantes, e coexistirem em condições
limitantes. Esta alta sobreposição durante a estação seca foi encontrada para outras espécies
sintópicas de Tropidurus, no Cerrado (Faria & Araujo 2004).
Quanto à diversidade de presas na dieta, T. semitaeniatus teve a diversidade
semelhante, quanto ao número de itens, nas estações seca e chuvosa; já quanto ao volume, a
diversidade na estação chuvosa foi bem maior. O mesmo foi observado para T. hispidus,
sendo que na estação chuvosa a diversidade quanto ao volume reflete a maior disponibilidade
de presas maiores durante a estação chuvosa, reforçando o caráter generalista e oportunista
desta espécie.
Comportamento de forrageio
As espécies de Tropidurus são consideradas forrageadoras de tocaia (sit-and-wait
lizards), e apesar de existir um gradiente de variação na intensidade de forrageio e uma
mudança de estratégia de acordo com a disponibilidade de alimento no ambiente proposto por
Huey & Pianka (1981). Neste estudo, T. semitaeniatus se mostrou mais ativo do que T.
hispidus, principalmente durante a estação chuvosa onde as diferenças na intensidade de
forrageio foram significativas.

Para as categorias comportamentais analisadas, apenas para a estação chuvosa
ocorreram diferenças significativas. Na estação seca é possível que as condições mais
adversas possam afetar de igual modo as duas espécies, no que se refere à escassez de
disponibilidade de diferentes recursos alimentares, a ponto de não mostrarem diferenças
significativas em suas estratégias de caça. Já na estação chuvosa, T. semitaeniatus apresentou
maior número de deslocamentos, permaneceu menos tempo parado e ainda exibiu maior
número de tentativas de captura de presas do que T. hispidus. Nesta estação, em que há maior
oferta de alimento, as espécies utilizaram estratégias de forrageio diferentes, de modo que T.
semitaeniatus se alimenta de presas menores do que T. hispidus. Aliado a isso, o maior
tamanho de T. hispidus o habilita a capturar presas maiores.
Observações de comportamento de forrageio para T. hispidus realizadas por Vitt et al.
(1996) durante estação chuvosa, na região amazônica, corroboram estes resultados, uma vez
que a média de deslocamentos e de tentativas de capturas de presas foram similares,
respectivamente 4,9 e 0,36.
Quanto à distância percorrida pelos lagartos, não ocorreu diferença significativa entre
as espécies em nenhuma das estações. Na estação seca, T. semitaeniatus e T. hispidus
realizaram número similar de deslocamentos, percorrendo a mesma distância, ao passo que na
estação chuvosa os espécimes de T. hispidus realizaram menor número de deslocamentos,
entretanto mais longos, mantendo a distância percorrida semelhante àquela de T.
semitaeniatus.

CONCLUSÕES
Tropidurus semitaeniatus e Tropidurus hispidus são generalistas, pois se alimentam
dos itens mais disponíveis no ambiente, embora possuam um certo grau de especialização em
suas dietas como o consumo maior de folhas, formigas e cupins na estação seca. Estas
espécies se sobrepõem amplamente quanto à composição da dieta em ambas as estações; no
entanto, durante a estação chuvosa, com o aumento da disponibilidade de alimento, T.
hispidus se alimenta de presas maiores e T. semitaeniatus é mais ativo, apesar de ambas as
espécies serem forrageadoras de tocaia; estas estratégias contribuem para a coexistência
destas espécies. Durante a estação seca, a alta sobreposição da dieta e a semelhança nas
estratégias de forrageio, constituem fatores limitantes.

REFERÊNCIAS BIBLIOGRÁFICAS
Ab’Sáber, A. 1974. O Domínio Morfoclimático Semiárido das Caatingas Brasileiras. Geomorfologia, 43, 1 – 139.
Altmann, J. 1974. Observational study of behaviour: sampling methods. Behaviour, 49: 227-267.
Bergallo, H.G. & Rocha, C. F. D. 1994. Spatial and trophic niche differentiation in two sympatric lizards (Tropidurus torquatus and Cnemidophorus ocellifer) with different foraging tactics. Australian Journal of Ecology, 19: 72-75.
Colli, G. R. Araújo, A. F. B. Silveira, R. & Roma, F. 1992. Niche partitioning and morphology of two syntopic Tropidurus (Sauria: Tropiduridae) in Mato Grosso, Brazil. Journal of Herpetology. 26 (1): 66-69.
Cooper, W. E. J. 1995. Foraging mode, prey chemical discrimination, and phylogeny in lizards. Animal Behaviour, 50 (4): 973-985.
Costa, T. B. G. & Freire, E. M. X. in press. Estrutura da comunidade de serpentes de uma área de Caatinga do Nordeste do brasileiro.
Dunham, A. E. 1983. Realized Niche Overlap, Resource Abundance, and Interespecifc Competition. pp. 261-280. In: R. B. Huey, E. R. Pianka & T. W. Schoener (Eds.), Lizard Ecology: Studies of a Model Organism., Harvard University Press, Harvard Massachusetts.
Faria, R. G. 2006. Estudo da interação entre Tropidurus oreadicus e Tropidurus itambere(Iguanidae: Tropidurinae), em áreas de cerrado rupestre do Brasil Central: uma abordagem comparativa e experimental. Tese de Doutorado. Universidade de Brasília. Brasilia.
Faria, R. G. & Araújo, A. F. B. 2004. Sintopy of two Tropidurus species (Squamata: Tropiduridae) in a rocky Cerrado habitat in central Brazil. Brazilian Journal of Biology, 64(4): 775-786.
Freire, E. M. X. 2001. Composição, Taxonomia, Diversidade e considerações Zoogeográficas sobre a Fauna de Lagartos e Serpentes de Remanescentes da Mata Atlântica do Estado de Alagoas, Brasil. Tese de Doutorado. Universidade Federal do Rio de Janeiro. Rio de Janeiro.
Freitas, M. A. & Pavie, I. 2002. Guia de répteis da região metropolitana de Salvador e litoral norte da Bahia. Malha-de-Sapo-Publicações, Lauro de Freitas/BA, 72 pp.
Gasnier, T. R. Magnusson, W. E. & Lima, A. P. 1994. Foraging activity and diet of four sympatric lizard species in a tropical rainforest. Journal of Herpetology, 28(2): 187-192.
Huey, R. B. & Pianka, E. R. 1981. Ecological consequences of foraging mode. Ecology, 62(4): 991-999.

Huey, R. B. Pianka, E. R. & Schoener, T. W. (Eds.) 1983. Lizard Ecology: Studies of a Model Organism. Harvard University Press, Cambridge, Massachusetts, 501 pp.
Lima, A. F. B. & Rocha, P. L. B. 2006. Ontogenetic change in plant consumption by Tropidurus psammonastes, Rodrigues, Kasahara & Yonenaga-Yassuda, 1988 (Tropiduridae), a lizard endemic to the dunes of the São Francisco River, Bahia, Brasil. Revista Brasileira de Zoociências, 8 (1): 67-75.
Maggi, B. S. Freire, E. M. X. & Rodrigues, L. S. in press. Resource partitioning and foraging behavior in two species of geckos(Squamata, Gekkonidae) in syntopy in the Caatinga of Northeast Brazil.
Magnusson, W. E. Paiva, L. J. Rocha, R. M. Franke, C. R. Kasper, L. A. & Lima, A. P.1985. The correlates of foraging mode in a community of Brazilian lizards. Herpetologica,41(3): 324-332.
Magurram, A. E. 1988. Ecological Diversity and its Measurement. Cambridge, University Press, London, 179 pp.
Martin, P. & Bateson, P. (1993). Measures of behaviour; pp. 63-83. In: Measuring Behaviour: an introductory guide. Cambridge University press.
Menezes, V. A. Amaral, V. C. Van Sluys, M. & Rocha, C. F. D. 2006. Diet and foraging of the endemic lizard Cnemidophorus littoralis (Squamata, Teiidae in the resting de Jurubatiba, Macaé, RJ. Brasilian Journal of Biology, 66 (3): 803-807.
Pianka, E. R. 1966a. Convexity, Desert Lizards, and Spatial Heterogeneity. Ecology, 47,1055-1059.
Pianka, E. R. 1966b. On Optimal Use of a Patchy Environment. The American Naturalist,100, 916, 603-609.
Pianka, E. R. 1969. Habitat Specificity, Speciation, and Species Density in Australian Deserts Lizards. Ecology, 50, 498-502.
Pianka, E. R. 1971. Lizard Species Density in the Kalahari Desert. Ecology, 52, 1024-1029.
Pianka, E. R. 1973. The Structure of Lizard Communities. Annual Reviews of Ecology and Systematics, 4, 53-74.
Pianka. E. R. 1986. Ecology and natural history of desert lizards. Princenton University Press. 208pp.
Pough, F. H. Janis, C. M. & Heiser, J. B. 2003. A Vida dos Vertebrados. 3ª Ed., Editora Atheneu, São Paulo, 699 pp.
Rocha, C. F. D. 1989. Diet of a tropical lizard (Liolaemus lutzae) of Southeastern Brazil. Journal of Herpetology, 23(3): 292-294.

Rocha, C. F. D. 1992. Liolaemus lutzae (sand lizard): Cannibalism. Herpetological Review,23(2): 60.
Rocha, C. F. D. 1994 A ecologia de lagartos no Brasil: Status e perspectivas In: Herpetologia no Brasil,1, pp. 35-57. Universidade Católica de Minas Gerais, MG.
Rocha, C. F. D. 1996. Seasonal shift in lizard diet: the seasonality in food resources affecting the diet of Liolaemus lutzae (Tropiduridae). Ciência e Cultura, 48(4): 264-269.
Rocha, C. F. D. 1998. Ontogenetic shift in the rate of plant consumption in a tropical lizard (Liolaemus lutzae). Journal of Herpetology, 32(2): 274-279.
Rocha, C. F. D. & Anjos, L. A. 2007. Feeding ecology of a nocturnal invasive alien lizard species, Hemidactylus mabouia Moreau de Jonnès, 1818 (Gekkonidae), living in na outcrop rocky área in southeastern Brasil. Brazilian Journal of Biology, 67 (3): 485-491.
Rodrigues, M. T. 1986. Uma Nova Espécie do Gênero Phyllopezus de Cabaceiras: Paraíba: Brasil; com comentários Sobre a Fauna de Lagartos da Área (Sáuria Gekkonidae). PapeisAvulsos de Zoologia, 36, 20, 237-250.
Rodrigues, M. T. 1987. Sistemática, ecologia e zoogeografia dos Tropidurus do grupo torquatus ao Sul do Rio Amazonas (Sauria, Iguanidae). Arquivos de Zoologia do Estado de São Paulo, 31(3): 105-230.
Rodrigues, M. T. 1991 a. Herpetofauna das dunas interiores do Rio São Francisco: Bahia: Brasil: I. Introdução à área e descrição de um novo gênero de microteiídeos (Calyptommatus)com notas sobre sua ecologia, distribuição e especiação (Sauria, Teiidae). Papeis Avulsos de Zoologia, São Paulo, 37 (19): 285-320.
Rodrigues, M. T. 1991 b. Herpetofauna das dunas interiores do Rio São Francisco: Bahia: Brasil: II. Psilophthalmus: um novo gênero de microteídeos sem pálpebras (Sauria, Teiidae). Papeis Avulsos de Zoologia, São Paulo, 37 (20): 321-327.
Rodrigues, M. T. 1991 c. Herpetofauna das dunas interiores do Rio São Francisco: Bahia: Brasil: III. Procellosaurinus: um novo gênero de microteiídeos sem pálpebras, com a redefinição do gênero Gymnophthalmus (Sauria, Teiidae). Papeis Avulsos de Zoologia, SãoPaulo, 37 (21): 329-342.
Rodrigues, M. T. 1996. Lizards, snakes and amphisbaenians of the quaternary sand dunes of the rio São Francisco: Bahia: Brazil. Journal of Herpetology, 30 (04): 513-523.
Rodrigues, M. T. 2000. A new specie of Mabuya (squamata: Scincidae) from the semiarid Caatingas of northeastern Brazil. Papeis Avulsos de Zoologia, São Paulo, 41 (21): 313-328.
Rodrigues, M. T. (coord.). 2002. A fauna de Répteis e Anfíbios das Catingas. In: Biodiversidade Brasileira – Avaliação e identificação de áreas e ações prioritárias para a Conservação, Utilização Sustentável e Repartição de Benefícios da Biodiversidade Brasileira. Ministério do Meio Ambiente, Secretaria de Biodiversidade e Florestas, Brasília-DF. 404 pp.

Rodrigues, M. T. 2003. Herpetofauna da Caatinga. In: Ecologia e Conservação da Caatinga. Ed. Universitária, UFPE, Recife-PE.
Rodrigues, M. T. 2005. Conservação dos répteis brasileiros: os desafios de um país megadiverso. Megadiversidade (1): 87-94.
Schoener, T. W. 1977. Competition and niche. Pp. 35-136. In: Biology of the Reptilia. C. Gans & D.W. Tinkle (eds.), Academic Press, New York.
Strüsmann, C. Vale, M. B. R. Meneghini, M. H. & Magnusson, W. E. 1984. Diet and foraging mode of Bufo marinus and Leptodactylus ocellatus. Journal of Herpetology, 18(2):138-146.
Teixeira, R. L. & Giovanelli, M. 1999. Ecologia de Tropidurus torquatus (Sauria: Tropiduridae) da restinga de Guriri, São Mateus, ES. Revista Brasileira de Biologia, 59(1):11-18.
Uetz, P. Etzold, T. & Chenna, R. 2008. The EMBL Reptile Data Base, http://www.reptile-database.org/ acessado em 06/08/2008.
Van Sluys, M. 1991. Dieta de Tropidurus itambere Rodrigues (Sauria; Iguanidae) na Fazenda Manga, município de Valinhos, São Paulo. Dissertação de Mestrado, Inst. Biologia, Universidade Estadual de Campinas, 65 pp.
Van Sluys, M. 1993. Food habits of the lizard Tropidurus itambere (Tropiduridae) in Southeastern Brazil. Journal of Herpetology, 27(3): 347-351.
Van Sluys, M. Rocha, C. F. D. Vrcibradic, D. Galdino, C. A. B. & Fontes, A. F. 2004. Diet, activity and microhabitat use of two syntopic Tropidurus species (Lacertilia: Tropiduridae) in Minas Gerais, Brazil. Journal of Herpetology, 38(4): 606-611.
Vanzolini, P. E. 1974. Ecological and geographical distribution of lizards in Pernambuco, northeastern Brazil (Sauria). Papéis Avulsos de Zoologia, 28: 61-90.
Vanzolini, P. E. 1976. On the lizards of a Cerrado-Caatinga contact: evolutionary and zoogeographical implications (Sauria). Papeis Avulsos de Zoologia, 29: 111-119.
Vanzolini, P. E. Ramos-Costa, A. M. M. & Vitt, L. J. 1980. Répteis das Caatingas. Academia Brasileira de Ciências, Rio de Janeiro, 161 p.
Varela-Freire, A. A. 2002. A Caatinga Hiperxerófila Seridó, a sua caracterização e estratégias para sua conservação Academia de ciências do estado de São Paulo.
Velloso A. L. Sampaio, E. V. S. B. & Pareyn, F. G. C. (Eds). 2002. Ecorregiões propostas para p Bioma Caatinga. Instituto de Conservação Ambiental The Nature Conservancy do Brasil, 76 pp.
Vitt, L. J. 1990. The influence of foraging mode and phylogeny on seasonality of tropical lizard reproduction. Papéis Avulsos de Zoologia, 37(6): 107-123.

Vitt, L. J. 1993. Ecology of isolated open-formation Tropidurus (Reptilia: Tropiduridae) in Amazonian lowland rain forest. Canadian Journal of Zoology. 71: 2370-2390.
Vitt, L. J. 1995. The ecology of tropical lizards in the Caatinga of Northeast Brazil. Occasional Papers of the Oklahoma Museum of Natural History, 1: 1-29
Vitt, L. J. & Vangilder, L. D. 1983. Ecology of a snake community in Nordeasthern Brazil. Amphibia-Reptilia, 4, 273-296.
Vitt, L. J. Zani, P. A. & Caldwell, J. P. 1996. Behavioural ecology of Tropidurus hispiduson isolated rock outcrops in Amazonia. Journal of Tropical Ecology, 12: 81-101.
Vitt, L. J. Zani, P. A. & Ávila-Pires, T. C. S. 1997. Ecology of the arboreal tropidurid lizard Tropidurus (= Plica) umbra in the Amazon region. Canadian Journal of Zoology, 75(11):1876-1882.
Vitt, L. J. Sartorius, S. S. Ávila-Pires, T. C. S. & Espósito, M. C. 2001. Life at river`s edge: ecology of Kentropyx altamazonica in Brazilian Amazonia. Canadian Journal of Zoology. (79): 1855-1865.
Vrcibradic, D. & Rocha, C. F. D. 1995. Variação sazonal na dieta de Mabuyamacrorhyncha (Sauria, Scincidae) na restinga da Barra de Maricá, RJ. Oecologia Brasiliensis,1: 143-153.
Vrcibradic, D. & Rocha, C. F. D. 1996. Ecological differences in tropical sympatric skinks (Mabuya macrorhyncha and Mabuya agilis) in Southeastern Brazil. Journal of Herpetology,30 (1): 60-67.
Vrcibradic, D. & Rocha, C. F. D. 1998. The ecology of the skink Mabuya frenata in an area of rock outcrops in Southeastern Brazil. Journal of Herpetology, 32(2): 229-237.
Zar, J. H. 1999. Biostatistical analysis. 4ª ed., Upper Saddle River, Prentice-Hall Inc., 663 pp.


















