MAPEANDO O CURSO DE DESENVOLVIMENTO DO OVARIO EMBRIONARIO EM VERTEBRADOS
14
MAPEANDO O CURSO DE DESENVOLVIMENTO DE OVÁRIOS EM VERTEBRADOS KELLY A. LOFFLER E PETER KOOPMAN RESUMO: A decisão da gônada embrionária se diferenciar em testículo ou ovário é um passo critico no desenvolvimento de vertebrados. A base molecular desta decisão tem sido foco de muitos estudos, particularmente na década passada. Aqui nós vamos contrastar o conhecimento sobre o desenvolvimento gonadal recente e o desvio de diferenciação dos testículos com a falha da compreensão molecular de desenvolvimento do ovário em estágios recentes. Nós examinamos os conhecimentos correntes, considerando a morfogênese ovariana e propondo um modelo para controle hierárquico do ovário fetal, incorporando os poucos genes já conhecidos como vários sinais importantes, ou fatores que existem hipoteticamente no ovário recém formado. Introdução Um dos mais impressionantes aspectos de muitas espécies é a existência de dois sexos distintos, masculino e feminino, uma distinção que serve a uma função reprodutiva. Em grande número de espécies animais, incluindo humanos, esses aspectos levaram a uma significação psicológica e social complexa. A natureza dessas diferenças, e como elas surgem tem exercitado as mentes de cientistas e filósofos por milhares de anos. Só recentemente, com o desenvolvimento de tecnologias genetico-moleculares,os mecanismos da determinação sexual e do desenvolvimento gonadal começaram a ficar claros. A determinação do sexo dos mamíferos acontece em três fases distintas (fig 1). Na primeira, o sexo genético é determinado na fertilização pelo complemento cromossômico do espermatozóide fertilizador. Depois, durante o desenvolvimento embrionário, esta informação genética é traduzida para o sexo gonadal, isto é, ela determina o crescimento de cada testículo ou ovário a partir da gônada bipotencial indiferente recém formada. O terceiro estágio é a determinação sexual fenotípica, que começa na vida fetal ou pós-natal e continua através da puberdade, um período no qual a produção das gônadas direcionam a diferenciação dos ductos acessórios sexuais e da genitália externa. De acordo com os biólogos do desenvolvimento, nós nos interessamos mais pela segunda fase, cujos aspectos formaram foco deste artigo.
Transcript of MAPEANDO O CURSO DE DESENVOLVIMENTO DO OVARIO EMBRIONARIO EM VERTEBRADOS

MAPEANDO O CURSO DE DESENVOLVIMENTO DE OVÁRIOS EM VERTEBRADOS
KELLY A. LOFFLER E PETER KOOPMAN
RESUMO: A decisão da gônada embrionária se diferenciar em testículo ou ovário é um passo critico no desenvolvimento de vertebrados. A base molecular desta decisão tem sido foco de muitos estudos, particularmente na década passada. Aqui nós vamos contrastar o conhecimento sobre o desenvolvimento gonadal recente e o desvio de diferenciação dos testículos com a falha da compreensão molecular de desenvolvimento do ovário em estágios recentes. Nós examinamos os conhecimentos correntes, considerando a morfogênese ovariana e propondo um modelo para controle hierárquico do ovário fetal, incorporando os poucos genes já conhecidos como vários sinais importantes, ou fatores que existem hipoteticamente no ovário recém formado.
Introdução
Um dos mais impressionantes aspectos de muitas espécies é a existência de dois sexos distintos, masculino e feminino, uma distinção que serve a uma função reprodutiva. Em grande número de espécies animais, incluindo humanos, esses aspectos levaram a uma significação psicológica e social complexa. A natureza dessas diferenças, e como elas surgem tem exercitado as mentes de cientistas e filósofos por milhares de anos. Só recentemente, com o desenvolvimento de tecnologias genetico-moleculares,os mecanismos da determinação sexual e do desenvolvimento gonadal começaram a ficar claros. A determinação do sexo dos mamíferos acontece em três fases distintas (fig 1). Na primeira, o sexo genético é determinado na fertilização pelo complemento cromossômico do espermatozóide fertilizador. Depois, durante o desenvolvimento embrionário, esta informação genética é traduzida para o sexo gonadal, isto é, ela determina o crescimento de cada testículo ou ovário a partir da gônada bipotencial indiferente recém formada. O terceiro estágio é a determinação sexual fenotípica, que começa na vida fetal ou pós-natal e continua através da puberdade, um período no qual a produção das gônadas direcionam a diferenciação dos ductos acessórios sexuais e da genitália externa. De acordo com os biólogos do desenvolvimento, nós nos interessamos mais pela segunda fase, cujos aspectos formaram foco deste artigo. O desenvolvimento gonadal é o único exemplo embrionário de todo o alelo que tem dois destinos organogênicos possíveis – testículos versus ovário, dependendo mais de um ultimato genético do que de uma informação posicional. A despeito disso, os mecanismos celulares e morfogenéticos – migração celular, proliferação, determinação, diferenciação e comunicação, deposição dos componentes da matriz, e apoptose – que apóiam a formação das gônadas são similares àqueles usados no desenvolvimento de qualquer outro órgão no sistema. É esta marca única de aspectos típicos do desenvolvimento gonadal que continua a fascinar biólogos do desenvolvimento. Observações citogenéticas por volta da segunda metade do século passado levaram à conclusão de que, pelo menos em mamíferos, o desenvolvimento de um testículo está normalmente relacionado à presença do cromossomo Y (Ford et al., 1959; Jacobs and Strong, 1959). Experimentos de embriologia clássica por Jobs (1947), um pouco antes, estabeleceram que o fenótipo sexual de um mamífero recém-nascido está relacionado com a presença de um testículo no embrião. Testes de remoção cirúrgica fetal em coelhos resultaram em um desenvolvimento de genitália e características sexuais fenotípicas femininas, enquanto ovariectomia e implantação de um testículo ectópico em um embrião fêmea resultaram num

desenvolvimento de estruturas normalmente vistas em machos. A junção dessas duas informações levaram à conclusão de que o desenvolvimento de mamíferos machos é desengatilhado por sinais ativos no cromossomo Y. Assim, o desenvolvimento feminino foi considerado por meio século como um caminho de falha, a ser seguido apenas quando na ausência dos sinais promotores da masculinidade. Não surpreendentemente, na última metade do último século, as pesquisas focalizaram mais na busca do sinal determinante masculino codificado pelo cromossomo Y dos mamíferos.Casos de sexo reverso em humanos provaram ser especialmente úteis nesta pesquisa, desde indivíduos XX que se desenvolveram como machos, ou fêmeas convertidamente XY, que puderam ser mapeadas por pequenas duplicações ou deleções. Isso significa que a busca pelo gene determinante do testículo foi restrita a uma pequena região de cromossomos Y, que, por sua vez, foi confirmado conter um gene SRY (Gubbay et al., 1990; Sinclair et al., 1990). SRY foi confirmada ser a única sequência Y codificada necessária e suficiente para direcionar o desenvolvimento da gônada ao longo do pathway dos testículos, e consequentemente, para o desenvolvimento fenotípico masculino (Koopman et al., 1991). Desde a identificação giratória SRY, há mais de uma década, o trabalho de muitos laboratórios pelo mundo todo, incluindo o nosso, tem sido focado em desenvolvimento do testículo, e, em particular, no mecanismo molecular de ação do SRY. Muitos reguladores chave do desenvolvimento masculino foram identificados durante este tempo, de modo que o alvo direto do SRY continua notadamente difícil de se compreender. Por exemplo, o fator Sox9 relativo ao SRY foi implicado na diferenciação testicular como uma peça chave na decisão inicial das células somáticas da gônada em desenvolvimento ao longo do pathway masculino (Kent et al., 1996; Morais da Silva et al., 1996). O gene Dmrt1 relativo ao duplosexo/mab-3 implicou em ainda mais diferenciações dos testículos (Raymond et al., 1998; Raymond et al., 2000). Outros numerosos fatores foram identificados em passos cruciais do desenvolvimento gonadal primitivo em ambos os sexos, incluindo WT1, Sf1, Lim1, Lhx9, Emx2 e M33 (fig2). Não vem ao caso aqui uma discussão mais detalhada desses fatores, mas podem ser encontradas considerações acerca deles em Swain and Lovell-Badge (1999) e Capel (2000). Em contraste aos avanços na compreensão do desenvolvimento dos testículos, o desenvolvimento do ovário continua comparativamente misterioso. Muitos pesquisadores têm focado seu trabalho no desenvolvimento pós-natal do ovário e da função dos folículos ovarianos em fêmeas adultas. A partir desses estudos nós sabemos que a unidade funcional do ovário de mamíferos é o folículo, compreendido como uma célula germe ou um oócito envolto em camadas de células somáticas. Nós sabemos pouco acerca dessa unidade e da coordenação dos vários tipos de células necessários ao seu crescimento. A estrutura do ovário fetal e sua histogênese foram estudadas em detalhes na primeira metade do século passado, mas as observações foram de alguma maneira subjetivas, baseadas em teorias não muito acuradas das observações da origem e destino de vários tipos de células ovarianas. Alem disso, muito poucos casos de observações de reversão sexual produziram informações de sinais ativos que promovam o desenvolvimento ovariano, e muito pouco é conhecido acerca da genética molecular do desenvolvimento embrionário do ovário. Dois fatores foram identificados e são preferencialmente expressivos no desenvolvimento de ovários. São os chamados regulador transcripcional Dax1 e o sinalizador molecular Wnt4 (Vainio et al., 1999). Contudo, análises transgênicas conclusivas em ratos indicam que esta função é mais um fator anti-testículo repressor do pathway masculino em desenvolvimento, do que um determinante ovariano per se (Swain et al., 1998; Vainio et al., 1999). Neste paper nós examinamos o conhecimento corrente sobre determinação sexual e desenvolvimento gonadal embrionário, mas escolhemos o caminho menos percorrido, focando-nos no desenvolvimento ovariano.

Rua de Sentido Único: O Desenvolvimento Prematuro da Estrutura Genital
A embriologia e a genética molecular do desenvolvimento sexual de vertebrados tem sido estudada mais intensamente com ratos. Este estudo terá como referência principal as informações obtidas a partir de estudos de ratos.
Formação da Mesoderma IntermediáriaOs embriões primários tanto masculino quanto feminino não têm distinção segundo
critério morfológico. Em ratos, as diferenças genéticas entre machos e fêmeas não são expressas em diferenças estruturais até meados da gestação (Kaufman and Bard, 1999); em humanos, por volta de sete semanas de gestação (Moore, 1988). As populações mesodermais aparecem no tronco do embrião depois da gastrulação, entre oito e nove dias após o coito (dpc), e se reportam obrigatoriamente para a sua posição, entre o mesoderma somítico e o mesoderma lamino-lateral. Essa população mesodermal forma o precursor primitivo do sistema urogenital do animal adulto.
Desenvolvimento dos MesonefrosO sistema urogenital na maioria dos vertebrados forma-se em três estágios.
Primeiramente temos o estágio pronefro, mais cranial, que deve ter uma função excretora temporária no embrião, mas que não permanece no adulto. Logo em seguida temos o mesonefro, que tem uma função excretora em algumas espécies, mas que mantém outras importantes funções no desenvolvimento urogenital, como veremos a seguir. Finalmente, num estágio mais caudal, o metanefro se forma, diferenciando-se na função excretora do rim adulto. Invaginações do epitélio superficial formam os ductos de Wolffian (mesonéfrico) e Müllerian (paramesonéfrico). Ambos os ductos se formam a despeito do sexo do embrião. No embrião macho, os testículos em desenvolvimento produzem dois reguladores endócrinos chave. A testosterona dá suporte para a diferenciação dos ductos de Wolffian para formar a maior parte da genitália do macho, incluindo o vaso deferencial e o epididimídio. Enquanto os ductos de Müllerian regridem sob a influência do hormônio anti-Müllerian (AMH, também conhecido por substância inibidora de Müllerian, MIS). Na ausência desses fatores, na fêmea, os ductos Wolffian se degeneram e os ductos Müllerian dão origem aos oviductos, útero e vagina superior. A diferenciação pré-natal da genitália interna das fêmeas parece ser independente da influência hormonal esteróide (Couse et al., 2000), mas a função genital no adulto é altamente dependente dos esteróides sexuais produzidos pelo ovário. O sistema interno de tubulações no mesonefro não tem nenhuma função em mamíferos, mas é retido em machos para dar origem à rede vascular, linfática e nervosa dos testículos, que forma a comunicação entre o tubo seminal e o epidídio. A existência de uma região correspondente no ovário, já foi discutida por outros autores e tem uma origem embriológica correspondente em fêmeas (por exemplo Byskov, 1978).
Indução da Borda UrogenitalUm importante papel do mesonefro é induzir o desenvolvimento da gônada. A gônada
primitiva de ambos os sexos é observável morfologicamente primeiro como celoma epitelial da superfície ventromedial do mesonefro paralelo à linha mediana, por volta de 10 dpc no rato. Essa estrutura é chamada de crista gonadal (ou genital); a combinação da crista gonadal e do mesonefro sublinear é conhecida como crista urogenital. A crista urogenital prolifera e se diferencia para o ponto onde a gônada ovóide estendida pode ser vista anexada ao mesonefro tubular.
Três tipos principais de células somáticas formam a crista urogenital: as células de suporte, as células esteroidogênicas e as células estrômalas. Esses tipos de célula se

localizam originalmente na gônada bipotencial indiferente (Merchant, 1975). Cada uma se desenvolve tanto no pathway do macho como no da fêmea para formar o testículo ou o ovário (Koopman et al., 1991; Bishop et al., 2000) (fig3). Tradicionalmente, pensa-se que esta escolha dependa inteiramente da presença ou ausência de sinais promotores masculinos, mas o papel dos reguladores positivos na diferenciação das células ovarianas não deve ser negligenciado.
A linhagem das células de suporte formam a crista genital, que imediatamente circunda e garante o desenvolvimento das células germinativas, referentes às células de Sartoli no testículo, ou as células granulosas no ovário. As células da linhagem esterodoigênica se diferenciam em células Leydigs em embriões machos e, analogamente, em células teca dos ovários em embriões femininos. Células do estrômalo, tanto em testículos como em ovários, são responsáveis por padronizar os componentes estruturais do órgão, incluindo a produção da matriz extracelular e a formação dos vasos sanguíneos.
A maioria das células do estrômalo não são identificavelmente especializadas entre os dois sexos, com uma notável exceção, as celulas mioide peritubulares dos testículos, que formam a camada que envolve as células Sertoli. Essas células interagem com as células Sertoli para direcionar o desenvolvimento estrutural da corda dos testículos, que dá origem aos túbulos seminíferos do testículo maduro (Skinner et al., 1985). A formação da corda é uma diferença embriológica muito importante no desenvolvimento de ovários e testículos, sendo assim, bem significativo que as células mioides, originadas da migração específica masculina para a gônada, a partir dos mesonefros adjacentes (Martineau et al., 1997), dependam da ação do Sry (Capel et al., 1999).
A Colonização da Crista Genital Pelas Células germinativas PrimordiaisEnquanto as células somáticas da gônada em desenvolvimento se diferenciam a partir tanto do mesêncrima mesonéfrico como do epitélio celômico, as células germinativas primordiais têm uma origem embriológica diferente. As células germinativas primordiais são discerníveis primeiramente como um pequeno grupo de células positivas fosfatase-alcalina aglomeradas no mesoderma extra embrionário, logo após o início da gastrulação, por volta de 7 dpc no rato (Ginsburg et al., 1990). Essa aglomeração de células forma um escudo a partir de rearranjos que ocorrem no próprio embrião, e as células germinativas ficam presumidamente protegidas pela influência de diferenciação dos fatores de crescimento e morfogênese.
Entre 8,5 e 12 dpc, as células germinativas primordiais migram cranialmente da base do alantoide através do hindgut (origem embrionária do intestino grosso) até o mesoderma do mesonefro, e dali para a gônada. Enquanto este movimento pode ser trazido à tona por rearranjos que moldam todo o embrião, evidências sugerem que o alvo dessas células para a crista genital em desenvolvimento é ativamente controlado por um sinal químico-atrativo (Godin et al., 1990), e é, ao menos parcialmente, dependente do sistema de sinalização c-Kit/Steel (Buehr et al., 1993) e integrins (Anderson et al., 1999). Durante essa migração, a população de células germinativas primordiais prolifera por mitose, milhares das quais atingem a gônada indiferente.
Uma Bifurcação no Caminho: Determinação Sexual e Histogênese do Ovário
Muitas mudanças celulares e estruturais ocorrem no ovário em desenvolvimento entre a aparência inicial da crista genital e a formação dos primeiros folículos reconhecíveis, entre 1 e 3 dias após o parto. Enquanto essas mudanças não são tão rápidas e impactantes como as observadas no testículo em desenvolvimento, elas são, não obstante, importantes para o desenvolvimento e a função apropriada do ovário adulto.
O Começo da Estrutura: Formação das Cordas Ovígeras

O primeiro rearranjo histológico observável visto no ovário embrionário é a formação gradual, entre 12 e 13,5 dpc, de estruturas cordais livres chamadas classicamente de cordas ovígeras. (Odor and Blandau, 1969; Konishi et al., 1986). Essas cordas não são definidas pela lâmina basal, como acontece durante o desenvolvimento testicular, e não são observáveis a nível microscópico, mas são vistos em seções histoquímicas e imuno-histoquímicas (fig 4 e Frojdman et al., 1993).
No centro dessas cordas está um aglomerado de células germinativas primordiais e estas são envolvidas por células somáticas com uma aparência mesenquimal. Observações mais detalhadas das células germinativas mostraram que elas se interconectam fisicamente por uma ponte citoplásmica para formar o sincício ou o cito (Pepling and Spradling, 1998). Essas pontes são parecidas com as vistas nas células germinativas do cito de ovário de drosófilas, designadas canais anelares, e uma aposição próxima é observada entre as membranas plasmáticas das células germinativas e as células pré-granulosas somáticas que circulam os sítios desses canais. Observações mais recentes indicam que o colapso destes citos para formar os folículos primordiais individuais é programado e ocorre rapidamente entre um e três dias pós-parto (Pepling and Spradling, 2001).
Essas observações fazem surgir a possibilidade de que a comunicação direta célula-célula entre as células germinativas em desenvolvimento pode ser uma forma de coordenação de sua diferenciação temporal, e as pontes podem ser importantes para a distribuição da mitocôndria entre as células germinativas em desenvolvimento. Um contato próximo entre as células germinativas e as células pré-granulosas nos sítios dos canais anelares quando do colapso dos citos é consistente com um papel ativo na rápida reorganização do citos para formar folículos primordiais.
O Papel Central das Células germinativas MeióticasÉ questionável que as células germinativas sejam a razão da existência das gônadas.
Desse modo, é surpreendente que, no desenvolvimento embrionário masculino, a presença das células germinativas seja desnecessária para a apropriada diferenciação do complemento da célula somática da gônada. A falta de células germinativas em indivíduos XY desenvolve-se de maneira fenotipicamente normal como albeito estéril em machos (Merchant, 1975). Isso indica que a determinação do destino das células e a diferenciação das células somáticas gonadais masculinas independem da sinalização das células germinativas.
No ovário em desenvolvimento, contudo, a situação é marcadamente diferente; a presença das células germinativas é essencial para a construção e/ou a manutenção de um ovário normal. A disgênese ovariana é observada em células germinativas deficientes em ratos mutantes como W e Steel (ver Russel, 1979 e referências). Células germinativas meióticas exercem uma função reguladora importante sobre o desenvolvimento das células somáticas imediatamente circundantes do ovário. Em outros mutantes nas quais populações de células germinativas da ponte genital normalmente, mas subsequentemente, são depletadas, as células somáticas adjacentes se transdiferenciam para tomar a forma de fenótipo Sertoli (Behringer et al., 1990; Hashimoto et al., 1990; Couse et al., 2000). Isso sugere que o desenvolvimento de vários tipos de célula devem ser coordenados numa forma precisa temporal e espacialmente para a histogênese embrionária e para a diferenciação futura dos folículos e ocitos no ovário adulto. É interessante notar que no ciclo fisiológico normal do ovário, a perda do oócito do folículo desencadeia a diferenciação das células circundantes para formar o corpo lúteo, enfatizando a importância das interconexões entre oócitos e células granulosas.

Epitelialização Das Células Pré-Granulosas – Folículos DefinitivosA diferenciação coordenada das células das cristas genitais XX traz à tona a
organização gradual do ovário dentro das características da estrutura folicular observada no organismo adulto. Interações entre as células mesenquimais da crista genital, células da camada superior do epitélio celômico, e as células germinativas primordiais migratórias dirigem a morfogênese do ovário em crescimento. O desenvolvimento estrutural do ovário fetal e neonatal de mamíferos, incluindo a classificação morfológica dos estágios embrionários do desenvolvimento do folículo, está esquematizada na fig 5.
Microscópio eletrônico mostrou que as células somáticas , presumidamente células pré-granulosas, circulando as células germinativas do ovário embrionário, ampliam gradativamente o processo citoplásmico fino entre as células germinativas das cordas ovígeras ou citos de células germinativas (Odor and Blandau et al., 1969; Byskov, 1978; Pepling and spradling, 1998). Esses processos de células somáticas eventualmente vão formar uma camada completa que separa os aglomerados em oócitos individuais, cada um circundado por uma monocamada de células escamosas granulosas epitelializadas (Merchant-Larios and Chimal-Monroy, 1989). Simultaneamente, uma lâmina basal é depositada na periferia do folículo, envolvendo as células granulosas e separando-as das células do estrômalo (Merchant-Larios and Chimal-Monroy, 1989; Rajah et al., 1992). Esses dois aspectos definem o folículo primordial.
Como exposto acima, as interações entre oócitos em desenvolvimento e células pré-granulosas parecem ser criticas para a formação da monocamada celular do folículo. As interações entre as células pré-granulosas diferenciadas e as células do estrômalo (pré-tecais) são com certeza responsáveis pelo depósito da membrana basal que envolve o folículo definitivo (Rajah et al., 1992), de uma maneira análoga à cooperação entre as células Sertoli e as células mioides para a formação da membrana basal que envolve as cordas testiculares no macho embrionário (Skinner et al., 1985). Evidências existem em outros sistemas de desenvolvimento que sugerem que a membrana basal , uma vez formada, influencia o comportamento e a futura diferenciação das células adjacentes (Klein et al., 1988). A rápida progressão da diferenciação do folículo depois da formação inicial sugere que isso ocorra no desenvolvimento do ovário.
A aposição proximal entre as membranas plasmáticas dos oócitos presumptivos e as células granulosos sugere que esse processo histogenético é guiado por pistas suplementares para direcionar o contato célula-célula.; talvez um evento sinalizatório disparado por um sensor marginal da membrana ou ligado aos integrins envolvidos no contato celular (discutido por Brown et al., 2000), ou via fator sinalizador secretado em quantidades mínimas. Alternativamente, junções intercelulares podem estar envolvidas nesse estágio da morfogênese ovariana. Suapeitou-se durante muito tempo que junções gap entre o oócito e suas células granulosas são necessárias para manter a detenção meiótica como observado em folículos comandantes ou por coordenação de desenvolvimento em folículos ativados. Estudos conclusivos confirmam que o papel dos connexins, o principal componente das junções gap nesses processos (Carabatsos et al., 2000; Ackert et al., 2001). É possível que junções ainda descaracterizadas sejam responsáveis pela coordenação de desenvolvimento observada no ovário fetal.
Sinalizadores: Regulação Molecular do Desenvolvimento Fetal Ovariano
Muitos pesquisadores do século passado analisaram o desenvolvimento do ovário a nível morfológico. Por outro lado, a compreensão do desenvolvimento e da função do ovário fetal, nos níveis molecular e endócrino ainda é muito obscura. Não apenas os sinais de diferenciação celular são completamente misteriosos nesse estágio, como também nós

sofremos de uma carência muito grande de conhecimento sobre marcadores moleculares que tracejam o desenvolvimento, movimento e interação dos vários tipos de células no ovário em desenvolvimento.
Reguladores Restritivos do Destino Das células EmbrionáriasMuito pouco é sabido sobre a regulação molecular do desenvolvimento embrionário do
ovário fetal. O desenvolvimento pré-natal do sistema reprodutivo feminino é aparentemente independente da ação de hormônios esteróides (Course et al., 2000), mesmo que a função do órgão reprodutor adulto seja altamente dependente de reguladores esteróides. Dessa maneira,outros fatores devem controlar a diferenciação ovariana fetal e pré-púbere.
No embrião macho, o Sry atua automaticamente na célula como um fator de transcrição para disparar e regular a diferenciação do testículo em desenvolvimento. (Palmer and Burgoyne, 1991; Gubbay et al., 1990; Ferrari et al., 1992). Supõe-se também que Sox9 aja como um fator de transcrição no desenvolvimento de células Sertoli (Kent et al., 1996; Morais da Silva et al., 1996). Pode-se assim especular que fatores correspondentes muito provavelmente existam também na fêmea, e esses fatores agem nas células pré-granulosas para regular seu crescimento e desenvolvimento. Contudo, nenhum fator ainda foi identificado para identificar uma informação.
Interações Celulares Para Sinalização Recíproca e Controle do Desenvolvimento Contrariamente à nossa falta de conhecimento relacionada ao controle de diferenciação
celular no ovário embrionário em desenvolvimento, existem pelo menos alguns candidatos a efetivadores de sinalização celular. Entre esses, o fator de transcrição FIG (fator in germline alfa) foi identificado em células germinativas em desenvolvimento de ovários embrionários, inicialmente caracterizadas por seu papel na expressão da caracterização de genes decodificativos de proteínas estruturais da zona pelúcida (Liang et al., 1997). Análises conclusivas de genes em camundongos que a função deste fator é crucial para a formação dos folículos primordiais (Soyal et al., 2000). Esta função embrionária do FIG, principal na atividade de seus genes alvos conhecidos, indica que outros alvos ainda não identificados devem existir. Entre eles, alguns provavelmente são os fatores que mediam os sinal de sobrevivência da célula granulosa discutido acima. Muitos membros do fator beta de transformação do crescimento (TGFß), mostram-se importantes na regulação da função reprodutiva do ovário adulto, e um membro dessa família, fator 9 de diferenciação e crescimento (GDF9) é necessário para a diferenciação do ovário embrionário. Na ausência desta molécula sinalizadora, o desenvolvimento do folículo é bloqueado ao estágio de folículo primário , no qual as células granulosas não diferenciam alem de uma monocamada cuboidal e o crescimento do oócito é desregulado (Elvin et al., 1999b). A observação de que esses oócitos são maiores que o normal e se diferenciam como aberrações na ausência desse fator , indicam que as células granulosas em questão produzem um “fator regulador de crescimento do oócito”, via evento indutivo recíproco. A natureza desse sinal regulador permanece desconhecida.
Um modelo simples para integração e dos fatores autonômicos celulares discutidos acima está descrita na fig 6.
O Caminho À Frente
Claramente, a diferenciação do ovário de vertebrados é um processo ativo e complexo, que envolve diferenciação coordenada, migração e comunicação entre diversos tipos de células. A identificação de fatores reguladores desses processos no ovário em desenvolvimento será um desafio para os próximos anos. É provável que a projeção dessa expressão, a clonagem

posicional e procedimentos de identificação de genes candidatos revelem os genes responsáveis pela diferenciação das células granulosas e de outros tipos no ovário em desenvolvimento.
Recentemente, um gene candidato para a síndrome humana de blefarofimosis ptosis epicanthus inversus, tipo I e II foi identificado. A mutação nesse gene, decodificada como fator FOXL2 de transcrição, foi identificada em pacientes BPES (Crisponi etal., 2001; De Baere et al., 2001). O FOXL2 corresponde ao gene murine caracterizado parcialmente como PFrk, que é conhecido por se expressar no desenvolvimento da pituitária, no desenvolvimento craniofacial e ovariano, todos afetados em pacientes BPES, ainda em detalhamento.
Outra síndromes e modelos animais existem nas quais o desenvolvimento ovariano é anormal e nas quais a disgênese da reversão sexual ou gonadal é observada, e para as quais nenhum gene candidato foi identificado. A avaliabilidade da sequência do genoma humano, e os organismos desses vários modelos, vão fornecer valiosas ferramentas para a análise desses casos. Um a investigação cuidadosa desses casos sindrômicos podem dar-nos pistas relacionadas aos mecanismos de determinação sexual normal. Adicionalmente, investigações e um conhecimento crescente das causas moleculares de falhas no desenvolvimento de ovários podem fornecer insights relacionados ao pleno desenvolvimento desses órgãos. Por sua vez, avanços na compreensão do desenvolvimento de ovários podem dar pistas relacionadas ao tratamento ou prevenção de falhas ovarianas.
Insights muito úteis surgiram a partir da investigação da determinação sexual e mecanismos de diferenciação em outras espécies. Enquanto a morfologia e funcionalidade de ovários adultos são muito similares em vertebrados, os mecanismos moleculares que delineiam estas semelhanças são ainda desconhecidos, e as diferenças entre as espécies, com diferentes mecanismos de determinação sexual são potencialmente informativas. Em particular, a análise de determinação sexual e desenvolvimento gonadal em mamíferos não-vertebrados é provavelmente uma maneira produtiva de identificação de temas pertinentes na evolução do desenvolvimento sexual de vertebrados.
A investigação das funções moleculares são alvos reguladores de poucos genes implicados no desenvolvimento ovariano embrionário, Como a transcrição do fator FIG, e a molécula sinalizadora GDF9, que podem fornecer descobertas sobre o complexo mecanismo do desenvolvimento do ovário e sua regulação. A forma como a diferenciação em vários tipos de células é integrada para gerar um ovário adulto funcional, não resta dúvida, será um trabalho cheio de desafios e oportunidades para os pesquisadores nos anos vindouros.
FIG 1
Sexo Geneético XX XY
Sexo Gonadal Ovário Testículo
Sexo Fenotípico Fêmea Macho
FIG 2
Muitas chaves reguladoras de desenvolvimento gonadal são conhecidas. WT1, Sf1, Lim1, Lhx9, Emx2 e M33 são fatores conhecidos e necessários para o desenvolvimento urogenital embrionário em ambos os sexos. O Sry (cromossoma Y de determinação sexual) age como o desvio inicial para disparar o desenvolvimento sexual masculino. O Dax1 e Wnt4 são expressivos em desenv de ovários embrionários, mas análises transgênicas conclusivas indicam um papel mais importante para esses fatores, na repressão do padrão masculino de desenvolvimento. Reguladores ativos de desenv. De ovários embrionários não são ainda muito bem caracterizados

FIG 3
Determinação sexual nos principais tipos de células da gônada A gônada indiferente se forma em ambos os sexos na meia-gestação (topo), e pode se desenvolver tanto em ovários como em testículos dependendo da informação genética e do milieu hormonal (a base da figura representa ovários e testículos neonatais). Células-germe (amarelas) fazem surgir oócitos em fêmeas ou esperma em machos. As células de apoio (violeta) se tornam células Sartoli nos testículos e células granulosas nos ovários. Células esteroidogênicas (rosa) se tornam células Leydig em machos ou células theca em fêmeas. Em machos, o Sry induz à migração de células a partir dos mesonefros (verde) a qual inclui precursores das células peritubular mioides.
FIG 4
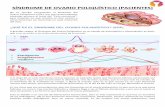
Desenvolvimento das Cordas OvígerasLâminas de ovários fetais de ratos com 16,5 dpc demonstram a presença de aglomerados de células-germe definidas em deposições laminares. (A) Uma secção de 8 µm formaldeido fixa, embebida em parafina colorida com hematoxilina e eosina, de acordo com método padrão. As células-germe são células redondas e grandes com núcleo largo e citoplasma pálido (setas). (B) Secção de 12µm congelada não-fixa de ovário de rato após detecção de imunoflorescência laminar. A escala indica 45 µm.
FIG 5
Desenvolvimento de Folículos em Ovário Neonatal e Fetal de RatosAglomerados de células-germe, ou citos de células-germe são envolvidos proximamente por céulas somáticas do ovário em desenvolvimento durante o crescimento fetal. Próximo ao dia do nascimento esses citos são rapidamente reorganizados como aqueles oócitos individuais são envolvidos por uma monocamada de células epiteliais pré-granulosas lisas que depositam uma membrana basal definitiva, formando os folículos primordiais. A ativação dos folículos pode ocorrer a qualquer hora poucos dias após o nascimento. Caracteriza-se pelo crescimento e diferenciação das células granulosas para formar primeiramente uma monocamada cuboidal, e então se expande para formar múltiplas camadas. O futuro desenvolvimento dos folículos ovarianos em preparação para a ovulação não é discutido aqui.
FIG 6
Controle Molecular Do Ovário Embrionário Em DesenvolvimentoO modelo esquemático dos passos reguladores conhecidos e propostos envolvidos na organização dos folículos primordiais mostra células germinativas (preto, Oo), células somáticas pré-granulosas, e células pré-teca (S). (A) Ovário embrionário onde os aglomerados de células germinativas são livremente envolvidos por células granulosas. Uma interação muito próxima e uma sinalização recíproca (setas) entre essas células, incluindo os chamados “sinais de sobrevivência de células granulosas” são essenciais para o desenvolvimento do folículo. (B) Representação de um estágio posterior do desenvolvimento fetal, onde o fator de transcrição das células germinativas FIG é essencial e o chamado fator específico de transcrição de células granulosas (TF). As setas representam os ainda não identificados eventos sinalizadores posteriores desses fatores de transcrição necessários à coordenação da diferenciação, como o “fator regulador do crescimento oócito” descrito no texto. (C) Ovário neonatal, no qual as interações entre oócitos e células pré-granulosas dividiram os aglomerados em folículos individuais. Um sinal de natureza desconhecida de células pré-granulosas para as células esteroidogênicas do estroma facilita a interação entre elas e, assim, a deposição de
uma membrana basal (BM, linha pontilhada).