
l - pantheon.ufrj.brpantheon.ufrj.br/bitstream/11422/3012/1/278225.pdf · Apterigota do...
109
IOOLOGIAiFCC 71 l ___ 1_0_4 _37 __ _ . ' .' ADESlÇ -'-· OA DE FOLHI' Dissertação apresentada à . ' do Curso de Pós-Graduação da Universidade Federal Co�:frd_ e nação em Zoologia do Rio de . .- Janeiro· como requisito para .obtenç�o d�-grau de Mestre em Ci�ncias Biológ! cas (Zoologia). Rio de Janeiro 1984 l
Transcript of l - pantheon.ufrj.brpantheon.ufrj.br/bitstream/11422/3012/1/278225.pdf · Apterigota do...

1
IOOLOGIAiFCC 71
l ___ 1_0_4_37 __ _
. '
.' ADECOMPOSlÇÀO -'EM- ·UM AMONTOADO DE FOLHIÇf)'
Dissertação apresentada à . '
do Curso de Pós-Graduação
da Universidade Federal
Co�:fr d_e nação
em Zoologia
do Rio de . .-
Janeiro· como requisito para .obtenç�o
d�-grau de Mestre em Ci�ncias Biológ!
cas (Zoologia).
Rio de Janeiro 1984
l

•
TRABALHO REALIZADO NO SETOR DE APTERIGOTA
DO DEPARTAMENTO DE ENTOMOLOGIA DO MUSEU
NACIONAL DA UNIVERSIDADE FEDERAL DO
RIO DE JANEIRO
Orientador

j
7
7
--,
.. i
Aos meus filhos,
trico e Raphaela
Dedico este trabalho.

7
7
GUIMAR�ES, Anthony trico da Gama
Exemplo de uma sucessão faunísticr. durante à
decomposição em um amontoado de folhiço. Rio
de Janeiro, Museu Nacional - UFRJ, 1984.
viii, 101 p.
Tese: Mestre em Ciências {Zoologia)
1. Sucessão 2. Zoologia 3. Humificação
J) Museu Nacional - UFRJ
II) Titulo
BANCA EXAMINADORA
Professor Alceu Lemos de Castro Presidente
Professor Johan Becker
Professora Anna TimÕtheo da Costa
Rio de Janeiro, 11 de outubro de 1984

l
AGRADECIMENTOS
Ao Professor Rogen Pienne Hipolyte Ante, nosso Orie�
tador, que sugeriu o assunto, nos facultou parte da bibliografia,
discutiu o andamento e a apresentação do trabalho. Agradeço-lhe
ainda, os valiosos ensinamentos transmitidos durante todo o tem
po em que estive diretamente ligado ao Departamento de Entomolo
gia do Museu Nacional. Alem da inestimável colaboração ao reali
zar os desenhos que compõem as pranchas desta dissertação.
Ao Professor Annaldo Campo� do� Santo� Coelho, coor
denador do Curso de PÕs Graduação em Zoologia do Museu Nacional,
pelo apoio e compreensao nas horas difíceis da vida Acadêmica.
Ao Professor Alceu Lemo� de Ca�tno pe]a identificação
das espécies de Isopoda e Amphipoda, e o apoio dispensado duran
te a realização do Curso.
A Professora Ana Timotheo da Co�ta pe]a identificação
das espécies de Araneida, e interesse demonstrado no desenvolvi
mento deste trabalho.
Ao Professor Joaquim Machado pela identificação das
espécies de Dermaptera.
A equipe do Liboratõrio de Malacologia do Museu Naci�
nal pela identificação das espécies de Gastropoda durante as fa
ses preliminares dos nossos estudos.
, A Professora Miniam Mende� de Oliveina da Gama Guima
nae� pelo auxílio na triagem do material coletado.
v.

7
Ao Senhor tnieo da Gama Guimanãe-0 que colaborou decl
sivamente nas atividades de campo, mantendo sempre a ãrea estuda
da nas condições adequadas a realização do experimento.
A Professora Monique Anlê.: pelo auxílio na confecção
dos grãficos.
à Sra. Neide Cannana Fennande-0 que, com sua boa von
tade e eficiência, realizou exímio trabalho datilogrãfico.
Ao Professor Homeno Figueinedo de Oliveina pela ver
sao dd sumãrio para o Inglês.
A Profeisora I-0olda Rocha e Silva pelo apoio, intere!
se e incentivo a todas as atividades que culminaram na conclusão
deste trabalho.
Ao Dr. Leonida-0 de Mello Veane pelas facilidades con
cedidas e o apoio dispensado durante as· fases de montagem e reda
ção desta dissertação.
Ao Conselho Nacional de Desenvolvimento Científico e
TecnolÕgico (CNPq) pela Bolsa concedida.
As Bibliotecas do Museu Nacional, da Fundação Oswaldo
Cruz e da Universidade Federal do Rio de Janeiro e a seus funcio
nãrios pela atenção e paciência com que nos atenderam em diversas
oportunidades.
Aos Professores do Curso de Mestrado em Zoologia do
Museu Nacional, aos colegas do Horto Bótânico e do Departamento
de Entomologia do Instituto Oswaldo Cruz, que de alguma forma con
tribuiram na realização e aperfeiçoamento deste trabalho.
vi.

r N D I e E
Pãg.
I) INTRODUÇÃO ..................................... 2
I I) M ETODOLO GIA . . . . . . . . . ;. . . . . . . . . . . . . . . . . . . . . . . . . . 5
I I. l) Região Estudada . . . . . . . . . . . . . . . . . . . . . . . . . . 5
I I. 2) Descrição do Biõtopo . . . . . . . . . . . . . . . . . . . . . 6
I I. 3) Material e Metodos . . . . . . . . . . . . . . . . . . . . . . . 7
I I I) REVISAO DA L ITERATURA . . . . . . . . . . . . . . . . . . . . . . . . . . 11
IV) RESULTADOS OBTIDOS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . l 8
V)
IV. l) Anãlise da Participação de Todos os Grupos
na Sucessão . . . . . . . . . . . . . . . . . . . :. . . . . . . . . . . 18
IV. 2) Anãlise da Participação da Fauna Colemboli
g i ca na Sucessão . . . . . . . . . . . . . . . . . . . . . . . . . 2 7
IV. 3) Formas Biológicas e Regime Alimentar dos
Collembola .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
D ISCUSSJl:O 35
VI) CON CL USÕ ES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 9
V II) RESUMO 43
VI I I) SUMMARY 45
IX) REFERtNCIAS BIBL IOGRÃF ICAS 47
vii.

7
7
Pãg.
X) TABELAS • • • • • • • • • • • • • • • • • • • • • • • • • . • • • . • • . • . . • . • • 55
XI) ILUSTRAÇOES • • . • • . • . . . • • . • . . . • . • . • • .. • • • . • • • .. • 63
XI. 1) Pranchas • • • • • • • • . • • • • . • • • • • . • • • • . • . • . . . • • 64
XI.2) Figuras .................................. 68
XI. 3) Fotografias .. • • • .. • .. • • .. .. . .. .. .. .. .. . .. 99
vii i.

7
1, INTRODUÇAO
--,

I) INTRODUÇÃO
2.
Se observarmos o solo das florestas tropicais, perc!
bemos uma constante cobertura de folhas sobre este estrato. Esta
camada de folhas caídas das ãrvores, o folhiço ( 11litter 11 em lín
gua Inglesa) , e a base do processo de humificação dos solos e o
estrato protetor de todo o sistema edãfico.
Este processo de humificação a partir de uma diversi
ficada massa de matéria vegetal formada de folhas, galhos, frutos,
etc. , ao qual se juntam animais mortos, é de extrema importância
e de grande complexidade ecológica.
O resultado final deste processo, com a mineração dos
elementos permitinuo a sua absorção pelos vegetais, representa o
ciclo crítico de todos os ecossistemas terrestres.
Numa primeira fase, inicia-se a degradação das subs
tâncias hidrossolúveis por decomposição bacteriana. A seguir, em
uma fase mais prolongada, dã-se a decomposição das substâncias
nao solúveis em ãgua, pela ação simultânea da microflora, da mi
crofauna e da mesofauna edãfica. Esta ultima, toma importância
muito grande e o seu estudo permite acompanhar a evolução da bio
cenose, pondo a vista a sucessão dos vãrios grupos durante a for
mação do solo.
E precisamente o estudo da sucessao dos elementos da
mesofauna, sucessão reveladora das diversas fases de degradação
do folhiço e da formação das biocenoses edâficas, que apresent!
mos neste trabalho.

7
7
3,
Este tipo de abordagem foi realizado por HE�MM:· GISIN
(1949) e por GISE[.A GISIN (1952) na Suíça, para o estudo do desen
volvimento da biocenose de Collembola com e sem adição de fertili
zante. Em nosso caso, estudamos o desenvolvimento da mesofauna
pelo processo de concentração e localização de uma massa de folhi
ço por varreduras de quintal para dentro de um buraco, como e ex
plicado adiante.
Não foi estudado a estratificação como ocorre na for
maçao natural do solo biolõgico, analisamos sim o que se passa nos
e s t r a to s s u p e r f i c i a i s o u s e j a n a s c a ma d a s A O ( f o r n a , l i ti ê re , ma n
tillo, serrapilheira, folhiço) e A 1 jã humificada.

n
II, METODOLOGIA
i,·

.l
n
n
li
n
1
í
n
r
II) METODOLOGIA
II.l) REGIAO EST UDA DA
s.
Na escolha da ãrea para a realização dos nossos estu
dos, dois aspectos principais foram levados em consideração. Alem
de nos permitir um controle constante sobre o biõtopo, tínhamos
que ter mãxima segurança possível de que poderíamos seguir sempre
a mesma metodologia pelo espaço de tempo que julgãssemos necessã
rio.
Assim sendo, optamos pela realização do experimento no
quintal d� casa que nos servia de residência por ocasião dos nos
sos estudos. Localizada ã Rua Américo Brasiliense, numero 135,
em Madureira, SubÜrbio da Zona Norte da Cidade do Rio de Janeiro,
o local preenchia todos os requisitos estabelecidos: localização
prõxima ao nosso laboratõrio no Horto Botânico do Museu Nacional
(cerca de 20 Km) , ãrea f.artamente arborizada, possibilidade de um
controle direto do biótopo isolado para o experimento e as amos
tragens poderiam ser feitas seguindo a mesma rotina e nos interva
los previamente determinados.
Integrante dos domínios morfoestruturais do litoral
do Estado do Rio de Janeiro, a topografia geral da região encon
tra-se inserida em ãreas do Complexo Cristalino Atlântico, segu�
do MOREIRA e CARMEl1ER (1977) , com solos argila-arenoso que se
originaram basicamente de granitos e gnaisses.
NIMER (197 9) considera que o carater de clima trop! , cal e mais marcante que o temperado para a Região Sudeste e refe
re-se ao clima da ãrea em questão como quente e umido com de um
a dois meses de seca durante o ano.

6.
Segundo as medições realizadas durante os 12 meses,
julho de 1977 a junho de 1978, em que analisamos a sucessão fau
nística ocorrida, as medias anuais de temperatura e umidade rela
tiva do ar estiveram em torno de 25, 5 ° C e 82%, com um total de
1023,2 mm de precipitações pluviom�tricas (Tabela I e Figuras 23
a 26) .
A cobertura vegetal do local específico onde foram
levadas a efeito as nossas observações,caracterizou-se pela co�
figuração quase que primordial de estratos arbõr.eo e arbustivo,
constituindo-se das seguintes espécies: Mu-0a panadi-0iaea (ban�
neira) , Spondia-0 dulci-0 (cajã-manga) , Bambu-0a vulgani-0 (bambu j�
ponês) , Avennhoa canambola (carambola) , Puniea gnanatum (romã) ,
Mangióena indica (manga) , P-0idium guajava (goiaba) , S�enoealix
michelii (pitanga) , Peh-0ea ghati-0-0ima (abacate) , Lacuna mammo-0a
· (sapotí) , Chny-0alidocanpu-0 .lu�e-0cen-0 (palmeira bambu), distribu_!.
das em uma ãrea de aproximadamente 600 m 2 •
11. 2) DESCR IÇAO DO B IOTOPO
O biótopo onde estudamos a sucessao faunística ao lon
go de 360 dias consecutivos, julho de 1977 a junho de 1978, cons :.• -
tituiu-se de um buraco, com cerca de 60 cm de profundidade e 110
cm de diâmetro, escavado na ãrea descrita no item anterior, ao
qual denominaremos na presente dissertação de 11 fosso de folhiço 11 •
Por meio de varreduras em toda ãrea arborizada, trans
portamos para o interior do buraco as folhas, pequenos galhos, fr�
tos em d eco m p os i ç ão, ex creme n to d e a v e s e d e p e q u e no s ma m í f e r os d�
mésticos (cães e gatos) que casualmente encontravam-se no local,
acumulados em toda a extensão do quintal ao longo dos 15 dias
anteriores, formando assim o 11fosso de folhiço 11 �Fotos l e 2) .

--,
1
í
7.
Apõs a realização das varreduras, consequentemente o
início das observações, não foi mais aumentado ou renovado o fo
lhiço formador do biõtopo, possibilitando assim o acompanhamento
de todo o processo sucessõrio ocorrido.
Durante o experimento, o "fosso de folhiço 11 nao este
ve em nenhum momento sob a incidência direta de raios solares, en
centrando-se permanentemente ã sombra da cobertura vegetal exis
tente no local.
II. 3) MATERIAL E METODOS
Todo o material estudado no desenvolvimento do exper.!_
mente provem dos exemplares coletados no período de julho de 1977
a junho de 1978 no "fosso de folhiço 11 descrito detalhadamente no
item anterior e encontra-se depositado nas coleções do Setor de
Apterigota do Departamento de Entomologia do Museu Nacional do
Rio de Janeiro.
A metodologia utilizada desde a coleta do material ate
a fase de triagem e identificação pode ser dividida em duas eta
pas. A primeira prende-se as atividades de campo, isto e, a cole
ta do folhiço em quantidades suficientes para completar um saco
plãstico com capacidade de 3 litros, sem comprimi-lo. Concomita�
temente a cada uma dessas amostragens anotavamos a temperatlJra e
a umidade relativa do ar do momento. Para tal, utilizamo-nos de
termômetro de mãxima e mínima, marca Incoterm e higrômetro de
ponteiro, marca Huger, que se encontravam fixados no local de co ·-leta. As medições das precipitações pl�viom�tricas eram realiz!
das, apõs cada amostragem, pela leitura do pluviômetro ali inst!
lado. Esta, correspondia ao total de precipitações
desde a coleta anterior, aproximadamente 15 dias.
acumulados

r
n
8.
A etapa seguinte consistiu das atividades realizadas
em nosso laboratõrio, onde a fauna contida no folhiço era extrai
da por metodologia dinâmica, através de funis de Berlese-Tullgrem
(TU L LGREM, 1917) .
Basicamente, o funil de extração e formado por uma C_!:!
ba cilíndrica, aberta em ambas extremidades, com 20 centímetros
de altura, tendo na parte inferior uma peneira, onde é deposit�
do o folhiço, cujo tamis possui 5 milímetros de diâmetro. Abai
xo, um funil com 50 centímetros de altura converge para um pequ�
no frasco onde colocamos ãlcool ·ã 96 ° GL.
Este método mostrou-se bastante eficaz para a extra
çao da fauna. No interior do reservatório, destinado ao folhiço,
forma-se um gradiente totalmente desfavorãvel fazendo com que a
fauna migre verticalmente para o interior do funil -a procura de
condições de umidade mais satisfatórias ã suas exigências.
Ultrapassando a peneira que sustenta o folhiço, a fau
na desliza pelo funil indo depositar-se no frasco com ãlcool co
locado na extremidade inferior do aparelho. Quando houver nece!
sidade de acelerar o processo, utiliza-se uma lâmpada de 25 Watts
que, colocada centralmente e a cerca de 20 centímetros acima do
aparelho, irã provocar a dessecação do folhiço mais rapidamente.
Logo ao primeiro dia de observação começa a depositar-se
no ãl cool a fauna menos resistente a dessecação. Levando aproximad�
mente l O dias para que toda a fauna migre pelo i'nteri or do funil.
Os vãrios grupos faunisticos selecionãveis segundo e!
te processo, apresenta-se diferentemente quando entram em contacto
com o ãlcool. Enquanto os representantes da ordem Collembola e

r
r
r
r
7 7
9.
alguns Acarina mantêm a maioria dos especimes flutuando na supe�
fície líquida, os demais dirigem-se rapidamente para o fundo do
frasco. No primeiro caso, ê necessãrio que vertamos cuidadosa
mente a película de alcool com estes especimes para outro frasco
e os submeta a ação imediata de ãlcool quente, para que possamos
romper a tensão superficial existente entre eles. Quanto aos que
permaneceram no fundo do primeiro frasco, são acondicionados nor
malmente em ãlcool ã 9 6 ° GL.
Não tendo como um dos objetivos do nosso trabalho a
identificação a níveis mais elevados que o de ordem para os gr�
pos faunísticos encontrados, somente os Collembola, por ser o de
maior afinidade com a rotina das pesquisas em nosso laboratõrio,
mereceram um estudo taxionÔmico a nível generico. Para tal, ba
seamo-nos nos criterio-s estabelecidos por SA LMON (19 6 4) para o
grupo.
Com objetivo de demonstrarmos a participação efetiva
de cada um dos grupos faunísti cos na sucessão, todos os resultados
computados sao apresentados sob a forma de tabelas e gráficos .
Essas tabelas permitem a observação das ocorrências segundo dois
aspecto-s principais: anãlise dos percentuais de cada grupo ou g�
nero de Collembola em diferentes fases da sucessão (Tabelas V e
V I I I) e quais os grupos ou generos de Collembola que atuaram em
cada etapa deste processo (Tabelas IV e V I I).

7
7
III, REVISAO DA LITERATURA

11.
III) REVISÃO DA LITERATURA
Durante anos os estudos sobre a biologia do solo re�
tringiram-se a aspectos da s ua composição química, particularme�
te relacionados com a fertilidade e a presença de potãs sio, nitr�
gênio e fos fatos . Posteriormente, a estrutura e a unidade dos s o
1 o s d e s p e r ta r a m o i n te r e s s e d o s F í s · i c o s e c o n com i t a n tem e n te o s Mi
c r o b i o 1 o g i s t a s i n i c i a r a m e s tu d o s s o b r e o s p r o b 1 em a s g e r a i s d e c re s
cimento dos vegetais.
Somente no final do século XIX os BiÕlogos começaram
a estudar não sõ as Bactérias, os Fungos e os Oligochaeta, mas
também as populações de diversos grupos faunísticos do solo.
Na realidade os estudos sobre a Zoologia do Solo ti
veram início com um trabalho de DARWIN (1881) , no qual chama a
a t e n ç ão p a r a o s v e r me s d a t e r r a , e com o s e s t u d o s d e M () L L E R ( 1 884 )
que publicou Bodenzoologie.
ApÕs os primeiros estudos em Zoologia do Solo, obser
vou-se que ê de vital importância para a humificação do folhiço
a atividade de diversos grupos da mes ofauna.
Baseado nes ses fatos, HURSH (1928) comentou em seu
trabalho: "Litter l<'eeps forest soil productive 11•
"The. 6011.v.:,t bu,í,.f_d6 up and ó�ze6 ili own -6oil ••• Foull. -
ó,<..óth-6 oó the. nt.dJu.e.n:t.6 a.J.i-6,Ún,U,a;le.d e.ac.h ye.aJL by 601tv.:,t tlte.eó aJte. .tz.e.tull.ne.d
to the. -6 oi.f.. aJ.i .f..e.a.vv.:, and de:t.Jú:tu.6 . • • The. time., .tz.e.qU,U.ie.d 60.tz. a.de.qua.te.
de.c.ompo.6,l;ü on VafL,{,e6 6.tz.om one to many ye.a.lL6. Fo.tz. the. a.ve..tz.a.ge oó Southe..tz.n
haJc.dvJood óoJz.ut c.oncli....tÃ..on.6 a. pvúod o 6 ;two to th.tz.e.e. ye.a.lL6 ,{,-6 ptz.obab.ty
.6 ú.6 6,lue.nt . . . I t ,{,}., be.üeve.d tha.t 60.tz.v.:,t a.dd to the. .6 oil mo.tz.e n,i.,tJtoge.n tha.n

12.
t.he.y Jz.emove • • • The c.Mpe,t oó fe.a.ve/2, .:twig.6, baJLk, úc.., ha..6 a. ma.Jtk.ed eóóect
upon t.he c.ond.U,i,,on oó the .6oil undeJz.ne.a.t.h. The. wa.teJz. c.on.te.nt a.nd tempeJz.a.tuJte.
o ó t.h.e .6 oil Me paJLtic.ui..o.Jc1.y a.ó óe.c.ted. Ut:te1t not o nly .6 eJz.ve/2 to a.b.6 otr.b a.nd
1te,ta.,i,n mol6tuJte, d wo tenció to p!te.ve.n.t too Jz.a.pid .:tlr.a.n.6po1ta.t,i,on ó1tom be.fow
a.nd thl6 .6 eJz.ve/2 to JtúaJLd the. dltying o ó t.h.e .6 uJtóa.c.e. .6 oil • • • Wdhin the. 1.:, oil
w e.ló, de.c.a.ye.d fdteJz. ha.1.:, a.n a.me.füolta.ting e.ó óe.ct upon t.he. .6 oil .6tltuc.tuJte..
Whe.n mix.e.d wdh .6a.ndy .6oil, 01t9a.nic. ma.tteJz. te.nd!.:, to inc.Jte.a..6e. wa.te11..-hofdln9
c.a.pa.c.dy and to bind t.he. .6oil pMtiueJ.i toge.t.he.Jz.. I n a he.a.vy ua.y 1te9ion,
01t9a.nic. ma.twaf Jte.nde.M the 1.:,oil molte. 6JU.a.b.te, malte. pe.Jtme.a.bfe., be.tteJz.
a.e/ta.te.d, a.nd c.on.õe.que.n.t.ey malte. óa.voJta.bfe. to pfa.n.t 91towt.h • • • The. a.nnuaf
Jte.tuJtn 06 te.a.ó fdteJz. in a. we.U-óoJte/2:t.e.d hMdwood Me.a. ma.y be. c.on.6ide.1te.d a..6
a.p pito ma.te.f y .:two to n.6 pe!t a.Cll.e. . F 1to m :t.he. b e.ó :t. da.ta. a.va.ila.b.te. :t.hl6 a.mo un.t o ó
ha11.dwood fd:t.eJz. c.on.tMn.6 25 to 35 pound6 06 N,i,tJtoge.n, 12 :to 15 pound..6 oó
phMphoJU.c. a.ud, a.nd 5 :to 7 pound6 oó po:t.Mh . . . Ve.c.ompo.6,i_;t,i_on oó fd:t.eJz.
undeJz. a pU!te c.onióeJz. .6.:ta.nd ,Í_I.:, .6fow a.nd inc.ompfe,t".
Excetuando-se os estudos realizados por G.GISIN (1952),
esta abordagem da sucessão faunística no folhiço que ora aprese�
tamos não foi levada a efeito pela maioria dos especialistas em
pedozoologia. Por este motivo, apresentaremos a seguir uma bre
ve s1ntese dos principais trabalhos, que embora não tratem dire
tamente deste aspecto, tenham de alguma forma contribuído para o
melhor conhecimento das comunidades atuantes no processo de de
composição desta camada de folhas que recobre e protege os solos.
BORNEBUSCH(l930, 1932) e TR�GARDH (19 33) realizaram
importantes estudos sobre a mesofauna do solo na Europa, utiliza�
do o Funil de Berlese para as amostragens do folhiço.
JACOB (19 35, 1936a, 1936b, 1940) publicou informações
sobre a composição da mesofauna do solo em regiões dos Estados
Unidos da America do Norte.

13.
F R E N Z E L ( l 9 3 6 ) d i v i d i u a f a u n a d.o s o l o em t rê s g r u p o s
quanto ao seu tempo de permanência nos vãrios estratos: Geobion
tes - animais que passam toda a vida no solo; GeÕfilos - animais
que passam pelo menos uma fase da sua vida no solo; Geoxenos - ani
mais que acidentalmente são encontrados no solo.
K I N G ( i · 9 3 9 ) o b se r v o u q u e o s i n se tos d o s o l o p o .d em s e r
caracterizados pela pouca mobilidade, pequena resistência aos f�
tores físicos extremos e não são suceptiveis a inimigos naturais
como os parasitas especificas.
NIKIFOROFF (1941) vem reafirmar a importância de es
tudos minuciosos sobre pedozoologia, ressaltando que o solo e um
sistema dinâmico e harmoniosamente organizado, possuindo fatores
bióticos e abiÕticos característicos.
H.G ISIN (19 43) assinalou que as variações de pH do
solo podem limitar a ocorrência dos representantes da mesofauna,
sendo alguns específicos dos solos ãcidos e outros dos bãsicos.
Entretanto, ressalta ainda, existirem espécies de Collembola que
podem ser encontradas em ambas situações. O mesmo autor (19 49,
19 56) realizou estudos comparativos entre a fauna de Collembola
em ambiente natural e no laborat6rio tratada com fertilizantes.
M URPHY (1953) estudou a distribuição da mesofauna do
solo em diferentes tipos de florestas Europeias.
NEF (19 57) analisou a atuação da fauna do solo em
geral na decomposição do folhiço e sugere que juntamente a açao
desses animais ocorra uma espécie de atpque químico.
RAPOPORT (19 5 9, 1960) realizou trabalhos sobre a ati
vidade dos Collembola na humificação dos solos, acentuando a im
portância do grupo neste processo.

14.
E DWARDS & HEALTH (19 63) ressaltam a importância dos
organismos do solo na decomposição do folhiço, chamando a atenção
pa·ra o fato de que na ausência desses animais este processo fica
sensivelmente prejudicado.
SURGES (1971) observou que a atuação da mesofauna no
folhiço e variãvel em função da sua estrutura e do intervalo de
queda das folhas.
COINEAU (1974) visando facilitar a rãpida identific�
çao dos organismos do solo, apresenta em seu trabalho diversas
pranchas mostrando o aspecto de cada um desses grupos no momento
em que são observados ao microscõpio no laboratório.
N IGNOLET & LEBRUN (197 5) relacionaram a evolução de
cinco diferentes tipos de folhiço em função da decomposição pelos
microartropodos do solo.
Como jã ressaltamos anteriormente, praticamente nao
encontramos na literatura referências sobre a sucessão da mesofau
na do solo em folhiço. Entretanto, nos parece que as observações
sobre a distribuição vertical desta fauna nos vãrios estratos do
solo, apresentam, sob determinad_os aspectos, uma certa relação com
o s n o s s o s e s t u d o s . N o s s o l ·o s f l o r e s t a i s , a e ama da de f o l h i ç o, a p õ s
sofrer as primeiras etapas do processo de decomposição ainda na
superficie, vai sendo paulatinamente encoberta pelas folhas que
continuam a despreender-se da cobertura vegetal, passando assim
a incorporar-se ao horizonte do solo imediatamente abaixo deste.
Em nosso caso, mesmo não havendo a reestruturação do folhiço ini
eia , podemos considerar que com o decorrer da sucessão dos vãrios
grupos faunísticos e a respectiva evolução do estado de humifica
çao, o biótopo considerado vã apresentando gradativamente caracte

-,
15.
r í s t i c a s c o r r e s p o n d_e n t e s a o q u e o c o r r e ri a n a c o n t í n u a e s t r a t f f i
cação da camada inicial de folhiço em ambiente florestal.
G L A S G O W ( l 9 3 9 ) , G I S I N ( l 9 4 3 ) , M U R P H Y ( l 9 5 3 ) , BEL L I N GE R
(19 54) e KVHNELT (1 965) em estudos sobre a distribuição vertical
dos microartropodas do solo, observam que os níveis supériores
sao os mais abundantes e o primeiro desses autores, seguido por
JACOB (1936�, 1940) e BAWEJA (1939), constatou que alguns desses
grupos, como os Acarina e os Collembola, podem migrar de um estr�
to do solo para outro com o decorrer das estações do ano.
KEVAN (1968) e WAL LWORK (1970) observam que a fauna en . . -
contrada no folhiço é formada por indivíduos de maior porte, com
quetotaxia mais intensa e mais pigmentado.s do que os de horizontes
mais profundos do solo.
L EBRUN (1971) ao comparar a fauna de solos profundos
com a de superfície, conclui que a primeira e mais rica em formas
primitivas em relação ãs encontradas no folhiço.
WAL LWORK (1971) ressalta que em solos de bosques a
incidência dos Acarina e muito maior nos horizontes mais prõximos
dos níveis de fermentação ou nas zonas de transição entre o fol h..:!_
ço e o humus. Asso ia ainda este fato ao elevado grau de umida
de desses estratos .
DUNGER (197 5) estudou a distribuição vertical dos mi
croartropodos em vãrios estratos do solo.
No que tange aos estudos levados a efeito em flores
tas da Região Tropical, estes deixam bastante a desejar, pois r�
ras sao as suas evidências na literatura. WIL LIANS (19 41), est�
dando a fauna do Panamã, cita a ocorrência de todas as classes de

l 6.
artropodos: Miriapoda, Crustacea; Aracnida e Insecta. Sendo os
dois ultimas os mais abundantes, com os Acarina e os Collembola
perfazendo cerca de 80% da fauna total encontrada.
No Brasil, as considerações sobre a incidência de re
presentantes da mesofauna do solo referem-se quase que exclusiv�
mente a estudos de âmbito taxionômico. Sendo a maioria desses
trabalhos referentes aos Collembola e da autoria do Prof. Roger
Arle, nosso orientador na presente dissertação.
Da extensa lista publicada ao longo dos Ültimos A O
anos, em pelo menos duas oportunidades (ARLE'., 19 57 e 1959) ressa..!_
ta a importância ecolÕgica dos Collembola na composição estrutu
ral da mesofauna do solo. Em um desses estudos (ARLE, 1957) de
dica especial atenção a anãlise das formas biológicas e a sua va
riação em função do horizonte do solo em que são encontradas, as
pecto que nos deteremos na discussão deste trabalho.
Quanto aos demais grupos faunísticos aqui relatados,
cremos ganhar em importância a citação de L EMOS DE CASTRO (197 6)
que estudando exemplares de Amphipoda, coletados por nõs na mesma
ãrea onde ora realizamos este experimento, assinala pela primei
ra vez no Brasil a especie Talit�u� (Tafit�oide�) aífuadi
(Chevreux) , 18 96.

IV, RESULTADOS OBTIDOS

IV) RESULTADOS OBTIDOS
18.
IV.1) AN�LISE DA PARTICIPAÇ71:0 DE.TODOS OS GRUPOS NA SUCESS;li;O
Ao longo dos 360 dias, julho de 1977 a junho de-1978,
em que estudamos a sucessão ocorrida no amontoado de folhiço a
que nos referimos na presente dissertação, encontramos 19 grupos
faunísticos diferentes, perfazendo um total de 79. 916 espécimes
(Tabela I I) .
Excetuando-se os Collembola, que foram definidos ta
xionomicamente a nível de gênero, todos os demais grupos coleta
dos são considerados segundo as respectivas ordens como um todo.
A ocorrência dos grupos faunísticos na sucessao estu
dada, nao nos pareceu estar relacionada diretamente com alguns
aspectos do clima local durante o experimento. Entretanto, deve
mos considerar os altos índices de precipitação pluviométrica ocor
ridos em alguns períodos do desenvolvimento da sucessão (Tabela I
e Figura 25) .
Durante o mes de outubro de 1977, cerca de 90 dias
apos o início dos nossos estudos, constatamos que concomi tanteme!!_
te ao significativo aumento das populações de Isopoda e Amphip�
da houve uma brusca queda no numero de Collembola coletados (T�
bela I I I e Figuras 2,8 e 9) . Se sobrepormos estes dados aos ín
dices de p ecipitação pluviométrica no período imediatamente an
terior a este (setembro de 1977 - Tabela I) observamos que, emb�
ra a presença desses grupos deva estar relacionada com o estãgio
em que se encontrava a humificação do folhiço, tanto a alta inci
ciência dos dois primeiros como a excessiva diminuição deste Ülti

7
7
7
7
7
7
7
1
l
íl
l
l
l
19.
mo devem ter recebido alguma influência dessas precipitações. F�
vorãvel no caso dos Isopoda e Amphipoda e. desfavorãvel no que
tange aos Collembbla. O mesmo raciocinio nos parece vãlido para
as ocorrências de Isopoda em março de 1978, ã 270 dias do inicio
do estudo, e as precipitações inerentes a fevereiro do mesmo ano.
Outros fatores como temperatura e umidade relativa
(Tabela I e Figuras 23 e 24) exerceram pouca e atê certo ponto
desconsiderãvel influência na continuidade da sucessao. Por ou
tro lado, ê importante ressaltarmos que embora não atuando dire
tamente na seqüência percentual dos indices populacionais encon
trados, estes dois fatores mantiveram-se dentro de limites favo
rãveis as comunidades do solo. A amplitude térmica das medias no
período ficou entre 22° C e 29° C, variando as de umidade relativa
de 80% a 85% ( Figura 26) .
Os três mais importantes quantitativamente foram, nes
ta ordem, Collembola, Isopoda e Aca�ina , c�da um deles com res
pectivamente 33, 38%, 33,15% e 22,22% do total geral coletado (T�
bela I I) . Apesar disto, o periodo de atividade de cada um deles
na sucessao deu-se em fases diferentes, (Figura 14) .
Passados os primeiros 30 dias da formação do "fosso
de fol hi ço", observamos um niti do predomínio dos representantes
das ordens Collembola e Isopoda, seguidos a distância por Acarj_
na, Amphipoda e Dermaptera. Neste periodo o percentual de abun
dância relativa mensal de cada um deles esteve na seguinte ordem:
Collembola 6 4, 5 4%, Isopoda 22,88%, Acarina 3,73%, Amphipoda 3,22%
e Dermaptera 2, 41%. Esta proporção manteve-se relativamente cons
tante em igual periodo subseqüente a este, dando ao biótopo uma
aparente estabilidade faunística ao longo desses 60 dias em que
os primeiros decompositores ali se instalaram (Tabela IV) .

7
20.
A partir de setembro de 1977, com cerca de um trimes
tre inteiro de observações, ocorre um significativo aumento do
numero de espécimes de Amphipoda e diminuição no de Dermaptera,
Coleoptera (larva) , Diptera (larva), Dictyoptera e Araneida, sen
do que estes tornariam aos percentuais anteriores nas coletas se
guintes a esta.
A esta altura das observações, constatamos que o fo
lhiço jã apresentava sinais de avanço no processo de humificação.
As folhas inicialmente intactas, algumas ainda com coloração es
verdeada, jã apresentavam um aspecto amarronzado e atê certo po�
to sofrendo o ataque de fungos. No geral, o biõtopo apresentou
se bastante Ümido neste periodo. As precipitações pluviométricas ..
aumentaram significativamente (Tabela I, Figura 25) e a·.parti.r das
prõximas amostragens este fato pode ser notado no aumento da inci
ciência de alguns grupos.
Grandes percentuais de Isopoda e Amphipoda, 61, 71% e
30, 98% respectivamente do total mensal, caracterizaram as coletas
com cerca de 120 dias do início da sucessão, outubro de 1977. Foi
evidente neste periodo a queda repentina dos Collembola, pass�
ram de 44, 8% do total mensal no mês anterior para 2, 8 6% neste
(Tabela IV) . Os Acarina também apresentaram uma pequena diminuj
çao. Entretanto, como a sua participação nesta fase da sucessão
ainda é pequena este declínio nas suas populações deve ser pouco
significativo no estágio atual. Os demais grUpos mantiveram - se
inalterados, podemos ressaltar entretanto o aumento do numero de
Gastropoda, Diplopoda e Oligochaeta durante este período (Tabela
I I I, Figura 6, 12 e 13).

-,
7
21.
Nas amostragens seguintes a estas, duas em novembro
de 1977 e outras tantas em dezembro do mesmo ano, o folhiço apr�
sentou grande atividade de vãrios organismos do solo. Nesta fa
se, em que observamos a sucessão entre 150 e 180 dias do início
do processo, 17 grupos foram encontrados. Constatamos uma relati
va estabilidade nas populações de Collembola, Isopoda e Acarina,
estes últimos ainda com pequena participação nos percentuais de
abundância mensal. Apõs o pico alcançado em outubro de 1977,
120 dias do início do estudo, os Amphipoda apresentaram nesta fa
se um sensível declínio populacional, passando de 30, 98% para
16,38% e 10,21% do total mensal, respectivamente em outubro, no
vembro e dezembro do referido ano (Tabela IV, Figura 8) .
Este período apresentou também um relativo aumento
das populações de Diplopoda. Durante todo o estudo foi nesta fa
se em que seus espécimes ocorreram em percentuais mais signific�
tivos, 34, 02% e 38, 14% do total de Diplopoda coletados ( Tabela V,
Figura 12) .
A partir de janeiro de 1978, a sucessao dos grupos
faunisticos no folhiço apresentou características diferentes dos
primeiros seis meses observados, julho-dezembro de 1977. Consta
tamos prontamente, nas amostragens realiza as nesta fase, uma si�
nificativa incidência das populações de Collembola, com 80,19% da
fauna coletada (Tabela IV, Figura 2) .
Pela Tabela I I I, podemos observar que os Acarina apr�
sentam tambem um pequeno aumento neste período, embora as suas
maiores ocorrências viessem a recair em abril do mesmo ano (Fig�
ra 10) , Os Isopoda apresentaram-se estabiliz�dos e os Amphipoda
estiveram totalmente ausentes nesta fase. Apõs 210 ..dias, o biõtopo

22.
encontra-se bastante modificado na sua composição orgânica . Ap�
sar de poucas chuvas terem ocorrido neste mês de janeiro de 1 978
(5 6 mm - Tabela I) , o folhiço apresentou-se relativamente umido
e em adiantado grau de decomposição . A sua aparência geral era
de uma massa de matéria vegetal escurecida e levemente compact�
da, estando inclusive cerca de 20 cm abaixo do nível mãximo do
" fosso de folhiço 11 do início dos nossos estudos ( Foto 3) .
Ao completar 240 dias em fevereiro de 1978, o biõtopo
foi atingido pelas maiores precipitações pluviométricas em todos
os 12 meses analisados, junho de 1977 a julho de 1978 (Tabela I,
Figura 25). No folhiço que normalmente forma a camada protetora
dos solos das florestas tropicais, estas chuvas fatalmente teriam
"lavado " esta cobertura . Em nosso caso entretanto, cremos que,
apesar dos elevados indices (16 5 mm) , esta ação esteve limitada
ao aumento do humus e consequentemente maiores teores de algumas
substâncias resultantes da atividade dos microo�ganismos ali exi�
tentes. O "fosso" onde foi depositado o folhiço, jã descrito a!!_
teriormente, provavelmente assumiu características de retenção
deste em seu interior, fazendo com que a ãgua das chuvas escorre�
sem, por entre a materia vegetal, em direção ã horizontes mais
profundos .
Talvez como consequencia dessas precipitações pluvi�
métricas, observamos nesta etapa da sucessão uma queda nas pop�
lações de Collembola e um aumento nas de Isopoda, alem do ressur
gimento de alguns Amphipoda (Tabela III , Figuras 2, 8 e 9) . Es
tas variações nos Índ i ces desses três grupos torna-se mais evid e.!:1_
te ao completarmos 270 dias de iniciadas as amostragens . Nesta
fase, março de 1978, o declínio dos Collembola e bastante acentuado,

23.
80 , 19% - 44, 0 5% e 6,35% de participação na fauna mensal total ,
respectivamente em janeiro, fevereiro e março de 1978 (Tabela IV) .
Atingi ndo a 77, 15% da fauna coletada, os Isopoda fo
ram os mais abundantes neste estãgio da sucessao. E, da mesma
forma que no mês anterior, fevereiro de 1 978, os Amphipoda apr�
sentaram-se com cerca de uma centena de indivíduos (Tabela I I I e
Figuras 8 e 9) .
Em abril de 1978 completou-se 300 dias que os grupos
faunísticos vêm se sucedendo no folhiço reunido a cerca de 10 me
ses atrãs . Tendo se colocado em numero bastante inferior aos gr�
pos mais abundantes ate esta etapa dos nossos estudos, os Acari
na passaram a índices muitas vezes superior aos demais a ;partir
desta data. Ocorrendo em percentuais de abundância relativa me�
sal de 92,22%, 88,78% e 93,8%, respectivamente em abril, maio e
j unho de 1978, deixaram de ser o terceiro grupo quantitativame�
te, quarto em algumas oportunidades, para ocupar a liderança abs�
luta nesta fase final do processo sucessõrio por nõs estudado (T�
bela IV, Figura 10) .
A amplitude para o segundo grupo mais abundante nes
te período, os Collembola , foi da seguinte ordem : 17, 5 - 10,27 e
19,43 vezes maior ao completar respectivamente 300 ( a.bril de 1978)
e 360 (junho de 1978) dias de iniciada a anãlise da sucessão fau
nística no folhiço (Tabela I I I e Figura 14) .
Nestes Ül ti mos três meses de observações evidenciamos
também a brusca diminuição, atê o desaparecimento total, dos Is�
poda e Amphipoda, que ocuparam lugar de destaque na maior parte
das amostragens ao longo da sucessão.

2 4 .
Pela Tabela I I I observamos que outros grupos t i veram
queda acentuada ou se ausentaram por completo nesta fase f i nal do
experi mento. Dentre estes, podemos ressaltar : Coleoptera (lar
va e adulto) , Dermaptera, D i ctyoptera, Hemi ptera, Psocoptera, Th�
sanoptera, Arane i da, D i plopoda e Gastropoda (F i guras 1, 3,4, 6, 7,
11 e 12) . Outros, como por exemplo, Pauropoda, Embi optera e lar
vas de Lepi doptera ocorreram em percentuai s mu i to pequenos ao lon
go dos meses anali sados.
Das amostragens reali zadas quando a sucessao estuda
da datava de 210 d i as do seu i níci o até os 360 d i as em que encer
ramos as observações (de jane i ro a junho de 1978), o folhi ço de
posi tado no i nteri or do "fosso" sofreu v i sualmente um s i gin i f i ci
t i vo processo de hum if i cação. Nesta oportuni dade, cess adas as
amostragens, constatamos que a matéri a orgâni ca apresentavJ. - se
quase que totalmente decomposta no que tange a parti c i pação da
mesofauna do solo . De coloração enegreci da e com odor caracterís
ti co de humus molhado, o aspecto geral do ."fosso de folh iço " é de
que a matéri a vegetal que para ali fo i carreada passou por exte�
s i vos mecan i smos de tri turação e compactação, no qual lhes fo i
reti rado quase toda a matéri a v i va de que era prov i da, restando
apenas um conjunto de restos orgân i cos fortemente envolv idos por
fungos e suceptível ao ataque de bactér i as e outros orga n i smos al
tamente saproxíli cos � Foto 4) .
Ate alcançar este grau de decompos i ção do fo l. h i ço, os
grupos fauníst i cos se sucederam em d i ferentes períodos a o longo
dos 360 d i as estudados. A Tabela V nos permi te anali sar em qual
período, durante a sucessão ocorr i da, cada grupo apresen tou os
ma i ores percentua i s de i nci dênc i a.

-,
25.
pentre os mais abundantes, observamos que os Collemb.2_
la apresentaram uma nítida preferência pela fase em que o folhiço
encontrava-se a cerca de 210 dias do início da sucessão. Embora,
no início do experimento os seus percentuais tenham sido também
bastantes significativos: 14,22% - 10,47% e 11,�2% respectiváme.!!_
te ã 30, 60 e 90 dias do inicio das observações ( Figura 2) .
Os Acarina por outro lado, somente tiveram uma particj_
pação mais incisiva no final da sucessao, com 27,58% - 21,39% e
23,08% nos três últimos meses analisados, isto e , transcorridos
300, 330 e 360 dias de iniciado o estudo (Figura 10) .
Amphipoda e Isopoda demonstraram as suas preferências
no mes de outubro de 1 977, quando o folhiço jã era atacado pelos
decomponedores a cerca de 120 dias. Neste período as incidências
a t i n g i r a m 5 3 , 7 7 % p a r a o p r i m e i r o e 2 6 , 7 3 % .P a r .a o s e g u n d o , d o t o
tal encontrado ao longo do experimento para esses grupos. Os Isa
poda ainda voltariam a ocorrer em elevados percentuais, 20,9%,
apos transcorridos 270 dias de observações, março de 1 978 (Fig�
ra s 8 e 9 ) .
Os demais grupos coletados, embora tenham particip�
do do total geral em numero inferior aos quatro anteriores, tam
bem apresentaram maio res incidências em determinadas fases da su
cessão e desta maneira atuando diferentemente no processo de de
composição do folhiço.
Os Coleoptera ( larvas e adultos) , Dermaptera e T hys�
noptera instalaram-se no biÕtopo logo ri os primeiros 60 dias do e�
perimento, julho e agosto de 1977. Os seus percentuai s neste p�
ríodo foram, nesta ordem, para os 30 e 60 dias iniciais : 12,5% e

-,
26 .
2 5 , 5 9 % em Coleoptera adultos (aihda ocorreram com 1 0 , 4 1 % e 1 3 , 6 9%
apõs 2 4 0 e 2 7 0 dias de estudo) e l 2 1 , 0 5 % e 1 2 , 7 8% para o estãgio
de larva deste mesmo grupo, 3 9 , 3 3% e 3 8 , 7 8% em Dermaptera e 22 ,85%
e 5 1 , 4 2% em Thysanoptera, do total coletado de cada um desses gr�
pos (Figuras 1, 3 e 4) .
Na realização das amostragens com cerca de 1 2 0 e 1 5 0
dias de estudos encontramos os mais altos índices de Oligochaeta,
2 2 , 0 9 % aos 1 2 0 dias (outubro de 1 9 7 7) , e Gastropoda, 3 1 , 7 9 % nes
te mes e 2 0 , 6 3% ã 1 5 0 d i as (novembro de 1 9 7 7) de todo experime�
to (Figuras 6 e 1 3) .
Dois períodos marcaram as maiores ocorrências de Dic
tyoptera, agosto de 1 9 7 7 e dezembro do mesmo ano, com 6 0 e 1 80
dias de observações. No primeiro caso estas marcas alcançaram
2 1 , 3 3% e no segundo 2 6 , 6 6% do total coletado para este grupo (Fj__
g u ra 4) . -� · ..
Dentre as larvas de Diptera as maiores i1 1cidências
( 2 1 , 2 6 %) foram notadas 2 1 0 dias apõs o início das amostragens e
no trimestre final para os adultos ápteros que apresentaram 37 , 5% ,
2 5 % e 2 0 % respectivamente com 3 0 0 , 3 3 0 e 3 6 0 dias sucedidos ( Fj__
gura 5).
O período compreendido entre 2 1 0 , 2 4 0 e 2 7 0 dias de
evolução da sucessao, janeiro, fevereiro e março de 1 9 7 8 , foi o
preferido pelos Hemiptera e Psocoptera, que nesta fase obtiveram
os seus maiores percentuais de ocorrência com respecti·vamente
2 6 , 9 2% - 3 0 , 7 6 % e 3 0 , 7 6 % para primeiro e 2 5 , 2 5 % - 3 0 , 3 0% e 3 3 , 3 3 %
para o segundo (Figuras 6 e 7) .

27.
Araneida e Chilopoda participaram do folhiço em maior
abundância a partir de fevereiro de 1 978, quando e.st.e . se encontr�
va com 24 0 dias de iniciado o processo de decomposição . Esta in
cidência dos Araneida foi observada entre 240 e 27 0 dias, com pe!
centuais de 19% e 21% , enquanto os Chilopoda apresentaram-se em
níveis semelhantes de fevereiro de 1 978 ã abril do mesmo ano, sen
do respectivamente 24% com 240 dias, 28% com 27 0 dias e 20% com
300 dias de realizadas as primeiras amostragens (Figuras 11 e 12) .
A 1 guns grupos como L epi doptera (Figura 7 ), Embi optera
e Pauropoda, principalmente esses dois ultimes, tiveram particl
pações muito pouco significativas ao longo dos 36 0 dias analisa
dos . Restringiram-se a somente 8 espécimes no caso dos L e pido�
tera e apenas 1 espécime para cada um dos outros dois grupos. Den
tre os L epidoptera, todos em fase de larva, 62, 5% dos espécimes
ocorreram em abril de 1978 com cerca de 300 dias de experimento .
Os unices Embioptera e Pauroptera, ocorreram respectivamente com
30 dias (julho de 1977) e com 270 dias (março de 1978) de estudos.
IV. 2) ANÃL ISE DA PART ICIPAÇ�O DA FAUNA COL EMBOL Õ G ICA NA SU
CESS�O
Encontramos ao longo dos 36 0 dias em que observamos
a sucessao no 11 fosso de folhiço 11 17 espécies de Collembola perf�
zendo um total de 26 677 espécimes, o mais abundante de todos os
grupos coletados (Tabela I I) . Devido a grande variedade de for
mas biológicas (Prancha B) e por ser o grupo de maior afinidades
com as atividades desenvolvidas em nosso laboratório no Museu Na
cional do Rio de Janeiro, os Collembola terão a sua participação
na sucessão analisada segundo os gêneros identificados ao longo do processo.

-,
2 8.
O genero Lepidocy4tu.6 foi nitidamente o mais abundan
te, tendo sido encontrado tanto no início da sucessão como nas
ultimas amostragens. Dos Collembola coletados, 6 6,39% pertenciam
a este gênero, tendo sido seguido a distância pelos Ibotoma com
25, 46% e por Cyphode4ub com 3, 75%.
Nos primeiros 30 dias de observações, constatamos que
o s e s pé c i me s d e C o 1 1 em b o 1 a e r a m o s m a i s a b u n d a n t e s . Dentre esses ,
as maiores participações ficaram por conta de Lepidocy4tub e l b o
toma, respectivamente com 5 8, 87% e 40, 59% das incidências neste
mês de julho de 1977 (Tabela VI I, Figuras 19 ·e 20). Além destes,
outros cinco generos também estiveram representados neste início
de decomposição do folhiço, a saber : A4l u ia - 1,23%, B4ach y.6 tE._
mella - 2, 95%, Cyphode4u.6 - 0,23%, Fo l.6omide.6 - 0,05% e Sminthu
4ide.6 - 0,05% (Tabela VI I, Figuras 16,17, 18 e 22) .
Nas amostragens realizadas ao longo dos 30 dias que
se seguiram, agosto de 1977, deparamos com a introdução de repr�
sentantes de F4iebia e com a ausência dos espécimes de Sminthu4i
de.ó , tendo os demais generos mantido as proporções do período an
terior (Tabela VI) . Devemos ressaltar entretanto, o ligeiro au
mento nas populações de Cyphode4u.6 e o declínio dentre os I.6oto
ma , que passaram respectivamente de 0,23% e 40, 59% no mes ante
rior para 7, 89% e 19 , 57% neste ultimo (Tabela VII) .
Ao completarmos, no final de setembro de 1977, o pr!
meiro trimestre de estudos da sucessão no folhiço, constatamos o
maior numero de espécies durante todo o experimento, 15 das 17 co
letados ao longo dos 360 dias de estudo (Tabela VI) . Contudo, ain
da desta feita, a maior abundância ficou por conta do gênero Le
pidocy4tu.6 com 84, 60% do total de espécimes coletados neste mês.

-,
29.
o segundo mais incidente foi Cyphodenué com 9, 0 3 %, ficando os de
mais muito abaixo desses percentuais (Tabela V I I) .
Foi tambem durante esta fase da sucessão que observ�
mos as ünicas ocorrências de espécimes de Campylothonax e de Vi
enanoeentnué { Tabela V I) .
Como jã nos referimos por ocasião da anãlise dos gr�
pos na sucessão no tõpico anterior a este, o biÕtopo foi bastante
castigado por intensas precipitações pluviométricas ao completar
120 dias de observações em outubro de 1977 . Dentre os Collembola
esta atuação é sentida nao sõ no numero absoluto de espécimes co
letados neste período, mas também pela ausência de alguns dos 9!
neros que jã haviam sido encontrados nas amostragens anteriores.
Apesar desta sensível diminuição, constatamos pelas Tabelas VI e
V I I que pelo menos o genero Bnaehy-6tomella não foi significativ�
mente afetado, pois além de um pequeno aumento no numero de esp!
cimes (Tabela VI) a sua participação percentual na sucessão den
tre os Col lembola passou de 1, 0 5% no mes anterior para 11, 5 5% ne�
te ultimo (Tabela V I I) . Por outro l ado, embora a Tabela VII nos
mostre uma maior participação percentual também para o genero e�
phoden u-6 neste fase do experimento, a observação dos valores ab
s o 1 u t o s r e p r e s e n t a d o s n a Ta b e l a V I , no s p e rm i t e c o n s t a t a r um a c e.!:!.
tuado declínio no seu numero de espécimes, cerca de cinco vezes
menos.
Em novembro de 1977 , com 150 dias de observações, as
amostragens apresentaram a ocorrênc i a pela primeira vez em nosso
estudo do gênero Anlea. Esta incidência contudo, restri'ngiu - se
a poucos espécimes que concederam-1 he urna participação de 0, 30%
entre os Collernbola coletados nesta etapa da sucessão no folhiço

7
3U .
(Tabela VI I, Figura 16) . -
Os demais generos mantiveram-se nas prE_
porçoes jã constatadas em amostragens anteriores, com o predomi
nio dos Lepidocy1t:tuh - 5 9,22%, seguido pelos I h o :toma - 17,70%, C�
phodeJz.U-6 - 13, 16% (Tabela VI I) .
A p õ s l 8 O d i as d e anã l i se d a s u c e s s a o , c o n s ta ta mos que
o biõtopo apresentava-se mais uma vez com um elevado numero de es
pecies de Collembola, 13 dentre as 17 encontradas em todo o exp!
rimento (Tabela V I) . Os gêneros que mais ocorreram neste mês de
dezembro de 1977 ainda foram LepidocyJz.:tU-6 - 56, 0 5% e I.t, o:toma -
33, 81%. Entretanto, observamos uma ligeira queda nos espécimes
de CyphodeJz.U-6 em relação aos 30 dias anteriores, de 13, 16% para
3,6 5% (Tabela V I I, Figuras 16 e 17) .
O início do 2 9 semestre de amostragens no "fosso de
f o l h i ç o II f o i ma r c a d o p e l a g r a n d e i n c i dê n c i a d e e s p e c i me s de Co 11 e!!!_
bola, que participaram com 80,19% da fauna coletada nesta fase em
que o biõtopo encontrava-se com 210 dias de estudos . Pela Tabe
la V I I I e Figuras 19 e 20 constatamos que os dois generos mais
abundantes ao longo de todo experimento, Lepidocy1t:tuh e 1 .6 0:toma ,
obtiveram os seus maiores percentuais justamente neste período,
respectivamente com 35,68% e 38, 63% dos espécimes coletados nes
te mês de janeiro de 1978. Ainda pela Tabela V I I I, percebemos
que o gênero Cyph odeJz.u.6 , o terceiro mais abundante, não acompanhou
os dois primeiros tendo inclusive significativa queda nesta eta
pa da sucessão no folhiço. Por outro lado, os representantes de
F1tie.6ia aumentaram a sua participação nesta oportunidade, muito
embora esta incidência não nos pareça proporcional mente compat�
vel com os gêneros citados anteriormente (Tabela VII I, Figura 18).

!
7
7
-,
3 1 .
Como jã nos referimos anteriormente, o mes de feverei
ro de 1978 foi marcado pelas maiores precipitações pluviométricas
de todo período estudado. Estas precipitações e o jã elevado es
tado de decomposição em que se encontrava o biótopo, fizeram com
que alguns . espécimes de Collembola diminuí ssem significativame�
te a sua ocorrência, c hegando a desaparecer totalmente em alguns
casos. Esta queda torna-se bastante evidente dentre os Lepidoc.y!!:_
.tu-0 que embora continuassem a ser os mais abundantes (Tabela VII)
tiveram uma queda de 35, 68% no mês anterior para 12, 02% neste Ül
timo {Tabela VII I, Figura 20) no que se refere a sua incidência
em todo o experimento. Fenômeno semelhante observamos nas pop�
lações de I-00.toma e B 1tac.hy-0.tomeffa que passaram respectivamente
de 38, Q;3% e 28, 0 4% para 6,81% e 7,35%, (Tabela V I I I) . Por outro
lado, ao atingir esses 240 dias de estudos em fevereiro de 1978,
o biõtopo pareceu-nos tornar-se favorãvel aos gêneros A1tfea , C�
phode1tu-0 , Fof-0omide-0 , Neefu-0 , P.teno.th1tix , Smin.thu1tide� e Sphae1tf
dia , que apresentaram sensivei s aumentos nesta fase do processo
sucessõrio (Tabelas VI, V I I I, Figuras 16, 17,18,21 e 22) .
A partir do mês de março de 1978 até final das nossas
observações em junho do mesmo ano, correspondendo as amostragens
com 270, 300, 330 e 360 dias transcorridos desde o início do ex
perimento, as populações de Collembola foram declinando acentua
damente· a sua participação na sucessão. Apenas alguns generos,
como Lepidoc.y1t.tu-0 , f a o.toma , F1tie-0ia , Cyphod e.1tu-0 , A1tfe-0ia e. N e. e.fu-0
conseguiram apresentar representantes atê o final do estudo. As
sim mesmo, se analisarmos as suas ocorrências em fases anteriores
a esta, constataremos que essas incidências tendiam ao desapar�
cimento total (Tabela V I I I e Figuras 16, 17, 1 8, 19,20 e 21) .

--, 1
7
32.
IV. 3) FORMAS B IO L O G ICAS E R EG IM E A L IMENTA R DOS COL L EMBOL A
Os Collembola formam um grupo zoológico extremamente
diversificado , apesar de intimamente ligado ao solo e seus ane
xos. São encontrados desde o litoral marinho até milhares de me
tros de altitude.
Hã uma primeira classificação ecológica, de acordo
com a sua distribuição vertical , distinguimos espécies atmobiõti
cas - acima da superfície do solo (vegetação , etc . . ) , epiedãfj_
cas - superfície do solo (folhiço) , hemiedãficas (povoam camadas
de folhiço e detritos vegetais) e euedãficas (povoam as camadas
mais profundas do solo atê onde as raizes alcançam) .
Estas quatro formas representam um gradiente de esp!
cialização edãfica. Partindo das espécies superficiais de tama
nho relativamente grande , com apêndices bem desenvolvidos (ant!
nas , patas e furca) , as vezes hipertêlicas , pigmentadas e com o
numero mãximo de cornêulas (8+8) , passa-se a espêcies menores , com
apendices mais curtos, atê chegar a formas muito pequenas, total
mente despigmentadas, com olhos reduzidos até a ausência total e
assim como a supressão da furca (Prancha B) .
Em nosso caso , os Collembola foram importantes indi
cadores do estado de humificação, 22 677 espécimes (33,38% do to
tal, sendo o grupo mais abundante) foram coletados durante o ano.
As Tabelas V I , V I I e V I I I e as Figuras de 16 ã 22 , mostram a evo
lução da população durante o estudo. Pode-se notar um decresci
mo geral das espécies hemiedaficas durante o experimento (que o�
tiveram um forte pico durante a estação chuvosa) - 'Btz.a.c.hy.6tome,U,a.,
Cyphode.Jtu.6 , 1 .6 0:toma. e. L e.p..i..doc.yn:tu.6 . Por outro lado , espécies com

7
1
3 3 .
tendência mais edã fica permaneceram com incidências relativamen
te significativas at e as ultimas amostragens - Atr..le�ia , Ftr..ie�ia ,
Neelu� e P.te n.o.thni x.
Existe assim uma certa relação entre a dist ribuição
vertical - os estratos inferiores constituindo-se num estãgio mais
avançado do processo de humificação - e a sucessão aqui . estudada,
onde esta evolui para um estado cada vez mais adiantado de decom
posição é humificação do folhiço inicial, acompanhada pela dimi
nuição do numero de espécimes de Collembola.
Os Collembola alimentam-se essencialmente de :hifas de
fungos � algas unicelulares, grãos de pÕlem, esporos, materia ve
getal mais ou menos decomposta, excrementos de outros animais e
bactérias. Existe uma especie (Smi n..thutr..u� vitr..idi� ) , que na Euro
pa, Ãfrica, Austrãlia e Argentina come folhas verdes de alfafa,
trevo e lotus, podendo constituir-se numa praga. Entretanto, a
especificidade alimentar dos Collembola decorre das exigências eco
lÕgicas e do micro habitat ocupado.

7
V , DI SCUSSAO

7
Y ) D IS CUSSÃO
35.
Em nos sos estudos mostramos o desenvolvimento de uma
biocenose limitada num amontoado de folhiço e detritos vegetais
diversos durante 12 meses consecutivos. Os invertebrados comp�
nentes desta biocenose encontram-se em dependência mutua, em equ.:!_
librio dinâmico e permanente evolução.
Podemos falar em s uces sao j ã que observamos uma sequê.!!_
eia de comunidades acompanhando as modificações do meio, sendo e�
tas, em grande parte, causadas pela ação destas mesmas comunida
des. As relações entre os divers os animais envolvidos são varia
das e as vezes antagônicas . A maior parte da fauna do solo aqui
estudada é constituída de espécies detritivoras que ingerem gra.!!_
de quantidade de material orgânico somente em parte digerido, se.!!_
do os excretas profundamente modificados fisicamente e em condi
ções Õtimas para serem decompostos quimicamente pelos fungos e
bactérias. Mas hã também espécies predadoras e parasitas e, no
quadro natural de equilibrio dinâmico onde se desenvolvem lado a
lado, ocorrem reações antagônicas e reações benéficas , competições
e as sociações.
Antes de tudo trata-se de uma suces sao secundãria ra
pida, uma microsuces são, que ocorre ao se formar uma biocenose no
acumulo de material morto. Suces são semelhante podemos observar
em um tronco de ãrvore caido, em excrementos de herbívoros ou em
cadãveres de vertebrados abandonados sobre o solo. Todas es sas
microsuces sões contribuem para a formação do humus.
Em nos so caso realizamos propositadamente a maneira
mais simples de se formar um composto de tipo "leaf mold", sem

-,
36.
estratificação e sem adição de fertilizante, como qualquer j ardl
neiro amador faria no fundo do seu quintal . Apenas tomamos os
cuidados necessãrios para não haver pertubações, salvo as nossas,
durante o período de experimentação. Isto nos permitiu compar�
çoes interessantes com a formação natural de "leaf mold " na mata,
onde se forma uma estratificação vertical, e podemos observar a
e v o l u ç ão d a b i o c e n o s e em r e l a ç ão a o t em p o p a s s a d o e as i n fl uê nci as
climatolõgicas, sendo este o nosso principal obj etivo.
São numerosos os trabalhos sobre sucessao ecolõgica
a longo prazo, principalmente em Collembola. Neste caso, ê toma
da como escala de tempo a sucessão da vegetação na impossibilid�
de de se obter uma escala absoluta ( MURPHY, 1 9 53) . Em certos ca
sos foram estudados as populações edãficas em plantações de coni
feras de idade conhecida ( GRETSCHY, 1949 e JOHN, 1944) . Este .ül
timo a·utor achou um pequeno aumento de Collembola e um muito maior
de Acarina ao longo de 78 anos. Diversos trabalhos foram reali
zados acompanhando a evolução de turfeiras altas. S-endo este,s es
tudos de grande interesse e de grande complexidade sendo difí cil
isolar o fenômeno sucessão. Ao contrãrio, no caso de um acumulo
de material vegetal morto onde o meio se altera rapidamente (ml
crosucessão) a verdadeira sucessão ê apenas pertubada (ou molda
da) pelas variações do ciclo anual .
Somente dois trabalhos nesta modalidade constam da li
teratura (H. GISIN, 1 949 e G. G IS IN 1 9 52) , realizados na Suí ça e
limitados aos Collembola. Em nosso estudo abordamos pela pri mei
ra vez este aspecto em clima tropical e, entre outras coisas� apo�
ta um caminho a seguir em um campo fértil de pesquisas, qual seja
o estudo cada vez mais aprofundado das microsucessões em :compo�
tos, variando o material original, acrescentando fertilizantes

37.
diversos e outros elementos, utilizando métodos precisos de ava
li ação e com um tratamento taxionômico adequado e rigoroso da fau
na, essas pesquisas poderão dar resultados de alto valor científi
co e prãtico, revelando alguns aspectos fascinantes da vida no
solo.

-VI, CONCLUSOES

VI) CONCLUSffES
39 .
Ao longo dos 3 60 dias consecutivos, julho de 1977 a
junho de 1978, que estudamos a sucessão faunística ocorrida, ob
servamos que quatro grupos participam decisivamente do processo
de decomposição do folhiço : Collembola, Isopoda, Acarina e Amphj_
poda, com respectivamente 3 3, 58%, 3 3,15%, 22,22% e 8,27% da fau
na total coletada (Tabela I I) .
Inicialmente, o folhiço ê atacado por populações de
Collembola e Isopoda em primeiro plano e pelos Acarin� Amphipoda
e Dermaptera em numeras inferiores a estes dois (Tabela IV) .
A partir do terceiro mes de observações ocorre um si1
nificati vo aumento no numero de Amphipoda, que passam a dividir
com os Collembola e os Isopoda os maiores percentuais de incidên
eia no folhiço (Tabela IV) .
As condições do "fosso de folhiço " apos 120 dias de
estudos favorecem sobremaneira as presenças de Isopoda (61,71%) e
Amphipoda (30, 98%) no processo de decomposição (Tabela IV) .
O início do penúltimo trimestre de estudos da suces
sao, marca a supremacia quantitativa quase que total de represe�
tantes de Collembola, 80,19% da fauna total neste período (Tab�
la VI) .
Fato semelhante ocorre com os Isopodas (77,15%) ã 270
dias de iniciado o estudo da sucessao.
O ultimo trimestre ê caracterizado pelo acentuado au
menta de espécimes de Acarina, com 92,22%, 88,78% e 9 3, 8% respe�
tivamente aos 300, 330 e 360 dias transcorridos no processo sucessório.

-,
40.
A anãlise da Figura 1 5, nos permite concluir que ª P!
sar dos Collembola e os Acarina terem grande participação na de
composição do folhiço, a · atuação desses grupos e feita em perí�
dos diferentes. Enquanto os Collembola ocorrem principalmente
n a s f a s e· s em q u e o f o 1 h i ç o n ão s e e n c o n t r a a i n d a m u i to h um f f i c a
do, os Acarina tem uma nítida preferência pelas etapas finais do
processó. fste fato inclusive, tem dado margem a certas distor
ç5es na literatura sobre qual dos dois grupos e o mais abundante
no solo. Pois, estudos levados a efeito em épocas diferentes ou
considerando apenas a fauna de determinado horizonte do solo � Pr!
sentam resultados tendenciosos para o grupo que na oportunidade ei
tiver mais bem identificado com as condições gerais do biõtopo.
A única espécie de Amphipoda encontrada no 11 fosso de
folhiço 11, Ta..titll.u.6 (Ta..ti:t.11.oid e..6 ) a..t.tua.udi, assim como as espécies
de Isopoda coletadas, aumenta significativamente o seu numero de
espécimes apõs a incidência de grandes chuvas (Tabela I e I I I).
Muito embora o ciclo anual de precipitações pluviomI
tricas, temperatura e umidade atue diretamente na incidência da
mesofauna, em nossos estudos, a sucessão ocorrida esteve ligada
principalmente ao regime alimentar exercido por cada grupo e o
corres·pondente avanço do grau de decomposição do folhiço ao lon
go do experimento.
Na maciça presença dos Collembola ao completar 210
dias de observações em janeiro de 1978, sal ienta-se a incidência
dos gêneros L e.pido �y11.tu.6 e I.õotom a . , qu� alem de serem os mais
abundantes em todo o experimento com respectivamente 17 712 e
6 7 92 especimes dentre os 26 677 Collembola coletados, foram os
responsãveis diretos pelos elevados Índices durante este
do (Tabela VI) .
perí�

-----,
4 ) .
Os gêneros Cyphode1tu.1.i , 1 .õ o.toma e Lepido c.y1t.tu.1.i, sao os
únicos Collembola presentes em todas as amostragens durante
360 dias de observações { Tabela V I) .
os
Excetuando-se o gênero Cyphode1tu.1.i , que apresenta as
maiores incidências nos primeiros 120 dias do processo de decom
posição do folhiço, os _ d�mai s� , obtiveram incidência superior
a 10% em pelo menos um dos 12 meses analisados (Tabela VI I) , a s�
ber : B1ta. c.hy1.i .tó.me.l.la. , 1 1.i o.toma e Le.pido c.y1t.tu.1.i , ocorrem preferencial
mente durante as fases intermediãrias, entre 150 e 240 dias, no
vembro de 1977 ã fevereiro de 1978 (Tabela V I I I, Figuras 17 , 1 9,
20) .
Concomitante a sucessao dos grupos no folhiço e ao
consequente avanço da decomposição, ocorre também a variação adaf
tativa das formas biolõgicas dos representantes da mesofauna nas
diferentes etapas deste processo .
Pelos Grafices e Tabelas, conclui-se que a sucessao
faunística observada apresenta um paralelismo com a distribuição
vertical em ambientes florestais. Os grupos presentes em nossas
coletas iniciais, são os encontrados nos estratos superficiais dos
solos florestais e, a medida que avançamos as amostragens ao lon
go dos meses, deparamos com a maior incidência daqueles que nor
malmente habitam os horizontes mais profundos desses solos.

VI I , RESUMO

43 .
V I I ) RESUMO
,/" '· . . , Visando estabelecer a sucessao existente entre os gr�
pos formadores da mesofauna do solo durante a decomposição do fo
lhiço, coletas sistemãticas foram realizadas em um biõtopo previ�
mente estabelecido, por ngs denominado de "fosso de folhiço '' , du
rante 12 meses consecutivos - jul ho de 1977 i junho de 1 978.
Ao longo dos 360 dias de estudos identificamos 19 gr�
pos faunísticos, �erfazendo um total de 7 9 916 especimes . Para
tal, realizamos 24 amostragens com um intervalo de 15 dias entre
elas.
Pelos dados, constatamos que, nesta oportunidade, os
grupos mais diretamente relacionados com a decomposição do folhi
ço foram respectivamente : Collembola, Isopoda e Acarina.
Os 19 grupos faunisticos encontrados sao definidos
taxionomicamente a nivel de ordem. Dentre estes, os Collembola
mereceram uma análise mais pormenorizada e, desta forma, os seus
especimes encontram-se ident i ficados segundo os generos
dentes, 17 ao todo . Dos quais, destacamos como os mais
tes Lepidocy�tu� e I�otoma.
correspo.!:!_
abundan
Estabelecemos os períodos de atuação de cada grupo
na sucessao e discutimos a diversidade das formas biolÕgicas en
contradas em relação ao período do processo de decomposição em
que ocorreram.

V I II , SlH1ARY
7

45.
VI I I ) SUMMARY
Aiming at the establishment of an existing succession
between the soil mesofauna developing groups during the decomposition
of the litter, systematic sampling were conducted in a previously
selected biotope - called by the author "fosso de folhiço 11 (*) -
during 12 consecutive months (�une 1977 to July 1978) .
Throughout 360 days of studies, we identified 19
faunistic groups, adding up a total of 79 916 specimens. To
accomphish that, there were made 24 samplings with intervals of
15 days.
From the data obtained, we realize that, in the
opportunity, the most directly related groups with decomposition
of the litter were respectively: Collembola, Isopoda and Acarina.
The 19 faunistic groups found were taxionomically
defined at level order. Among them, Collembola deserved a more
detailed analysis and, consequently, their specimens are identified
in accordance with the corresponding genus, 17 in the whole.
Out of these, we considered Lepidoey�tu� and I �otoma as the most
abundant.
We have established the periods of activity of each
group in the succession and we have discussed the diversity of
biol.ogica.l forms found in the relation to the period of the
decomposition process in which they have occurred.
(*) - hole full of litter.

7
IX , REFERENC I AS BIBLIOGRAFICAS

4 7.
IX ) REFER[NCIAS BIBLIOGRÃFI CAS
A RLt , R. , 1957. Alguns aspectos da ecologia e biogeografia dos
Collembola. Ata4 S o e.. Biol. , Rio de J aneir o , 1 ( 6) : 33-35.
, 1959. Generalidades e impor tâ ncia eco l Õgi ca da 0 rdem ----Collembola. AtrH S o e.. Biol. , Rio de Janeiro , 3 ( 2) : 4 - 7.
BAWEJ A , K. D. , 1937. The calculation of s oil population figures.
J. Animal E c. ol. 6 : 266 -26 7 .
, 1939. Studies of t he soil faun a , wit h s pecial reference ----to t he recolonization of s terilized s oil. J. A nimal Ec.ol. 8 :
120 - 161.
BELLIN GER , P. F. , 19 54. Studies of soil fauna wit h special reference
to t he Collembola. Buli. C o n n. Agfl.i c.. Exp , S:t,an. 583 : 1- 67.
BERTHET, P. , 195 5. L ' Echan tillon nage de la litie re des fores ts
pour l ' etude quan t itative des populations d ' acariens et
collemboles. Ann. S o e.. R. Z o ol. B e.lg. 8 5 ( 1) : 5-22.
BORNEBUSCH , C. H. , 1930. The fauna of fores t s oil. FoM.th FoM og4 v.
Va nm . l l : l -2 2 4 .
----'---- , 1932. Das Tierleben der Waldboden. F ofl.4 t. wi4 4.
C e.n:t,fl.albl. 54 : 253-266.
BURGES , A. , 197 1. " La decompos iciõ n de la materia orgãn ica en el
suelo" , l n : BURGES , A. & RAW , F. , Biologia del Suelo. ·Ed.
Omega S/ A , Barcelona. 557 - 573.

48.
CLARK , L.R. , GEIER , P.W. , HUGHES � R.D. & MORRIS , R. F. , 19 74.
The ecologica of insect population in theory and practice.
Science Paparbac ks , Methuen & CO. Ltd. , London. XIII : 232 pp.
COINEAU , Y. , 19 74. Introduction a l ' étude des microarthropodes du
s ol , et de ses annexes. Doin rditeurs , Paris. 117 pp.
DARWIN , e . , 1881. The formation of vegetable moü; d through the
action of worms , with observations of their habits. Murray ,
London. 326 pp.
DELAMARE DEBOUTTEVILLE , C. , 1 9 51. Recherches écologiques sur la
microfauna du s ol des pays temperes et tropicaux. The.ó e..ó Fa.c..
S c_,i_ • U n.,i_ v • , P Q.fl..,Í_.ó • ( A ) 2 4 l 6 : l -3 6 O •
DOWDY , W.W. , 1944. The influence of temperature on vertical
migration of invertebrates inhabiting different s oil tipes.
E c.o lo g y. 25 : 44 9 - 4 60.
DUNGER , W. , 19 7 5. " On the delimitation of soil microarthropod
coenoses in time and s pace" , l n : Progress in Soil Zoology.
Organized by the Soil Zoology Committee of the International
Society of Soil Science , 59 International Colloquium on Soil
Zoology. Publi. Hous e of Czechos lovak Acad�my of Sciences ,
Prague. : 43-4 9.
EDWARDS , C.A. & HEATL H , G.W. , 19 63. " The role of s oil animals in
breakdown of leaf material " , l n : DOERSEN , J. & DRIFT , J. van
der , Soil Organisms . North-Holland Publ. , CO. , Amsterdam.
: 7 6 -84.
FARB , P. , 19 59. Living earth. Constable and Company Ltd., London.
178 pp.

-,
7
FORD, J. , 1937a. Fluctuations i n natural populati on of Collembola
and Acari na. J. Animal Eeol. 6: 9 8-111.
, l 9 3 7 b . S oi l e o mun i t i e s i n Central Euro p e . J. Animal ----Eeol. 6 : 197-198.
FRENZ É L, G. , 1936. Untersuchungen uber die T i er welt des
Wiesenbodens. Jena . 130 pp.
GHIL AROV, M. S. , 1 944. Correlation between si ze and number of
soi l ani mals. Comp.t. R e.nd. Aead. Sei. URSS . 43 (6) : 267-26 9.
G IS IN, G. , 19 52. O kologi e studi en über d i e Collembolen de
blattkomposts. R e.v. Sui1.> 1.> e. Zool. 5 9: 5 43-578.
G ISIN, H. , 1943. O kologie und Lebensgeme i nschaften der Collembolen
in schwei zerischen Exkursionsgebiet Basels. R e.v. SU,Ú>1.>e ZooL
50 (4) : 131-224.
----, 19 49. Exemple du developpement d ' une biocenose dans un
ta s d e f e u i l l e e n d e e o m p o s i t i o n . B ull • S o e . E n.t • S ui1.> 1.> e. • 2 2
(4) : 422 pp.
----, 19 56. L ' evolution du peuplement des Collemboles
( Insectes Apteri gotes) dans deux tas de feui lles compostes
dans de condi tions differents. V I Congn. In.te.n. Sei. Sol. ,
Pari s. : ll-14.
GLASGOW, J. P. , 1939. A population study of subterranean soi l
Collembola. J. Animal Eeol. 8: 323-353.
GRETSCHY, G. , 19 49. Die Sutzessione der bodenti ere auf T i chten
Schlagen. D i ssertation, Vienna. 16: 723.

--,
5 0 .
HALE , H.G. , 19 7 1. 11 Colembolos 11 , In : BU R GES , A. & RAW , F. , Biologia
del Suelo. Ed. Omega S/A , Barcelona. : 4 63-4 79.
H U RSH , C.R. , 1928. Litter keeps forest soil productive. Southern
Lumberman. 134 : 1-3.
J A e o B , A . P • , l 9 3 5 • W i l d 1 i f e o f t h e f o r e s t e a r p e t. S c.,l • M o • 4 O :
24 5-430.
, 1936a. So il populatios . Si c. . M o . 17 : 24 7 -251. ----, 1936 b. Soil structure and soil biology. E c.o lo g y . 1 7 : ----
359-379.
, 1940. The fauna of the soil. Qua.11.,t . R e. v. Bio l . 1 5 : ----
28 - 58.
J EANSON , C. , 19 7 1. " ttude experimental e de l ' act i on des vers de
terre sur les s ols artificiels " , l n : PESSON , P. , La Vie dans
les Sols , as pects nouveaux e tude experimentales. Gauthier-
Villars Ed. , Paris . : 2 13-27 7.
J OHN , E. , 1944. Bodentieruntersuchungen in den FlUgs andboden des
Marchfeldes Hochschule f. Bodenkult , Vienna.
KEVAN , D.K. Mc E. , 1968. So i l Animal. H.F. & G. Witherby Ltd. ,
1 London. X + 244 pp .
KING, K.M. , 1939. Populations s tudies of s oil ins ects. E c. o l .
M o n o g 11. . 9: 2 7 0 -300.
K UHN ELT , W. , 196 1. Soi l biology , with s pecial reference to the
animal kingdom. Faber & Faber Ed. , London. 39 7 pp.

----
5 1 •
, l 9 6 5 . 11 A n i n t r o d u e ti o n to t h e s tu dy o f s oi l a n i ma l s 11 ,
l n : Kevan , D. K . McE. , Soil Zool ogy . Butterworths, London.
: 3-22 .
LEBRUN , P . , 1971 . Ecol ogie et biocé notique de que l ques peuplements
d ' Artropodes edaph iques . S e. . Nat . B elgiq ue M ém. :165-203 .
LEMO S DE CASTRO, A . , 19 7 6 . Anfípodos terrestres do Gê ner,o TaLi;tJc.uL>
introduzi dos n o Bras i l (Amphi poda , Ta 1 i tri dae) . Atai.> Soe. • . 6io-l,
Rio de Janeiro . 19 : 4 7 - 49.
MAS S O U D , Z • , l 9 7 l • 11 U n é l em e n t c a r a c te. r i s t i q u e d e l a p e d o f a u n e :
l es Col l embol es " , l n : P ES S ON , P . , La Vie dan s Sol s , as pects
nouveaux études expérimental es. Gauthier - Vil l ars Ed. , Paris .
: 213-27 7 .
MIGNOLET, R. & LEBRUN, P., 1 9 7 5 . " Col on is ation par l es
microarthropodes du so l de cin q types de l it i ere en
decompos ition 11 , l n : Progres s in Soil Zool ogy. Organized by
the Soil Zool ogy Committe of the International Society of
Soil Science , 59 Intern ational Col l oquium on Soil Zool ogy .
Pub l . House of Czechos l ovak Academy of Sciences, Prague .
: 261-281 .
MOREIRA , A . A.N. & CAMELIER , C. , 197 7 . " Re l evo" , l n : Geografia do
Bras il . Reg ião Sudeste. Fundação Instituto Bras il eiro de Geo
grafia e Estatística, Rio de Janeiro . 3 : 1-50 .
MULLER , P . E., 1884. Studier over S kovj ord, s om Bridrag til
S kovdy k n igen s Theori. Tid1.i 1.> k..tr.. . S k. o v b.tr..ug . 7 : 1-232.
MURPHY, P . W. , 1953. The biol ogy of forest soil s with s pecial
reference to the mes ofauna or meiofaun a . J . S oil Sic. . 4 : 155-
1 9 3 .

52.
NEF, L . , 1957. ttat actuel des connaissances sur le rôle des
animaux dans la decomposition des litiêres de forêts.
Ag Jz.ieuf.:tuJz.e . 5: 245-316 .
N I KI FOROFF, C. C. , 1941. Soil dynamics. S ei. Mo. 52: 422-429.
NIMER, E. , 1979. Climatologia do Brasil. Fundação do I nstituto
Brasileiro de Geográfia e Estattstica, Rio de Janeiro. Serie
Recursos Naturais e Meio Ambiente. 4: 421 pp.
PEARSE, A. S. , 194.6. Observations on the microfauna of the Duke
forest. E eof. Monog Jz.. 16 (2) : 127-150.
RAPO PORT, E . H. , 1959 . Algunos aspectos de 1·a biologia del suelo.
Univ. Nac. Sur. Extensiõn Cultural, Bahia Blanca. 23 pp.
, 1960. Formaciõn de humus por los insectos colembolos. ----Idia , Supl. l: 80 pp.
RAW, F. , 1971. 1 1 Artrõpodos (exceto ãcaros y colembolos) 11 , In:
BUR GES, A. & RAW, F. , Biologia del Suelo. Ed. Omega S/A,
Barcelona. : 379-423.
SAL MON, J. T. , 196 4. An index to the Collembola. TJz.an.6 . R. Soe.
N. Z. 7 (1-2-3): 1-6 44.
T RA GARD� I. , 1933. Methods of automatic cbllecting for studying
the fauna of the soil. Bufl. En.:tomol. Ru. 24: 203-214.
T UL L GREN, A. , 1917. Ein sehr einfacher ausleseappara·t fUr
terricole Tierformen. Zei.:t.6 ehJz.. an gew. En .:tomofogiae. 4:149-150.
VAN N I E R , . G • , l 9 71 . 1 1 Te c h. n i q u e s d ' e tu d e 'd e s p o p u l a t i o n s d e s
microarthropodes du sol " , I n: PESSON, P. , La Vie dans les Sols,
aspects nouveaux êtudes experimentales. Gauthier-Villars
Ed. , Paris. : 85-109.

7
WALLW ORK , J . A . , 1970. Ecology of soil animals . Mc Graw -Hill
Publis hing Company Limited , London . VII + 283 pp.
53 .
, 19 71. 11 11:caros " , l n : BURGES, A . & RAW , F . , Biologia del ----sue l o • E d • Omega S / A , Bar e e 1 o na • : 4 2 5 -4 61 •
WEBB , D . P. , 197 7 . " Regulation of deciduous forest litter
decomposition by s oil arth ropod feces " , l n : MATTSON , W.J., The
role of Arth ropods in forest ecosystems . Springer- Verlag Ed . ,
New Yor k. : 57 - 69 .
WILLIAMS , E.C., 1941. An ecological study of the floor fauna of
the Panama rain forest . Bull . Chicag o A cad . S ei . 6 : 63-124.

-,
X , TABELAS

55 . .
TABELA I
Variaç5es midtas mensais de temperatura, umidade
relativa e precipitação pluviométrica, ao longo
das observações da sucessão faunística em - um
"fosso .de folhiço 11, de j ulho de 1977 a junho de
1978.
M E S E S 1
J U L H O
A G O S T O
S E T E M B R O
O U T U B R O
N O V E M B RO
D E Z E M B R O
J A N E I R O
F E V E R E I R O
MA R Ç O
A B R I L
M A I O
J U N H O
Tempera tu r a
Mé d i a º e
2 4 , 5
2 3 , 5
2 4 , 0
2 5 , 5
2 6 , 0
2 6 , 0
2 7 , 5
2 8 , 5
3 0 , O
2 8 , 5
2 5 , 0
2 3 , 0
U m i d a d e Re l a ti v a P re c ipi ta ç ão
M é d i a % Plu viom é t ri ca
8 3 4 3 , 8
8 1 5 3 , 2
8 1 1 2 8 , 6
8 1 4 5 , 2
8 2 1 1 8 , 5
8 1 9 0 , 4
8 0 5 6 , 0
8 2 1 6 4 , 9
8 3 5 1 , 4
8 5 1 1 O , 3
8 5 1 2 7 , 9
8 5 3 3 , 0

7
e:(
1-
u
w
U')
z:
.....
e:( w
u e:(
U') =::, o:::: u
e:( C) ..... u e:( o:::: e:(
U') o o.. =::, o:::: (.!,
U') o o::::
o
TABELA l i
Grupós faunisticos coletados em um "fosso de folhiço " , com o respectivo percentual de pa rticipa ção no total , de j ulho de 1 977 a j unho de 1 978.
G R U P o s N<? %
Coleopter a ( a d. ) 336 0 , 42 Coleoptera ( 1 a . ) 133 0 , 16 Collembola 26677 33 , 38 Dermapter a 3 61 0, 4 5 Dictyoptera ( n i . ) 7 5 0, 09 Dipter a ( a d. ) 40 0 , 05 Dipter a ( 1 a . ) 221 0, 27 Embiopter a 1 0 , 00 Hemipter a ( n i . ) 52 0, 06 Lepidopter a ( 1 a . ) 8 O , 01 Psocopter a 9 9 O , 1 2 Thysanoptera 3 5 0 , 04
T o t a 1 28038 35 , 08
Amphipoda 6611 8, 27 Isopoda 26494 3 3 , 1 5
T o t a 1 33105 41 , 4 2
Ac a rina 17763 22, 22 A raneida 200 0, 25
T o t a 1 17963 22 , 47
Diplopoda 2 91 0 , 36 Chilopoda 25 0 , 03 Gastropoda 63 0 , 07 Oligochaeta 430 0 , 53 P auropoda 1 0 , 00
T o t a 1 810 1 , O l ,
T O T A L G E R A L 79 916 - 100 , 00 =
56.

GRUP
OS
COLE
TADO
S
Cole
opte
ra
(ad.
) Co
leop
tera
(l
a.)
Coll
embo
la
Derm
apt
era
Dic
tio
pter
a Di
pter
a (a
d.)
Dipt
era
(la.
) Em
bio
pter
a He
mipt
era
Lepi
dopt
era
(la.
) Ps
ocop
tera
�
Thys
ano
pter
a Am
phi
poda
Is
opo
da
Acar
ina
Aran
eida
Di
plo
poda
Ch
ilo
poda
Ga
str
opo
da
Olig
ocha
eta
Pa
uro
poda
T O
T A
L
J J
TA
BELA
Ili
J
Nume
ro d
e es
péci
mes
cole
tado
s em
cad
a gr
upo
, a
o lo
ngo
da
suce
ssão
fau
nist
ica
em u
m·
"fos
so
de f
olhi
ço",
de
ju
lho
de
1977
a
jun
ho d
e 19
78.
Nume
ro
de
dias
a
pos
o in
icio
da
s ob
serv
açõ
es
30
60
90
l 20
1
50
180
210
240
270
300
330
360
I TO
TAL
Jul.
l Ag
o.l
Set.
1 Ou
t.1
Nov.
l De
z.l
Jan.
l Fe
v.l
Mar.
l Ab
r.l
Mai.
l Ju
n.
42 28
3796
14
2 l 3 3
2 l 8 l 9
0 13
47
220 9
2
62
86
l 7
27 9
5 14
0 l 6 4 25
l
l 8
78
2856
58
1 l 7 l 6
31
36 2
3 021
18
l l
120
2 l 9
27
466
l 2 4 53
20
l 8
329
3
3
l
11
2
355
5 70
82
271 27
11
20
95
21
l 6
1322
l
2 l
2 8 l 2 86
1 24
50
367 l 9
99
13
52
l 3
l 3
2023
2 20 5 l l 3 6
487
1476
53
0 22
11 l 9
46
9 4 92
14 4 5
47
14
25 l
l 27
9 84
8 11
21
l 2
35 11
2860
4 9 lo
l 6 l
30
l 08
2500
83
1 38
20
6 3 l
o
46
lo
4
56 4 l o
l 2
l 6 l
33
l 20
55
39
849 42 20 7 2
11 l
21
5 28
0 2 1 5
25 1 5 4 23
4900
2 4 5 21
5 6 37
0 10
28 2 10
1 5
3800
l 2 4 2 25
7 3 2
11 8 l 7 2
4100
2 · 2 24
5 8 8 3
1 6
6 7 2
I 6 7
4 3
111 4
7 5
I 5 2
5 5
I 4 7 6
8 I 1
1 485
I 6
4 9
2 I 7
1 7 9
I 5
3 l 3
I 4
2 8 O
I 4 3 7
1
336
-l
-33
2667
7 36
1 7
5 40
22
1 1 52
8 9
9
35
6611
26
494
177
63
200
291 25
63
43
0 1
799
16
u,
'-1

GR
UPO
S
CO
LETA
DOS
Col
eop
tera
(
ad.)
C
ole
opte
ra
(la
.)
Col
lem
bola
De
rma
pte
ra
Dic
tiop
tera
D
ipte
ra
(ad
.)
Dip
ter
a
(la
.)
Emb
iop
tera
He
mip
tera
Le
pid
opte
ra
Psoc
opte
ra
.. T
hys
anop
tera
Am
phi
pod
a
Isop
oda
Ac
arin
a Ar
ane
ida
Di
plop
oda
Ch
ilop
oda
Ga
str
opod
a Ol
igoc
hae
ta
Paur
opod
a
T O
T A
L
J
30
Jul.
o, 7
1 0
,47
64,5
4 2
,41
0,0
1 0
,05
0,5
4 0
,01
0,1
3
3,2
2 22
,88
3,7
3 0
,15
0,0
3
1,0
5
J j
J
TABE
LA
IV
Abu
ndâ
ncia
re
lat
iva
me
nsal
en
tre
os
grup
os
col
eta
dos
a
o
lon
go
da
suc
ess
ão
faun
isti
ca e
m um
"f
oss
o de
fo
lh
iço
",
de
jul
ho d
e 19
77
a ju
nho
de
1978
.
J
Num
ero
de
dia
s ap
os
o in
icio
da
s o
bse
rvaç
ões
60
Ago.
l ,2
8 0
,25
41,
89
2,0
9 0
,23
0,0
5 0
,37
O ,0
1
0,2
6 l
, 17
42
,80
8
,71
0,2
5 0
,01
0,0
8 0
,46
90
Set.
0,5
3 0
,02
44,8
0
0,2
6 0
,01
0,0
1
17,8
2 28
,57
6,9
1 O
, 17
0,0
5 0
,78
120
Out
.
O, 1
7
0,1
5
2,8
6 0
,28
0,0
0
0,0
9
O ,0
1
30,9
8 61
, 71
2,3
6 0
,23
0,0
9
o, 1
7
0,8
2
150
Nov
.
0,39
0
,30
2
5,1
5 0
,22
0,2
2
O, 1
5
0,0
1
0,0
3 16
,38
46,6
2 6
,98
0,3
6 1,
88
0,2
4 0
,98
180
Dez.
0,2
7 0
,27
42,4
2 0
,04
0,4
1
o, 1
0
0,0
2 0
,02
0,0
6 O
, 12
l 0,2
1 30
,95
11, 1
1 0
,46
2,3
2
o, 1
8 0
,96
210
Jan .
0,0
7 0
,03
80, 1
9
0,0
3 0
,04
0,4
0
o, 1
4
0,2
0 0
,00
11 , 1
3
7,38
0
,09
o, 1
8
0,0
0 0
,01
240
Fev
.
0,5
3 0
,16
44,0
5 0
,06
o, 1
3
0,1
5
0,2
4 0
,01
0,4
6
l ,6
6 38
,50
12
,80
0
,58
0,3
0
0,0
9 0
,04
o, 1
5
270
Mar.
0,6
4 o, 1
3
6,3
5 0
,05
o, 1
3
o, 1
6
0,2
2 0
,01
0,4
5
1,67
77
, 15
11
,82
0,5
8 0
,27
0,0
9 0
,02
o, 1
5
0,0
1
300
Abr
.
0,3
9
0,09
5,
27
0,0
3
0,2
8 0
,47
0,0
1 0
,09
0,0
7
0,4
3 92
,22
0,0
3 0
,07
0,09
0,39
330
Mai.
o, 1
1 o, 1
4
8,6
4
0,2
3 0
,65
0,0
4
0,2
3 0
,35
88,7
8 0
,02
0,0
4 0
,09
0,0
4 0
,58
J
360
Jun.
0,0
4 0
,06
4,8
2
o, 1
8
0,3
8
0,0
4
93,
80
0,0
4 0
,04
0,5
4
99,9
3 1 9
9,9
1 1
99,9
3 1
99,9
2 I 9
9,9
1 I
99,9
2 I
99,8
9 I
99,9
1 I
99,9
0 I
99,9
3 1 9
9,9
4 1
99,9
4 _
__
__
__
__
__
..__ _
__
_.__ _
__
_,__ _
__
_._ _
__
---1. _
__
---1. _
__
__Ji___
__
_ .1__
__
_ .1__
__
_ .J_
__
_ _1.
__
_ -l
__
_ _
J u,
co

GRU
POS
COL
ETA
DOS
Cole
opt
er
a
(ad
.)
Co
leop
te
ra
(l
a.)
C
olle
mbo
la
Derm
apt
era
Oi
ctio
pter
a Di
pter
a (a
d.)
D
i_pte
ra
(la
.)
Emb
iop
te
ra
Hem
i pte
ra
• Le
pid
opt
era
Ps
oco
pte
ra
Thys
ano
pter
a Am
phi
po
da
Isop
oda
A
car
ina
Ara
neid
a Di
plo
poda
C
hil
opo
da
Gast
rop
oda
Ol
igo
cha
et
a Pa
urop
oda
. l
TABE
LA
V
J J
J
Per
cent
ual
de
oco
rrê
ncia
de
ca
da
grup
o,
em
dif
ere
nte
s et
apas
da
suc
ess
ão
faun
ist
ica
em
um
"fos
so
de
fol
hiç
o"
, de
ju
lho
dé
19
77
a
jun
ho
de
197
8.
Nume
ro
de
dia
s ap
os
o in
ici
o da
s o
bser
vaçõ
es
30
60
90
120
15
0
180
2
10
240
2
70
300
330
360
Jul
. 1
Ago
. 1
Set
. 1
Out
. 1
Nov.
1
Dez.
1
Jan.
1
Fev
. 1
Mar
. 1
Abr
. 1
Mai.
1
Jun.
12,5
0
25
,59
10,7
1 5
,95
6,
25
3
,86
2,6
7
10,4
1 21
,05
12
,78
1,50
13
,53
12
,03
9
,77
3,
00
8,2
7
12 ,6
9
7, 51
6
,25
1,
48
2,0
8 3
,75
4,5
1 2,
25
14,2
2 10
,47
11,
32
1,2
3
4,9
5
7,5
8 3
4,5
3
10,7
2 l,
70
l , 1
O 1,
04
1,
38
0,7
9
39,3
3
38
,78
4,9
8 1
,33
2
1 ,3
3
1,3
3
7, 5
0
1 O
,00
-
9,1
41
3,3
21
0,5
5
1,3
3
16
,00
26
,66
1,1
01
1,1
0
6,66
12,
00
13,3
3
0,5
5,
-1
-
37
,50
1 2
5,0
01
20,0
0
14,4
7,
ll,3
1 0
,45
4
,97
3,8
4
3 ,6
1
1,9
2
2,2
61
21,
26
4
,52
5
,42
11
,31
1 1
2,7
6 7
,69
10
0,0
0
-
22
,85
2,8
7 5
,08
1, 23
4,5
0
0,6
8
1,92
51,4
2
1 , 1
7 1 o
, 77
3,2
7
8,50
0
,34
18
, 18
7,2
7 2
,62
6,00
53
,77
26
,73
1,5
2
13
,50
3
,78
5,7
1
13
,02
9, 2
4
2,0
6 9
,50
3
4,0
2
1,9
2
12,9
5
3,0
3 1
7, 1
4
7,3
6
5,5
7
2,9
8 11
, 00
38
, 14
14,4
1 9
,52
1 6,
34
1 31
,74
1 20
,63
11
4,2
8 7
,20
12
,32
2
2,09
1
2,0
9
10,69
26
,92
30,7
6
30,7
6
1,9
2
-12
,25
12
,25
62
,50
2
5,2
5
30,3
0
33
,33
4
,04
2,8
5
--
-
4,8
2
4, 7
7
5,5
0
7,2
1
4,0
0 3
,17
1 ,63
9
,43
4,6
7
19,0
0 6,
87
24,0
0 4
,76
2,3
2
1 , 8
1
20,9
0 4
,77
21
,00
6,
87
28,
00
3,1
7
2,5
5
100
,00
0,0
8 2
7,5
8 1
,00
1,
37
20,0
0
4,8
8
2,0
2
o, 1
5
0,0
5
21
,39
0,5
0
0,6
8 16
,00
3
, 17
5,8
1
2,0
2
23,0
8
8,0
0 3
,17
5,5
8
J
TOTA
L
l 01,
44
99,9
5 99
,93
99,9
5
99,9
7
100
,00
99
,93
100
,00
99
,96
99,2
5
99,99
99
,97
99,9
6 99
,94
99,9
4
100
,00
99,9
6 10
0,0
0 99
,99
99
,88
100
,00
u,
I.O

J
Gt
NE
RO
S
CO
LETA
DOS
A1tl
e.a.
A1tl
e.1.i
ia.
Bita.e.
hy1.i
tom
e.li
a. Ca.m
pylo
tho
Jta.X
e yp
ho
d e.
Jtu
.6
V ,i, c.Jta.
no e.
e. YLtJr.uJ.i
Fal
i.> o
mi
de.1.i
F1t
iu
ia.
L
� o-t
oma.
I .6 o
to
mi
e.l
la.
Le.pi
doc.y
lttu.6
..
Me.g
a.lot
hoJta.X
Ne.
e.lu
.6
Pt
e.no
.th/t
ix
s e
.i/t
a.
Smi
nthu/ti
du
s p
ha. e.
/ti
d,i,
a.
_I
30
.1
TABELA
VI
J J
1 J
Num
er
o
de
es
pec
ime
s d
e
Col
lem
bola
co
let
ad
os
em
ca
da
gên
ero
, ao
lo
ng
o
do
pro
ces
so
su
ce
ssó
rio
em
um
"fo
sso
de
f
ol
hiç
o"
, de
j
ulh
o d
e
1977
a
ju
nho
de
19
78.
Num
ero
de
d
ias
apo
s o
ini
cio
d
as
ob
serv
aç
õe
s
60
90
120
150
180
210
240
270
300
330
360
Jul
. 1
Ago
. 1
Set
. 1
Out
. 1
Nov
. 1
Dez
. 1
Jan
. 1
Fev
. 1
Mar
. 1
Abr
. 1
Mai.
1
Jun
.
47
112
9 2
1541
2083
2
3
20
220 1 36
547
1968
7 32 7
273 5 16
6 29
2556
l l 3 25
13
47
38
51 2 2
230 6
4 6 59
174 24
34
234
783 2 2
6 4 64
74
12
28
684 l
1134
5 l 2 8
3 l 14
5 36 8 63
2624
6321
l 3 3 6
8 2 38
114
44
29
463
2130
9 6 12
5
2 9 26 1 2
154
259 2
3 4 2 17
4 85 6 5
12
22
20
217 98 l
5 1 16
123 65
l
TOT
AL
21
90
517 7
1003
5 11
2
238
6792
1
1771
2 1 28
13
38
47
52
---
--
--
--
--
--
-t--
--
-t--
--
-11--
--
--t-
--
--t-
--
--
1--
--
-t--
--
-+-
--
---,1--
--
--t-
--
---+-
--
--
+--
--
-+--
--
--
--
-
T O
T A
L 37
96
I 27
95
I 30
21
329
1 13
22 1
2023
I
92T4
1
2860
45
6 28
0 37
0 21
1 26
677
O)
o

J J
GtN
EROS
CO
LETA
DOS
A1tl
e.a
A1t
le.
.6-ta
BM
.c.hy
.6tom
ella
Ca
mpyl
otho
1ta.x
e
yp h
o d
e.Jtu
.6
V-tc.Jt
a.no c.
e.ntluv.i
Fo l
.6 o
mi
d e.
.6
F1ti
e..6-t
a
I.6
ot
om
a
I .6 ot
omi
ella
Le.pi.
doc.y
ltt.u..6
Me.g
alo:t
hoJtaX
N e.
e.l
u.6
Pt
e.n
ot1t
ix
Se.i
1ta
Sm-i.n;t
hwúd
e..6
Sp
ha.
e.Jti
di
a
T O
T A
L
J
30
_J
.l
TABE
LA
VII
J · .1
Abu
ndâ
ncia
re
lati
va m
ens
al
ent
re o
s ge
nero
s de
Co
llem
bola
co
leta
dos
ao
. lo
ngo
da
suce
ssio
fau
nis
tica
em
um
''fos
so
de
fo
lhi
ço"
, de
ju
lho
de
1977
a
junh
o de
19
78.
Nume
ro
de
dia
s ap
os
o
60
9
0
120
15
0
180
• -r
•
1n1c
10
210
das 240
obs
erva
ções
.
270
30
0
330
J 36
0
---
-+
--
--+
--
·---+
--
--l
l---
--
.---
--
+--
--
.+--
--
-4--
--
--f--
--
-J-
--
�1---
--
--
Jul.
1, 23
2
,95
0,2
3
0,0
5
40,5
9
54,8
7
0,0
5
Ago.
o, 1
O
o, 7
1
7,87
0,0
3 1,
28
19, 5
7
70,4
1
Set.
0,2
3 1 ,
05
0,2
3 9
,03
o, 1
6
0,5
2
o, 1
9
0,9
5
Out
.
11 ,5
5
15,5
0
0,6
0
0,6
0
Nov
.
0,3
0
0,4
5 4,
46
13, 1
6
1 ,81
2
,57
17,7
0
84,6
0
1 6
9,9
0
1 59
,22
0,0
3 -
-
o ,0
3 -
1 o, 1
5
0,0
9
--
0,8
2
1,82
1 0
,15
0
,43
--
l ,55
1
-1
-
Dez
. 1
·Jan
.
0,29
o, 1
9
3, 1
6
3,6
5
0,5
9
1,38
33
,81
0,0
4
0,0
3 0
,01
1,57
0,3
9
0,0
8 0
,68
28
,47
56,0
5 I
68,
60
0,2
4 0
,01
0,0
4 0
,03
0,0
9
0,0
3 0
,39
0
,06
Fev.
0,2
7 0
,06
1,32
3,9
8
1 , 5
3 0
,06
16, 1
8
Mar
.
0,4
3 1,
97
5,7
0
0,2
1 0
,43
33,7
7
74,4
7 I
56,7
9
0,3
1 I
0,4
3 0
,20
1
-
0,4
1 I
0,2
1 0
,17
1 -
Abr.
0,3
5
1 ,0
7
1,4
2
0,7
1 62
, 14
30,3
5
2, 1
4
1,78
Mai.
3,2
4
5,9
4
5,4
0
58,6
4
Jun.
2,3
6
0,4
7
7,58
58
,29
26
,48
I 30
,80
0,2
7 I
0,4
7
99
,97
I 9
9,9
7 1
99
,91
I 99
,97
I 9
9,9
7 I
99
,92
I 9
9,9
6 1
98
,96
I
99
,94
I 99
,96
I
99,9
7 I
99
,97
.J
O"I

Gt
NE
RO
S
CO
LET
ADO
S
A1r.
le.
a
A1r.l
e.1.>
..i..a.
B1r.a.c.
htj1.>
tom
elta
Campy
loth
oJr.aX
Cy
ph
od
e.Jt
u'->
V ..i..cJt
ano c.
e.nbr..u.-6
Fo.l
1.>om
..i..d
e.!>
F1t..i..
u..i..
a
I '-> o
tom
a
U, o
tom
..i.. e.
li
a.
Le.p..i..
doc.y
Jc;Ó.L6
Me.g
alot
hoJta.
x
Ne.
e.fu
.t, P
te.
no
th1t
ix
s e.
..i..Jr.
a.
SmW
hwúdu
S
ph
a.e.
1r.i
di
a.
J
_)
.l -�
1
TAB
ELA
V
II
I
.l ._
J _
J
Per
cent
ual
de
o
corr
ên
cia
d
e
cada
gê
ner
o
de
Co
lle
mbo
la,
em
dif
er
en
te
s e
ta
pa
s d
a
su
ces
são
f
aun
ist
ica
em
um
"f
os
so d
e f
olh
iço
",
de
j
ul
ho
de
19
77
a j
un
ho
de
19
78.
Núm
ero
de
d
ias
apo
s o
ini
cio
d
as
ob
serv
açõ
es
30
60
90
120
150
180
210
24
0 27
0 30
0 33
0 36
0 -
--
--
-+-
--
---l
r--
--
-+-
--
--
--+-
--
--
+--
--
---¼
--
--
"-""r-
--
--
+-
--
---1
r--
--
-+-
--
--
--¼-
--
--
Jul
. 1
Ago
. 1
Set
. 1
Out
. 1
Nov
. 1
Dez
. 1
Jan
. 1
Fev
. 1
Mar.
1
Abr.
1
Mai
. 1
Jun
.
.52
, 22
21,6
6
3,3
3 7
,77
3,8
6 6
,18
-10
0,0
0
0,8
9 21
,93
27,2
1
--
l 00
,00
1,78
0
,89
14,2
8 -
15,1
2
2,5
2
22,6
8 8
,05
0,4
2
11 , 7
6 1
11 , 1
1 1
14, 4
3
4,2
5
l 00
,00
3,5
7 23
,07
65,Z
S 27
,65
90,3
8
7,35
19,0
4 28
,57
14,2
8 38
,09
6,6
6 4
,44
1,11
2
,22
11,4
1 12
,37
28,0
4 7
,35
5,08
1 17
,34
1 7
,37
1 3
,58
1 11
,36
-21
,42
10
,71
7,14
39
,28
0,8
4 14
,28
11,7
6 26
,47
12,1
8
0,0
2 3
,44
10,0
7 38
,63
6,81
-
•100
,00
1,29
1 4
,42
1 6
,40
1 35
,68
1 12
,02
15,7
8
7, 1
4
17 ,8
5 -
7,69
5
,26
5,26
-
17,0
2
3,57
32
,14
23
,07
46,1
5
7,8
9 -
12,7
6 25
,53
-9,
61
2,22
1,74
2,59
0,8
9
0,8
4
2,2
6
l ,46
1, 11
0,2
9
3,5
7
0,8
4
2,5
6
0,4
7
7, 14
I 21
,42
2, 12
I l O
, 63
13,3
3
2, 19
8,4
0
3, 19
0,5
5
3,5
7
5,5
5
0,0
9
6,7
2
l ,8
1
0,3
6
3,5
7
TO
TAL
99,9
8
99,9
6
99,9
6
100
,00
99,9
2
100
,00
99,9
6
99,9
7
99,9
4
100
,00
99,9
5
100
,00
99,9
7 99
,97
99,9
7 99
,96
99,9
9
1 .,
°'
N

XI , I LUSTRAÇOES

-,
64.
X I. l) P RANCHAS
P RANCHA : A - Principais grupos faunísticos encontra
dos no folhiço.
l. a) Collembola (Entomobryomorpha)
l . b) Collembola (Symphypleona)
2) Isopoda
3) Acarina
4) 01 i gochaeta
5 ) Amphipoda
6 ) Pauropoda
7) Coleoptera
8) Diptera (larva)
9 . a) Thysanoptera (Terebrantia)
9 . b) Thysanoptera (Tubulifera)
1 O) Di ctyoptera
11) Embi optera
12) Homoptera
13) Psocoptera
l 4 • a ) D i p l o p o d a ( P o f J., x. e. n u.-6 )
14. b) Diplopoda (Julidae)
15) Chilopoda
16) Araneida
17) Gastropoda
18) Dermaptera

b 5 .
6
9 . a
1 0
1 2
1 3
1 6
P r a n c h a A

�, P RAN C H A : B - F o rm a s b i o 1 6 g i c a s em C o l l em bo l a
ATMOB I ÕT l COS OU E P I EDÃFI COS
l . L e.pi do c.y1t:tu.6
2 • M a.6 ;t,i. g o e. e.Jt a.6
3 . T e.m e.Jti:ta.6
HEMIEDÃF I COS
4 . N e. o :t!to pi e..f...f..a
5 . I.6 o:toma
6 . Smin:thu!tu-6
E UEDÃFI COS
7 . K e. nyuJta
8 . Tu.f...f..b e.1tgia
9 . N e.anu!ta
l o . P .6 e. udo.6 :ta c.hia
1 1 . N e. e..f.u-6
66.

1
4 6
7
1 1 �
.Pra n c h a B

7
7
68.
XI . 2 ) F I G U RAS
Figura 1 :
Figura 2 :
Figura 3:
Figura 4 :
Figura 5 :
Figura 6 :
F r e q tJ ê n c i a r e l a t i v a em n um e r o d e i n d i v í d u o s d e Co l e o .e.
téra · (adulto e larva) ao longo dos 360 dias de ob
servaçoes do processo de decomposição do folhiço,
julho de 1977 a junho de 1978 .
Freqüência relativa em numero de indivíduos de - Collem
bola ao longo dos 360 dias de observações do proce�
so de decomposição do folhiço, julho de 1977 a junho
de 1978.
Freqüência relativa em nümer de indivíduos de Der
maptera ao longo dos 360 dias de observações do pr�
cesso de decomposição do folhiço, julho de 1977 a
junho de 1978.
Freqüência relativa em numero de indivíduos de Thf
sanoptera e Dictyoptera ao longo dos 360 dias de ob
servaçoes do processo de decomposição do folhiço,
julho de 1977 a junho de 1978.
Freqüência relativa em numero de indivíduos de DiE
tera (adulto e larva) ao longo dos 360 dias de obser
vaçoes do processo de decomposição do folhiço, julho
de 1977 a junho de 1978.
Freqüência relativa em numero de indivíduos de Gas
tropoda e Hemiptera ao longo dos 360 dias de obser
vaçoes do processo de decomposição do folhiço, julho
de 1977 a junho de 1978. ·

Figura 7 :
Figura 8 :
Figura 9 : 7
Figura 1 0 :
Figura 1 1 :
Figura 1 2 :
Fi gura 1 3 :
69.
Fre q fl ê n c i a re l at i v a em n umero d e i n d i v íd u os d e Ps o
c o p tera e L e p i d o p t era ( l arva) ao l o ngo d os 3 6 0 d i as
d e o bs ervaç õ es d o pro cess o d e d e c om p os i çio d o fo l h i
ç o , j u l h o d e 1 9 7 7 a j u n h o d e 1 9 78 .
Fre q fl ê n c i a re l at i va em n umero de i n d i v í d u os de Amph..:!_
p o da ao l o ngo d os 3 6 0 d i as d e o bs ervações d o pro c el
s o d e d e c o m p os i ç ã o d o f o l h i ç o , j u l h o d e 1 9 7 7 a j u n h o
d e 1 9 78 .
Fre q U ê n c i a re l a t i va em n um ero d e i n d i v í d uos d e I s o
p o da a o l o ngo d os 3 6 0 d i as d e o bs ervações d o pro c el
s o d e d e c o m p os i ç ã o d o f o l h i ç o , j u l h o d e 1 9 7 7 a j u n h o
d e 1 9 78 .
Fre q � ê n c i a re l at i va em n umero d e i n d i v í d u os d e A ca
ri na ao l o ngo d os 3 6 0 d i as d e o bs ervações d o pro cel
s o d e d e c o m p os i çã o d o fo l h i ç o . j u l h o d e 1 9 7 7 a j u n h o
d e 1 9 78 .
Fre q � ê n c i a re l at i va em n um ero d e i n d i v í duos d e Ara
n e i da ao l o ngo d os 3 6 0 d i as d e o bs ervaç ões d o pro c el
s o d e d e co m pos i ç ã o d o fo l h i ç o , j u l h o d e 1 9 7 7 a j u n ho
d e 1 9 78 .
Fre q Uê n c i a re l a t i va em n umero d e i n d i v í d uos d e D i p l �
p o da e C h i l o p o da ao l o ngo d os 3 6 0 d i as d e observações
d o pro c ess o d e d e c o m p os i ç ã o d o fo l h i ç o , j u l ho de 1 9 7 7
a j u n h o d e 1 9 78 .
Fre q U ê n c i a re l at i va em n um ero d e i n d i v í d uos d e 0 l i
go c hae ta ao l o ngo dos 3 6 0 d i as d e observações d o pro c el
s o d e decompos i ção do fol h i ço, j u l h o d e 1 977 a j unho de l 978.

Figura 14:
7
Figura 15:
7
Figura 16 :
Figura 17:
Figura 18:
Figura 19:
70.
Anilise comparativa intre as freqilências re l ativas
em numero de indivíduos de Collembola, Isopoda. e Ac!
rina ao longo dos 360 dias de observações do proce�
so de decomposição do folhiço, julho de 1977 a junho
de 1 978.
Comparação entre as tendências populacionais das fr�
qtJências relativas em numero de indivíduos de Collem
bola e Acarina ao longo dos 360 de observações do
processo de decomposição do folhiço, julho de 1977
a junho de 1978.
Freqüência relativa em numero de indivíduos de AJc_R...e_a.
e Afl_R... e,�..i..a. (Collembola) ao longo dos 360 dias de ob
servaçoes do processo de decomposição do folhiço,
julho de 1977 a junho de 1978.
Freqüência relativa em numero de indivíduos de Bfl_a.
Qhy��om e,R...R...a. e Cyphode_fl_u� ( Collembola) ao longo dos
360 dias de observações do processo de decomposição
do folhiço, julho de 1977 a junho de 1978.
Freqüência relativa em numero de indivtduos de FQR...
�om..i..de,� e fJc_..i_ e_�..i..a. ( Collembola) ao longo dos 360 dias
d e observações d o processo d e d eco m p os i ç ão do f o l h i
ço, julho de 1977 a junho de 1978.
Freqüência relativa e m numero de indivíduos de I�o
�orna. ( Collembola) ao longo dos 360 dias de bbserva
çoes do processo de decomposição do folhiço, julho
de 1978.

-,
Figura 20:
Figura 21:
Fi.gura 22:
Figura 23:
Figura 24:
Figura 25:
7 1 .
FreqfJência re l ativa em numero de indivíduqs de Le.p:!:
doc.1p1.tu-0 { Co l l embol a) ao l ongo dos 3 60 dias de ,obser
vações do processo de decomposição do fol hiço, jul ho
de 1977 a junho de 1978.
F reqüência re ] à tiva em n umero de indivíduos de Ne.ei.U-O ,
S e.ina e Pte.nothnix (Col l embol a) ao l ongo dos 3 60
dias de observações do processo de decomposição do
fol hiço, ju l ho de 1977 a junho de 1978.
Freqüência rel ativa em numero de indivíduos de Smin
thunidu e Sphae.nidia (Col l embol a) ao l ongo dos 3 60
di as de observações do processo de decomposição do
fol hiço, jul ho de 1977 a junho de 1978.
Variação mensa l das medias de temperatura ( º C) ao
l ongo dos 3 60 di as de observações do processo de de
composição do fol hiço, jul ho de 1977 a junho de 1978.
Medições tomadas durante as amostragens.
Variação mensa l das medias de umidade re l ativa do ar
ao l ongo dos 3 60 dias de observações do processo de
decomposição do fo l hiço, jul ho de 1977 a junho de
1978. Medições tomadas durante as amostragens.
Variação mensal das medias de precipitação pl uviom�
tri ca ao l ongo dos 3 60 dias de observações do proce�
so de decomposição do fol hiço, jul ho de 1977 a junho
de 1978. Medições tomadas durante as amostragens. ,

Figura 26 :
---,
J
72 .
Anãlise comparativa .entre as variações mensais de .tem
peratura, umidade relativa do ar e precipitação pl�
viometrica ao longo dos 360 dias de observações do
processo de decomposição do folhiço , julho de 1 977
a junho de 1978.

Nº de i nd i v íd uos
1 00
80
60
40
20
J/ 30 A s o N
N O d e i n d i v íd u o s
50
40
3 0
2 0
1 0
J /30 A s o N
D J F M A
D J
F M A
F i g u ra
M
M
G o l e o p t e ra
a d u l to s
73.
m e ses/ /d i a s
C o l e o p t e ra
l a r v as
J/360 m e sex d i a s

10 . 000 NQ. de i n d i v íd u o s
9 . 000
8 . 000
7 . 000
6 . 000
5 . 000
4 . 000
3 . 000
2 . 000
1 . 000
A s o N D j F M
F i g u ra 2
A
74 .
C o l l e m b o l a
meses/ /d i a s

140
120
1 00
80
60
40
20
N O d e i n d i v Íd uo s
J/ 3 0 A J
F ig u r a
75 .
D e r m a p t e r a
F M A M J/360 m e s e¾ d ias
3
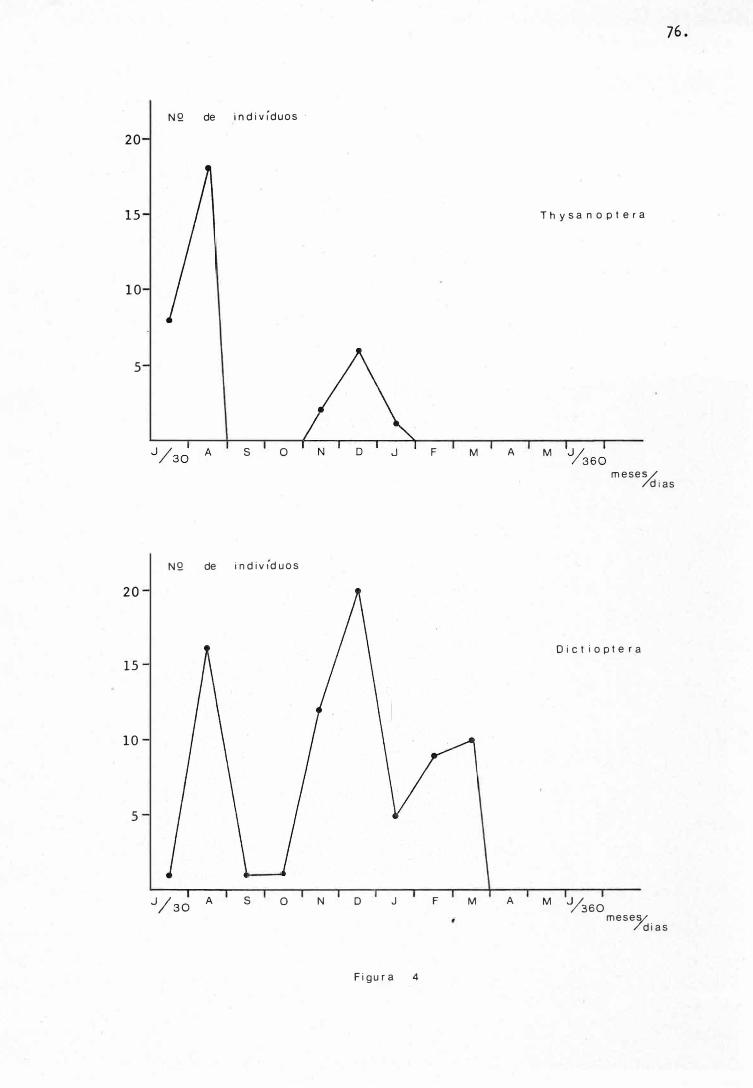
N Q de i n d i v íd u o s ·
20
15
1 0
5
s o
N Q de i n d i v íd u o s
20
15
10
5
J/so A s o N
F M A
D J F M A
F i g u r a 4
76 .
T h y s a n o p t e r a
M
M
m ese¾ d i as
O i c t i o p t e r a
j/360 meses/
/d i a s

N O d e i n d i v íd u o s
3 0
20
10
J /30 s o N
N º de i n d i v i'd u o s
60
40
20
J/30 A s o N
D J
D J
F i g u ra 5
F M
F M
A M
A
D i p t e r a a p t e r a
'1/360
7 7 .
m eses/ /d i a s
D i p t e r a l a r v a
mese¾ d i as

20
15
10
5
20
15
1 0
5
N� de i n d i v (d u os
J /30 s
N º d e i n d i v id ues
J/30
N
N
D J
D J
F i g u ra 6
F A M
F M
G a s t r o p o d a
J/4so m eses/
/d i a s
H e m i p t e r a
J/360 m e seo/, /d i as
78 .

---, '
40
30
20
1 0
1 0
5
NQ de i n d i v íd uos
J/30 A s o
NQ de i n d i v i'd u os
J/ 30 A s o
N
N
D J
D
F ig u ra 7
F M A
P s o c o p t e r a
meses/. /d i a s
L e p i d o p t e r a l a r v a
meses/ /d i as
79 .

N Q de i n d i v íd u o s
4 . 500
4 . 000
3 . 500
3 . 000
2 . 500
2 . 000
1 . 500
1 . 000
500
A s o N
F i g u r a 8
80 .
A m p h i po d a
J/360 m e se s/
/d i a s

N� de i nd i v íd u o s
9 . 000
8 . 000
7 . 000
6 . 000
5 . 000
4 . 000
3 . 000
2 . 000
1 . 000
J/30 A s o N D J
F i g u r a 9
F M
8 1 .
l s o p o d a
m ese¼ d i a s

82 .
N Q d e i n d i v íd uos A c a r i n a
5 . 000
4 . 500
4 . 000
3 . 500
3 . 000
2 . 500
2 . 000
1 . 500
1 . 000
500
J/30 A s o N D J F M A M J/260 m e se¼ d i a s
F i g u r a 1 0

7
7 N Q. de i n d i v íd u o s
5 0
4 0
3 0
2 0
1 0
J/30 A s o N D J F
F i g u r a 1 1
M A
83 .
A r a n e i d a
meses/ /d ias

NQ de i n d i v í d uos
1 00
80
60
40
20
N
N Q de i nd i v í d uos
1 0
5
J / 30 A s o N
D J
D J F M
F i g u ra 1 2
A M
84 .
D i p l o p o d a
m e se¼ d ia s
C h i l o p o d a
J/3 6 0 m ese¼ d ia s

N Q de i n d i v íd u o s
100
80
60
40
2 0
J/ 30 A s o N D J
F i g u r a 1 3
M A M
85 .
O l i g o c h a e t a
J/36 0 m e s e¼ d ia s

N Q d e i n d i v íd u os
1 0 . 000
8 . 000
6 . 000
4 . 000
2 . 000
N
N Q de i n d i v íd u o s
4 . 000
2 . 000
Fi g u ra 1 4
8 6 .
C o l l e m b o l a
'1/360 mese¾ d i as
A c a r i n a
meses/ /d ias

10 . 000 N Q
9 . 000
8 . 000
7 . 000
6 . 000
5 . 000
4 . 000
3 . 000
2 . 000
1 . 000
de
A
i n d i v i'd u o s
s
. . . . : :
o
I
, , I ,
·. 1 1 1 1
·. ,,• •(
I •
1 1 1 1
1 1 1 1 1
1 1 1 1
;. :, , : , , 1 1 1 1 1 1
I • • .l . I a,., , .'
•' · · - . ..• -
N D J
1
1 1 1 1 1
1 1 1 ' 1 1 1 1 1
�=-,. J
&' • 1 . ' : ' : '
.-· 1 .-· 1
F
1 1
F i g u r a 1 5
' 1
• · . .
M
•- - -·
·· · · · · · ··
• •
' +- - - -• ..... .... .... ,. .. . . . . . ... . ... A M J/460
87 .
C o l l e m b o l a
l s o p o d a
A c a r i n a
m e s e¼ d i as

7 10
50
40
3 0
20
10
N Q d e i nd i v íd u o s
J/ 3 0 A s o
Nº de i n d i v i'd u o s
N M A
F M A
F i g u ra 1 6
M
M
88 .
A r l e a
A r l e s i a
J/360 m e s e s; /d i as

150 N 2 de i n d i v íd u os
1 00
50
A s o N
3 00 N º d e i n d i v íd u o s
250
7 200
7 150
1 00
50
A s o N
D J F M
D J F M
F i g u r a 1 7
A
A
89 .
B r a c h y s t o m e l l a
M ¾eo meses/ /d i a s
C y p h o d e r u s
½someses/ /d i a s

60
40
20
80
60
40
20
N� d e i n d i v Íd u os
o
N � de i n d i v íd u o s
N D J
F i g u r a 1 8
F
F , M A
M
90.
F o i s o m i d e s
m e ses/ /d i a s
F r i e s e a
M ¾60 m e s e s/
/d ias

91.
N º d e i n d i v íd u o s
3 . 200
1 s o t o m a
2 . 800
7 2 . 4 00
2 . 000
1 . 600
1 . 200
800
400
7 A s J F M A
m e se¼ d i a s
F i g u r a 1 9

N<2 de i n d i v íd u o s
7 . 000
6 . 000
5 . 000
4. 000
7 3 , 000
2 . 000
1 . 000
A s o N D j A
F ig u ra 2 0
92 .
L e p i d o c y r t u s
M 3/360
meses/ /d ;as

20
1 0
7 7 20
10
20
1 0
7
7
NQ de i n d i v íd uos
J/30 A
N � de i n d i v í d u o s
o
NQ de i n d i v íd u os
A s o
N M A M
N D F M A
F i g u ra 2 1
93 .
N e e l u s
J/2so mese¼ d i a s
P t e n o t h r i x
J/260 meses/
/d i a s
S e i r a
meses/ /d ias

N Q d e i nd iv i'd uos
7 20
10
o N
NQ de i n d i v íd u os
40
30
20
1 0
A s o N
j
D j
F i g u r a 2 2
A M
A M
94 .
S m i n t h u r i d e s
J/36 0 mese¼ d ias
S p h a e r i d i a
J/460 meses/
/d ias

30
2 9
2 8
2 7
26
25
24
23
22
21
20
J/30 A s o N D J F M
F i gu r a 2 3
95 .
Te m p e r a t u r a m é d i a
A M J /360 m eses/ /d ias

¾
86
85
84
7 83
82
81
80
J / 30 A s o
7
N D J F M
F i g u ra 2 4
96 .
U m i d a d e r e l a t i v a
A M
m é d i a
meses/ . /d i a s

m m
160
140
120
1 00
80
60
40
2 0
s o N D J F
F i g u ra 2 5
9 7 .
P r e c i p i t a ç ã o p l u v i o m é t r i ca
M
m é d i a
meses/ /d ; as

J _
) _
J _
,
..-Pr
ecip
i t a
çao
pl
uvio
mét
r;ca
m
édia
•---·
Te
mpe
ratu
ra
méd
ia
mm
º
e
%
•····
····Â
U
mid
ad
e
méd
ia
180
] 30
88
I '
' 16
0
... A
29
87
I
'
' '
140
-, '
,. --
- -28
86
'
1 '
I
120 1
,' 27
�
85
100
� __
_ __ •
____
/
26
l ª4
80 -1
•
11
�
i 2 5
�
83
' ' 60
-I
·-� · ...
:,,
,- ' V . .... .... · ·· · ·...
\J ....
V
,_\ \
1-2 4
�
82
4 o -1
..
. .... .
......
...... .
�.-·
··• ..
\ \
1-23
r 8
1 \
\
20 -l
·-·
· •
r 22 r ªº
I J
I A
I S
I O
I
N I
i J
I F
I I
A I
M IJ
/
i /
30
D
M
/36
0
me
ses/
/
di a
s \O
F
igu
ra
26
co

99.
X I . 3 ) F O T O G RA F I AS
Fo t o l :
Fo t o 2 :
F o t o 3 :
Fo t o 4 :
V i s t a p a r c i a l d a ã r e a o n d e r e a l i z a m o s a s v a r re d u r a s p�
r a a fo rm a ç ã o d o " fo s s o d e fo l h i ç o "
A s p e c t o d o " fo s s o d e f o l h i ç o " a o i n i c i a rmo s a s a mo s t r a
g e n s , j u l h o d e 1 9 7 7 .
A s p e c t o d o " f o s s o d o fo l h i ç o " a p õ s s e i s m e s e s d e a m o s
t ra g e n s , d e z e m b r o d e 1 9 7 7 .
A s p e c t o d o 11 f o s s o d e fo l h i ç o " a o f i n a l d o s n o s s o s e s
t u d o s , j u n h o d e 1 9 78 .

1 0 0 .
F O TO
-

F O T O 2
1 O 1 •


















