
Instituto Nacional de Pesquisas da Amazônia INPA Programa de … · 2016-09-13 · Amazonas –...
90
i Instituto Nacional de Pesquisas da Amazônia – INPA Programa de Pós-Graduação em Genética, Conservação e Biologia Evolutiva - GCBEv Composição, abundância, diversidade e mapeamento cromossômico de DNA repetitivo em ciclídeos neotropicais utilizando dados de sequenciamento de nova geração Edika Sabrina Girão Mitozo Tavares MANAUS – AM 2016
Transcript of Instituto Nacional de Pesquisas da Amazônia INPA Programa de … · 2016-09-13 · Amazonas –...

i
Instituto Nacional de Pesquisas da Amazônia – INPA
Programa de Pós-Graduação em Genética, Conservação e Biologia Evolutiva -
GCBEv
Composição, abundância, diversidade e mapeamento cromossômico de DNA
repetitivo em ciclídeos neotropicais utilizando dados de sequenciamento de
nova geração
Edika Sabrina Girão Mitozo Tavares
MANAUS – AM
2016

ii
Instituto Nacional de Pesquisas da Amazônia – INPA
Programa de Pós-Graduação em Genética, Conservação e Biologia Evolutiva -
GCBEv
Composição, abundância, diversidade e mapeamento cromossômico de DNA
repetitivo em ciclídeos neotropicais utilizando dados de sequenciamento de
nova geração
Edika Sabrina Girão Mitozo Tavares
Projeto apresentado ao Instituto
Nacional de Pesquisas na Amazônia
como parte dos requisitos para obtenção
do título de Mestre em Genética,
Conservação e Biologia Evolutiva.
Orientador: Carlos Henrique Schneider, Dr.
Coorientador: Cesar Martins, Dr.
Fontes Financiadoras: FAPEAM/ CNPq/ Fapesp
MANAUS – AM
2016

iii
T231c Tavares, Edika Sabrina Girão Mitozo
Composição, abundância, diversidade e mapeamento cromossômico de
DNA repetitivo em ciclídeos neotropicais utilizando dados de
sequenciamento de nova geração / Edika Sabrina Girão Mitozo Tavares. ---
Manaus : [s.n.], 2016.
xvii, 90f. : il., color.
Dissertação (mestrado) --- INPA, Manaus, 2016.
Orientador: Carlos Henrique Schneider.
Coorientador: Cesar Martins .
Área de concentração: Genética, Conservação e Biologia Evolutiva.
1. Mapeamento cromossômico. 2. Ciclídeos Neotropicais. 3. Peixes –
água doce.
I Título
CDD
639.309
Sinopse:
Análise da composição da fração repetitiva genômica de quatro
espécies de ciclídeos neotropicais amazônicos a partir de dados de
sequenciamento de nova geração e mapeamento cromossômico
comparativo de elementos transponíveis (L2, R4/Dong, RTE e Tc1) entre
as espécies com suporte de sequências nucleotídicas. As análises
evidenciaram ampla diversidade de elementos repetitivos na composição
genômica em ciclídeos neotropicais e diferentes padrões de organização
cromossômica de elementos transponíveis.
Palavras-chave: composição genômica, DNA repetitivo e
transponível, FISH
FICHA CATALOGRÁFICA

iv
Dedico essa dissertação aos meus pais
Edil e Cássia, ao meu amor Paulo e à
todos que acreditaram, confiaram e
participaram dessa conquista.

v
“Para achar a solução? Continue a nadar”
Dory, Procurando o Nemo

vi
A realização deste projeto foi possível devido:
Ao Programa de Pós-graduação em Genética, Conservação e Biologia
Evolutiva, do INPA.
Ao Laboratório de Citogenômica da UFAM, Departamento de Genética, onde
a maior parte deste trabalho foi desenvolvida, com financiamento proporcionado pela
Fundação de Amparo à Pesquisa do Amazonas (FAPEAM FIXAM 017/2014).
Aos laboratórios Genômica Integrativa da Universidade Estadual Paulista
“Júlio de Mesquita Filho” – Botucatu/SP, onde análises do Repeat Explorer foram
realizadas e ao Laboratório de Tecnologias de DNA da Universidade Federal do
Amazonas – Manaus/AM, onde as clonagens dos DNAs transponíveis foram
desenvolvidas.
À Fundação de Amparo à Pesquisa do Amazonas (FAPEAM) pela concessão
da bolsa de estudo (Projeto 007/2014) durante a realização deste trabalho.

vii
Agradecimentos
Agradeço a todas as pessoas, colegas e amigos que, direta ou indiretamente,
possibilitaram a realização deste trabalho.
Aos meus amados pais Edil Tavares e Cássia Mitozo, que sempre com muito
amor e carinho, me deram o suporte necessário para o meu desenvolvimento, me
ensinando a fazer sempre o meu melhor e que, para mim, são fontes de inspiração e
admiração. À minha mãe, que com seu amor e ternura, sempre tolerou a pior parte
de mim e mesmo assim continuava sendo sempre gentil e bondosa. Ao meu pai, que
nunca limitou minhas decisões e sempre me apoiou cegamente em todas elas. Aos
meus irmãos que com orgulho falam de mim. Amo todos vocês, minha família.
Ao meu grande amor Paulo Rodrigues, sem o qual muitas vezes teria
desistido. Agradeço por sempre acreditar no meu potencial e por inúmeras vezes me
convencer que eu conseguiria. Por ser um cara fantástico, o qual me motiva
cotidianamente e me lembra do valor que tenho, mesmo quando esqueço. Por ser
alguém que incansavelmente me instiga a ser grande, especialmente na minha
profissão e que ensinou, com seu próprio exemplo, que não há limites quando se
tem um sonho e o quer de verdade. Agradeço também por ser sempre tão amoroso
e carinhoso comigo, mesmo nos dias difíceis, por sempre me fazer rir como suas
brincadeiras inusitadas, me poupando de cair na rotina e por sempre fazer eu me
sentir especial. Amo você.
Ao meu excelentíssimo orientador, Doutor Carlos Schneider, que antes de
tudo foi um impulsionador da minha carreira acadêmica e a quem sempre serei grata
por ter acreditado em mim. Carlão, mano, você é um orientador extraordinário e eu
não poderia ter feito melhor escolha ao decidir trabalhar com você, ou o contrário
hahahaha, fato que é um grande privilégio para mim. Você é muito mais que um
orientador, é um pai acadêmico que de forma muita sábia sabe conduzir pelo
caminho certo e sempre me instiga a pensar mais. É alguém: em quem se pode
confiar, com quem se tem prazer em trabalhar, que estimula e dá ânimo, que
convence de forma gentil e respeitosa e prova que existem pessoas brilhantes no
meio cientifico. Quero muito ser como você um dia!
À Doutora Maria Claudia, que mesmo não sendo oficialmente ligada a mim,
emocionalmente assumiu esse posto, me recebendo de braços abertos, sempre me

viii
dando suporte, sendo provedora e fornecendo ajuda quando precisei. Você é
fantástica e jamais esquecerei toda ajuda que me deu, sem olhar pra trás e sem
nem me conhecer tão bem. O mundo precisa de mais Marias-Cláudias para ser um
lugar melhor.
Ao Dr. Cesar Martins, que abriu as portas de seu laboratório e concordou em
ser meu co-orientador, participando desse trabalho da melhor forma possível.
À Érica Ramos, que forneceu grandes contribuições nesse trabalho.
Aos meus amigos do LACA, sem os quais eu não teria conseguido: Nathy, a
quem devo um agradecimento especial por ter toda a paciência e zelo de me ensinar
tudo o que eu precisava saber de bancada e fora dela, e que sempre se apresenta
prestativa e disposta a ajudar, dando sempre o melhor de si; ao Léo, cujas
conversas são sempre agradáveis, instigantes e carregadas de ensinamentos
valiosos e que muitas vezes mudaram minha forma de pensar e acrescentaram
conhecimento que carregarei sempre comigo; Francy (minha patroa), que com seu
jeito divertido e espontâneo, faz todo dia ser mais legal; Francy (minha patroa), que
com seu jeito divertido e espontâneo, faz todo dia ser mais legal (a repetição não foi
um erro!); Sabrina (minha querida xará), com a qual vivi experiências exclusivas e a
quem tenho muito respeito e admiração; Érika, que com seu jeito todo meigo e gentil
me ajudou nessa conquista; Marcos, com quem sempre pude contar, mesmo em
cima da hora; Phamela, que conquistou seu espaço e nos alegra com sua presença,
especialmente nas tardes de conversa; Vanessa Pinheiro, que com seu jeito
carinhoso sempre busca estar de boa com tudo e com todos. Agradeço a todos
vocês.
Ao doutorando Edson, que nos auxiliou com as clonagens e o fez com muito
zelo.
Às minhas queridas amiga Israela e Paola, sem as quais eu não teria nem me
graduado, nem feito a seleção do mestrado, nem conseguido várias outras
conquistas. Meninas eu estaria bem perdida sem vocês. Agradeço por fazerem parte
disso ao ouvirem minhas queixas e alegrias, me auxiliarem nas minhas indagações
e estarem sempre comigo em todas as circunstâncias. Vocês são demais!

ix
Resumo
Os ciclídeos apresentam diferentes adaptações morfológicas, ecológicas e
comportamentais, promovendo sua adaptação em diversos ambientes ao longo da
evolução. Essa diversidade é refletida na composição citogenômica do grupo, que
apresenta disparidade na fórmula cariotípica e número diploide entre espécies
próximas filogeneticamente. As diferenças na composição e estrutura cariotípica
dessas espécies estão diretamente relacionadas à fração repetitiva do genoma, pois
correspondem à maior parte, e favorecem eventos de rearranjo cromossômico, que
podem ser observados pela técnica de hibridização fluorescente in situ. O
mapeamento físico cromossômico de ciclídeos neotropicais amazônicos limita-se a
elementos pontuais que são os principais alvos de estudos citogenéticos, como os
retroelementos não-LTR da família Rex e DNAs ribossomais. Contudo, outros
participantes da porção repetitiva do genoma ainda não foram mapeados até o
presente estudo. Além disso, estudos em ciclídeos africanos, grupo filogenético
irmão dos neotropicais, indicam que a composição genômica repetitiva dessas
espécies é muito mais abrangente. Graças à recente disponibilidade do
sequenciamento genômico de machos e fêmeas das espécies amazônicas Cichla
vazzoleri, Astronotus ocellatus, Pterophyllum scalare e Symphysodon discus, o
presente estudo identificou, a partir de uma pequena fração do genoma total destas
espécies, outras classes de elementos repetitivos a fim de mapeá-los nos
cromossomos. Os elementos repetitivos identificados foram repetições simples e
DNA satélites, além de diferentes classes de retrotransposons não-LTRs do tipo
LINE, retrotransposons LTRs do tipo ERV e transposons do tipo Tc1. Além disso, foi
possível identificar a estimativa da proporção genômica e relacionar a abundância
do número de cópias de cada elemento com a filogenia do grupo hospedeiro,
fornecendo dados preliminares sobre a participação de transponíveis na evolução. A
partir dessa abordagem inicial, foram selecionados os elementos transponíveis mais
abundantes para serem utilizados como sondas no mapeamento físico
cromossômico: retrotransposons não-LTRs lines L2, RTE e Dong.R4 e o transposon
Tc1. As marcações foram conspícuas e dispersas e não se restringiram às regiões
de heterocromatina. Algumas peculiaridades foram observadas, como a do
retroelemento L2 em Astronotus ocellatus, cujas marcações foram majoritariamente

x
conspícuas nas regiões centroméricas e terminais, com exceção do par
cromossômico 10 que apresentou fortes marcações dispersas ao longo de todo o
segmento cromossômico. Uma investigação adicional foi realizada a fim de
identificar possíveis variações dos elementos utilizados como sondas, pela técnica
de clonagem in vivo. De maneira geral, as sequências são relativamente
conservadas, apresentando alto grau de identidade entre si. Dados genômicos dos
elementos transponíveis combinado ao mapeamento cromossômico e análise
comparativa de suas sequências nucleotídicas fornecem dados preliminares sobre a
participação na composição, organização e evolução citogenômica de ciclídeos
neotropicais amazônicos.

xi
Abstract
Cichlids have different morphological, ecological and behavioral adaptations,
promoting their adaptation in different environments throughout evolution. This
diversity is reflected in the composition of cytogenomics group, which shows disparity
in karyotype formula and diploid number of phylogenetically related species. The
differences in composition and structure variations of these species are directly
related to the repetitive fraction of the genome, since they correspond to most of the
genome, and promotes chromosomal rearrangement events that can be observed by
fluorescent in situ hybridization technique. Chromosome physical mapping of
neotropical amazonian cichlid limited to specific elements that are the main targets of
cytogenetic studies, such as non-LTR retrotransposons Rex and ribosomal DNA
family. However, other participants in the repetitive portion of the genome have not
yet been mapped until this present study. In addition, studies in African cichlids,
brother phylogenetic group of neotropical, indicate that repetitive genomic
composition of these species is much broader. Thanks to the recent availability of
genomic sequencing of males and females of Amazonian species Cichla vazzoleri,
Astronotus ocellatus, Pterophyllum scalare and Symphysodon discus, this study has
identified, from a small fraction of the total genome of these species, other classes of
repetitive elements in order to map them on chromosomes. The repetitive elements
were identified as simple repeat and satellites DNA, and different classes of non-
LTR retrotransposons LINE-like, LTR retrotransposons ERV-like and transposons
Tc1-like. Moreover, it was possible to identify the proportion of each element in
genomic sample used and relate the abundance of the number of copies of each
element with the phylogeny of the host group, providing preliminary data on the
participation of transposable throughout evolution. From this initial approach, the
transposable elements more abundant were selected for use as probes in
chromosomal physical mapping: retrotransposons non-LTR lines L2, RTE and
Dong.R4 and transposons Tc1. The markings were conspicuous and widespread and
not restricted to regions of heterochromatin. Some peculiarities were observed, such
as retroelement L2 in Astronotus ocellatus, whose markings were mostly
conspicuous in the centromeric and terminal regions, with the exception of
chromosome pair 10 which showed strong marks scattered throughout the
chromosome segment. Further investigation was conducted to identify possible

xii
variations of elements used as probes, the technique of in vivo cloning. In general,
the sequences are relatively conserved, showing high degree of identity each other.
Genomic data of transposable elements combined with its chromosomal mapping
and their comparative nucleotide sequences provide preliminary data on the
participation in the composition, organization and cytogenomics evolution of
neotropical Amazonian cichlids.

xiii
SUMÁRIO
1. Introdução ............................................................................................................. 18
1.1 Ciclídeos como modelo de estudos comparativos ........................................ 18
1.2 DNA repetitivo e Elementos transponíveis .................................................... 21
1.3 Sequenciamento de nova geração no estudo de elementos transponíveis .. 25
1.4 Mapeamento físico cromossômico ................................................................ 26
1.5 Justificativa ................................................................................................... 27
1.6. Objetivo Geral .............................................................................................. 27
1.6.1 Objetivos Específicos ........................................................................ 27
2. Material e Métodos ................................................................................................ 28
2.1 Coleta das amostras ..................................................................................... 28
2.2 Extração de ADN e Sequenciamento de Nova Geração............................... 28
2.3 Preparação cromossômica ........................................................................... 30
2.4 Identificação de elementos transponíveis ..................................................... 30
2.5 Primers e amplificação por reação em cadeia da polimerase (PCR) ............ 31
2.6 Clonagem ...................................................................................................... 33
2.7 Hibridização fluorescente in situ (FISH – Fluorescence in situ hybridization)34
2.7.1 Tratamento das lâminas .................................................................... 34
2.7.2 Fixação .............................................................................................. 34
2.7.3 Pré-hibridização ................................................................................. 35
2.7.4 Solução de hibridização ..................................................................... 35
2.7.5 Hibridização ....................................................................................... 35
2.7.6 Lavagens ........................................................................................... 35
2.7.7 Detecção ........................................................................................... 35
2.7.8 Montagem das lâminas ...................................................................... 36
2.8 Processamento microscópico ....................................................................... 36
3. Resultados e Discussão ........................................................................................ 36
3.1 Artigo 1: Composição e Representatividade de Elementos Repetitivos no
Genoma de Ciclídeos Neotropicais ..................................................................... 37
3.1.1 Introdução .......................................................................................... 37
3.1.2 Material e Métodos ............................................................................ 40
3.1.2.1 Extração de DNA e Sequenciamento de Nova Geração .......... 40

xiv
3.1.2.2 Identificação de elementos transponíveis ................................ 41
3.1.3 Resultados ......................................................................................... 42
3.1.3.1 Diferença de 500 reads entre as espécies ............................... 45
3.1.3.2 Diferença de 500 reads entre os sexos da mesma espécie ..... 48
3.1.3.3 Altamente representativo em uma das espécies ...................... 48
3.1.4 Discussão .......................................................................................... 49
3.1.4.1 Diferença na composição genômica entre as espécies ........... 50
3.1.4.1.1 Retrotransposons não LTR ................................................ 50
3.1.4.1.2 Transposons ...................................................................... 52
3.1.4.2 Diferença na composicão genômica entre os sexos da mesma
espécie ................................................................................................. 52
3.1.4.2.1 Transposons e Retrotransposons não LTR ....................... 53
3.1.4.2.2 Repetições simples............................................................ 54
3.1.4.3 Altamente Representativo em uma das Espécies .................... 54
3.1.4.3.1 Retrotransposons não LTR ................................................ 54
3.1.4.3.2 Retrotransposos LTR ......................................................... 54
3.2 Artigo 2: Composição, Diversidade e Mapeamento Cromossômico de
Elementos Transponíveis no Genoma de Ciclídeos Neotropicais ...................... 56
3.2.1 Introdução .......................................................................................... 56
3.2.2 Material e Métodos ............................................................................ 58
3.2.2.1 Preparação cromossômica ....................................................... 58
3.2.2.2 Extração de DNA e amplificação por reação em cadeia da
polimerase (PCR) ................................................................................. 59
3.2.2.3 Clonagem, sequenciamento e análise das sequências ........... 59
3.2.2.4 Hibridização fluorescente in situ (FISH – Fluorescence in situ
hybridization) ........................................................................................ 60
3.2.2.5 Processamento microscópico .................................................. 60
3.2.3 Resultados ......................................................................................... 60
3.2.3.1 Clonagem ................................................................................. 60
3.2.3.2 FISH ......................................................................................... 62
3.2.4 Discussão .......................................................................................... 68
3.2.4.1 Diversidade de ETs no genoma de ciclídeos amazônicos ....... 68

xv
3.2.4.2 Distribuição de elementos transponíveis no genoma de
ciclídeos amazônicos ........................................................................... 72
4. Conclusões Gerais .......................................................................................... 77
5. Referências Bibliográficas ............................................................................... 78

xvi
Lista de figuras
Introdução
Figura 1: Gêneros de ciclídeos amazônicos evidenciando variações morfológicas. 18
Figura 2: Árvore filogenética de máxima parcimônia com identificação a partir de
família, seguido por subfamília, tribo, subtribo e gênero (Smith et al., 2008). ........... 20
Figura 3: Adaptado de Klug et al., 2013. Organização dos elementos repetitivos por
tipo e unidade de repetição. ...................................................................................... 22
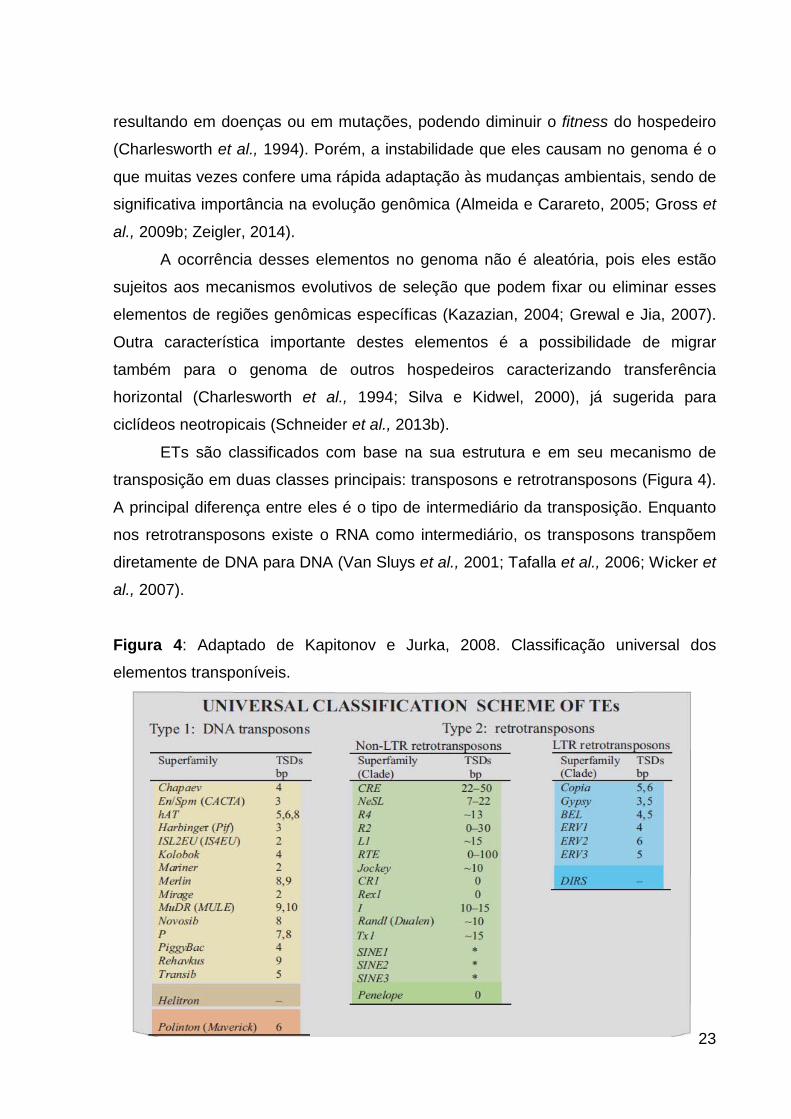
Figura 4: Adaptado de Kapitonov e Jurka, 2008. Classificação universal dos
elementos transponíveis. .......................................................................................... 23
Artigo 1
Figura 1: Clusters organizados por critério de escolha ............................................. 44
Figura 2: Elementos repetitivos e superfamílias de transponíveis identificados e suas
proporções em uma amostragem genômica de ciclídeos neotropicais amazônicos. 44
Artigo 2
Figura 1 Cariótipos das espécies estudadas hibridizadas com sonda de Mariner/Tc1.
Os cromossomos foram contra corados com DAPI. Barra 10 um. ............................ 64
Figura 2 Cariótipos das espécies modelo hibridizadas com sonda de Line/R4/Dong.
Os cromossomos foram contra corados com DAPI. Barra 10 um. ............................ 65
Figura 3 Cariótipos das espécies estudadas hibridizadas com sonda de Line/L2. Os
cromossomos foram contra corados com DAPI. Barra 10 um................................... 65
Figura 4: Cariótipos das espécies estudadas hibridizadas com sonda de Line/RTE.
Os cromossomos foram contra corados com DAPI. Barra 10 um. ............................ 67

xvii
Lista de tabelas
Introdução
Tabela 1: Lista de primers sintetizados para o isolamento dos elementos repetitivos.
.................................................................................................................................. 32
Artigo 1
Tabela 1: Dados do sequenciamento genômico total e da amostragem utilizada na
investigação da fração repetitiva de quatro espécies de ciclídeos neotropicais
amazônicos. (CV) Cichla vazolleri, (AO) Astronotus ocellatus, (PS) Pterophyllum
scalare, (SD) Symphysodon discus, (F) fêmeas e (M) machos................................. 43
Tabela 2: Clusters que apresentam diferença de pelo menos 500 reads entre
espécies. ................................................................................................................... 45
Tabela 3: Número de reads por elemento e por espécie. (CV) Cichla vazolleri, (AO)
Astronotus ocellatus, (PS) Pterophyllum scalare, (SD) Symphysodon discus, (F)
fêmeas e (M) machos. ............................................................................................... 46
Tabela 4: Espécies que apresentaram diferença no número de cópias de elementos
transponíveis e outros repetitivos entre sexos. ......................................................... 48
Tabela 5: Clusters compostos predominantemente por reads de uma das espécies.
.................................................................................................................................. 49
Artigo 2
Tabela 1: Lista de primers utilizados. ........................................................................ 59

18
1. Introdução
1.1 Ciclídeos como modelo de estudos comparativos
Os ciclídeos são a maior família de peixes acantomorfos e o maior clado de
euteleósteos dulcícolas e pertencem a família Cichlidae (Smith et al., 2008). Sua
distribuição abrange a África, América do Sul e Central, Ilha das Antilhas,
Madagascar, Índia e Sri Lanka (Smith et al., 2008). Os ciclídeos englobam uma
ampla gama de espécies de peixes, sendo o número de espécies neotropicais
superior a 350 somente na América do Sul (Smith et al., 2008).
Os ciclídeos apresentam morfologia bem variada, tanto na forma corpórea
quanto na coloração e, por isso, são massivamente usados como peixes
ornamentais na aquariofilia, principalmente os gêneros Pterophyllum e
Symphysodon (Figura 1). A maioria desses peixes habitam ambientes lênticos de
rios e córregos, alimentam-se de plânctons, invertebrados e matéria vegetal, embora
alguns gêneros como Cichla e Astronotus, se alimentem também de outros peixes,
apresentam dimorfismo sexual e cuidado parental. Já foram registrados em
praticamente todas as bacias hidrográficas da América do Sul, mas raramente
ocorrem em altitudes superiores a 500 m e ocorrem preferencialmente até 200 m de
altitude (Kullander 2003).
Nesse contexto, é fácil notar que os ciclídeos são um importante grupo de
estudos evolutivos devido à sua expressiva radiação adaptativa ao longo da
evolução, à diversidade de habitats que povoam e às suas adaptações morfológicas
e comportamentais (Kocher, 2004; Fan et al., 2012; Schneider et al., 2013a).
Figura 1: Gêneros de ciclídeos amazônicos evidenciando variações morfológicas.
Symphysodon sp. Pterophyllum sp. Astronotus sp. Cichla sp.

19
Os ciclídeos amazônicos destacam-se como fonte protéica, para fins
ornamentais e pesca de subsistência e esportiva. O tucunaré (gênero Cichla)
movimenta um forte mercado econômico relacionado à prática de pesca desportiva,
o qual atrai praticantes devido ao seu comportamento agressivo (Figura 1). Este tipo
de pesca tem aumentado no decorrer dos anos, principalmente na região que
abrange o médio rio Negro e seus afluentes (Freitas e Rivas, 2006). No que diz
respeito à alimentação, os peixes são a base alimentar em regiões ribeirinhas e nos
grandes centros amazônicos, sendo espécies de ciclídeos amplamente consumidas
o acará-açu (Astronotus spp.) e o tucunaré (Cichla spp.), onde dados pesqueiros
chegam a registrar um desembarque de mais de 5.000 toneladas por ano da última
espécie (Figura 1) (Cabral Júnior e Almeida, 2006).
A bacia do rio Negro, principalmente na região média do seu curso, ocorre a
exploração de peixes ornamentais, e os ciclídeos correspondem a grande parte dos
animais exportados para mercados da Ásia e Europa (Anjos et al., 2007). Duas
espécies se destacam, são elas o acará-bandeira (Pterophyllum spp.), com mais de
6.000 exemplares exportados legalmente em 2007 e o acará-disco (Symphysodon
spp.), com mais de 42.000 exportações na mesma época (Anjos et al., 2007).
A família Cichlidae é dividida em 4 subfamílias, Etroplinae, Ptychochrominae
Pseudocrenilabrinae e Cichlinae sendo que as duas últimas correspondem aos
ciclídeos africanos e neotropicais, respectivamente (Sparks e Smith, 2004; Smith et
al., 2008). A partir de análises moleculares combinadas às características
morfológicas, esses dois clados são considerados monofiléticos e irmãos (Figura 2).
Vale ressaltar que os ciclídeos neotropicais apresentam um nível significativamente
maior de variabilidade genética do que seu grupo irmão africano. Essa hipótese é
defendida pelo fato destes peixes apresentarem taxas aceleradas de evolução
molecular em relação à outra subfamília (Smith et al., 2008).
Na árvore filogenética mais recente proposta por Smith et al. (2008), os
gêneros amazônicos Cichla e Astronotus são considerados basais e Pterophyllum e
Symphysodon gêneros derivados na subfamília Cichlinae (Figura 2). Essa proposta
filogenética corrobora as descrições citogenéticas propostas para a subfamília. O
número diploide mais frequente para espécies de Cichlinae é 2n=48 cromossomos,
número considerado ancestral para esta subfamília e também para a ordem
Perciformes (Brum, 1995; Feldberg et al., 2003). Porém, de acordo com Feldberg et

20
al., (2003), três tendências de evolução cromossômica são evidenciadas nos
ciclídeos neotropicais: (i) manutenção do 2n=48 cromossomos considerado basal,
(ii) diminuição do 2n ou (iii) aumento do número diploide. Em contrapartida, dados
recentes tem demonstrado que estes caminhos evolutivos ocorrem
independentemente entre os gêneros (Schneider et al., 2013a).
Figura 2: Árvore filogenética de máxima parcimônia com identificação a partir de
família, seguido por subfamília, tribo, subtribo e gênero (Smith et al., 2008).

21
As espécies Cichla vazzoleri, Astronotus ocellatus e Pterophyllum scalare
apresentam 2n=48 cromossomos, entretanto existem diferenças em sua fórmula
cariotípica. Cichla vazzoleri possui apenas cromossomos subtelo-acrocêntricos,
enquanto A. ocellatus e P. scalare possuem cariótipo com 16 cromossomos meta-
submetacêntricos + 32 subtelo-acrocêntricos (Schneider et al., 2013a). Por outro
lado os acarás-disco do gênero Symphysodon (S. aequifasciatus, S. discus e S.
haraldi) apresentaram número diploide igual a 2n=60 cromossomos, com a maioria
dos cromossomos meta-submetacêntricos, confirmando sua posição derivada
proposta pela filogenia vigente (Mesquita et al., 2008; Gross et al., 2009a; Schneider
et al., 2013b; Smith et al., 2008) (Figura 1).
As diferenças na composição e estrutura cariotípica dessas espécies estão
diretamente relacionadas a eventos de rearranjos cromossômicos, já reportados em
ciclídeos (Gross et al., 2010; Poletto et al., 2010) que podem estar relacionados à
porção repetitiva do genoma por serem alvo de quebra cromossômica,
principalmente aos elementos transponíveis.
1.2 DNA repetitivo e Elementos transponíveis
A porção codificante do DNA de eucariotos correspondente a uma parte muito
pequena em relação ao genoma total, cerca de 1,5-2% do genoma humano (Martins,
2007). Desta forma, a maior parte do genoma é composta por regiões intergênicas e
repetitivas, as quais por muito tempo foram consideradas dispensáveis ao DNA,
sendo chamadas de “DNA lixo” (Charlesworth et al., 1994; Martins, 2007).
Uma das possíveis razões para haver uma quantidade tão elevada de porção
repetitiva no genoma seriam eventos de duplicação, inserção e amplificação
oriundas de recombinação desigual de cromossomos homólogos (Martins, 2007). A
região repetitiva do genoma pode ser classificada de acordo com o número de
unidades de repetição, o comprimento dessas unidades e a frequência que ocorrem
(Wicker et al., 2007). Eles podem ser altamente repetitivos, representados por DNA
satélite ou moderadamente repetitivos com repetições in tandem, como genes de
cópias múltiplas, mini e microssatélites, ou dispersos, que são os elementos
transponíveis (Figura 3) (Charlesworth et al., 1994; Phillips e Reed, 1996; Martins,
2007). Os elementos transponíveis (ETs), também chamados de “jumping genes”, se
distinguem dos outros elementos repetitivos pela habilidade de replicar-se no

22
genoma independente do ciclo celular, além serem capazes de migrar para outras
regiões diferentes da origem, apresentarem sequências nucleotídicas codificadoras
e não ocorrer em amplas repetições (in tandem) (McClintock, 1950; Charlesworth et
al., 1994; Feschotte et al., 2008).
Inicialmente esses elementos eram considerados parasitas do DNA que se
tornaram não funcionais ao longo do tempo. Entretanto, uma pequena parcela pode
apresentar papéis funcionais no genoma (Zeigler, 2014). Diversos trabalhos têm
demonstrado que essa parte não codificante do DNA desenvolve um papel
fundamental na manutenção celular e estão envolvidos em mecanismos de
regulação determinantes na expressão gênica, podendo resultar em mudanças
fenotípicas, além de estarem envolvidos em processos de especiação e atuando na
evolução do genoma do hospedeiro (Kazazian, 2004; Martins, 2007; Vitte et al.,
2014; Zeigler, 2014). Em certas situações, os ETs podem se inserir em éxons,
Figura 3: Adaptado de Klug et al., 2013. Organização dos elementos repetitivos
por tipo e unidade de repetição.
23
resultando em doenças ou em mutações, podendo diminuir o fitness do hospedeiro
(Charlesworth et al., 1994). Porém, a instabilidade que eles causam no genoma é o
que muitas vezes confere uma rápida adaptação às mudanças ambientais, sendo de
significativa importância na evolução genômica (Almeida e Carareto, 2005; Gross et
al., 2009b; Zeigler, 2014).
A ocorrência desses elementos no genoma não é aleatória, pois eles estão
sujeitos aos mecanismos evolutivos de seleção que podem fixar ou eliminar esses
elementos de regiões genômicas específicas (Kazazian, 2004; Grewal e Jia, 2007).
Outra característica importante destes elementos é a possibilidade de migrar
também para o genoma de outros hospedeiros caracterizando transferência
horizontal (Charlesworth et al., 1994; Silva e Kidwel, 2000), já sugerida para
ciclídeos neotropicais (Schneider et al., 2013b).
ETs são classificados com base na sua estrutura e em seu mecanismo de
transposição em duas classes principais: transposons e retrotransposons (Figura 4).
A principal diferença entre eles é o tipo de intermediário da transposição. Enquanto
nos retrotransposons existe o RNA como intermediário, os transposons transpõem
diretamente de DNA para DNA (Van Sluys et al., 2001; Tafalla et al., 2006; Wicker et
al., 2007).
Figura 4: Adaptado de Kapitonov e Jurka, 2008. Classificação universal dos
elementos transponíveis.

24
De maneira geral, os transposons mobilizam-se no genoma pelo mecanismo
“corta-e-cola”, no qual transposases e ligases promovem a excisão do ET de uma
região e inserem-no em outra (Wicker et al., 2007; Kapitonov e Jurka, 2008). Fazem
parte dos transposons as superfamílias Mariner/Tc1, hAT, MuDR, EnSpm/CACTA,
PiggyBac, P, Merlin, Harbinger, Transib, Novosib, Kolobok, ISL2EU, Helitron,
Polinton, Mirage, Rehavkus e Chapaev (Figura 4) (Kapitonov e Jurka, 2008).
Os transposons são divididos em duas subclasses e ambas apresentam
genes codificantes para a transposase. A subclasse 1, que é a maior em número de
superfamílias, contém também repetições terminais invertidas (TIRs). Já a subclasse
2 transpõem-se sem a quebra das duas hélices do DNA e somente uma das fitas é
clivada e inserida em outra região do genoma e engloba as superfamílias Helitron e
Polinton (Maverick) e Crypton (Wicker et al., 2007).
Os retrotransposons mobilizam-se no genoma gerando uma cópia de si
mesmos e inserindo-a em outra região gerando um intermediário de RNA, com a
participação de transcriptase reversa. Esse mecanismo é conhecido como “copia-e-
cola”, onde o número de cópias desse retroelemento sempre aumenta no genoma
(Kapitonov e Jurka, 2008). São classificados nas classes LTR (long terminal
repeats), da qual fazem parte as superfamílias Copia, Gypsy, BEL, DIRS e retrovírus
endógenos e a classe não-LTR composta pelas superfamílias CRE, NeSL, R4, R2,
L1, RTE, Jockey, CR1, Rex1, I, RandI (Dualen), Tx1, SINEs e Penelope (Figura 4)
(Kapitonov e Jurka, 2008).
Elementos LTR são estruturalmente similares. Além das longas regiões
terminais, normalmente eles contêm open read frame (ORF) para GAGs (codificam
proteínas do capsídeo), proteinase aspártica (AP), transcriptase reversa (RT),
RNase H (RH) e integrase DDE (INT). São separados em clados distintos por
diferenças na filogenia da transcriptase reversa e pela presença de ENV (codificam
proteínas envelope), no caso de ERVs. Os DIRS apresentam uma estrutura muito
similar a essa, salvo pela ausência de INT e presença de tirosina recombinase (TR)
(Wicker et al., 2007).
A maior parte dos não-LTRs é composta pela ordem LINE que apresenta
constituição bem diferenciada entre as superfamílias, salvo pela transcriptase
reversa e nuclease que estão presente em todas elas (Wicker et al., 2007).

25
Independente da classe, os ETs são abundantes no genoma e amplamente
diversificados, e com o advento de novas tecnologias de sequenciamento genômico,
novas descrições só tendem a aumentar (Wicker et al., 2007).
1.3 Sequenciamento de nova geração no estudo de elementos transponíveis
Com o advento de tecnologias de sequenciamento de nova geração (next
generation sequencing - NGS), os dados gerados com essas ferramentas possuem
dimensões astronômicas, com o melhor custo-benefício (Wicker et al., 2007; Novak
et al., 2010). Entretanto, a abundância e diversificação dos ETs no genoma, pode
representar um grande problema no sequenciamento genômico total, devido aos
procedimentos necessários para executá-lo (Wicker et al., 2007). Para o
sequenciamento, é necessário fragmentar o DNA em regiões menores que depois
serão organizadas em sequência, a partir da sobreposição dos fragmentos - reads -
(Lapidus, 2009). O grande problema encontra-se nas regiões repetitivas, pois essa
fragmentação produz segmentos que podem conter mais de uma combinação e,
muitas vezes, essas combinações possuem muitas possibilidades formando grandes
agrupamentos - clusters – (Novak et al., 2010).
Apesar de a abundância de ETs no genoma representar um problema de
anotação e identificação em NGS, trabalhos recentes têm apresentado ferramentas
desenvolvidas para análises de ETs a partir de dados massivos obtidos por NGS e
vem apresentando resultados satisfatórios. Uma delas é o Repeat Explorer (Novak
et al., 2010) que constrói gráficos baseados em agrupamento e caracterização de
elementos repetitivos em dados de NGS. Esta análise é baseada na comparação all-
to-all dos reads a fim de identificar similaridades entre eles e construir clusters pela
sobreposição (overlaps) dos reads representando diferentes elementos repetitivos.
O tamanho dos clusters (número de reads dentro dos clusters) é usado para
quantificar famílias distintas de elementos transponíveis (Novak et al., 2010).
Essa ferramenta apresenta-se satisfatória na caracterização de ETs de dados
de NGS de estudos em plantas e apresenta-se promissora para outros grupos de
organismos. Seus resultados proporcionam informações necessárias para inferência
da evolução das famílias de elementos transponíveis, descoberta e caracterização
de novos elementos e ajuda na montagem de sequências consenso (Novak et al.,
2010).

26
A identificação massiva dos ETs presentes no genoma proporciona matéria
prima para o mapeamento físico cromossômico, possibilitando assim a identificação
de rearranjos cromossômicos, embasando estudos evolutivos e permitindo melhor
interpretações do panorama genômico de diferentes espécies.
1.4 Mapeamento físico cromossômico
A citogenética molecular é utilizada para mapeamento físico de sequências de
DNA específicas no cromossomo, consistindo basicamente no pareamento de
determinado segmento de DNA ou RNA com uma sequência de nucleotídeos
complementar situado na célula alvo (Guerra, 2004).
Na citogenética de ciclídeos neotropicais a hibridização fluorescente in situ
(FISH) revela-se importante, pois tem permitido novas interpretações da diversidade
cariotípica e sobre a organização genômica de segmentos cromossômicos tanto de
DNAs ribossomais e sequências teloméricas, quanto de elementos transponíveis
(Vicari et al., 2006; Gross et al., 2009b; Mazzuchelli e Martins, 2009; Teixeira et al.,
2009; Gross et al., 2010; Poletto et al., 2010; Valente et al., 2011; Schneider et al.,
2013a; Schneider et al., 2013b).
Diferentes classes de ETs foram mapeadas no genoma dos ciclídeos
neotropicais, tais como transposon Tc1 que foi mapeado em Cichla kelberi
evidenciando blocos conspícuos em regiões centroméricas e marcações dispersas
nos cromossomos (Teixeira et al., 2009). Três novas classes de retroelementos
foram descritas: RCk que apresenta marcações dispersas nos cromossomos de C.
kelberi; AoRex3 e AoLINE que são agrupados na heterocromatina centromérica de
todos os cromossomos de Astronotus ocellatus (Mazzuchelli e Martins, 2009).
A maioria dos estudos focam no mapeamento dos retroelementos Rex (Rex 1,
3 e 6) e os resultados têm mostrado um padrão de distribuição compartimentalizado
na heterocromatina pericentromérica, entretanto sinais dispersos e agrupados em
regiões eucromáticas também foram observados (Gross et al., 2009b; Mazzuchelli e
Martins, 2009; Teixeira et al., 2009; Valente et al., 2011, Schneider et al., 2013b).
Desta forma, faz-se necessário identificar outras classes classe de elementos
transponíveis, a fim de conhecer sua sequência e utilizá-las como sondas para o
mapeamento físico cromossômico, dando suporte para identificação de rearranjos
cromossômicos, compreensão da organização cariotípica e interpretações sobre a

27
participação de elementos transponíveis na dinâmica evolutiva dos ciclídeos
neotropicais amazônicos.
1.5 Justificativa
Ciclídeos têm importante relevância ecológica, uma vez que correspondem à
família mais rica em número de espécies de peixes de água doce e exibem
expressiva radiação adaptativa distribuída em quase todos os continentes. Também
exercem impacto social e econômico, na pesca esportiva, aquariofilia e economia
regional de muitas comunidades ribeirinhas no Amazonas, além de serem descritos
como um excelente grupo para estudos evolutivos. Algumas espécies de ciclídeos
africanos já tiveram seu genoma sequenciado (Cichlid Genome Consortium Program
em http://cichlid.umd.edu/cichlidlabs/kocherlab/bouillabase.html), bem como algumas
espécies sul americanas, abrindo novos horizontes para integração entre
sequências nucleotídicas e mapeamento físico cromossômico, uma vez que os
dados sobre esta ainda são incipientes.
Considerando que a grande parte do mapeamento físico cromossômico de
ETs é focada em retroelementos não-LTRs da família Rex, a identificação e o
mapeamento físico de outras classes de ETs em ciclídeos neotropicais permitirá
uma melhor compreensão da organização genômica bem como o entendimento da
dinâmica dos retroelementos na evolução cromossômica deste grupo de peixes.
1.6. Objetivo Geral
Comparar elementos repetitivos das espécies Cichla vazzoleri, Astronotus
ocellatus, Pterophyllum scalare e Symphysodon discus a fim de compreender sua
organização genômica e sua evolução cariotípica, bem como inferir a dinâmica
desses elementos.
1.6.1 Objetivos Específicos
- Isolar e caracterizar comparativamente os elementos repetitivos das quatro
espécies de ciclídeos neotropicais;
- Identificar e mapear elementos transponíveis destas espécies, utilizando as
sequências repetitivas isoladas;

28
- Analisar comparativamente as sequências nucleotídicas dos elementos
transponíveis entre as espécies modelo.
2. Material e Métodos
2.1 Coleta das amostras
Quatro espécies de ciclídeos neotropicais da Amazônia Central foram
analisados, sendo elas basais Cichla vazzoleri (3 machos e 3 fêmeas) e Astronotus
ocellatus (3 machos e 5 fêmeas), e derivadas Pterophyllum scalare (3 machos e 3
fêmeas) e Symphysodon discus (2 machos e 2 fêmeas). Amostras de tecido foram
retiradas para realização do sequenciamento genômico e para preparações
cromossômicas. As espécies foram coletas com autorização do ICMBio (ICMBio
SISBIO 22984-1/2011).
Exemplares de Cichla vazzoleri foram coletados nos meses de março e abril
de 2011 no lago da Usina Hidroelétrica de Balbina (Presidente Figueiredo-AM),
bacia do rio Uatumã̃ afluente do rio Amazonas (3°09’57”S, 59°54’44”W).
Espécimes de Symphysodon discus foram coletados em igarapés próximos a
Barcelos AM (bacia do rio Negro) no período de abril de 2010 a março de 2011
(0°52'2.29"S, 62°48'35.61"W).
Os indivíduos de Astronotus ocellatus foram coletados no mês de abril de
2010 e os indivíduos de Pterophyllum scalare foram coletados no mês de fevereiro
de 2010. Ambas as espécies, A. ocellatus e P. scalare, foram coletadas no lago
Catalão, região de confluência entre os rios Negro e Solimões (3°09'45,53''S e
59°55'05,6''W).
Dados citogenéticos clássicos tais como número diploide, localização de
heterocromatina e regiões organizadoras de nucléolo já foram previamente descrita
(Schneider et al., 2013b).
2.2 Extração de ADN e Sequenciamento de Nova Geração
Os dados utilizados nesse trabalho foram fornecidos pela parceria
estabelecida com o Laboratório Genômica Integrativa do Instituto de Biociências de
Botucatu, São Paulo, o qual realizou o sequenciamento genômico total das quatro

29
espécies investigadas e onde estão sendo montados e anotados os genomas das
mesmas.
Os espécimes foram sacrificados pela imersão em água gelada e tecidos
muscular e hepático foram recolhidos para extração de DNA genômico, tanto para o
sequenciamento genômico quanto para as preparações cromossômicas. O DNA
total foi extraído usando fenol-clorofórmio (Sambrook et al., 1989) e quantificado
pelo espectrofotômetro NanoDrop 2000 (Thermo Scientific, Wilmington, DE, USA).
Reações de sequenciamento foram realizadas para 2 indivíduos (macho e
fêmea) de cada espécie (A. ocellatus, C. vazzoleri, P. scalare e S. discus). As
amostra de DNA de cada espécie foram fragmentadas em tamanhos com cerca de
500 pb usando um ultrasonicador S220 (Covaris Inc., Woburn, MA, USA). As
bibliotecas genômicas foram construídas pelo Sequencing Facility da Universidade
de Maryland, USA (http://www.ibbr.umd.edu/facilities) utilizando o kit TruSeq DNA
Sample Preparation (Illumina Inc., San Diego, California, USA). Os sequenciamentos
genômicos foram realizados utilizando a plataforma HiSeq 1000 Illumina, sendo dois
lanes por espécie em uma abordagem paired-end de acordo com as recomendações
do fabricante e geraram 8 conjuntos de reads genômicos.
O output foi composto por reads paired-end com tamanho médio aproximado
de 500 pares de base (pb) e a cobertura de cada genoma foi de ~32 a 40 vezes o
genoma de cada sexo e espécie. Todas as sequências foram submetidas à
avaliação de qualidade e de presença de adaptadores utilizando o software
FASTQC (website:http://www.bioinformatics.babraham.ac.uk/projects/fastqc/) a partir
dos valores de Phred quality score obtidos para cada base nucleotídica durante o
sequenciamento. Foram eliminados reads que não apresentassem 90% de seus
nucleotídeos com qualidade igual ou superior a 30 (Phred quality score). O número
de reads obtido foi: 267.797.946 para fêmea de A. ocellatus; 226.726.662 para
macho de A. ocellatus; 231.369.922 para fêmea de C. vazolleri; 227.201.454 para
macho de C. vazolleri; 248.087.060 para fêmea de P. scalare; 240.672.242 para
macho de P. scalare; 216.066.104 para fêmea de S. discus; e 251.236.834 para
macho de S. discus. O genoma completo de cada espécie estará disponível no
banco de dados público SaciBase (www.lgi.edu.br/sacibase).

30
2.3 Preparação cromossômica
Cromossomos mitóticos foram obtidos a partir de células renais usando o
protocolo de Bertollo et al.,. (1978) que consiste em injetar intraperitonealmente
colchicina 0,0125% na proporção de 1 mL para cada 100 g de peso do animal. Após
40 minutos, os espécimes foram sacrificados pela imersão em água gelada e a
porção anterior do rim foi removida e transferida para uma solução hipotônica de
KCL 0,075 M (6-8 mL). O tecido foi divulsionado com o auxílio de uma seringa de
vidro. Em seguida, o sobrenadante (suspensão celular) foi retirado com o auxílio de
uma pipeta Pasteur, colocado em tubo de centrífuga e incubado em estufa a 37 °C
por 30 minutos. A suspensão foi pré-fixada com 6 gotas de metanol:ácido acético
(3:1) e ressuspendida com cuidado. Após 5 minutos, foi adicionado 8ml de fixador
Carnoy e centrifugado por 10 minutos a 900 rpm. O sobrenadante foi descartado e 6
mL de fixador Carnoy foi adicionado. O material foi novamente ressuspendido e
centrifugado por 10 minutos a 900 rpm, repetindo essa lavagem por mais duas
vezes. Após a última centrifugação e a eliminação do sobrenadante, 1,5 mL de
fixador foram adicionados e o material ressuspendido com cuidado. A suspensão
celular foi então estocada em tubo “eppendorf” e mantida em freezer -20°C. Para a
preparação das lâminas as mesmas foram lavadas, secas ao ar e posteriormente
imersas em água destilada a 50 ºC, em banho-maria. Após cinco minutos, as
lâminas foram retiradas da água de forma a manter uma película de água sobre a
sua superfície, na qual foi gotejada a suspensão celular em diferentes regiões. As
lâminas secaram diretamente ao ar.
2.4 Identificação de elementos transponíveis
Após as filtragens iniciais e o preparo das bibliotecas para a análise de
clusterização, cerca de 700.000 reads (correspondente a 0,1x de cobertura do
genoma completo, cujo tamanho estimado é de 1Gb) de cada sexo e espécie foram
selecionados para identificar a composição da fração repetitiva dos genomas destas
espécies. Os reads desta amostragem de sequências de cópias repetitivas foram
filtrados, agrupados em clusters e identificados e caracterizados utilizando a
ferramenta Repeat Explorer - website http://www.repeatexplorer.org/ - (Novak et al.,
2013). Neste pipeline os reads de elementos repetitivos são agrupados por
identidade em uma comparação all-to-all formando clusters. Para que um read seja

31
incorporado ao cluster, ele precisa ter 55% de cobertura e 90% de identidade em
relação a outro read. Desta forma, cada clusters representa um grupo de sequências
mais similares entre si do que em relação às sequências de outro cluster. Os
clusters foram anotados utilizando o algoritmo Repeat Masker em conjunto com o
banco de dados Repbase Update (Jurka et al., 2005). A caracterização de cada
cluster contém informações específicas e revela a identidade do elemento
transponível de acordo com o Repbase Update e o número de hits, número de reads
total e discriminado por sexo e espécie, além de gráficos, domínios de proteínas e
outras informações. Os elementos foram identificados ao nível de superfamília
(Kapitonov e Jurka, 2008), portanto a ocorrência de mais de um cluster identificado
como o mesmo elemento, não significa que eles sejam iguais em níveis mais
específicos. A análise estatística é feita pela ferramenta SeqGrapheR, que distingue
tipos básicos de repetições e investiga a variabilidade de sequências dentro das
famílias de elementos repetitivos (Novak et al., 2010).
Com o intuito de estudar elementos repetitivos de dinâmicas diferentes entre
as espécies e entre os indivíduos de sexos diferentes, foram selecionados para as
próximas etapas os clusters com diferenças nos números de reads de cada espécie
(utilizados na montagem do mesmo elemento). Os clusters selecionados deveriam
apresentar diferença de no mínimo 500 reads: i) entre as espécies; ii) entre os sexos
da mesma espécie; iii) os altamente representativos em uma única espécie.
2.5 Primers e amplificação por reação em cadeia da polimerase (PCR)
Após a identificação e caracterização dos elementos repetitivos,
oligonucleotídeos iniciadores (primers) foram sintetizados a partir das sequências
consenso de cada cluster no programa Geneious ® 8.0.5 (Biomatters Limited -
http://www.geneious.com/), uma ferramenta que permite organizar, analisar e
manipular as sequências do output do Repeat Explorer, além de permitir
alinhamentos múltiplos e aos pares, inferência de árvores filogenéticas, assembly
dos contigs, desenho de primers para mapeamento genético das sequências de
interesse (Kearse et al., 2012). A lista dos primers desenhados que amplificam os
elementos repetitivos de cada cluster podem ser observados na Tabela 1.

32
Tabela 1: Lista de primers sintetizados para o isolamento dos elementos repetitivos.
ID ELEMENTO PRIMER FORWARD SEQUÊNCIA PRIMER REVERSE SEQUÊNCIA
07 LINE DONG R4 CL07R4-F WGGAATCTGCTGGAGCCAYT CL07R4-R ACTCTCAGTGGCAGGGGAAR
08 DNA.TcMar.Tc1 CL08Tc1-F CAATAAGTCAAGGACAACAC CL08Tc1-R CAAACCATCATGGCCCTGTGT
09 Satélite CL09Sat-F AGTCAGCCATCTTGGCGTGA CL09Sat-R TTGTGGTTCATGCTRTAGGG
10 LINE L2 CL10L2-F GCTTTTCAGCATCACTCCTG CL10L2-R ATGCAGATGATACCCAGGCT
11 LINE RTE CL11RTE-F ATGATGGYDGTYCTGTTGGC CL11RTE-R GTAGGACATGCCCKGGAACA
13 LINE L2 CL13L2-F TCGTGCTTGGAACTATAAGG CL13L2-R GCTTGTYTCAACAGAGGATG
16 LINE RTE CL16RTE-F CAGCAGCCTCAAACAATGGA CL16RTE-R CGGACCGMATCCCCGGCTAT
21 LINE L2 CL21L2-F AGGAAACATGATGTGGATGC CL21L2-R TAAAGTGAAAGGAACAGAGC
22 LINE L2 CL22L2-F CTTAGTTACAGCACTTTCAG CL22L2-R STGCCTTACAGAAACCTGGT
23 DNA.TcMar.Tc1 CL23Tc1-F CAAGTCCTGCACAGTGGTCA CL23Tc1-R TCATGGCTAAATTGGACCAG
24 Satélite CL24Sat-F CAACATCATCTAGAGAAGTG CL24Sat-R TGCTGCTGTGTATAATGCCT
28 DNA.TcMar.Tc1 CL28Tc1-F TGTGAACAGCAGTCTTCAGC CL28Tc1-R GTTTGAAGCCTGAAATGTGGC
33 LINE RTE CL33RTE-F TGCTGACTTCAACTGAGGCT CL33RTE-R GTAGACTGGWTGRGCAAACT
35 LINE Rex.Babar CL35Rex.Babar-F ATTTGGTTCACCCTGTGGCT CL35Rex.Babar-R TCAGAAGAAGTYGAAGAAGA
38 LINE Penelope CL38Penelope-F GGGAGAATAAGAATGACAAG CL38Penelope-R GATGGTTGTTCACAGTGGAC
41 LTR ERV1 CL41ERV1-F TCTGAGAAGATGTGTCCAGT CL41ERV1R TAAGAATCATCTGGCTTCTG
48 Satélite CL48Sat-F CCTTACCCTCCCATTAAGCC CL48Sat-R CATACCCCAAACCCTACAGG
63 LINE Penelope CL63Penelope-F AATGTTTGGCGGTCCTGCGT CL63Penelope-R CTCACAAGCAGGACTCCAGA
66 LINE Rex.Babar CL66Rex.Babar-F ACAATGTTACTAACTACTGG CL66Rex.Babar-R GGGTAAAGGTGACACTTGG

33
O software Geneious 8.0.5 ® (Biomatters Limited - http://www.geneious.com/)
permite ainda a busca por sequências similares públicas em banco de dados
genômicos online, como NCBI BLAST (Kearse et al., 2012). Desta forma, é possível
realizar comparações dos elementos transponíveis das espécies modelo com o
genoma de outros peixes.
A amplificação dos elementos repetitivos pela reação em cadeia da
polimerase (Polymerase Chain Reaction - PCR) foi realizada utilizando os primers
desenhados a partir das sequências consenso dos elementos repetitivos. A reação
de PCR foi realizada para o volume final de 25 µL, contendo DNA genômico (200
ng), tampão 1x com 1,5 mM de magnésio, Taq DNA polimerase (5 U/µL), dNTPs (1
mM), primers (5 mM) e água Milli-Q. As condições dos ciclos para todos os
elementos incluíram os seguintes passos: 95°C por 5 min; 34 ciclos de 95°C por 1
min, 55°C por 40 seg e 72°C por 2 min; e a extensão final a 72°C por 5 min. Os
produtos de PCR foram analisados por eletroforese em gel de agarose à 1%,
quantificados pelo espectrofotômetro NanoDrop 2000 (Thermo Scientific,
Wilmington, DE, USA) e usados na clonagem e como sonda na técnica de FISH.
De todos os 19 elementos transponíveis isolados por PCR, selecionamos as
quatro superfamílias mais abundantes no genoma para as etapas seguintes de
clonagem e mapeamento cromossômico, que foram: os elementos transponíveis
Mariner/Tc1 (ID08), Line/L2 (ID22), Line/R4/Dong (ID07) e Line/RTE (ID11).
2.6 Clonagem
A fim de investigar possíveis variações nas sequências amplificadas por estes
quatro primers, realizamos a clonagem, onde cada vetor só incorpora um produto de
PCR por vez que posteriormente é sequenciado e comparado aos outros fragmentos
do mesmo primer. Os produtos de PCR dos elementos transponíveis Mariner/Tc1
(ID 08), Line/L2 (ID 22), Line/R4/Dong (ID 07) e Line/RTE (ID 11) foram inseridos no
plasmídeo pGEM-T Easy (Promega). Os produtos de ligação foram inseridos em
células competentes de Escherichia coli linhagem DH5 por eletro e
quimiocompetência. Clones carregando o inserto de interesse foram sequenciados
pelo Sequenciador de DNA ABI 3130 XL (Perkin-Elmer) para confirmar a
amplificação da região de interesse e os resultados das sequências foram
submetidos no banco de dados Repbase Update. Cada clone foi identificado na

34
busca contra o banco de dados Repbase Update no Genetic Information Researche
Institute (GIRI) (http://www.girinst.org/repbase/) utilizando o software CENSOR
(Jurka et al., 2005). As sequências nucleotídicas foram alinhadas usando o
programa ClustalW implementado pelo programa Geneious ® 8.0.5 (Biomatters
Limited).
2.7 Hibridização fluorescente in situ (FISH – Fluorescence in situ hybridization)
Os produtos de PCR dos elementos transponíveis Mariner/Tc1 (ID08), Line/L2
(ID22), Line/R4/Dong (ID07) e Line/RTE (ID11) foram utilizados como sondas para o
mapeamento cromossômico por hibridização fluorescente in situ (FISH) homólogas e
heterólogas, de acordo com o protocolo de Pinkel et al,. (1986). Os produtos de PCR
desses elementos transponíveis foram marcados com digoxigenin-11-dUTP (mix
Dig-Nick Translation; Roche) por nick translation de acordo com as recomendações
do fabricante e a anti-digoxigenina rodamina (Roche) foi usada para a detecção do
sinal das sondas.
2.7.1 Tratamento das lâminas
As lâminas contendo preparações cromossômicas foram lavadas em tampão
PBS 1x por 5 minutos em temperatura ambiente. As lâminas foram desidratadas em
uma série alcoólica gelada (70%, 85% e 100%) durante 5 minutos cada. Em
seguida, tratadas com 90 μL de RNase 10 μg/mL (5 μL de RNase 10 mg/mL e 975
μL de 2XSSC) por 1 hora em câmara úmida a 37 ºC. As lâminas foram lavadas três
vezes em 2xSSC durante 5 minutos cada. Após isso, foram lavadas em PBS 1x
durante 5 minutos.
2.7.2 Fixação
As lâminas foram fixadas em formaldeído 1% em PBS 1X/50mM MgCl2
durante 10 minutos à temperatura ambiente. Posteriormente lavadas em PBS 1x por
5 minutos. Após, as lâminas foram desidratadas em uma série alcoólica gelada
(70%, 85% e 100%) durante 5 minutos cada.

35
2.7.3 Pré-hibridização
As lâminas foram desnaturadas em formamida 70% em 2xSSC a 70 ºC por 5
minutos e novamente desidratadas em etanol gelado 70%, 85% e 100% por 5
minutos cada.
2.7.4 Solução de hibridização
Em um tubo eppendorf foram adicionados 4 μL de sonda, 20 μL de
formamida, 8 μL de sulfato de dextrano 50% e 4 μL de 20xSSC. A sonda foi
desnaturada a 99 °C por 10 minutos e transferida imediatamente ao gelo.
2.7.5 Hibridização
Foram colocados 40 μL de solução de hibridização sobre uma lamínula e a
lâmina foi invertida sobre a lamínula. As lâminas foram mantidas com o material
voltado para baixo em câmara úmida (H2O destilada) a 37 °C por cerca de 14 horas.
2.7.6 Lavagens
As lamínulas foram removidas das lâminas. Em seguida foram lavadas duas
vezes em formamida 15% a 42°C durante 10 minutos cada. Em seguida, lavadas
novamente em solução Tween 0,5% durante 5 minutos a temperatura ambiente.
2.7.7 Detecção
As lâminas foram incubadas em tampão NFDM (Non Fat Dry Milk) por 15
minutos. Em seguida, lavadas duas vezes com solução Tween 5% por 5 minutos a
temperatura ambiente. Foram colocadas sobre cada lâmina 20 µL de anti
digoxigenina-rodamina e 100 µL de solução contendo tampão NFDM e
estreptavidina (2 μL de estreptavidina estoque em 998 μL de NFDM). Em seguida,
cobertas com lamínula e deixadas por 60 minutos em câmara úmida com água
destilada. Posteriormente as lamínulas foram removidas e as lâminas lavadas três
vezes em solução Tween 5% por 2 minutos a temperatura ambiente cada. As
lâminas foram desidratadas em uma série alcoólica 70, 85 e 100% durante 5 min
cada, secando ao final.

36
2.7.8 Montagem das lâminas
Foi adicionado a cada lâmina solução de DAPI diluído em antifade
VectaShield Vector (20 μL de antifade e 1 μL de DAPI).
2.8 Processamento microscópico
Quatro lâminas e no mínimo 20 metáfases foram analisadas por espécie para
cada ET. Cromossomos foram analisados em objetiva de 100x usando o
microscópio de epifluorescência Leica DM 2000 e as imagens foram capturadas com
uma câmera digital (Olympus DP 71) usando o software Leica LAS 4.0. Metáfases
mitóticas foram processadas pelo programa Adobe Photoshop CS6. Os cariótipos
foram organizados pela ordem decrescente de tamanho dos cromossomos (Levan et
al.,. 1964).
3. Resultados e Discussão
Os resultados obtidos com a análise genômica e cromossômica comparativa
de ciclídeos amazônicos e a discussão destes dados estão apresentados na forma
de dois artigos científicos, mencionado a seguir:
Artigo 1 – Composição e Representatividade de Elementos Repetitivos no
Genoma de Ciclídeos Neotropicais.
Artigo 2 – Composição, Diversidade e Mapeamento Cromossômico de
Elementos Transponíveis no Genoma de Ciclídeos Neotropicais.

37
3.1 Artigo 1: Composição e Representatividade de Elementos Repetitivos no
Genoma de Ciclídeos Neotropicais
Edika Sabrina Girão Mitozo Tavares, Érica Ramos, Maria Claudia Gross, Cesar Martins,
Carlos Henrique Schneider
3.1.1 Introdução
A porção codificante do genoma de eucariotos corresponde a uma pequena
parte em relação às regiões intergênicas e repetitivas. As regiões repetitivas do
genoma podem ser classificadas de acordo com o número de unidades de repetição
e o comprimento dessas unidades. Os dois maiores grupos de sequências
repetitivas em eucariotos são repetições in tandem e dispersas (Charlesworth et al.,
1994). Repetições in tandem são sequências curtas de DNA não codificantes que se
repetem uma após outra por várias vezes e incluem os DNAs satélites (altamente
repetitivas), mini e microssatélites, DNAs ribossomais e repetições simples
(moderadamente repetitivas) (Charlesworth et al., 1994; Martins, 2007; Padeken et
al., 2015).
As sequências repetitivas dispersas correspondem aos elementos
transponíveis, que se distinguem das outras repetições pela habilidade de se mover
no genoma, independente do ciclo celular (McClintock, 1950; Charlesworth et al.,
1994; Feschotte, 2008; Kazazian et al., 2004). Essa mobilidade pode conferir
vantagens evolutivas ou ser deletéria ao hospedeiro (Kazazian et al., 2004; Kidwell e
Lisch, 2000). Os elementos transponíveis tem o potencial de conduzir a evolução do
genoma onde estão inseridos e influenciam diretamente o tamanho e estrutura do
genoma (Kazazian, 2004; Imai et al., 2007; Metcalfe et al., 2012), conferindo
estabilidade à molécula de DNA (Chalopin et al., 2015), participando dos
mecanismos de regulação gênica (Feschotte, 2008; Indjeian et al., 2016),
determinação sexual (Faber-Hammond et al., 2015), diversidade, organização e
evolução genômica (Kazazian, 2004; Warren et al., 2015) e cariotípica (Schneider et
al., 2013b).
Os elementos transponíveis são classificados quanto à sua estrutura,
tamanho e mecanismo de transposição (Kapitonov e Jurka, 2008; Wicker et al.,
2007). Todos os elementos transponíveis descritos para eucariotos estão unificados
em um banco de dados exclusivo de sequências repetitivas, o Repbase Update

38
(Jurka et al., 2005; Bao et al., 2015). Nele, os ETs são organizados em dois tipos: 1)
Transposons, cuja transposição é realizada pela excisão desse elemento do genoma
e inserção do mesmo em outra região, permanecendo assim com o mesmo número
de cópias, 2) Retrotransposons, no qual uma transcriptase reversa converte uma
cópia de RNA do retrotransposon em cDNA e a insere em outra porção do genoma,
aumentando o número de cópias desse elemento (Bao et al., 2015; Finnegan, 1989;
Kazazian, 2004; Kapitonov e Jurka, 2008; Markova e Mason-Gamer, 2015; Warren
et al., 2015).
Os transposons são divididos em três subclasses, de acordo com seu
mecanismos de transposição e similaridade estrutural: i) Cut-and-paste, na qual
ambas as fitas de DNA são clivadas e constituem a ordem mais abundante dentre os
transposons, os TIRs (Kapitonov e Jurka, 2008). Nela estão inseridas as
superfamílias Mariner/Tc1, hAT, MuDR, EnSpm/CACTA, PiggyBac, P, Merlin,
Harbinger, Transib, Novosib, Kolobok, ISL2EU e os descritos somente no Repbase
Update Mirage, Rehavkus e Chapaev (Kapitonov e Jurka, 2008); ii) Rolling-circle que
inclui os Helitrons e iii) Self-synthesizing que inclui os Polintons/Maverick (Bao et al.,
2015; Kapitonov e Jurka, 2008).
Os retrotransposon são divididos em duas subclasses, são elas: i) LTR (long
terminal repeat) composta pelas superfamílias Copia, Gypsy, BEL, DIRS e retrovírus
endógenos (ERV1, ERV2, ERV3) e ii) não-LTR composta pelas ordens LINE (long
interspersed nuclear elements), SINEs (short interspersed nuclear elements) e
Penelope. A ordem LINE é a mais variável dentre os retrotransponíveis e inclui as
superfamílias CRE, NeSL, R4, R2, L1, RTE, Jockey, CR1, Rex1, I, RandI (Dualen) e
Tx1 (Kapitonov e Jurka, 2008).
Os ETs podem mobilizar-se produzindo sua própria maquinaria de
transposição (considerados autônomos) ou utilizando a maquinaria de elementos
autônomos ou do hospedeiro (Capy, 1998; Friedli e Trono, 2015). A transposição
pode ocorrer tanto entre cromossomos homólogos e não-homólogos, quanto entre
indivíduos e, até mesmo, entre espécies (Capy et al., 1994; Schaack et al., 2010).
Todos os tipos de ETs são amplamente difundidos nos vertebrados (Warren
et al., 2015). Em uma comparação geral, os peixes teleósteos são os que
apresentam a maior diversidade de ETs entre os vertebrados (Chalopin et al., 2015).
Dentre as espécies desse grupo, destacam-se os ciclídeos que caracterizam um

39
grupo de enorme diversidade morfológica, ecológica e comportamental, que os torna
extremamente atraentes para o estudo de questões biológicas, particularmente
aquelas relacionadas à evolução genômica (Lowe-McConnell, 1991; López-
Fernández et al., 2005; Turner, 2007).
A família Cichlidae é dividida em 4 subfamílias: Etroplinae, Ptychochrominae,
Pseudocrenilabrinae e Cichlinae, sendo que as duas últimas correspondem aos
ciclídeos africanos e neotropicais, respectivamente (Sparks e Smith, 2004; Smith et
al., 2008). A partir de análises moleculares combinadas às características
morfológicas, ciclídeos africanos e neotropicais foram considerados clados
monofiléticos e irmãos (Smith et al., 2008). Embora os ciclídeos africanos sejam alvo
de muitos estudos em função da sua ampla gama de espécies, vale ressaltar que os
ciclídeos neotropicais apresentam taxas aceleradas de evolução molecular que
condizem com eventos de radiação adaptativa e contemplam mais de 350 espécies
somente na América do Sul (López-Fernández et al., 2010; Smith et al., 2008;
Turner, 2007). Desta forma, eles representam um grupo de estudo muito
interessante do ponto de vista evolutivo, sendo necessárias investigações a cerca de
sua composição genômica associadas às adaptações do grupo.
Estudos relacionados à composição genômica de ciclídeos neotropicais
caracterizam elementos transponíveis específicos, como Rex1, Rex3, Rex6 e Tc1
(Schneider et al., 2013b; Teixeira et al., 2009) e sítios ribossomais (Gross et al.,
2009b; Schneider et al., 2013a). A abordagem mais abrangente foi a identificação
geral da composição genômica da porção repetitiva em Cichla vazolleri e
Symphysodon discus obtida pela cinética de reassociação do DNA. Foram
identificados satélites, microssatélites, mais de 4 superfamílias de retrotransposons
e representante das superfamílias de transposons (Schneider et al., 2013a).
Entretanto, técnicas atuais permitem a identificação mais precisa e a detecção
da representatividade e uma diversidade muito maior de elementos que constituem
esses genomas. Novas tecnologias de Sequenciamento de Nova Geração (Next
Generation Sequence - NGS) geram dados em larga escala em um período de
tempo reduzido à baixo custo (Novak et al., 2010; Wicker et al., 2007). Contudo, a
abundância de sequências repetitivas ainda é um dos grandes desafios que
dificultam a anotação correta e completa desses genomas (Treangen e Salzberg,
2012; Wicker et al., 2007). Nesse sentido, ferramentas como o Repeat Explorer

40
(Novak et al., 2013), voltadas especificamente para os componentes repetitivos do
genoma, fornecem informações necessárias como a identificação, caracterização,
quantificação, representatividade e proporção de elementos repetitivos no genoma,
possíveis domínios proteicos, descrição de novos elementos, layout do grau de
similaridade entre as sequências (Novak et al., 2010).
Tendo em vista os processos evolutivos aos quais estão sujeitas as espécies
de ciclídeos neotropicais e a importância de ETs na evolução e na dinâmica dos
genomas de eucariotos, o presente estudo identificou e caracterizou os elementos
da fração repetitiva do genoma de quatro espécies de ciclídeos neotropicais
utilizando dados de Sequenciamento de Nova Geração.
3.1.2 Material e Métodos
Quatro espécies de ciclídeos neotropicais da Amazônia Central foram
analisados, sendo elas basais Cichla vazzoleri (3 machos e 3 fêmeas) e Astronotus
ocellatus (3 machos e 5 fêmeas), e derivadas Pterophyllum scalare (3 machos e 3
fêmeas) e Symphysodon discus (2 machos e 2 fêmeas). As espécies foram coletas
com autorização do ICMBio (ICMBio SISBIO 22984-1/2011).
Exemplares de Cichla vazzoleri foram coletados nos meses de março e abril
de 2011 no lago da Usina Hidroelétrica de Balbina (Presidente Figueiredo-AM),
bacia do rio Uatumã̃ afluente do rio Amazonas (3°09’57”S, 59°54’44”W).
Espécimes de Symphysodon discus foram coletados em igarapés próximos a
Barcelos AM (bacia do rio Negro) no período de abril de 2010 a março de 2011
(0°52'2.29"S, 62°48'35.61"W).
Os indivíduos de Astronotus ocellatus foram coletados no mês de abril de
2010 e os indivíduos de Pterophyllum scalare foram coletados no mês de fevereiro
de 2010. Ambas as espécies, A. ocellatus e P. scalare, foram coletadas no lago
Catalão, região de confluência entre os rios Negro e Solimões (3°09'45,53''S e
59°55'05,6''W).
3.1.2.1 Extração de DNA e Sequenciamento de Nova Geração
Os espécimes foram sacrificados pela imersão em água gelada e tecidos
muscular e hepático foram recolhidos para extração de DNA genômico a ser
utilizado no sequenciamento genômico. O DNA total foi extraído usando fenol-

41
clorofórmio (Sambrook et al., 1989) e quantificado pelo espectrofotometro NanoDrop
2000 (Thermo Scientific).
Reações de sequenciamento foram realizadas para 2 indivíduos (macho e
fêmea) de cada espécie (A. ocellatus, C. vazzoleri, P. scalare e S. discus). As
amostras de DNA de uma fêmea e um macho de cada espécie foram fragmentadas
em tamanhos com cerca de 500 pb usando um ultrasonicador S220 (Covaris Inc.,
Woburn, MA, USA). As bibliotecas genômicas foram construídas pelo Sequencing
Facility da Universidade de Maryland, USA (http://www.ibbr.umd.edu/facilities)
utilizando o TruSeq DNA sample preparation kit ver.2 rev. C (Illumina Inc., San
Diego, California, USA). Os sequenciamentos genômicos foram realizados utilizando
a plataforma HiSeq 1000 Illumina, sendo dois lanes por espécie em uma abordagem
paired-end de acordo com as recomendações do fabricante e geraram 8 conjuntos
de reads genômicos.
O output foi composto por reads paired-end com tamanho médio aproximado
de 500 pares de base (pb) e a cobertura de cada genoma foi de ~32 a 40 vezes o
genoma de cada sexo e espécie. Todas as sequências foram submetidas à
avaliação de qualidade e de presença de adaptadores utilizando o software
FASTQC (website:http://www.bioinformatics.babraham.ac.uk/projects/fastqc/) a partir
dos valores de Phred quality score obtidos para cada base nucleotídica durante o
sequenciamento. Foram eliminados reads que não apresentassem 90% de seus
nucleotídeos com qualidade igual ou superior a 30 (Phred quality score).
3.1.2.2 Identificação de elementos transponíveis
Após as filtragens iniciais e o preparo das bibliotecas para a análise de
clusterização, cerca de 700.000 reads (correspondente a 0,1x de cobertura do
genoma completo, cujo tamanho estimado é de 1Gb) de cada sexo e espécie foram
selecionados para identificar a composição da fração repetitiva dos genomas destas
espécies. Os reads desta amostragem de sequências de cópias repetitivas foram
filtrados, agrupados em clusters e identificados e caracterizados utilizando a
ferramenta Repeat Explorer - website http://www.repeatexplorer.org/ - (Novak et al.,
2013). Neste pipeline os reads de elementos repetitivos são agrupados por
identidade em uma comparação all-to-all formando clusters. Para que um read seja
incorporado ao cluster, ele precisa ter 55% de cobertura e 90% de identidade em

42
relação a outro read. Desta forma, cada clusters representa um grupo de sequências
mais similares entre si do que em relação às sequências de outro cluster. Os
clusters foram anotados utilizando o algoritmo Repeat Masker em conjunto com o
banco de dados Repbase Update (Jurka et al., 2005). A caracterização de cada
cluster contém informações específicas e revela a identidade do elemento
transponível de acordo com o Repbase Update e o número de hits, número de reads
total e discriminado por sexo e espécie, além de gráficos, domínios de proteínas e
outras informações. Os elementos foram identificados ao nível de superfamília
(Kapitonov e Jurka, 2008), portanto a ocorrência de mais de um cluster identificado
como o mesmo elemento, não significa que eles sejam iguais em níveis mais
específicos. A análise estatística é feita pela ferramenta SeqGrapheR, que distingue
tipos básicos de repetições e investiga a variabilidade de sequências dentro das
famílias de elementos repetitivos (Novak et al., 2010).
Com o intuito de estudar elementos repetitivos de dinâmicas diferentes entre
as espécies e entre os indivíduos de sexos diferentes, foram selecionados para as
próximas etapas os clusters com diferenças nos números de reads de cada espécie
(utilizados na montagem do mesmo elemento). Os clusters selecionados deveriam
apresentar diferença de no mínimo 500 reads: i) entre as espécies; ii) entre os sexos
da mesma espécie; iii) os altamente representativos em uma única espécie.
3.1.3 Resultados
O número de reads totais gerados pelo Sequenciamento de Nova Geração, a
cobertura dos genomas, o número de reads total utilizados como amostragem para
identificação de elementos transponíveis e de o número dos que foram agrupados
em clusters ou não podem ser observados na Tabela 1. A quantidade de reads
agrupados em clusters variou de 51,3% a 63% dessa amostragem. Nem todas as
sequências foram consideradas na composição dos clusters ou por se tratarem de
sequências de cópia única ou por serem sequências repetitivas que não
apresentaram similaridade com outras sequências e, portanto, não foram agrupados
em nenhum cluster e variaram de 37% a 48,7% (Tabela 1).

43
Tabela 1: Dados do sequenciamento genômico total e da amostragem utilizada na
investigação da fração repetitiva de quatro espécies de ciclídeos neotropicais
amazônicos. (CV) Cichla vazolleri, (AO) Astronotus ocellatus, (PS) Pterophyllum
scalare, (SD) Symphysodon discus, (F) fêmeas e (M) machos.
Ao total, 30 clusters dos 69 gerados foram utilizados nas etapas
subsequentes. Os selecionados pelo critério 1 foram os clusters 06, 07, 08, 10, 11,
15, 16, 21, 22, 23, 27, 28, 33, 35, 38; pelo critério 2 foram 02, 05, 06, 08, 11, 51, 63;
e pelo critério 3 foram 01, 02, 03, 05, 09, 13, 24, 41, 44, 48, 54, 61, 63, 66 (Figura 1).
Alguns clusters atendem a dois critérios, como exemplo dos clusters 02, 05, 06, 08,
11, 63. Os elementos repetitivos identificados foram: as superfamílias DONG.R4, L2,
RTE, PENELOPE e REX.BABAR da ordem LINE da classe dos retrotransposons
não-LTR, superfamília ERV1, da classe dos retrotransposons LTRs, superfamília
Tc1 da classe dos transposons, repetições simples e DNA satélites (Figura 2).
Grupo
N° total de
reads obtidos
no SNG
Cobertura
do
genoma
Amostragem
utilizada
(reads)
Reads agrupados
em clusters Singlets
CV - F 231.369.922 ~34x 700.000 359.229 (51.3%) 340.771 (48.7%)
CV - M 227.201.454 ~34x 700.000 372.638 (53.2%) 327.362 (46.8%)
AO - F 267.797.946 ~40x 700.000 378.617 (54.1%) 321.383 (45.9%)
AO - M 226.726.662 ~34x 700.000 392.116 (56%) 307.884 (44%)
PS - F 248.087.060 ~37x 700.000 397.237 (56.7%) 302.763 (43.3%)
PS - M 240.672.242 ~36x 700.000 427.087 (61%) 272.913 (39%)
SD - F 216.066.104 ~32x 700.000 433.776 (62%) 266.224 (38%)
SD - M 251.236.834 ~37x 700.000 441.146 (63%) 258.854 (37%)
Total 1.909.158.224 5.600.000 3.201.846 2.405.154
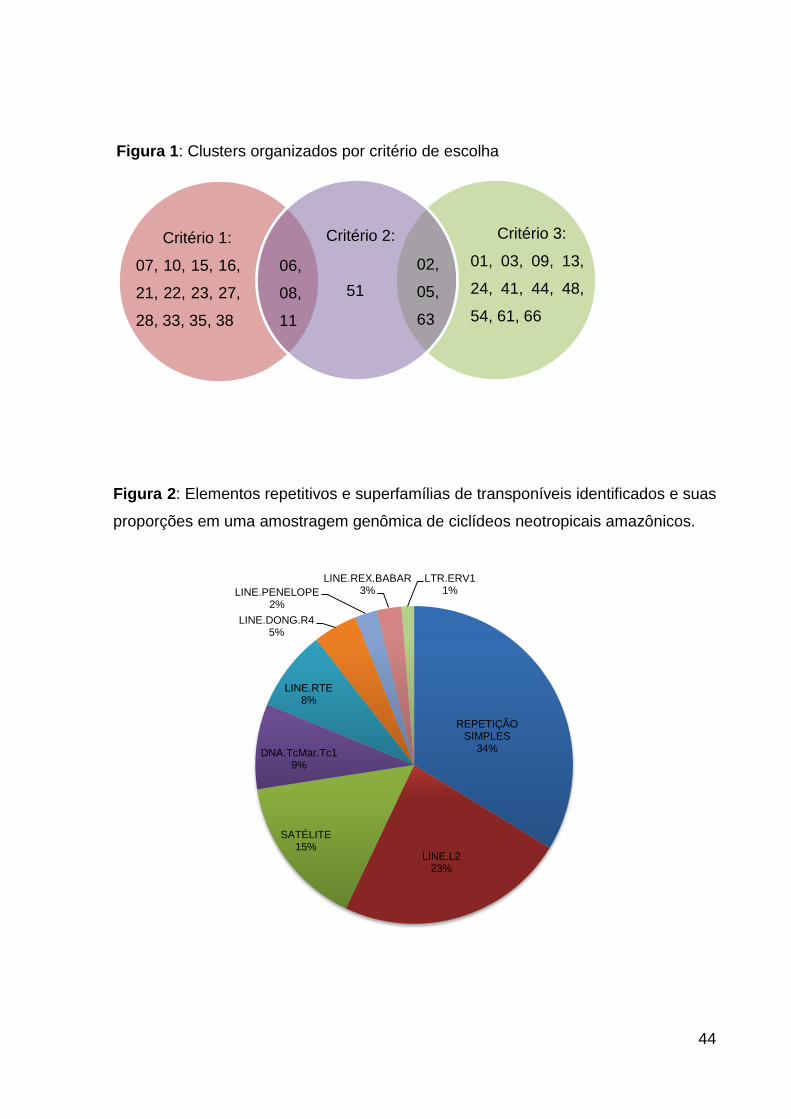
44
Figura 2: Elementos repetitivos e superfamílias de transponíveis identificados e suas
proporções em uma amostragem genômica de ciclídeos neotropicais amazônicos.
REPETIÇÃO SIMPLES
34%
LINE.L2 23%
SATÉLITE 15%
DNA.TcMar.Tc1 9%
LINE.RTE 8%
LINE.DONG.R4 5%
LINE.PENELOPE 2%
LINE.REX.BABAR 3%
LTR.ERV1 1%
02,
05,
63
06,
08,
11
Critério 1:
07, 10, 15, 16,
21, 22, 23, 27,
28, 33, 35, 38
Critério 2:
51
Critério 3:
01, 03, 09, 13,
24, 41, 44, 48,
54, 61, 66
Figura 1: Clusters organizados por critério de escolha
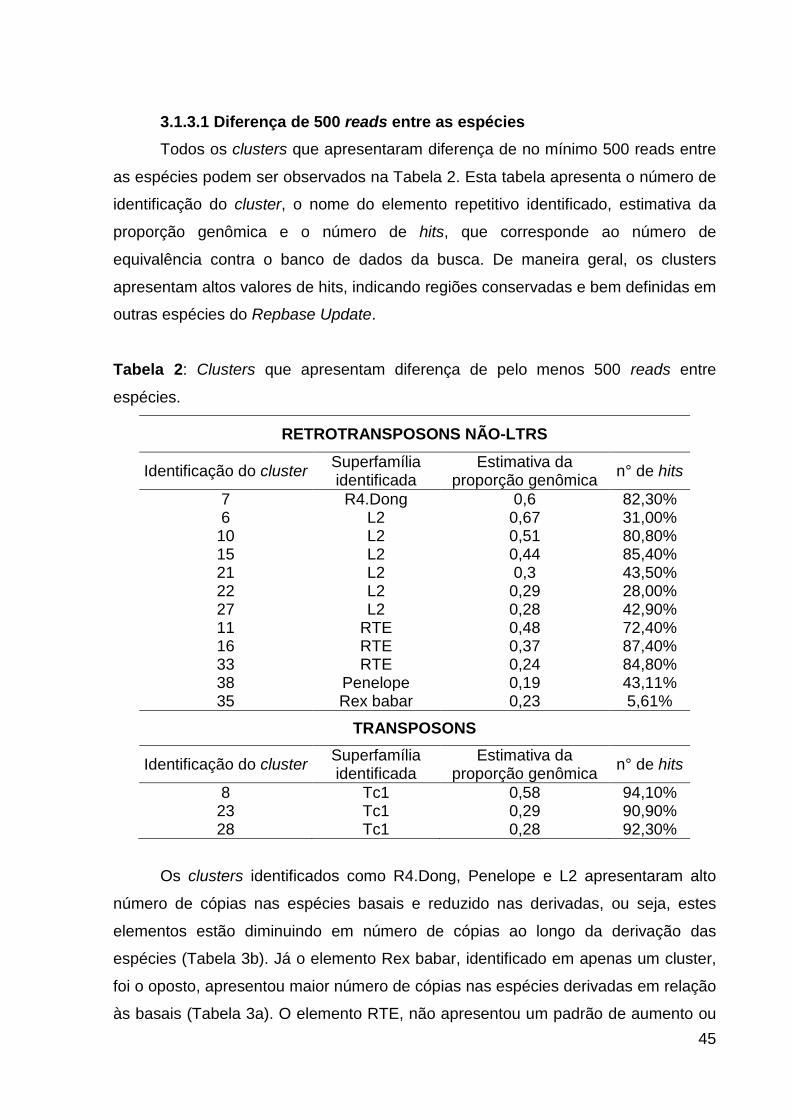
45
3.1.3.1 Diferença de 500 reads entre as espécies
Todos os clusters que apresentaram diferença de no mínimo 500 reads entre
as espécies podem ser observados na Tabela 2. Esta tabela apresenta o número de
identificação do cluster, o nome do elemento repetitivo identificado, estimativa da
proporção genômica e o número de hits, que corresponde ao número de
equivalência contra o banco de dados da busca. De maneira geral, os clusters
apresentam altos valores de hits, indicando regiões conservadas e bem definidas em
outras espécies do Repbase Update.
Tabela 2: Clusters que apresentam diferença de pelo menos 500 reads entre
espécies.
RETROTRANSPOSONS NÃO-LTRS
Identificação do cluster Superfamília identificada
Estimativa da proporção genômica
n° de hits
7 R4.Dong 0,6 82,30% 6 L2 0,67 31,00%
10 L2 0,51 80,80% 15 L2 0,44 85,40% 21 L2 0,3 43,50% 22 L2 0,29 28,00% 27 L2 0,28 42,90% 11 RTE 0,48 72,40% 16 RTE 0,37 87,40% 33 RTE 0,24 84,80% 38 Penelope 0,19 43,11% 35 Rex babar 0,23 5,61%
TRANSPOSONS
Identificação do cluster Superfamília identificada
Estimativa da proporção genômica
n° de hits
8 Tc1 0,58 94,10% 23 Tc1 0,29 90,90% 28 Tc1 0,28 92,30%
Os clusters identificados como R4.Dong, Penelope e L2 apresentaram alto
número de cópias nas espécies basais e reduzido nas derivadas, ou seja, estes
elementos estão diminuindo em número de cópias ao longo da derivação das
espécies (Tabela 3b). Já o elemento Rex babar, identificado em apenas um cluster,
foi o oposto, apresentou maior número de cópias nas espécies derivadas em relação
às basais (Tabela 3a). O elemento RTE, não apresentou um padrão de aumento ou
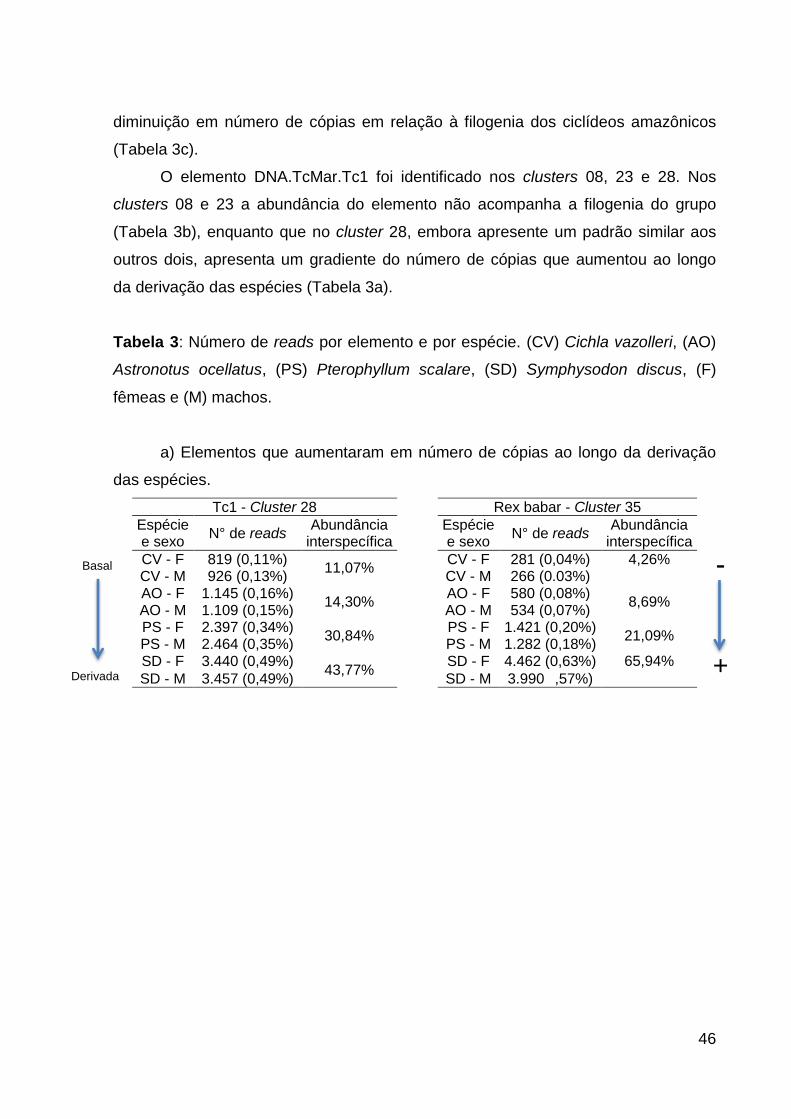
46
diminuição em número de cópias em relação à filogenia dos ciclídeos amazônicos
(Tabela 3c).
O elemento DNA.TcMar.Tc1 foi identificado nos clusters 08, 23 e 28. Nos
clusters 08 e 23 a abundância do elemento não acompanha a filogenia do grupo
(Tabela 3b), enquanto que no cluster 28, embora apresente um padrão similar aos
outros dois, apresenta um gradiente do número de cópias que aumentou ao longo
da derivação das espécies (Tabela 3a).
Tabela 3: Número de reads por elemento e por espécie. (CV) Cichla vazolleri, (AO)
Astronotus ocellatus, (PS) Pterophyllum scalare, (SD) Symphysodon discus, (F)
fêmeas e (M) machos.
a) Elementos que aumentaram em número de cópias ao longo da derivação
das espécies.
Tc1 - Cluster 28
Rex babar - Cluster 35
Espécie e sexo
N° de reads Abundância
interspecífica
Espécie e sexo
N° de reads Abundância
interspecífica
CV - F 819 (0,11%) 11,07%
CV - F 281 (0,04%) 4,26%
CV - M 926 (0,13%)
CV - M 266 (0.03%) AO - F 1.145 (0,16%)
14,30%
AO - F 580 (0,08%) 8,69%
AO - M 1.109 (0,15%)
AO - M 534 (0,07%) PS - F 2.397 (0,34%)
30,84%
PS - F 1.421 (0,20%) 21,09%
PS - M 2.464 (0,35%)
PS - M 1.282 (0,18%) SD - F 3.440 (0,49%)
43,77%
SD - F 4.462 (0,63%) 65,94%
SD - M 3.457 (0,49%)
SD - M 3.990 ,57%)
Basal
Derivada
-
+
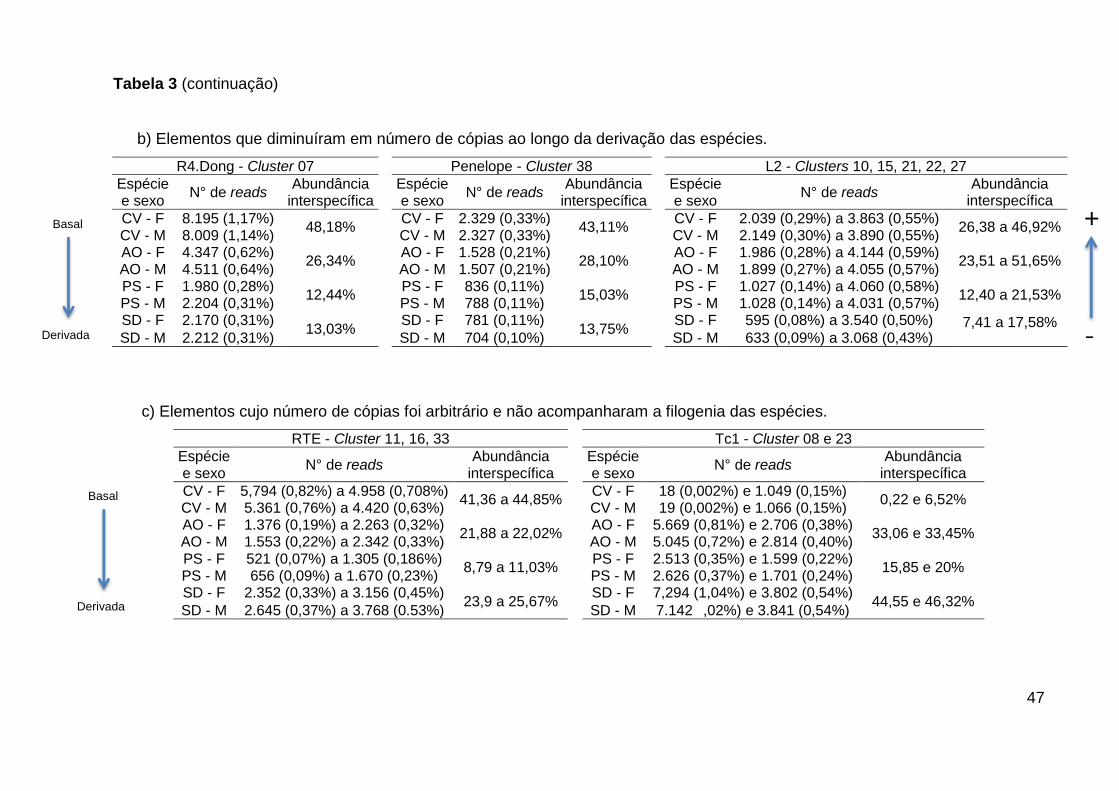
47
Tabela 3 (continuação)
b) Elementos que diminuíram em número de cópias ao longo da derivação das espécies.
R4.Dong - Cluster 07
Penelope - Cluster 38
L2 - Clusters 10, 15, 21, 22, 27
Espécie e sexo
N° de reads Abundância
interspecífica
Espécie e sexo
N° de reads Abundância
interspecífica
Espécie e sexo
N° de reads Abundância
interspecífica
CV - F 8.195 (1,17%) 48,18%
CV - F 2.329 (0,33%)
43,11%
CV - F 2.039 (0,29%) a 3.863 (0,55%) 26,38 a 46,92%
CV - M 8.009 (1,14%)
CV - M 2.327 (0,33%)
CV - M 2.149 (0,30%) a 3.890 (0,55%) AO - F 4.347 (0,62%)
26,34%
AO - F 1.528 (0,21%) 28,10%
AO - F 1.986 (0,28%) a 4.144 (0,59%)
23,51 a 51,65% AO - M 4.511 (0,64%)
AO - M 1.507 (0,21%)
AO - M 1.899 (0,27%) a 4.055 (0,57%)
PS - F 1.980 (0,28%) 12,44%
PS - F 836 (0,11%)
15,03%
PS - F 1.027 (0,14%) a 4.060 (0,58%) 12,40 a 21,53%
PS - M 2.204 (0,31%)
PS - M 788 (0,11%)
PS - M 1.028 (0,14%) a 4.031 (0,57%) SD - F 2.170 (0,31%)
13,03%
SD - F 781 (0,11%) 13,75%
SD - F 595 (0,08%) a 3.540 (0,50%) 7,41 a 17,58%
SD - M 2.212 (0,31%)
SD - M 704 (0,10%)
SD - M 633 (0,09%) a 3.068 (0,43%)
c) Elementos cujo número de cópias foi arbitrário e não acompanharam a filogenia das espécies.
RTE - Cluster 11, 16, 33
Tc1 - Cluster 08 e 23
Espécie e sexo
N° de reads Abundância
interspecífica
Espécie e sexo
N° de reads Abundância
interspecífica
CV - F 5,794 (0,82%) a 4.958 (0,708%) 41,36 a 44,85%
CV - F 18 (0,002%) e 1.049 (0,15%)
0,22 e 6,52% CV - M 5.361 (0,76%) a 4.420 (0,63%)
CV - M 19 (0,002%) e 1.066 (0,15%)
AO - F 1.376 (0,19%) a 2.263 (0,32%) 21,88 a 22,02%
AO - F 5.669 (0,81%) e 2.706 (0,38%)
33,06 e 33,45% AO - M 1.553 (0,22%) a 2.342 (0,33%)
AO - M 5.045 (0,72%) e 2.814 (0,40%)
PS - F 521 (0,07%) a 1.305 (0,186%) 8,79 a 11,03%
PS - F 2.513 (0,35%) e 1.599 (0,22%)
15,85 e 20% PS - M 656 (0,09%) a 1.670 (0,23%)
PS - M 2.626 (0,37%) e 1.701 (0,24%)
SD - F 2.352 (0,33%) a 3.156 (0,45%) 23,9 a 25,67%
SD - F 7,294 (1,04%) e 3.802 (0,54%)
44,55 e 46,32% SD - M 2.645 (0,37%) a 3.768 (0.53%)
SD - M 7.142 ,02%) e 3.841 (0,54%)
Basal
Derivada
+
-
Basal
Derivada

48
3.1.3.2 Diferença de 500 reads entre os sexos da mesma espécie
As espécies que apresentaram diferença no número de reads entre os sexos
foram: i) Astronotus ocellatus nos clusters 08 (Tc1) e 06 (L2), sendo 5.669 e 5.906
reads no genoma de fêmeas e, 5.049 e 6.591 no de machos, respectivamente; e ii)
Symphysodon discus nos clusters 06 (L2 – 3.540 reads de fêmeas e 3.068 reads de
macho), 51 (L2 - 2.894 reads de fêmeas e 2.324 reads de machos), 63 (Penelope -
3.019 reads de fêmeas e 2.483 reads de machos), 11 (RTE - 3.156 reads de fêmeas
e 3.768 reads de machos) e nos clusters 02 e 05, ambos repetições simples com
17.014 reads de fêmeas e 22.476 reads de machos no primeiro e 50.588 reads de
fêmeas e 37.768 reads de machos no segundo (Tabela 4).
Tabela 4: Espécies que apresentaram diferença no número de cópias de elementos
transponíveis e outros repetitivos entre sexos.
Espécie e cluster Sequências repetitivas e
superfamílias identificadas Sexo com maior
n° de reads
Astronotus ocellatus
Cluster 08 Tc1 fêmea Cluster 06 L2 macho
Symphysodon discus
Cluster 06 L2 fêmea Cluster 51 L2 fêmea Cluster 63 Penelope fêmea Cluster 11 RTE macho Cluster 05 Repetição simples macho Cluster 02 Repetição simples fêmea
3.1.3.3 Altamente representativo em uma das espécies
Alguns clusters foram compostos quase exclusivamente por reads de uma
única espécie. Um cluster de LTR.EV1 foi composto majoritariamente por reads de
C. vazzoleri, um de Penelope por reads de S. discus e clusters de L2 e Rex babar
por reads de A. ocellatus. Além destes, diferentes clusters identificados como
repetições simples e DNA satélite foram compostos por reads de uma das quatro
espécies (Tabela 5).
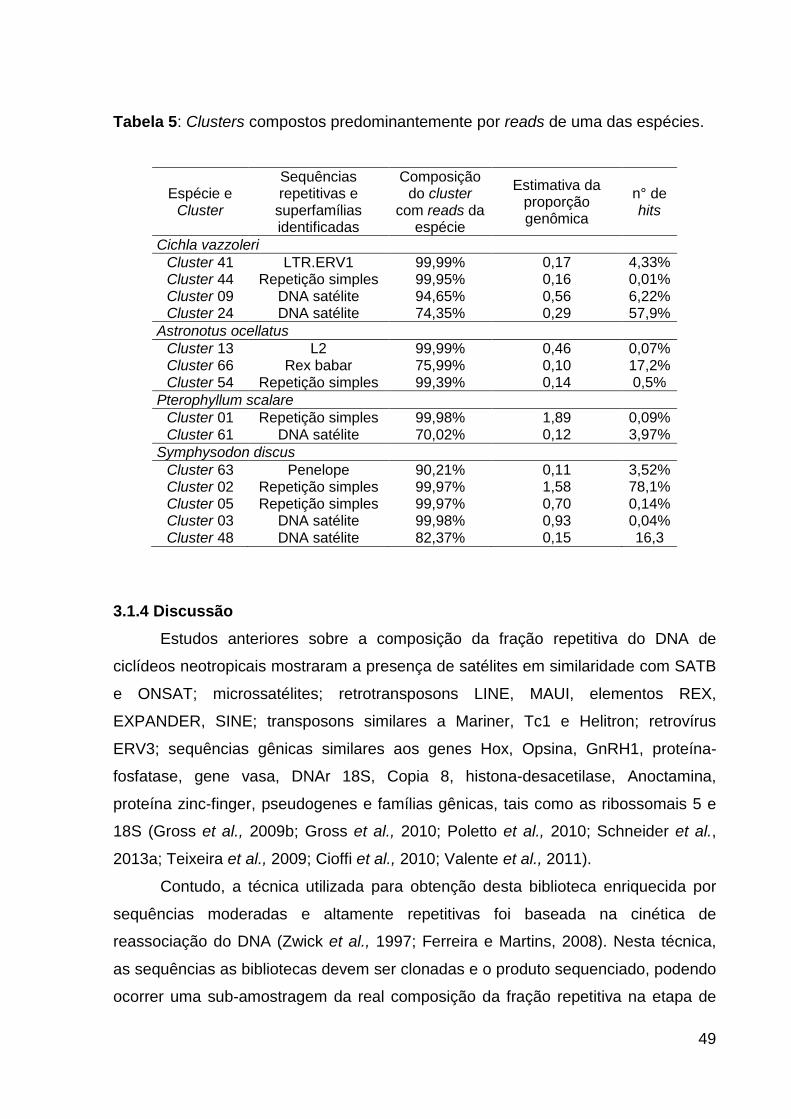
49
Tabela 5: Clusters compostos predominantemente por reads de uma das espécies.
Espécie e Cluster
Sequências repetitivas e superfamílias identificadas
Composição do cluster
com reads da espécie
Estimativa da proporção genômica
n° de hits
Cichla vazzoleri
Cluster 41 LTR.ERV1 99,99% 0,17 4,33% Cluster 44 Repetição simples 99,95% 0,16 0,01% Cluster 09 DNA satélite 94,65% 0,56 6,22% Cluster 24 DNA satélite 74,35% 0,29 57,9%
Astronotus ocellatus
Cluster 13 L2 99,99% 0,46 0,07% Cluster 66 Rex babar 75,99% 0,10 17,2% Cluster 54 Repetição simples 99,39% 0,14 0,5%
Pterophyllum scalare
Cluster 01 Repetição simples 99,98% 1,89 0,09% Cluster 61 DNA satélite 70,02% 0,12 3,97%
Symphysodon discus
Cluster 63 Penelope 90,21% 0,11 3,52% Cluster 02 Repetição simples 99,97% 1,58 78,1% Cluster 05 Repetição simples 99,97% 0,70 0,14% Cluster 03 DNA satélite 99,98% 0,93 0,04% Cluster 48 DNA satélite 82,37% 0,15 16,3
3.1.4 Discussão
Estudos anteriores sobre a composição da fração repetitiva do DNA de
ciclídeos neotropicais mostraram a presença de satélites em similaridade com SATB
e ONSAT; microssatélites; retrotransposons LINE, MAUI, elementos REX,
EXPANDER, SINE; transposons similares a Mariner, Tc1 e Helitron; retrovírus
ERV3; sequências gênicas similares aos genes Hox, Opsina, GnRH1, proteína-
fosfatase, gene vasa, DNAr 18S, Copia 8, histona-desacetilase, Anoctamina,
proteína zinc-finger, pseudogenes e famílias gênicas, tais como as ribossomais 5 e
18S (Gross et al., 2009b; Gross et al., 2010; Poletto et al., 2010; Schneider et al.,
2013a; Teixeira et al., 2009; Cioffi et al., 2010; Valente et al., 2011).
Contudo, a técnica utilizada para obtenção desta biblioteca enriquecida por
sequências moderadas e altamente repetitivas foi baseada na cinética de
reassociação do DNA (Zwick et al., 1997; Ferreira e Martins, 2008). Nesta técnica,
as sequências as bibliotecas devem ser clonadas e o produto sequenciado, podendo
ocorrer uma sub-amostragem da real composição da fração repetitiva na etapa de

50
clonagem. Os resultados do presente estudo não apresentam este tipo de viés
técnico, uma vez que os dados utilizados foram gerados diretamente pela
amostragem de dados do sequenciamento de nova geração (SNG), sendo uma
abordagem mais robusta.
Nas quatro espécies de ciclídeos analisadas foram encontrados LINEs
pertencentes a 5 superfamílias: L2, RTE, Penelope, Rex.babar e Dong.R4. Com
relação aos transposons, somente um foi identificado, o Tc1. Ainda, foram
identificados Retrotransposons LTR do tipo ERV1, DNA satélites e outras repetições
simples.
Todos os elementos identificados no genoma de ciclídeos neotropicais foram
encontrados em ciclídeos africanos. Entretanto, uma gama mais diversificada foi
evidenciada nos ciclídeos africanos, tais como: SINEs, L1, Rex1, Crack, Daphne,
Nimb, Hero, Gypsy, Pao, Copia, DIRs, hAT, Harbinger, piggyBac, dada, Polinton,
Helitron (Brawand et al., 2014). Provavelmente isto está relacionado à dois possíveis
viés deste trabalho: o de i) sub-amostragem, que, embora utilize diretamente dados
do SNG, analisou apenas uma pequena fração do genoma total das espécies,
refletindo uma resultado inicial da porção repetitiva; e o de ii) sub-agrupamento, pois
os elementos repetitivos ou superfamílias de transponíveis identificados nos clusters
são abrangentes e correspondem ao maior número de sequências envolvidas na
formação daquele cluster. Para minimizar esse viés, seria necessário fazer
reagrupamentos subsequentes (reclustering), a fim de favorecer identificações mais
específicas dos mesmos. Desta forma, é possível que a diversidade de ETs em
ciclídeos neotropicais seja tão vasta quanto a encontrada em seu grupo irmão
africano em uma abordagem mais abrangente e refinada com identificações mais
específicas (Smith et al., 2008; Warren et al., 2015).
3.1.4.1 Diferença na composição genômica entre as espécies
3.1.4.1.1 Retrotransposons não LTR
O elemento LINE foi o ET mais representativo encontrado no genoma de
ciclídeos neotropicais, correspondendo a mais de 40% dos ETs encontrados. Isso
confronta os principais relatos na literatura, pois os ETs mais abundantemente
distribuídos nos genomas de peixes teleósteos são os transposons (Chalopin et al.,
2015). Em ciclídeos africanos, os LINEs são apenas o segundo elemento mais

51
abundante. O primeiro é o transposon Tc1 que correspondem a mais de 60% do
genoma (Brawand et al., 2014).
Nos ciclídeos neotropicais, os elementos DONG.R4 (cluster 07), L2 (clusters
06, 10, 15, 21, 22 e 27) e PENELOPE (cluster 38) apresentaram maior
representatividade nas espécies basais do que nas derivadas. LINE.REX.BABAR foi
o único que apresentou menor representatividade nas espécies basais do que nas
derivadas, possivelmente indicando um acúmulo deste elemento ao longo da
evolução destas espécies.
Quando retrotransposons estão ativos no genoma, o número de cópias tende
a aumentar ao longo da derivação das espécies hospedeiras em virtude do
mecanismo de retrotransposição que apresentam, como observado no elemento
LINE.REX.BABAR. Ainda, eventos de duplicação ou crossing over desigual podem
ter ocorrido, contribuindo com o aumentando de cópias deste ET. Se esse elemento
não interfere na dinâmica celular ou acaba favorecendo o hospedeiro, ele é mantido
no genoma e pode apresentar alto grau de polimorfismo.
No entanto, se os elementos adquirem função, se tornam ativos, ou alocam-
se próximos a gene, eles ficam sob um processo restritivo, que é o que
provavelmente acontece com DONG.R4, L2 e PENELOPE. Isso significa que eles
estão sob alta pressão seletiva, pois qualquer mudança mais acentuada em suas
sequências, pode interferir significativamente na expressão de genes adjacentes
(Feschotte, 2008; Kidwell e Lisch, 2000; Warren et al., 2015). Para isso, ocorre o
processo de seleção purificadora, que elimina esses elementos do genoma do
hospedeiro a fim de prevenir maiores danos, tais como sugerido para plantas
(Hollister et al., 2011), invertebrados como para Drosophila (Casola et al., 2007), e
mamíferos (Lowe et al., 2007; Kamal et al., 2006).
LINE.PENELOPE, LINE 1 e 2, Rex1/BABAR aparentemente estão presente
em todos os vertebrados, mas apresentam distribuição desigual entre eles, enquanto
que DONG.R4 e Rex6 estão em quase todos os peixes Actinopterygii e ausentes
Sarcopterygii, com exceção de celacanto (Chalopin et al., 2015). O movimento e o
acúmulo de ETs exercem grande influência na trajetória evolutiva do hospedeiro e
parece evidente que TEs são ferramentas regulatórias do genoma, com papel
importante no direcionamento da expressão gênica (Brookfield, 2005; Slotkin e
Martienssen, 2007).

52
3.1.4.1.2 Transposons
Os elementos Tc-Mariner e hAT são superfamílias mais representativas de
transposons e estão presentes em todos os eucariotos de forma heterogênea
(Brunet et al., 1994; Wallau et al., 2014). O elemento Tc1 foi o segundo elemento
transponível mais representativo no genoma de ciclídeos neotropicais,
correspondendo a 9% dos ETs encontrados e o transposon mais abundante. Sua
representatividade foi oscilante em ciclídeos neotropicais, sendo maior em A.
ocellatus e S. discus e menor em C. vazolleri e P. scalare. A ocorrência descontínua
e não concatenada com a filogenia dos ciclídeos pode ser resultado de i)
polimorfismo ancestral ii) diferentes taxas de evolução dos ETs, incluindo perdas
estocásticas de cópias de ETs e iii) eventos de transferência horizontal ao longo da
evolução da espécie hospedeira.
Inconsistências entre a filogenia de transposons e espécies hospedeiras já
foram relatadas em estudos evolutivos (Arkhipova e Meselson, 2000; Capy et al.,
1994; Lohe et al., 1995; Yamada et al., 2015), sendo também explicadas por essas
hipóteses, tais como em plantas para o transposon Pong do tipo Harbinger (Markova
e Mason-Gamer, 2015), em Drosophila melanogaster (Dias e Carareto, 2012), entre
crustáceos e insetos (Dupeyron et al., 2014), entre mamíferos e insetos (Oliveira et
al., 2012), em Cichla kelberi (Teixeira et al., 2009), entre peixes teleósteos e
lampreias, os quais apresentam comportamento parasita-hospedeiro (Kuraku et al.,
2012). Existem evidências de elementos transponíveis procarióticos em eucariotos
(Gilbert e Cordaux, 2013). Esses eventos já foram sugeridos para outros ETs como
os elementos da família Rex em espécies ciclídeos neotropicais filogeneticamente
distantes (Schneider et al., 2013b).
3.1.4.2 Diferença na composicão genômica entre os sexos da mesma
espécie
A variabilidade em abundância e diversidade de ETs entre os organismos é
fortemente influenciada pelo seu modo de reprodução (Wright e Finnegan, 2001).
Linhagens assexuadas, mesmo as derivadas de linhagens sexuadas, tendem a
apresentar baixas ou nulas probabilidades de propagação e reduzida abundância de
ETs no genoma (Wright e Finnegan, 2001; Arkhipova e Meselson, 2000).

53
Além do modo de reprodução, diferenças na composição genômica entre os
sexos da mesma espécie também estão relacionados à determinação sexual,
especialmente pela composição diferenciada dos cromossomos sexuais
(Charlesworth, 2012). ETs geralmente estão ativos em células germinativas e
promovem rearranjos cromossômicos em diferentes populações, conduzindo ao
isolamento reprodutivo e especiação (Volff, 2005). A mobilidade de ET em
salmonídeos, por exemplo, está associada às regiões de determinação sexual em
machos (Faber-Hammond et al., 2015). Em mamíferos, a presença do elemento
LINE L1 leva à condensação de um dos cromossomos X de fêmeas (Bailey et al.,
2000), que resulta na compensação de dosagem hormonal nas fêmeas, uma vez
que possuem o dobro da informação genética em relação aos machos. Ainda em
mamíferos, uma família de retrogenes presentes no cromossomo Y está envolvida
na fertilidade masculina e é expressa exclusivamente nos testículos (Volff, 2005).
3.1.4.2.1 Transposons e Retrotransposons não LTR
A espécie Astronotus ocellatus foi a única espécie que apresentou diferença
no número de reads entre machos e fêmeas para o transposon Tc1.Mariner, sendo
maior em fêmeas do que em machos. Por outro lado, a maior representatividade do
retrotransposons não-LTR LINE.L2 para essa espécie foi em machos. A espécie
Symphysodon discus apresentou diferença no número de reads dos elementos L2,
Penelope e RTE entre sexos, sendo L2 e Penelope superior em fêmeas e RTE em
machos.
Em salmonídeos, o elemento Tc1 e os retroelementos não LTR são
candidatos responsáveis pela determinação sexual (Yano et al., 2012). Pois
sequências deste transposon, bem como domínios de endo e exonucleases e
transcriptase reversa típicos do LINE-1 foram encontradas entre éxons de genes de
determinação sexual (Faber-Hammond et al,. 2015). As transposases responsáveis
pela transposição do Tc1 estão mobilizando, além do ET situado nas regiões
flanqueadoras, grandes porções das regiões de determinação sexual. Transposons
e elementos do tipo Rex foram detectados duplicando regiões de regulação de
genes de determinação sexual em medaka, Oryzias latipes (Herpin et al., 2010).
Pode ser que haja uma relação como essa em Astronotus ocellatus, portanto, é
importante que uma investigação acerca da constituição do transposon Tc1.Mariner

54
seja realizada a fim de detectar a presença ou ausência de sequências de
transposases inseridas em genes de determinação de sexo nesta espécie.
3.1.4.2.2 Repetições simples
Diferenças no número de repetições simples entre machos e fêmeas foram
observadas somente em Symphysodon discus. Vários podem ser os motivos que
justifiquem essa característica e um deles pode estar atrelado à determinação
sexual ou aos mecanismos de proteção das regiões de determinação sexual.
Repetições simples tendem a se acumular em regiões de recombinação desigual de
cromossomos sexuais e isso, de certa forma, previne que regiões codificadoras de
produto funcional sejam perdidas ou alteradas (Charlesworth, 2012).
3.1.4.3 Altamente Representativo em uma das Espécies
O genoma de peixes teleósteos é constituído em grande parte por elementos
transponíveis, enquanto que o genoma de mamíferos e aves não, sugerindo que a
evolução de cada linhagem é conduzida por mecanismos evolutivos diferentes
(Chalopin et al., 2015; Warren et al., 2015).
3.1.4.3.1 Retrotransposons não LTR
O elemento LINE.L2 foi altamente representativo em Astronotus ocellatus,
apresentando um único read em Cichla vazolleri. O elemento L1, semelhante ao L2,
apresenta-se ativo no genoma de mamíferos, podendo mobilizar-se e promover
doenças (Kazazian, 2004)
O elemento LINE.Rex.Babar também foi altamente representativo em A.
ocellatus, embora as outras espécies tenham apresentado cópias desse elemento
em menor proporção, diferente do ocorrido para o LINE.L2. Elementos do tipo Rex3
e AoLINE semelhante ao CR1 (Mazzuchelli e Martins, 2009) já foram identificados
em Astronotus ocellatus previamente.
3.1.4.3.2 Retrotransposos LTR
O elemento LTR.ERV1 foi altamente representativo em Cichla vazolleri. Esse
elemento é o mais ativo em ciclídeos africanos e corresponde a metade dos LTRs
no genoma (Brawand et al., 2014). Embora ele seja altamente representativo em C.

55
vazolleri, provavelmente esse elemento surgiu em um ancestral comum e foi
eliminado nas outras espécies. Isso porque sua representatividade nas outras
espécies foi baixa.
Além do ERV1, Cichla vazolleri também apresentou repetição simples e
satélite espécie-específicos; LINE.L2, LINE.REX.BABAR e repetições simples foram
específicas de Astronotus ocellatus; Pterophyllum scalare apresentou uma única
repetição simples espécie-específico; Em contraste a Symphysodon discus que
apresentou LINE.PENELOPE, LINE.L2, repetições simples e satélites.
Desta forma, nossos dados indicam que os genomas de ciclídeos neotropicais
são constituídos por repetições em tandem e diferentes superfamílias de elementos
transponíveis que se distribuem de forma desigual entre os genomas e fornecem
suporte para melhor compreensão da composição e organização genômica.

56
3.2 Artigo 2: Composição, Diversidade e Mapeamento Cromossômico de
Elementos Transponíveis no Genoma de Ciclídeos Neotropicais
Edika Sabrina Girão Mitozo Tavares, Natália Dayane Moura Carvalho, Edson Junior do Carmo,
Marcos Andrei de Almeida Barbosa, Maria Claudia Gross, Cesar Martins, Carlos Henrique Schneider
3.2.1 Introdução
O genoma dos vertebrados é composto majoritariamente por sequências
repetitivas. Elementos transponíveis (ETs) são sequências repetitivas de DNA
capazes de mobilizar-se dentro e entre os genomas (Fedoroff, 2012; Kapitonov e
Jurka, 2008; Warren et al., 2015; Wicker et al., 2007). Vários trabalhos evidenciam a
participação direta desses ETs na estabilidade, tamanho e estrutura do genoma
(Bennetzen e Wang, 2014; Imai et al., 2007), mudanças fenotípicas (Lisch, 2013;
Piacentini et al., 2014), diversidade e evolução genômica (Schneider et al., 2013b;
Bourque, 2009), determinação sexual (Faber-Hammond et al., 2015) e processos de
especiação (Bonchev e Parisod, 2013). Ainda, a transposição promove diversidade e
aumento no número de cópias, mudanças de posição de importantes sequências de
DNA e consequente alteração da expressão gênica (Indjeian et al., 2016; Kazazian,
2004; Rebello et al., 2012). Dessa forma, é clara a participação de elementos
transponíveis nos processos evolutivos que conduzem a evolução do hospedeiro
(Kazazian, 2004; Schneider et al., 2013b; Bourque, 2009; Kidwell e Lisch, 2000;
Warren et al., 2015).
A transposição de um ET não é aleatória e está sujeita aos mecanismos de
defesa do genoma onde estão inseridos, como RNAs de interferência (RNAi),
atividades epigenéticas, repressão transcricional e pós-transcricional (Bourc’his &
Bestor, 2004; Obbard et al., 2009; Kidwell e Lisch, 2001; Lippman et al., 2004;
Cowley & Oakey, 2013). A fim de evitar sua eliminação, os ETs também
desenvolveram estratégias que viabilizam sua permanência no genoma, como a
tendência a se acumular em regiões de heterocromatina (Kidwell e Lisch, 2000;
Kidwell e Lisch, 2001; Lippman et al., 2014). Alguns desses mecanismos se
tornaram tão específicos que coevoluiram e contribuem com benefícios ao
hospedeiro, como o silenciamento mediado por RNA que fornecem um complexo
regulatório adicional aos genes do próprio hospedeiro (Jordan e Miller, 2008). Além
disso, os ETs são considerados fontes de biodiversidade em vertebrados e a

57
instabilidade que eles causam no genoma é o que muitas vezes confere uma rápida
adaptação às mudanças ambientais (Böhne et al., 2008; Schrader et al., 2014, Volff,
2005).
Os peixes teleósteos são detentores da mais alta diversidade de
superfamílias, conteúdo e atividade de ETs entre os vertebrados (Böhne et al., 2011;
Duvernell et al., 2004; Kojima e Jurka, 2011; Ravi e Venkatesh, 2008; Volff et al.,
2003; Volff, 2005). Takifugu rubripes e Tetraodon nigroviridis, por exemplo,
apresentam baixo número de cópias de ETs, mas uma vasta diversidade de
retrotransposons, superando com vantagem as encontradas em mamíferos (Volff et
al., 2003). O mesmo cenário se repete para o elemento Line 1, que em mamíferos
apresenta grande número de cópias de uma única linhagem, enquanto que em
teleósteos existem várias linhagens antigas desse elemento mas restrito número de
cópias (Duvernell et al., 2004).
Dentre os teleósteos, destacam-se os ciclídeos que são um importante grupo
de estudo devido à sua ampla radiação adaptativa, evidenciada pela expressiva
diversidade morfológica, comportamental e ecológica (Kocher, 2004; Fan et al.,
2012; Schneider et al., 2013b). Diferentes superfamílias de elementos transponíveis
já foram identificadas no genoma de ciclídeos neotropicais, tais como Tc1 da
superfamília Mariner e retrotransposons não-LTR L2, RTE e R4 (Tavares et al.,
2016). A manutenção desses elementos nas espécies Cichla vazzoleri, Astronotus
ocellatus, Pterophyllum scalare e Symphysodon discus pode ocorrer por diferentes
mecanismos evolutivos, uma vez que ocorrem de maneira heterogênea, com
diferentes números de cópias entre as espécies (Tavares et al., 2016).
No que tange a mapeamento cromossômico, várias sequências repetitivas já
foram mapeadas no genoma de ciclídeos neotropicais, sendo elas do tipo satélites e
microssatélites, Rex1, Rex3, Rex6 e Tc1 (Gross et al., 2009b; Mazzuchelli e Martins,
2009; Schneider et al., 2013b; Schneider et al., 2015; Teixeira, 2009; Valente et al.,
2011). Ainda que os diferentes ETs tenham sido mapeados nos genomas de
ciclídeos neotropicais, um enfoque comparativo, incluindo espécies basais e
derivadas, está restrito apenas aos retroelementos Rex1, Rex3 e Rex6 (Schneider et
al., 2013b). Diante disso a identificação e mapeamento de outros ETs são
fundamentais no entendimento da dinâmica evolutiva destes ETs, bem como o seu
papel na evolução genômica dos ciclídeos neotropicais.

58
Recentemente genomas de diferentes espécies de ciclídeos neotropicais
foram totalmente sequenciados (Ramos, 2016; Tavares et al., 2016), abrindo assim
a possibilidade de integração entre Next Generation Sequence e mapeamento e
identificação de ETs. Diante disso, o presente estudo tem como objetivo caracterizar
molecularmente e mapear fisicamente diferentes ETs presentes no genoma de
ciclídeos neotropicais, incluindo espécies basais (Cichla vazzoleri e Astronotus
ocellatus) e derivadas (Pterophyllum scalare e Symphysodon discus), gerando assim
dados sobre o papel destes ETs na evolução dos ciclídeos neotropicais.
3.2.2 Material e Métodos
Quatro espécies de ciclídeos neotropicais da Amazônia Central foram
analisados, sendo elas basais Cichla vazzoleri (3 machos e 3 fêmeas) e Astronotus
ocellatus (3 machos e 5 fêmeas), e derivadas Pterophyllum scalare (3 machos e 3
fêmeas) e Symphysodon discus (2 machos e 2 fêmeas). As espécies foram coletas
com autorização do ICMBio (ICMBio SISBIO 22984-1/2011). Dados citogenéticos
clássicos tais como número diploide, localização de heterocromatina e regiões
organizadoras de nucléolo já foram previamente descrita (Schneider et al., 2013a).
3.2.2.1 Preparação cromossômica
Cromossomos mitóticos foram obtidos a partir de células renais usando o
protocolo de Bertollo et al., (1978) que consiste em injetar intraperitonealmente
colchicina 0,0125% na proporção de 1 mL para cada 100 g de peso do animal. Após
40 minutos, os espécimes foram sacrificados pela imersão em água gelada e a
porção anterior do rim foi removida e transferida para uma solução hipotônica de
KCL 0,075 M (6-8 mL). O tecido foi divulsionado com o auxílio de uma seringa de
vidro. Em seguida, o sobrenadante (suspensão celular) foi retirado com o auxílio de
uma pipeta Pasteur, colocado em tubo de centrífuga e incubado em estufa a 37 °C
por 30 minutos. A suspensão foi pré-fixada com 6 gotas de metanol:ácido acético
(3:1) e ressuspendida com cuidado. Após 5 minutos, foi adicionado 8ml de fixador
Carnoy e centrifugado por 10 minutos a 900 rpm. O sobrenadante foi descartado e 6
mL de fixador Carnoy foi adicionado. O material foi novamente ressuspendido e
centrifugado por 10 minutos a 900 rpm, repetindo essa lavagem por mais duas
vezes. Após a última centrifugação e a eliminação do sobrenadante, 1,5 mL de

59
fixador foram adicionados e o material ressuspendido com cuidado. A suspensão
celular foi então estocada em tubo “eppendorf” e mantida em freezer -20°C.
3.2.2.2 Extração de DNA e amplificação por reação em cadeia da
polimerase (PCR)
DNA total foi extraído dos tecidos muscular e hepático usando fenol-
clorofórmio (Sambrook et al., 1989) e quantificado pelo espectrofotometro NanoDrop
2000 (Thermo Scientific). A amplificação dos ETs pela reação em cadeia da
polímerase (PCR) foi realizada utilizando os primers da tabela 1 (Tavares, 2016). A
reação de PCR foi realizada para o volume final de 25 µL, contendo DNA genômico
(200 ng), tampão 1x com 1,5 mM de magnésio, Taq DNA polimerase (5 U/µL),
dNTPs (1 mM), primers (5 mM) e água Milli-Q. As condições dos ciclos para todos
os elementos incluíram os seguintes passos: 95°C por 5 min; 34 ciclos de 95°C por 1
min, 55°C por 40 seg e 72°C por 2 min; e a extensão final a 72°C por 5 min. Os
produtos de PCR foram analisados por eletroforese em gel de agarose à 1%,
quantificados pelo espectrofotometro NanoDrop 2000 (Thermo Scientific) e usados
na clonagem e como sonda na técnica de FISH.
Tabela 1: Lista de primers utilizados.
Nome do primer
Primer forward Nome do
primer Primer reverse
Transponível amplificado
C07R4-F WGGAATCTGCTGGAGCCAYT C07R4-R ACTCTCAGTGGCAGGGGAAR Line/R4/Dong
C11RTE-F TAGGAGCTGCCTGACACAGT C11RTE-R CAAACCATCATGGCCCTGTGT Line/RTE
C22L2-F CTTAGTTACAGCACTTTCAG C22L2-R STGCCTTACAGAAACCTGGT Line/L2
C08Tc1-F ATGATGGYDGTYCTGTTGGC C08Tc1-R GTAGGACATGCCCGGGAACA Mariner/Tc1
3.2.2.3 Clonagem, sequenciamento e análise das sequências
Os produtos de PCR dos elementos transponíveis Mariner/Tc1, Line/L2,
Line/R4/Dong e Line/RTE foram inseridos no plasmídeo pGEM-T Easy (Promega).
Os produtos de ligação foram inseridos em células competentes de Escherichia coli
por eletro e quimiocompetência. Clones carregando o inserto de interesse foram
sequenciados pelo Sequenciador de DNA ABI 3130 XL (Perkin-Elmer) e os
resultados das sequências foram submetidos no banco de dados Repbase Update.
Cada clone foi identificado utilizandos na busca contra o banco de dados Repbase

60
Update no Genetic Information Researche Institute (GIRI)
(http://www.girinst.org/repbase/) utilizando o software CENSOR (Jurka et al., 2005).
As sequências nucleotídicas foram alinhadas usando o programa ClustalW
implementado pelo programa Geneious ® 8.0.5 (Biomatters Limited).
3.2.2.4 Hibridização fluorescente in situ (FISH – Fluorescence in situ
hybridization)
Os produtos de PCR de todos os ET foram marcados com digoxigenin-11-
dUTP (mix Dig-Nick Translation; Roche) por nick translation de acordo com as
recomendações do fabricante e a anti-digoxigenina rodamina (Roche) foi usado para
a detecção do sinal das sondas. Hibridizações fluorescentes in situ homólogas e
heterólogas foram tratadas com 77% de extringência (2,5 ng/µL de DNA, 50%
formamida deionizada, 10% sulfato dextrano e 2x SSC a 37°C por 18 horas) (Pinkel
et al., 1986). Os cromossomos foram contra-corados com DAPI (2 mg/ml) em um
meio de montagem com Vectashield (Vetor). Quatro lâminas e no mínimo 20
metáfases foram analisadas por espécie para cada ET.
3.2.2.5 Processamento microscópico
Quatro lâminas e no mínimo 20 metáfases foram analisadas por espécie para
cada ET. Cromossomos foram analisados em objetiva de 100x usando o
microscópio de epifluorescência Leica DM 2000 e as imagens foram capturadas
usando o software Leica LAS 4.0. Metáfases mitóticas foram processadas pelo
programa Adobe Photoshop CS6. Os cariótipos foram organizados pela ordem
decrescente de tamanho dos cromossomos (Levan et al., 1964).
3.2.3 Resultados
3.2.3.1 Clonagem
Foram sequenciados 14 clones do Transposon Mariner/Tc1. Foram obtidas 2
sequências de Astronotus ocellatus, 9 de Pterophyllum scalare e 3 de Symphysodon
discus. Todas as sequências foram identificadas como TC1_RP e apresentaram de
85 a 100% de identidade quando comparadas ao banco de dados Repbase Update
(Tabela 2).

61
Foram sequenciados 34 clones de Line/R4/Dong. Foram obtidas 12
sequências de Cichla vazzoleri, 7 de Astronotus ocellatus, 9 de Pterophyllum scalare
e 6 de Symphysodon discus. Todas as sequências foram identificadas como
DONG_FR2, salvo 7 sequências identificadas como Rex6 (2 de A. ocellatus, 3 de P.
scalare e 2 de S. discus) e 3 como Dong-1_AFC (2 sequências de P. scalare e 1 de
S. discus). As sequências apresentaram de 71 a 93% de identidade quando
comparadas ao banco de dados Repbase Update (Tabela 2).
Foram sequenciados 27 clones de Line/L2. Foram obtidas 9 sequências de
Cichla vazzoleri, 5 de Astronotus ocellatus, 1 de Pterophyllum scalare e 12 de
Symphysodon discus. No total, 19 sequências foram identificadas como MAUI (9 de
Cichla vazzoleri, 5 de A. ocellatus, 1 de P. scalare e 4 de S. discus), as outras 8
sequências de S. discus foram identificadas como L2-5_OL. As sequências
apresentaram de 70 a 82% de identidade quando comparadas ao banco de dados
Repbase Update (Tabela 2).
Foram sequenciados 13 clones de Line/RTE. Foram obtidas 4 de Astronotus
ocellatus, 5 de Pterophyllum scalare e 4 de Symphysodon discus. Não foram obtidos
clones de Cichla vazzoleri. No total, 10 sequências foram identificadas como
Expander (3 de A. ocellatus, 3 de P. scalare e 4 de S. discus), 2 sequências de A.
ocellatus foram identificadas como Rex3 e RTE-1_OL cada e 1 sequência de P.
scalare como Rex3. As sequências apresentaram de 72 a 87% de identidade
quando comparadas ao banco de dados Repbase Update (Tabela 2).
Tabela 2: Número e identificação das sequências isoladas na clonagem
Elemento transponível
Total de clones
Número de clones em cada
identificação Identificação
Identidade com o banco dados
Tc1 14 14 Tc1_RP 85 a 100%
Dong 34 24 DONG_FR2
71 a 93% 7 Rex 6
3 DONG-1_AFC
L2 27 19 MAUI 70 a 82%
8 L2-5_OL
RTE 13 10 Expander
72 a 87% 2 Rex 3
1 RTE-1_OL

62
3.2.3.2 FISH
Os mapeamentos dos ETs evidenciaram localizações dispersas e/ou
conspícuas no genoma das quatro espécies de ciclídeos. As marcações ocorreram
ao longo de todo o segmento cromossômico e não se restringiram às regiões de
heterocromatina, embora em algumas situações elas ocorram preferencialmente nas
regiões centroméricas, pericentroméricas ou terminais.
A hibridização da sonda Mariner/Tc1 apresentou marcações dispersas e
conspícuas. As marcações dispersas ocorreram em todos os cromossomos, com
exceção dos pares: 14, 19 e 21 de Cichla vazolleri e 18 e 24 de Pterophyllum
scalare. Symphysodon discus apresentou marcações dispersas somente nos pares
3, 5, 6, 9, 10, 11, 16, 18, 19 e 22 e não apresentou blocos conspícuos. As
marcações conspícuas foram menos recorrentes que as dispersas e observadas
somente nos pares: 4 de C. vazzoleri tanto na região centromérica quanto na região
terminal e um sinal intenso no par 6; nos pares 2, 3, 6, 8, 10 e 15 as marcações
ocorrem nas regiões centroméricas e terminais em A. ocellatus, já nos pares 5, 7 e
17 ocorrem fora dessas regiões; nos pares 3, 4, 5, 9 e 11 nas regiões, centromérica,
pericentromérica e terminais de P. scalare, enquanto que os pares 14, 16 e 17 as
marcações conspícuas ocorrem fora dessas regiões. S. discus apresentou fracas
marcações conspícuas nos pares 6, 10, 11 e 18.
A sonda Line/R4/Dong apresentou marcações dispersas em C. vazzoleri
(pares 1, 2, 4, 8, 9, 11, 14 e 17), em todos os pares P. scalare e em S. discus (nos
pares 2, 7-13, 16-18, 24-30). As marcações dispersas apresentam-se bem reduzidas
e fracas nas espécies P. scalare e S. discus. Em A. ocellatus, os pares 5, 8, 10, 12,
14, 16, 21 e 22 apresentaram tanto marcações dispersas quanto conspícuas. Já os
pares 2, 3, 4, 6, 7, 9, 11, 13, 15, 17, 18, 19, 20, 23 e 24 apresentaram somente
marcações conspícuas, mas que não se restringiram às regiões heterocromáticas.
Em S. discus, alguns pares apresentaram marcações conspícuas, tais como o par
12 e o 27. Os outros não apresentaram marcações.
A sonda Line/L2 apresentou marcações dispersas em todos os cromossomos
de C. vazolleri, P. scalare e S. discus e nos pares 8-10 e 23 de A. ocellatus. Em S.
discus, as marcações dispersas dos cromossomos subtelo/acrocêntricos
apresentam um sinal forte e ocorrem ao longo de todo o segmento cromossômico.
As marcações conspícuas foram variantes entre as espécies e não restringiram-se

63
às regiões de heterocromatina. Enquanto que C. vazolleri apresenta marcações sutis
nos pares 1, 2, 5, 6, 9, 10, 11, 14, 17 e 22, S. discus evidencia marcações
conspícuas bem definidas nos pares 1-6, 10, 13-17, 19-21, 24 e 25. O par 1, por
exemplo, apresentou muitas marcações conspícuas, um diferencial em relação aos
outros pares dessa espécie e em relação às outras espécies. As principais
marcações de Line/L2 em A. ocellatus são conspícuas e ocorrem preferencialmente
nos centrômeros e regiões terminais. Os pares que exibiram esse perfil foram 1-5, 7,
11-22 e 24. Em contrapartida, o par número 6 não exibiu qualquer marcação. P.
scalare também apresentou blocos conspícuos evidenciados nos pares 2, 5, 6, 7,
11, 13 e 18.
A sonda Line/RTE apresentou marcações dispersas em todos os
cromossomos de C. vazolleri, nos pares 9, 12-14, 22-24 de A. ocellatus, nos pares
13, 17-24 de P. scalare e nos pares 8, 13, 14, 16-19, 24, 26, 29 e 30 de S. discus.
As marcações dispersas em C. vazzoleri foram sutis e nenhum dos cromossomos
apresentou blocos conspícuos. Para essa sonda, todos os cromossomos que
apresentaram marcações conspícuas também apresentaram marcações dispersas.
São eles: os pares 1-8, 10, 11, 15, 17-21 de A. ocellatus e em todos os pares de S.
discus e P. scalare, exceto os pares 12, 14, 16 desse último.

64
Figura 1 Cariótipos das espécies estudadas hibridizadas com sonda de Mariner/Tc1. Os cromossomos foram contra corados com
DAPI. Barra 10 um.

65
Figura 2 Cariótipos das espécies modelo hibridizadas com sonda de Line/R4/Dong. Os cromossomos foram contra corados com
DAPI. Barra 10 um.

66
Figura 3 Cariótipos das espécies estudadas hibridizadas com sonda de Line/L2. Os cromossomos foram contra corados com
DAPI. Barra 10 um.
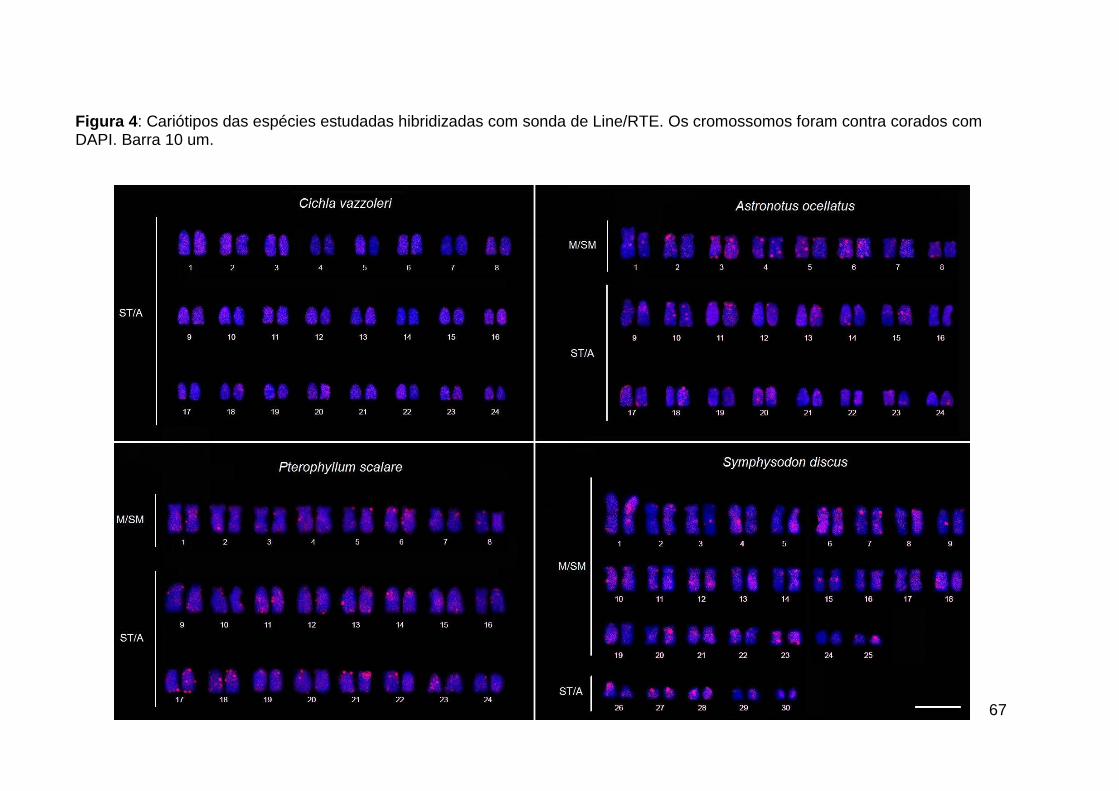
67
Figura 4: Cariótipos das espécies estudadas hibridizadas com sonda de Line/RTE. Os cromossomos foram contra corados com DAPI. Barra 10 um.

68
3.2.4 Discussão
Elementos transponíveis constituem uma fração importante no genoma dos
ciclídeos (Tavares et al., 2016). Pelo menos 4 superfamílias – Line/L2,
Line/R4/Dong, Line/RTE e Mariner/Tc1 - foram identificadas nesse trabalho, uma
diversidade relativamente alta comparada a outros grupos de vertebrados (Chalopin
et al., 2015). Todas as 4 superfamílias são amplamente difundidas entre os peixes
teleósteos (Terai et al., 1998; Volff et al., 2001; Volff et al., 1999; Zupunski et al.,
2001; Radice et al., 1994), enquanto que estão ausentes em outros vertebrados e
provavelmente foram perdidas ao longo da evolução (Böhen et al., 2011; Volff et al.,
2003; Chalopin et al., 2014; Kojima e Jurka, 2011).
3.2.4.1 Diversidade de ETs no genoma de ciclídeos amazônicos
A dinâmica evolutiva dos ETs é claramente distinta entre os organismos
(Ferreira et al., 2011; Chalopin et al., 2015) e isso pode ser exemplificado pela
diversidade das sequências dos ETs. O elemento Mariner/Tc1 foi o único cujas
sequências apresentaram alta similaridade com apenas uma linhagem de ET do
Repbase Update, o TC1_RP. O elemento TC1_RP é um transposon mariner do tipo
Tc1 isolado de Rana pipiens (Miskey et al., 2003). Embora as sequências de
Mariner/Tc1 estejam representadas por uma única linhagem, a variação das
sequências entre as espécies foi de 35%, já considerado uma divergência par-a-par
alta, típica de atividade transponível antiga (Blass et al., 2012).
Quando representantes de uma família de ET estão presentes em espécies
hospedeiras próximas ou distantes filogeneticamente, contendo pequenas regiões
com altos níveis de identidade, considera-se que: ou são elementos transponíveis
muito antigos que acumularam mutações ao longo do tempo antes da separação
das espécies (Capy, 1994), ou são elementos oriundos de transferência horizontal
(Capy, 1994), hipóteses como estas já foram suportadas para peixes teleósteos e os
vertebrados de forma geral (Radice et al., 1994).
O elemento Mariner/Tc1 já havia sido identificado e mapeado em ciclídeos
neotropicais (Teixeira et al., 2009). Ele teria originado MITEs (miniature inverted
repeat transposable elements) no genoma desses peixes e possivelmente sofre
transferência horizontal ao longo da evolução de Cichla kelberi. Estes eventos
possivelmente ocorreram entre esta espécie e Pleuronichthys cornutus e Eptatretus

69
burgeri (Ahn et al., 2013). A alta similaridade do elemento Mariner/Tc1 com as
linhagens isoladas de Rana pipiens somado às evidências de transferência
horizontal em peixes reforça a ideia de que esse elemento pode ter sofrido
transferência horizontal em outros grupos de vertebrados adicionalmete (Miskey et
al., 2003; Kidwell, 1993; Kuraku et al., 2012; Schaack et al., 2010). Desta forma,
Mariner/Tc1 teria uma participação especial, atuando ativamente a dinâmica
evolutiva dos genomas dos vertebrados (Kuraku et al., 2012).
Diferente do Mariner/Tc1, os elementos Line/RTE, Line/L2 e Line/R4/Dong
tiveram suas sequências identificadas em mais de uma família de elemento
transponível pelo Repbase Update. Embora sejam distintos, esses tipos (linhagens)
estão agrupados na mesma superfamília na classificação dos ETs, correspondem ao
elemento esperado e possuem sequências semelhantes entre si. O elemento
Line/R4/Dong apresentou similaridade com três linhagens do banco de dados do
Repbase, mas todas elas pertencem a superfamília Line/R4/Dong e diferem em 30%
entre as espécies. A maioria das sequências apresentou similaridade com a
linhagem DONG_FR2 isolada de Takifugu rubripes da ordem tetraodontiformes
(Smit, 2002), 7 de 32 apresentaram similaridade com a linhagem Rex6-1_OL isolado
de Oryzias latipes da ordem beloniformes (Kojima e Jurka, 2014) e apenas 3
sequências apresentaram similaridade com a linhagem Dong-1_AFC de ciclídeo
africano (Brawand et al., 2014). O elemento DONG_FR2 é considerado um ancestral
de DONG_FR (Smit, 2002), que, quando comparado ao elemento Rex6, revela ser o
mesmo elemento (Volff et al., 2001).
As sequências de C. vazzoleri apresentaram similaridade somente com a
linhagem DONG_FR2. Possivelmente, o controle deste ETs no genoma de C.
vazzoleri é mais rigoroso, pois além de apresentar alta similaridade com apenas
uma linhagem de ET considerada ancestral, as sequências são altamente
conservadas dentro dessa espécie, com menos de 17% de variação, sendo ela uma
espécie basal. Uma possível explicação para baixa variabilidade deste elemento no
genoma de C. vazzoleri, seria que provavelmente este esteja associado a genes e
relacionado aos seus mecanismos de regulação (Rebello et al., 2012), como
verificado em Oryzias latipes, onde o elemento Rex6_Ola, muito similar ao
DONG_FR2, que está truncado ao gene para guanilil-ciclase ou ainda está ativo no
genoma (Volff et al., 2001).

70
Em relação às outras três espécies, a variação entre as sequências dentro de
cada espécie foi superior a 40%, evidenciando um dinamismo menos controlado. A
alta variação entre as sequências dentro de uma espécie e entre as espécies e a
identificação com diferentes linhagens pode ser explicado por algumas hipóteses: i)
esse elemento não apresenta atividade de transposição recente (Capy, 1994) nas
espécies derivadas deste trabalho, contrário ao sugerido para Rex1, Rex3 e Rex6
(Volff et al., 2001; Schneider et al., 2013b); ii) as sequências podem estar alocadas
em regiões de baixa pressão seletiva facilitando essa diversificação, como já foi
sugerido para ciclídeos (Valente et al., 2011); iii) as forças de seleção atuam de
formas diferentes entre as famílias de ETs, como a diferença resultante do sucesso
de transposição e as taxas de eliminação resultando na taxa de evolução distinta em
cada espécie hospedeira (Böhne et al., 2008; Capy, 1994); iv) transferência
horizontal em uma linhagem específica (Böhne et al., 2008; Capy, 1994; Kuraku et
al., 2012); e v) surgimento de novos elementos em clados filogenéticos específicos
(Böhne et al., 2008), como é o caso do elemento Zisupton, presente somente em
peixes teleósteos entre todos os vertebrados (Böhne et al., 2012).
Os teleósteos apresentam a mais alta diversidade de superfamílias de ETs
entre os vertebrados sendo que os mais amplamente encontrados no genoma são
os retrotransposons Line/L2, juntamente com hAT e Tc-mariner (Chalopin et al.,
2015). As sequências de Line/L2 apresentam-se altamente conservadas,
apresentando similaridade com apenas duas linhagens que são intimamente
relacionadas: MAUI e L2-5_OL. MAUI é um retrotransposon não-LTR, superfamília
CR1 e família Line/L2 (Poulter et al., 1999). Inicialmente isolado em Takifugu
rubripes, representa o clado mais abundante nessa espécie (Poulter et al., 1999),
assim como em Gasterosteus aculeatus (Blass et al., 2012). Enquanto o elemento
L2-5_OL também faz parte da família Line/L2 e foi isolado primeiramente do genoma
de Oryzias latipes (Kojima e Jurka, 2014).
Todos os clones do elemento Line/L2 sequenciados das espécies C.
vazzoleri, A. ocellatus e P. scalare foram similares ao retroelemento MAUI, com
menos de 16% de variação entre as espécies. Sequências que apresentam baixa
variação entre si, sendo muito similares ou idênticas, indicam fortemente atividade
recente da família de ET e ainda podem estar ativos no genoma (Blass et al., 2012),
como na espécie Anolis carolinensis (Novick et al., 2009) e Gasterosteus aculeatus

71
(Blass et al., 2012) ou já estiveram, como sugerido para celacanto (Chalopin et al.,
2014) e ciclídeos (Schneider et al., 2013b). Ainda, existem evidências que Line/L2
atua como potencializador do gene GDF6 em peixes de água doce, fornecendo um
provável mecanismo de regulação para modelagem da estrutura óssea (Indjeian et
al., 2016).
S. discus foi o único que apresentou similaridade com duas linhagens de
Line/L2, ou seja, suas sequências foram mais divergentes dentro da espécie que
nas demais. Essa ocorrência pode ser resultado de sua complexa evolução
genômica (Gross et al., 2009b; Gross et al., 2010) associada aos elementos Rex1 e
Rex3 que apresentam sequências mais variantes em S. discus do que em espécies
basais (Schneider et al., 2013b). A filogenia do elemento Rex1 é tão diferente entre
S. discus e outras espécies de ciclídeos neotropicais (Cichla vazzoleri, Astronotus
ocellatus, Pterophylum scalare, Geophagus proximus) que a distância genética entre
eles é elevada (aproximadamente 36%) e, por isso, são separados em dois clados
filogenéticos distintos, sendo que S. discus apresenta-se mais próximo de ciclídeos
africanos do que de ciclídeos neotropicais (Schneider et al., 2013b).
As sequências de Line/RTE apresentaram similaridade com 3 linhagens do
Repbase Update. A maioria das sequências foram similares à linhagem Expander,
que trata-se de um retrotransposon não-LTR pertencente a superfamília Line/RTE
isolado inicialmente em Takifugu rubripes (Kapitonov & Jurka 1999). A linhagem
Expander inclui uma open reading frame (ORF) codificadora de endonuclease e
transcriptase reversa, possivelmente inativa por apresentar várias repetições
tetranucleotídicas dispostas em tandem na extremidade 3’ UTR, característico de
elementos Line/RTE (Kapitonov e Jurka, 1999). Apenas uma sequência de A.
ocellatus foi similar à linhagem RTE-1_OL também pertence à superfamília RTE, foi
isolado primeiramente de Fugu rubripes e 81% similar ao Expander (Kojima e Jurka,
2014). O elemento Expander é o segundo mais abundante no genoma de
Gasterosteus aculeatus e apresenta recente atividade retrotransponível nessa
espécie, devido ao alto nível de similaridade entre os elementos (Blass et al., 2012).
Duas sequências de A. ocellatus e uma de P. scalare foram identificadas
como Rex3, descrito originalmente em Xiphophorus maculatus (Volff et al., 1999). O
elemento Rex3 foi o primeiro Line/RTE descrito nas linhagens de peixes e está
amplamente distribuído entre os teleósteos (Volff et al., 1999). Fujimura et al., (2011)

72
sugerem que Expander e Rex3 tratam-se do mesmo ET. Contudo, quando as
sequências consenso de ambos são alinhadas no presente estudo, elas diferem em
20%, demonstrando que embora sejam muito similares, não são idênticas e portanto
não correspondem ao mesmo elemento. Rex3 apresenta evidências de atividade
retrotransponível recente, sucesso na inserção nos genomas de peixes e várias
cópias estão associadas às regiões codificadoras, o que pode conferir à este ET um
importante papel na evolução genômica (Volff et al., 1999), ou cariotípica dos peixes
teleósteos, ainda que não estejam mais ativos no genoma de ciclídeos tropicais
(Schneider et al., 2013b).
3.2.4.2 Distribuição de elementos transponíveis no genoma de ciclídeos
amazônicos
A ocorrência dos ETs no genoma pode estar associada tanto à
heterocromatina quanto à eucromatina, embora haja uma tendência preferencial
pelas regiões de heterocromatina centroméricas e telomérica, onde existe a
predominância de elementos repetitivos e replicação tardia (Ferreira et al., 2011;
Lippman et al., 2004; Schneider et al., 2013b; Teixeira et al., 2009). Na realidade,
uma das teorias é que o silenciamento da heterocromatina teria como objetivo
evolutivo original o controle dos ETs (Grewal e Jia, 2007). Ainda, presença desses
elementos no genoma já confere estabilidade à molécula (Kazazian, 2004), tais
como os telômeros de Drosophila melanogaster, os quais são alvos de transposição
de retrotransposons e isto seria fundamental para manutenção desta porção
cromossômica, superior às cópias repetitivas geradas pela telomerase (Grewal e Jia,
2007).
A locação prefencial de ETs na heterocromatina pode ocorrer por três
motivos: i) são regiões de baixa pressão seletiva (Valente et al., 2011; Teixeira et al.,
2009), ii) exercem função regulatória ou estrutural no genoma trazendo vantagens
ao hospedeiro (Schneider et al., 2013b; Valente et al., 2011; Kazazian, 2004; Volff et
al., 1999) e iii) são mantidos no genoma hospedeiro por neutralismo (Chalopin et al.,
2015; Schneider et al., 2013b; Kazazian, 2004; Volff et al., 1999). Cada elemento
teve um padrão de distribuição particular. Os padrões variaram tanto quando o
mesmo elemento é comparado entre as espécies, quanto quando são comparados
os diferentes elementos dentro da mesma espécie. Essa grande variação entre as

73
espécie pode ser resultado de um longo processo de divergência entre as
sequências a ponto de elas não serem mais detectadas na hibridização, como já foi
sugerido para Line2 em Oreochromis niloticus e Batrachocottus baikalensis (Oliveira
et al., 1999; Volff et al., 2011). Ainda, é possível que os cariótipos que tenham
marcações fracas ou que aparentemente não apresentem marcação, possuam na
verdade, baixo número de cópias do ET no genoma ou que estes esteja dispersos,
não sendo detectado pela técnica de FISH, como sugerido para o elemento Line/L2
em Oreochromis niloticus (Oliveira et al., 1999), ou foram perdidos em processos de
erosão molecular, já reportado para peixes, répteis e mamíferos (Lovsin et al., 2001;
Schneider et al., 2013b).
Os mapeamentos físicos das diferentes classes de ETs apresentaram
marcações compartimentalizadas coincidentes com as áreas de heterocromatina
constitutiva e dispersas ou conspícuas ao longo do segmento cromossômico
coincidentes com as áreas de eucromatina descritas previamente (Schneider et al.,
2013a). Os que se apresentaram dispersos estão na porção heterocromática do
genoma alternativa às posições centroméricas, pericentroméricas e teloméricas.
Sendo assim, eles não estão na eucromatina necessariamente, só não estão
localizados nas regiões heterocromáticas mais evidentes. Além disso, genes ativos
em blocos heterocromáticos já foram descritos em Drosophila melanogaster
(Yasuhara et al., 2005), em Arabidopsis thaliana (Copenhaver et al., 1999), no
genoma de arroz (Nagaki et al., 2004), tomate (Wang et al., 2006) e em humanos
(Mudge e Jackson, 2005). Por outro lado, alguns trabalhos relatam a presença de
ETs em regiões de eucromatina e ainda inferem que esses elementos podem estar
envolvidos na regulação gênica, como é o caso os elementos Rex1 e Rex3 cujo
mapeamento pela técnica de hibridização fluorescente in situ indicou que a
compartimentalização desses elementos não está restrita somente à
heterocromatina no genoma de Cichla kelberi (Teixeira et al., 2009).
O elemento transponível Mariner/Tc1 foi mais frequentemente localizado em
marcações dispersas ao longo dos cromossomos. Os blocos conspícuos mais
evidentes são nas espécies Astronotus ocellatus e Pterophyllum scalare, entretanto
as marcações dispersas são as mais numerosas. A espécie Cichla vazzoleri
apresentou algumas marcações dispersas enquanto Symphysodon discus quase
não apresentou marcação. O mapeamento cromossômico do Mariner/Tc1 ainda é

74
limitado a poucas espécies de vertebrados, tais como Gymnotus paraguensis (da
Silva et al., 2011) e Cichla kelberi (Teixeira et al., 2009). Em C. kelberi, Mariner/Tc1
apresentou mais marcações compartimentalizadas na heterocromatina
centroméricas (Teixeira et al., 2009). Isso evidencia que a organização genômica do
elemento Mariner/Tc1 é dinâmica e variável entre as espécies de ciclídeos
amazônicos. Já existem registros de transferência horizontal para esse elemento em
outras espécies de ciclídeos (Teixeira et al., 2009), entre peixes teleósteos e
lampréias (Kuraku et al., 2012).
O elemento Line/R4/Dong apresentou muitas marcações dispersas nas
espécies C. vazzoleri e P. scalare e quase nenhuma em S. discus. As marcações de
Astronotus ocellatus foram diferenciais, pois também apresentaram muitos blocos
conspícuos distribuídos ao longo do cromossomo. Este é o primeiro mapeamento
cromossômico do elemento transponível do tipo Line/R4/Dong. As sequências
utilizadas como sonda para Line/R4/Dong foram identificadas em três linhagens:
Dong_FR2, Dong-1_AFC e Rex6. O elemento Rex6 é um membro da família R4 e é
similar às outras linhagens encontradas (Volff et al., 2001). Portanto, de todos os
elementos já mapeados cromossomicamente em vertebrados, Rex6 é o mais similar
ao esperado, ainda que seus dados sejam incipientes. O elemento Rex6 foi
mapeado pela primeira vez em Cichla kelberi onde apresentou marcações
clusterizadas tanto nos centrômeros quanto ao longo dos cromossomos (Teixeira et
al., 2009). Já nas espécies Cichla vazzoleri, Astronotus ocellatus, Geophagus
proximus, Pterophyllum scalare e Symphysodon discus o elemento Rex6 está
organizado em blocos conspícuos localizados principalmente nas regiões
centroméricas e terminais (Schneider et al., 2013b). Essa diferença na composição e
localização indica que esses elementos evoluíram de forma independente e/ou sua
mobilidade no genoma traz vantagens ao hospedeiro, como já sugerido para Rex6
em ciclídeos neotropicais (Schneider et al., 2013b).
O elemento Line/L2 apresenta-se disperso e bem compartimentalizado nos
centrômeros na espécie basal A. ocellatus, enquanto que em S. discus apresenta
muitas marcações clusterizadas ao longo de todo o segmento cromossômico. Além
disso, apresenta também muitas pequenas marcações dispersas, principalmente
nos cromossomos subtelo/acrocêntricos. C. vazzoleri e P. scalare apresentaram
poucas marcações dispersas. Pode ser que esse elemento tenha aumentado em

75
número de cópias nos cromossomos de espécies derivadas (S. discus), enquanto
que nas espécies basais (A. ocellatus) esse elemento tende a estar restrito à
heterocromatina como forma de controle de expressão, como já foi sugerido para os
elementos Rex1 e Rex3 nas espécies C. vazzoleri e S. discus (Schneider et al.,
2013b). Uma alta concentração do Line/L2 está presente no par um de S. discus e
em outros pares cromossômicos. O elemento Line/L2 foi mapeado em Oreochromis
niloticus que exibiu pequenos blocos conspícuos dispersos ao longo dos
cromossomos e evidentes marcações teloméricas na maioria dos cromossomos
(Oliveira et al., 1999). Assim como os resultados para S. discus, o par cromossômico
1 de O. niloticus apresentou uma alta concentração de marcações que, somado à
uma possível associação desse elemento com repetições teloméricas, seria a
confirmação de que o cromossomo 1 originou-se da fusão de três pequenos
cromossomos (Oliveira et al., 1999).
Assim como o elemento Line/R4/Dong, o elemento Line/RTE foi identificado
como três linhagens: Rex3, Expander e RTE1_OL, mas desses três somente o
elemento Rex3 foi mapeado cromossomicamente em ciclídeos até a presente data
(Schneider et al., 2013b; Teixeira et al., 2009; Valente et al., 2010). O elemento
Line/RTE apresentou apenas marcações dispersas na espécie basal C. vazzoleri,
enquanto as demais espécies exibiram tanto marcações dispersas quanto
marcações conspícuas agrupadas preferencialmente nos centrômeros. O padrão de
blocos conspícuos presente nos nossos resultados foi observado em outras
espécies de ciclídeos neotropicais e africanos (Gross et al., 2009b; Schneider et al.,
2013b; Valente et al., 2011), em loricarídeos (Ferreira et al., 2010) e em peixes
antárticos (Ozouf-Costaz et al., 2004). Ainda, a localização e distribuição de Rex1 e
Rex3 coincidiram em Cichla kelberi, estando relacionadas à evolução do grupo
(Teixeira et al., 2009). O mesmo foi observado em outros ciclídeos neotropicais, com
a diferença que as localizações de Rex 1, 3 e 6 correspondiam à ITS (interstitial
telomeric sites), evidenciando rearranjos cromossômicos, e aos sítios ribossomais
5S e 18S (Schneider et al., 2013b).
A expansiva radiação adaptativa dos ciclídeos pode estar relacionada à
divergência genômica promovida pelas diferentes atividades e evolução dos ETs,
como já sugerido para os vertebrados de forma geral (Kidwell e Lisch, 1997; Warren
et al., 2015). Sabe-se, por exemplo, que o dinamismo cariotípico de peixes

76
neotropicais está intimamente relacionado à ocorrência de ETs (Schneider et al.,
2013a). Os peixes teleósteos apresentam várias famílias de ETs ativos no genoma e
isso tem forte relação com as diversas especiações do grupo (Volff, 2005). ETs
inseridos próximos às regiões 5’ e 3’ UTR de genes aumentam significativamente a
expressão gênica em todos os tecidos de diferentes ciclídeos africanos, salvo no
cérebro e músculo esquelético no segundo caso (Brawand et al., 2014).
Portanto, diante da diversidade molecular e a distribuição cromossômica
diferenciada de elementos transponíveis no genoma de ciclídeos neotropicais fica
evidente que as forças evolutivas às quais estão sujeitos agem de formas distintas
para cada um deles. Não há um padrão exclusivo de organização cromossômica dos
ETs no genoma dos ciclídeos amazônicos. Isso porque a dinâmica evolutiva dos
elementos transponíveis é independente da evolução genômica do hospedeiro e da
evolução de outros ETs e, portanto, cada um participa de forma singular na evolução
do genoma hospedeiro.

77
4. Conclusões Gerais
Os genomas das espécies de ciclídeos amazônicos são compostos por
diferentes superfamílias de elementos transponíveis e outros elementos repetitivos,
que se organizam no genoma de diferentes formas.
Os elementos transponíveis identificados, ainda não haviam sido reportados
para ciclídeos neotropicais amazônicos e compreendem as superfamílias Line/L2,
Line/RTE, Line/R4/Dong, Line/Penelope, Line/Rex.babar, Mariner/Tc1, ERV1, e
repetições em tandem do tipo simples e satélites.
O elemento transponível mais representativo em todos os genomas foi o
Line/L2, mas os genomas são compostos em sua maioria por repetições simples.
A ocorrência e representatividade de cada elemento ocorrem de forma
heterogênea entre as espécies, resultando em diferentes organizações genômicas.
Possivelmente, cada elemento esteve ou ainda está sujeito a diferentes tipos de
mecanismos evolutivos que modelam o genoma das espécies de forma
independente entre elas.
A distribuição cromossômica dos elementos transponíveis divergente reforça
a hipótese de que o mesmo elemento, está sujeito a modelagens evolutivas distintas
entre as espécies, podendo ser utilizado na detecção de rearranjos cromossômicos
e contribuir com estudos evolutivos. Maiores proporções dessa perspectiva são
observadas quando diferentes elementos são comparados entre si.
A diversidade molecular dos ETs apresentou certo padrão de similaridade
entre as sequências para cada espécie, sugerindo que esses elementos podem ser
resultado de atividade transponível recente, sem tempo suficiente para terem
acumulado mutações e que ainda podem estar em atividade no genoma das
espécies hospedeiras, abrindo a possibilidade para estudos posteriores.

78
5. Referências Bibliográficas
Ahn, S.J.; Kim, J.Y.; Kim, M.S.; Lee, H.H. 2013. Cloning and characterization of
Tc1 family-derived PPTN related transposons from ridged-eye flounder
(Pleuronichthys cornutus) and inshore hagfish (Eptatretus burgeri). Genes &
Genomics, 35(1), 47-58.
Almeida, L.M.; Carareto, C.M.A. 2005. Multiple events of horizontal transfer of the
Minos transposable element beTween Drosophila species. Molecular phylogenetics
and evolution, 35(3): 583-594.
Anjos, H.D.B.; Siqueira, J.A.; Amorim, R.M.S. 2007. Comércio de peixes
ornamentais do Estado do Amazonas, Brasil. Boletim Sociedade Brasileira de
Ictiologia, 4-5.
Arkhipova, I.; Meselson, M. 2000. Transposable elements in sexual and ancient
asexual taxa. Proceedings of the National Academy of Sciences, 97(26), 14473-
14477.
Bailey, J.A.; Carrel, L.; Chakravarti, A.; Eichler, E.E. 2000. Molecular evidence for
a relationship beTween LINE-1 elements and X chromosome inactivation: the Lyon
repeat hypothesis. Proceedings of the National Academy of Sciences, 97(12), 6634-
6639.
Bao, W.; Kojima, K.K.; Kohany, O. 2015. Repbase Update, a database of
repetitive elements in eukaryotic genomes. Mobile DNA, 6(1), 1.
Bennetzen, J.L.; Wang, H. 2014. The contributions of transposable elements to
the structure, function, and evolution of plant genomes. Annual review of plant
biology, 65, 505-530.
Bertollo, L.A.C.; Takahashi, C.S.; Moreira-Filho, O. 1978. Cytotaxonomic
considerations on Hoplias lacerdae (Pisces, Erythrinidae). Brazilian Journal of
Genetics, 1:103–120.
Blass, E.; Bell, M.; Boissinot, S. 2012. Accumulation and rapid decay of non-LTR
retrotransposons in the genome of the three-spine stickleback. Genome biology and
evolution, 4(5), 687-702.
Blankenberg, D.; Kuster, G. V.; Coraor, N.; Ananda, G.; Lazarus, R.; Mangan, M.;
Nekrutenko, A.; Taylor, J. 2010. Galaxy: a web‐based genome analysis tool for
experimentalists. Current protocols in molecular biology, 19-10.

79
Böhne, A.; Brunet, F.; Galiana-Arnoux, D.; Schultheis, C.; Volff, J.N. 2008.
Transposable elements as drivers of genomic and biological diversity in vertebrates.
Chromosome Research, 16(1), 203-215.
Böhne, A., Zhou, Q., Darras, A., Schmidt, C., Schartl, M., Galiana-Arnoux, D., &
Volff, J. N. (2011). Zisupton—a novel superfamily of DNA transposable elements
recently active in fish. Molecular biology and evolution, msr208.
Bonchev, G.; Parisod, C. 2013. Transposable elements and microevolutionary
changes in natural populations. Molecular ecology resources, 13(5), 765-775.
Bourc'his, D.; Bestor, T. H. 2004. Meiotic catastrophe and retrotransposon
reactivation in male germ cells lacking Dnmt3L. Nature, 431(7004), 96-99.
Bourque, G. 2009. Transposable elements in gene regulation and in the evolution
of vertebrate genomes. Current opinion in genetics & development, 19(6), 607-612.
Brawand, D.; Wagner, C.E.; Li, Y.I.; Malinsky, M.; Keller, I.; Fan, S.;...; Turner-
Maier, J. 2014. The genomic substrate for adaptive radiation in African cichlid fish.
Nature, 513(7518), 375-381.
Brookfield, J. 2005. The ecology of the genome-mobile DNA elements and their
hosts. Nature Reviews Genetics, 5:128–136.
Brum, M.J.I. 1995. Correlações entre a filogenia e a citogenética dos peixes
Teleósteos. Séries Monografias, 2: 5-42. Sociedade Brasileira de Genética, Ribeirão
Preto, SP, Brasil.
Brunet, F.; Godin, F.; David, J.R.; Capy, P. 1994. The mariner transposable
element in the Drosophilidae family. Heredity, 73(4), 377-385.
Cabral Júnior, W.; Almeida, O.T. 2006. Avaliação do mercado da indústria
pesqueira na Amazônia. In: Almeida, O.T. (Ed). A indústria pesqueira na Amazônia.
Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais
Renováveis/ProVárzea. p. 17-40.
Capy, P.; Login, T.; Bigot, Y.; Brunet, F.; Daboussi, M.J.; Periquet, G.; David, J.;
Hartl, D. 1994. Horizontal transmission versus ancient origin: mariner in the witness
box. Genetica, 93:161-170.
Capy, Pierre. Dynamics and evolution of transposable elements. Landes
Bioscience; North American distributor Chapman & Hall, 1998.

80
Casola, C.; Lawing, A.M.; Betrán, E.; Feschotte, C. 2007. PIF-like transposons are
common in Drosophila and have been repeatedly domesticated to generate new host
genes. Molecular biology and evolution, 24(8), 1872-1888.
Chalopin, D.; Fan, S.; Simakov, O.; Meyer, A.; Schartl, M.; Volff, J. N. 2014.
Evolutionary active transposable elements in the genome of the coelacanth. Journal
of Experimental Zoology Part B: Molecular and Developmental Evolution, 322(6),
322-333.
Chalopin, D.; Naville, M.; Plard, F.; Galiana, D.; Volff, J.N. 2015. Comparative
analysis of transposable elements highlights mobilome diversity and evolution in
vertebrates. Genome biology and evolution, 7(2), 567-580.
Charlesworth, B.; Sniegowski, P.; Stephan, W. 1994. The evolutionary dynamics
of repetitive DNA in eukaryotes. Nature Review Article, 371: 215-220.
Charlesworth, B. 2012. The effects of deleterious mutations on evolution at linked
sites. Genetics, 190(1), 5-22.
Cioffi, M.B.; Martins, C.; Bertollo, L.A. 2010. Chromosome spreading of associated
transposable elements and ribosomal DNA in the fish Erythrinus erythrinus.
Implications for genome change and karyoevolution in fish. BMC Evolutionary
Biology, 10(1), 1.
Copenhaver, G.P.; Nickel, K.; Kuromori, T.; Benito, M.I. Kaul, S.; Lin, X.;...;
McCombie, W.R. 1999. Genetic definition and sequence analysis of Arabidopsis
centromeres. Science, 286(5449), 2468-2474.
Cowley, M.; Oakey, R.J. 2013. Transposable elements re-wire and fine-tune the
transcriptome. PLoS Genet, 9(1), e1003234.
Da Silva, M.; Matoso, D.A.; Vicari, M.R.; De Almeida, M.C.; Margarido, V.P.;
Artoni, R.F. 2011. Physical mapping of 5S rDNA in two species of knifefishes:
Gymnotus pantanal and Gymnotus paraguensis (Gymnotiformes). Cytogenetic and
genome research, 134(4), 303-307.
Dias, E.S.; Carareto, C.M.A. 2012. Ancestral polymorphism and recent invasion of
transposable elements in Drosophila species. BMC evolutionary biology, 12(1), 1.
Dupeyron, M.; Leclercq, S.; Cerveau, N.; Bouchon, D.; Gilbert, C. 2014. Horizontal
transfer of transposons beTween and within crustaceans and insects. Mobile DNA,
5(1), 1.

81
Faber-Hammond, J.J.; Phillips, R.B.; Brown, K.H. 2015. Comparative analysis of
the shared sex-determination region (SDR) among salmonid fishes. Genome biology
and evolution, 7(7), 1972-1987.
Fan, S.; Elmer, K.; Meyer, A. 2012. Genomics of adaptation and speciation in
cichlid fishes: recent advances and analyses in African and Neotropical lineages.
Philosophical Transactions of the Royal Society B-Biological Sciences, 367(1587):
385-394.
Fedoroff, N.V. 2012. Transposable elements, epigenetics, and genome evolution.
Science, 338(6108), 758-767.
Feldberg, E.; Porto, J.I.R.; Bertollo, L.A.C. 2003. Chromosomal changes and
adaptation of cichlid fishes during evolution. In: Val, A.L., Kapoor, B.G. (Eds). Fish
adaptation. Science Publishers, Enfield, USA. p. 285-308.
Ferreira, I.A.; Martins, C. 2008. Physical chromosome mapping of repetitive DNA
sequences in Nile tilapia Oreochromis niloticus: Evidences for a differential
distribution of repetitive elements in the sex chromosomes. Micron, 39:411–418.
Ferreira, I.A.; Poletto, A.B.; Kocher, T.D.; Mota-Velasco, J.C.; Penman, D.J.;
Martins, C. 2010. Chromosome evolution in African cichlid fish: contributions from the
physical mapping of repeated DNAs. Cytogenetic and genome research, 129(4),
314-322.
Ferreira, D.C.; Porto-Foresti, F.; Oliveira, C.; Foresti, F. 2011. Transposable
elements as a potential source for understanding the fish genome. Mobile genetic
elements, 1(2), 112-117.
Feschotte, C. 2008. Transposable elements and the evolution of regulatory
networks. Nature Reviews Genetics, 9(5), 397-405.
Finnegan, D.J. 1989. Eukaryotic transposable elements and genome
evolution. Trends in genetics, 5, 103-107.
Freitas, C.E.C.; Rivas, A.A.F. 2006. A pesca e os recursos pesqueiros na
Amazônia Ocidental. Ciência e Cultura, 58 (3): 30-32.
Friedli, M.; Trono, D. 2015. The Developmental Control of Transposable Elements
and the Evolution of Higher Species. Annual review of cell and developmental
biology, 31, 429-451.
Fujimura, K.; Conte, M.A.; Kocher, T.D. 2011. Circular DNA intermediate in the
duplication of Nile tilapia vasa genes. PLoS One, 6(12), e29477.

82
Giardine, B.; Riemer, C.; Hardison, R.C.; Burhans, R.; Elnitski, L.; Shah, P.;
Zhang, Y; Blankenger, D.; Albert, I.; Taylor, J.; Miller, W.; Kent, W.J.; Nekrutenko, A.
2005. Galaxy: a platform for interactive large-scale genome analysis. Genome
research, 15(10), 1451-1455.
Gilbert, C.; Cordaux, R. 2013. Horizontal transfer and evolution of prokaryote
transposable elements in eukaryotes. Genome biology and evolution, 5(5), 822-832.
Grewal, S.I.S.; Jia, S. 2007. Heterochromatin revised. Nature Reviews Genetics,
8:35–46.
Goecks, J.; Nekrutenko, A.; Taylor, J. 2010. Galaxy: a comprehensive approach
for supporting accessible, reproducible, and transparent computational research in
the life sciences. Genome Biol, 11(8), R86.
Gross, M.C.; Feldberg, E.; Cella, D.M.; Schneider, M.C.; Schneider, C.H.; Porto,
J.I.R.; Martins. C. 2009a. An intriguing evidence of translocations in discus fish
(Symphysodon, Cichlidae) and report of the biggest meiotic chromosomal chain in
vertebrates. Heredity, 102:435–441.
Gross, M.C.; Schneider, C.H.; Valente, G.T.; Porto, J.I.R.; Martins, C.; Feldberg,
E. 2009b. Comparative Cytogenetic Analysis of the Genus Symphysodon (Discus
Fishes, Cichlidae): Chromosomal Characteristics of Retrotransposons and Minor
Ribosomal DNA. Cytogenetic and Genome Research, 127:43-53.
Gross, M.C.; Schneider, C.H.; Valente, G.T.; Porto, J.I.R.; Martins, C.; Feldberg E.
2010. Variability of 18S rDNA locus among Symphysodon fishes: chromosomal
rearrangements. Journal of Fish Biology, 76:1117–1127.
Guerra, M. 2004. Hibridização in situ: Princípios básicos. In: Guerra, M. (Ed) FISH:
Conceitos e Aplicações na Citogenética. Editora da Sociedade Brasileira de
Genética, p. 1-32.
Herpin, A.; Braasch, I.; Kraeussling, M.; Schmidt, C.; Thoma, E.C.;Nakamura,
S.;...; Schartl, M. 2010. Transcriptional rewiring of the sex determining dmrt1 gene
duplicate by transposable elements. PLoS Genet, 6(2), e1000844.
Hollister, J.D.; Smith, L.M.; Guo, Y.L.; Ott, F.; Weigel, D.; Gaut, B.S. 2011.
Transposable elements and small RNAs contribute to gene expression divergence
beTween Arabidopsis thaliana and Arabidopsis lyrata. Proceedings of the National
Academy of Sciences, 108(6), 2322-2327.

83
Imai, S.; Sasaki, T.; Shimizu, A.; Asakawa, S.; Hori, H.; Shimizu, N. 2007. The
genome size evolution of medaka (Oryzias latipes) and fugu (Takifugu
rubripes). Genes & genetic systems, 82(2), 135-144.
Indjeian, V.B.; Kingman, G.A.; Jones, F.C.; Guenther, C.A.; Grimwood, J.;
Schmutz, J.; Myers, R.M.; Kingsley, D.M. 2016. Evolving New Skeletal Traits by cis-
Regulatory Changes in Bone Morphogenetic Proteins. Cell, 164(14), 1–12.
Jordan, I.K.; Miller, W.J. 2008. Genome defense against transposable elements
and the origins of regulatory RNA. In Transposons and the Dynamic Genome (pp.
77-94). Springer Berlin Heidelberg.
Jurka, J.; Kapitonov, V. V.; Pavlicek, A.; Klonowski, P.; Kohany, O.; Walichiewicz,
J. 2005. Repbase Update, a database of eukaryotic repetitive elements. Cytogenetic
and genome research, 110(1-4), 462-467.
Kamal, M.; Xie, X.; Lander, E.S. 2006. A large family of ancient repeat elements in
the human genome is under strong selection. Proceedings of the National Academy
of Sciences of the United States of America, 103(8), 2740-2745.
Kapitonov, V.V.; Jurka, J. 1999. The long terminal repeat of an endogenous
retrovirus induces alternative splicing and encodes an additional carboxy-terminal
sequence in the human leptin receptor. Journal of molecular evolution, 48(2), 248-
251.
Kapitonov, V.V.; Jurka, J. 2008. A universal classification of eukaryotic
transposable elements implemented in Repbase. Nature Reviews Genetics, 9(5),
411-412.
Kazazian, H.H.Jr. 2004. Mobile elements: drivers of genome evolution. Science,
303: 1626-1632.
Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.;
Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; Thierer, T.; Ashton, B.; Meintjes, P.;
Drummond, A. 2012. Geneious Basic: an integrated and extendable desktop
software platform for the organization and analysis of sequence
data. Bioinformatics, 28(12), 1647-1649.
Kidwell, M.G. 1993. Lateral transfer in natural populations of eukaryotes. Annual
review of genetics, 27(1), 235-256.

84
Kidwell, M.G.; Lisch, D. 1997. Transposable elements as sources of variation in
animals and plants. Proceedings of the National Academy of Sciences, 94(15), 7704-
7711.
Kidwell, M.G.; Lisch, D.R. 2000. Transposable elements and host genome
evolution. Trends in Ecology & Evolution, 15(3), 95-99.
Kidwell, M.G.; Lisch, D.R. 2001. Perspective: transposable elements, parasitic
DNA, and genome evolution. Evolution, 55(1), 1-24.
Kocher, T. 2004. Adaptive evolution and explosive speciation: The cichlid fish
model. Nature Reviews Genetics, 5(4): 288-298.
Kojima, K.K.; Jurka, J. 2014. Non-LTR retrotransposons from the medaka
genome. Repbase Reports 14(10), 2847-2862.
Kullander, S.O. 2003. Family Cichlidae. In: Reis, R.R.; Kullander, S.O.; Ferraris.
C.J. (Eds). Check list of the freshwater of South and Central America. Editora da
Universidade Católica, Porto Alegre, Brasil. p. 605-654.
Kuraku, S.; Qiu, H.; Meyer, A. 2012. Horizontal transfers of Tc1 elements
beTween teleost fishes and their vertebrate parasites, lampreys. Genome biology
and evolution, 4(9), 929-936.
Lapidus, A.L. 2009. Genome Sequence Databases: Sequencing and Assembly. 3
ed. Academic Press, 2009, p.196-210.
Levan, A.; Fredga, K.; Sandberg, A.A. 1964. Nomenclature for centromeric
position on chromosomes. Hereditas, 52:201–220.
Lippman, Z.; Gendrel, A.V.; Black, M.; Vaughn, M.W.; Dedhia, N.; McCombie,
W.R.; ...; Carrington, J.C. 2004. Role of transposable elements in heterochromatin
and epigenetic control. Nature, 430(6998), 471-476.
Lisch, D. 2013. How important are transposons for plant evolution?. Nature
Reviews Genetics, 14(1), 49-61.
Lohe, A.R.; Moriyama, E.N.; Lidholm, D.A.; Hartl, D.L. 1995. Horizontal
transmission, vertical inactivation, and stochastic loss of mariner-like transposable
elements. Molecular biology and evolution, 12(1), 62-72.
López-Fernández, H.; Honeycutt, R.L.; Winemiller, K.O. 2005. Molecular
phylogeny and evidence for an adaptive radiation of geophagine cichlids from South
America (Perciformes: Labroidei). Molecular and Phylogenetics Evolution, 34:227–
244.

85
López-Fernández, H., Winemiller, K. O., & Honeycutt, R. L. 2010. Multilocus
phylogeny and rapid radiations in Neotropical cichlid fishes (Perciformes: Cichlidae:
Cichlinae). Molecular Phylogenetics and Evolution, 55(3):1070-1086.
Lovšin, N.; Gubenšek, F.; Kordi, D. 2001. Evolutionary dynamics in a novel L2
clade of non-LTR retrotransposons in Deuterostomia. Molecular biology and
evolution, 18(12), 2213-2224.
Lowe-McConnell, R.H. 1991. Ecology of cichlids in South American and African
waters, excluding the African Great Lakes. Cichlid fishes: behaviour, ecology and
evolution, 60-85.
Lowe, C.B.; Bejerano, G.; Haussler, D. 2007. Thousands of human mobile
element fragments undergo strong purifying selection near developmental genes.
Proceedings of the National Academy of Sciences, 104(19), 8005-8010.
Markova, D.N.; Mason-Gamer, R.J. 2015. Diversity, abundance, and evolutionary
dynamics of Pong-like transposable elements in Triticeae. Molecular phylogenetics
and evolution, 93, 318-330.
Martins, C. 2007. Chromosomes and repetitive DNAs: a contribution to the
knowledge of fish genome. In: Pisano, E.; Ozouf-Costaz, C., et al., (Ed.). Fish
Cytogenetics. New Hampshire, EUA: Science Publisher,. cap. 4.4, p.421-453.
Mazzuchelli, J.; Martins, C. 2009. Genomic organization of repetitive DNAs in the
cichlid fish Astronotus ocellatus. Genetica, 136:461–469.
McClintock, B. 1950. The origin and behavior of mutable loci in maize. Nature,
36:344-355.
Mesquita, D.R.; Porto, J.I.R.; Feldberg, E. 2008. Chromosomal variability in the
wild ornamental variability fish Symphysodon spp. (Perciformes, Cichlidae) from
amazon. Neotropical Ichthyology, 6:181–190.
Metcalfe, C.J.; Filée, J.; Germon, I.; Joss, J.; Casane, D. 2012. Evolution of the
Australian lungfish (Neoceratodus forsteri) genome: a major role for CR1 and L2
LINE elements. Molecular biology and evolution, mss159.
Miskey, C.; Izsvák, Z.; Plasterk, R.H.; Ivics, Z. 2003. The Frog Prince: a
reconstructed transposon from Rana pipiens with high transpositional activity in
vertebrate cells. Nucleic acids research, 31(23), 6873-6881.
Mudge, J.M.; Jackson, M.S. 2004. Evolutionary implications of pericentromeric
gene expression in humans. Cytogenetic and genome research, 108(1-3), 47-57.

86
Nagaki, K.; Cheng, Z.; Ouyang, S.; Talbert, P.B.; Kim, M.; Jones, K.M.; Jiang, J.
2004. Sequencing of a rice centromere uncovers active genes. Nature genetics,
36(2), 138-145.
Novak, P.; Neumann, P.; Macas, J. 2010. Graph-based clustering and
characterization of repetitive sequences in next-generation sequencing data. Bmc
Bioinformatics, 11:378-390.
Novak, P.; Neumann, P.; Pech, J.; Steinhaisl, J.; Macas, J. 2013. RepeatExplorer:
a Galaxy-based web server for genome-wide characterization of eukaryotic repetitive
elements from next-generation sequence reads. Bioinformatics, 29(6), 792-793.
Novick, P.A.; Basta, H.; Floumanhaft, M.; McClure, M.A.; Boissinot, S. 2009. The
evolutionary dynamics of autonomous non-LTR retrotransposons in the lizard Anolis
carolinensis shows more similarity to fish than mammals. Molecular biology and
evolution, 26(8), 1811-1822.
Obbard, D.J.; Gordon, K.H.; Buck, A.H.; Jiggins, F.M. 2009. The evolution of RNAi
as a defence against viruses and transposable elements. Philosophical Transactions
of the Royal Society of London B: Biological Sciences, 364(1513), 99-115.
Oliveira, C.; Chew, J.S.; Porto-Foresti, F.; Dobson, M.J.; Wrigth, J.M. 1999. A
LINE2 repetitive DNA sequence from the cichlid fish, Oreochromis niloticus:
sequence analysis and chromosomal distribution. Chromosoma 108:457–468.
Oliveira, S.G.; Bao, W.; Martins, C.; Jurka, J. 2012. Horizontal transfers of Mariner
transposons beTween mammals and insects. Mobile Dna, 3(1), 1.
Ozouf-Costaz, C.; Brandt, J.; Körting, C.; Pisano, E.; Bonillo, C.; Coutanceau, J.P.;
Volff, J.N. 2004. Genome dynamics and chromosomal localization of the non-LTR
retrotransposons Rex1 and Rex3 in Antarctic fish. Antarctic Science 16: 51-57.
Padeken, J.; Zeller, P.; Gasser, S.M. 2015. Repeat DNA in genome organization
and stability. Current opinion in genetics & development, 31, 12-19.
Phillips, R.B.; Reed, K.M. 1996. Application of fluorescence in situ hybridization
(FISH) techniques to fish genetics: a review. Aquaculture, 140.3:197-216.
Piacentini, L.; Fanti, L.; Specchia, V.; Bozzetti, M.P.; Berloco, M.; Palumbo, G.;
Pimpinelli, S. 2014. Transposons, environmental changes, and heritable induced
phenotypic variability. Chromosoma, 123(4), 345-354.

87
Pinkel, D.; Straume, T.; Gray, J.W. 1986. Cytogenetic analysis using quantitative,
high-sensitivity, fluorescence hybridization. Proceedings of the Natural Academy of
Science, 83: 2934-2938.
Poletto, A.B.; Ferreira, I.A.; Cabral-de-Mello, D.C.; Nakajima, R.T.; Mazzuchelli, J.;
Ribeiro, H.B.; Venere, P.C.; Nirchio, M.; Kocher, T.D.; Martins, C. 2010.
Chromosome differentiation patterns during cichlid fish evolution. BMC Genetics,
11:50-62.
Poulter, R.; Butler, M.; Ormandy, J. 1999. A LINE element from the pufferfish
(fugu) Fugu rubripes which shows similarity to the CR1 family of non-LTR
retrotransposons. Gene 227(2), 169-179.
Radice, A.D.; Bugaj, B.; Fitch, D.H.; Emmons, S.W. 1994. Widespread occurrence
of the Tc1 transposon family: Tc1-like transposons from teleost fish. Molecular and
General Genetics MGG, 244(6), 606-612.
Ramos, E. 2016. Genômica comparativa para estudos de evolução em ciclídeos
sul-americanos. Dissertação de mestrado, Instituto de Biociências de Botucatu,
Botucatu, São Paulo. 41p.
Ravi, V.; Venkatesh, B. 2008. Rapidly evolving fish genomes and teleost diversity.
Current opinion in genetics & development, 18(6), 544-550.
Rebollo, R.; Romanish, M.T.; Mager, D.L. 2012. Transposable elements: an
abundant and natural source of regulatory sequences for host genes. Annual review
of genetics, 46, 21-42.
Sambrook, J.; Fritsch, E.F.; Maniatis, T. 1989. Molecular Cloning: a Laboratory
Manual. Cold Springs Harbor Laboratory Press, Cold Springs Harbor, NY.
Schaack, S.; Gilbert, C; Feschotte, C. 2010. Promiscuous DNA: horizontal transfer
of transposable elements and why it matters for eukaryotic evolution. Trends in
ecology & evolution, 25(9), 537-546.
Schneider, C.; Gross, M.C.; Terencio, M.L.; Artoni, R.F; Vicari, M.R.; Martins, C.;
Feldberg, E. 2013a. Chromosomal evolution of neotropical cichlids: the role of
repetitive DNA sequences in the organization and structure of karyotype. Reviews in
Fish Biology and Fisheries, 23(2): 201-214.
Schneider, C.; Gross, M.C.; Terencio, M.L.; do Cromo, E.J.; Martins, C.; Feldberg,
E. 2013b. Evolutionary dynamics of retrotransposable elements Rex1, Rex3 and
Rex6 in neotropical cichlid genomes. Bmc Evolutionary Biology, 13:152-167.

88
Schrader, L.; Kim, J.W.; Ence, D.; Zimin, A.; Klein, A.; Wyschetzki, K.;...; Wurm, Y.
2014. Transposable element islands facilitate adaptation to novel environments in an
invasive species. Nature communications, 5.
Silva, J.C.; Kidwel, M.G. 2000. Horizontal transfer and selection in the evolution of
P elements. Molecular Biology and Evolution, 17:1542-1557.
Slotkin, R.K.; Martienssen, R. 2007. Transposable elements and the epigenetic
regulation of the genome. Nature Reviews Genetics, 8:272-85.
Smit, A.F. 2002. Initial survey of interspersed repeats in Takifugu rubripes.
Repbase Reports, 2(1), 14-14.
Smith, W.; Chakrabarty, P.; Sparks, J. 2008. Phylogeny, taxonomy, and evolution
of Neotropical cichlids (Teleostei : Cichlidae : Cichlinae). Cladistics, 24(5):625-641.
Sparks, J.; Smith, W. 2004. Phylogeny and biogeography of cichlid fishes
(Teleostei : Perciformes : Cichlidae). Cladistics, 20(60):501-517.
Tafalla, C.; Estepa, A.; Coll, J.M. 2006. Fish transposons and their potential use in
aquaculture. Journal of Biotechnology, 123(4): 397-412.
Tavares, E.S.G.M.; Ramos, E.; Martins, C.; Schneider, C.H. 2016. Composição e
Representatividade de Elementos Repetitivos no Genoma de Ciclídeos Neotropicais.
Em preparação.
Teixeira, W.G.; Ferreira, I.A.; Cabral-de-Mello, D.C.; Mazzuchelli, J.; Valente, G.T.;
Pinhal, D.; Poletto, A.B.; Venere, P.C.; Martins, C. 2009. Organization of repeated
DNA elements in the genome of the cichlid fish Cichla kelberi and its contributions to
the knowledge of fish genomes. Cytogenetic and Genome Research, 125:224–234.
Terai, Y.; Takahashi, K.; Okada, N. 1998. SINE cousins: the 3'-end tails of the two
oldest and distantly related families of SINEs are descended from the 3'ends of
LINEs with the same genealogical origin. Molecular biology and evolution, 15(11),
1460-1471.
Treangen, T.J.; Salzberg, S.L. 2012. Repetitive DNA and next-generation
sequencing: computational challenges and solutions. Nature Reviews Genetics,
13(1), 36-46.
Turner, G.F. 2007. Adaptive radiation of cichlid fish. Current Biology,17(19), R827-
R831.
Valente, G.T.; Mazzuchelli, J.; Ferreira, I.A.; Poletto, A.B.; Fantinatti, B.E.A.;
Martins, C. 2011. Cytogenetic mapping of the retroelements Rex1, Rex3 and Rex6

89
among cichlid cish: new insights on the chromosomal distribution of transposable
elements. Cytogenetic and Genome Research, 133:34-42.
Van Sluys, M.A.; Scortecci, K.C.; Costa, A.P.P. 2001. O genoma instável,
sequências genéticas móveis. In: Matioli, S.R. (Ed) Biologia Molecular e Evolução.
Holos Editora, Ribeirão Preto, São Paulo. p. 70-81.
Vicari, M.R.; Artoni, R.F.; Moreira-Filho, O.; Bertollo, L.A.C. 2006. Basic and
molecular cytogenetics in freshwater Cichlidae (Osteichthyes, Perciformes).
Karyotypic conservationism and divergence. Caryologia, 59:260-266.
Vitte, C.; Fustier, M.A.; Alix, K.; Tenaillon, M.I. 2014. The bright side of
transposons in crop evolution. Brief Funct Genomics, 13(4): 276-295.
Volff, J.N.; Körting, C.; Sweeney, K.; Schartl, M. 1999. The non-LTR
retrotransposon Rex3 from the fish Xiphophorus is widespread among teleosts.
Molecular biology and evolution, 16(11), 1427-1438.
Volff, J.N.; Körting, C.; Froschauer, A.; Sweeney, K.; Schartl, M. 2001. Non-LTR
retrotransposons encoding a restriction enzyme-like endonuclease in vertebrates.
Journal of Molecular Evolution, 52(4), 351-360.
Volff, J.N.; Laurence, B.; Zouf-Costaz, C.; Fischer, C. 2003. Diversity of
retrotransposable elements in compact pufferfish genomes. Trends in Genetics,
19(12): 674-678.
Volff, J.N. 2005. Genome evolution and biodiversity in teleost fish. Heredity, 94,
280–294.
Yamada, K.; Yamada, A.; Kawanishi, Y.; Deep Gurung, R.; Tokuda, G.; Maekawa,
H. 2015. Widespread distribution and evolutionary patterns of mariner-like elements
among various spiders and insects. Journal of Insect Biotechnology and Sericology,
84(2), 2_029-2_041.
Yano, A.; Guyomard, R.; Nicol, B.; Jouanno, E.; Quillet, E.; Klopp, C.;...; Guiguen,
Y. 2012. An immune-related gene evolved into the master sex-determining gene in
rainbow trout, Oncorhynchus mykiss. Current Biology, 22(15), 1423-1428.
Yasuhara, J.C.; DeCrease, C.H.; Wakimoto, B.T. 2005. Evolution of
heterochromatic genes of Drosophila. Proceedings of the National Academy of
Sciences of the United States of America, 102(31), 10958-10963.

90
Wallau, G.L.; Capy, P.; Loreto, E.; Hua-Van, A. 2014. Genomic landscape and
evolutionary dynamics of mariner transposable elements within the Drosophila
genus. BMC genomics, 15(1), 1.
Wang, Y.; Tang, X.; Cheng, Z.; Mueller, L.; Giovannoni, J.; Tanksley, S.D. 2006.
Euchromatin and pericentromeric heterochromatin: comparative composition in the
tomato genome. Genetics, 172(4), 2529-2540.
Warren, I.A.; Naville, M.; Chalopin, D.; Levin, P.; Berger, C.S.; Galiana, D.; Volff,
J.N. 2015. Evolutionary impact of transposable elements on genomic diversity and
lineage-specific innovation in vertebrates. Chromosome Research, 23(3), 505-531.
Wicker, T.; Sabot, F.; Hua-Van, A.; Bannetzen, J.L.; Capy, P.; Chalhoub, B.; et al,.
2007. A unified classification system for eukaryotic transposable elements. Nature
Reviews Genetics, 8(12): 973-982.
Wright, S.; Finnegan, D. 2001. Genome evolution: sex and the transposable
element. Current Biology, 11(8), R296-R299.
Zeigler, D. 2014. Transposable elements, viruses, and genomes. In: Zeigler, D.
(Ed). Evolution: Components and Mechanisms. v.1. Academic Press, p.55-60.
Župunski, V.; Gubenšek, F.; Kordis, D. 2001. Evolutionary dynamics and
evolutionary history in the RTE clade of non-LTR retrotransposons. Molecular
Biology and Evolution, 18(10), 1849-1863.
Zwick, M.S.; Hanson, R.E.; McKnight, T.D.; Nurul-Islam-Faridi, M.; Stelly, D.M.
1997. A rapid procedure for the isolation of Cot-1 DNA from plants. Genome, 40:138-
142.


















