
MARIANA BARROS FRANCO DE ANDRADE VILÉLA - uel.br Viléla Monografia.pdf · 2 MARIANA BARROS FRANCO...
59
1 MARIANA BARROS FRANCO DE ANDRADE VILÉLA ESTRESSE OXIDATIVO EM Prochilodus lineatus EXPOSTOS AO CHUMBO Londrina – Paraná 2007
Transcript of MARIANA BARROS FRANCO DE ANDRADE VILÉLA - uel.br Viléla Monografia.pdf · 2 MARIANA BARROS FRANCO...

1
MARIANA BARROS FRANCO DE ANDRADE VILÉLA
ESTRESSE OXIDATIVO EM Prochilodus lineatus EXPOSTOS AO CHUMBO
Londrina – Paraná
2007

2
MARIANA BARROS FRANCO DE ANDRADE VILÉLA
ESTRESSE OXIDATIVO EM Prochilodus lineatus EXPOSTOS AO CHUMBO
Supervisor de Estágio: Cláudia Bueno dos Reis Martinez
Londrina – Paraná 2007
Monografia apresentada ao Curso de Graduação em Ciências Biológicas da
Universidade Estadual de Londrina como um dos requisitos à obtenção do
título de Bacharel em Ciências Biológicas.

3
BANCA EXAMINADORA
Profª. Dra. Cláudia Bueno dos Reis Martinez
Universidade Estadual de Londrina
Profª Dra. Estefânia Gastaldello Moreira Universidade Estadual de Londrina
Ms. Lindalva Pereira Maduenho
Universidade Estadual de Londrina
Londrina, 07 de dezembro de 2007

4
DEDICATÓRIA
A todos que durante este ano
me auxiliaram de alguma forma e
tornaram meus dias mais felizes...

5
AGRADECIMENTOS
A Deus, pela persistência, orientação e pelo grande aprendizado proporcionado por mais essa experiência. À minha orientadora, Profª. Dra. Cláudia Bueno dos Reis Martinez por sua dedicação, paciência, sensibilidade e apoio ao longo desta jornada. À Profa. Dra. Estefânia Gastaldello Moreira e a Mestre Lindalva Pereira Maduenho que fizeram parte da banca examinadora deste trabalho, pelas correções, críticas e sugestões. À Káthya, pelas explicações e companhia na hora dos ensaios e ainda nos passeios ao shopping. A todos que fazem parte do LEFA, que de alguma forma contribuíram para o meu crescimento e me deram apoio. A meus pais, que mesmo com todas as dificuldades não pouparam esforços para me educar e oferecer as condições necessárias para que eu estudasse, aos quais devo, em grande parte, o que hoje sou. À minha irmã Liliam, pela companhia nos momentos mais difíceis. Ao Gui, pelo seu jeitinho terno que me acalma. Ao Lucas, pela paciência, compreensão e principalmente pela companhia, te amo muito! À Cássia, Véssia e á Ericka, que juntas dividimos não só a nossa casa, mas as alegrias e tristezas. Ao tio Paulo, que mesmo distante também me auxiliou com valiosos conselhos e ainda me proporcionou o estudo de inglês que foi muito útil nesse trabalho. À Luciana Festti, minha grande amiga, que me confortou durante todo esse tempo via e-mails, e faz muita falta aqui do meu lado. Companheira de trabalhos, exames, risadas e muitas decisões acertadas, essencial durante toda a faculdade. A todos os meus amigos e familiares que acreditaram que eu era capaz. Aos funcionários da Estação de Piscicultura (EPUEL), pelo fornecimento dos peixes. E a todos aqueles que de alguma forma contribuíram para esta dissertação tornar-se realidade, MUITO OBRIGADA!

6
“Hoje em dia, o ser humano apenas tem ante si três grandes problemas que foram ironicamente provocados por ele próprio:
a super povoação, o desaparecimento dos recursos naturais e a destruição do meio ambiente. Triunfar sobre estes problemas, visto sermos nós a sua causa, deveria ser a nossa mais profunda
motivação.” Jacques Yves Cousteau

7
RESUMO
ESTRESSE OXIDATIVO NO PEIXE Prochilodus lineatus APÓS EXPOSIÇÃO AGUDA AO CHUMBO.
Mariana Barros Franco de Andrade Viléla
Devido às diversas ocorrências de contaminação de ambientes aquáticos com chumbo, este trabalho foi desenvolvido com Prochilodus lineatus, peixe de hábito alimentar detritívoro, que pode ser contaminado por metais que se acumulam no sedimento. Assim, foram feitos testes de toxicidade estáticos agudos para avaliar os possíveis efeitos do chumbo em parâmetros de estresse oxidativo da espécie de peixe neotropical P. lineatus. Peixes jovens foram expostos a 0,5 ppm de chumbo dissolvido (EXP) ou apenas à água (CTR) durante 6, 24 e 96h em aquários de vidro de 100L com uma média de sete peixes em cada. Os experimentos foram feitos com réplica. Após exposição os peixes foram anestesiados com benzocaína e mortos por secção medular para retirada do cérebro, músculo e fígado. Os órgãos foram congelados a –80º C para análises posteriores. Foram determinadas as atividades das enzimas acetilcolinesterase, catalase (CAT), glutationa peroxidase (GPx) e glutationa-S-transferase (GST), as concentrações de glutationa reduzida (GSH), metalotioneína e a lipoperoxidação (ensaio FOX). Após 6 horas de exposição ao Pb, a CAT, GSH e a MET aumentaram significativamente, enquanto a GPx, GST e FOX apresentaram tendência ao aumento. Em 24 h os peixes apresentaram decréscimo significativo na atividade da GST e tendência de redução da CAT e GSH, acompanhadas de aumento significativo da GPx e MET, e tendência da FOX. E no tempo de 96 horas, houve decréscimo significativo da GST, a GSH e a FOX apresentaram aumento significativo e a CAT, GPx e MET também tiveram uma tendência a aumentar. A acetilcolinesterase apresentou uma diminuição significativa em todos os tempos experimentais. Esses resultados indicam que o Pb na concentração testada agiu como anti-colinesterásico; promoveu a ativação das defesas antioxidantes enzimáticas e não-enzimáticas; inibição da enzima de conjugação GST, provavelmente devido ao uso da GSH na associação com o Pb; aumentou a peroxidação lipídica, sugerindo que houve danos ao fígado do animal e as defesas antioxidantes não foram suficientes para evitar o estresse oxidativo. Palavras-chave: chumbo, estresse oxidativo, fígado, Prochilodus lineatus.

8
SUMÁRIO
Pág.
1. INTRODUÇÃO
1.1 Contaminação ambiental 01
1.2 Toxicologia aquática 02
1.3 Exposição ao xenobiótico 03
1.4 Chumbo 07
1.5 Prochilodus lineatus 09
1.6 Biomarcadores 10
2. OBJETIVOS 17
3. ARTIGO: Estresse oxidativo em Prochilodus Lineatus expostos ao chumbo 18
Resumo 19
Introdução 20
Materiais e Métodos 22
Resultados 26
Discussão 32
Conclusão 37
Referências Bibliográficas 38
4. REFERÊNCIAS BIBLIOGRÁFICAS GERAIS 43

1
1. INTRODUÇÃO
1.1 Contaminação ambiental
Grande parte dos estudos do século XXI baseia-se principalmente na
problemática dos recursos hídricos em nível nacional e internacional. A água é
indispensável para a sustentação da vida e o que vem ocorrendo é a degradação da
sua qualidade, ressaltando a contaminação química pela agricultura e também por
indústrias. Segundo Heath (1995) a poluição aquática consiste na adição à água de
qualquer substância que altere sua composição química, temperatura ou
composição microbiológica.
Os efluentes gerados a partir de atividades industriais, muitas vezes contém
metais pesados que contaminam as águas. As principais fontes de contaminação
com metais derivam de diversas atividades humanas, tais como metalurgia,
mineração, fundição e galvanoplastia, indústrias de couro, gases liberados pela
queima de combustíveis fósseis como a gasolina e a fabricação e descarte de
baterias, entre outros (CHAOUI et al., 1997; JORDÃO et al., 1999; GIMENO-
GARCIA et al., 1996; SCHICKLER & CASPI, 1999). Nas últimas cinco décadas,
foram liberadas na biosfera 22.000 toneladas de cádmio, 939.000 t de cobre,
783.000 t de chumbo e 1.350.000 t de zinco, como produto da industrialização global
(SINGH et al., 2003). Estes contaminantes podem causar efeitos adversos aos
organismos. Além disso, causam prejuízos aos ecossistemas aquáticos, pois tendem
a se acumular na biota aquática.

2
1.2 Toxicologia aquática
A avaliação dos efeitos de contaminantes nos ecossistemas aquáticos vem
sendo alvo de várias pesquisas na área da ecotoxicologia. Os peixes estão
representados por mais de 21.000 espécies entre as espécies de água doce e
marinha, e estão presentes em quase todos os ecossistemas aquáticos incluindo os
mais poluídos. Devido à sua capacidade de adaptação às variações ambientais, os
peixes são considerados modelos interessantes para estudos em ecotoxicologia. Em
áreas poluídas, os peixes encontram-se submetidos ao estresse químico que pode
ocasionar alterações em órgãos alvos afetando diferentes sistemas, dentre eles o
sistema imune (BETOULLE, 1998).
Os estudos em toxicologia aquática são qualitativos e quantitativos em
relação aos efeitos tóxicos sobre os organismos aquáticos. Os efeitos tóxicos podem
incluir tanto a letalidade (mortalidade) e efeitos subletais, como alterações no
crescimento, desenvolvimento, reprodução, respostas farmacocinéticas, patologia,
bioquímica, fisiologia e comportamento. Os efeitos podem ser expressos através de
critérios mensuráveis como o número de organismos mortos, alterações no tamanho
e peso, porcentagem de inibição de enzima, incidência de tumor, dentre outros. A
toxicologia aquática também está relacionada com as concentrações ou quantidades
dos agentes químicos que podem ocorrer no ambiente aquático (água, sedimento ou
alimento) (RAND & PETROCELLI, 1985). O efeito tóxico de um composto químico
depende da exposição, da suscetibilidade do organismo, das características
químicas do agente e de fatores ambientais.
A legislação brasileira determina que, quando apropriado, a qualidade dos
ambientes aquáticos pode ser avaliada por indicadores biológicos, utilizando-se
organismos e comunidades aquáticas (CONAMA 357, 2005).

3
1.3 Exposição ao xenobiótico
A exposição é o contato entre o organismo e o composto químico, sendo que
os fatores mais importantes relacionados à exposição são: o tipo, duração,
freqüência da exposição e a concentração do agente químico. (RAND &
PETROCELLI, 1985).
Na exposição aguda, os organismos entram em contato com o composto
químico em um evento único ou em eventos múltiplos que ocorrem em pequenos
períodos de tempo, geralmente variando de horas a dias. Nas exposições agudas
onde o agente químico é rapidamente absorvido normalmente os efeitos são
imediatos, embora seja possível a produção de efeitos retardados similares àqueles
resultantes da exposição crônica. Na exposição crônica normalmente os organismos
são expostos a baixas concentrações do agente tóxico que é liberado continuamente
ou com alguma periodicidade em um longo período de tempo (semanas, meses ou
anos). Exposição crônica a compostos químicos pode também induzir a efeitos
rápidos e imediatos, como os efeitos agudos, em adição aos efeitos que se
desenvolvem lentamente (OLIVEIRA et al., 2002).
A freqüência da exposição também afeta o efeito tóxico dos compostos
químicos. Uma exposição aguda a uma única concentração pode resultar em efeito
adverso imediato no organismo, enquanto duas exposições sucessivas cumulativas
iguais à exposição aguda única podem ter efeito pequeno ou nenhum efeito, devido
ao metabolismo (detoxificação) do organismo entre as exposições ou à adaptação
do organismo ao composto (SPRAGUE, 1985).
O efeito tóxico também depende da suscetibilidade dos organismos ao
composto químico. Diferentes espécies possuem suscetibilidades diferentes de
acordo com seu aparato metabólico, de acordo com seus hábitos alimentares, seu

4
comportamento, fase de desenvolvimento, dentre outros aspectos. Organismos
estressados em função de exposição prévia a outros tóxicos podem ser mais
suscetíveis aos compostos químicos, cenário comum na realidade dos
ecossistemas, pois normalmente há a presença simultânea de diferentes produtos
(RAND & PETROCELLI, 1985).
As características do composto químico também influenciam grandemente no
efeito tóxico como, por exemplo, sua composição, ou grau de pureza, pois as
impurezas ou contaminantes que são consideravelmente mais tóxicos do que o
agente propriamente dito, podem estar presentes. As propriedades físicas e
químicas como solubilidade, pressão de vapor e pH afetam a biodisponibilidade,
persistência, transformação, e o destino do agente químico no ambiente e também
são fatores importantes nos testes de toxicidade. Existem compostos químicos não
seletivos em seu modo de ação e que provocam efeitos indesejáveis em numerosas
células e tecidos dos organismos aquáticos. Em contraste, há os compostos com
modo de ação seletivo que afetam adversamente apenas um tipo de célula ou
tecido, sendo inofensivo para os demais com os quais esteve em contato direto,
assim, o modo de ação dos compostos químicos também afetam o efeito tóxico
(MALAVOLTA, 1994).
Os fatores ambientais definidos pelas características bióticas e abióticas
também podem alterar o efeito tóxico de compostos químicos no ambiente aquático.
Os fatores bióticos incluem o tipo de organismo (alga, inseto ou peixe), estágio de
desenvolvimento (larva, juvenil, adulto), tamanho, estado nutricional e de saúde,
alterações sazonais no estado fisiológico, sendo que estes fatores bióticos
influenciam a resposta ao poluente de diferentes maneiras. Os fatores abióticos que
podem atuar modificando o efeito tóxico incluem todas as características físicas e

5
químicas da água que circunda o organismo vivo, como a temperatura, o pH, o teor
de oxigênio dissolvido na água, a salinidade e a dureza, conteúdo de matéria
orgânica e material particulado em suspensão, a velocidade do fluxo da água, dentre
outros (SPRAGUE, 1985).
Para assegurar sua sobrevivência, os organismos desenvolveram
mecanismos para a proteção de suas células na presença xenobióticos. Estes
envolvem: a absorção, a distribuição, a biotransformação e a excreção das
substâncias estranhas. A absorção em peixes ocorre por várias rotas, incluindo a
alimentação e o transporte direto através das membranas externas, sendo então
distribuídos para as diversas partes do corpo.
A biotransformação (metabolização) geralmente leva à formação de
compostos mais polares, portanto, mais hidrofílicos, conseqüentemente mais
facilmente excretados do que o composto original. O órgão mais comumente
envolvido neste processo é o fígado. O fígado atua na emulsão de gorduras através
da produção da bile, na metabolização de substâncias presentes na corrente
sanguínea e produção de vários compostos, como por exemplo, proteínas
(LIVINGSTONE, 1998; VAN DER OOST, 2003).
A biotransformação geralmente envolve dois estágios distintos, referidos
como reações da Fase I e da Fase II. Na Fase I, a biotransformação de xenobióticos
ocorre através de reações de redução, hidrólise e/ou oxidação, sendo responsáveis
por introduzir grupos funcionais no xenobiótico. A principal reação é a oxidação,
catalisada principalmente pelo citocromo P-450 monooxigenase, pelas monoamino
oxidases (MAO) e pelas flavinas monooxigenase (FMO) (LIVINGSTONE, 1998).
Além disso, em múltiplas reações metabólicas de transferência de elétrons em
células aeróbicas são formadas espécies reativas de oxigênio (ERO). Estas

6
espécies reativas também podem resultar do metabolismo de certos xenobióticos e
causam peroxidação de lipídios, alterações em proteínas e em ácidos nucléicos,
produzindo danos às células. A proteção contra estas ERO é feita pelas enzimas,
catalase, GST, glutationa peroxidase e enzimas de reparo do DNA. Em geral, o
aumento da polaridade da molécula leva à perda do potencial tóxico do composto
sendo biotransformado, levando a desintoxicação. Porém, em alguns casos, pode
ocorrer a ativação metabólica da molécula e o xenobiótico pode ser convertido em
um produto com maior potencial tóxico (bioativação).
A longo prazo, de acordo com o agente estressor e tempo de exposição,
estas alterações podem gerar, por exemplo, redução na capacidade reprodutiva ou
tolerância aos fatores de estresse. A contaminação do animal ocorre através da
superfície respiratória branquial, responsável pela absorção direta de substâncias
presentes na água, ou através da ingestão de água e alimento contaminado,
podendo comprometer assim o funcionamento de brânquias, trato gastrointestinal,
fígado e rins. Em geral, os compostos tóxicos acabam gerando uma resposta
integrada, envolvendo diversos sistemas e a liberação de várias substâncias para
conter ou minimizar os efeitos tóxicos no metabolismo. Perturbações internas no
organismo dos peixes, como resultado de alterações do ambiente, geram respostas
compensatórias que, se não forem suficientes podem levar à morte do animal
(KENNEDY & FARRELL, 2005; MARTÍNEZ-ÁLVAREZ et al., 2002).
Adicionalmente, o metabolismo de compostos tóxicos freqüentemente resulta
na formação EROs e, quando a taxa de formação de ERO é excessiva, acaba
superando a capacidade fisiológica dos organismos de eliminá-las ou anulá-las,
criando a situação de estresse oxidativo. Esse processo pode também ser definido
como o reflexo do desequilíbrio existente entre a produção e a remoção destes

7
produtos oxidantes. Quando os mecanismos de defesa do animal, responsáveis pela
remoção destes compostos são inadequados ou insuficientes, aumentam o nível de
estresse oxidativo de tecidos animais, com dano às suas macromoléculas, o que
pode até acarretar a morte (CAZENAVE et al., 2006; GUERREIRO et al., 2002;
HERMES-LIMA & ZENTENO-SAVÍN, 2002; MARTÍNEZ-ÁLVAREZ et al., 2002).
Para combater essas espécies reativas de oxigênio existe um sistema de
defesa antioxidante, que utiliza mecanismos enzimáticos e não-enzimáticos.
(STOREY, 1996; VAN DER OOST et al., 2003).
1.4 Chumbo
O chumbo é um metal cinza-azulado de peso atômico 207.19, ponto de fusão
327.502° C e ponto de ebulição 1740° C (IPCS, 1995).
Este metal era conhecido pelos antigos egípcios, que devido ao seu baixo
ponto de fusão, durabilidade e facilidade em formar ligas metálicas era utilizado na
fabricação de armas, adornos e utensílios. Os antigos romanos usavam o chumbo
para fabricar manilhas, e alguns compostos do metal já eram usados na fabricação
de cosméticos e de tintas (MELLOR, 1967).
O chumbo é considerado um contaminante ambiental e as concentrações no
meio ambiente cresceram de acordo com o aumento do seu uso industrial. Com o
advento da Revolução Industrial, as concentrações de chumbo no meio ambiente
elevaram-se de forma alarmante, principalmente devido à introdução de compostos
orgânicos de chumbo (chumbo tetraetila) como aditivo para gasolina, mas em alguns
países, incluindo o Brasil, o uso se tornou restrito (WHO, 1995).
E nos últimos anos a demanda de chumbo tem sofrido uma mudança quanto
ao tipo de utilização. Seu emprego em aditivos para gasolina e tintas tem diminuído

8
bastante, porém existe um aumento significativo nos processos industriais. Usa-se o
chumbo na fabricação de canos, revestimentos, cabos elétricos, chapas para pias,
cisternas e telhados, na indústria de acumuladores, e outros.
O chumbo é tóxico para toda a biota aquática, e, embora não seja
considerado um dos metais de maior mobilidade no meio ambiente, ainda há
evidências consideráveis mostrando a biodisponibilidade de chumbo associado a
sedimentos para espécies que se alimentam de depósitos (BRYAN & LANGSTON,
1992).
Além disso, o chumbo pode ser acumulado diretamente de águas salgadas e
doces, especialmente em organismos filtradores que utilizam as brânquias como a
principal rota de ingestão de nutrientes. Estudos toxicológicos apontaram efeitos
subletais em peixes, incluindo mudanças na morfologia, no metabolismo e em
atividades enzimáticas. O comportamento de esquiva (comportamento através do
qual alguns animais aquáticos são capazes de evitar áreas com concentrações
anormais de substâncias nocivas) também foi observado em peixes adultos
expostos a níveis que variam de 10 a 100 mg.L-1 (SADIQ, 1992; WHO, 1989).
A legislação brasileira (CONAMA, 1989) admitia um valor máximo permitido
de 0,5 mg de chumbo total por litro de água. Após uma revisão, este valor foi
reduzido para 0,01 mg.L-1, com adequação ao valor proposto pela Organização
Mundial de Saúde (CONAMA 357, 2005).
Um estudo conduzido por Yabe & Oliveira (1998) aponta a presença de
metais em rios de Londrina-PR. No local mais crítico, onde eram descartados os
efluentes da fábrica de baterias, encontrou-se um valor máximo de 4,5 mg de
chumbo por litro de água.

9
A exposição ao chumbo é perigosa uma vez que se trata de um poluente
acumulável, e, mesmo em concentrações relativamente baixas, o chumbo está
associado a alterações no desempenho das enzimas, transferência de energia e
outros processos bioquímicos (RAND et al., 1995).
A presença desse poluente no ambiente aquático pode causar letalidade em
massa nas populações aquáticas ou causar diversos efeitos em todos os níveis da
organização biológica, desde o subcelular e orgânico até níveis de comunidade e
ecossistema. Isso porque as concentrações subletais podem causar alterações no
organismo que comprometem a estrutura dos tecidos, o comportamento, o
crescimento, o desenvolvimento e a reprodução, e isso pode comprometer também
as comunidades e ecossistema aquático (JOBLING, 1995; RAND et al., 1995).
1.5 Prochilodus lineatus
O curimba (Prochilodus lineatus), um peixe de região neotropical, pertencente
à ordem Characiformes, família Prochilodontidae é considerado uma das espécies
mais abundantes e importantes da região Sul e Sudeste do Brasil, sendo bastante
utilizado na alimentação humana. São encontrados ainda na Bacia Amazônica,
Araguaia-Tocantins, Prata, São Francisco, açudes do Nordeste, onde foram
introduzidos.
São peixes de escamas, e a principal característica da família é a boca
protrátil em forma de ventosa, com lábios carnosos, sobre os quais estão
implantados numerosos dentes diminutos dispostos em fileiras. As escamas são
ásperas e a coloração é prateada. Pode alcançar de 30 a 80 cm de comprimento
total. Realizam longas migrações reprodutivas. São capturados em grandes

10
cardumes, sendo espécies importantes comercialmente, principalmente para as
populações de baixa renda.
Este peixe demonstra elevada tolerância a variações de temperatura e de pH,
apresentando um ótimo potencial para a piscicultura. Entretanto, devido ao hábito
alimentar detritívoro a chance de serem contaminados por metais que se acumulam
nos sedimentos é grande (BARRINUEVO e FERNANDES, 1995; MARTINEZ et al.,
2006; TAKASUSUKI et al., 2004).
Essa espécie é muito sensível, não sobrevive em locais poluídos e por isso
pode ser considerada como espécie bioindicadora da qualidade do ambiente.
Possui, ainda, uma biologia bastante conhecida, sendo utilizada em muitos trabalhos
da área.
Figura 1: Exemplar jovem de Prochilodus lineatus
1.6 Biomarcadores
A utilização de alguns parâmetros fisiológicos e morfológicos pode ser uma
ferramenta bastante útil para a avaliação dos efeitos da contaminação ambiental nas
populações de peixes. Vários são os parâmetros biológicos que podem ser alterados
como conseqüência da interação entre o agente químico e o organismo; entretanto,
a determinação quantitativa desses parâmetros pode ser usada como biomarcador
somente se existir uma correlação com a intensidade da exposição e/ou com o efeito
1 cm

11
biológico da substância. Estes biomarcadores são excelentes ferramentas para
monitorar a saúde do ecossistema aquático e têm sido incluídos em vários
programas modernos de monitoramento ambiental de países desenvolvidos. O uso
de biomarcadores bioquímicos em programas de monitoramento oferece algumas
vantagens porque são normalmente os primeiros a serem detectados, são bastante
sensíveis à presença de determinados poluentes, apresentam alta especificidade,
possuem baixo custo de análise e fornecem informações a respeito do efeito
metabólico causado pelo xenobiótico (AMORIM, 2003; STEGEMAN et al., 1992;
WALKER et al., 1996).
Os aspectos comuns entre organismos diferentes se acentuam principalmente
ao nível molecular e, por isso, biomarcadores moleculares possuem a vantagem de
poderem ser aplicados a uma ampla variedade de organismos vivos. Estes tipos de
biomarcadores são de grande importância como sinalizadores iniciais da exposição
aos contaminantes e de seus efeitos, sobretudo antes que as populações e
comunidades, e até mesmo todo o ecossistema, sejam afetados ou encontrem-se
comprometidos (LAMA & GRAY, 2003).
Quando um xenobiótico entra no organismo, no caso dos peixes de água
doce, mais comumente pelas brânquias, este é transportado via sanguínea até os
órgãos, onde são armazenados ou metabolizados para sua eliminação. Como a
maioria dos xenobióticos tem propriedade lipofílica, possuem grande capacidade de
atravessar as membranas biológicas, podendo causar danos aos componentes
celulares. O organismo tem duas maneiras principais de eliminar um composto
xenobiótico, ou pode excretá-lo em sua forma original ou pode ocorrer a
biotransformação, que geralmente leva à formação de compostos mais hidrofílicos

12
que são mais facilmente eliminados pelo organismo (JOBLING, 1995; MARTINEZ,
2006; VAN DER OOST et al., 2003).
O fígado é considerado um órgão-alvo e de grande importância para os
peixes, pois participa dos processos de biotransformação e eliminação de
xenobióticos e, por isso, pode ser utilizado em biomonitoramentos devido à sua alta
sensibilidade a poluentes (HINTON et al., 1992; MYERS et al., 1998; THOPHON et
al., 2003).
O processo de metabolização de xenobióticos é uma importante fonte de
espécies reativas de oxigênio, que correspondem aos produtos da redução do
oxigênio molecular, como o ânion radical superóxido (O2-●), o radical hidroxil (OH●) e
o peróxido de hidrogênio (H2O2). As espécies reativas de oxigênio passam a ter um
efeito prejudicial ao organismo quando ocorre um aumento excessivo na sua
produção ou diminuição de agentes antioxidantes. Em qualquer uma dessas
situações predomina EROS no organismo, o que é denominado estresse oxidativo.
Essa situação pode derivar tanto de fatores internos como de fatores externos
(STEGEMAN et al., 1992).
Para combater essas espécies reativas de oxigênio existe um sistema de
defesa antioxidante, composto por mecanismos enzimáticos e não-enzimáticos. As
principais enzimas antioxidantes são superóxido dismutase (SOD), a catalase e a
glutationa peroxidase (GPx), todas elas abundantes nos tecidos de peixes
(STOREY, 1996; VAN DER OOST et al., 2003).
A catalase descrita em 1901 por Loew (FRUGOLI et al., 1996), é uma enzima
tetramérica que contém grupos heme e é encontrada em todos os organismos vivos.
A CAT é a única entre as enzimas degradantes de H2O2 que não consome
equivalentes redutores celulares e possui um mecanismo muito eficiente para

13
remover o H2O2 formado nas células sob condições de estresse (MALLICK &
MOHN, 2000). Ocorrem primariamente nos peroxissomos e a proliferação
peroxissômica é a principal causa para a elevação da atividade catalásica nos
tecidos (HALLIWELL & GUTTERIDGE, 2005).
A glutationa peroxidase (GPx), é a principal peroxidase de peixes, uma
enzima citosólica tetramérica, dependente de selênio, que emprega a glutationa
reduzida (GSH) como co-fator. A GPx catalisa o metabolismo de um grande número
de hidroperóxidos orgânicos (ROOH) e do H202 para água, envolvendo a oxidação
concomitante da GSH para sua forma oxidada (GSSG). A GPx pode estar localizada
no citosol, bem como na matriz mitocondrial, e divide com a catalase a habilidade
para detoxificar o H2O2 (MARTINEZ, 2006).
A enzima glutationa-S-transferase (GST) desempenha um papel importante
na detoxificação e eliminação de compostos eletrofílicos, incluindo agroquímicos.
Sua estimulação envolve reações de conjugação na presença de glutationa. Assim,
animais aquáticos que habitam ambientes poluídos podem estar expostos a
xenobióticos os quais sofrem detoxificação mediada pela glutationa na sua forma
reduzida, catalisada pela enzima GST. Esta enzima de biotransformação tem sido
estudada em trabalhos de campo no monitoramento de poluentes de origem
industrial e agrícola (CHO et al., 1999; FENET et al., 1998; KANTONIEMI et al.,
1996).
A GSH está envolvida em várias funções fisiológicas: mantém os grupos SH
das proteínas no estado reduzido, participa no transporte de aminoácidos e
detoxificação de toxinas, atua enzimaticamente degradando peróxidos endógenos,
forma moléculas bioativas e atua como coenzima em várias reações enzimáticas
(STAMLER & SLIVKA, 1996). A GSH participa ainda na decomposição do H2O2,

14
potencialmente tóxico, que é convertido em H2O em reação catalisada pela GPx, às
custas da glutationa reduzida; a glutationa oxidada resultante é reciclada à forma
reduzida pela glutationa redutase e NADPH (GUL et al., 2000). O NADPH é
regenerado pela via das pentoses fosfato, em reação catalisada pela glicose 6-
fosfato desidrogenase, a qual é particularmente importante nos eritrócitos. Dessa
forma, este processo de reciclagem e a manutenção de níveis adequados de GSH
podem prevenir o dano celular causado pelo estresse oxidativo (STAMLER &
SLIVKA, 1996).
Nos vertebrados é bastante conhecido o fato de que a exposição a metais
induz a síntese nos tecidos do animal, de um tipo particular de proteína, as
metalotioneínas (MTs). As MTs compreendem uma família de proteínas de baixo
peso molecular, ricas em cisteína que tem como propriedades fundamentais a
detoxificação de metais, proteção celular contra a citotoxidade da radiação ionizante
e captura de radicais livres. Estas proteínas possuem uma elevada afinidade por
íons metálicos dos grupos IB e IIB da tabela periódica, especialmente Zn e Cu. Sua
função principal está relacionada com a regulação das concentrações intracelulares
de metais-traço essenciais. Servem como reserva intracelular destes íons para o
metabolismo de ácidos nucléicos, síntese de proteínas e outros processos
metabólicos (CANLI et al., 1997).
A peroxidação lipídica é um processo fisiológico regular, pois possui um
importante papel na manutenção celular e mobilização de lipídeos (FEUSSNER et
al., 1995 e 2001; MATSUI et al., 1997; SCHEWE et al., 1986; VAN LEYEN et al.,
1998). Porém, determinadas classes de contaminantes podem conduzir a efeitos
deletérios que podem resultar no mau funcionamento celular, destruição da função
das membranas celulares e organelas essenciais tais como o processo de

15
transporte, a manutenção de gradiente de metabólitos e íons e a transdução de
sinais mediada por receptores, com conseqüentes modificações estruturais dos
complexos lipoprotéicos das membranas celulares (BAKER & KRAMER, 1999;
BENZIE, 1996; GIROTTI, 1998; KOZAR et al., 1994; MASON et al., 1997;
MEAGHER & FITZGERALD, 2000; SEVANIAN & URSINI, 2000).
Durante a lipoperoxidação os grupos hidroperóxidos ligam-se aos sítios
hidrofóbicos dos ácidos graxos insaturados, tendo este processo uma dupla
conseqüência: perturbação nas interações lipídicas, que levam às alterações
estruturais das biomembranas e das lipoproteínas; e a formação de espécies
reativas de oxigênio, que podem induzir a modificação secundária de outros
constituintes da membrana (GIROTTI, 2002).
Estas espécies reativas de oxigênio, produzidas em sistemas biológicos, são
detoxificadas por mecanismos antioxidantes do organismo. Estes são encontrados
com freqüência em organismos aquáticos (LIVINGSTONE, 1991; LIVINGSTONE et
al., 1992; AVCI et al., 2005). A peroxidação lipídica apresenta expressivas respostas
frente a danos oxidativos, possuindo também estreita correlação com dados
genéticos.
A acetilcolinesterase (AChE) é um outro biomarcador que pode ser utilizado
para a avaliação da contaminação ambiental. Esta enzima catalisa a clivagem do
neurotransmissor acetilcolina nas sinapses colinérgicas bloqueando a propagação
contínua do impulso nervoso. Vários são os agentes anticolinesterásicos
conhecidos, mas sua inibição é diretamente ligada às ações tóxicas de
organofosforados e carbamatos, praguicidas largamente utilizados em atividades
agrícolas e campanhas de saúde pública. A ação tóxica de alguns compostos inibe a
atividade de acetilcolinesterase (HUGGETT et al., 1992).

16
Os parâmetros bioquímicos como as enzimas de biotransformação, os
antioxidantes enzimáticos e não enzimáticos e os danos oxidativos podem ser
utilizados como biomarcadores em peixes, pois fornecem importantes informações a
respeito da capacidade de defesa destes organismos, bem como a capacidade de
metabolização de compostos tóxicos. Entretanto, é importante que seja feita uma
análise conjunta desses parâmetros para uma melhor interpretação dos resultados.

17
2. OBJETIVOS
● Analisar as possíveis alterações bioquímicas e fisiológicas em Prochilodus
lineatus submetidos a uma concentração subletal de chumbo em testes de
toxicidade aguda.

18
3. ARTIGO
ESTRESSE OXIDATIVO EM Prochilodus lineatus EXPOSTOS AO
CHUMBO
Viléla, M.B.F. de A; Ribeiro, A. M. e Martinez, C.B.R.

19
ESTRESSE OXIDATIVO EM Prochilodus lineatus EXPOSTOS AO CHUMBO
Viléla, M.B.F. de A; Ribeiro, A. M. e Martinez, C.B.R.
Departamento de Ciências Fisiológicas, Universidade Estadual de Londrina. Correspondência para: Mariana Barros F. de A. Viléla, Depto de Ciências Fisiológicas,
Universidade Estadual de Londrina, C.P. 6001, CEP 86051-990, Londrina, PR. e-mail: [email protected]
RESUMO
A exposição de peixes aos contaminantes em ecossistemas aquáticos pode aumentar a formação intracelular de espécies reativas ao oxigênio (EROs). Mas possuem um sistema de defesa antioxidante, responsável por combater essas EROs. As enzimas antioxidantes têm um papel crucial na manutenção da homeostase celular, e sua indução reflete uma resposta específica aos poluentes. O presente estudo investigou os efeitos do chumbo em Prochilodus lineatus. Os animais foram expostos a uma concentração subletal de 0,5 ppm de chumbo dissolvido, durante 6, 24 e 96 horas. Foram determinadas as atividades das enzimas acetilcolinesterase muscular e cerebral, catalase (CAT), glutationa peroxidase (GPx) e glutationa-S-transferase (GST) no fígado, as concentrações de glutationa reduzida (GSH), metalotioneína (MET) e a lipoperoxidação (ensaio FOX). Após 6 horas de exposição ao Pb, a CAT, GSH e a MET aumentaram significativamente, enquanto a GPx, GST e FOX apresentaram tendência ao aumento. Em 24 h os peixes apresentaram decréscimo significativo na atividade da GST e tendência de redução da CAT e GSH, acompanhadas de aumento significativo da GPx e MET, e tendência da FOX. E no tempo de 96 horas, houve decréscimo significativo da GST, a GSH e a FOX apresentaram aumento significativo e a CAT, GPx e MET também tiveram uma tendência a aumentar. A acetilcolinesterase apresentou uma diminuição significativa em todos os tempos experimentais. Esses resultados indicam que o Pb na concentração testada agiu como anti-colinesterásico; promoveu a ativação das defesas antioxidantes enzimáticas e não-enzimáticas; inibição da enzima de conjugação GST, provavelmente devido ao uso da GSH na associação com o Pb; aumentou a peroxidação lipídica, sugerindo que houve danos ao fígado do animal e as defesas antioxidantes não foram suficientes para evitar o estresse oxidativo. Palavras-chave: chumbo, Prochilodus lineatus, estresse oxidativo, fígado.

20
INTRODUÇÃO
A água é indispensável para a sustentação da vida e o que vem ocorrendo é
a degradação da sua qualidade, ressaltando a contaminação química pela
agricultura e também por indústrias. Os efluentes gerados a partir de atividades
industriais, muitas vezes contêm metais pesados que contaminam o ambiente. Estes
contaminantes podem causar efeitos adversos aos organismos e, além disso,
causam prejuízos aos ecossistemas aquáticos, pois tendem a se acumular na biota
aquática (WHO, 1987).
A exposição ao chumbo é perigosa uma vez que se trata de um poluente
acumulável, e mesmo que em doses relativamente baixas, o chumbo está associado
a alterações na atividade das enzimas, transferência de energia e outros processos
bioquímicos (RAND et al., 1995).
Os biomarcadores nos níveis mais baixos da organização biológica, como os
bioquímicos, fisiológicos e histológicos, têm sido considerados como medidas viáveis
de alterações biológicas em virtude da presença de agentes estressores. Esses
biomarcadores podem permitir uma avaliação rápida do estado de saúde do
organismo (HUGGETT et al., 1992).
Nesse contexto, a toxicologia aquática, ciência que estuda os efeitos dos
compostos químicos produzidos pelo homem sobre os organismos aquáticos, pode
ser uma ferramenta bastante útil para avaliação de impactos de poluentes sobre a
biota aquática (LOMBARDI, 2004; RAND et al., 1995).
O curimba (Prochilodus lineatus), um peixe de região neotropical, é
considerado uma das espécies mais abundantes e importantes da região Sul e
Sudeste do Brasil, sendo bastante utilizado na alimentação humana. Este peixe

21
demonstra elevada tolerância a variações de temperatura e de pH, apresentando um
ótimo potencial para a piscicultura. Entretanto, devido ao hábito alimentar detritívoro
a chance de serem contaminados por metais que se acumulam nos sedimentos é
grande (BARRINUEVO & FERNANDES, 1995; MARTINEZ et al., 2006;
TAKASUSUKI et al., 2004).
Os efeitos dos contaminantes em peixes podem se manifestar em vários
níveis de organização biológica, incluindo disfunções fisiológicas, alterações
estruturais em órgãos e tecidos e modificações comportamentais que levam ao
prejuízo do crescimento e reprodução (ADAMS, 1990). Os aspectos comuns entre
organismos diferentes se acentuam principalmente ao nível molecular e, por isso,
biomarcadores moleculares possuem a vantagem de poderem ser aplicados a uma
ampla variedade de organismos vivos. Estes tipos de biomarcadores são de grande
importância como sinalizadores iniciais da exposição aos contaminantes e de seus
efeitos (LAMA & GRAY, 2003).
A absorção de metais, pose se dar diretamente através da água ou
indiretamente através da cadeia alimentar (BENTLEY, 1992; HAMILTON et al.,
1998). Nos peixes, as brânquias são alvos de metais, mas estes também podem ser
absorvidos pelo trato gastrointestinal e pele (WATANABE et al., 1997; SCHLENK &
BENSON, 2001).
O fígado é considerado um órgão-alvo e de grande importância para os
peixes, pois participa dos processos de biotransformação e excreção de
xenobióticos e por isso pode ser utilizado em biomonitoramentos devido a sua alta
sensibilidade a poluentes. Alterações em sua estrutura podem ser significativas na
avaliação da saúde de peixes e refletem os efeitos da exposição a uma variedade de

22
poluentes ambientais (HINTON et al., 1992; MYERS et al., 1998; THOPHON et al.,
2003).
Assim, considerando-se as ocorrências de contaminação aquática por
chumbo, como por exemplo, o estudo conduzido por Yabe & Oliveira (1998) em que
encontraram concentrações de até 4,5 mg.L-1 em Londrina-PR, onde eram
descartados os efluentes da fábrica de baterias, e por conseqüência a exposição
dos peixes ao xenobiótico, torna-se necessário a realização de testes de toxicidade
para avaliação do efeito toxicológico desse metal. Portanto, esse trabalho visou
avaliar as possíveis alterações bioquímicas e fisiológicas de Prochilodus lineatus,
após exposição aguda a concentração subletal de chumbo.
MATERIAIS E MÉTODOS
Organismo-teste
Os experimentos foram realizados com exemplares jovens de Prochilodus
lineatus (VALENCIENNES, 1847), com massa corpórea de 9,26 ± 3,22 g e
comprimento total de 9,71 ± 1,10 cm (média ± DP), fornecidos pela Estação de
Piscicultura da Universidade Estadual de Londrina (EPUEL). No laboratório os
animais foram aclimatados em tanques de 300 litros durante uma semana, com água
desclorada e aeração constante, alimentados em intervalos de 48 horas com ração
comercial peletizada, sendo esta suspensa 24h antes do início da amostragem.
Parâmetros como temperatura, pH, condutividade, oxigênio e fotoperíodo de 12/12
claro/escuro foram observados durante todo o período de aclimatação e exposição.

23
Testes de toxicidade
Depois da aclimatação os peixes foram submetidos a testes de toxicidade
estáticos agudos de 6, 24 e 96h. Os animais foram transferidos para aquários de
100 litros e divididos em dois grupos, o controle, com água limpa, e o experimental,
que recebe uma concentração de 4 gramas de nitrato de chumbo, o que
corresponde a 0,5 mg.L-1 de chumbo dissolvido. Cada aquário apresentou uma
média de 7 animais, obedecendo a proporção de 1 grama de peixe por litro de água.
Após os experimentos amostras de água foram coletadas para a
quantificação da concentração de chumbo, por espectrofotometria de absorção
atômica. Amostras de água filtrada (filtro com poro de 45 µm) foram utilizadas para a
determinação da concentração de chumbo dissolvido, que corresponde à fração do
metal que está livre na água, disponível para o peixe. Amostras não filtradas foram
utilizadas para a análise da concentração de chumbo total, que corresponde à
concentração total do metal, incluindo o chumbo que está complexado com outros
compostos.
Amostragem
Após a retirada dos peixes dos aquários, foram anestesiados imediatamente
com benzocaína (0,1 g.L-1). Na amostragem os animais foram medidos
(comprimento total e padrão) e pesados, e então mortos por secção medular para
retirada do cérebro, músculo e fígado, e as amostras congeladas a - 80 C até o
momento do ensaio.
Análise de parâmetros de estresse oxidativo

24
As amostras de fígado congeladas a -80ºC foram utilizadas para
determinação das atividades das enzimas catalase (CAT), glutationa peroxidase
(GPx) e glutationa-S-transferase (GST), para a determinação das concentrações de
glutationa reduzida (GSH) e metalotioneína e para a quantificação da
lipoperoxidação (ensaio FOX). Os órgãos foram pesados e homogeneizados com
mini-homogeneizador TE-103 em 4000 rpm por cerca de 1 minuto, em tampão
fosfato 0,1 M e pH 7,0 (10 vezes o volume), e então centrifugados, durante 20
minutos a 4°C e 12.000 g. O sobrenadante foi separado para as análises
enzimáticas.
A atividade da catalase foi determinada de acordo com a técnica descrita por
Beutler (1975), seguindo-se a velocidade de decomposição do peróxido de
hidrogênio, sendo fornecido um tampão para catalase com pH 8,0 e o meio de
reação. A leitura da absorbância foi feita em 240 nm, e a atividade foi expressa em
µmol.min-1.mg de proteína hepática-1.
A atividade da glutationa-S-transferase (GST) foi determinada de acordo com
a metodologia descrita por Keen et al., (1976), seguindo-se a complexação da
glutationa reduzida (GSH) com o 1-cloro-2,4-dinitrobenzeno (CNDB), em 340 nm e
expressa em nmol.min-1.mg de proteína hepática-1.
A GPx foi determinada de acordo com a técnica de Nakamura (1984), através
da oxidação do NADPH + H+ em presença de peróxido, em 340 nm. Ou seja, a GPx
dismuta o H2O2 do ensaio, gerando para isso uma ponte de dissulfeto entre duas
GSH (GS-GS), que por sua vez volta ao estado reduzido (2 GSH), pela ação da
glutationa redutase (GR). A GR age mediante a oxidação de NADPH. Assim, o
ensaio é uma medida que consiste em registrar a diminuição de NADPH. Expressa
em nmol.min-1.mg de proteína hepática-1.

25
A lipoperoxidação foi determinada utilizando-se o ensaio FOX (JIANG et al.,
1991), que corresponde à reação química de auto oxidação de lipídios (LH) que
conduz a lipoperoxidação (LOOH). O ensaio FOX está baseado na oxidação do Fe
(II) por LOOH em pH ácido na presença de um pigmento complexador de Fe (III), o
xilenol laranja. A formação deste complexo pode ser quantificada pelo aumento da
absorção em 560 nm e expressa em µM CHP (hidroperóxido de cumeno) por mg de
proteína hepática.
O princípio para medida da GSH corresponde à reação da glutationa com o
DNTB, formando um tiolato (TNB) de cor amarelada, quanto maior a presença de
GSH, mais acentuada a cor. Leitura em 412 nm e expressa em nmol.mg de proteína
hepática-1.
A concentração das metalotioneínas foi estimada através da concentração de
grupamentos sulfidrilas (VIARENGO et al., 1997). O fígado foi homogeneizado em
tampão TRIS-HCL 20 mM com pH 8,6, contendo sacarose 0,5M e ß-mercaptoetanol
0,01%, e centrifugado a 12000 g por 20 minutos a 4º C. A concentração de
metalotioneína foi quantificada espectrofotometricamente utilizando-se o reagente de
Ellman (NaCl 2M, DTNB 0,43M em tampão fosfato 0,2M e pH 8,0) em 412 nm e
expressa em µM GSH de proteína hepática-1.
A concentração de proteínas do sobrenadante foi determinada de acordo com
o método descrito por Lowry et al. (1951), para expressar a atividade das enzimas. A
leitura foi feita em 700 nm, utilizando-se como padrão protéico albumina sérica
bovina.
Análise da atividade de acetilcolinesterase muscular e cerebral

26
As amostras de músculo e cérebro foram homogeneizadas em tampão fosfato
com pH 7,5 e centrifugadas a 10.000 g a 4ºC, durante 20 minutos, para a
determinação da atividade da acetilcolinesterase e de proteína cerebral e muscular.
A atividade da acetilcolinesterase foi determinada de acordo com a técnica de Silva
de Assis (1998), modificada de Ellman et al., (1961), a partir da reação de iodeto de
acetilcolina com o reagente de cor ditionitrobenzoato (DTNB), em leitora de
microplacas a 415 nm, e expressa em nmol.min-1.mg de proteína-1. A concentração
da proteína foi determinada de acordo com Lowry et al. (1951).
Análises estatísticas
Os grupos experimentais foram comparados com seus respectivos controles
empregando-se testes estatísticos apropriados. Foram considerados significativos
valores de P ≤ 0,05.
RESULTADOS
Análise da água
Os parâmetros físico-químicos da água se mantiveram estáveis durante os
experimentos. Os valores médios (média ± DP) foram: Temperatura 23,02 ± 2,1 ºC;
pH 7,41 ± 0,22; OD 7,05 ± 0,61 mg O2.L-1; condutividade 63,55 ± 7,3 µS.cm-1. Nos
aquários experimentais, a concentração média de chumbo total foi de 2,25 mg.L-1, e
a de chumbo dissolvido, 0,5 mg.L-1. Enquanto que nos controles foi menor que 0,1
mg.L-1.

27
Parâmetros de estresse oxidativo
Os exemplares de Prochilodus lineatus expostos à concentração de 0,5 ppm
de chumbo dissolvido apresentaram aumento significativo na atividade hepática da
enzima catalase no tempo experimental de 6h, quando comparado com o grupo
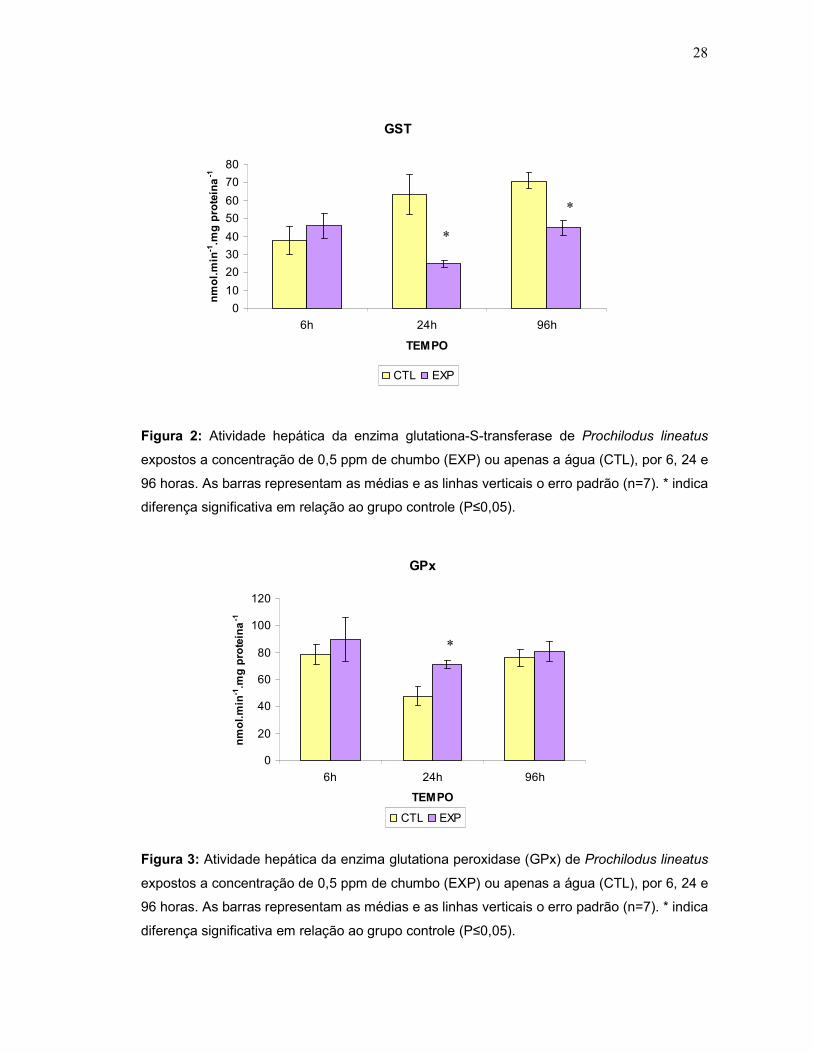
controle (Fig.1). Já a glutationa-S-transferase (GST) apresentou uma diminuição
significativa nos tempos de 24 e 96h (Fig.2) enquanto a glutationa peroxidase (GPx)
aumentou significativamente em 24h (Fig.3). Dentre os antioxidantes não
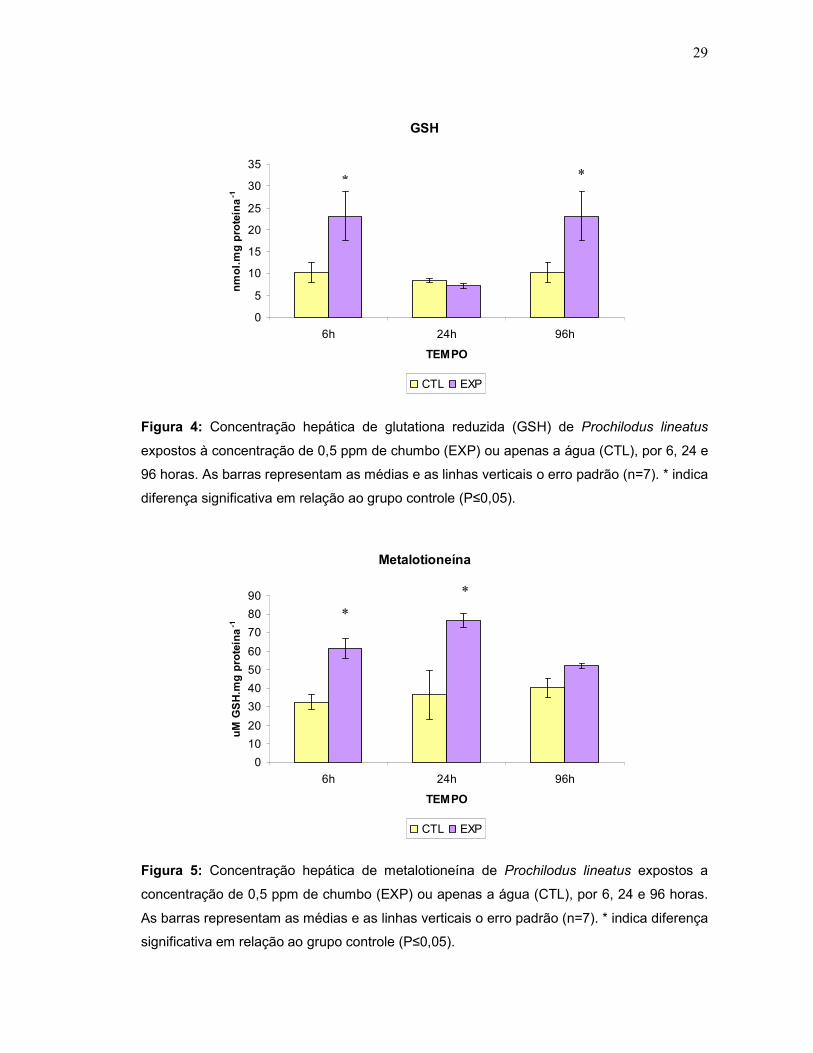
enzimáticos, a concentração hepática de glutationa reduzida (GSH) aumentou
significativamente após 6 e 96h (Fig. 4) e a de metalotioneína hepática apresentou
um aumento significativo após 6 e 24h de exposição ao metal. (Fig.5).
CAT
0
10
20
30
40
50
60
70
80
6h 24h 96h
TEMPO
nmol.min
-1.m
g proteína-1
CTL EXP
Figura 1: Atividade hepática da enzima catalase (CAT) de Prochilodus lineatus expostos a
concentração de 0,5 ppm de chumbo (EXP) ou apenas a água (CTL), por 6, 24 e 96 horas.
As barras representam as médias e as linhas verticais o erro padrão (n=7). * indica diferença
significativa em relação ao grupo controle (P≤0,05).
*
28
Figura 2: Atividade hepática da enzima glutationa-S-transferase de Prochilodus lineatus
expostos a concentração de 0,5 ppm de chumbo (EXP) ou apenas a água (CTL), por 6, 24 e
96 horas. As barras representam as médias e as linhas verticais o erro padrão (n=7). * indica
diferença significativa em relação ao grupo controle (P≤0,05).
GPx
0
20
40
60
80
100
120
6h 24h 96h
TEMPO
nmol.min
-1.m
g proteína-1
CTL EXP
Figura 3: Atividade hepática da enzima glutationa peroxidase (GPx) de Prochilodus lineatus
expostos a concentração de 0,5 ppm de chumbo (EXP) ou apenas a água (CTL), por 6, 24 e
96 horas. As barras representam as médias e as linhas verticais o erro padrão (n=7). * indica
diferença significativa em relação ao grupo controle (P≤0,05).
GST
0
10
20
30
40
50
60
70
80
6h 24h 96h
TEMPO
nmol.min
-1.m
g proteína-1
CTL EXP
*
*
*
29
GSH
0
5
10
15
20
25
30
35
6h 24h 96h
TEMPO
nmol.mg proteína-1
CTL EXP
Figura 4: Concentração hepática de glutationa reduzida (GSH) de Prochilodus lineatus
expostos à concentração de 0,5 ppm de chumbo (EXP) ou apenas a água (CTL), por 6, 24 e
96 horas. As barras representam as médias e as linhas verticais o erro padrão (n=7). * indica
diferença significativa em relação ao grupo controle (P≤0,05).
Metalotioneína
0
10
20
30
40
50
60
70
80
90
6h 24h 96h
TEMPO
uM GSH.m
g proteína-1
CTL EXP
Figura 5: Concentração hepática de metalotioneína de Prochilodus lineatus expostos a
concentração de 0,5 ppm de chumbo (EXP) ou apenas a água (CTL), por 6, 24 e 96 horas.
As barras representam as médias e as linhas verticais o erro padrão (n=7). * indica diferença
significativa em relação ao grupo controle (P≤0,05).
* *
*
*

30
Lipoperoxidação
A concentração de lipoperóxidos no fígado de P. lineatus expostos ao
chumbo, estimada pelo ensaio FOX e expressa pela concentração de hidroperóxido
de cumeno (CHP), apresentou aumento significativo após 96 h de exposição.
FOX
0
20
40
60
80
100
120
140
6h 24h 96h
TEMPO
uM CHP/m
g proteína-1
CTL EXP
Figura 6: Concentração de lipoperóxidos no fígado de Prochilodus lineatus expostos à
concentração de 0,5 ppm de chumbo (EXP) ou apenas a água (CTL), por 6, 24 e 96 horas.
As barras representam as médias e as linhas verticais o erro padrão (n=7). * indica diferença
significativa em relação ao grupo controle (P≤0,05).
Atividade da enzima acetilcolinesterase em músculo e cérebro
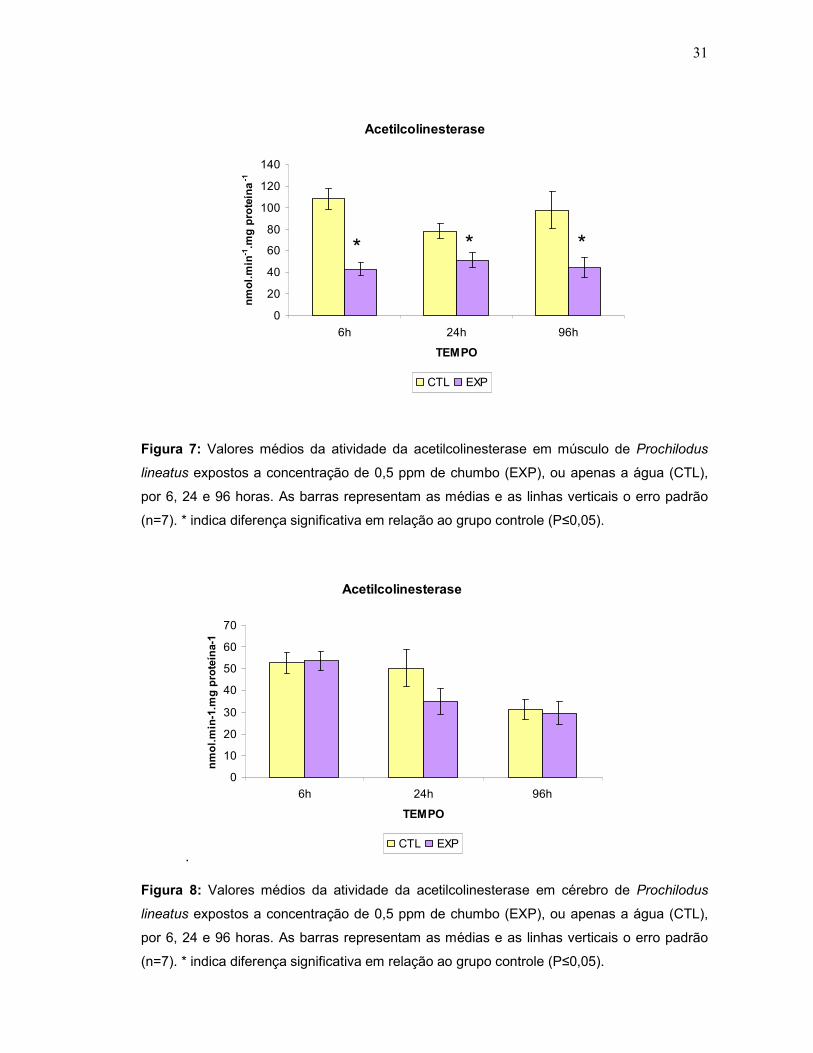
A atividade da acetilcolinesterase muscular foi significativamente menor nos
curimbas expostos ao chumbo, em todos os tempos experimentais (6, 24 e 96 h)
quando comparados aos seus respectivos grupos controles (Fig.7). Entretanto, para
atividade da acetilcolinesterase cerebral não foram detectadas diferenças entre os
animais do grupo controle e aqueles expostos ao chumbo (Fig.8).
*
31
Acetilcolinesterase
0
20
40
60
80
100
120
140
6h 24h 96h
TEMPO
nmol.min
-1.m
g proteína-1
CTL EXP
Figura 7: Valores médios da atividade da acetilcolinesterase em músculo de Prochilodus
lineatus expostos a concentração de 0,5 ppm de chumbo (EXP), ou apenas a água (CTL),
por 6, 24 e 96 horas. As barras representam as médias e as linhas verticais o erro padrão
(n=7). * indica diferença significativa em relação ao grupo controle (P≤0,05).
.
Acetilcolinesterase
0
10
20
30
40
50
60
70
6h 24h 96h
TEMPO
nmol.min-1.m
g proteína-1
CTL EXP
Figura 8: Valores médios da atividade da acetilcolinesterase em cérebro de Prochilodus
lineatus expostos a concentração de 0,5 ppm de chumbo (EXP), ou apenas a água (CTL),
por 6, 24 e 96 horas. As barras representam as médias e as linhas verticais o erro padrão
(n=7). * indica diferença significativa em relação ao grupo controle (P≤0,05).
* * *

32
DISCUSSÃO
As avaliações bioquímicas são ferramentas utilizadas para se verificar o
estado geral de um organismo (MCGEER et al., 2000; ROMÉO et al., 2000). No
presente estudo os parâmetros avaliados foram as atividades das enzimas
acetilcolinesterase, catalase (CAT), glutationa peroxidase (GPx) e glutationa-S-
transferase (GST), as concentrações de glutationa reduzida (GSH), metalotioneína e
de lipoperóxidos (ensaio FOX). Os parâmetros analisados mostraram-se de uma
maneira geral bastante sensíveis a presença do chumbo na água.
A catalase é uma hemeproteína que fica nos peroxissomos e catalisa a
redução do H2O2 a H2O e O2. É encontrada no sangue, medula óssea, mucosas, rim
e fígado. A suplementação de catalase exógena previne a oxidação da GSH
mediada pelo H2O2, em eritrócitos humanos normais, e também inibe as lesões
oxidativas do DNA (MAYES, 1990; SCOTT et al., 1991). Nesse trabalho a catalase
apresentou um aumento significativo em 6 horas, o que supõe o esforço do animal
para combater o peróxido de hidrogênio produzido em função da presença do
chumbo.
O aumento da atividade específica da CAT tem sido relatado como indicador
de estresse oxidativo. O estresse oxidativo encontrado é resultado da tentativa da
célula em eliminar espécies reativas de oxigênio, formadas na metabolização de
diversos compostos químicos, através de enzimas específicas tais como a catalase
e enzimas desidrogenases como as glutationas. A indução da catalase sugere uma
defesa do organismo frente a agentes oxidantes, pois é uma enzima chave para
remover peróxido de hidrogênio e evitar a formação de radicais hidroxil que podem

33
causar danos celulares. (GIULIO et al., 1989; REGOLI et al., 2000; STURVE et al.,
2005, STURVE et al., 2006; VAN DER OOST et al., 2003).
Aumento na atividade da CAT também foi evidenciado no mexilhão de água
doce (Dreissena polymorpha) coletados no Rio Sena, e no mexilhão marinho
(Mytilus edulis) na Baía Sena, França, em locais contaminados por metais Pb, Cd,
Hg e Ni. Outros estudos também já demonstraram o aumento da atividade da CAT
decorrente da exposição à metais (GRAVATO et al., 2006; HANSEN et al., 2006; LIU
& KUEH, 2005; ROCHER et al., 2006).
Além da CAT, a GPx também catalisa a redução do peróxido de hidrogênio
(H2O2), como também de peróxidos orgânicos para seus correspondentes alcoóis às
custas da conversão da GSH a GSSG. A GPx tem ação fundamentalmente
citosólica. O aumento significativo dessa enzima no fígado dos peixes após 24 h de
exposição ao chumbo deve estar relacionado com o combate ao peróxido de
hidrogênio, já que a catalase neste tempo não esteve aumentada, e também a
possível metabolização de peróxidos orgânicos (HEBELL, 1986; SHAN et al., 1990).
A GSH está presente na maioria das células e é o tiol mais abundante no
meio intracelular. Sua capacidade redutora é determinada pelo grupamento -SH,
presente na cisteína. A GSH pode ser considerada um dos agentes mais
importantes do sistema de defesa antioxidante da célula, protegendo-a contra lesões
resultantes da exposição a agentes oxidantes como íons ferro, oxigênio hiperbárico,
radiação e luz ultravioleta. Além disto, participa da detoxificação de agentes
químicos e da eliminação de produtos da lipoperoxidação e é requerida para a
síntese de DNA, de proteínas e de algumas prostaglandinas. No presente trabalho, o
aumento significativo da GSH nos tempos experimentais de 6 e 96 horas indicam o
aumento da proteção do organismo contra o estresse oxidativo (DENEKE et al.,

34
1989; GALLEANO & PUNTARULO, 1994; MEISTER et al., 1983). Além disto, após a
entrada de metais nas células esses tendem a ser complexados com compostos de
baixo peso molecular, como o tripeptídeo glutationa (GSH), de modo a inibir ou
reduzir suas ações deletérias (GIULIO et al., 1989).
A GST é considerada uma importante enzima de desintoxicação, por
metabolizar grande variedade de compostos xenobióticos orgânicos, por meio da
conjugação destes com a glutationa reduzida (GSH), formando substâncias de baixa
toxicidade. No presente trabalho, o chumbo promoveu inibição da enzima desta
enzima de conjugação após 24 e 96 horas de exposição, provavelmente devido ao
uso da GSH na associação com o chumbo, quando o metal entra na célula do
animal.
A indução de metalotioneínas em organismos aquáticos após exposição a
metais pesados é um fato bem documentado (CANLI et al., 1997). E comprovado
com este trabalho, onde ocorreu um aumento significativo nos tempos de 6 e 24
horas. Após a entrada do chumbo na célula hepática o metal deve ter se
complexado com a metalotionéina existente, e ainda estimulado a transcrição de
genes específicos para a produção de mais metalotioneína.
A enzima acetilcolinesterase tem se mostrado sensível a alguns tipos de
contaminantes, como praguicidas, metais, produtos originados da combustão de
hidrocarbonetos e detergentes, que contem amônia terciária e quaternária, entre
outros. Neste trabalho verificou-se que curimbas expostos ao chumbo apresentaram
diminuição significativa na atividade muscular da acetilcolinesterase em todos os
tempos experimentais, indicando a ação inibitória da acetilcolinesterase pelo metal.
A inibição da AChE por poluentes tem sido amplamente estudada e já foi
considerada um biomarcador específico para carbamatos e organofosforados (OP),

35
porém outros estudos tem indicado que outras substâncias como metais pesados,
organoclorados e PAHs também podem vir a inibí-la, mas a concentração
necessária para promover este efeito é relativamente mais alta. Essa inibição
causada pelo anticolinesterásicos pode ser reversível ou irreversível. Compostos
organofosforados, por exemplo, formam um complexo inibidor enzimático muito
estável e que não sofre dissociação espontânea significativa, e desta forma, a
enzima fica permanentemente fosforilada e a recuperação de sua atividade depende
da síntese de nova enzima (AKAISHI et al., 2004; GOODMAN & GILMAN, 1996;
MARTINEZ-TABCHE et al., 1997; PAYNE et al., 1996; RANGE & DALE, 2000;
STURM et al., 1999).
A AChE é muito importante à manutenção e equilíbrio da transmissão de
estímulos nervosos, pois catalisa a hidrólise do neurotransmissor acetilcolina, que é
transformado em colina e acetato que reage com uma molécula de água e produz
ácido acético, e por fim a enzima é reativada e a fração colina é prontamente
reabsorvida por um processo de transporte sódio-dependente. Quando a atividade
da AChE é inibida ocorre um bloqueio na transmissão dos impulsos nervosos, o que
pode paralisar algumas funções vitais do organismo (ADAMS, 1990; BEYERS &
SIKOSKI, 1994; HABIG & DI GIULIO, 1991; KLEMZ, 2002; STENESH, 1998).
Estudos indicam que a relação entre a inibição da acetilcolinesterase cerebral e
muscular é específica para cada espécie, podendo o animal apresentar igual, maior
ou menor sensibilidade entre estes tecidos (FULTON & KEY, 2001).
Além disto, a inibição da atividade da acetilcolinesterase no músculo pode
estar associada ao aumento da produção de espécies reativas de oxigênio no
organismo, em função de um aumento da demanda energética, e, portanto aumento
do metabolismo energético. Isto se justifica pelo fato de que, com a inibição da

36
acetilcolinesterase, o animal gasta mais energia devido a manifestações como
hiperestesia, espasmos intermitentes e tremores musculares e o aumento da
demanda de oxigênio, em função do gasto energético devido à ação colinérgica,
pode levar ao estresse oxidativo.
Considerando-se de forma conjunta todos os resultados obtidos neste
trabalho, pode-se propor um modelo ainda bastante preliminar para a ação do
chumbo em células hepáticas de Prochilodus lineatus (Fig.9): após entrar nas
células do organismo, o chumbo associou-se a moléculas de baixo peso molecular,
como as proteínas metalotioneínas, ou o tripeptídeo GSH, que tornam o metal
inativo. O Pb também deve ter induzido o aumento da produção da MET para
combater os efeitos desse metal. Porém, a quantidade do metal foi maior do que a
proteção dada pela MET e GSH, e o deve ter interferido na cadeia respiratória da
mitocôndria, levando ao aumento da produção do ânion superóxido (O2●), que é
muito tóxico para a célula e pode se transformar em radical hidroxil (HO●), que é
ainda mais nocivo. O peróxido de hidrogênio, resultante da ação da SOD, foi
quebrado em água e oxigênio através da ação das enzimas catalase e GPx, que
apresentaram aumento significativo no fígado dos peixes expostos ao chumbo. A
diminuição significativa da GST deve estar relacionada com a utilização da GSH
para a associação inicial com o metal. A inibição da acetilcolinesterase muscular
também deve ter contribuído para a geração de mais EROs. Entretanto, apesar da
ativação das defesas antioxidantes do animal, houve o aumento da lipoperoxidação
no fígado. Isto demonstra que nos animais expostos à concentração de chumbo
testada (0,5 mg de chumbo dissolvido por litro de água) as defesas antioxidantes
foram insuficientes para prevenir a ocorrência de lesões oxidativas, resultando no
estresse oxidativo.

37
Figura 9: Modelo explicativo sobre a ação do chumbo nas células hepáticas de Prochilodus
lineatus. Ver texto para explicações.
CONCLUSÕES
• O chumbo na concentração testada inibiu a acetilcolinesterase muscular o
que pode ter contribuído para o aumento da geração de EROs no organismo.
• Promoveu a ativação das defesas antioxidantes enzimáticas (CAT e GPx) e
não-enzimáticas (GSH e MT);
• Promoveu a inibição da enzima de conjugação GST, provavelmente
associada ao uso da GSH na associação com o Pb;
• Aumentou a peroxidação lipídica no fígado, indicando a ocorrência de danos
oxidativos provavelmente porque as defesas antioxidantes não foram
suficientes para evitar os efeitos das EROS.
Pb
MT +
O2- H2O2 +
SOD
H2O + O2
CAT
GPx
2GSH
GSSH
GST
AChE
+
HO•
-
-
PUFA
Peroxidação lipídica
Pb

38
REFERÊNCIAS BIBLIOGRÁFICAS
ADAMS, S. M. Status and use of biological indicators for evaluating the effect of stress in fish. Amer. Fish. Soc. Symp. 8: 1-8. 1990.
AKAISHI, F.M.; SILVA DE ASSIS, H.C.; JAKOBI, S.C.G.; STJEAN, S.; COUTERNAY, S.C.; LIMA, E.;WAGNER, A.L.R.; SCOFIELD, A.; OLIVEIRA RIBEIRO, C.A. Morphological and neurotoxicological findings in tropical freshwater fish (Astyanax sp.) after waterborne and acute exposure to water soluble fraction (WSF) of crude oil. Arch. Environ. Contam. Toxicol. 46: 244–253. 2004.
BARRINUEVO, W.R.; FERNANDES, M.N. Critical thermal maxima and ninima for curimbatá, Prochilodus scrofa (Steindachner) of two different sizes. Aqua. Res. 26: 447-450, 1995.
BENTLEY, P.J. Influx of zinc bt channel catfish (Ictalurus ounctatus):uptake from external environmental solutions. Com. Biochem. Phys. 1C: 215-217, 1992.
BEUTLER, E. Red cell metabolism: A manual of biochemical a methods. Grune & Stron, New York, 1975.
BEYERS, D.W.; SIKOSKI, P.J. Acethylcholinesterase inhibition in federally endangered Colorado squawfish exposed to cararyl and malathion. Environ. Toxicol. Chem. 13:935–939. 1994.
CANLI, M.; STAGG, R.M.; RODGER, E.G. The induction of metallothionein in tissues of the Norway lobster Nephrops norvegicus following exposure to cadmium, copper and zinc: the relationships between metallothionein and the metals. Environ. Poll. 96: 343-350, 1997.
DENEKE, S.M.; FANBURG, B.L. Regulation of cellular glutathione. Am. J. Physiol. 257: L163-73, 1989.
ELLMAN, G.L.; COUTNEY, K.; ANDRES, V.; FEATHERSTONE, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharm. 7: 88-95, 1961.
FULTON, M.H.; KEY, P.B. Acetylcholinesterase inhibition in estuarine fish and invertebrates as an indicator of organophosphorus inseticide exposure and effects. Environ. Toxicol. Chem. 20 (1): 37-75, 2001.
GALLEANO, M.; PUNTARULO, S. Effect of mild iron overload on liver and kidney lipid peroxidation. Braz. J. Med. Biol. Res. 27: 2.349-2.358, 1994.
GIULIO, R.T.D.; WASHBURN, P.C.; WENNING, R.J.; WINSTON, G.W.; JEWELL, C.S. Biochemical responses in aquactic animals: a review of determinants of oxidative stress. Environ. Toxicol. Chem. 9: 1103-1123, 1989.
GOODMAN, L.S.; GILMAN, A.G. As Bases Farmacológicas da Terapêutica. 9º Ed. Rio de Janeiro. McGraw Hill Interamericana Editores. 1435p, 1996.

39
GRAVATO, C.; TELES, M.; OLIVEIR, M.; SANTOS, M.A. Oxidative stress, liver biotransformation and genotoxic effects induced by copper in Anguilla anguilla L. – the influence of pre-exposure to b-naphthoflavone. Chem. in press, 2006.
HABIG, C.; DI GIULIO, R.T. Biochemical characteristics of cholinesterase in aquatic organisms. In: Mineau P (ed), Cholinesterase inhibition insecticides: their impact on wildlife in the environment. Elsevier, New York, 19–33, 1991.
HAMILTON, D.P.; MALIK, D.S.; SASTRY, K.V. Effects of zinc toxicity on biochemical composition of muscle and liver of murrel (Channa puntactus). Environ. Intern. 24(4): 433-438, 1998.
HANSEN, B.H.; ROMMA, S.; GARMO, O.A.; OLSVIK, P.A.; ANDERSEN, R.A. Antioxidative stress proteins and their gene expression in brown trout (Salmo trutta) from three rivers with different heavy metal levels. Comp. Biochem. Physiol. Part C: Toxicol Pharmacol 143: 263-274. 2006.
HEBBEL, R.P. Erythrocyte antioxidants and membrane vulnerability. J. Lab. Clin. Med. 107: 401-4, 1986.
HINTON, D.E.; BAUMANN, P.C.; GARDNER, G.R.; HAWKINS, W.E.; HENDRICKS, J.D.; MURCHELANO, R.A.; OKIHIRO, M.S. Histopathologic Biomarkers. In: Hugget, R.J.; Kimerle, R.A.; Mehrle, J.R.; Bergman, H.L. Biomarkers: Biochemical, Physiological and Histological Markers of Anthropogenic Stress. Lewis Pubishers, Boca Raton, 1992.
HUGGET, R.J.; KIMERLE, R.A.; MEHRLE JR, P.M.; BERGMAN, H.L. Biomarkers biochemical, physiological and histological markers os anthropogenic stress. Lewis Publishers, Boca Raton, 347p. 1992.
JIANG, Y.; LEE, A.; CHEN, J.; CADENE, M.; CHAIT, B.T. ; MACKINNON, R. Crystal structure and mechanism of a calcium-gated potassium channel. Nat. 417:515-522, 1991.
KEEN, J.H.; HABIG, W.H.; JAKOBY, W.B. Mechanism for several activities of the glutathione-S-transferase. J. Biol. Chem. 251: 6183-6188, 1976.
KLEMZ, C. Uso de biomarcadores de contaminação ambiental em peixes Ancistrus sp. (cascudo). Dissertação de Mestrado em Farmacologia. Universidade Federal do Paraná – UFPR, Curitiba, 2002.
LAMA, P.K.S.; GRAY, J. S. The use of biomarkers in environmental monitoring programmes. Mar. Pollut. Bull. 46(2): 182-186, 2003.
LIU, J.H.; KUEH, C.S.W. Biomonitoring of heavy metals and trace organics using the intertidal mussel Perna viridis in Hong Kong coastal waters. Mar Pollut Bull 51: 857–875, 2005.
LOMBARDI, J.V. Fundamentos de Toxicologia Aquática. In:RANZANI-PAIVA, M.J.T.; TAKEMOTO, R.M.; LIZAMA, M.A.P. Sanidade de Organismos Aquáticos. Ed. Varela, São Paulo, 2004.

40
LOWRY, O.H.; ROSENBROUGH, N.J.; FARR, A.L.; RANDALL, R.J. Protein measurements with the Folinphenol reagent. J. Biol. Chem. 193: 265-275, 1951.
MARTINEZ, C.B.R.; AZEVEDO, F.; WINKALER, E.U. Toxicidade e efeitos da amônia em peixes neotropicais. In: CYRINO, J.E.P.; URBINATI, E.C. Tópicos Especiais em Biologia Aquática e Aqüicultura, Jaboticabal – S.P: Sociedade Brasileira de Aqüicultura e Biologia Aquática, 81-95, 2006.
MARTINEZ-TABCHE, L.; RAMÍREZ, B.M.; GERMÁN-FAZ, C.; GALAR, C.I.; MADRIGAL, O.M.; ULLOA, G.V.; OROZCO, F.M. Toxic effect of sodium dodecilbencensulphonate, lead, petroleum and their mixtures on the activity of acetylcholinesterase of Moina macrocopa in vitro. Environ, Toxicol. Water 12: 1–5, 1997.
MAYES, P.A. Biologic oxidation. In Murray RK, Granner DK, Mayes PA, Rodwell VW (eds): Harper's biochemistry. San Mateo, Appleton & Lange, 105-11. 1990.
MCGEER, J.C.; SZEBEDINSZKY, C.; MCDONALD, D.G.; WOOD, C.M. Effects of chronic sublethal exposure to waterborne Cu, Cd or Zn in rainboe trout: Ionoregulatory disturbance and metabolic costs. Aquat. Toxicol. 50: 231-243, 2000.
MEISTER, A.; ANDERSON, M.E. Glutathione. Annu. Rev. Biochem. 52: 711-60, 1983.
MYERS, M.S.; JOHNSON, L.L.; OLSON, O.P.; STHER, C.M.; HORNESS, B.H.; COLLIER, T.K., MCCAIN, B.B. Toxicopathic hepatic lesions as biomarkers of chemical contaminant exposure and effects in marine bottomfish species from the northeastand Pacific Coast, USA. Mar. Pollut. Bull. 37, 1998.
PAYNE, J.F.; MATHIEU, A.; MELVIN, W. ; FANCEY, L.L.. Acetylcholinesterase, an old biomarker with a new future? Field trials in association whit two urban rivers and a paper mill in Newfoundland. Mar. Pollut. Bull. 32: 225-231, 1996.
RAND, G.M.; WELLS, P.G.; MCCARTY, L.S. Introdution to aquatic toxicology. In: Rand, G.M. Fundamentals of Aquatic Toxicology: Effects, Environmental Fate, and Risk Assessment. 2º ed. Ed. Taylor & Francis, 1995.
RANGE, H.P.; DALE, M.M. Farmacologia. 5ª ed. Guanabara Koogan, Rio de Janeiro, 2000.
REGOLI, F.; NIGRO, M.; BOMPADRE, S.; WINSTON, G.W. Total oxidant scavenging capacity (TOSC) of microsomal and cytosolic fractions from Antarctic, Arctic and Mediterranean scallops: differentiation between three potent oxidants. Aquat. Toxicol. 49: 13-25, 2000.
ROCHER, B.; LE GOFF, J. ; PELUHET, L.; BRIAND, M.; MANDUZIO, H.; GALLOIS, J.; DEVIER, M.H.; GEFFARD, O.; GRICOURT, L.; AUGAGNEUR, S.; BUDZINSKI, H.; POTTIER, D.; ANDRE, V.; PIKKARAINEN, A. Ethoxyresorufin-O-deethylase (EROD) activity and bile metabolites as contamination indicators in Baltic Sea perch: Determination by HPLC, 2006.

41
ROMÉO, M.; BENNANI, N.; GNASSIA-BARELLI, M.; LAFAURIE, M.; GIRARD, J.P. Cadmium and cooper display different responses towards oxidative stress in the kidney of the sea bass Dicentrarchus labrax. Aquat. Toxicol. 48: 185-194, 2000.
SCHLENK, D.; BENSON, W.H. Traget Organ Toxicity in Marine and Freshwater Teleosts. London, 1, 2001.
SCOTT, M.D.; LUBIN, B.H.; ZUO, L.; KUYPERS, F.A. Erythrocyte defense against hydrogen peroxide: preeminent importance of catalase. J. Lab. Clin. Med. 118: 7-16, 1991.
SHAN, X.; AW, T.Y.; JONES, D.P. Glutathione-dependent protection against oxidative injury. Pharm.Ther. 47: 61-71, 1990.
SILVA DE ASSIS, H.C. Der Einsatz von biomarkern zur summarischen erfassung von Gewässerverschmutzungen. Tese de doutorado apresentada na Universidade Tècnica de Berlim, Alemanha, 99p, 1998.
STENESH, J. Bioch.. Plenum. New York, EUA. p 596, 1998.
STURM, A.; SILVA DE ASSIS, H.C.; HANSEN, P.D. Cholinesterases of marine teleost fish: enzymological characterization and potential use in the monitoring of neurotoxic contamination. Mar. Environ. Res. 47: 389-398, 1999.
STURVE, J.; STEPHENSEN, E.; FÖRLIN, L. Effects of redox cycling compounds on DT diaphorase activity in the liver of rainbow trout (Oncorhynchus mykiss). Comp. Hepatol. 4: 4-10, 2005.
STURVE, J.; HASSELBERG, L.; FALTH, H.; CELANDER, M.; FÖRLIN, L. Effects of North Sea oil and alkylphenols on biomarker responses in juvenile Atlantic cod (Gadus morhua). Aquat. Toxicol. 78: 73-78, 2006.
TAKASUSUKI, J.; ARAUJO, M.R.R.; FERNANDES, M.N. Effect of water pH on copper toxicity in the neotropical fish, Prochilodus scrofa (Prochilodontidae). Bull. Environ. Contam. Toxicol. 72: 1075-1-82, 2004.
THOPHON, S.; KRUATRACHUE, M.; UPATHAM, E.S.; POKETHITIYOOK, P.; SAHAPHONG, S.; JARRITKHUAN, S. Histopathological alterations of White seabass, Lates calcarifer, in acute and subchronic cadmiun exposure. Environ. Pollut, 121, 2003.
VAN DER OOST, R.; BEYER, J.;VERMEULEN, N.P.E. Fish bioaccumulation and biomarkers in environmental risk assessment: a review. Environ. Toxicol. Pharmacol. 13: 57-149, 2003.
VIARENGO, A.; PONZANO, E.; DONDERO, F.E.R. A simple spectrophotometric method formetallothionein evaluation in marine organisms: na application to mediterranean and antartic molluscs. Mar. Environ. Res. 44: 69-84, 1997.

42
WATANABE, T.; KIRON, V.; SATOH, S. Trace minerals in fish nutrition. Aquac. 151: 158-207, 1997.
WHO: World Health Organization, Air Quality Guidelines for Europe, WHO Regional Office in Europe, Copenhagen. 1987.
YABE, M.J.S.; OLIVEIRA, E.O. Metais pesados em águas superficiais como estratégia de caracterização de bacias hidrográficas. Quim. Nova 21: 551-556, 1998.

43
4. REFERÊNCIAS BIBLIOGRÁFICAS GERAIS
ADAMS, S. M. Status and use of biological indicators for evaluating the effect of stress in fish. Amer. Fish. Soc. Symp. 8: 1-8, 1990.
AKAISHI, F.M.; SILVA DE ASSIS, H.C.; JAKOBI, S.C.G.; STJEAN, S.; COUTERNAY, S.C.; LIMA, E.;WAGNER, A.L.R.; SCOFIELD, A.; OLIVEIRA RIBEIRO, C.A. Morphological and neurotoxicological findings in tropical freshwater fish (Astyanax sp.) after waterborne and acute exposure to water soluble fraction (WSF) of crude oil. Arch. Environ. Contam. Toxicol. 46: 244–253, 2004.
AMORIM, L.C.A. O Uso dos Biomarcadores na Avaliação da Exposição Ocupacional a Substâncias Químicas. Rev. Brás. Med. Trab. Belo Horizonte 1: 124-132, 2003.
AVCI, A.; KAÇMAZ, M.; DURAK, I. Peroxidation in muscle and liver tissues from fish in a contaminated river due to a petroleum refinery industry. Ecotoxicol. Environ. Saf. 60: 101-105, 2005.
BAKER, R.C.; KRAMER, R.E. Cytotoxicity of short-chain alcohols. Annu. Rev. Pharmaco. Toxicol. 39: 127-150, 1999.
BARRINUEVO, W.R.; FERNANDES, M.N. Critical thermal maxima and ninima for curimbatá, Prochilodus scrofa (Steindachner) of two different sizes. Aqua. Res. 26: 447-450, 1995.
BENTLEY, P.J. Influx of zinc bt channel catfish (Ictalurus ounctatus):uptake from external environmental solutions. Comp. Bioch. Phys. 101C: 215-217, 1992.
BENZIE, I.F. Lipid peroxidation: a review of causes, consequences, measurement and dietary influences. Int. J. Food Sci. Nutr. 47: 223-261, 1996.
BETOULLE, S. Action du lindane sur l’activité sécrétrice (radicaux libres oxygénés et cytokines à activités « MAF ») des cellules phagocytaires de truite arc-en-ciel (Oncorhynchus mykiss) : implication du calcium intracellulaire. Thèse ( Doctorat en Biologie ,sciences et santé). Université de Limoges/France, 1998.
BEUTLER, E. Red cell metabolism: A manual of biochemical a methods. Grune & Stron, New York, 1975.
BEYERS, D.W.; SIKOSKI, P.J. Acethylcholinesterase inhibition in federally endangered Colorado squawfish exposed to cararyl and malathion. Environ. Toxicol. Chem. 13: 935–939, 1994.
BRYAN, G.W.; LANGSTON, W.J. Bioavailability, accumulation and effects of heavy metals in sediments with special reference to United Kingdom estuaries: a review. Environ. Poll. 76: 89-131, 1992.

44
CANLI, M.; STAGG, R.M.; RODGER, E.G. The induction of metallothionein in tissues of the Norway lobster Nephrops norvegicus following exposure to cadmium, copper and zinc: the relationships between metallothionein and the metals. Environ. Poll. 96: 343-350, 1997.
CAZENAVE, J.; BISTONI, M.L.A.; PESCE, S.F.; WUNDERLIN, D.A. Differencial detoxification and antioxidant response in diverse organs of Corydoras paleatus experimentally exposed to microcystin-RR. Aquat. Toxicol. 76: 1-12, 2006.
CHAOUI, A.; GHORBAL, M.H.; EL FERJANI, E. Effects of cadmium-zinc interations on hydroponically grown bean (Phaseolus vulgaris L.). Plan. Scien. 26: 21-28, 1997.
CHO J.R.; KIM Y. J.; HONG K. J.; YOO J. K.; LEE J.O.; AHN Y. J.; CHO J.R.; KIM Y.J.; HONG K.J.; YOO J.K.; LEE J.O.; AHN Y.J. Resistance monitoring and enzyme activity in field-collected populations of the spiraea aphid, Aphis citricola Van der Goot. J. Asian Pacific Entomol. 2: 113-119, 1999.
CONAMA, Conselho Nacional do Meio Ambiente, Resolução n.357 de 17/03/2005.
DENEKE, S.M.; FANBURG, B.L. Regulation of cellular glutathione. Am. J. Physiol. 257: 163-73, 1989.
ELLMAN, G.L.; COUTNEY, K.; ANDRES, V.; FEATHERSTONE, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharm. 7:88-95, 1961.
FENET, H.; CASELLAS, C.; BONTOUX, J. Laboratory and field caging studies on hepatic enzymatic activities in european eel and rainbow trout. Ecotox. Environ. Safety. 40:137-143, 1998.
FEUSSNER, I.; WASTERNACK, C.; KINDI, H.; KUHN, H. Lipoxygenase catalysed oxygenation of storage lipids is implicated in lipid mobilization during germination of cucumber. Proc. Nati. Acad. Sci. USA. 92: 11849-11853, 1995.
FEUSSNER, I.; KUHN, H.; WASTERNACK, C. The lipoxygenase dependent degradation of storage lipids. Trends Plant. Sci. 6: 268-273, 2001.
FRUGOLI, J.A.; ZONGH, H.H.; NUCCIO, M.L.; MCCOURT, P.; MCPEEK, M.A.; THOMAS, T.L.; MCCLUNG, C.R. Catalase is encoded by a multigene family in Arabidopsis thaliana (L.) Heynh. Plant Physiol. 112: 327-336, 1996.
FULTON, M.H.; KEY, P.B. Acetylcholinesterase inhibition in estuarine fish and invertebrates as an indicator of organophosphorus inseticide exposure and effects. Environ. Toxic. Chem. 20 (1): 37-75, 2001.
GALLEANO, M.; PUNTARULO, S. Effect of mild iron overload on liver and kidney lipid peroxidation. Braz. J. Med. Biol. Res. 27: 2.349-2.358, 1994.

45
GIMENO-GARCIA, E.; ANDREU, V.; BOLUDA, R. Heavy metals incidence in the application of inorganic fertilizers and pesticides to rice farming soils. Environ. Poll. 92: 19-25, 1996.
GIROTTI, A.W. Lipid hydroperoxide generation, turnover, and effector action in biological systems. J. Lipid. Res. 39: 1529-1542, 1998.
GIROTTI, A.W. Serial Review: Regulatory and Cytoprotective Aspects of Lipid Hydroperoxide Metabolism. Free Radic. Biol. Med. 33: 154-172, 2002.
GIULIO, R.T.D.; WASHBURN, P.C.; WENNING, R.J.; WINSTON, G.W.; JEWELL, C.S. Biochemical responses in aquactic animals: a review of determinants of oxidative stress. Environ. Toxicol. Chem. 9: 1103-1123, 1989.
GOODMAN, L.S.; GILMAN, A.G. As Bases Farmacológicas da Terapêutica. 9º Ed. Rio de Janeiro. McGraw Hill Interamericana Editores. 1435p, 1996.
GRAVATO, C.; TELES, M.; OLIVEIR, M.; SANTOS, M.A. Oxidative stress, liver biotransformation and genotoxic effects induced by copper in Anguilla anguilla L. – the influence of pre-exposure to b-naphthoflavone, 2006.
GUERREIRO, G.; DI FINIZIO, A.; CIARCIA, G. Stress-induced changes of plasma antioxidants in aquacultured sea bass, Dicentrarchus labrax. Comp. Biochem. Physiol. 132: 205-211, 2002.
GUL, M.; KUTAY, F. Z.; TEMOCIN, S.; HAANNINEN, O. Cellular and Clinical implications of glutathione. Ind. J. Exper. Biol. 38: 625-634, 2000.
HABIG, C.; DI GIULIO, R.T. Biochemical characteristics of cholinesterase in aquatic organisms. In: Mineau P (ed), Cholinesterase inhibition insecticides: their impact on wildlife in the environment. Elsevier, New York, pp 19–33. 1991.
HALLIWELL, B.; GUTTERIDGE, J.M.C. Free Rad. Biol. Med. 3º ed. Oxford, Oxford University Press, 936p, 2005.
HAMILTON, D.P.; MALIK, D.S.; SASTRY, K.V. Effects of zinc toxicity on biochemical composition of muscle and liver of murrel (Channa puntactus). Environ. Intern. 24(4): 433-438, 1998.
HANSEN, B.H.; ROMMA, S.; GARMO, O.A.; OLSVIK, P.A.; ANDERSEN, R.A. Antioxidative stress proteins and their gene expression in brown trout (Salmo trutta) from three rivers with different heavy metal levels. Comp. Biochem. Physiol. Part C: Toxicol Pharmacol 143: 263-274, 2006.
HEATH, A.G.; Water Pollution and Fish Physiology. 2º ed. Lewis Publishers, Boca Raton, p.357, 1995.
HEBBEL, R.P.Erythrocyte antioxidants and membrane vulnerability. J. Lab. Clin. Med. 107: 401-4, 1986.

46
HERMES-LIMA, M.; ZENTENO-SAVÍN, T. Animal response to drastic changes in oxygen availability and physiological oxidative stress. Comp. Bichem. Physiol. 133C: 537-556, 2002.
HINTON, D.E.; BAUMANN, P.C.; GARDNER, G.R.; HAWKINS, W.E.; HENDRICKS, J.D.; MURCHELANO, R.A.; OKIHIRO, M.S. Histopathologic Biomarkers. In: Hugget, R.J.; Kimerle, R.A.; Mehrle, J.R.; Bergman, H.L. Biomarkers: Biochemical, Physiological and Histological Markers of Anthropogenic Stress. Lewis Pubishers, Boca Raton, 1992.
HUGGET, R.J.; KIMERLE, R.A.; MEHRLE JR, P.M.; BERGMAN, H.L. Biomarkers biochemical, physiological and histological markers os anthropogenic stress. Lewis Publishers, Boca Raton, 347p, 1992.
IPCS. International Programme on Chemical Safety. Environ. Health Criteria 165 for Inorganic Lead, 1995.
JIANG, Y.; LEE, A.; CHEN, J.; CADENE, M.; CHAIT, B.T. ; MACKINNON, R. Crystal structure and mechanism of a calcium-gated potassium channel. Nat. 417: 515-522, 1991.
JOBLING, M. Human impacts on aquatic environments. In: Environ. Biol. Fishes. Chapman & Hall, London, 1995.
JORDÃO, C.P.; DA SILVA, A.C.; PEREIRA, J.L.; BRUNE, W. Contaminação por crômio de rios provenientes de curtumes em MInas Gerais. Quím. Nova, 47-52, 1999.
KANTONIEMI, A.; KIRSI, V.; OIKARI, A. The capacity of liver microsomes to form benzo(a)pyrene-diolepoxide DNA adducts and induction of cytochrome P450 1A in feral fish exposed to pulp mill effluents. Ecotox. Environ. Safety, 35: 136-141, 1996.
KEEN, J.H.; HABIG, W.H.; JAKOBY, W.B. Mechanism for several activities of the glutathione-S-transferase. J. Biol. Chem. 251: 6183-6188, 1976.
KENNEDY, C.J.; FARRELL, A.P. Ion homeostasis and interregnal stress responses in juvenile Pacific herring, Clupea pallasi, exposed to the water-soluble fraction of crude oil. J. Exp. Mar. Biol. Ecol. 323: 43-56, 2005.
KLEMZ, C. Uso de biomarcadores de contaminação ambiental em peixes Ancistrus sp (cascudo). Dissertação de Mestrado em Farmacologia. Universidade Federal do Paraná – UFPR, Curitiba. 2002.
KOZAR, R.A.; McKEONE, B.J.; POWNALL, H.J. Free radical induced alterations in endothelial cell function. J. Surg. Res. 56: 32-36, 1994.
LAMA, P.K.S.; GRAY, J. S. The use of biomarkers in environmental monitoring programmes. Mar. Poll. Bull. 46(2): 182-186, 2003.

47
LIU, J.H.; KUEH, C.S.W. Biomonitoring of heavy metals and trace organics using the intertidal mussel Perna viridis in Hong Kong coastal waters. Mar. Pollut. Bull. 51: 857–875, 2005.
LIVINGSTONE, D.R. Organic xenobiotic metabolism in marine invertebrates. In: Gilles, R. (Ed), Advances in Comparative and Environmental Physiology, 7. Springer, Berlin, pp. 45-185, 1991.
LIVINGSTONE, D.R.; LIPS, F.; GARCIA-MARTINEZ, P.; PIPE, R.K. Antioxidant enzymes in digestive gland of the common mussel, Mytilus edulis L. Mar. Biol. 112: 265-276, 1992.
LIVINGSTONE, D.R. The fate of organic xenobiotics in aquatic ecosystems: quantitative and qualitative differences in biotransformation by invertebrates and fish. Comparative Biochemistry and Physiology - Part A: Molecular & Integrative Physiology 120: 43-49, 1998.
LOMBARDI, J.V. Fundamentos de Toxicologia Aquática. In:RANZANI-PAIVA, M.J.T.; TAKEMOTO, R.M.; LIZAMA, M.A.P. Sanidade de Organismos Aquáticos. Ed. Varela, São Paulo, 2004.
LOWRY, O.H.; ROSENBROUGH, N.J.; FARR, A.L.; RANDALL, R.J. Protein measurements with the Folinphenol reagent. J. Biol. Chem. 193: 265-275, 1951.
MALAVOLTA, E. Fertilizantes e seu impacto ambiental: micronutrientes e metais pesados, mitos, mistificação e fatos. São Paulo: Petroquímica, 153p, 1994.
MALLICK, N.; MOHN, F.H. Reactive oxygen species: response of algal cells. J. Plant Physiol. 157: 183-193, 2000.
MARTINEZ-TABCHE, L.; RAMÍREZ, B.M.; GERMÁN-FAZ, C.; GALAR, C.I.; MADRIGAL, O.M.; ULLOA, G.V.; OROZCO, F.M. Toxic effect of sodium dodecilbencensulphonate, lead, petroleum and their mixtures on the activity of acetylcholinesterase of Moina macrocopa in vitro. Environ. Toxicol. Water 12: 1–5, 1997.
MARTINEZ-ÁLVAREZ, R.M.; HIDALGO, M.C.; DOMEZAIN, A.; MORALES, A.E.; GARCIA- GALLEGO, M.; SANZ, A. Physiological changes of sturgeon Acipenser naccarii caused by increasing environmental salinity. J. Exp. Biol, 205: 3699-3706, 2002.
MARTINEZ, C.B.R.; AZEVEDO, F.; WINKALER, E.U. Toxicidade e efeitos da amônia em peixes neotropicais. In: CYRINO, J.E.P.; URBINATI, E.C. Tópicos Especiais em Biologia Aquática e Aqüicultura, Jaboticabal – S.P: Sociedade Brasileira de Aqüicultura e Biologia Aquática, p.81-95, 2006.
MARTINEZ, C.B.R. Parâmetros bioquímicos de peixes para avaliação da qualidade da água. In: SILVA-SOUZA, A.T (Org) Sanidade de organismos aquáticos no Brasil. Maringá, P.R. Abrapoa, 2006.

48
MASON, R.P.; WALTER, M.F.; MASON, P.E. Effect of oxidative stress on membrane structure: small-angle X-ray diffraction analysis. Free Radic. Biol. Med. 23: 419-425, 1997.
MATSUI, K.; SHIBATA, Y.; TATEBA, H.; HATANAKA, A.; KAJIWARA, T. Changes of lipoxygenase and fatty acid hydroperoxide lyase activities in bell pepper fruits during maturation. Biosci. Biotechnol. Biochem. 61: 199-201, 1997.
MAYES, P.A. Biologic oxidation. In Murray RK, Granner DK, Mayes PA, Rodwell VW (eds): Harper's biochemistry. San Mateo, Appleton & Lange, 105-11. 1990.
MCGEER, J.C.; SZEBEDINSZKY, C.; MCDONALD, D.G.; WOOD, C.M. Effects of chronic sublethal exposure to waterborne Cu, Cd or Zn in rainboe trout 1: Ionoregulatory disturbance and metabolic costs. Aquat. Toxic. 50: 231-243, 2000.
MEAGHER, E.A. ; FITZGERALD, G.A. Indices of lipid peroxidation in vivo : Strengths ans limitation. Free Radic. Biol. Med. 26: 202-226, 2000.
MEISTER, A.; ANDERSON, M.E. Glutathione. Annu. Rev. Biochem. 52: 711-60, 1983.
MELLOR, J. W. Química Inorgânica Moderna; Vol II, Editora Globo, Porto Alegre, p 843, 1967.
MYERS, M.S.; JOHNSON, L.L.; OLSON, O.P.; STHER, C.M.; HORNESS, B.H.; COLLIER, T.K., MCCAIN, B.B. Toxicopathic hepatic lesions as biomarkers of chemical contaminant exposure and effects in marine bottomfish species from the northeastand Pacific Coast, USA. Mar. Poll. Bull. 37, 1998.
OLIVEIRA, J.P.; SIQUEIRA, M.E.P.B.; SILVA, C.S. Urinary nickel as bioindicator of workers Ni exposure in a Galvanizing plant in Brazil. Intern. Archives Occup. Environ. Health. 73: 65-68, 2002.
PAYNE, J.F.; MATHIEU, A.; MELVIN, W. ; FANCEY, L.L.. Acetylcholinesterase, an old biomarker with a new future? Field trials in association whit two urban rivers and a paper mill in Newfoundland. Mar. Poll. Bull. 32: 225-231, 1996.
RAND, G.M.; PETROCELLI, S.R. Fundamentals of aquatic toxicology: methods and applications. Hemisphere Publishing Corporation, Washington, 1985.
RAND, G.M.; WELLS, P.G.; MCCARTY, L.S. Introdution to aquatic toxicology. In: Rand, G.M. Fundamentals of Aquatic Toxicology: Effects, Environmental Fate, and Risk Assessment. 2º ed. Ed. Taylor & Francis, 1995.
RANGE, H.P.; DALE, M.M. Farmacologia. 5ª ed. Guanabara Koogan, Rio de Janeiro, 2000
REGOLI, F.; NIGRO, M.; BOMPADRE, S.; WINSTON, G.W. Total oxidant scavenging capacity (TOSC) of microsomal and cytosolic fractions from Antarctic,

49
Arctic and Mediterranean scallops: differentiation between three potent oxidants. Aquat. Toxicol. 49: 13-25, 2000.
ROCHER, B.; LE GOFF, J. ; PELUHET, L.; BRIAND, M.; MANDUZIO, H.; GALLOIS, J.; DEVIER, M.H.; GEFFARD, O.; GRICOURT, L.; AUGAGNEUR, S.; BUDZINSKI, H.; POTTIER, D.; ANDRE, V.; PIKKARAINEN, A. Ethoxyresorufin-O-deethylase (EROD) activity and bile metabolites as contamination indicators in Baltic Sea perch: Determination by HPLC. 2006.
ROMÉO, M.; BENNANI, N.; GNASSIA-BARELLI, M.; LAFAURIE, M.; GIRARD, J.P. Cadmium and cooper display different responses towards oxidative stress in the kidney of the sea bass Dicentrarchus labrax. Aquat. Toxicol. 48: 185-194, 2000.
SADIQ, M. Toxic metal chemistry in marine environments. Marcel Dekker Inc., New York, Basel, Hong Kong, 1992.
SCHEWE, T.; RAPOPORT, S.M.; KUHN, H. Enzymology and physiology of reticulocyte lipoxygenase: comparison with other piloxygenases. Adv. Enzymol. Relat. Areas Mol. Biol. 58, 191-272, 1986.
SCHICKLER, H.; CASPI, H. Response of antioxidative enzymes to nickel and cadmium stress in hyperaccumulator plants of the genus Alyssum. Phys. Plant. 105: 39-44, 1999.
SCHLENK, D.; BENSON, W.H. Traget Organ Toxicity in Marine and Freshwater Teleosts. London, v. 1, 2001.
SCOTT, M.D.; LUBIN, B.H.; ZUO, L.; KUYPERS, F.A. Erythrocyte defense against hydrogen peroxide: preeminent importance of catalase. J. Lab. Clin. Med. 118: 7-16, 1991.
SEVANIAN, A.; URSINI, F. Lipid peroxidation in membranes and low-density lipoproteins: similarities and differences. Free Radic. Biol. Med. 29: 306-311, 2000.
SHAN, X.; AW, T.Y.; JONES, D.P. Glutathione-dependent protection against oxidative injury. Pharm. Ther. 47: 61-71, 1990.
SILVA DE ASSIS, H.C. Der Einsatz von biomarkern zur summarischen erfassung von Gewässerverschmutzungen. Tese de doutorado apresentada na Universidade Tècnica de Berlim, Alemanha, 99p. 1998.
SINGH, O.V.; LABANA, S.; PANDEY, G.; BUDHIRAJA, R.; JAIN, R.K. Phytoremediation of toxic aromatic polluants from soil. Applied Microb. Biotech. 63:128-135, 2003.
SPRAGUE, J.B. Factors that modify toxicity. In: RAND, G.M. & PETROCELLI , S.R., (Ed.). Fundamentals of aquatic toxicology: methods and applications, New York: Hemisphere, p.124-163, 1985.

50
STEGEMAN, J.J.; BROWER, M.; DI GIULIO, R.T.; FORLIN, B.; FOWLER, B.; SANDERS, B.; VAN VELD, P. Molecular responses to environmental contamination: Proteins and enzymes as indicators of contamination exposure and effects. In Biomarkers: Biochemical, Physiological and Histological Markers of Anthropogenic Stress (R. J. Huggett, Ed.), pp. 235–335. Lewis Publishers, Boca Raton, Fl, 1992.
STENESH, J. Biochemistry. Plenum. New York, EUA. p 596, 1998.
STOREY, K.B. Oxidative stress: animal adaptations in nature. Braz. J. Med. Hist. Res. 96:1715-1733, 1996.
STURM, A.; SILVA DE ASSIS, H.C.; HANSEN, P.D. Cholinesterases of marine teleost fish: enzymological characterization and potential use in the monitoring of neurotoxic contamination. Mar. Environ. Res. 47: 389-398, 1999.
STURVE, J.; STEPHENSEN, E.; FÖRLIN, L. Effects of redox cycling compounds on DT diaphorase activity in the liver of rainbow trout (Oncorhynchus mykiss). Comp. Hepatol. 4: 4-10, 2005.
STURVE, J.; HASSELBERG, L.; FALTH, H.; CELANDER, M.; FÖRLIN, L. Effects of North Sea oil and alkylphenols on biomarker responses in juvenile Atlantic cod (Gadus morhua). Aquat. Toxicol. 78: 73-78, 2006.
TAKASUSUKI, J.; ARAUJO, M.R.R.; FERNANDES, M.N. Effect of water pH on copper toxicity in the neotropical fish, Prochilodus scrofa (Prochilodontidae). Bull. Environ. Contamin. Toxic. 72: 1075-1082, 2004.
THOPHON, S.; KRUATRACHUE, M.; UPATHAM, E.S.; POKETHITIYOOK, P.; SAHAPHONG, S.; JARRITKHUAN, S. Histopathological alterations of White seabass, Lates calcarifer, in acute and subchronic cadmiun exposure. Environ. Poll. 2003.
VAN DER OOST, R.; BEYER, J.;VERMEULEN, N.P.E. Fish bioaccumulation and biomarkers in environmental risk assessment: a review. Environ. Toxicol. Pharmacol. 13: 57-149, 2003.
VIARENGO, A.; PONZANO, E.; DONDERO, F.E.R. A simple spectrophotometric method formetallothionein evaluation in marine organisms: na application to mediterranean and antartic molluscs. Mar. Environ. Res. 44: 69-84, 1997.
WALKER, C. H.; HOPKIN, S. P.; SIBLY, R. M.; PEAKALL, D. B. Principles of ecotoxicology. Bristol: Taylor & Francis, 1996.
WATANABE, T.; KIRON, V.; SATOH, S. Trace minerals in fish nutrition. Aquac. 151: 158-207, 1997.
WHO: World Health Organization, Air Quality Guidelines for Europe, WHO Regional Office in Europe, Copenhagen. 1987.

51
WHO: World Health Organization. Lead- environmental aspects. Environmental Health Criteria 85. ISBN 9241542853. 1989.
YABE, M.J.S.; OLIVEIRA, E.O. Metais pesados em águas superficiais como estratégia de caracterização de bacias hidrográficas. Quim. Nova, 21: 551-556, 1998.


















