
LÍVIA SCHEUNEMANN DOS SANTOS ESTUDOS SOBRE AS ...
108
LÍVIA SCHEUNEMANN DOS SANTOS ESTUDOS SOBRE AS PROTEÍNAS FERRITINA E OsNRAMP7 EM PLANTAS DE ARROZ (Oryza sativa L.) Tese submetida ao Programa de Pós- Graduação em Botânica da Universidade Federal do Rio Grade do Sul, como requisito parcial à obtenção do título de Doutor em Ciências. Orientador: Prof. Dra. Janette Palma Fett Professor Associado Centro de Biotecnologia & Departamento de Botânica Universidade Federal do Rio Grande do Sul Co-orientador: Dr. Rinaldo Pires dos Santos Professor Adjunto Departamento de Botânica Instituto de Biociências Universidade Federal do Rio Grande do Sul Porto Alegre, 2012
Transcript of LÍVIA SCHEUNEMANN DOS SANTOS ESTUDOS SOBRE AS ...

1
LÍVIA SCHEUNEMANN DOS SANTOS
ESTUDOS SOBRE AS PROTEÍNAS FERRITINA E OsNRAMP7
EM PLANTAS DE ARROZ (Oryza sativa L.)
Tese submetida ao Programa de Pós-
Graduação em Botânica da Universidade
Federal do Rio Grade do Sul, como
requisito parcial à obtenção do título de
Doutor em Ciências.
Orientador: Prof. Dra. Janette Palma Fett
Professor Associado
Centro de Biotecnologia & Departamento de Botânica
Universidade Federal do Rio Grande do Sul
Co-orientador: Dr. Rinaldo Pires dos Santos
Professor Adjunto
Departamento de Botânica
Instituto de Biociências
Universidade Federal do Rio Grande do Sul
Porto Alegre, 2012

2
There will come a time when you believe everything is finished.
Yet that will be the beginning.
Louis L'Amour

3
Agradecimentos
Começo agradecendo às mulheres de minha vida: minha mãe Ana Lore Scheunemann,
minha madrinha Inguelore Scheunemann e minha avó Eny Neutzling Scheunemann pelo
amor, carinho, amizade, compreensão e dedicação, bem como por tudo que me ensinaram. E à
minha família, que, de uma forma ou de outra, está sempre comigo.
À minha orientadora, Professora Janette Palma Fett, pela oportunidade de realizar o
doutorado em seu grupo. E claro, pela sua orientação, dedicação e confiança depositada.
Aos pedaços do meu coração que eu conheci no Laboratório de Fisiologia Vegetal e
sem os quais não sou completa: Paloma Menguer, Carolina Ruedell, Joséli Schwambach e
Naíla Cannes pelo amor, apoio, risadas, abraços e conversas intermináveis que acalmam o
coração e fazem sumir a distância.
Cíntia Pereira Barenho, obrigada por seres tu, amiga. Contigo sempre estarei em casa,
e acho que isso diz tudo.
Aos amados amigos, mais que colegas – Raul, Felipe, Fernanda, Anna, Hélio, Luiza,
Kaka, Edilena, Márcia e Cibele, pela ajuda, pelas conversas, chimarrão, churrasco e tudo de
bom que esse laboratório sempre teve.
Ao PPGBCM por ter pessoas especiais como a Lívia, Charley e Diogo, sempre
dispostos a ajudar. E a todos que, direta ou indiretamente, contribuíram de alguma forma para
a realização deste trabalho.
Ao meu orientador estrangeiro, Dr. Tony Miller, e demais membros do Disease and
Stress Biology Lab no John Innes Centre, por fazerem eu me sentir como parte da família em
apenas quatro meses de convívio.
De coração, muito obrigada a todos.

4
Resumo
O arroz é um dos cereais mais produzidos e consumidos no mundo, cultivado em
aproximadamente 156 milhões de hectares, com uma produção mundial de mais de 600
milhões de toneladas por ano. O arroz é, hoje, alimento básico para mais de dois terços da
população mundial. Contudo, minerais como ferro e zinco são perdidos durante o processo de
beneficiamento dos grãos para comercialização. Uma vez que a deficiência de ferro afeta
cerca de três bilhões de pessoas e é a deficiência mineral mais comum em humanos, diversos
esforços têm sido feitos para aumentar a concentração deste mineral em grãos de arroz.
Diversos projetos têm como objetivo compreender o mecanismo de translocação de nutrientes
para grãos de arroz, visando o aumento de sua concentração com fins de biofortificação do
alimento. Para melhor compreender a homeostase de ferro em plantas de arroz, conduzimos
experimentos para analisar possíveis funções de duas proteínas. Proteínas da família NRAMP
(Natural Resistance Associated Macrophage Protein) foram descritas como tendo
envolvimento na homeostase de ferro em diferentes organismos. OsNRAMP7 apresenta
propriedades características da família, como os motivos DPGN e MPH, possivelmente
envolvidos no transporte de metais. Oócitos de Xenopus injetados com o mRNA de
OsNRAMP7 apresentaram aumento significativo na concentração de ferro. A expressão
heteróloga da proteína em oócitos indica o envolvimento da proteína no transporte
transmembrana de ferro. Ferritina é outra proteína envolvida na homeostase de ferro nas
células. Ferritinas são proteínas esféricas, capazes de armazenar ferro no seu interior, agindo
também como um estoque de ferro nas células. O armazenamento de ferro dentro desta
proteína pode prevenir reações que levam a produção de radicais livres e, consequentemente,
estresse oxidativo. Duas cópias do gene da ferritina foram descritas em arroz. Respostas ao
estresse oxidativo em uma linhagem mutante de arroz para o gene OsFER2 foram estudadas.
Quando submetidas a excesso de ferro, plantas mutantes tiveram aumento na concentração de
MDA (malondialdeído) nas partes aéreas e da atividade da enzima APX (ascorbato
peroxidase) em raízes, revelando respostas ao dano oxidativo quando há baixa produção de
ferritina. Plantas mutantes acumulam menos biomassa do que plantas WT (wild type) mesmo
em condição controle de crescimento. Isso pode indicar um possível papel da ferritina na
homeostase de ferro em plantas de arroz, ainda que as mesmas não estejam em estresse por
excesso de ferro. Mecanismos compensatórios como o aumento da quantidade da proteína

5
frataxina e aumento do influxo de ferro para vacúolos também devem ser investigados. Mais
experimentos são necessários para melhor compreensão do papel da ferritina na homeostase
de ferro em arroz. Não obstante, com os experimentos aqui apresentados é possível
determinar o envolvimento da proteína OsNRAMP7 na homeostase de ferro em arroz.
Palavras-chave: Oryza sativa, ferro, OsNRAMP7, Xenopus, ferritina, estresse oxidativo.

6
Abstract
Rice is one of the most produced and consumed cereals in the world, cultivated in
approximately 156 million hectares, with a world production of over 600 million tons. It is a
staple food for two thirds of the world population. However, minerals such as iron and zinc
are lost during rice processing for commercialization. Since iron deficiency affects around
three billion people, and is the most common mineral deficiency in humans, several efforts
have been made in order to increase this nutrient’s levels in rice grains. Several projects have
as goal to understand translocation mechanisms of nutrients to rice grains as to increase their
levels for biofortification purposes. To better understand iron homeostasis in rice plants, we
conducted experiments in order to analyze the putative role of two proteins. The NRAMP
(Natural Resistance Associated Macrophage Protein) family was described as having an
important role in iron homeostasis in different organisms. OsNRAMP7 presents characteristic
features of the family, as motifs DPGN and MPH, said to be involved in metal transport.
Xenopus oocytes injected with OsNRAMP7 mRNA exhibited a significant increase in iron
content. Heterologous expression of the protein in oocytes indicated that the protein is
involved in transmembrane iron transport. Ferritin is another protein involved in intracellular
iron homeostasis. Ferritins are spherical proteins capable of storing iron in their core, also
acting as an iron buffer in cells. Storage of free iron inside this protein may prevent reactions
that lead to the formation of oxygen radicals and, therefore, to oxidative stress. Two ferritin
genes have been described in the rice genome. We studied the oxidative stress response of a
mutant line of rice with impaired expression of OsFer2. When subjected to iron excess,
mutant plants increased MDA (malondialdehyde) concentration in shoots and APX (ascorbate
peroxidase) enzyme activity in roots, revealing oxidative damage responses when ferritin
production is impaired. Mutant plants have lower weight than WT (wild type) even in control
growth condition. This may indicate a possible role of ferritin in iron homeostasis in rice
plants, even when they are not under iron stress. Compensative mechanisms such as increase
of frataxin levels and iron influx to the vacuole should be investigated. More experiments are
required for a proper understanding of ferritin role in iron homeostasis. Still, with these
experiments allowed to determine the involvement of the OsNRAMP7 protein in iron
homeostasis in rice.

7
Keywords: Oryza sativa, iron, OsNRAMP7, Xenopus, ferritin, oxidative stress.

8
Lista de abreviaturas
ABC – ATP Binding Cassete
APX – Ascorbate Peroxidase
AtFH – Arabidopsis thaliana frataxin
BSA – Bovine Serum Albumin
CAT – Catalase
cDNA – Complementary Deoxyribonucleic Acid
CTM – Consensus Transport Motif
DMA – 2’-Deoxymugineic Acid
DNA – Deoxyribonucleic Acid
DW – Dry Weight
EDX – Energy Dispersive X-ray
Et0/ABS – Quantum Yield of Electron Transport
Et0/TRO – Efficiency with which an electron can move to the PSI electron acceptor
FRD – Ferric Reductase
FRDL – Ferric Reductase Like
FRO – Ferric Chelate Reductase
FST – Flanquing Sequence Tags
Fv/Fm – Maximal Efficiency of PSII Photochemistry
FW – Fresh Weight
GFP – Green Fluorescent Protein
IRE – Iron Regulatory Elements
IRT – Iron Regulated Transporter
ITP – Iron Transport Protein
LEA – Late Embryogenesis Abundant
MA – Mugineic Acid
MATE – Multidrug and Toxic Compound Extrusion Transporter
MBS – Modified Barth’s Saline
MDA – Malondialdehyde
MIR – Mitochondrial Iron-Regulated
mRNA – Messenger Ribonucleic Acid
NA – Nicotianamine

9
NADPH – Nicotianamide Adenine Dinucleotide Phosphate
NRAMP – Natural Resistance Associated Macrophage Protein
OsFER – Oryza sativa Ferritin
OsUbq – Oryza sativa Ubiquitin
PCR – Polymerase Chain Reaction
PETIS – Positron-Emitting Tracer Imaging System
PIC1 – Permease In Chloroplast 1
PIC1ox – Permease In Chloroplast 1 overexpressing lines
PMSF – Phenylmethylsulphonylfluoride
PS – Phytosiderophores
PSII – Photosystem II
QTL – Quantitative Trait Loci
RGRC – Rice Genome Resource Center
RNA – Ribonucleic Acid
RNAi – RNA Interference
ROS – Reactive Oxygen Species
RT-PCR – Reverse Transcription Polymerase Chain Reaction
SOD – Superoxide Dismutase
SPR – Short Postembryonic Root
TBARS – Thiobarbituric Reactive Species
TCA – Trichloroacetic Acid
TMD – Transmembrane Domain
WT – Wild Type
YSL – Yellow Stripe Like

10
Sumário
Introdução ............................................................................................................................... 11
Justificativa ............................................................................................................................. 18
Objetivos .................................................................................................................................. 19
Geral ..................................................................................................................................... 19
Específicos ........................................................................................................................... 19
Capítulo 1: Artigo a ser submetido como revisão a periódico indexado ........................... 20
Introduction ......................................................................................................................... 21
Iron uptake ........................................................................................................................... 21
Translocation ....................................................................................................................... 23
Storage ................................................................................................................................. 29
Transcription factors related to iron homeostasis in rice ..................................................... 30
Biofortification ..................................................................................................................... 31
Conclusion and prospects .................................................................................................... 31
References ............................................................................................................................ 33
Figures ................................................................................................................................. 41
Capítulo 2: Iron Transport by the Rice OsNRAMP7 Protein............................................ 44
Abstract ................................................................................................................................ 46
Background .......................................................................................................................... 47
Results and Discussion ........................................................................................................ 48
Conclusions .......................................................................................................................... 51
Material and Methods .......................................................................................................... 51
References ............................................................................................................................ 56
Legends to figures ................................................................................................................ 62
Supplementary data .............................................................................................................. 64
Figures ................................................................................................................................. 65
Capítulo 3: Dual impact on rice plants bearing OsFer2 mutation .................................... 70
Abstract ................................................................................................................................ 71
Introduction .......................................................................................................................... 72
Results .................................................................................................................................. 73
Discussion ............................................................................................................................ 75
Material and Methods .......................................................................................................... 78
References ............................................................................................................................ 81
Figures ................................................................................................................................. 84
Considerações finais ............................................................................................................... 91
Referências .............................................................................................................................. 93

11
Introdução
O arroz é um dos cereais mais produzidos e consumidos no mundo, sendo cultivado
em aproximadamente 150 milhões de hectares, com produção mundial de 610 milhões de
toneladas em 2004 (IRRI, 2007), servindo assim, de alimento básico para dois terços da
população do planeta (Guidolin, 1993).
O arroz, Oryza sativa L., é uma planta anual, da família Poaceae, da qual são
conhecidas diferentes variedades cultivadas. Segundo Terres (1998), o arroz cultivado,
embora pareça ter sido originado de uma forma perene, é considerado uma gramínea semi-
aquática anual. O caule é um colmo, formado por nós e entrenós. De cada nó surge uma folha.
A estrutura básica vegetativa é o fitômero, o qual é constituído por um entrenó, um nó, uma
folha e uma gema. Freqüentemente, os entrenós basais são muito curtos, proporcionando uma
maior concentração de folhas na base da planta. As folhas envolvem o colmo e, têm
disposição alternodística e constam de bainha, lígula e lâmina. A bainha é a parte alongada da
folha, em forma de cartucho envolvendo o colmo, que nasce em cada nó e, assim, recobre o
entrenó. A lígula se constitui na estrutura membranosa situada na face adaxial da folha, na
região limítrofe entre a bainha e a lâmina, a qual é linear, paralelinérvia (Boldrini et al., 2005).
A inflorescência do arroz é uma panícula de espiguetas, constituídas de um par basal de
brácteas estéreis, denominadas glumas, e um eixo denominado ráquila, o qual sustenta as
glumas e os antécios (composto por pálea e lema). As glumas e os antécios têm disposição
alterno-dística sobre a ráquila. A flor é bissexuada, constando assim dos órgãos sexuais,
androceu e gineceu, e de um perianto rudimentar, representado pelas lodículas. É protegida
por duas brácteas, a pálea e a lema, que constituem o conjunto chamado antécio. O androceu
consta de seis estames e o ovário é bicarpelar, unilocular, unisseminado, com estilete curto e
dois estigmas plumosos (Silva, 1975; Boldrini et al., 2005). O fruto é uma cariopse, a qual
apresenta o pericarpo soldado em toda a sua extensão à testa da semente, deixando ver na
base, do lado dorsal, o embrião superficial, e do lado ventral o hilo. Outra característica é
apresentar endorsperma abundante (Boldrini et al., 2005).
A duração do ciclo vegetativo do arroz, isto é, o número de dias que decorre desde a
emergência até a maturação, é muito variável segundo a variedade considerada e as condições
de solo e de clima, podendo, no entanto fixar-se de 80 a 220 dias para as variedades da

12
subespécie indica e de 120 a 180 dias para as da subespécie japonica (Silva, 1975).
A nutrição mineral é um fator importante envolvido no crescimento e
desenvolvimento vegetal e, portanto, na sua produtividade. Entre os elementos minerais
essenciais, o Ferro (Fe) é um micronutriente de grande importância devido à sua implicação
em processos fundamentais como fotossíntese, respiração, fixação de nitrogênio e síntese de
DNA e às suas propriedades físico-químicas, participando em grande parte das reações
redutivas básicas (Briat et al., 1995; Briat & Lobréaux, 1997). Além disso, o ferro tem papel
essencial como componente de diferentes enzimas envolvidas na transferência de elétrons
(reações redox) como citocromos e age como um co-fator de enzimas essenciais envolvidas
na síntese de fithormônios (enzimas formadoras de etileno, por exemplo) (Bouzayen et al.,
1991; Siedow, 1991). Cerca de 75% do ferro na folha está presente nos cloroplastos, como
fitoferritina e ferredoxina, proteína que se sabe estar envolvida na transferência de elétrons no
processo fotossíntese, sendo reversivelmente oxidado de Fe2+
a Fe3+
durante a transferência de
elétrons. Portanto, a deficiência de ferro afeta em muito a fotossíntese (Taiz & Zeiger, 2004).
Em nível celular a alta reatividade desse metal pode vir a causar severos problemas.
As mesmas propriedades físicas que permitem que o ferro funcione como um eficiente cofator
e permita reações de catalisação em reações redox controladas permitem que o mesmo
funcione como uma potente toxina quando não é protegido de biomoléculas suscetíveis.
Várias reações intracelulares utilizam oxigênio molecular como aceptor de elétrons
produzindo superóxido ( 2O ) ou peróxido de hidrogênio (H2O2). Essas espécies não são
prejudiciais per se, mas contribuem para a geração de espécies reativas de oxigênio, no caso,
o radical hidroxila (•OH). Sua formação é catalisada por ferro através da Reação de Fenton
(Hell & Stephan, 2003):
2
2
2
3 OFeOFe
OHOHFeOHFe 3
22
2
Resumida:
OHOHOOHO 2222
Um dos mecanismos para a tolerância ao excesso de ferro em plantas pode ser a
capacidade de tornar o ferro absorvido indisponível. O armazenamento de ferro pode ocorrer
no espaço apoplástico, formado pelo continuum de paredes celulares de células adjacentes

13
bem como o espaço extracelular, em mitocôndrias (Zancani et al. 2004), plastídios (Seckback,
1982) e também no vacúolo, onde o baixo pH e altas concentrações de ácidos orgânicos
representam condições ótimas para o depósito de ferro, dependendo do órgão vegetal e da
espécie em questão (Briat & Lobréaux, 1998). O vacúolo é capaz de seqüestrar ferro, além de
outros metais, tanto como um mecanismo de desintoxicação celular como de armazenamento,
permitindo o crescimento celular quando em um ambiente com baixa disponibilidade de ferro
(Santos & Costa de Oliveira, 2007). Diferentes proteínas presentes no tonoplasto são
responsáveis pela translocação de metais através desta membrana. Em mitocôndrias e
plastídios, a proteína ferritina, é utilizada como meio de armazenamento de ferro, sendo
comum em plantas (Briat & Lobréaux, 1998).
A ferritina parece ser um componente importante para o controle da homeostase do
ferro em eucariotos, pois constitui uma classe de proteínas armazenadoras de ferro
amplamente distribuídas, que consiste em esferas formadas por 24 subunidades
simetricamente relacionadas que formam uma cavidade oca, sendo capaz de armazenar até
4500 átomos de ferro por molécula em seu interior (Harrison & Arosio, 1996). Ferritinas são
encontradas em diversos organismos, como animais, vegetais e bactérias (Briat et al., 2010).
Ferritinas vegetais têm sua sequência de aminoácidos altamente conservada com a de
mamíferos (Andrews et al., 1992). Contudo, diferentemente de ferritinas encontradas em
outros organismos, em plantas sua regulação se dá de maneira diferente. Enquanto em animais
essa regulação é traducional (Arosio et al., 2008), em plantas se dá pelo controle
transcricional (Lescure et al. 1991). A síntese de ferritina também é controlada pelo status de
ferro no interior da célula, podendo ser acumulada no caso de excesso de ferro para que o
metal seja armazenado de maneira atóxica para a célula, de forma que não reaja com oxigênio
(Briat et al., 1995).
A ferritina parece estar intimamente relacionada com o desenvolvimento vegetal,
contudo podendo exercer diferentes papéis. Em ervilhas, o ferro armazenado em ferritinas
corresponde a 92% do total de ferro encontrado em embriões maduros (Marentes & Grusak,
1998), evidenciando o importante papel da ferritina na germinação. Contudo, a mesma parece
estar envolvida com outros processos em sementes de Arabidopsis. Estima-se que não mais de
5% do ferro dessas sementes esteja armazenado em ferritinas (Ravet et al., 2009).
Arabidopsis thaliana possui quatro genes que codificam ferritina (Petit et al., 2001).
Apesar de serem estruturalmente conservados, são expressos sob diferentes condições. Em

14
situações de excesso de ferro, AtFer1 e AtFer3 são majoritariamente expressos (Petit et al.,
2001). AtFer4 codifica uma ferritina mitocondrial em plantas tratadas com ferro (Tarantino et
al., 2009) e AtFer2 é expresso em sementes e sua expressão responsiva a ácido abscísico
(Petit et al., 2001).
Dois genes que codificam para a proteína ferritina foram identificados no genoma de
arroz, OsFER1 e OsFER2 (Gross et al., 2003). Em plantas expostas a tratamentos com cobre,
Paraquat, nitroprussiato de sódio e excesso de ferro foi observado aumento na abundância de
transcritos de ferritina, particularmente de OsFER2 (Stein et al., 2009).
Dentre as famílias envolvidas na homeostase de metais, membros da família NRAMP
(Natural-Resistance-Associated Macrophage Protein) foram descritos como tendo um papel
importante na homeostase de ferro em diferentes organismos. Contudo, os membros desta
família gênica possuem amplo espectro de especificidade no transporte de metais, incluindo
ferro, manganês, cobalto, zinco, cobre, cádmio, níquel (Ňuňuková et al., 2010) e vanádio
(Ueki et al., 2011). Alguns membros desta família foram caracterizados utilizando a técnica
de expressão em oócitos de Xenopus (e.g. Agranoff et al., 2005, Ueki et al., 2011). Em
Arabidopsis foram isolados seis genes homólogos à família NRAMP (Mäser et al., 2001),
sendo que destes, AtNRAMP3 e AtNRAMP4 se mostraram capazes de complementar uma
linhagem de levedura mutante, defectiva para a absorção de ferro (Thomine et al., 2000). Em
estudo subseqüente, foi demonstrado que AtNRAMP3 é um transportador vacuolar (Thomine
et al., 2003). Foi demonstrado também que estes transportadores são capazes de mediar
transporte de ferro em Arabidopsis, estando envolvidos no efluxo de ferro dos vacúolos na
semente durante a germinação (Lanquar et al., 2005).
Através de análises in silico, foi possível observar que, dentre as oito proteínas
NRAMP previstas com base no genoma de arroz (Gross et al., 2003), o transportador
OsNRAMP7 possui a maior homologia com as proteínas AtNRAMP3 e AtNRAMP4, 68%
para ambas. Em um trabalho recente, nosso grupo observou correlação negativa significativa
entre a expressão de OsNRAMP7 em folhas bandeira durante o período de enchimento do
grão e a concentração final de ferro e zinco em grãos (Sperotto et al., 2010). Esse resultado
sugere o papel da proteína OsNRAMP7 no seqüestro de metais para o vacúolo em folhas,
resultando em uma menor disponibilidade do metal a ser transportado para panículas. O papel
do gene OsNRAMP7 na alocação de metais para o grão também foi sugerido por sua co-
localização com um QTL, capaz de explicar a variação fenotípica da concentração de ferro
(Stangoulis et al., 2007) e zinco (Garcia-Oliveira et al., 2009) nos grãos em 14% e 13%,

15
respectivamente.
Em um trabalho pioneiro, Boorer et al. (1992) utilizaram a técnica de análise funcional
de uma proteína de Arabidopsis thaliana em oócitos de Xenopus, definindo a afinidade da
proteína transportadora ao substrato. Nessa técnica, cRNA referente ao gene em estudo é
injetado em oócitos de Xenopus laevis, os quais possuem grande quantidade de enzimas,
organelas e proteínas, produzindo assim a proteína de interesse. Uma das grandes vantagens
do método é que há baixíssima atividade de transporte na membrana plasmática dessas
células, o que garante background inexistente ou muito baixo da atividade de transporte para
interferir com os resultados obtidos pela proteína de interesse, além de estudos de
eletrofisiologia serem facilitados por se tratarem de células grandes (Miller & Zhou, 2000).
Outra vantagem desta técnica se dá ao fato de a célula poder ser exposta a diferentes metais,
ligados ou não a diferentes quelantes (Koike et al., 2004), permitindo investigar não somente
a especificidade do transportador em relação ao metal, como também a forma química
preferencial de transporte do metal através do transportador em estudo. Proteínas vegetais
vacuolares foram expressas em oócitos e a direção do transporte in planta foi identificada
avaliando a diferença entre o transporte do metal em questão entre plantas mutantes que não
continham a proteína transportadora e plantas do tipo selvagem (Chopin et al., 2007).
Membros da família NRAMP já foram caracterizados utilizando esta técnica (Gunshin
et al., 1997; Okubo et al., 2003; Agranoff et al., 2005; Ueki et al., 2011), contudo nenhum
deles pertencente ao reino vegetal. Em um trabalho recente, realizado em colaboração com
Dr. Anthony Miller (John Innes Centre, Reino Unido), foi possível observar que oócitos de
Xenopus injetados com o mRNA de OsNRAMP7 foram capazes de absorver ferro na forma
Fe2+
(Capítulo 2 desta tese). Esta foi a primeira tentativa de caracterização de transporte desta
proteína e os resultados obtidos sugerem não só sua viabilidade, como também o
envolvimento da proteína no transporte de ferro e, consequentemente, na homeostase do metal
em plantas de arroz.
Em Arabidopsis, a importância de vacúolos no armazenamento de ferro em sementes e
sua remobilização durante a germinação foi documentada em nível molecular pela
caracterização da atividade de transportadores vacuolares de efluxo, NRAMP3 and NRAMP4,
e de influxo, VIT1. Foi então realizada a análise do gene de ferritina AtFER2 em diferentes
backgrounds genéticos que possuem a homeostase ferro dos compartimentos plastídicos e
vacuolares afetados (mutantes knockout fer, nramp e vit, e plantas superexpressando as
proteínas NRAMP e VIT) (Ravet et al., 2009). Tais estudos revelaram que a estabilidade da

16
ferritina em sementes depende da alocação apropriada do ferro do vacúolo para os plastídios,
evidenciando uma possível comunicação entre os compartimentos de armazenamento
vacuolares e plastidiais de ferro em sementes. Esses resultados indicam uma resposta
integrada quanto à homeostase de ferro nas células.
A compreensão dos mecanismos de regulação da homeostase do ferro em plantas é de
fundamental importância tanto do ponto de vista agronômico (possibilitando mais
produtividade em plantas que não sofram os efeitos danosos da deficiência ou do excesso de
ferro) quanto do ponto de vista da nutrição humana (possibilitando a produção de alimentos
de origem vegetal com altos níveis de ferro disponíveis para absorção pelo sistema digestivo
humano) (Gura, 1999; Grotz & Guerinot, 2002).
A caracterização de tais mecanismos em arroz é igualmente importante do ponto de
vista científico, uma vez que o arroz desponta como planta modelo (fisiológico e genético)
para as monocotiledôneas, fazendo par à dicotiledônea Arabidopsis thaliana. O arroz foi a
segunda Angiosperma a ter o seu genoma completamente seqüenciado (Burr et al., 2005),
sendo escolhido como organismo modelo para seqüenciamento entre as monocotiledôneas por
sua importância agronômica, pequeno tamanho do genoma (392 Mpb – o menor das
gramíneas) e sua relação de sintenia com outras espécies de cereais (IRGSP, 2005). O arroz
tem sido extensamente manipulado geneticamente, uma vez que é visto como modelo de
pesquisa para outras culturas (Devos & Gale, 2000). Dessa forma, permite que seja utilizado
para a realização de estudos de colinearidade molecular em outras espécies de gramíneas, e
assim, com base na sintenia, identificar e caracterizar genes de interesse em espécies
relacionadas.
Além disso, como alimento básico de grande parte da população humana (e
indiscutivelmente da brasileira) o arroz foi escolhido para experimentos pioneiros de
fortificação alimentar através da engenharia genética, já tendo sido obtidas plantas
transgênicas com maiores teores de betacaroteno (precursor da vitamina A), uma vez que a
deficiência de vitamina A também é um problema grave de saúde pública em países não
desenvolvidos (Ye et al., 2000; Paine et al., 2005).
Visto que a homeostase de ferro parece resultar de uma resposta integrada na célula, é
necessário que o estudo abranja não apenas uma das formas de armazenamento de ferro nas
células, dado que ambos, ferritina e vacúolo, se mostraram intimamente ligados em estudos
anteriores (Ravet et al., 2009).

17
Justificativa
Estudos acerca do desenvolvimento vegetal destacam o papel da ferritina como um
reservatório de ferro transiente para importantes processos ferro-dependentes como
fotossíntese e fixação de nitrogênio. Trabalhos recentes (Silveira et al., 2009, Stein et al.,
2009) propõem o envolvimento da ferritina na proteção ao estresse oxidativo em cultivares de
arroz submetidas a tratamento de excesso de ferro. Plantas de arroz são expostas
frequentemente a excesso de ferro, devido às condições resultantes do cultivo em condições
de alagamento. A produção de ferritina pode também ser induzida por outros fatores, visto
que a luz pode vir a induzir a produção da proteína (Stein et al., 2009). Contudo, mais
trabalhos são necessários para determinar o papel da ferritina na resposta ao estresse
oxidativo. Estudos utilizando mutantes são ferramentas úteis para a identificação ou a
confirmação da função de um gene. A utilização de uma linhagem de arroz mutante para
OsFer2 no presente trabalho tem como intuito gerar dados que contribuam para a
compreensão do papel da proteína ferritina na homeostase de ferro em plantas de arroz.
O arroz é um dos principais alimentos da população humana, mas contém baixas
concentrações de minerais essenciais, como o ferro, nos grãos. Como a principal deficiência
mineral em humanos é a de ferro, vários esforços tem sido feitos visando compreender e
manipular os mecanismos responsáveis pela alocação de ferro para os grãos de arroz. A
proteína OsNRAMP7 foi identificada dentre as proteínas co-localizadas com um Quantitative
Trait Loci (QTL) que explica parte da variação fenotípica da concentração de ferro
(Stangoulis et al., 2007) e zinco (Garcia-Oliveira et al., 2009) nos grãos de arroz. Além disso,
foi observada correlação negativa significativa entre a expressão de OsNRAMP7 em folhas
bandeira durante o período de enchimento do grão e a concentração final de ferro e zinco em
grãos (Sperotto et al., 2010), em trabalho realizado no Laboratório de Fisiologia Vegetal da
UFRGS. Por meio de análises in silico, foi constatado que OsNRAMP7 tem identidade de
68% com as proteínas AtNRAMP3 e AtNRAMP4 de Arabidopsis thaliana. Foi demonstrado
que AtNRAMP3 e AtNRAMP4 são proteínas transportadoras de ferro em vacúolos (Lanquar
et al., 2005). Até o momento, pouco se sabe sobre o transporte intracelular de ferro em arroz.
Determinar se OsNRAMP7 é, de fato, um transportador de ferro, poderá aumentar a
compreensão dos mecanismos necessários para a manutenção da homeostase deste metal em

18
plantas de arroz.
Desta forma justifica-se o presente trabalho, pelo qual pretende-se contribuir para o
aprofundamento, expansão e difusão das pesquisas sobre a homeostase de ferro em plantas de
arroz. Os resultados obtidos neste trabalho também poderão ser úteis para o desenvolvimento
de estratégias visando aumentar os teores de ferro no grão de arroz e a tolerância de plantas de
arroz ao excesso de ferro.

19
Objetivos
Geral
Pelo presente trabalho tem-se como objetivo principal contribuir para a elucidação de
mecanismos de regulação da homeostase de ferro em plantas de arroz (Oryza sativa L.). Para
tanto, o papel da proteína de armazenamento ferritina será investigado em relação aos
mecanismos de defesa e proteção contra o estresse oxidativo gerado pelo excesso de ferro. Da
mesma forma, determinar se a proteína transportadora OsNRAMP7 está envolvida com o
transporte de ferro em plantas de arroz pode esclarecer aspectos importantes da homeostase
deste metal.
Específicos
1. Analisar a presença dos transcritos de OsFer1 e OsFer2, bem como da proteína
ferritina, em plantas mutantes de arroz contendo inserção do transposon Tos17 no gene
OsFer2 (linhagem NG0250) e plantas do tipo selvagem (WT).
2. Investigar a propriedade protetora da proteína ferritina frente ao estresse oxidativo
gerado pelo excesso de ferro livre intracelular.
3. Expressar o mRNA de OsNRAMP7 em oócitos de Xenopus laevis isolados,
caracterizando a capacidade da proteína transportar ferro como potencial substrato.
4. Analisar características estruturais da proteína OsNRAMP7 e motivos relacionados ao
transporte de metais.

20
Capítulo 1
Artigo a ser submetido como revisão a periódico indexado

21
Review
Iron homeostasis in Plants: current knowledge on mechanisms and genes.
1. Introduction
Mineral nutrition is an important factor involved in plant development. Among the
essential mineral elements, iron is a micronutrient of great importance due to its physical-
chemical properties, participating in most of the basic reductive reactions. It is also essential
for basic processes as photosynthesis, respiration, nitrogen fixation and DNA synthesis (Briat
et al., 1995; Briat & Lobréaux, 1997). Iron has an essential role as component of different
enzymes involved in electron transfer (redox reactions) such as cytochromes and acting as a
co-factor of essential enzymes involved in phytohormone synthesis (eg ethylene synthesis)
(Bouzayen et al., 1991; Siedow, 1991). About 75% of the iron in leaves is in chloroplasts, as
phytoferritin and ferredoxin, proteins involved in electron transfer in photosynthesis,
reversibly oxidized from Fe2+
to Fe3+
during electron transfer. Therefore, iron deficiency
deeply affects photosynthesis (Msilini et al., 2011).
Owing to such important characteristics, we aimed to review the current knowledge on
iron uptake mechanisms, trafficking and storage in plants.
2. Iron uptake
Despite the fact that iron is the second most abundant metal in soils (first being
aluminum), it may not be available or easily absorbed by plants. For that reason, plants have
developed two strategies to assure iron absorption when exposed to iron deficiency
conditions. Two mechanisms underlying iron deficiency responses have been characterized
and in this situation iron is absorbed either by chelation or reduction strategies.
2.1 Reduction strategy
Iron absorption in non-grass plants is directly related to the root capacity to reduce
iron from Fe3+
to Fe2+
(Figure 1a). Also called Strategy I, it relies on the capacity of H+-
ATPases, located in the epidermis, to release protons to the rhizosphere (Santi et al., 2005).
Increment of H+ in the soil lowers the pH, increasing iron solubility. To be transported into
the cells, iron must be reduced from Fe3+
to Fe2+
. In Arabidopsis, this is carried out by a

22
NADPH-dependent ferric chelate reductase, AtFRO2 (Robinson et al., 1999). This reduction
is crucial for iron absorption in non-grass plants. Heterologous expression of AtFRO2 in
Soybean (Glycine max Merr.) led to an increase in Fe+3
reduction in both roots and leaves.
This enhanced activity reduced chlorotic phenotype and increased chlorophyll concentration
(Vasconcelos et al., 2006).
It is only after this step that roots can absorb iron. AtIRT1 is a divalent metal
transporter, with affinity not only for iron (Eide et al., 1996), but to Zn, Mn, Co, Cd
(Korshunova et al., 1999) and Ni (Schaff et al., 2006) as well. Mutant irt1 plants present
increased photosensitivity and altered chlorophyll fluorescence parameters. Plants are also
chlorotic and growth and fertility are significantly reduced, symptoms related to impaired iron
transport (Varotto et al., 2002). These evidences suggest a primary role of IRT in iron
absorption under iron-deficiency conditions.
2.2 Chelation strategy
Grasses make use of a different mechanism to absorb iron, called Strategy II. This
process, based on chelation of the iron molecule, occurs when grass roots release
phytosiderophores (PS) into the rhizosphere (Figure 1b). Of the PS molecules, maize (Zea
mays L) and rice (Oryza sativa L.) secrete 2’-deoxymugineic acid (DMA). However, other
grass species secrete hydroxylate DMA, releasing different mugineic acids (MAs) into the
soil. DMA is a molecule able of chelating Fe3+
, while transporters of the YSL family transport
this complex through the root (Ueno et al., 2007).
The first Fe-PS transporter identified was YS1 in maize. Mutant ys1 plants are
deficient in Fe3+
-MA uptake, leading to a constitutive iron-deficiency response. Due to the
lack of iron, leaves display interveinal chlorosis (Curie et al., 2001). In rice, OsYSL15 is
expressed in root epidermis and stele, being induced by iron-deficiency (Inoue et al., 2009). It
has also been determined that this protein has high affinity for Fe3+
-DMA (Inoue et al., 2009).
Rice osysl15 mutant plants showed chlorotic phenotype under iron deficiency conditions and
reduced iron concentration in all organs (Lee et al., 2009).
Since this strategy is not based on iron solubility in the rhizosphere, it is less sensitive
to pH in the soil. The capacity of plants to release PS and the availability of iron in the
environment are the limiting factors for this strategy.
2.3 Combined strategy

23
A combination of the strategies described above was also reported. Rice presents not
only the PS release to the rhizosphere, but also to reduce Fe3+
to Fe2+
and to transport the
latter using the OsIRT1 transporter (Ishimaru et al., 2006) (Figure 1c). When Fe3+
-DMA and
Fe2+
were supplied to rice plants, a Positron-Emitting Tracer Imaging System (PETIS)
experiment allowed to observe that both forms were absorbed. When OsIRT1 was over-
expressed in rice plants, iron and zinc content was elevated in shoots, roots and mature seeds
(Lee & An, 2009).
Rice is less tolerant to calcareous soils then barley or maize, since it releases less PS
into the rhizosphere than these species (Nagasaka et al., 2009). Compensating this deficit with
the expression of a divalent metal transporter, such as OsIRT1, may enable rice to sustain
normal growth under iron-limiting conditions.
3. Translocation
A sequence of processes that involve several metal chelators and transporters are
required for the safe translocation of iron within the plant. Chelators are used by plants due to
the metal’s chemical properties, such as high reactivity and poor solubility. It aims to prevent
formation of reactive oxygen species (ROS) and hydroxyl radical (OH), a formation
catalyzed by iron through the Fenton Reaction and a possible precipitation of the metal (Hell
& Stephan 2003).
3.1 Citrate
Fe3+
-citrate is known to be the major form of iron present in xylem exudates (Grotz &
Guerinot 2006) and is involved in long-distance transport of iron (Yokosho et al., 2009).
FRD3, a multidrug and toxic compound extrusion (MATE) transporter, is a gene mainly
expressed in roots and is involved with the translocation of iron-citrate chelates (Durrett et al.,
2007). Due to its localization in the plasma membrane of cells in the pericicle and vasculature
(Green & Rogers, 2004), FRD3 seems to be responsible for the iron-citrate transport to the
xylem (Durrett et al., 2007).
The Arabidopsis thaliana ferric reductase defective3 (frd3) mutant exhibits a chlorotic
phenotype and constitutive expression of iron uptake responses. As a result, the mutant
accumulates iron in roots, however not being able to translocate the metal via the xylem to
aerial parts, leading to iron deficiency in leaves and a chlorotic phenotype. Of the six

24
orthologs found in rice, OsFRDL1 is also expressed in the pericicle of root cells and is found
to transport citrate when expressed in Xenopus oocytes (Yokosho et al., 2009).
Also, the osfrdl1 loss of function insertion mutant has a similar phenotype to frd3 –
leaf chlorosis, lower leaf iron concentration and precipitation of iron in the root. Although
there was a decrease in the Fe3+
concentration in xylem sap, the same was not observed for
Fe2+
, suggesting the use of another chelating molecule by the plant to transport Fe2+
through
the xylem (Yokosho et al., 2009).
3.2 Nicotianamine
NA is a non-proteogenic amino acid ubiquitous in plants that chelates both Fe2+
and
Fe3+
, in addition to other divalent metals (Haydon et al., 2007). The chelation properties of
NA, such as affinity and stability, are the highest at neutral and mild basic pHs, making the
molecule more suitable for phloem transport than other compounds, such as organic acids
(Curie et al., 2009).
Most of the information regarding NA has come from the tomato chloronerva (chl)
knock-out mutant, defective in NA synthase. Plants lacking NA show interveinal chlorosis in
young growing leaves and constitutively activate their root iron-uptake systems, despite their
mature leaves containing a high amount of iron (Conte & Walker, 2011). As a result of the
apparently immobility of iron in the phloem, younger leaves lack iron. This contrasts with the
total iron transported to older leaves, supposedly via xylem, which remains normal (Conte &
Walker, 2011).
Both Arabidopsis and rice have proteins described as being involved in transporting
Fe-NA complexes. Of the eight members in Arabidopsis, YSL1 and YSL2 have been
described as transporters of Fe-NA (DiDonato et al., 2004). Out of the 18 members of the YS
Like family in rice, OsYSL2 has been demonstrated to transport Fe-NA complexes through
heterologous expression in Xenopus oocytes (Koike et al., 2004), but not Fe3+
-PS.
3.3 2’-deoxymugineic acid (DMA)
DMA is a PS responsible for chelating Fe3+
molecules in the rhizosphere to enable its
absorption by YSL family members located in roots of graminaceous plants (Inoue et al.,
2009). DMA may also be involved in iron translocation within the plant. It was detected in the
phloem sap of rice leaves (Mori et al., 1991; Higuch et al., 2001) and OsNAAT1 (Inoue et al.,
2009), OsNAS1–3 (Inoue et al., 2003) and OsDMAS1 (Bashir et al., 2006) are expressed in the

25
phloem companion cells of iron deficient leaves. These genes encode key enzymes in the
biosynthetic pathway of MAs.
3.4 ITP
Another iron-binding protein was identified when analyzing the phloem-mediated
transport of micronutrients during the germination of Ricinus communis (Kruger et al., 2002).
The Iron Transport Protein (ITP) was identified in phloem exudates, where it appears
associated to the radio labeled iron supplied to the plantlets. The protein showed high affinity
to Fe3+
but not to Fe2+
in vitro, where it also complexes Cu2+
, Zn2+
and Mn2+
(Kruger et al.,
2002).
The ITP from castor bean shows high similarity to the stress-related family of late
embryogenesis abundant (LEA) proteins. The most similar annotated sequences in both
Arabidospsis and rice are related to stress induced responses. The Arabidopsis sequences are
apparently involved in responding to water stress, while rice’s are to both to water and saline
stress (Kruger et al., 2002).
3.5 Intercellular
Once absorbed by roots, iron is likely complexed by chelating molecules (Figure 2)
due to the metal’s chemical properties. It is then translocated to the xylem as part of the
distribution process within the plant. An important molecule for xylem movement of iron is
citrate. Iron transport into xylem cells can be made by transporters such as AtFPN1 (Morrisey
et al., 2009) or by proteins known to transport citrate, such as AtFRD3 (Durret et al., 2007)
and OsFRDL1 (Yokosho et al., 2009).
Mechanisms involved in iron loading from xylem to phloem vessels have already been
described. Is has been reported that OsIRT1 is involved in iron transport in the stele and that it
is specific for Fe2+
(Ishimaru et al., 2006).
Members of the OligoPeptide Transporter (OPT) family were also described in iron
translocation in plants, including genes of the YSL subfamily. Among 18 putative YSL genes
identified in the rice genome, OsYSL2 had its expression observed in phloem cells of the
vascular bundles of leaves and leaf sheaths. It has been recently demonstrated that the protein
encoded by this gene is vital for the long-distance transport of not only Fe, but also Mn. The
RNAi (OsYSL2i) line increased iron concentration in roots while decreasing Fe and Mn
concentrations in shoots. When the gene expression was driven by the sucrose transporter

26
promoter, iron concentration in the polished grain was increased by 4.4 fold (Ishimaru et al.,
2010). These results indicate the importance of the OsYSL2 protein in Fe-NA translocation in
rice plants.
Arabidopsis has eight members of the YSL gene family identified in its genome. Of
them, YSL1, YSL2 and YSL3 have been characterized as important for metal homeostasis.
Although it is thought that AtYSL2 is involved in Fe transport, conflicting results were
obtained when groups attempted to establish its function. While DiDonato et al. (2004)
reported restored growth of fet3fet4 yeast mutant complemented with OsYSL2 only when
supplied with Fe-NA, Schaaf et al. (2005) claim that the observation could not be made in
their experiment. However, additional experiments done by the latter group support the
protein’s involvement in iron homeostasis.
The Arabidopsis ysl1ysl3 double mutant exhibited Fe deficiency symptoms, such as
interveinal chlorosis, low concentration of iron in leaves and impaired mobilization of metals
from leaves during senescence (Waters et al., 2006). Although the ysl1ysl3 mutant showed
interveinal chlorosis, as the chl mutant, its Fe deficiency response remained unaltered. This
could be an indication of a tissue-specific response in Arabidopsis. These proteins are also
involved in translocation of Fe into seeds, since the concentrations of Fe, Zn and Cu were
lower when the proteins were impaired. Seed fertility was also reduced in the double mutant,
since anthers and embryos had defective development (Waters et al., 2006).
Of the same family, AtOPT3 has an essential role in embryo development. A mutation
on this gene induced continued Fe deficiency responses in roots, high level of Fe in tissues
due to continuous absorption, and development of necrotic areas. Despite the high amount of
Fe in plants, atopt3 mutants showed less Fe in seeds, indicating an important role for the
protein in Fe translocation to developing seeds (Stacey et al., 2008). Among the 15 putative
members of the OPT family in Oryza sativa cv. japonica (Gomolplitinant & Saier, 2011), the
one with highest identity to AtOPT3 is Osa13 (GenBank accession number 115455379).
Further studies are required to determine if this rice putative protein has indeed functions
related to its Arabidopsis homologue.
3.6 Intracellular
Once inside the cell, iron must be compartmentilized, to avoid toxic effects possibly
generated by oxidative stress. Iron storage compartments and molecules are of great
importance for supplying iron to essential processes, maintaining cell functions. An overview

27
of mechanisms involved in compartimentalization are described in Figure 3.
3.6.1 Vacuole
The largest iron storage compartment, the vacuole, is essential for maintaing iron
homeostasis. It is of special importance in seeds, where it provides iron before the organism is
capable of aquiring the metal from the environment (Lanquar et al., 2005, Kim et al., 2006).
Two proteins, AtFPN2 and AtVIT1, are known to be involved in iron loading to the vacuole.
AtFPN2 has its expression localized at the outermost layers of Arabidopsis roots (Morrisey et
al., 2009). It also appears to be involved in translocation of other metals besides iron (Schaaf
et al., 2006). This is coherent with the fact that fpn2 mutants are more sensitive to Co and Ni
than the wild type (WT) (Morrisey et al., 2009).
AtVIT1 influx protein is involved in iron loading to seeds and is highly expressed
during germination and development of young seedlings (Kim et al., 2006). Heterologous
expression of AtVIT1 was able to complement the iron-sensitive phenotype of the yeast
mutant line ccc1. Upon expression, an increase in vacuolar iron content was observed,
confirming the protein role as a vacuolar iron transporter (Kim et al., 2006).
In Arabidopsis seeds, iron is mainly located in the provascular strands of the
developing embryo. However, when the AtVIT1 protein had its function impaired this was no
longer observed. Although mutant seeds had the same content of iron as WT, its seeds didn’t
have the same germination development as WT seeds in alkaline pH conditions (Kim et al.,
2006).
The same was observed in mutants of the vacuolar transporters AtNRAMP3 and
AtNRAMP4. Arabidopsis nramp3 nramp4 double mutants have impaired germination in low
iron conditions, despite having the same seed iron content as WT. This happens due to seed
incapacity of mobilizing iron from vacuoles. Analysis using Energy Dispersive X ray (EDX)
showed that, after two days of germination, the vacuole of mutant seedlings still contained
iron, while WT had remobilized the iron for a proper germination. This is especially
important due to the fact that AtIRT1, an iron uptake transporter, is expressed only after the
third day of germination (Lanquar et al., 2005).
Our group identified eight NRAMP family members in the rice genome (Gross et al.,
2003). In a recent work, a significant negative correlation was found between OsNRAMP7
(LOC_Os12g39180) expression in flag leaves during grain filling and final Fe and Zn
concentrations in the grain (Sperotto et al., 2010). To investigate OsNRAMP7 influence on

28
iron homeostasis, heterologous expression of the protein was conducted in Xenopus oocytes.
We observed that oocytes expressing the protein presented a significant increase in iron
content, in two iron concentrations tested (Santos et al., data not published). This
demonstrates that OsNRAMP7 is able to perform transmembrane iron transport and is,
indeed, involved in cellular iron homeostasis.
3.6.2 Chloroplast
The vacuole is the major iron storage in seeds. In leaves, however, plastids are
responsible for up to 80% of the iron content (Shikanai et al., 2003). Iron concentration must
be tightly regulated in chloroplasts, since the photosynthetic electron transport chain produces
ROS, which may react with iron leading to oxidative damage. Although not much is known
about iron trafficking in this organelle, recent findings have shed light on the transport
mechanism involved.
To be transported into chloroplasts, Fe must first be reduced from Fe3+
to Fe2+
.
AtFRO7, a member of the ferric reductase oxidase family, is localized in chloroplasts.
Chloroplasts of Arabidopsis fro7 mutants had 33% less iron than WT, and defects in
photosynthetic transport were also observed (Jeong et al., 2008). This could be an indication
of the existence of an influx iron transporter specific for divalent metals.
Recent findings in sugar beet (Beta vulgaris L.) have demonstrated that uptake
transporters present in intact chloroplasts have a preference for ferric iron complexes as
substrate. It also supports the existence of an active ferric chelate reductase, localized in the
inner membrane and that uses NADPH. Once inside the inner envelop, these ions are
incorporated into Fe-S/heme cofactors (Solti et al., 2012).
In a screen for metal transporters in plastids, Arabidopsis’ Permease In Chloroplasts1
(PIC1) was identified. Arabidopsis PIC1 mutants had chloroplast development impaired and
an increase in ferritin clusters. They also presented dwarfism and chlorotic phenotype (Duy et
al., 2007). Yeast complementation assay confirmed that AtPIC1 transports iron (Duy et al.,
2007), although it is not certain if Fe3+
or Fe2+
. PIC1ox (PIC1-overexpressing lines), however,
resembled ferritin knock-out plants. These plants presented symptoms as oxidative stress and
leaf chlorosis, which could be due to an increase of iron concentration in chloroplasts. PIC1
function in iron homeostasis could also be indicated by the results obtained when it was
overexpressed, leading to impaired plant growth, especially in fruit development (Duy et al.,
2011).

29
3.6.3 Mitochondria
As chloroplasts, mitochondria have a large demand for iron, used as a cofactor in the
respiratory electron transfer chain (Balk & Pilon, 2011). An iron influx transporter has not
been characterized, but it is proposed that AtSTA1 and AtSTA2 are involved in Fe-S cluster
efflux from the organelle. AtSTA1 belongs to a subfamily of Arabidopsis half-ABC
transporters, is localized in the inner membrane and its ABC domains face the mitochondrial
matrix. Chlorotic phenotype and stunted growth tendency were observed in sta1 mutants
(Kushnir et al., 2001). A defect in the maturation of Fe-S proteins seems to be related to this
phenotype (Kispal et al., 1999).
Proteome analysis of mitochondria revealed the presence of a ferric chelate reductase
AtFRO8 (Heazlewood et al., 2004). This could be an indication that the reduction strategy
observed in chloroplast is also present in mitochondria.
4. Storage
Iron is an essential metal, required for several metabolic processes both in chloroplast
and mitochondria. Iron is also stored in these organeles, as well as in the vacuole. As part of
iron homeostasis in the whole plant, iron levels and compartmentalization are tightly
regulated. Sequestration and chelation strategies are used by plants to prevent suffering from
toxic effects of iron.
Once loaded to the vacuoles, molecules containing iron are stored complexed to
globoids. The presence of globoids was directly linked to levels of iron in vacuoles (Lanquar
et al., 2005). Both NA and phytates (PA) are molecules that can complex iron in globoids. If
indeed iron is transported to or from the vacuole complexed with NA, members of the YSL
family could be involved in the transport. In a proteomic analysis of isolated vacuoles of A.
thaliana, AtYSL4 and AtYSL6 were identified as being present in the tonoplast (Jaquinod et
al., 2007). Iron-phytate globoids are common in vacuoles due to a high binding capacity
between PA-O-Fe. Each molecule of PA is capable of binding 2 to 4 Fe3+
ions (Bohn et al.,
2008).
Different functions have been assigned to ferritin, depending on the species analyzed.
In peas, ferritin seems to be the most important iron storage site in seeds, where it releases and
provides the metal to the iron-containing proteins after germination (Becker et al., 1998).

30
However, no more than 5% of the iron present in Arabidopsis seeds is estimated to be stored
in ferritins (Ravet et al., 2009). It would seem that in Arabidopsis the protein’s most
significant feature would be related to oxidative stress prevention.
Ferritins are found in chloroplasts, mitochondria and cell walls (Becker et al., 1998).
They are composed of 24 subunits that may allocate from 2,000 to 4,500 Fe3+
atoms per
protein. Iron is oxidized by the ferroxidase centre of the protein before being stored as Fe3+
inside the mineral core. Not only the protein prevents iron from reacting with compounds that
might generate ROS, it actually consumes oxygen and hydrogen peroxide during the
oxidation reaction (Arosio et al., 2009). Ferritins provide bio-available iron inside the cell and
have yet some potential detoxification properties.
Unlike ferritin, frataxin (Fh) is found solely in mitochondria. Since it is strongly
conserved, it is proposed that it should have similar roles in different organisms (Ramirez et
al., 2011). Putative functions for this protein include assisting in Fe-S cluster assembly (Chen
et al., 2002) and involvement in energy conversion and oxidative phosphorylation (Ristow et
al., 2000). The protein appears to have an essential role in seed development, as it was
observed that Arabidopsis knockout mutants (atfh-2 and atfh-3) have an embryo lethal
phenotype. A protective role against oxidative damage was proposed for frataxin as the
mutants showed increased content of ROS and higher levels of transcripts of proteins known
to be involved in oxidative stress responses (Busi et al., 2006). The frataxin from the
monocots Triticum aestivum, Oryza sativa and Zea mays present 77%, 76% and 75%
similarity with AtFH, respectively (Busi et al., 2004).
5. Transcription factors related to iron homeostasis in rice
Two transcription factors were shown to have influence either in sensing cellular iron
status or iron accumulation in rice plants. IDEF1 has a particular characteristic that allows it
to sense iron status and, therefore, positively regulate most genes involved in iron uptake or
utilization. It has His-Asp repeats and Pro-rich regions, known to bind Fe2+
. When these
metal-binding regions are deleted from IDEF1, the plant fails to have a normal response to
iron-deficiency and early iron-deficiency genes regulated by the transcription factor are not
activated (Kobayashi et al., 2012).
OsARF12, also a transcription factor, has an influence in iron homeostasis in rice (Qi
et al., 2012). Knockout plants showed lower concentrations of iron in leaves, roots and seeds

31
when compared to WT plants. It was also observed in these plants an alteration in the
abundance of mitochondrial iron-regulated (OsMIR), iron-regulated transporter (OsIRT) and
short postembryonic root1 (OsSPR1) transcripts. OsMIR, which encodes a mitochondrial
protein involved in iron homeostasis, has its transcription increased both in roots and shoots
under iron deficiency conditions. In the lack of OsARF12, OsMIR was up-regulated and
OsSPR1 down-regulated. OsIRT1 and OsIRT2 transcripts were lower in roots when compared
to WT. However, transcript levels were increased in leaves. Despite having different spatial
expression, both roots and shoots showed lower iron concentrations than WT (Ishimaru et al.,
2009).
Unlike the transcription factors described above, OsWRKY80 is induced by iron excess
instead of iron deficiency conditions (Ricachenevsky et al., 2010). The increase of transcripts
is found throughout the plant, indicating a systemic response to the stress. OsWRKY80 also
responds to other stresses, such as drought and dark-induced senescence. This particular
transcription factor is a member of the WRKY family, which is mostly, but not exclusively,
found in plants. They are related to several processes such as senescence, plant defense and
response to abiotic stresses (for review, see Rushton et al., 2012).
6. Biofortification
It has been previously described that rice possesses a combination of both iron
absorption strategies. This enables rice to adapt more easily to diverse iron-limiting
conditions. Molecules that are essential for iron chelation and transport within the plant have
been described in rice as well. Among them, NA has been demonstrated to be of great value
concerning rice biofortification. The overexpression of a single NAS gene, OsNAS2,
increased iron concentration in rice grains by four-fold (Johnson et al., 2011). This appears to
be the most successful attempt to increase iron and zinc concentrations in rice grains so far.
7. Conclusion and prospects
Due to conjunct and continuous efforts of several teams worldwide, we are now
unraveling the processes regarding iron homeostasis in plants. Much progress was made
toward understanding physiological and molecular mechanisms underlying the phenomena. A
better understanding on release of PS to the rhizosphere will allow a complete

32
characterization of the iron absorption Strategy II, used by many economically important
crops. Also, research is still needed to understand several mechanisms vital for intracellular
iron transport, where knowledge is still at surface. It is known that chloroplasts and
mitochondria have particular iron requirements. However, little is known about Fe intake by
these organelles, how they interact regarding iron homeostasis and how the metal status is
sensed and the signal distributed to maintain adequate levels within the cell.
Clarifying ferritin and vacuole functions both in vegetative tissues as in seeds provided
important information about iron detoxification and storage. Findings in Arabidopsis were
crucial for understanding the vacuole role as primary iron source in the seeds, enabling
germination. Revealing aspects of signaling in the plant, how it adjusts metal uptake to the
current condition, translocates it throughout the plant and then proceeds to storage will help
fully understand iron homeostasis in plants. Disclosing this information will be of great value
for agriculture and human nutrition.
Acknowledgements
The authors would like to thank Vinícius Waldow for help with figures.

33
References
AROSIO P, INGRASSIA R, CAVADINI P (2009) Ferritins: a family of molecules for iron
storage, antioxidation and more. Biochim Biophys Acta 1790:589–599
BALK J & PILON M (2011) Ancient and essential: the assembly of iron–sulfur clusters in
plants. Trends Plant Sci. 16:218-226
BASHIR K, INOUE H, NAGASAKA S, TAKAHASHI M, NAKANISHI H, MORI S,
NISHIZAWA NK (2006) Cloning and characterization of deoxymugineic acid synthase genes
from graminaceous plants. J Biol Chem. 281:32395–32402
BECKER R, MANTEUFFEL R, NEUMANN D, SCHOLZ G (1998) Excessive iron
accumulation in the pea mutants dgl and brz: subcellular localization of iron and ferritin.
Planta. 207:217–223
BOHN L, MEYER AS, RASMUSSEN SK (2008) Phytate: impact on environment and
human nutrition A challenge for molecular breeding. JZUS-B. 9(3):165-191
BOUZAYEN M, FELIX G, LATCHÉ A, PECH JC, BOLLER T (1991) Iron: an essential
cofactor for the conversion of 1-aminocy-clopropane-1-carboxylic acid to ethylene. Planta.
184:244-247
BRIAT JF & LOBRÉAUX S (1997) Iron transport and storage in plants Trends Plant Sci
Reviews 2:187–193
BRIAT JF, FOBIS-LOISY I, GRIGNON N, LOBREAUX S, PASCAL N, SAVINO G,
THOIRON S, VON WIRÉN N, VAN WUYTSWINKEL O (1995) Cellular and molecular
aspects of iron metabolism in plants. Biol Cell. 84:69–81.
BUSI MV, MALIANDI MV, VALDEZ H, CLEMENTE M, ZABALETA EJ, ARAYA A,
GOMEZ-CASATI DF (2006) Deficiency of Arabidopsis thaliana frataxin alters activity of
mitochondrial Fe–S proteins and induces oxidative stress. Plant J. 48:873–882
BUSI MV, ZABALETA EJ, ARAYA A, GOMEZ-CASATI DF (2004) Functional and
molecular characterization of the frataxin homolog from Arabidopsis thaliana. FEBS Lett.

34
576:141–144
CHEN OS, HEMENWAY S, KAPLAN J (2002) Inhibition of Fe-S cluster biosynthesis
decreases mitochondrial iron export: Evidence that Yfh1p affects Fe-S cluster synthesis. Proc.
Natl. Acad. Sci. USA. 99(19): 12321-12326
CONTE SS & WALKER EL (2011) Transporters contributing to iron trafficking in plants.
Mol. Plant. 4:464-476
CURIE C, CASSIN G, COUNCH D, DIVOL F, HIGUCHI K, LE JEAN M, MISSON J,
SCHIKORA A, CZERNIC P, MARI S (2009) Metal movement within the plant: contribution
of nictotianamine and yellow stripe 1-like transporters. Ann. Bot. 103:1–11
CURIE C, PANAVIENE Z, LOULERGUE C, DELLAPORTA SL, BRIAT JF, WALKER
EL (2001) Maize yellow stripe1 encodes a membrane protein directly involved in Fe(III)
uptake. Nature. 409:346-349
DIDONATO RJ, ROBERTS LA, SANDERSON T, EISLEY RB, WALKER EL (2004),
Arabidopsis Yellow Stripe-Like2 (YSL2): a metal-regulated gene encoding a plasma
membrane transporter of nicotianamine–metal complexes. Plant J. 39:403–414
DURRETT TP, GASSMANN W, ROGERS EE (2007) The FRD3-mediated efflux of citrate
into the root vasculature is necessary for efficient iron translocation. Plant Physiol. 144:197-
205
DUY D, STÜBE R, WANNER G, PHILIPPAR K (2011) The Chloroplast Permease PIC1
regulates plant growth and development by directing homeostasis and transport of iron. Plant
Physiol. 155(4):1709-1722
DUY D, WANNER G, MEDA AR, VON WIREN N, SOLL J, PHILIPPAR K (2007) PIC1,
an ancient permease in Arabidopsis chloroplasts, mediates iron transport. Plant Cell. 19:986–
1006
EIDE D, BRODERIUS M, FETT J, GUERINOT ML (1996) A novel iron-regulated metal
transporter from plants identified by functional expression in yeast. PROC. NATL. ACAD.
SCI. USA.. 93:5624-5628
GOMOLPLITINANT KM & SAIER MH JÚNIOR (2011) Evolution of the oligopeptide

35
transporter family. J Membr Biol. 240(2):89–110
GREEN LS & ROGERS EE (2004) FRD3 controls iron localization in Arabidopsis. Plant
Physiol. 136:2523-2531
GROSS J, STEIN RJ, FETT-NETO AG, FETT JP (2003) Iron homeostasis related genes in
rice. Genet. Mol. Biol. 26 (4) 477-497.
GROTZ N & GUERINOT ML (2006) Molecular aspects of Cu, Fe and Zn homeostasis in
plants. Biochim Biophys Acta. 1763:595-608
HAYDON M J & COBBETT C S (2007) Transporters of ligands for essential metal ions in
plants. New Phytol. 174:499–506
HEAZLEWOOD JL, TONTI-FILIPPINI JS, GOUT AM, DAY DA, WHELAN J, MILLAR
AH (2004) Experimental analysis of the Arabidopsis mitochondrial proteome highlights
signaling and regulatory components, provides assessment of targeting prediction programs,
and indicates plant-specific mitochondrial proteins. Plant Cell. 16:241–256
HELL R & STEPHAN UD (2003) Iron uptake, trafficking and homeostasis in plants. Planta.
216:541–551
HIGUCH K, WATANABE S, TAKAHASHI M, KAWASAKI S, NAKANISHI H,
NISHIZAWA NK, MORI S (2001) Nicotianamine synthase gene expression differs in barley
and rice under Fe-deficient conditions. Plant J. 25:159-167
INOUE H, KOBAYASHI T, NOZOYE T, TAKAHASHI M, KAKEI Y, SUZUKI K,
NAKAZONO M, NAKANISHI H, MORI S, NISHIZAWA NK (2009) Rice OsYSL15 is an
iron-regulated iron(III)-deoxymugineic acid transporter expressed in the roots and is essential
for iron uptake in early growth of the seedlings. J Biol Chem. 284:3470–3479
ISHIMARU Y, BASHIR K, FUJIMOTO M, AN G, ITAI RN, TSUTSUMI N, NAKANISHI
H, NISHIZAWA NK (2009) Rice-specific mitochondrial iron-regulated gene (MIR) plays an
important role in iron homeostasis. Mol Plant. 2:1059–1066
ISHIMARU Y, MASUDA H, BASHIR K, INOUE H, TSUKAMOTO T, TAKAHASHI M,
NAKANISHI H, AOKI N, HIROSE T, OHSUGI R, NISHIZAWA NK (2010) Rice metal-
nicotianamine transporter, OsYSL2, is required for the long-distance transport of iron and

36
manganese. Plant J. 62:379–390
ISHIMARU Y, SUZUKI M, TSUKAMOTO T, SUZUKI K, NAKAZONO M, KOBAYASHI
T, WADA Y, WATANABE S, MATSUHASHI S, TAKAHASHI M, NAKANISHI H, MORI
S, NISHIZAWA NK (2006) Rice plants take up iron as an Fe3+
-phytosiderophore and as Fe2+
.
Plant J. 45:335–346
JAQUINOD M, VILLIERS F, KIEFFER-JAQUINOD S, HUGOUVIEUX V, BRULEY C,
GARIN J, BOURGUIGNON, J (2007) A proteomics dissection of Arabidopsis thaliana
vacuoles isolated from cell culture. Mol Cell Proteomics. 6:394-412
JEONG J, COHU C, KERKEB L, PILN M, CONNOLLY EL, GUERINOT ML (2008)
Chloroplast Fe(III) chelate reductase activity is essential for seedling viability under iron
limiting conditions. PROC. NATL. ACAD. SCI. USA.. 105:10619–10624
JOHNSON AAT, KYRIACOU B, CALLAHAN DL, CARRUTHERS L, STANGOULIS J,
LOMBI E, TESTER M (2011) Constitutive Overexpression of the OsNAS Gene Family
Reveals Single-Gene Strategies for Effective Iron- and Zinc-Biofortification of Rice
Endosperm. PLoS ONE. 6(9):e24476
KIM SA, PUNSHON T, LANZIROTTI A, LI L, ALONZO JM, ECKER JR, KAPLAN J,
GUERINOT ML (2006) Localization of iron in Arabidopsis seed requires the vacuolar
membrane transporter VIT1. Science. 314:1295–1298
KISPAL G, CSERE P, PROHL C, LILL R (1999) The mitochondrial proteins Atm1p and
Nfs1p are essential for biogenesis of cytosolic Fe/S proteins. EMBO J. 18:3981-3989
KOBAYASHI T, ITAI RN, AUNG MS, SENOURA T, NAKANISHI H, NISHIZAWA, NK
(2012) The rice transcription factor IDEF1 directly binds to iron and other divalent metals for
sensing cellular iron status. Plant J. 69:81–91
KOIKE S, INOUE H, MIZUNO D, TAKAHASHI M, NAKANISHI H, MORI S,
NISHIZAWA NK (2004) OsYSL2 is a rice metal–nicotianamine transporter that is regulated
by iron and expressed in the phloem. Plant J. 39:415-424
KORSHUNOVA YO, EIDE D, CLARK WG, GUERINOT ML, PAKRASI HB (1999) The
IRT1 protein from Arabidopsis thaliana is a metal transporter with a broad substrate range.

37
Plant Mol. Biol. 40:37-44
KRUGER C, BERKOWITZ O, STEPHAN UW, HELL R (2002) A metal-binding member of
the late embryogenesis abundant protein family transports iron in the phloem of Ricinus
communis L. J. Biol. Chem. 277:25062–25069
KUSHNIR S, BABIYCHUK E, STOROZHENKO S, DAVEY MW, PAPENBROCK J, DE
RUCKE R, ENGLER G, STEPHAN UW, LANGE H, KISPAL G, LILL R, VAN MONAGU
M (2001) A mutation of the mitochondrial ABC transporter Sta1 leads to dwarfism and
chlorosis in the Arabidopsis mutant starik. Plant Cell. 13:89–100
LANQUAR V, LELIEVRE F, BOLTE S, HAMES C, ALCON C, NEUMANN D,
VANSUYT G, CURIE C, SCHRODER A, KRAMER U, BARBIER-BRYGOO H,
THOMINE S (2005) Mobilization of vacuolar iron by AtNRAMP3 and AtNRAMP4 is
essential for seed germination on low iron. EMBO J. 24(23):4041-51
LEE S & AN G (2009) Over-expression of OsIRT1 leads to increased iron and zinc
accumulations in rice. Plant Cell Environm. 32:408-416
LEE S, CHIECKO JC, KIM SA, WALKER EL, LEE Y, GUERINOT ML, AN G (2009)
Disruption of OsYSL15 leads to iron inefficiency in rice plants. Plant Physiol. 150:786-800
MORI S, NISHIZAWA N, HAYASHI H, CHINO M, YOSHIMURA E, ISHIHARA J (1991)
Why are young rice plants highly susceptible to iron deficiency? Plant Soil. 130:1-2
MORRISSEY J, BAXTER IR, LEE J, LI L, LAHNER B, GROTZ N, KAPLAN J, SALT DE,
GUERINOT ML (2009) The ferroportin metal efflux proteins function in iron and cobalt
homeostasis in Arabidopsis. Plant Cell. 21:3326-3338
MSILINI N, ZAGHDOUDI M, GOVINDACHARY S, LACHAÂL M, OUERGHI Z,
CARPENTIER R (2011) Inhibition of photosynthetic oxygen evolution and electron transfer
from the quinone acceptor QA- to QB by iron deficiency. Photosynth. Res. 107(3):247-56
NAGASAKA S, TAKAHASHI M, NAKANISHI-ITAI R, BASHIR K, NAKANISHI H,
MORI S, NISHIZAWA NK (2009) Time course analysis of gene expression over 24 hours in
Fe-deficient barley roots. Plant Mol. Biol. 69:621–631
QI Y, WANG S, SHEN C, ZHANG S, CHEN Y, XU Y, LIU Y, WU Y, JIANG D (2012)

38
OsARF12, a transcription activator on auxin response gene, regulates root elongation and
affects iron accumulation in rice (Oryza sativa). New Phytol. 193:109–120
RAMIREZ L, SIMONTACCHI M, MURGIA I, ZABALETA E, LAMATTIN E (2011)
Nitric oxide, nitrosyl iron complexes, ferritin and frataxin:A well equipped team to preserve
plant iron homeostasis. Plant Sci. 181 (5):582–592
RAVET K, TOURAINE B, KIM SA, CELLIER F, THMINE S, GUERINOT ML, BRIAT J-
F, GAYMARD F (2009) Post-translational regulation of AtFER2 ferritin in response to
intracellular iron trafficking during fruit development in Arabidopsis. Mol. Plant. 2:1095–
1106
RICACHENEVSKY FK, SPEROTTO RA, MENGUER PK, FETT JP (2010) Identification
of Fe-excess-induced genes in rice shoots reveals a WRKY transcription factor responsive to
Fe, drought and senescence. Mol. Biol. Rep. 37:3735–3745.
RISTOW M, PFISTER MF, YEE AJ, SCHUBERT M, MICHAEL L, ZHANG CY, UEKI K,
MICHAEL MD, LOWELL BB, KAHN CR (2000) Frataxin activates mitochondrial energy
conversion and oxidative phosphorylation. PROC. NATL. ACAD. SCI. USA.. 97:12239–
12243
ROBINSON NJ, PROCTER CM, CONNOLLY EL, GUERINOT ML (1999) A ferric-chelate
reductase for iron uptake from soils. Nature. 397:694-697
RUSHTON DL, TRIPATHI P, RABARA RC, LIN J, RINGLER P, BOKEN AK, LANGUM
TJ, SMIDT L, BOOMSMA DD, EMME NJ, CHEN X, FINER JJ, SHEN QJ, RUSHTON PJ
(2012) WRKY transcription factors: key components in abscisic acid signaling. Plant Biotech.
J. 10:2–11.
SANTI S, CESCO S, VARANINI Z, PINTON R (2005) Two plasma membrane H+-ATPase
genes are differentially expressed in iron-deficient cucumber plants. Plant Physiol. Bioch.
43(3):287-292
SCHAAF G, HONSBEIN A, MEDA AR, KIRCHNER S, WIPF D, VON WIREN N (2006)
AtIREG2 encodes a tonoplast transport protein involved in iron-dependent nickel
detoxification in Arabidopsis thaliana roots. J. Biol. Chem. 281:25532–25540

39
SCHAAF G, SCHIKORA A, HÄBERLE J, VERT G, LUDEWIG U, BRIAT JF, CURIE C,
VON WIRÉN N (2005) A putative function for the arabidopsis Fe-Phytosiderophore
transporter homolog AtYSL2 in Fe and Zn homeostasis. Plant Cell Physiol. 46:762-774
SHIKANAI T, MÜLLER-MOULÉ P, MUNEKAGE Y, NIYOGI KK, PILON M (2003)
PAA1, a P-type ATPase of Arabidopsis, functions in copper transport in chloroplasts. Plant
Cell. 15:1333–1346
SIEDOW JN (1991) Plant lipoxygenase:structure and function. Annu. Rev. Plant Physiol.
Plant Mol. Biol. 42, 145-188
SOLTI A, KOVÁCS K, BASA B, VÉRTES A, SÁRVÁRI É, FODOR F (2012) Uptake and
incorporation of iron in sugar beet chloroplasts. Plant Physiol. Biochem. 52:91-97
SPEROTTO RA, BOFF T, DUARTE GL, SANTOS LS, GRUSAK MA, FETT JP (2010)
Identification of putative target genes to manipulate Fe and Zn concentrations in rice grains. J.
Plant Physiol. 167 (17):1500-1506
STACEY MG, PATEL A, MCCLAIN WE, MATHIEU M, REMLEY M, ROGERS EE,
GASSMANN W, BLEVINS DG, STACEY G (2008) The Arabidopsis AtOPT3 protein
functions in metal homeostasis and movement of iron to developing seeds. Plant Physiol.
146:589-601
UENO D, ROMBOLÀ AD, IWASHITA T, NOMOTO K, MA JF. (2007) Identification of
two novel phytosiderophores secreted by perennial grasses. New Phytol. 174: 304–310
VAROTTO C, MAIWALD D, PESARESI P, JAHNS P, SALAMINI F, LEISTER D (2002)
The metal ion transporter IRT1 is necessary for iron homeostasis and efficient photosynthesis
in Arabidopsis thaliana. Plant J. 31:589-599
VASCONCELOS M, ECKERT H, ARAHANA V, GRAEF G, GRUSAK MA, CLEMENTE
T (2006) Molecular and phenotypic characterization of transgenic soybean expressing the
Arabidopsis ferric chelate reductase gene, FRO2. Planta. 224(5):1116-1128
WATERS BM, CHU HH, DIDONATO RJ, ROBERTS LA, EISLEY RB, WALKER EL
(2006) Mutations in Arabidopsis Yellow Stripe-Like1 and Yellow Stripe-Like3 reveal their
roles in metal ion homeostasis and loading of metal ions in seeds. Plant Physiol. 141:1446–

40
1458
YOKOSHO K, YAMAJI N, UENO D, MITANI N, MA JF (2009) OsFRDL1 is a citrate
transporter required for efficient translocation of iron in rice. Plant Physiol. 149:297-305

41
Figures
Figure 1
Figure 1: Strategies by which plants absorb iron from soil in iron deficiency conditions. a.
Reduction strategy used by non-graminaceous plants. Protons are expelled by the H+-ATPase.
FRO, a ferric chelate reductase, is responsible for reducing Fe3+
to Fe2+
which is then
transported inside the plant by the divalent metal transporter IRT. b. Chelation strategy, used
by grasses, where phytosiderophores (PSs) are secreted by roots. These molecules easily
chelate Fe3+
present in the soil, and the complexed molecule can be transported by Yellow
Stripe (YS) proteins into the root. c. Combined strategy present in rice. Both acidification
system and chelation by a PS are present in rice roots in iron limited environments.
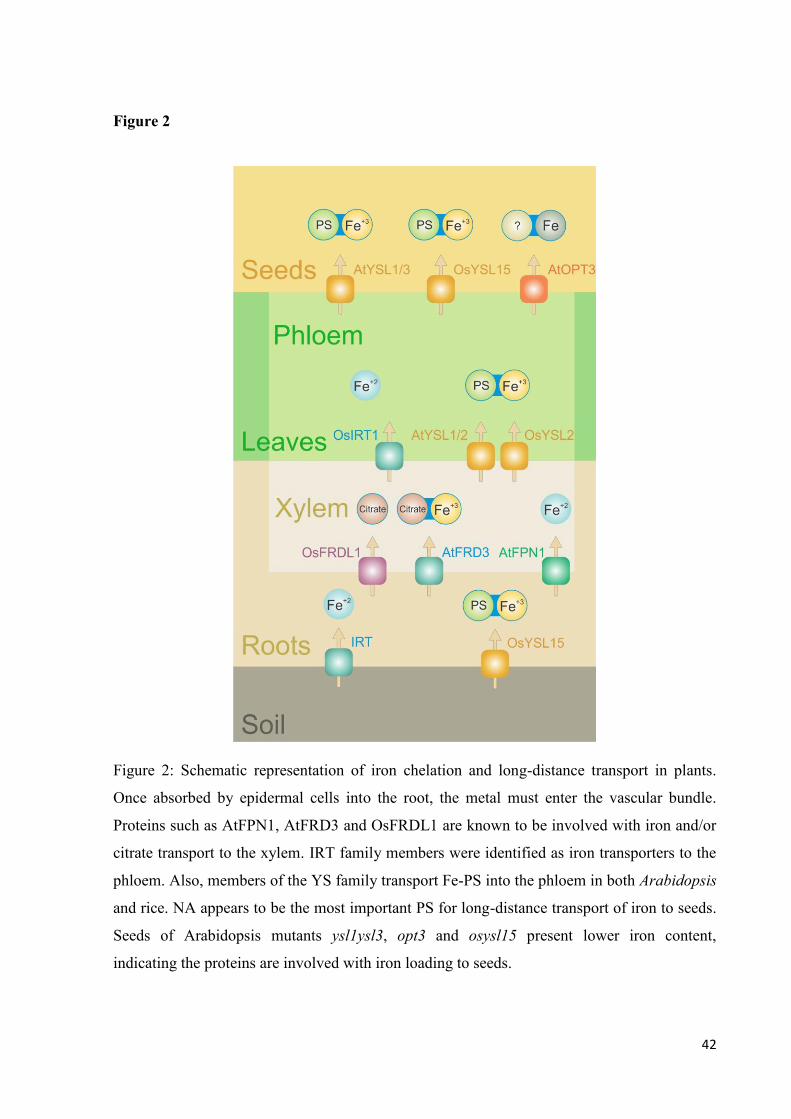
42
Figure 2
Figure 2: Schematic representation of iron chelation and long-distance transport in plants.
Once absorbed by epidermal cells into the root, the metal must enter the vascular bundle.
Proteins such as AtFPN1, AtFRD3 and OsFRDL1 are known to be involved with iron and/or
citrate transport to the xylem. IRT family members were identified as iron transporters to the
phloem. Also, members of the YS family transport Fe-PS into the phloem in both Arabidopsis
and rice. NA appears to be the most important PS for long-distance transport of iron to seeds.
Seeds of Arabidopsis mutants ysl1ysl3, opt3 and osysl15 present lower iron content,
indicating the proteins are involved with iron loading to seeds.
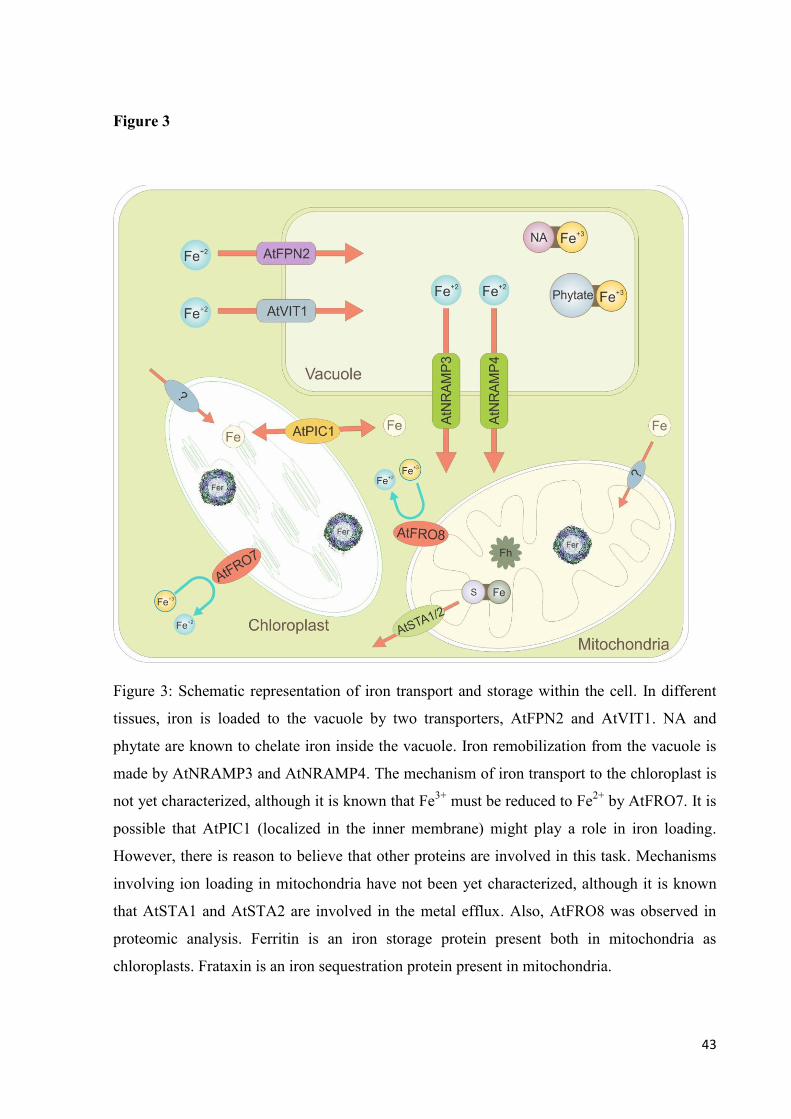
43
Figure 3
Figure 3: Schematic representation of iron transport and storage within the cell. In different
tissues, iron is loaded to the vacuole by two transporters, AtFPN2 and AtVIT1. NA and
phytate are known to chelate iron inside the vacuole. Iron remobilization from the vacuole is
made by AtNRAMP3 and AtNRAMP4. The mechanism of iron transport to the chloroplast is
not yet characterized, although it is known that Fe3+
must be reduced to Fe2+
by AtFRO7. It is
possible that AtPIC1 (localized in the inner membrane) might play a role in iron loading.
However, there is reason to believe that other proteins are involved in this task. Mechanisms
involving ion loading in mitochondria have not been yet characterized, although it is known
that AtSTA1 and AtSTA2 are involved in the metal efflux. Also, AtFRO8 was observed in
proteomic analysis. Ferritin is an iron storage protein present both in mitochondria as
chloroplasts. Frataxin is an iron sequestration protein present in mitochondria.

44
Capítulo 2
Iron Transport by the Rice OsNRAMP7 Protein
Artigo submetido como Short Communication ao Journal of Molecular Biology

45
Iron Transport by the Rice OsNRAMP7 Protein
Lívia Scheunemann dos Santos1, Anthony John Miller
2, Janette Palma Fett
1,3§.
1 Departamento de Botânica, Instituto de Biociências, Universidade Federal do Rio Grande do
Sul, 91501-970, Porto Alegre, RS, Brazil.
2 Department of Disease and Stress Biology, John Innes Centre, Norwich Research Park, NR4
7UH, UK.
3 Centro de Biotecnologia, Universidade Federal do Rio Grande do Sul, 91501-970 Porto
Alegre, RS, Brazil.
§Corresponding author
E-mail addresses:
LSS: [email protected]
AJM: [email protected]
JPF: [email protected]
Corresponding author
Centro de Biotecnologia, Universidade Federal do Rio Grande do Sul, P.O. Box 15005,
91501-970 Porto Alegre, RS, Brazil. Tel.: +55 51 3308 7643; Fax: +55 51 3308 7309.

46
Abstract
Proteins from the NRAMP family from diverse organisms are known to transport a wide
range of metals, playing important roles in cellular metal homeostasis. Some members of the
family have been described as particularly participating in plant iron homeostasis. Of special
interest in rice plants, the OsNRAMP7 protein was investigated in this work. Characteristic
features, motifs DPGN and MPH, said to be involved in metal transport, were identified in the
protein structure. To investigate the involvement of the rice OsNRAMP7 protein in iron
transport, a complete version of the protein as well as a truncated form missing the first 15
amino acids, were expressed in Xenopus oocytes. These amino acids were indicated, through
in silico analysis, as a putative endomembrane targeting signal. Only oocytes injected with the
complete form of the protein exhibited a significant increase in iron content, in two iron
concentrations tested, demonstrating the protein’s ability to perform transmembrane iron
transport.
Keywords: Iron, metal transport, NRAMP, Oryza sativa L., rice, Xenopus laevis.

47
Background
Metals are essential for the maintenance of living cells. They are required for many
biological processes such as gene regulation and free-radical homeostasis, catalyzing electron
transfer in mitochondria and chloroplast or acting as co-factors for enzymes. Since
accumulation of high amounts of metals might lead to toxic effects, metal homeostasis must
be tightly regulated. It has been shown that the excess of a metal such as iron, in its free form,
may lead to increased generation of reactive oxygen species (ROS) through the Fenton
reaction1, and that this iron-mediated oxidative stress can cause damage to several biological
macromolecules such as lipids, proteins and nucleic acids, being ultimately deleterious to cell
integrity2.
The Natural-Resistance-Associated Macrophage Protein (NRAMP) gene family was
shown to play important roles in metal homeostasis. Its importance was demonstrated in
several organisms, from bacteria to human3. Protein members of this family have a broad
spectrum of specificity in metal transport including Fe, Mn, Co, Zn, Cu, Cd, Ni4 and V
5.
Heterologous expression in yeast and molecular physiological studies in plants are some of
the experiments that have characterized the family6, 7, 8
, providing information on which
metals are transported and how the family is linked to metal homeostasis.
Techniques which allow high-resolution of the structures of transporters shed light on
the mechanisms of ion transport, since conformational changes are part of the transport
mechanism9. However, typical eukaryotic cells have a large number of transporters in the
plasma membrane, and studying the transport mechanism of a single specific transporter is a
hard task. Thus, important information about transporter proteins has come from studies of
metal transporters expressed in Xenopus laevis oocytes6, 10, 11, 12, 13
. One of the advantages of
this method is that there is little background transport activity to interfere with the
characteristics of the foreign protein and the large cell size facilitates the analysis14
.

48
Our group identified eight NRAMP gene family members in the rice genome15
. In a
recent work, we found significant negative correlation between OsNRAMP7
(LOC_Os12g39180) expression in flag leaves during grain filling and final Fe and Zn
concentrations in the grain16
. The influence of OsNRAMP7 in metal allocation to the grain
could also be supported by the co-localization of the gene with a QTL that explains around
14% of the phenotypic variation in Fe 17
and Zn concentration18
in the grain.
Based on these previous findings, we hypothesized that the OsNRAMP7 protein is
able to perform iron transport in rice plants, and in the present study we further investigated
this possibility.
Results and Discussion
An in silico analysis of the OsNRAMP7 protein was conducted to help investigate
some of its properties and its anchorage in the membrane. The topology algorithm HMMTOP
(http://www.enzim.hu/hmmtop/) predicted the existence of twelve significant putative
transmembrane domains (TMDs) (Fig. 1a), consistent with findings on other members of the
family, which show from 10 to 12 TMDs3. Conserved regions, such as the signature sequence
of the NRAMP family, DPGN (Asp-Pro-Gly-Asn) and the MPH motif (Met-Pro-His)19,20
are
also present in OsNRAMP7. When analyzing the protein for the presence of a transport motif
to define NRAMP family membership, OsNRAMP7 shows the Consensus Transport Motif
(CTM)19,20
at the hydrophilic loop between TMDs 7 and 8 (Fig. 1). This conserved motif is a
feature present in all members of the NRAMP family in rice (Supplementary Figure 1). The
rice NRAMP7 protein shares important features with the isoform I of the mammalian DCT13,
the first member of the NRAMP family to have its role in metal homeostasis described10
: both
proteins share the same number of TMDs (12) and the same number of amino acids (18) at the
C-terminus domain located after TMD12 (Fig. 1a).

49
A three-dimensional representation of the protein was constructed (Fig. 1b-f) where all
three conserved regions are in evidence (Fig. 1c-f). It is possible to observe that that the CTM
is present in the protein’s widest loop (Fig. 1c) and adjacent to a large cavity (Fig. 1c-d).
Also, among the cavities present, that seems to be the largest. This could be evidence of a
structural property of this feature. Adjacent to this structure are the DPGN and MPH motifs
(Fig. 1e-f). Structures as the DPGN site are likely to be involved in divalent metal (Me+2
)
uptake, since it bears analogy to a known functional signature for Me+2
transport21
. In E. coli
MntHAsp mutants, it was shown that the DPGN site is also implicated in Me+2
binding and
coupling of Me+2
transport to proton-motive force22
. The MPH motif participates in pH-
dependent regulation by deprotonation, which could favor a conformation that enables Me+2
transport22
.
The localization and orientation of the DPGN and MPH motifs (seen in Fig. 1f) are
shared by several cation transporter families23
. Another important feature involving these
motifs is the formation of a pair of extended peptides by their anti-parallel orientation,
interrupting the TMDs. This characteristic is present in transporter structures, and the
presence of discontinuous TM helices with extended peptides seems to be directly correlated
with the ion transport reaction22, 24
.
OsNRAMP7 expression was observed in all rice organs evaluated. When analyzing
hydroponically grown rice plants, expression in leaves was almost eight times higher of what
was found in roots (Fig. 2a). In field grown plants, organs involved in grain filling (panicles
and flag leaves) presented higher expression of OsNRAMP7 then vegetative organs (stem and
non-flag leaves) at the grain filling stage (Fig. 2b). Previous experiments revealed that
OsNRAMP7 expression was not altered by plant treatments leading to Fe excess or Fe, Zn or
Mn deficiency, both in roots and shoots (Ricardo Stein and Marta Spohr, personal
communication).

50
The protein sequence of OsNRAMP7 has high identity to Arabidopsis thaliana’s
AtNRAMP3 and AtNRAMP4 (68%, Supplementary Figure 2a). All three proteins also belong
to the Group II in a phylogenetic analysis (Supplementary Figure 2b). Such similarity is
relevant since these proteins were demonstrated to be involved in Fe transport by
complementing a mutant strain of yeast defective in Fe uptake7 and, also, by mediating iron
transport in Arabidopsis, being involved in iron efflux from vacuoles during germination8.
AtNRAMP3’s subcellular localization on the vacuolar membrane25
suggests a function in
intracellular metal homeostasis. Other more distantly related NRAMP proteins have been
shown to transport Al26
. Although recent publications presented phylogenetic trees of the rice
NRAMPs27, 28
, none were complete, not including all members of the family.
To test the ability of the OsNRAMP7 protein to mediate transmembrane iron
transport, we expressed the protein in Xenopus oocytes. Before starting the experiments, we
investigated the OsNRAMP7 protein sequence for the presence of a possible targeting signal
peptide cleavage site using the SignalP 3.0 Server (http://www.cbs.dtu.dk/services/SignalP/).
The presence of a targeting sequence could impair targeting of the protein to the plasma
membrane in heterologous expression systems like Xenopus oocytes. SignalP described a
possible targeting signal peptide, with a maximum cleavage site probability between amino
acids 16 and 17. Targeting sequences were removed in previous reports, as for expression of
chloroplastic membrane proteins (e.g. Mariscal et al.29
; Maughan et al.30
). For this reason, not
only the full length cDNA of OsNRAMP7 was used for oocyte expression, but also the N-
terminal truncated form, missing the first 15 amino acids of the protein.
A barley aquaporin mRNA was used as a control in order to test for the quality of the
injections and translation of the injected RNA into protein31
. A simple swelling assay can be
used in these positive control experiments. A second control group, of oocytes injected with

51
water, was also used. Therefore, five groups of oocytes were analyzed: uninjected oocytes,
oocytes injected with water, aquaporin, OsNRAMP7 and the truncated form of OsNRAMP7.
The optimal timing for protein expression was established by exposing aquaporin mRNA
injected oocytes to swelling assays (where they were exposed to 50% MBS) 3, 4 or 5 days
after injections. The functional aquaporin protein resulted in the oocytes swelling more
rapidly (Fig. 3a). These experiments established that a four day incubation period was optimal
for expression of the aquaporin. Hence, four days after mRNA injections oocytes from all
treatments were exposed to MBS medium containing 50 or 100 µM FeCl3, where they were
kept for 24 hours. These iron concentrations are used in hydroponic solutions for rice growth
experiments26
.
Single cells were examined for the absorption of iron by quantification of total iron
content. A significant increase of iron content in oocytes injected with the full-length form of
OsNRAMP7 was observed (Fig. 3b), while cells that were not injected or injected with water,
aquaporin or the truncated form of OsNRAMP7 did not show a significant increase of iron
content (Fig 3b). The presence of the functional form of the protein OsNRAMP7 resulted in
increased iron uptake.
These results suggest that the cleaved fraction of the OsNRAMP protein may be
essential for function and not necessarily an endomembrane targeting signal. The lack of
function of the truncated form of OsNRAMP7, when compared to the complete form, may be
explained by a possible change in conformational structure of the protein due to the alteration
of the N-terminal portion32, 33
, which could impair its activity. Previous studies showed that,
in certain proteins, alteration by deletion of a fraction of the N-terminal portion might lead to
decreased activity or even, as seen here, to protein loss of function33, 34
.
Conclusions

52
The increased iron content in oocytes upon expression of OsNRAMP7 indicates that
this protein is able to mediate iron transport. Together with its high similarity to other
NRAMP proteins shown to be iron transporters and its high expression in reproductive
organs, the presumed function of OsNRAMP7 in iron homeostasis during grain filling is
confirmed. The protein could have a role in the internalization of metals in vacuoles, resulting
in lower metal availability to be transported to panicles. Therefore, it is possible to confirm an
active role of OsNRAMP7 in iron homeostasis. Further studies, such as sub-cellular
localization through Green Fluorescent Protein (GFP) fusion and complementation of yeast
strains defective in metal transport, are being conducted in order to better understand the role
of this transporter in metal homeostasis in rice plants.
Materials and Methods
For the analysis of panicles, stem, flag leaf and other leaves, rice plants (Oryza sativa
L. cv. Nipponbare) were field-grown as described16
up to the reproductive stage 5 (R5)
according to the classification established by Counce et al35
. For analysis of OsNRAMP7
expression in roots and leaves, seeds were germinated for 4 days in an incubator (28° C, first
2 days in the dark and last 2 days in the light) on paper soaked with distilled water. After
germination and growth in vermiculite and nutrient solution (0.1 mM KCl, 0.1 mM KH2PO4,
0.7 mM K2SO4, 2 mM Ca(NO3)2 4 H2O, 0.5 mM MgSO4 7 H2O, 0.5 µM MnSO4 4 H2O,
0.01 µM (NH4)6 Mo7 O24 4 H2O, 10 µM H3BO3, 0.5 µM ZnSO4 7 H2O, 0.2 µM CuSO4 5
H2O, 100 µM FeSO4 7 H2O, 100 µM EDTA) for 14 days (28° C, with 16 h of light), plants
were transferred to pots containing 2.5 L of nutrient solution. All solutions were replaced
every 3 days. Plants were cultivated in a growth room at 26 ± 1° C under white light with a
photoperiod of 16/8 h light/dark cycle (irradiance of approximately 100 µmol m–2
s–1
at the

53
plant tops) for another period of 14 days, when analyses were conducted. RNA extraction was
performed with pooled material from three plants in each replicate.
Total RNA was extracted using Concert® (Invitrogen Life Technologies) reagent.
First-strand cDNA synthesis was performed after DNAse treatment with reverse transcriptase
(M-MLV®, Invitrogen Life Technologies) using 1 g of RNA, quantified using Qubit
®
(Invitrogen Life Technologies). An Applied Biosystem 7500 real-time cycler was used to
carry out the qRT-PCR analysis. A 240 bp region was amplified using gene-specific primers
(5’-GCTGCCAAATCAGATCATCA-3’ e 5’-GCTTCAGGACGACACAGTCA-3’). Reaction
settings were performed according to Sperotto et al36
. Gene expression was evaluated by the
2−ΔCT
method37, 38
. Each data point corresponds to three biological replicates, which were
analyzed in four technical replications. A ΔCT value was obtained by subtracting the
Ubiquitin CT value from the CT obtained for the gene of interest. When appropriate, data were
subjected to analyses of variance (ANOVA) and means were compared by the Duncan test
(P≤0.05) using the SPSS Base 17.0 for Windows (SPSS Inc., USA).
Three-dimensional structure of OsNRAMP7 was built using homology modeling,
since its crystal structure is not available. The protein sequence (Accession No.
NP_001067135, length: 541 amino acid) was retrieved from NCBI GenBank. A 3D model
based on multiple-threading alignments by LOMETS and iterative TASSER assembly
simulations was built at the I-TASSER webserver39
. The comparison between template and
target sequence resulted in C-score equal to -3.74, TM-score of 0.39±0.13 and RMSD of
14.5±3.7Å. The pre-refined model was examined under the Ramachandran plot and the
residue properties plot was evaluated using the PROCHECK server40
. Final adjustments and
molecular visualization were performed using PyMOL (http://www.pymol.org).
Two forms of OsNRAMP7 cDNA were produced by PCR, one corresponding to the
full coding region of OsNRAMP7 cDNA and one where a putative signal peptide present on

54
the N-Terminal portion of the protein was removed. The full coding region was amplified
using two specific primers (Forward 1, 5’- AATAGATCTACCCCCATCTCCAAATC-3’;
Reverse, 5’- AATGGTACCTGAAGCGTATAGTTGTG-3’). The amplification product of
the truncated version of OsNRAMP7 was obtained using a different forward primer (Forward
2, 5’- AGATCTATGGCCACCCCCGCC-3’), while maintaining the same Reverse primer. A
BglII restriction site was added at the 5’ end of both forward primers. The resulting PCR
product was cloned into the Xenopus laevis oocyte expression vector pT7TS’ BglII and
EcoRV sites. Plasmid DNA was digested with EcoRI, and mRNA was produced with the
Ambion In Vitro Transcription kit (mMessage mMachine Ultra®) following the
manufacturer’s instructions. Oocytes were prepared as previously described41
. A 50 ng
aliquot of mRNA in 50 nl of water was injected into stage V and VI oocytes obtained from X.
laevis. The oocytes were incubated in Modified Barth’s Saline (MBS) (8.8 mM NaCl, 1 mM
KCl, 2.4 mM NaHCO3, 10 mM HEPES, CaCl·2H2O) at 20° C for 4 days. Swelling tests were
carried out by transferring to 0.82 mM MgSO4·7H2O, 0.33 mM Ca(NO3)2·4H2O, 0.41 mM
CaCl·2H2O at 20°C for 4 days.
Swelling tests were carried out by transferring oocytes to 50% MBS (MBS 1:1 water),
and time lapse images were taken every five minutes with a digital camera attached to a
dissection microscope31
. The swelling rate was established by comparing the mean diameter
measure of six independent oocytes, for four times each, during a twenty minute interval. Iron
uptake assays were performed on oocytes four days after the mRNA injections, when they
were incubated in MBS solution containing 50 or 100 µM FeCl3, added of Ascorbic Acid in
an equimolar concentration to maintain the Fe in an available form5 or maintained in the MBS
solution without the addition of iron. Oocytes were kept in these solutions for 24 hours before
uptake analysis. After the incubation period the oocytes were washed in DCB solution (0.03
M Na3C6H5O7·2H2O, 0.125 M NaHCO3 and 0.06 M Na2S2O442
), prepared in MBS for 10

55
minutes under slight agitation to remove any iron that might have been deposited on the
plasma membrane. The colorimetric ferrozine-based assay for Fe2+
quantification was
performed as described43
. Measurements were conducted in 10 single oocytes for each
treatment. Data was subjected to statistical analysis using Student’s t test (P = 0.05) on the
SPSS Base 12.0 for Windows (SPSS Inc., USA).
Funding
This work was supported by CNPq (Conselho Nacional de Desenvolvimento Científico
e Tecnológico), Brazil, and CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível
Superior), Brazil. The John Innes Centre receives grant-aided support from Biotechnology
and Biological Sciences Research Council (BBSRC) of the United Kingdom.
Authors’ contributions
JPF conceived the work. LSS and JPF performed initial studies. LSS performed
sequence analyses, RT-qPCR assays, DNA constructions and 3D modeling. LSS and AJM
performed the experimental procedures with Xenopus oocytes and analyzed the data. LSS
drafted the original manuscript. JPF was responsible for the final editing. All authors read and
approved the final manuscript.
Acknowledgements
We thank the Gurdon Institute (Cambridge, UK) for kindly providing the oocytes,
Juliana Reis for assistance with the figures, Paloma Koprovisky Menguer for valuable
discussions and Raul Antonio Sperotto for providing the RNAs used for RT-qPCR.

56
References
1. Hell, R. & Stephan, U.D. (2003) Iron uptake, trafficking and homeostasis in plants. Planta.
216: 541–551.
2. Briat, J.-F., Duc, C., Ravet, K., Gaymard, F. (2010) Ferritins and iron storage in plants.
Biochim. Biophys. Acta - General Subjects. 1800 (8): 806-814.
3. Nevo, Y. & Nelson, N. (2006) The NRAMP family of metal-ion transporters. Biochim.
Biophys. Acta - Molecular Cell Research. 1763 (7): 609-620.
4. Ňuňuková, V., Urbánková, E., Jelokhani-Niaraki, M., Chaloupka, R. (2010) Ion channel
activity of transmembrane segment 6 of Escherichia coli proton-dependent manganese
transporter. Biopolymers. 93: 718–726.
5. Ueki, T., Furunob, N., Michibata, H. (2011) A novel vanadium transporter of the Nramp
family expressed at the vacuole of vanadium-accumulating cells of the ascidian Ascidia
sydneiensis samea. Biochim. Biophys. Acta - General Subjects. 1810 (4): 457-464.
6. Chen, X.Z., Peng, J.B., Cohen, A., Nelson, H., Nelson, N., Hediger, M.A. (1999) Yeast
SMF1 mediates H+-coupled iron uptake with concomitant uncoupled cation currents. J Biol.
Chem. 274: 35089–35094.
7. Thomine, S., Wang, R.C., Ward, J.M., Crawford, N.M., Schroeder, J.I. (2000) Cadmium
and iron transport by members of a plant metal transporter family in Arabidopsis with
homology to Nramp genes. P. Natl. Acad. Sci. USA. 97: 4991-4996.
8. Lanquar, V., Ramos, M.S., Lelièvre, F., Barbier-Brygoo, H., Krieger-Liszkay, A., Krämer,
U., Thomine, S. (2005) Mobilization of vacuolar iron by AtNRAMP3 and AtNRAMP4 is
essential for seed germination on low iron. EMBO J. 24 (23): 4041-51.

57
9. Abramson, J., Smirnova, I., Kasho, V., Verner, G., Iwata, S., Kaback, H.R. (2003) The
lactose permease of Escherichia coli: Overall structure, the sugar-binding site and the
alternating access model for transport. FEBS Letters. 555 (1): 96-101.
10. Gunshin, H., Mackenzie, B., Berger, U. V., Gunshin, Y., Romero, M. F., Boron, W. F.,
Nussberger, S., Gollan, J. L., Hediger, M. A. (1997) Cloning and characterization of
mammalian proton-coupled metal-ion transporter. Nature. 388: 482–488.
11. Ueki, T., Uyama, T., Kanamori, K., Michibata, H. (2001) Subunit C of the vacuolar-type
ATPase from the vanadium-rich ascidian Ascidia sydneiensis samea rescued the pH
sensitivity of yeast vma5 mutants. Mar. Biotechnol. 3: 316–321.
12. Okubo, M., Yamada, K., Hosoyamada, M., Shibasaki, T., Endou, H. (2003) Cadmium
transport by human Nramp 2 expressed in Xenopus laevis oocytes. Toxicol. Appl. Pharmacol.
187: 162–167.
13. Agranoff, D., Collins, L., Kehres, D., Harrison, T., Maguire, M., Krishna, S. (2005) The
Nramp orthologue of Cryptococcus neoformans is a pH-dependent transporter of manganese,
iron, cobalt and nickel. Biochem. J. 385: 225-232.
14. Miller, A.J. & Zhou, J.-J. (2000) Xenopus oocytes as an expression system for plant
transporters. Biochim. Biophys. Acta. 1465: 343-358.
15. Gross, J., Stein, R. J., Fett-Neto, A. G., Fett, J. P. (2003) Iron homeostasis related genes in
rice. Genet. Mol. Biol. 26 (4) 477-497.
16. Sperotto, R.A., Boff, T., Duarte, G.L., Santos, L.S., Grusak, M.A., Fett, J.P. (2010)
Identification of putative target genes to manipulate Fe and Zn concentrations in rice grains. J.
Plant Physiol. 167 (17): 1500-1506.
17. Stangoulis, J.C.R., Huynh, B.L., Welch, R.M., Choi, E.Y., Graham, R.D. (2007)
Quantitative trait loci for phytate in rice grain and their relationship with grain micronutrient
content. Euphytica. 154: 289-294.

58
18. Garcia-Oliveira, A.L., Tan, L., Fu, Y., Sun, C. (2009) Genetic identification of
quantitative trait loci for contents of mineral nutrients in rice grains. J. Integr. Plant Biol. 51:
84-92.
19. Gruenheid, S., Cellier, M., Vidal, S., Gros, P. (1995) Identification and characterization of
a second mouse Nramp gene. Genomics. 25 (2): 514-525.
20. Cellier, M., Privé, G., Belouchi, A., Kwan, T., Rodrigues, V., Chia, W., Gros, P. (1995)
Nramp defines a family of membrane proteins. PROC. NATL. ACAD. SCI. USA.. 92 (22):
10089-10093.
21. Argüello, J. M. (2003) Identification of Ion-Selectivity Determinants in Heavy-Metal
Transport P 1B-type ATPases. J. Membrane Biol. 195 (2): 93-108.
22. Haemig, H.A.H. & Brooker, R.J. (2004) Importance of Conserved Acidic Residues in
MntH, the Nramp homolog of Escherichia coli. J. Membrane Biol. 201: 97–107.
23. Screpani, E. & Hunte, C. (2007) Discontinuous membrane helices in transport proteins
and their correlation with function. J Struct Biol. 159, 261–267
24. Courville, P., Urbankova, E., Rensing, C., Chaloupka, R., Quick, M., Cellier, M.F. (2008)
Solute carrier 11 cation symport requires distinct residues in transmembrane helices 1 and 6. J
Biol Chem. 283: 9651–9658
25. Thomine, S., Lelièvre, F., Debarbieux, E., Schroeder, J.I., Barbier-Brygoo, H. (2003)
AtNRAMP3, a multispecific vacuolar metal transporter involved in plant responses to iron
deficiency. Plant J. 34 (5): 685-95.
26. Kobayashi, T., Suzuki, M., Inoue, H., Itai, R.N., Takahashi, M., Nakanishi, H., Mori, S.,
Nishizawa, N.K. (2005) Expression of iron-acquisition-related genes in iron-deficient rice is
co-ordinately induced by partially conserved iron-deficiency-responsive elements. J. Exp.
Bot. 56 (415):1305-1316.

59
27. Xia, J., Yamaji N., Kasai, T., Ma, J.F. (2010). Plasma membrane-localized transporter for
aluminum in rice. PROC. NATL. ACAD. SCI. USA.. 107 (43): 18381-18385.
28. Takahashi, R., Ishimaru, Y., Senoura, T., Shimo, H., Ishikawa, S., Arao, T., Nakanishi,
H., Nishizawa, N.K. (2011) The OsNRAMP1 iron transporter is involved in Cd accumulation
in rice. J. Exp. Bot. 62 (14): 4843-4850.
29. Mariscal, V., Moulin, P., Orsel, M., Miller, A.J., Fernández, E., Galván, A. (2006).
Differential regulation of the Chlamydomonas Nar1 gene family by carbon and nitrogen.
Protist. 157 (4): 421-433.
30. Maughan SC, Pasternak M, Cairns N, Kiddle G, Brach T, Jarvis R, Haas F, Nieuwland J,
Lim B, Müller C, Salcedo-Sora , E., Kruse, C., Orsel, M., Hell, R., Miller, A.J., Bray, P.,
Foyer, C.H., Murray, J.A.H., Meyer, A.J., Cobbett, C.S. (2010). Plant homologs of the
Plasmodium falciparum chloroquine-resistance transporter, PfCRT, are required for
glutathione homeostasis and stress responses. PROC. NATL. ACAD. SCI. USA.. 107 (5):
2331-2336.
31. Besse, M., Knipfer, T., Miller, A.J., Verdeil, J.-L., Jahn, T.P., Fricke, W. (2011)
Developmental pattern of aquaporin expression in barley (Hordeum vulgare L.) leaves. J.
Exp. Bot. 62 (12): 4127-4142.
32. Sokal, I., Otto-Bruc, A.E., Surgucheva, I., Verlinde, C.L., Wang, C.K., Baehr, W.,
Palczewski, K. (1999) Conformational changes in guanylyl cyclase-activating protein 1
(GCAP1) and its tryptophan mutants as a function of calcium concentration. J. Biol. Chem.
274 (28): 19829–19837.
33. Wang, X.C., Yang, J., Huang, W., He, L., Yu, J.T., Lin, Q.S., Li, W., Zhou, H.M. (2002)
Effects of removal of the N-terminal amino acid residues on the activity and conformation of
firefly luciferase. Int. J. Biochem. Cell Biol. 34 (8): 983-91.

60
34. Li, W., Srinivasula, S.M., Chai, J., Li, P., Wu, J.W., Zhang, Z., Alnemri, E.S. Chi, Y.
(2002) Structural insights into the pro-apoptotic function of mitochondrial serine protease
HtrA2/Omi. Nat. Struct. Biol. 9: 436 – 441.
35. Counce P.A., Keisling T.C., Mitchell A.J. (2000) A uniform, objective and adaptative
system for expressing rice development. Crop Sci. 40: 436–43.
36. Sperotto, R. A., Ricachenevsky, F. K., Duarte, G. L., Boff, T., Lopes K. L., Sperb, E. R.,
Grusak, M. A., Fett, J.P. (2009) Identification of up-regulated genes in flag leaves during rice
grain filling and characterization of OsNAC5, a new ABA-dependent transcription factor.
Planta. 230: 985–1002.
37. Livak, K.J. and Schmittgen, T.D. (2001) Analysis of relative gene expression data using
real-time quantitative PCR and the 2-ΔΔCt
method. Methods. 25: 402-408.
38. Schmittgen, T.D. and Livak, K.J. (2008) Analyzing real-time PCR data by the
comparative CT method. Nat Protoc. 3 (6): 1001-1008.
39. Roy, A., Kucukural, A., Zhang, Y. (2010) I-TASSER: a unified platform for automated
protein structure and function prediction. Nature Protocols, vol 5, 725-738.
40. Laskowski, R.A., MacArthur, M.W., Moss, D.S., Thornton, J.M. (1993). PROCHECK: a
program to check the stereochemical quality of protein structures. J. Appl. Cryst. 26, 283-291.
41. Ai, P., Sun, S., Zhao, J., Fan, X., Xin, W., Guo, Q., Yu, L., Shen, Q., Wu, P., Miller, A. J.
and Xu, G. (2009) Two rice phosphate transporters, OsPht1;2 and OsPht1;6, have different
functions and kinetic properties in uptake and translocation. Plant J. 57: 798–809.
42. Chen, Z., Zhu, Y.G., Liu, W.J., Meharg, A.A. (2005) Direct evidence showing the effect
of root surface iron plaque on arsenite and arsenate uptake into rice (Oryza sativa) roots. New
Phytol. 165: 91-97.

61
43. Riemer, J., Hoepken, H.H., Czerwinska, H., Robinson, S.R., Dringen, R. (2004)
Colorimetric ferrozine-based assay for the quantitation of iron in cultured cells. Anal
Biochem. 331: 370–375.
44. Spyropoulos, I.C., Liakopoulos, T.D., Bagos, P.G., Hamodrakas, S.J. (2004) TMRPres2D:
high quality visual representation of transmembrane protein models. Bioinformatics. 20 (17):
3258–3260.
45. Guindon, S. & Gascuel, O. (2000) A simple, fast, and accurate algorithm to estimate large
phylogenies by maximum likelihood. Syst. Biol. 52 (5): 696-704.
46. Gouy, M., Guindon, S., Gascuel, O. (2010) SeaView version 4: a multiplatform graphical
user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 27 (2):
221-224.

62
Legends to figures
Figure 1: (a) Schematic representation of OsNRAMP7 protein constructed with the software
TMRPres2D44
. The predicted 12 TMDs topology is based on HMMTOP algorithm. The
conserved motifs DPGN and MPH and the putative transit peptide are indicated. (b)
Schematic representation of transport in the three-dimensional model of OsNRAMP7
depicted by the secondary structure. (c) and (d) Transversal view of OsNRAMP7. In (d) the
asterisk indicates a large cavity present in the protein. (e) Parallel view to the membrane with
highlighted structures. (f) Detail of conserved structures. (c – f) Extended peptides MPH and
DPGN are colored yellow and magenta, respectively. CTM is depicted in red.
Figure 2. (a) Relative expression levels (RT-qPCR, relative to Ubiquitin expression) of
OsNRAMP7 in hydroponically grown 28-day old rice plants. (b) Relative expression levels of
OsNRAMP7 in distinct rice plant organs harvested at the R5 (grain filling) stage from field-
grown plants. Values are averages of three replicates ± SE. Means indicated by different
letters in (a) are different by t-test (P ≤ 0.05), while in (b) they differ by the Duncan test (P ≤
0.05).
Figure 3. Plant mRNA expression assays in Xenopus laevis oocytes. (a) Swelling assay of
aquaporin injected oocytes. Six independent oocytes injected with aquaporin mRNA were
analyzed on three consecutive days starting at the third day after the injections, when they
were subjected to 50% MBS and had the diameter size determined five times during twenty
minutes to establish an increase in volume. (b) Relative variation of iron content in single
oocytes injected or not with mRNA (aquaporin, OsNRAMP7 and truncated OsNRAMP7)
when exposed to two different concentrations of iron in MBS solution. Values are the average

63
of ten single oocytes and data was subjected to statistical analysis by Student’s t test (P =
0.05).

64
Supplementary data
Supplementary Figure 1: Alignment of members of the NRAMP family of rice made with
ClustalW (http://www.clustal.org/). Conserved amino acids are shaded and the Consensus
Transporter Motif (CTM) is highlighted in the box. Residues marked in darker shades of grey
belong to a conserved group of amino acids defined as strong by ClustalW.
Supplementary figure 2: (a) Alignment of AtNRAMP3, AtNRAMP4 and OsNRAMP7 made
with ClustalW (http://www.clustal.org/). Conserved amino acids are shaded. 68% of the
amino acids are identical in the three proteins. (b) Phylogenetic tree of NRAMP members in
Arabidopsis and rice. Sequences we aligned and the tree constructed using the PhyML’s
maximum likelihood method45
computed by Seaview46
.
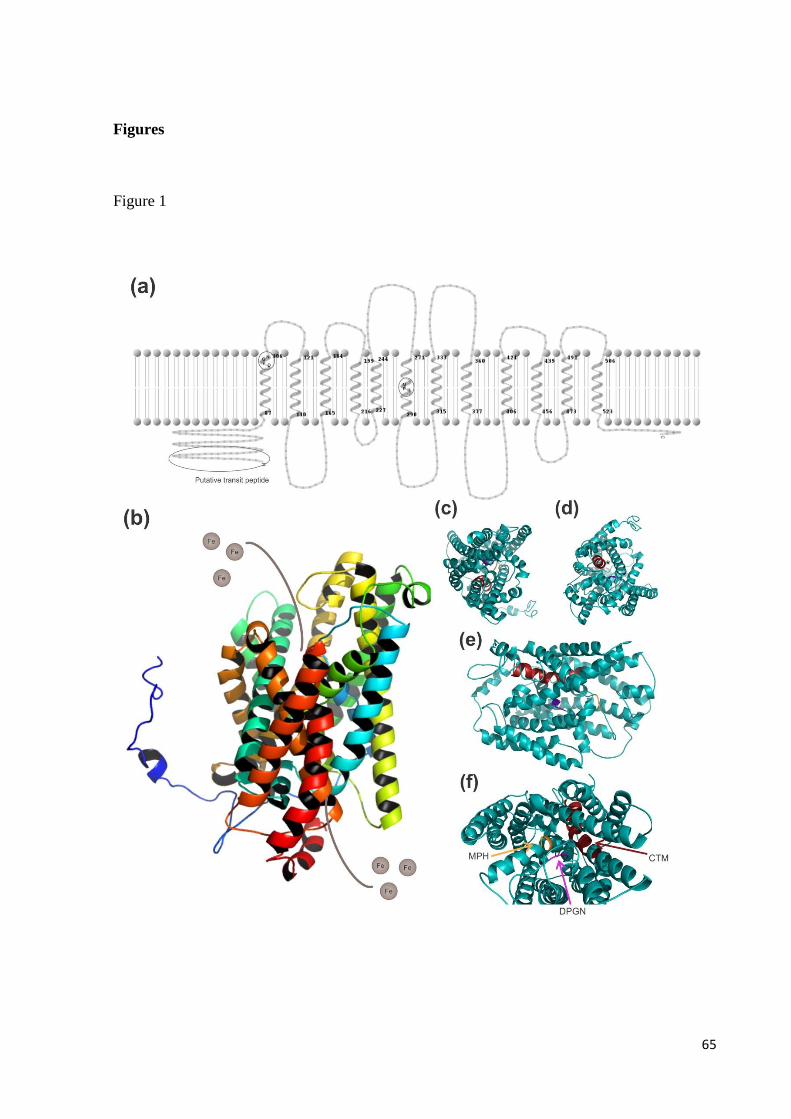
65
Figures
Figure 1
66
Figure 2
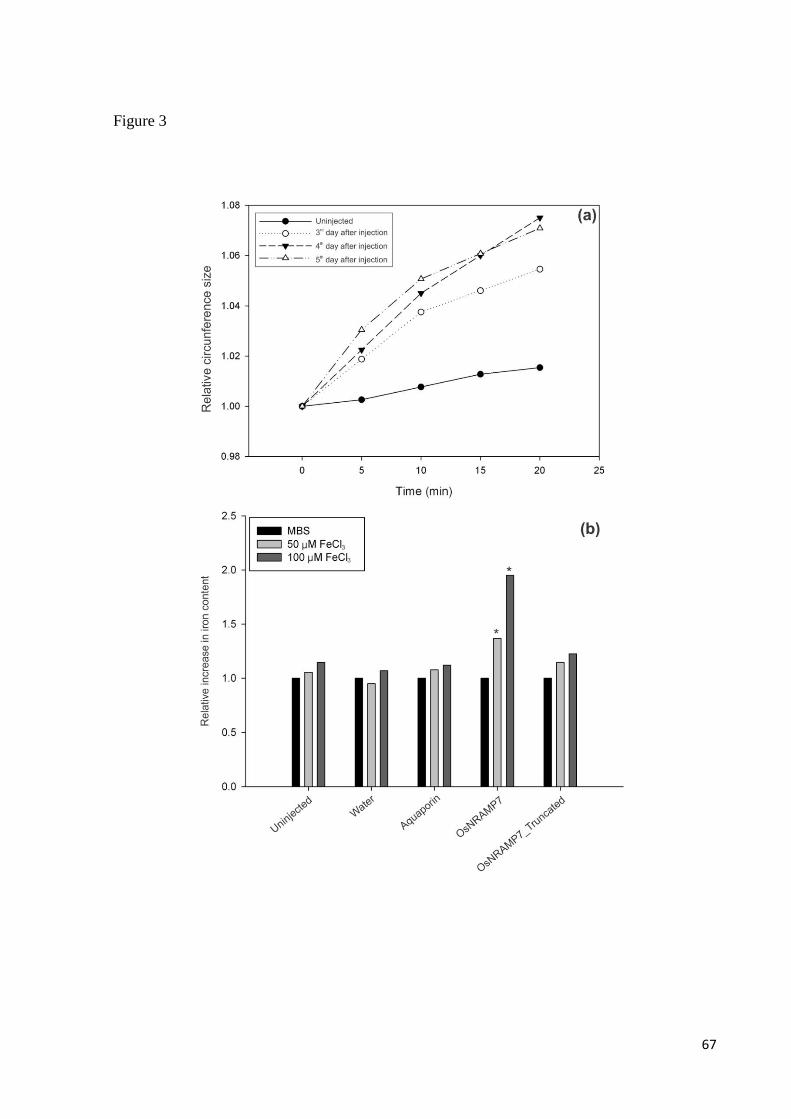
67
Figure 3

68
Supplementary figure 1

69
Supplementary figure 2

70
Capítulo 3
Dual impact on rice plants bearing OsFer2 mutation

71
Dual impact on rice plants bearing OsFer2 mutation
Lívia Scheunemann dos Santos, Luiza Monteavaro Mariath, Diogo Ribeiro Demartini,
Ricardo José Stein, Janette Palma Fett.
Abstract
Iron is an important micronutrient to plants, involved in several biological processes.
However, it is important that plants maintain control of iron homeostasis, since excess of iron
in the free form may catalyze the formation of oxygen radicals. One of the strategies to avoid
leaving iron available for such reactions is to store Fe inside ferritins. Ferritin is a spherical
protein capable of storing iron in its core, also acting as an iron buffer in cells. In this study, a
mutant line with impaired expression of OsFer2 was used to investigate a possible role of
ferritins in iron overload responses. Although rice has two genes encoding ferritin, the
OsFer2 mutation resulted in decreased ferritin transcript levels of both genes and less protein.
As mutant plants accumulated less biomass than WT also under the control treatment, rice
ferritin is apparently important for iron homeostasis and plant development even when iron
availability is considered normal. No photo-oxidative damage could be observed through
maximal efficiency of PSII photochemistry in mutant plants under iron excess conditions.
However, shoots of mutant plants had higher content of malondialdehyde (MAD), an
indicative of lipid peroxidation, and roots of mutant plants had increased APX activity, both
in the iron excess treatment, indicating a possible enhancement of oxidative stress in the
mutant under this condition. Further studies are required to establish if increasing iron stress
would further alter the oxidative stress response or if a compensating mechanism, such as
influx to vacuole and production of frataxin, are involved in oxidative stress responses in
OsFer2 rice mutants.
Keywords: ferritin, Oryza sativa L., oxidative stress.

72
Introduction
Iron is an essential nutrient for plant growth and development. Both physical and
chemical properties of iron enable this micronutrient to participate in most basic reduction
reactions in an organism. Iron is essential for a variety of processes such as photosynthesis
and DNA synthesis (Briat & Lobréaux, 1997). Iron is directly involved in the electron transfer
chain of respiration and acts a co-factor of enzymes involved in the synthesis of plant
hormones (Guerinot, 2011). Since iron homeostasis must be tightly regulated, plants have
different strategies to maintain cellular homeostasis. Among these strategies are allocation of
the metal to the vacuole, translocation to plastids and storage in ferritins.
Different functions have been described to ferritins depending on the species analyzed.
In peas it seems to have an important role in iron storage in seeds, releasing the metal when
necessary and enabling germination (Becker et al., 1998). A distinctive role has been
proposed to Arabidopsis seeds, since it is estimated that no more than 5% of the seed iron
content would be stored in ferritins (Ravet et al., 2009).
Ferritins have been found in mitochondria, chloroplasts and cell walls (Becker et al.,
1998). The functional ferritin protein is formed by 24 subunits, which can store up to 4000
ferric iron atoms each. Before being stored inside the mineral core of the protein as Fe3+
, iron
is oxidized by the protein ferroxidase center (Arosio et al., 2008). Ferritins provide an
intracellular easy-access form of iron and might have a putative detoxification property. This
could be due to the fact that the oxidation reaction necessary for iron storage actually
consumes oxygen and hydrogen peroxide (Arosio et al., 2008).
Two ferritin genes have been identified in rice, OsFER1 and OsFER2 (Gross et al.,
2003). Very little sets these two genes apart. OsFER1 possesses three deletions and 15 single
nucleotide changes within the coding region (Stein et al., 2009). Using specially designed
primers, Stein et al. (2009) was able to separate by electrophoresis the transcripts amplified
from each gene.
The present work uses a mutant OsFER2 line to study ferritin influence in protection
against oxidative stress in rice. This mutant is a result of the insertion of the Tos17
transposon, one of the most active retroelements in rice plants.

73
Results
A rice line containing a Tos17 insertion in the OsFER2 gene was identified in the Rice
Genome Resource Center (RGRC) bank. Seeds were obtained, plants segregating for the
mutation were screened by PCR analysis using specific primers and an homozygous line was
identified. Aligning the genomic sequence of OsFER2 with the respective Flanquing
Sequence Tags (FSTs) of the mutant line NG0250 allowed to determine that the insertion of
the retroelement occurred in the 3’UTR (untranslated region) of the gene (Figure 1).
There was the necessity to determine if the gene was being transcribed and the protein
produced even in the presence of the insertion. For that, the amount of transcripts of both
ferritin genes was analyzed. The expression pattern of the ferritin genes in both wild type
(WT) and mutant lineage were evaluated by semi-quantitative RT-PCR, using an ubiquitin
gene as control for loading and optical density measurements. It was possible to observe that
the level of ferritin transcripts (OsFER1 and OsFER2) was higher on wild type plants under
iron excess, while in mutants a low amount of transcripts was detected in either treatment
(Figure 2a).
Using an optical density analysis it was possible to observe an increase by two fold in
the amount of OsFER1 transcripts from the control to iron excess treatment in WT plants
(Figure 2a). An increase of 20% in the amount of OsFER2 transcripts was also observed in
WT plants. Mutant plants had fewer transcripts in both treatments. Under control conditions,
mutant plants had 25% less OsFER1 transcripts than WT. When subjected to iron excess,
mutant plants increased the level of OsFER1 transcripts by 30%, while WT plants more than
doubled the OsFER1 transcript abundance. The level of OsFER2 transcripts in mutant plants
was not sufficient to allow optical density analysis (Figure 2a).
To confirm that mutants were impaired in ferritin protein production, a Western Blot
analysis was performed. Only shoots of plants subjected to iron excess treatments were used
for this analysis (when more protein would be produced and, therefore, more easily detected).
Virtually no protein was detected in mutant plants (Figure 2b).
Once established that the mutant plants were in fact defective in the production of
ferritin, stress-related analysis were performed in order to assess its role in protection against
oxidative damage. Chlorophyll fluorescence was evaluated to establish if the iron excess
treatment had affected the photosynthetic apparatus. Plants with low content of ferritin did not
have a significant difference of the maximal efficiency of PSII photochemistry (Fv/Fm)

74
values when compared to WT plants under the same conditions (Figure 3). No significant
difference between treatments was observed in WT plants (Figure 3). Other measurements
related to photosynthetic efficiency, such as the quantum yield of electron transport
(ET0/ABS) and the efficiency with which an electron can move to the PSI electron acceptor
(ET0/TR0) also didn’t result in significant differences between genotypes or treatments (data
not shown).
Roots of mutant plants showed a significant decrease in length when subjected to iron
stress treatment (Figure 4). The same was not observed in WT plants. Although there was no
difference in shoot length, either between genotypes or treatments (Figure 4), there was a
significant difference in shoot fresh weight. While WT plants showed a significant decrease in
fresh weight under iron excess, the same could not be observed in mutant plants (Figure 5a).
Fresh weight of mutant plants was significantly smaller than in WT in the control condition,
however was not altered by the iron excess treatment. The same pattern was observed for the
fresh weight of roots (Figure 5b).
There was no difference in shoot dry weight either among genotypes or treatments
(Figure 5c). On the other hand, there was a significant decrease in root dry weight in WT
plants under iron excess. Despite not showing a significant difference between treatments,
root dry weight of mutant plants was much lower in control condition than in WT plants
(Figure 5d).
Both shoots and roots were used for oxidative damage analyses. Oxidative damage to
lipids was observed through malondialdehyde (MDA) quantification. MDA is produced when
polyunsaturated lipids are degraded by reactive oxygen species (ROS) and can be quantified
by Thiobarbituric Acid Reactive Substances (TBARS) analysis. Both genotypes increased
MDA content when subjected to iron excess (Figure 6a). However, shoots of mutant plants
seemed to accumulate more MDA than WT plants under excess iron (Figure 6a). A tendency
of increase in MDA concentration was also observed in roots of mutant plants (Figure 6b).
Despite a tendency of raise in H2O2 levels in roots of mutant plants under iron excess
treatment, no significant difference was observed in H2O2 assays (Figure 6c, d).
There was no significant difference in shoot ascorbate peroxidase (APX) activity
(Figure 7a), while roots of both genotypes had increased APX activity under iron excess
(Figure 7b). Of these, mutant plants had higher increase in APX activity than WT plants.
There was no difference in catalase (CAT) (Figure 7c, d) or superoxide dismutase (SOD)

75
(Figure 7e, f) activity in roots and shoots of both genotypes.
Discussion
Rice mutants with lower content of the ferritin protein accumulated less biomass than
WT plants. This can be stated by the fact that, despite having similar shoot lengths, shoot
weight of mutant plants were significantly lower than those observed in WT under control
conditions. This difference is even stronger when analyzing their roots. Although WT and
mutant plants had no significant difference in length, WT roots were much more abundant,
resulting in higher fresh weight.
Mutant plants accumulated less biomass than WT plants in the control treatment:
lower fresh weight in roots and shoots, lower dry weight in roots. These results indicate that,
besides a possible function in prevention of oxidative stress when plants are exposed to iron
excess, ferritins may also have an important role in iron homeostasis under normal iron
availability.
Both genotypes presented similar levels of peroxide production and antioxidant
enzyme activities when subjected to iron excess treatment. However, in the mutants, APX
activity in roots under iron excess was more evident. The increased MDA concentration in
shoots of mutant plants submitted to iron excess could be a sign of oxidative stress. The
combined results observed in OsFER2 mutant plants indicated that ferritin might have a
protective role related to iron stress in rice plants.
It has been described that different organs might have distinct physiological strategies
to respond to oxidative stress. In a work conducted by Fang et al. (2001) iron stress resulted in
decreased SOD activities, but increased APX activity in leaves. On a different note, Bode et
al. (1995) reported that APX activities under iron stress were unchanged. In a different
experiment, Majerus et al. (2007) described that sheaths and laminae respond differently to
iron stress. On their experiment, SOD activity was stimulated in the first and APX in the
latter. Variations in ferritin concentration among different plant organs could have an indirect
impact on the activities of these enzymes, since levels of free iron would be altered according
to ferritin availability.
Plants can resort to several detoxification strategies. The vacuole has an important role
when considering intracellular iron homeostasis. In Arabidopsis, AtFPN2 (Morrisey et al.,

76
2009) and AtVIT1 (Kim et al., 2006) were considered as iron influx proteins to the vacuole.
Since a vacuolar influx transporter of iron in rice is not known, it is virtually impossible to
determine if a particular protein with this function is up-regulated in this situation to
overcome the stress generated by iron overload.
Also involved in subcellular iron homeostasis is the mitochondrial protein frataxin.
Putative functions of this protein include assisting in iron–sulfur cluster assembly (Chen et al.,
2002) or involvement in energy conversion and oxidative phosphorylation (Ristow et al.,
2000). This protein is believed to act in conjunction with ferritins and the vacuole to prevent
oxidative stress. Frataxin mutants showed increase in the content of reactive oxygen species
and high levels of transcripts that encode proteins involved in oxidative stress response (Busi
et al., 2006). This supports the proposed role of the protein on prevention of oxidative
damage.
There might be an increase in the production of proteins such as frataxin or a vacuole
iron influx transporter in rice plants bearing OsFER2 mutation. This compensating
mechanism could be set into action in the presence of low amounts of ferritin, protecting the
plant from oxidative damage. Further studies in order to better understand iron homeostasis
and transport to the vacuole and in organelles such as chloroplasts and mitochondria will lead
to a better understanding of the mechanisms plants may resort to when under abiotic stress.
While knowledge in this area is still at surface, research in oxidative stress responses may
lead to a better understanding of possible compensating mechanisms.
It appears that ferritins act in iron homeostasis in rice plants in two ways: one
independent from the iron status inside the cell and a second one in response to iron excess.
Plants with impaired ferritin production accumulated less biomass than WT in the control
treatment and had increased stress responses (such as MDA concentration in shoots and APX
activity in roots) under iron stress. Exposing plants to an even higher concentration of iron
may help to further characterize ferritin involvement in oxidative stress responses in rice.

77
Material and Methods
Plant growth
Seeds of the rice (Oryza sativa L. cv. Hwayoung) mutant line NG0250, with a Tos17
insertion in the OsFER2 gene, were requested from the Rice Genome Resource Center –
RGRC (http://tos.nias.affrc.go.jp/~miyao/pub/tos17/index.html.em). Homozygous lines were
identified by PCR using specific primers (F: 5’ ACTTGCCAGGCTTCGAGTTA 3’, R: 5’
CGCAGTAGCAATGGAGTGAA 3’, Tos17 3’ tail: 5’ AGGTTGCAAGTTAGTTAAGA 3’).
Experiments were conducted with homozygous plants of this line and the WT background
line.
Seeds were germinated for 4 days in an incubator (28° C, first 2 days in the dark and
last 2 days in the light) on paper soaked with distilled water. After germination and growth in
vermiculite and nutrient solution (0.1 mM KCl, 0.1 mM KH2PO4, 0.7 mM K2SO4, 2 mM
Ca(NO3)2 4 H2O, 0.5 mM MgSO4 7 H2O, 0.5 µM MnSO4 4 H2O, 0.01 µM (NH4)6 Mo7
O24 4 H2O, 10 µM H3BO3, 0.5 µM ZnSO4 7 H2O, 0.2 µM CuSO4 5 H2O, 100 µM FeSO4
7 H2O, 100 µM EDTA) for 14 days (28° C, with 16 h of light), plants were transferred to pots
containing 2.5 L of nutrient solution. All solutions were replaced every 3 days. Plants were
cultivated in a growth room at 26 ± 1° C under white light with a photoperiod of 16/8 h
light/dark cycle (irradiance of approximately 100 µmol m–2
s–1
at the plant tops) for another
period of 14 days. After this period they were subjected to iron excess treatment, were 1.25
mM of FeSO4 was the final concentration of iron in solution. To maintain iron solubility an
equimolar concentration of EDTA was added to the solution (Sillanpää & Oikari, 1996).
Solutions were changed every three days and plants were under this condition for 9 days,
when analyses were conducted.
RNA extraction and cDNA synthesis
Rice leaves were harvested from plants grown under iron excess or control conditions
for 14 days (n = 4 per each group). RNA extractions and cDNA synthesis were performed as
described by Sperotto et al (2010).

78
Semi-quantitative RT-PCR analysis
Semi-quantitative RT-PCR was conducted as described by Stein et al (2009) with the
following modification: amplification of the ubiquitin cDNA was used to normalize the data.
Analysis was performed using three biological replicates, each consisting of three plants.
Ubiquitin optical density was designated 1.00 and the relative normalized optical density for
the remaining sample was expressed as percentage. The signal intensity of the stained bands
was photographed using a digital imager (Kodak DC120 Zoom Digital Camera) and analyzed
using the ImageJ 1.45 program (http://rsbweb.nih.gov/ij/index.html). Although only one
representative gel was shown, the quantified data represents the averages of three independent
replicates.
Protein extraction and Western Blot
Protein extraction was performed as described by Motta et al. (2001) with
modifications. 1 ml of extraction buffer (50 mM Tris pH 8, 10% sucrose, 1 mM EDTA, 1 mM
PMSF, 1 mM θ-phenanthroline, 0,1% SDS) was added to 100 mg of leaf powder
homogenized with liquid nitrogen. The mixture was pelleted by centrifugation at 10000 rpm
for 10 minutes at 4° C. The supernatant was transferred to 1.5 ml eppendorf tubes and protein
concentration was determined using the Quant-iT Protein Assay Kit® and the Qubit®
Fluorometer (Invitrogen). Samples (20 μg) were analyzed on 12.5% SDS-PAGE, according to
Laemmli (1970). Western blot analysis was performed according to Silveira et al. (2009) with
the following modifications: electrotransfer was used to relocate proteins to PVDF membrane
(140 V, 350 mA, 1 hour) and the membrane was incubated with serum diluted in Blotto
(1:200) overnight. Representative blots are shown.
Chlorophyll a fluorescence
Chlorophyll fluorescence was quantified using a portable OS-30p Chlorophyll
Fluorometer (Opti-Sciences Inc., Hudson, NH, USA) in a room at 28 ± 1°C under white light
with a photoperiod of 16/8 h light/dark cycle (irradiance of approximately 100 µmol m-2
s-1
at
plant tops). The OJIP rising transient measurements were made on attached first fully
expanded leaves which were dark-adapted for 12 hours. Eight plants from each genotype and
treatment were used. According to the model of energy fluxes in this test, the photon flux

79
absorbed by the antennae pigments is indicated as absorption flux (ABS).
Part of this excitation energy is dissipated, mainly as heat and less as fluorescence
emission, the excitation energy flux which reaches the reaction center (RC) and gets trapped
there (in the sense of leading to QA reduction) is indicated as trapping flux (TR). In the RCs,
the excitation energy is converted to redox energy by reducing primary quinine electron
acceptor of PSII (QA) to QA−, which is then reoxidized to QA, leading to an electron
transport flux (ET), which maintains the metabolic reactions of photosynthetic apparatus. The
derivation of the formulae for the various energy fluxes and for the flux ratios in the JIP test is
resultant from Strasser & Strasser (1995) and Krüger et al. (1997) (for a review, see Strasser
et al., 2000).
The trapping probability (TR0/ABS, the Fv/Fm ratio) was estimated, which is the
probability that an absorbed photon will be trapped by the Photosystem II reaction center with
the resultant reduction of QA (primary electron acceptor of Photosystem II). Means were
compared by Student’s t test (p ≤ 0.05) using the SPSS Base 12.0 for Windows (SPSS Inc.,
Chicago, IL, USA).
Oxidative Damage to Lipids and H2O2 determination
Lipid peroxides were extracted in ethanol 80% from fully expanded leaves and lipid
peroxidation determined by measuring the concentration of thiobarbituric acid-reacting-
substances (TBARS) as described by Du & Bramlage (1992). Hydrogen peroxide was
quantified spectrophotometrically (Cintra 5, GBC Scientific Equipment, Victoria, AU) after
extraction with 0.1% TCA and reaction with KI in the dark (Alexieva et al., 2001). The
amount of hydrogen peroxide was calculated using a standard curve prepared with known
concentrations. Analysis was performed using three biological replicates, each consisting of
three plants. Means were compared by Analysis of Variance (p ≤ 0.05) using the SPSS Base
12.0 for Windows (SPSS Inc., Chicago, IL, USA), and no significant differences were
detected.
Antioxidant enzymes activity
For all enzymatic activity determinations, fully expanded leaves were ground in cold
extraction buffer containing 50 mM of sodium phosphate buffer (pH 7.4), 1% PVP, 1 mM

80
EDTA, 1mM PMSF and 1mM benzamidine. The homogenate was centrifuged at 12000 g for
15 min at 4°C and the supernatant immediately used for enzymatic assays. Ascorbate
peroxidase (APX) activity was determined according to Klapheck et al. (1990), from the
decrease in absorbance at 290 nm, catalase (CAT) activity was determined following the
decrease of absorbance at 240 nm due to H2O2 consumption (Cakmak & Marschner, 1992)
and superoxide dismutase (SOD) activity was quantified as described by Beyer & Fridovich
(1987), using 15 min of illumination and recording the absorbance at 560 nm. All enzymatic
assays were performed at 25°C as initial activities, with no lag period, and protein
concentration was determined using the Quant-iT Protein Assay Kit® and the Qubit®
Fluorometer (Invitrogen). Analysis was performed using three biological replicates, each
consisting of three plants. Means were compared by Tukey test (p ≤ 0.05) using the SPSS
Base 12.0 for Windows (SPSS Inc., Chicago, IL, USA) where appropriate.
Acknowledgements
Authors would like to thank grants and scholarships from CNPq and FAPERGS.

81
References
Alexieva V, Sergiev S, Mapelli S, Karanov E (2001) The effect of drought and ultraviolet
radiation on growth and stress markers in pea and wheat. Plant Cell Environm. 24:1337-1344.
Arosio P, Ingrassia R, Cavadini P (2009) Ferritins: a family of molecules for iron storage,
antioxidation and more. Biochim Biophys Acta 1790:589–599
Becker R, Manteuffel R, Neumann D, Scholz G (1998) Excessive iron accumulation in the
pea mutants dgl and brz:subcellular localization of iron and ferritin. Planta. 207:217–223
Beyer WF & Fridovich I (1987) Assaying of superoxide dismutase activity: some large
consequences of minor changes in conditions. Anal Biochem. 161:559-566
Bode K, Döring O, Lüthje S, Neue HU, Böttger M (1995) The role of active oxygen in iron
tolerance of rice (Oryza sativa L.). Protoplasma 184:249-255
Briat JF & Lobréaux S (1997) Iron transport and storage in plants Trends Plant Sci. Reviews
2:187–193
Busi MV, Maliandi MV, Valdez H, Clemente M, Zabaleta EJ, Araya A, Gomez-Casati DF
(2006) Deficiency of Arabidopsis thaliana frataxin alters activity of mitochondrial Fe–S
proteins and induces oxidative stress. Plant J. 48:873–882
Cakmak I & Marschner H (1992) Magnesium deficiency and high light intensity enhance
activities of superoxide dismutase, ascorbate peroxidase and glutathione reductase in bean
leaves. Plant Physiol. 98:1222-1227
Chen OS, Hemenway S, Kaplan J (2002) Inhibition of Fe-S cluster biosynthesis decreases
mitochondrial iron export: evidence that Yfh1p affects Fe-S cluster synthesis. PROC. NATL.
ACAD. SCI. USA.. 99:12321–12326
Du Z & Bramlage WJ (1992) Modified thiobarbituric acid assay for measuring lipid oxidation
in sugar-rich plant tissue extracts. J. Agric. Food Chem. 40:1566-1570
Fang WC, Wang JW, Lin CC, Kao CH (2001) Iron induction of lipid peroxidation and effect
on antioxidative enzyme activities in rice leaves. Plant Growth Regul. 35:75-80
Gross J, Stein RJ, Fett-Neto AG, Fett JP (2003) Iron homeostasis related genes in rice. Genet.
Mol. Biol. 26 (4):477-497
Guerinot ML (2011) Iron. In: Cell Biology of Metals and Nutrients, Plant Cell Monographs.

82
17:75-94
Hodges D.M., DeLong J.M., Forney C.F., Prange R.K. (1999) Improving the thiobarbituric
acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing
anthocyanin and other interfering compounds. Planta. 207:604-611
Kim SA, Punshon T, Lanzirotti A, Li L, Alonzo JM, Ecker JR, Kaplan J, Guerinot ML (2006)
Localization of iron in Arabidopsis seed requires the vacuolar membrane transporter VIT1.
Science. 314:1295–1298
Klapheck S., Zimmer I., Cosse H. (1990) Scavenging of hydrogen peroxide in the
endosperm of Ricinus communis by ascorbate peroxidase. Plant Cell Physiol. 31:1005-1013
Krüger GHJ, Tsimilli-Michael M, Strasser RJ (1997) Light stress provokes plastic and elastic
modifications in structure and function of Photosystem II in Camellia leaves. Physiol Plant
101: 265-277
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of
bacteriophage T4, Nature. 227:680–685
Majerus V, Bertin P, Swenden V, Fortemps A, Lobréaux S, Lutts S (2007) Organ-dependent
responses of the African rice to short-term iron toxicity: ferritin regulation and antioxidative
responses. Biol Plant 51:303–331
Morrissey J, Baxter IR, Lee J, Li L, Lahner B, Grotz N, Kaplan J, Salt DE, Guerinot ML
(2009) The ferroportin metal efflux proteins function in iron and cobalt homeostasis in
Arabidopsis. Plant Cell. 21:3326-3338
Motta A, Bassoa B, Dell’Orto M, Briat JF, Soave C (2001) Ferritin synthesis in response to
iron in the Fe-inefficient maize mutant ys3. Plant Physiol. Biochem. 39:461−465
Ravet K, Touraine B, Kim SA, Cellier F, Thmine S, Guerinot ML, Briat J-F, Gaymard F
(2009) Post-translational regulation of AtFER2 ferritin in response to intracellular iron
trafficking during fruit development in Arabidopsis. Mol. Plant. 2:1095–1106
Ristow M, Pfister MF, Yee AJ, Schubert M, Michael L, Zhang CY, Ueki K, Michael MD,
Lowell BB, Kahn CR (2000) Frataxin activates mitochondrial energy conversion and
oxidative phosphorylation. PROC. NATL. ACAD. SCI. USA.. 97:12239–12243
Sillanpää M & Oikari A (1996) Assessing the impact of complexation by EDTA and DTPA
on heavy metal toxicity using microtox bioassay. Chemosphere. 32(8):1485-1497

83
Silveira VC, Fadanelli C, Sperotto
RA, Stein
RJ, Basso
LA, Santos
DS, Vaz Junior
IS, Dias
JF,
Fett JP (2009) Role of ferritin in the rice tolerance to iron overload. Sci. Agric. 66(4):549-555
Sperotto RA, Boff T, Duarte GL, Santos LS, Grusak MA, Fett JP (2010) Identification of
putative target genes to manipulate Fe and Zn concentrations in rice grains. J. Plant Physiol.
167 (17):1500-1506
Stein RJ, Ricachenevsky FK, Fett JP (2009) Differential regulation of the two rice ferritin
genes (OsFER1 and OsFER2). Plant Science 177:563–569
Strasser BJ & Strasser RJ (1995) Measuring fast fluorescence transients to address
environmental questions: The JIP-test. – In: Mathis, P. (ed.): Photosynthesis: From Light to
Biosphere. 977-980
Strasser RJ, Srivastava A, Tsimilli-Michael M (2000) The fluorescence transient as a tool to
characterize and screen photosynthetic samples. In: Yunus M, Pathre U, Mohanty P (eds)
Probing Photosynthesis: Mechanism, Regulation and Adaptation. Taylor & Francis, London.
443–480
Yoon JH, An SH, Kyeong IG, Lee MS, Kwon SC, Kang JH (2011) Oxidative modification of
ferritin induced by hydrogen peroxide. BMB Rep. 44(3):165-9

84
Figures
Figure 1
Figure 1: Structure of the OsFER2 gene. Introns are indicated by black lines and exons by
gray boxes. The position where the retroelement Tos17 was inserted in the mutant line
NG0250 is also indicated.

85
Figure 2
Figure 2: Analysis of ferritin protein and RNA expression in rice leaves. a. Semi-quantitative
RT-PCR of WT and mutant plants under control (CC) or iron excess treatment (+Fe). Semi-
quantitative RT-PCR was performed using standard conditions and the expression of
ubiquitin (OsUbq) was used as control for equal loading. Numbers below each lane represent
the percentage in relation to ubiquitin optical density (means of three independent replicates).
b. Western Blot of protein extracted from WT and mutant plants submitted to iron excess
treatment (1.25 mM) for nine days. Purified pea ferritin (3 µg) was used as positive control.
86
Figure 3
Figure 3: Analysis of maximum photochemical efficiency (Fv/Fm) on leaves of rice plants
exposed for 9 days to control (CC) or iron excess treatment (+Fe). Values are the averages of
five samples ± SE.

87
Figure 4
Figure 4: Shoot and root length of WT and mutant plants under control (CC) or iron excess
treatment (+Fe). Values are the averages of ten samples ± SE. Statistical differences by the
Student's t-test in comparison to control are shown by one asterisk (p ≤ 0.05).
88
Figure 5
Figure 5: Fresh and dry weight of WT and mutant plants under control (CC) or iron excess
treatment (+Fe). Values are the averages of ten samples ± SE. Means indicated by different
letters are different by the Tukey test (p ≤ 0.05).

89
Figure 6
Figure 6: Effect of iron excess in oxidative damage to lipids (MDA concentration) and H2O2
concentration. WT and mutant plants were analyzed under control (CC) or iron excess
treatment (+Fe). a and b. MDA concentration, quantified by TBARS. c and d. Effect of iron
stress in hydrogen peroxide content in shoots (c) and roots (d). Values are the averages of
three samples ± SE. Means indicated by different letters are different by the Tukey test (p ≤
0.05).
90
Figure 7
Figure 7: Antioxidant enzyme activity assays. APX (a, b), CAT (c, d) and SOD (e, f) activity
from shoots and roots of plants subjected to control (CC) or iron excess treatment (+Fe).
Values are the averages of ten samples ± SE. Different lower case letters above bars indicate
significant difference between means according to the Tukey test (p ≤ 0.05).

91
Considerações finais
Sendo o ferro um mineral essencial ao organismo vegetal, vários artigos ressaltam a
importância de se aprofundar o conhecimento a cerca de sua homeostase em plantas. Diversos
mecanismos envolvidos na homeostase de ferro já foram descritos em Arabidopsis thaliana.
Contudo, até o momento não temos a mesma quantidade de informação disponível para Oryza
sativa. Os resultados obtidos sobre a proteína OsNRAMP7 permitiram determinar que a
mesma é capaz de mediar o transporte de ferro através de membranas biológicas e, portanto,
acredita-se que o mesmo ocorra em células de plantas de arroz. OsNRAMP7 apresenta
características específicas da família NRAMP, como o CTM (Consensus Transport Motif) e a
sequência conservada DPGN, possivelmente envolvida na ligação de metais à proteína
transportadora. A expressão de OsNRAMP7 levou ao aumento na concentração de ferro em
oócitos de Xenopus. Portanto, é provável que OsNRAMP7 esteja envolvida na manutenção da
homeostase de ferro em plantas de arroz.
Como continuidade a este primeiro trabalho, já estão sendo realizados experimentos de
complementação de leveduras deficientes no influxo de metais como ferro e zinco. Mesmo a
proteína já tendo sido caracterizada como um transportador transmembrana, experimentos
com diferentes linhagens de levedura poderão fornecer indicações sobre a função de
OsNRAMP7 como transportadora de membrana plasmática ou tonoplasto. A realização de
testes de complementação em leveduras mutantes defectivas na absorção de outros metais,
como o zinco, se deve ao fato de a família de transportadores NRAMP estar associada à
translocação de outros metais, como manganês, cobalto, zinco, cobre, cádmio, níquel
(Ňuňuková et al., 2010) e vanádio (Ueki et al., 2011). A localização subcelular da proteína
OsNRAMP7, a ser investigada por meio de sua fusão a GFP e analisada em protoplastos de
arroz, deve ajudar a esclarecer se de fato trabalhamos com um transportador vacuolar.
Outras técnicas que possibilitam melhor compreensão do funcionamento da proteína
são análises eletrofisiológicas e mutações sítio-dirigidas. Em um projeto aprovado pelo
CNPq, a ser realizado em colaboração com o presente grupo, estão previstos tais
experimentos. Utilizando a técnica de voltage-clamp, correntes elétricas serão medidas na
membrana de oócitos injetados com mRNA de OsNRAMP7, determinando assim sua
atividade quando submetida a ferro e outros metais. Foram observados no presente trabalho,
na estrutura da proteína, motivos possivelmente importantes para a atividade de transporte de

92
metais pela proteína. Estudos utilizando mutação sítio-dirigida visam investigar a relação
entre estrutura e função, bem como a importância de aminoácidos conservados.
A redução na quantidade de ferritina em plantas mutantes OsFer2 pode estar
envolvida em mais do que respostas ao estresse oxidativo gerado por excesso de ferro. Plantas
mutantes parecem ter desenvolvimento atípico, caracterizado por plantas menores do que
aquelas do tipo selvagem. A ausência de ferritina na parte aérea levou ao aumento na
concentração de MDA, subproduto da degradação de lipídios poli-insaturados por espécies
reativas de oxigênio. Em raízes de plantas mutantes, o excesso de ferro levou ao aumento da
atividade da enzima APX, responsável pela detoxificação de peróxidos na célula. Os
resultados obtidos levam a crer que a proteína não é importante para a homeostase de ferro
apenas quando a planta se encontra em estresse por excesso do metal, mas também em
condições normais de crescimento.
De certa forma, os resultados relativos à proteína OsNRAMP7 podem vir a
complementar os estudos com as plantas mutantes para OsFER2. Caso a proteína
OsNRAMP7 seja mesmo um transportador responsável pelo influxo de ferro para o vacúolo,
esta proteína poderia atuar na detoxificação do excesso de ferro em situações de limitação da
disponibilidade de ferritina. Melhor compreensão sobre o papel do vacúolo no estresse por
excesso de ferro, bem como análises da importância da frataxina, deverão esclarecer os
mecanismos utilizados para lidar com situações de estresse. Até o presente momento não
existem trabalhos que relatem a função da proteína frataxina em plantas de arroz, apesar de
uma cópia ter sido descrita para a espécie (Busi et al., 2004).
Acreditamos que os dados aqui expostos, bem como os demais experimentos que já
estão sendo realizados, possam auxiliar na compreensão do intrincado mecanismo relacionado
à homeostase de ferro em plantas de arroz.

93
Referências
ABRAMSON J, SMIRNOVA I, KASHO V, VERNER G, IWATA S, KABACK HR (2003)
The lactose permease of Escherichia coli: Overall structure, the sugar-binding site and the
alternating access model for transport. FEBS Letters. 555 (1): 96-101
AGRANOFF D, COLLINS L, KEHRES D, HARRISON T, MAGUIRE M, KRISHNA S
(2005) The Nramp orthologue of Cryptococcus neoformans is a pH-dependent transporter of
manganese, iron, cobalt and nickel. Biochem. J. 385: 225-232
AI P, SUN S, ZHA, J, FAN X, XIN W, GUO Q, YU L, SHEN Q, WU P, MILLER AJ, XU G
(2009) Two rice phosphate transporters, OsPht1;2 and OsPht1;6, have different functions and
kinetic properties in uptake and translocation. Plant J. 57: 798–809
ALEXIEVA V, SERGIEV S, MAPELLI S, KARANOV E (2001) The effect of drought and
ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environm.
24:1337-1344
ANDREWS SC, HARRISON PM, YEWDALL SJ (1992) Structure, function and evolution
of ferritins. J Inorg Biochem. 47: 161–174
ARGÜELLO JM (2003) Identification of Ion-Selectivity Determinants in Heavy-Metal
Transport P 1B-type ATPases. J. Membrane Biol. 195 (2): 93-108
AROSIO P, INGRASSIA R, CAVADINI P (2009) Ferritins: a family of molecules for iron
storage, antioxidation and more. Biochim Biophys Acta 1790:589–599
BALK J & PILON M (2011) Ancient and essential: the assembly of iron–sulfur clusters in
plants. Trends Plant Sci. 16:218-226
BASHIR K, INOUE H, NAGASAKA S, TAKAHASHI M, NAKANISHI H, MORI S,
NISHIZAWA NK (2006) Cloning and characterization of deoxymugineic acid synthase genes
from graminaceous plants. J Biol Chem. 281:32395–32402
BECKER R, MANTEUFFEL R, NEUMANN D, SCHOLZ G (1998) Excessive iron
accumulation in the pea mutants dgl and brz: subcellular localization of iron and ferritin.

94
Planta. 207:217–223
BESSE M, KNIPFER T, MILLER AJ, VERDEIL JL, JAHN TP, FRICKE W (2011)
Developmental pattern of aquaporin expression in barley (Hordeum vulgare L.) leaves. J.
Exp. Bot. 62 (12): 4127-4142
BEYER WF & FRIDOVICH I (1987) Assaying of superoxide dismutase activity: some large
consequences of minor changes in conditions. Anal Biochem. 161:559-566
BODE K, DÖRING O, LÜTHJE S, NEUE HU, BÖTTGER M (1995) The role of active
oxygen in iron tolerance of rice (Oryza sativa L.). Protoplasma 184:249-255
BOHN L, MEYER AS, RASMUSSEN SK (2008) Phytate: impact on environment and
human nutrition A challenge for molecular breeding. JZUS-B. 9(3):165-191
BOLDRINI II, LONGHI-WAGNER HM, BOECHAT SC (2005) Morfologia e taxonomia de
gramíneas sul-riograndenses. Porto Alegre: Ed. UFRGS. 95p.
BOORER MJ, FORDE BG, LEIGH RA, MILLER AJ (1992) Functional expression of a plant
plasma membrane transporter in Xenopus oocytes. FEBS. 302 (2): 166-168
BOUZAYEN M, FELIX G, LATCHÉ A, PECH JC, BOLLER T (1991) Iron: an essential
cofactor for the conversion of 1-aminocy-clopropane-1-carboxylic acid to ethylene. Planta.
184:244-247
BRIAT JF & LOBRÉAUX S (1997) Iron transport and storage in plants Trends Plant Sci
Reviews 2:187–193
BRIAT JF, DUC C, RAVET K, GAYMARD F (2010) Ferritins and iron storage in plants.
BBA - General Subjects. 1800 (8): 806-814.
BRIAT JF, FOBIS-LOISY I, GRIGNON N, LOBREAUX S, PASCAL N, SAVINO G,
THOIRON S, VON WIRÉN N, VAN WUYTSWINKEL O (1995) Cellular and molecular
aspects of iron metabolism in plants. Biol Cell. 84:69–81.
BRIAT, J. F. & LOBRÉAUX, S. (1998). Iron storage and ferritin in plants. In: Iron Transport
and Storage in Microorganism, Plants and Animals, vol. 25, Metal Ions in Biological
Systems, pp. 563–584, Sigel A. and Sigel H. (eds), Marcel Dekker, New York.

95
BURR, B. et al. (2005) The map-based sequence of the rice genome. Nature 436: 793-800.
BUSI MV, MALIANDI MV, VALDEZ H, CLEMENTE M, ZABALETA EJ, ARAYA A,
GOMEZ-CASATI DF (2006) Deficiency of Arabidopsis thaliana frataxin alters activity of
mitochondrial Fe–S proteins and induces oxidative stress. Plant J. 48:873–882
BUSI MV, ZABALETA EJ, ARAYA A, GOMEZ-CASATI DF (2004) Functional and
molecular characterization of the frataxin homolog from Arabidopsis thaliana. FEBS Lett.
576:141–144
CAKMAK I & MARSCHNER H (1992) Magnesium deficiency and high light intensity
enhance activities of superoxide dismutase, ascorbate peroxidase and glutathione reductase in
bean leaves. Plant Physiol. 98:1222-1227
CELLIER M, PRIVÉ G, BELOUCHI A, KWAN T, RODRIGUES V, CHIA W, GROS P
(1995) Nramp defines a family of membrane proteins. PROC. NATL. ACAD. SCI. USA.. 92
(22): 10089-10093.
CHEN OS, HEMENWAY S, KAPLAN J (2002) Inhibition of Fe-S cluster biosynthesis
decreases mitochondrial iron export: Evidence that Yfh1p affects Fe-S cluster synthesis. Proc.
Natl. Acad. Sci. USA. 99(19): 12321-12326
CHEN XZ, PENG JB, COHEN A, NELSON H, NELSON N, HEDIGER MA (1999) Yeast
SMF1 mediates H+-coupled iron uptake with concomitant uncoupled cation currents. J Biol.
Chem. 274: 35089–35094.
CHEN Z, ZHU YG, LIU WJ, MEHARG AA (2005) Direct evidence showing the effect of
root surface iron plaque on arsenite and arsenate uptake into rice (Oryza sativa) roots. New
Phytol. 165: 91-97.
CHOPIN F, ORSEL M, DORBE M-F, CHARDON F, TRUONG H-N, MILLER AJ, KRAPP
A, DANIEL-VEDELE F (2007) The Arabidopsis ATNRT2.7 nitrate transporter controls
nitrate content in seeds. Plant Cell. 19:1590-1602
CONTE SS & WALKER EL (2011) Transporters contributing to iron trafficking in plants.
Mol. Plant. 4:464-476
COUNCE PA, KEISLING TC, MITCHELL AJ (2000) A uniform, objective and adaptative

96
system for expressing rice development. Crop Sci. 40: 436–43.
COURVILLE P, URBANKOVA E, RENSING C, CHALOUPKA R, QUICK M, CELLIER
MF (2008) Solute carrier 11 cation symport requires distinct residues in transmembrane
helices 1 and 6. J. Biol. Chem. 283: 9651–9658
CURIE C, CASSIN G, COUNCH D, DIVOL F, HIGUCHI K, LE JEAN M, MISSON J,
SCHIKORA A, CZERNIC P, MARI S (2009) Metal movement within the plant: contribution
of nictotianamine and yellow stripe 1-like transporters. Ann. Bot. 103:1–11
CURIE C, PANAVIENE Z, LOULERGUE C, DELLAPORTA SL, BRIAT JF, WALKER
EL (2001) Maize yellow stripe1 encodes a membrane protein directly involved in Fe(III)
uptake. Nature. 409:346-349
DEVOS KM & GALE MD (2000) Genome Relationships: The Grass Model in Current
Research. Plant Cell. 12: 637-646
DIDONATO RJ, ROBERTS LA, SANDERSON T, EISLEY RB, WALKER EL (2004),
Arabidopsis Yellow Stripe-Like2 (YSL2): a metal-regulated gene encoding a plasma
membrane transporter of nicotianamine–metal complexes. Plant J. 39:403–414
DURRETT TP, GASSMANN W, ROGERS EE (2007) The FRD3-mediated efflux of citrate
into the root vasculature is necessary for efficient iron translocation. Plant Physiol. 144:197-
205
DUY D, STÜBE R, WANNER G, PHILIPPAR K (2011) The Chloroplast Permease PIC1
regulates plant growth and development by directing homeostasis and transport of iron. Plant
Physiol. 155(4):1709-1722
DUY D, WANNER G, MEDA AR, VON WIREN N, SOLL J, PHILIPPAR K (2007) PIC1,
an ancient permease in Arabidopsis chloroplasts, mediates iron transport. Plant Cell. 19:986–
1006
EIDE D, BRODERIUS M, FETT J, GUERINOT ML (1996) A novel iron-regulated metal
transporter from plants identified by functional expression in yeast. PROC. NATL. ACAD.
SCI. USA.. 93:5624-5628
FANG WC, WANG JW, LIN CC, KAO CH (2001) Iron induction of lipid peroxidation and

97
effect on antioxidative enzyme activities in rice leaves. Plant Growth Regul. 35:75-80
GARCIA-OLIVEIRA AL, TAN L, FU Y, SUN C (2009) Genetic identification of
quantitative trait loci for contents of mineral nutrients in rice grains. J. Integr. Plant Biol. 51:
84-92.
GOMOLPLITINANT KM & SAIER MH JÚNIOR (2011) Evolution of the oligopeptide
transporter family. J Membr Biol. 240(2):89–110
GOUY M, GUINDON S, GASCUEL O (2010) SeaView version 4: a multiplatform graphical
user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 27 (2):
221-224.
GREEN LS & ROGERS EE (2004) FRD3 controls iron localization in Arabidopsis. Plant
Physiol. 136:2523-2531
GROSS J, STEIN RJ, FETT-NETO AG, FETT JP (2003) Iron homeostasis related genes in
rice. Genet. Mol. Biol. 26 (4) 477-497.
GROTZ N & GUERINOT ML (2006) Molecular aspects of Cu, Fe and Zn homeostasis in
plants. Biochim Biophys Acta. 1763:595-608
GRUENHEID S, CELLIER M, VIDAL S, GROS P (1995) Identification and characterization
of a second mouse Nramp gene. Genomics. 25 (2): 514-525.
GUERINOT ML (2011) Iron. In: Cell Biology of Metals and Nutrients, Plant Cell
Monographs. 17:75-94
GUIDOLIN AF (1993) Caracterização de genótipos de arroz irrigado por técnicas
eletroforéticas. Pelotas: UFPel, 92 p. Dissertação de Mestrado.
GUINDON S & GASCUEL O (2000) A simple, fast, and accurate algorithm to estimate large
phylogenies by maximum likelihood. Syst. Biol. 52 (5): 696-704.
GUNSHIN H, MACKENZIE B, BERGER UV, GUNSHIN Y, ROMERO MF, BORON WF,
NUSSBERGER S, GOLLAN JL, HEDIGER MA (1997) Cloning and characterization of
mammalian proton-coupled metal-ion transporter. Nature. 388: 482–488.

98
GURA T (1999) New genes boost rice nutrients. Science. 285: 994-995
HAEMIG HAH & BROOKER RJ (2004) Importance of Conserved Acidic Residues in
MntH, the Nramp homolog of Escherichia coli. J. Membrane Biol. 201: 97–107.
HARRISON PM & AROSIO P (1996) The ferritins: molecular properties, iron storage
function and cellular regulation. Biochem. Biophys. Acta. 1275: 161–203
HAYDON M J & COBBETT C S (2007) Transporters of ligands for essential metal ions in
plants. New Phytol. 174:499–506
HEAZLEWOOD JL, TONTI-FILIPPINI JS, GOUT AM, DAY DA, WHELAN J, MILLAR
AH (2004) Experimental analysis of the Arabidopsis mitochondrial proteome highlights
signaling and regulatory components, provides assessment of targeting prediction programs,
and indicates plant-specific mitochondrial proteins. Plant Cell. 16:241–256
HELL R & STEPHAN UD (2003) Iron uptake, trafficking and homeostasis in plants. Planta.
216:541–551
HIGUCH K, WATANABE S, TAKAHASHI M, KAWASAKI S, NAKANISHI H,
NISHIZAWA NK, MORI S (2001) Nicotianamine synthase gene expression differs in barley
and rice under Fe-deficient conditions. Plant J. 25:159-167
HODGES DM, DELONG JM, FORNEY CF, PRANGE RK (1999) Improving the
thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues
containing anthocyanin and other interfering compounds. Planta. 207:604-611
INOUE H, HIGUCHI K, TAKAHASHI M, NAKANISHI H, MORI S, NISHIZAWA NK
(2003) Three rice nicotianamine synthase genes, OsNAS1, OsNAS2, and OsNAS3 are
expressed in cells involved in long-distance transport of iron and differentially regulated by
iron. Plant J. 36:366–381
INOUE H, KOBAYASHI T, NOZOYE T, TAKAHASHI M, KAKEI Y, SUZUKI K,
NAKAZONO M, NAKANISHI H, MORI S, NISHIZAWA NK (2009) Rice OsYSL15 is an
iron-regulated iron(III)-deoxymugineic acid transporter expressed in the roots and is essential
for iron uptake in early growth of the seedlings. J Biol Chem. 284:3470–3479

99
IRGSP - International Rice Genome Sequencing Project. (2005) The map-based sequence of
the rice genome. Nature. 436: 793-800.
IRRI – International Rice Research Institute. Acessado em: 05 de dezembro de 2011.
Disponível em:
http://beta.irri.org/solutions/index.php?option=com_content&task=view&id=250
ISHIMARU Y, BASHIR K, FUJIMOTO M, AN G, ITAI RN, TSUTSUMI N, NAKANISHI
H, NISHIZAWA NK (2009) Rice-specific mitochondrial iron-regulated gene (MIR) plays an
important role in iron homeostasis. Mol Plant. 2:1059–1066
ISHIMARU Y, MASUDA H, BASHIR K, INOUE H, TSUKAMOTO T, TAKAHASHI M,
NAKANISHI H, AOKI N, HIROSE T, OHSUGI R, NISHIZAWA NK (2010) Rice metal-
nicotianamine transporter, OsYSL2, is required for the long-distance transport of iron and
manganese. Plant J. 62:379–390
ISHIMARU Y, SUZUKI M, TSUKAMOTO T, SUZUKI K, NAKAZONO M, KOBAYASHI
T, WADA Y, WATANABE S, MATSUHASHI S, TAKAHASHI M, NAKANISHI H, MORI
S, NISHIZAWA NK (2006) Rice plants take up iron as an Fe3+
-phytosiderophore and as Fe2+
.
Plant J. 45:335–346
JAQUINOD M, VILLIERS F, KIEFFER-JAQUINOD S, HUGOUVIEUX V, BRULEY C,
GARIN J, BOURGUIGNON, J (2007) A proteomics dissection of Arabidopsis thaliana
vacuoles isolated from cell culture. Mol Cell Proteomics. 6:394-412
JEONG J, COHU C, KERKEB L, PILN M, CONNOLLY EL, GUERINOT ML (2008)
Chloroplast Fe(III) chelate reductase activity is essential for seedling viability under iron
limiting conditions. PROC. NATL. ACAD. SCI. USA.. 105:10619–10624
JOHNSON AAT, KYRIACOU B, CALLAHAN DL, CARRUTHERS L, STANGOULIS J,
LOMBI E, TESTER M (2011) Constitutive Overexpression of the OsNAS Gene Family
Reveals Single-Gene Strategies for Effective Iron- and Zinc-Biofortification of Rice
Endosperm. PLoS ONE. 6(9):e24476
KIM SA, PUNSHON T, LANZIROTTI A, LI L, ALONZO JM, ECKER JR, KAPLAN J,
GUERINOT ML (2006) Localization of iron in Arabidopsis seed requires the vacuolar
membrane transporter VIT1. Science. 314:1295–1298

100
KISPAL G, CSERE P, PROHL C, LILL R (1999) The mitochondrial proteins Atm1p and
Nfs1p are essential for biogenesis of cytosolic Fe/S proteins. EMBO J. 18:3981-3989
KLAPHECK S, ZIMMER I, COSSE H (1990) Scavenging of hydrogen peroxide in the
endosperm of Ricinus communis by ascorbate peroxidase. Plant Cell Physiol. 31:1005-1013
KOBAYASHI T, ITAI RN, AUNG MS, SENOURA T, NAKANISHI H, NISHIZAWA, NK
(2012) The rice transcription factor IDEF1 directly binds to iron and other divalent metals for
sensing cellular iron status. Plant J. 69:81–91
KOBAYASHI T, SUZUKI M, INOUE H, ITAI RN, TAKAHASHI M, NAKANISHI H,
MORI S, NISHIZAWA NK (2005) Expression of iron-acquisition-related genes in iron-
deficient rice is co-ordinately induced by partially conserved iron-deficiency-responsive
elements. J. Exp. Bot. 56 (415):1305-1316.
KOIKE S, INOUE H, MIZUNO D, TAKAHASHI M, NAKANISHI H, MORI S,
NISHIZAWA NK (2004) OsYSL2 is a rice metal–nicotianamine transporter that is regulated
by iron and expressed in the phloem. Plant J. 39:415-424
KORSHUNOVA YO, EIDE D, CLARK WG, GUERINOT ML, PAKRASI HB (1999) The
IRT1 protein from Arabidopsis thaliana is a metal transporter with a broad substrate range.
Plant Mol. Biol. 40:37-44
KRÜGER GHJ, TSIMILLI-MICHAEL M, STRASSER RJ (1997) Light stress provokes
plastic and elastic modifications in structure and function of Photosystem II in Camellia
leaves. Physiol. Plant 101: 265-277
KRUGER C, BERKOWITZ O, STEPHAN UW, HELL R (2002) A metal-binding member of
the late embryogenesis abundant protein family transports iron in the phloem of Ricinus
communis L. J. Biol. Chem. 277:25062–25069
KUSHNIR S, BABIYCHUK E, STOROZHENKO S, DAVEY MW, PAPENBROCK J, DE
RUCKE R, ENGLER G, STEPHAN UW, LANGE H, KISPAL G, LILL R, VAN MONAGU
M (2001) A mutation of the mitochondrial ABC transporter Sta1 leads to dwarfism and
chlorosis in the Arabidopsis mutant starik. Plant Cell. 13:89–100
LAEMMLI UK (1970) Cleavage of structural proteins during the assembly of the head of

101
bacteriophage T4, Nature. 227:680–685
LANQUAR V, LELIEVRE F, BOLTE S, HAMES C, ALCON C, NEUMANN D,
VANSUYT G, CURIE C, SCHRODER A, KRAMER U, BARBIER-BRYGOO H,
THOMINE S (2005) Mobilization of vacuolar iron by AtNRAMP3 and AtNRAMP4 is
essential for seed germination on low iron. EMBO J. 24(23):4041-51
LASKOWSKI RA, MACARTHUR MW, MOSS DS, THORNTON JM (1993).
PROCHECK: a program to check the stereochemical quality of protein structures. J. Appl.
Cryst. 26, 283-291.
LEE S & AN G (2009) Over-expression of OsIRT1 leads to increased iron and zinc
accumulations in rice. Plant Cell Environm. 32:408-416
LEE S, CHIECKO JC, KIM SA, WALKER EL, LEE Y, GUERINOT ML, AN G (2009)
Disruption of OsYSL15 leads to iron inefficiency in rice plants. Plant Physiol. 150:786-800
LESCURE AM, PROUDHON D, PESEY H, RAGLAND M, THEIL EC, BRIAT JF (1991)
Ferritin gene transcription is regulated by iron in soybean cell cultures. PROC. NATL.
ACAD. SCI. USA. 88: 8222–8226
LI W, SRINIVASULA SM, CHAI J, LI P, WU JW, ZHANG Z, ALNEMRI ES, CHI Y
(2002) Structural insights into the pro-apoptotic function of mitochondrial serine protease
HtrA2/Omi. Nat. Struct. Biol. 9: 436 – 441.
LIVAK KJ & SCHMITTGEN TD (2001) Analysis of relative gene expression data using
real-time quantitative PCR and the 2-ΔΔCt method. Methods. 25: 402-408.
MAJERUS V, BERTIN P, SWENDEN V, FORTEMPS A, LOBRÉAUX S, LUTTS S (2007)
Organ-dependent responses of the African rice to short-term iron toxicity: ferritin regulation
and antioxidative responses. Biol Plant 51:303–331
MARENTES E & GRUSAK MA (1998) Iron transport and storage within the seed coat and
embryo of developing seeds of pea (Pisum sativum L.). Soil Science Res. 8: 367–375
MARISCAL V, MOULIN P, ORSEL M, MILLER AJ, FERNÁNDEZ E, GALVÁN A
(2006). Differential regulation of the Chlamydomonas Nar1 gene family by carbon and
nitrogen. Protist. 157 (4): 421-433.

102
MÄSER P, THOMINE S, SCHROEDER JI, WARD JM, HIRSCHI K, SZE H, TALKE IN,
AMTMANN A, MAATHUIS FJM, SANDERS D, HARPER JF, TCHIEU J, GRIBSKOV M,
PERSANS MW, SALT DE, KIM SA, GUERINOT, ML. (2001) Phylogenetic relationships
within cation transporter families of Arabidopsis. Plant Physiol. 126: 1646–1667
MAUGHAN SC, PASTERNAK M, CAIRNS N, KIDDLE G, BRACH T, JARVIS R, HAAS
F, NIEUWLAND J, LIM B, MÜLLER C, SALCEDO-SORA , E., KRUSE, C., ORSEL, M.,
HELL, R., MILLER, A.J., BRAY, P., FOYER, C.H., MURRAY, J.A.H., MEYER, A.J.,
COBBETT, C.S. (2010). Plant homologs of the Plasmodium falciparum chloroquine-
resistance transporter, PfCRT, are required for glutathione homeostasis and stress responses.
PROC. NATL. ACAD. SCI. USA.. 107 (5): 2331-2336.
MILLER AJ & ZHOU JJ (2000) Xenopus oocytes as an expression system for plant
transporters. Biochim. Biophys. Acta. 1465: 343-358.
MORI S, NISHIZAWA N, HAYASHI H, CHINO M, YOSHIMURA E, ISHIHARA J (1991)
Why are young rice plants highly susceptible to iron deficiency? Plant Soil. 130:1-2
MORRISSEY J, BAXTER IR, LEE J, LI L, LAHNER B, GROTZ N, KAPLAN J, SALT DE,
GUERINOT ML (2009) The ferroportin metal efflux proteins function in iron and cobalt
homeostasis in Arabidopsis. Plant Cell. 21:3326-3338
MOTTA A, BASSOA B, DELL’ORTO M, BRIAT JF, SOAVE C (2001) Ferritin synthesis in
response to iron in the Fe-inefficient maize mutant ys3. Plant Physiol. Biochem. 39:461−465
MSILINI N, ZAGHDOUDI M, GOVINDACHARY S, LACHAÂL M, OUERGHI Z,
CARPENTIER R (2011) Inhibition of photosynthetic oxygen evolution and electron transfer
from the quinone acceptor QA- to QB by iron deficiency. Photosynth. Res. 107(3):247-56
NAGASAKA S, TAKAHASHI M, NAKANISHI-ITAI R, BASHIR K, NAKANISHI H,
MORI S, NISHIZAWA NK (2009) Time course analysis of gene expression over 24 hours in
Fe-deficient barley roots. Plant Mol. Biol. 69:621–631
NEVO Y & NELSON N (2006) The NRAMP family of metal-ion transporters. BBA -
Molecular Cell Research. 1763 (7): 609-620.
ŇUŇUKOVÁ V, URBÁNKOVÁ E, JELOKHANI-NIARAKI M, CHALOUPKA R (2010)

103
Ion channel activity of transmembrane segment 6 of Escherichia coli proton-dependent
manganese transporter. Biopolymers. 93: 718–726.
OKUBO M, YAMADA K, HOSOYAMADA M, SHIBASAKI T, ENDOU H (2003)
Cadmium transport by human Nramp 2 expressed in Xenopus laevis oocytes. Toxicol. Appl.
Pharmacol. 187: 162–167.
PAINE J.A., SHIPTON C.A., CHAGGAR S, HOWELLS RH, KENNEDY MJ, VERNON G,
WRIGHT, SY, HINCHLIFFE E, ADAMS, JL, SILVERSTONE, AL, DRAKE R (2005)
Improving the nutritional value of Golden Rice through increased pro-vitamin A content.
Nature Biotech. 23: 482-487
PETIT JM, BRIAT JF, LOBRÉAUX S (2001) Structure and differential expression of the
four members of the Arabidopsis thaliana ferritin gene family. Biochem J. 359: 575-582
QI Y, WANG S, SHEN C, ZHANG S, CHEN Y, XU Y, LIU Y, WU Y, JIANG D (2012)
OsARF12, a transcription activator on auxin response gene, regulates root elongation and
affects iron accumulation in rice (Oryza sativa). New Phytol. 193:109–120
RAMIREZ L, SIMONTACCHI M, MURGIA I, ZABALETA E, LAMATTIN E (2011)
Nitric oxide, nitrosyl iron complexes, ferritin and frataxin:A well equipped team to preserve
plant iron homeostasis. Plant Sci. 181 (5):582–592
RAVET K, TOURAINE B, KIM SA, CELLIER F, THMINE S, GUERINOT ML, BRIAT J-
F, GAYMARD F (2009) Post-translational regulation of AtFER2 ferritin in response to
intracellular iron trafficking during fruit development in Arabidopsis. Mol. Plant. 2:1095–
1106
RICACHENEVSKY FK, SPEROTTO RA, MENGUER PK, FETT JP (2010) Identification
of Fe-excess-induced genes in rice shoots reveals a WRKY transcription factor responsive to
Fe, drought and senescence. Mol. Biol. Rep. 37:3735–3745.
RIEMER J, HOEPKEN HH, CZERWINSKA H, ROBINSON SR, DRINGEN R (2004)
Colorimetric ferrozine-based assay for the quantitation of iron in cultured cells. Anal
Biochem. 331: 370–375.
RISTOW M, PFISTER MF, YEE AJ, SCHUBERT M, MICHAEL L, ZHANG CY, UEKI K,

104
MICHAEL MD, LOWELL BB, KAHN CR (2000) Frataxin activates mitochondrial energy
conversion and oxidative phosphorylation. PROC. NATL. ACAD. SCI. USA.. 97:12239–
12243
ROBINSON NJ, PROCTER CM, CONNOLLY EL, GUERINOT ML (1999) A ferric-chelate
reductase for iron uptake from soils. Nature. 397:694-697
ROY A, KUCUKURAL A, ZHANG Y (2010) I-TASSER: a unified platform for automated
protein structure and function prediction. Nature Protoc. 5:725-738.
RUSHTON DL, TRIPATHI P, RABARA RC, LIN J, RINGLER P, BOKEN AK, LANGUM
TJ, SMIDT L, BOOMSMA DD, EMME NJ, CHEN X, FINER JJ, SHEN QJ, RUSHTON PJ
(2012) WRKY transcription factors: key components in abscisic acid signaling. Plant Biotech.
J. 10:2–11.
SANTI S, CESCO S, VARANINI Z, PINTON R (2005) Two plasma membrane H+-ATPase
genes are differentially expressed in iron-deficient cucumber plants. Plant Physiol. Bioch.
43(3):287-292
SANTOS LS DOS, COSTA DE OLIVEIRA A (2007) Rice iron metabolism: from source to
solution. J. Plant Sci. Biotech. 10: 64-72
SCHAAF G, HONSBEIN A, MEDA AR, KIRCHNER S, WIPF D, VON WIREN N (2006)
AtIREG2 encodes a tonoplast transport protein involved in iron-dependent nickel
detoxification in Arabidopsis thaliana roots. J. Biol. Chem. 281:25532–25540
SCHAAF G, SCHIKORA A, HÄBERLE J, VERT G, LUDEWIG U, BRIAT JF, CURIE C,
VON WIRÉN N (2005) A putative function for the arabidopsis Fe-Phytosiderophore
transporter homolog AtYSL2 in Fe and Zn homeostasis. Plant Cell Physiol. 46:762-774
SCHMITTGEN TD & LIVAK KJ (2008) Analyzing real-time PCR data by the comparative
CT method. Nature Protoc. 3 (6): 1001-1008.
SCREPANI E & HUNTE C (2007) Discontinuous membrane helices in transport proteins and
their correlation with function. J. Struct. Biol. 159, 261–267
SECKBACK JJ (1982) Ferreting out the secret of plant ferritin – a review. J. Plant Nutr. 5:
369–394.

105
SHIKANAI T, MÜLLER-MOULÉ P, MUNEKAGE Y, NIYOGI KK, PILON M (2003)
PAA1, a P-type ATPase of Arabidopsis, functions in copper transport in chloroplasts. Plant
Cell. 15:1333–1346
SIEDOW JN (1991) Plant lipoxygenase:structure and function. Annu. Rev. Plant Physiol.
Plant Mol. Biol. 42, 145-188
SILLANPÄÄ M & OIKARI A (1996) Assessing the impact of complexation by EDTA and
DTPA on heavy metal toxicity using microtox bioassay. Chemosphere. 32(8):1485-1497
SILVA MVE (1975) A Cultura do Arroz. Coleção técnica agrária. Clássica Editora. 171p.
SILVEIRA VC, FADANELLI C, SPEROTTO
RA, STEIN
RJ, BASSO
LA, SANTOS
DS,
VAZ JUNIOR IS, DIAS
JF, FETT JP (2009) Role of ferritin in the rice tolerance to iron
overload. Sci. Agric. 66(4):549-555
SOKAL I, OTTO-BRUC AE, SURGUCHEVA I, VERLINDE CL, WANG CK, BAEHR W,
PALCZEWSKI K (1999) Conformational changes in guanylyl cyclase-activating protein 1
(GCAP1) and its tryptophan mutants as a function of calcium concentration. J. Biol. Chem.
274 (28): 19829–19837.
SOLTI A, KOVÁCS K, BASA B, VÉRTES A, SÁRVÁRI É, FODOR F (2012) Uptake and
incorporation of iron in sugar beet chloroplasts. Plant Physiol. Biochem. 52:91-97
SPEROTTO RA, BOFF T, DUARTE GL, SANTOS LS, GRUSAK MA, FETT JP (2010)
Identification of putative target genes to manipulate Fe and Zn concentrations in rice grains. J.
Plant Physiol. 167 (17):1500-1506
SPEROTTO RA, RICACHENEVSKY FK, DUARTE GL, BOFF T, LOPES KL, SPERB ER,
GRUSAK MA, FETT JP (2009) Identification of up-regulated genes in flag leaves during rice
grain filling and characterization of OsNAC5, a new ABA-dependent transcription factor.
Planta. 230: 985–1002.
SPYROPOULOS IC, LIAKOPOULOS TD, BAGOS PG, HAMODRAKAS SJ (2004)
TMRPres2D: high quality visual representation of transmembrane protein models.
Bioinformatics. 20 (17): 3258–3260.
STACEY MG, PATEL A, MCCLAIN WE, MATHIEU M, REMLEY M, ROGERS EE,

106
GASSMANN W, BLEVINS DG, STACEY G (2008) The Arabidopsis AtOPT3 protein
functions in metal homeostasis and movement of iron to developing seeds. Plant Physiol.
146:589-601
STANGOULIS JCR, HUYNH BL, WELCH RM, CHOI EY, GRAHAM RD (2007)
Quantitative trait loci for phytate in rice grain and their relationship with grain micronutrient
content. Euphytica. 154: 289-294.
STEIN RJ, RICACHENEVSKY FK, FETT JP (2009) Differential regulation of the two rice
ferritin genes (OsFER1 and OsFER2). Plant Science 177:563–569
STRASSER BJ & STRASSER RJ (1995) Measuring fast fluorescence transients to address
environmental questions: The JIP-test. – In: Mathis, P. (ed.): Photosynthesis: From Light to
Biosphere. 977-980
Taiz, L. & Zeiger, E. (2004) Fisiologia Vegetal. 3ª ed. Porto Alegre: Artmed.
TAKAHASHI R, ISHIMARU Y, SENOURA T, SHIMO H, ISHIKAWA S, ARAO T,
NAKANISHI H, NISHIZAWA NK (2011) The OsNRAMP1 iron transporter is involved in
Cd accumulation in rice. J. Exp. Bot. 62 (14): 4843-4850.
TARANTINO D, CASAGRANDE F, SOAVE C, MURGIA I (2009) Knocking out of the
mitochondrial AtFer4 ferritin does not alter response of Arabidopsis plants to abiotic stresses.
J Plant Physiol. 167(6): 453-60
TERRES AL, GALLI J, FAGUNDES PPR, MACHADO M A, MAGALHÃES JR, A M de,
MARTINS, JF, NUNES, CDM, FRANCO, DF, AZAMBUJA, IHV (1998) Arroz irrigado no
Rio Grande do Sul: generalidades e cultivares. Pelotas: EMBRAPA – CLIMA TEMPERADO
(Embrapa Clima Temperado. Circular técnica, 14). 58p.
THOMINE S, LELIÈVRE F, DEBARBIEUX E, SCHROEDER JI, BARBIER-BRYGOO H
(2003) AtNRAMP3, a multispecific vacuolar metal transporter involved in plant responses to
iron deficiency. Plant J. 34 (5): 685-95.
THOMINE S, WANG RC, WARD JM, CRAWFORD NM, SCHROEDER JI (2000)
Cadmium and iron transport by members of a plant metal transporter family in Arabidopsis
with homology to Nramp genes. P. Natl. Acad. Sci. USA. 97: 4991-4996.

107
UEKI T, FURUNOB N, MICHIBATA H (2011) A novel vanadium transporter of the Nramp
family expressed at the vacuole of vanadium-accumulating cells of the ascidian Ascidia
sydneiensis samea. BBA - General Subjects. 1810 (4): 457-464.
UEKI T, UYAMA T, KANAMORI K, MICHIBATA H (2001) Subunit C of the vacuolar-
type ATPase from the vanadium-rich ascidian Ascidia sydneiensis samea rescued the pH
sensitivity of yeast vma5 mutants. Mar. Biotechnol. 3: 316–321.
UENO D, ROMBOLÀ AD, IWASHITA T, NOMOTO K, MA JF. (2007) Identification of
two novel phytosiderophores secreted by perennial grasses. New Phytol. 174: 304–310
VAROTTO C, MAIWALD D, PESARESI P, JAHNS P, SALAMINI F, LEISTER D (2002)
The metal ion transporter IRT1 is necessary for iron homeostasis and efficient photosynthesis
in Arabidopsis thaliana. Plant J. 31:589-599
VASCONCELOS M, ECKERT H, ARAHANA V, GRAEF G, GRUSAK MA, CLEMENTE
T (2006) Molecular and phenotypic characterization of transgenic soybean expressing the
Arabidopsis ferric chelate reductase gene, FRO2. Planta. 224(5):1116-1128
WANG XC, YANG J, HUANG W, HE L, YU JT, LIN QS, LI W, ZHOU HM (2002) Effects
of removal of the N-terminal amino acid residues on the activity and conformation of firefly
luciferase. Int. J. Biochem. Cell Biol. 34 (8): 983-91.
WATERS BM, CHU HH, DIDONATO RJ, ROBERTS LA, EISLEY RB, WALKER EL
(2006) Mutations in Arabidopsis Yellow Stripe-Like1 and Yellow Stripe-Like3 reveal their
roles in metal ion homeostasis and loading of metal ions in seeds. Plant Physiol. 141:1446–
1458
XIA J, YAMAJI N, KASAI T, MA JF (2010). Plasma membrane-localized transporter for
aluminum in rice. PROC. NATL. ACAD. SCI. USA.. 107 (43): 18381-18385.
YE X, AL-BABILI S, KLOTI A, ZHANG J, LUCCA P, BEYER PP (2000) Engineering the
provitamin A (β-carotene) biosynthetic pathway into (carotenoid-free) rice endosperm.
Science. 287: 303-305
YOKOSHO K, YAMAJI N, UENO D, MITANI N, MA JF (2009) OsFRDL1 is a citrate
transporter required for efficient translocation of iron in rice. Plant Physiol. 149:297-305

108
YOON JH, AN SH, KYEONG IG, LEE MS, KWON SC, KANG JH (2011) Oxidative
modification of ferritin induced by hydrogen peroxide. BMB Rep. 44(3):165-9
ZANCANI M, PERESSON C, BIROCCIO A, FEDERICI G, URBANI A, MURGIA I,
SOAVE C, MICALI F, VIANELLO A, MACRÝ F (2004) Evidence for the presence of
ferritin in plant mitochondria. Eur. J. Biochem. 271: 3657–3664


















