
FIT 5806 - BIOTECNOLOGIAS -...
111
1 UNIVERSIDADE FEDERAL DE SANTA CATARINA CENTRO DE CIÊNCIAS AGRÁRIAS DEPARTAMENTO DE FITOTECNIA FIT 5806 - BIOTECNOLOGIAS APOSTILA (v.2016) Rubens Onofre Nodari Doutor em Genética (UCDavis-CA), Prof. Titular do Dep. de Fitotecnia, Centro de Ciências Agrárias, Universidade Federal de Santa Catarina, C. Postal 476, Florianópolis, SC, 88040-900, e-mail: [email protected] Miguel Pedro Guerra Doutor em Ciências (USP), Prof. Titular do Dep. de Fitotecnia, Centro de Ciências Agrárias, Universidade Federal de Santa Catarina, C. Postal 476, Florianópolis, SC, 88040-900, e-mail: [email protected] Adriana Cibele de Mesuita Dantas Doutora em Ciências (UFPel, Profa. Adjunto em Biotecnologia UERGS – RS, e-mail [email protected] Valdir Marcos Stefenon Doutor em Ciências Florestais/Genética (Uni-Göttingen-Alemanha), Prof. Adjunto Universidade Federal do Pampa, São Gabriel - RS e-mail: [email protected] Sarah Zanon Agapito Tenfen Doutora, Genok, Center of Biosafety,Tromso, Noruega e-mail: [email protected] Gustavo Henrique Ferrerro Klabunde Doutor, Programa de Pós-graduação em Recursos Genéticos Vegetais, UFSC, Florianópolis, e-mail: [email protected] Maio de 2016
Transcript of FIT 5806 - BIOTECNOLOGIAS -...

1
UNIVERSIDADE FEDERAL DE SANTA CATARINA
CENTRO DE CIÊNCIAS AGRÁRIAS
DEPARTAMENTO DE FITOTECNIA
FIT 5806 - BIOTECNOLOGIAS APOSTILA (v.2016)
Rubens Onofre Nodari
Doutor em Genética (UCDavis-CA), Prof. Titular do Dep. de Fitotecnia, Centro de Ciências Agrárias, Universidade Federal de Santa Catarina, C. Postal 476,
Florianópolis, SC, 88040-900, e-mail: [email protected]
Miguel Pedro Guerra Doutor em Ciências (USP), Prof. Titular do Dep. de Fitotecnia, Centro de Ciências
Agrárias, Universidade Federal de Santa Catarina, C. Postal 476, Florianópolis, SC, 88040-900, e-mail: [email protected]
Adriana Cibele de Mesuita Dantas
Doutora em Ciências (UFPel, Profa. Adjunto em Biotecnologia UERGS – RS, e-mail [email protected]
Valdir Marcos Stefenon
Doutor em Ciências Florestais/Genética (Uni-Göttingen-Alemanha), Prof. Adjunto Universidade Federal do Pampa, São Gabriel - RS e-mail:
Sarah Zanon Agapito Tenfen
Doutora, Genok, Center of Biosafety,Tromso, Noruega e-mail: [email protected]
Gustavo Henrique Ferrerro Klabunde Doutor, Programa de Pós-graduação em Recursos Genéticos Vegetais, UFSC,
Florianópolis, e-mail: [email protected]
Maio de 2016

2
CONTEÚDO PARTE 1 – Princípios de Genética Molecular 1-Introdução às macromoléculas: proteínas e ácidos nucleicos 4 1.1-Proteínas 4 1.2-Ácidos nucleicos 6 2-Replicação 16 3-Transcrição 17 4-Tradução 18 5-Mutação e reparo 19 6-Metilação 22 7-Regulação gênica 8-Epigenética
22 23
PARTE 2 – Marcadores genéticos 1- Introdução 25 2-Marcadores morfológicos 25 3-Marcadores proteínas de sementes 25 4-Isoenzimas 27 5-RFLPs 29 6-Minissatélites 30 7-RAPDs 31 8-Microssatélites 33 9-AFLPs 35 10-SCARs 37 11-SNPs 38 12-Análise comparativa 39 13-Aplicações dos marcadores moleculares 39
PARTE 3 – Organismos Geneticamente Modificados 1-Introdução 44 2-Tecnologia do DNA recombinante 3-Genes marcadores e genes reporteres 4- Métodos de Transformação de plantas
45 46 46
5-Diferenças entre os métodos de melhoramento convencionais e biotecnológicos 49 6-Aplicações 49 7-Evolução do desenvolvimento e cultivo de plantas transgênicas 56 8-Limitações 61 9-Biossegurança – Regulamentação 62 10-Fiscalização 67 11-Análise de Risco 68 12-Princípio da Precaução 84 13-Rotulagem 86
14- Nova Tecnologia CRISPR 87
PARTE 4 – Direitos De Propriedade Intelectual 1-Direitos de proteção e patentes 89 2-Lei de proteção das cultivares 90 3- Implicações Das Normas De Propriedade Intelectual Sobre Transgênicos 92 4-Biodiversidade, Biotecnologia e Agricultura 93
PARTE 5 – Bioética 1-Introdução 95 2-Percepção Pública 97 3- Os interesses econômicos da transgênia 98 4- A Relação Da Comunidade Científica Com O Governo 100 5- A Necessidade De Um Debate Público Com A Sociedade 101 6- Implicações da clonagem de animais e humanos 102 7- Terapia Genética Com Vetores Recombinantes Na Espécie Humana 103 8- O Que Se Espera Dos Profissionais Da Biologia E Da Agronomia? 104 9- Conclusões 105
BIBLIOGRAFIA 106

3
APRESENTAÇÃO
Esta apostila reúne conteúdos básicos de biologia celular e molecular e suas decorrentes aplicações biotecnológicas e outras técnicas de uso freqüente, visando conhecer, conservar e melhorar a diversidade genética existente. O objetivo desta apostila é proporcionar ao estudante um conjunto de informações básicas e as principais aplicações das biotecnologias. Este conjunto de informações se constitui no ponto de partida para estudos mais aprofundados. As biotecnologias em seu sentido mais amplo compreendem a manipulação de microorganismos, plantas e animais, objetivando a obtenção de processos e produtos de interesse. Desta maneira, toda atividade que envolva a aplicação dos conhecimentos de fisiologia, bioquímica e genética, é considerada como técnica biotecnológica. Em seu senso mais restrito as biotecnologias compreendem a associação de técnicas mais sofisticadas de biologia molecular e celular, engenharia genética e manipulações celulares in vitro. Para o CNPq, biotecnologia pode ser conceituada como a utilização de sistemas celulares para a obtenção de produtos e desenvolvimento de processos. A FAO (1989) conceitua biotecnologia como a aplicação dos princípios científicos e de engenharia para o processamento de materiais por agentes biológicos proporcionando produtos ou serviços. Mais especificamente, a Tecnologia do DNA recombinante possibilitou a obtenção de organismos geneticamente modificados ou transgênicos. As primeiras atitudes do governo brasileiro em relação às biotecnologias tiveram inicio em meados da década de 1980, quando tanto o CNPq quanto o MCT iniciaram o apoio à formação de recursos humanos. Atualmente, o volume de recursos, o número de bolsas e o número de pesquisadores que trabalham com as biotecnologias na área agrícola e florestal atingem valores inferiores a 10% em relação às demais áreas de C&T no país. Contudo, é cada vez maior o número de pessoas envolvidas com as biotecnologias, as quais passam a ser utilizadas nas diversas disciplinas da área biológica. No estado de São Paulo, a FAPESP, a agência de fomento a pesquisa do estado de São Paulo, financiou um projeto para o sequenciamento da bactéria Xyllela fastidiosa, o agente causador da doença denominada de amarelinho em citrus. Outros programas de pesquisa em biotecnologia de plantas estão em progresso em café, cacau, soja, milho, trigo e outras espécies de importância econômica. Mais recentemente o governo vem destinando mais improtância e recursos para as modernos biotecbologias, que incluem a transgenia. A clonagem de mamíferos, obtidas em 1997, desencadeou uma discussão não só no seio da comunidade científica, mas também em toda a sociedade sobre as implicações do poder das biotecnologias. Toma corpo então a Bioética, que discute o modo de ser (ética) da vida. A bioética pergunta-se sobre a legitimidade dos projetos de efeitos biotecnológicos. Vários agrônomos estão desenvolvendo atividades na geração de processos e produtos, utilizando estas técnicas biotecnologias. O mercado tende a uma expansão nos próximos anos. Além dos conhecimentos técnicos necessários ao desempenho profissional, o Engenheiro Agrônomo tem um importante papel na discussão das questões relacionadas com as biotecnologias com a sociedade. A liberação da soja transgênica em setembro de 1998, resistente ao herbicida glifosate, constitui-se num marco da agricultura e exige que os profissionais formados tenham o conhecimento técnico e científico não só para o correto manuseio destes organismos como também para participar das decisões a respeito das mesmas. Agradecemos aos estudantes de pós-graduaçao pelas contribuições a esta apostila.
Os Autores

4
PARTE 1 – PRINCÍPIOS DE GENÉTICA MOLECULAR
1-INTRODUÇÃO ÀS MACROMOLÉCULAS: PROTEÍNAS E ÁCIDOS NUCLEICOS
1.1-Proteínas
Proteínas são cadeias de aminoácidos (aa). A estrutura básica é composta de um esqueleto e de grupos laterais variáveis (Figura 1.1). Uma série repetida de ligações peptídicas entre o carbono de um aa e o nitrogênio de outro aa formam moléculas grandes, as proteínas (Figura 1.2). Devido a natureza da ligação peptídica, uma das extremidades da proteína é H2N (H3N
+), denominada de N-terminal, e na outra extremidade encontra-se COOH (COO-), que é chamada de carboxi-terminal. Existem cerca de 20 aa, cada um com sua forma e constituição química característica. Dependendo da composição, as proteínas podem ter carga positiva, neutra ou negativa. Os aa lisina, arginina e histidina contribuem com carga positiva (denominados de básicos) enquanto que o ácido aspártico e o ácido glutâmico são carregados negativamente (denominados de ácidos). Os demais 15 aa são neutros com relação a carga elétrica. Destes, os polares são: serina, treonina, tirosina, triptofano, asparigina, glutamina e cisteína. Os demais apresentam propriedades hidrofóbicas (não polar): alanina, fenilalanina, glicina, isoleucina, leucina, metionina, prolina e valina. Tais propriedades (polaridade e a hidrofobia) também são incorporadas às proteínas.
Os tipos de aa incluídos e principalmente a sua sequência determinam a conformação tridimensional e portanto, as propriedades de todas as proteínas. O tamanho de uma proteína pode variar de alguns poucos até 30.000 aa. Trinta ou 40 aa são suficientes para proporcionar uma conformação terciária.
Figura 1.1: Estrutura geral de um aminoácido mostrando suas estruturas fixas e o radical variável, porção que diferencia os diferentes aminoácidos
Figura 1.2: oligopeptídeo formado por quatro aminoácidos unidos por ligações peptídicas (em vermelho). O primeiro aminoácido (glicina, com o radical H) apresenta a extremida N-terminal, enquanto o último aminoácido (alanina, com o radical CH3) apresenta a extremidade carboxi-termina.

5
A estabilidade das proteínas representa um equilíbrio entre a sua síntese e a sua degradação. Existe um processo contínuo de reposição (turnover) que pode ser caracterizado quando se conhece a meia-vida das proteínas, ou seja o tempo necessário para a renovação da metade da sua concentração. A meia-vida das proteínas pode variar de minutos a mais de 20 horas e sua degradação é catalisada por enzimas proteolíticas. Exemplos: proteínas com N-terminal arginina - 2 min; lisina, leucina e fenilalanina - 3 min; prolina - 7 min; tirosina e glutamina - 10 min.
Na maioria das vezes as proteínas para exercerem suas funções devem sofrer modificações, como fosforilação, glicosilação ou metilação. No processo de fosforilação é adicionado à proteína um grupo fosfato pelas kinases, tonando-se fosfoproteínas. A metilação ou acetilação consiste na incorporação de um metil ou acetil à proteína pelas metilases ou acetilases, respectivamente. A incorporação de carboidratos numa cadeia protéica denomina-se glicosilação, origina as moléculas denominadas de glicoproteínas.
Enzima é a denominação de uma proteína quanto esta apresenta a habilidade de acelerar uma reação fazendo ou quebrando uma ligação (covalente) específica. Para o exercício desta função, as proteínas devem apresentar a conformação terciária ou quaternária. A conformação quaternária é na realidade a agregação de duas ou mais sub-unidades, que nesta condição proporcionam a função catalisadora à uma proteína enzima. Exemplo: Rubisco ou ribulose-1,5-bifosfato carboxilase/oxigenase se torna uma enzima quando oito sub-unidades se agrupam, quatro delas codificadas por genes nucleares e as outras quatro por genes do cloroplasto. A Rubisco é responsável pela inclusão de CO2 numa
cadeia de carbono (1ª etapa no ciclo de Calvin). Tratando-se de enzimas, nem todos os aa
participam da reação catalítica. Existe um sítio ativo responsável pela catálise. Este sítio ativo é então um conjunto de aa denominado de motivo ou domain. A domain pode ser entendida como a unidade funcional de uma proteína, uma região relativamente independente da proteína. Nas interações com outras proteínas ou ácidos nucleicos apenas uma parte da proteína, o motivo (ou domain), é responsável pela função.
Quando diferentes proteínas desempenham funções semelhantes, constituem uma família de proteínas. A mesma seqüência formadora de uma determinada domain pode se encontrada em várias proteínas de espécies diferentes. Aparentemente, durante a evolução a domain se moveu dentro da sequência linear de aminoácidos sem perder sua função e especificidade de ligação. Estas domains variam quanto ao número de aa: 18 no Colágeno, mais de 250 aa Fibrinogênio. Freqüentemente, as domains podem se repetir (até mais de 30) numa mesma proteína, neste caso denominadas de motivo (motif) sendo que nem todas as repetições são exatamente idênticas. Estas duplicações provavelmente são devido a existência de elementos móveis ou transformação. As duplicações têm provocado a elongação de muitas proteínas. Estimativas admitem a existência de mais de 50 mil tipos de proteínas numa espécie eucariota.
As primeiras técnicas de separação de macromoléculas, foram desenvolvidas na década de 40. Nesta época foi desenvolvido os sistemas de cromatografia que permitem a separação das frações polares das não polares com base na solubilidade das diferentes moléculas. De acordo com este princípio, um solvente não polar move-se carregando solutos com ele. As substâncias migram a diferentes distâncias de acordo com a sua solubilidade no solvente. Atualmente existem uma dezena de diferentes técnicas de cromatografia, que possibilitam inclusive a identificação de moléculas presentes numa mistura.
Nos anos 80 foi descoberto que algumas doenças (desordens degenerativas) poderiam ser causadas por agentes infecciosos formados apenas por proteínas. Estas proteínas foram denominadas de príons ('proteinaceous infections particles'). O príon é uma forma alterada da proteína PrP que normalmente está presente no cérebro de vertebrados. Estas desordens degenerativas ocorrem com freqüência em animais e muito raro na espécie humana.

6
O sequenciamento de proteínas é uma técnica, desenvolvida por (Sanger, 1950), com a finalidade de conhecer a seqüência dos aa numa proteína. As implicações desta descoberta são inúmeras. A mais importante se relaciona com a saúde humana, pois a técnica permitiu a identificação de inúmeras doenças. Mutações ao nível de DNA podem provocar a substituição de um aa por outro numa determinada posição da seqüência de uma proteína e dependendo da posição a proteína perde sua função, causando então uma doença. Outra conseqüência foi a possibilidade de inferência da seqüência de bases ao nível de DNA que codifica para as proteínas sequenciadas. Isto permitiu o isolamento e a clonagem dos primeiros genes. Mais tarde, o próprio Sanger desenvolveu um método de sequenciamento de DNA. Por esta contribuição à ciência, Sanger foi agraciado com um segundo prêmio Nobel.
1.2-ÁCIDOS NUCLEICOS (Griffiths et al., 2015)
1.2.1-Ácido desoxirribonucleico - DNA
As moléculas de DNA têm estrutura em forma de dupla hélice, semelhante a de uma escada retorcida. Cada fita é formada por uma seqüência de nucleotídeos (dNTP). Cada dNTP é composto de uma base nitrogenada ligadas a uma molécula de açúcar (desoxirribose) e um grupo fosfato. As bases nitrogenadas ligadas a desoxirribose são quatro: adenina (A), citosina (C), guanina (G) e timina (T). Uma ligação fosfodiéster unindo o grupo fosfato de um dNTP e o açúcar desoxirribose de outro dNTP forma o esqueleto da fita (strand), como se fosse uma das laterais da escada. A outra fita (ou a outra lateral da escada) é formada da mesma maneira, mas com orientação da ligação fosfodiéster contrária, o que impõe a característica de antiparalelismo as duas fitas. Cada fita tem uma orientação (5'-3') em função da natureza da ligação fosfodiéster entre o carbono 3' e o 5' da desoxirribose, sendo que um nucleotídeo só pode ser incluído na cadeia através da ligação do fosfato com o carbono 3'OH da desorribose. Por isto, a orientação da cadeia é 5'-3', pois haverá sempre o carbono 3' numa das extremidade da fita.
Mais do que isto, estas duas fitas são complementares já que quando existir adenina de um lado, somente timina é encontrada na mesma posição na outra fita. O mesmo acontece com citosina e guanina. São estes os dois únicos tipos complementação de bases nitrogenadas possíveis no DNA. Como conseqüência o número de adeninas será igual ao número de timinas num organismo. O mesmo vale para C e G. Entretanto a quantidade de purinas (A e G) é característica de cada espécie. Assim a proporção entre A e G é de 0,7 em Bacillus, 1,56 no homem e 1,7 em Saccharomyces cereviseae. Isto é conhecido como regra de Chargaff.
Entre as bases nitrogenadas existem pontes de hidrogênio, duas entre A e T e três entre C e G. Tais pontes juntamente com outras forças, mantêm as duas fitas unidas. Cada par de bases é análogo a um degrau desta escada. O DNA funciona como um modelo para a síntese de novas fitas de DNA. O DNA é a molécula responsável pelo armazenamento e perpetuação do código genético. Apesar da ocorrência de 3 tipos de DNAs ('A', 'Z', 'B'), aparentemente desempenham a mesma função.
A prova definitiva de que o DNA é a molécula repositório do código genético foi obtida em 1952 por Hershey e Chase. Experimentalmente adicionou-se 32P numa colônia de bactérias infectadas por vírus, neste caso o fósforo radioativo foi incorporado no DNA, já que pouco ou quase nenhum fósforo é encontrado nas proteínas. Num experimento paralelo, foi feita a adição do isótopo 35S, que pode marcar radioativamente as proteínas, já que estas têm enxofre, mas não marca o DNA, pois este não contém enxofre. Como só o 32P foi detectado nas progênies dos vírus, conclui-se que o DNA passava de geração a geração. Na realidade, oito anos antes, outros três cientistas (Avery, MacLead e McCarty) haviam postulado que o agente transformador (possivelmente o DNA) era destruído pela desorribonuclease pancreática que por sua vez não afetava as proteínas.

7
A quantidade de DNA pode variar de 103 a 1013 nucleotídeos. Esta quantidade de DNA por célula haplóide é denominada de valor C. São aproximadamente 3 bilhões de pares de bases no núcleo de cada célula humana. Entretanto podem ser apenas 5387 no
vírus x174. A maioria das plantas tem uma quantidade de DNA que varia entre 109 a 1011. Nos mamíferos existem de 109 a 1010 pares de bases; já alguns peixes ou anfíbios podem ter até 1013 pares de bases. É muito DNA para pouca função (paradoxo do valor C). Enquanto nos procariotos praticamente quase todo o DNA carrega informações necessárias para a síntese de proteínas e RNAs, a maior parte da seqüência de bases dos eucariotos não codifica para produto algum. Assim apenas 3% (aproximadamente) do genoma humano é formado por genes (estimados em mais de 50 mil) sendo que a função do restante ainda não está suficientemente compreendida. A maior parte deste DNA sem função conhecida é composto por seqüências repetidas, de onde se originou o nome de DNA repetitivo (selfish, nos anos 80).
Quando esticada, uma molécula de DNA de qualquer célula humana mediria 1,80 m e teria a espessura de um trilionésimo de um centímetro (1 micrômetro = 1 milésimo de milímetro). Uma célula humana não comportaria tal estrutura. Dentro de uma célula as moléculas de DNA estão ligadas a proteínas e são retorcidas ou enroladas (supercoil). Quando completamente compactadas são possíveis de serem visualizadas no microscópio ótico e recebem a denominação de cromossomos. A compactação pode alcançar um fator de 7000 vezes. Vírus e bactérias contêm apenas um cromossomo. Já os eucariotos (fungos, plantas, animais) têm dois ou mais cromossomos que em geral, variam de tamanhos.
Figura 1.3: Nucleotídeos formados com as pentoses ribose (formam RNAs) ou desoxiribose (forma DNA). A diferença entre as pentoses está realçada em vermelho.
Figura 1.4: ligação entre dois desoxirribonucleotídeos (dNTPs), através de uma ligação fosfodiéster (em vermelho) entre o grupo fosfato de um dNTP e a pentose de outro dNTP. Os carbonos 5‟ e 3‟ estão realçados em azul.

8
Genoma e gene
A seqüência de pares de bases que formam o DNA pode ser chamada de genoma. A forma do genoma pode ser circular como nos vírus, bactérias, mitocôndria, cloroplasto e plasmídeos ou linear como nos cromossomos dos organismos eucariotos e alguns procariotos. O genoma da maioria absoluta dos organismos é de DNA. Poucos vírus são de RNA, como Influenza, HIV, TMV, poliomielite. A grande maioria também apresenta fita
dupla. Exceção a alguns vírus como (x174, M13 e f1, cujos genomas são constituídos de apenas uma fita de DNA. As características de um indivíduo como a cor dos olhos ou da pele são determinadas por um conjunto limitado de pares de bases contidas no DNA (ou no RNA, como já mencionado, trecho este, denominado de gene.
O conceito de gene evoluiu tanto quanto a biologia. Uma das primeiras observações sobre o tema foi feita por Leonardo da Vinci. Observando a cor dos filhos de mulheres brancas com homens pretos, ele sugeriu que a semente da mãe tinha o mesmo vigor que a do pai (Wallace, 1992). Mas foi Mendel em 1865 quem utilizou pela primeira vez a expressão fator para os componentes hereditários parentais responsáveis pelas características nas progênies. Só mais tarde (1908), Johannsen sugeriu o termo gene para designar os fatores hereditários.
Por gene entende-se a unidade de herança. Contudo, os diferentes textos de genética apresentam diferentes conceitos para gene. Segundo a maioria dos autores, o principal atributo do gene é sua relação com a proteína que codifica. Neste caso, define-se gene como sendo um segmento de DNA, que através da intermediação de uma molécula mensageira de RNA, é responsável pela especificação de uma cadeia peptídica (Wallace, 1992). Entretanto, outros geneticistas incluem, além das proteínas, os RNAs como produtos gênicos transcritos, mas não traduzidos. Neste caso, a definição de gene é um segmento de DNA responsável pela produção de um produto difusível (Lewin, 1994). Como um significativo grupo de RNAs exerce funções outras que a de mensageiro, como por exemplo, a regulação gênica, o segundo conceito de gene é mais realista.
Dentre os vários RNAs descobertos neste súltimos 20 anos, o RNAi, denominado de RNA interferência, da clase dos micros RNAs (miRNA) tem um papel fundamental da regulação gênica.
Por se tratar de uma seqüência de DNA, um gene pode ocorrer sob mais que uma alternativa ou alelo. Desta forma, basta uma alteração na seqüência de bases que cause uma mudança no produto, para que se configure uma alternativa (alelo) diferente. Para simplicidade, normalmente utiliza-se um modelo básico de um gene com dois possíveis alelos, já que a maioria dos seres vivos é diplóide, portanto, carregam dois alelos (um em cada cromossomo homólogo) para o mesmo gene. Mas na realidade, um gene pode ter muitas alternativas. Evidentemente que num indivíduo diplóide só ocorrem uma ou duas formas no máximo. Mas diferentes indivíduos podem apresentar formas alélicas diferentes uns dos outros. Um dos exemplos mais conhecido trata-se do tipo sanguíneo, sendo que numa população de indivíduos podem ser encontrados quatro diferentes alelos.
Sequenciamento de ácidos nucleicos
O sequenciamento consiste na identificação ordenada dos nucleotídeos que compõem um fragmento de DNA ou RNA. Existem duas técnicas que são utilizadas normalmente em laboratórios. Por outro lado, nos últimos anos foram desenvolvidos equipamentos sequenciadores de alta velocidade e que estão sendo utilizados no sequenciamento de espécies procariotas (bactécias) e eucariotas (fungos, vegetais e animais, incluindo Homo sapiens).
Conhecer a sequência de bases dos genomas das espécies tem sido um dos objetivos dos biólogos. A sequência completa de vários vírus já é conhecida há bastante tempo, devido ao fato do pequeno número de nucleotídeos participantes de seus genomas. Em 1995 foi finalizado o sequenciamento do genoma das duas primeiras bactérias pelo 'The
9
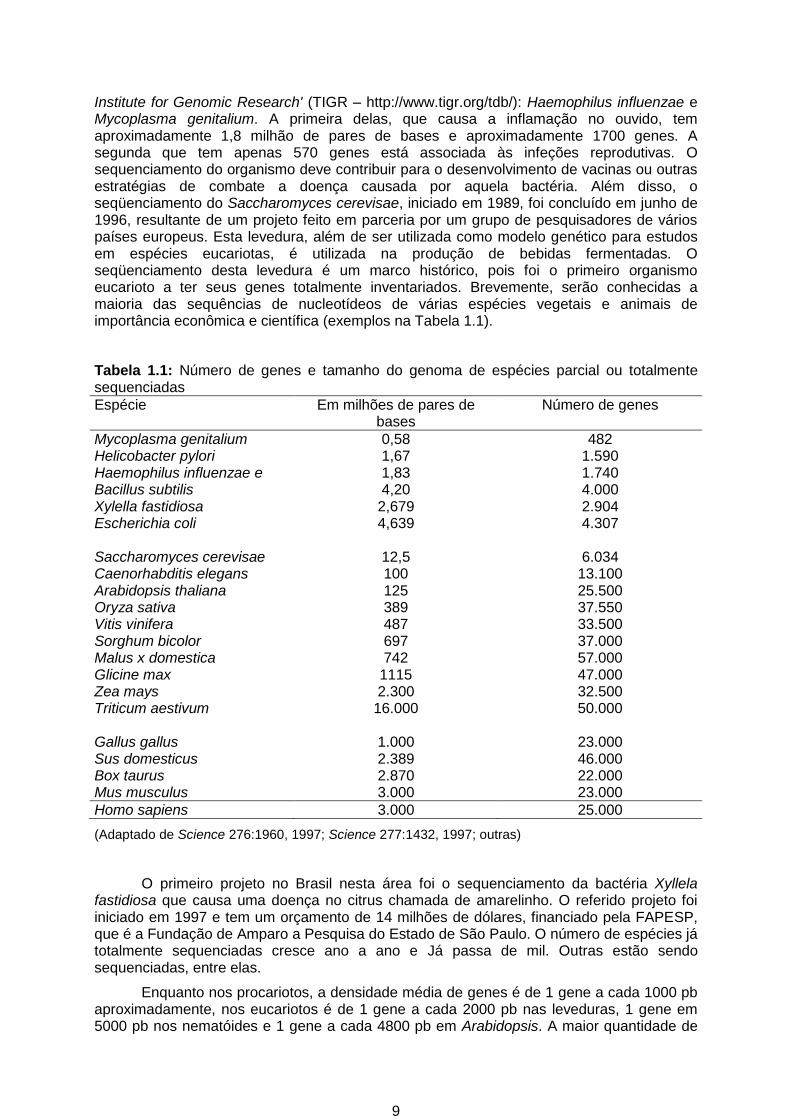
Institute for Genomic Research' (TIGR – http://www.tigr.org/tdb/): Haemophilus influenzae e Mycoplasma genitalium. A primeira delas, que causa a inflamação no ouvido, tem aproximadamente 1,8 milhão de pares de bases e aproximadamente 1700 genes. A segunda que tem apenas 570 genes está associada às infeções reprodutivas. O sequenciamento do organismo deve contribuir para o desenvolvimento de vacinas ou outras estratégias de combate a doença causada por aquela bactéria. Além disso, o seqüenciamento do Saccharomyces cerevisae, iniciado em 1989, foi concluído em junho de 1996, resultante de um projeto feito em parceria por um grupo de pesquisadores de vários países europeus. Esta levedura, além de ser utilizada como modelo genético para estudos em espécies eucariotas, é utilizada na produção de bebidas fermentadas. O seqüenciamento desta levedura é um marco histórico, pois foi o primeiro organismo eucarioto a ter seus genes totalmente inventariados. Brevemente, serão conhecidas a maioria das sequências de nucleotídeos de várias espécies vegetais e animais de importância econômica e científica (exemplos na Tabela 1.1).
Tabela 1.1: Número de genes e tamanho do genoma de espécies parcial ou totalmente sequenciadas
Espécie Em milhões de pares de bases
Número de genes
Mycoplasma genitalium 0,58 482 Helicobacter pylori 1,67 1.590 Haemophilus influenzae e 1,83 1.740 Bacillus subtilis 4,20 4.000 Xylella fastidiosa 2,679 2.904 Escherichia coli 4,639 4.307 Saccharomyces cerevisae 12,5 6.034 Caenorhabditis elegans 100 13.100 Arabidopsis thaliana 125 25.500 Oryza sativa 389 37.550 Vitis vinifera 487 33.500 Sorghum bicolor 697 37.000 Malus x domestica 742 57.000 Glicine max 1115 47.000 Zea mays 2.300 32.500 Triticum aestivum 16.000 50.000 Gallus gallus 1.000 23.000 Sus domesticus 2.389 46.000 Box taurus 2.870 22.000 Mus musculus 3.000 23.000
Homo sapiens 3.000 25.000
(Adaptado de Science 276:1960, 1997; Science 277:1432, 1997; outras)
O primeiro projeto no Brasil nesta área foi o sequenciamento da bactéria Xyllela fastidiosa que causa uma doença no citrus chamada de amarelinho. O referido projeto foi iniciado em 1997 e tem um orçamento de 14 milhões de dólares, financiado pela FAPESP, que é a Fundação de Amparo a Pesquisa do Estado de São Paulo. O número de espécies já totalmente sequenciadas cresce ano a ano e Já passa de mil. Outras estão sendo sequenciadas, entre elas.
Enquanto nos procariotos, a densidade média de genes é de 1 gene a cada 1000 pb aproximadamente, nos eucariotos é de 1 gene a cada 2000 pb nas leveduras, 1 gene em 5000 pb nos nematóides e 1 gene a cada 4800 pb em Arabidopsis. A maior quantidade de

10
DNA pode ser parcialmente explicada pelo fato de que, nos eucariotos a parte regulatória dos genes é muito maior que nos procariotos. Além disso, nos eucariotos existem sequências repetidas, que são ausentes nos procariotos. Embora se saiba o número de genes dos organismos sequenciados, ainda não se conhece as funções de 40 a 60% dos genes, dependendo da espécie. O conhecimento da sequência de bases de um genoma permite aos biólogos o entendimento do funcionamento dos organismos, as funções dos genes, que tipo, tamanho, quantidade e características das proteínas formadas. A maior parte das espécies de bactérias já sequenciadas causam doenças à espécie humana. A razão principal para se conhecer sua sequência relaciona-se com a possibilidade do seu controle, via desenvolvimento de vacinas ou outros medicamentos. As plantas são a base da vida na terra. Contudo, pouco se conhece de seu genoma. O genoma das angiospermas é altamente variável, mas ainda praticamente desconhecido. Desconhecemos também o número de espécies e o número de genes para a maioria das espécie. Na verdade, ainda não é conhecido o número de cromossomos de mais de 70% das espécies vegetais. O valor C de DNA só é conhecido em 1% das espécies. Desta forma, o projeto genoma é de fundamental importância para o aprofundamento do conhecimento das plantas, domesticadas ou não.
Muitos cientistas têm afirmado que o seqüenciamento completo do genoma humano (estimado em três bilhões de pares de bases) deverá revolucionar a medicina e poderá auxiliar na cura para as mais de 3000 doenças hereditárias que atingem a raça humana. Iniciado em 1985, o seqüenciamento do genoma humano que reúne cientistas e laboratórios dos Estados Unidos, Canadá, Japão, Inglaterra, França, Rússia, Itália, Austrália e Brasil entre outros, foi completado antes da data prevista (2005). Quando pronto, o arquivo necessário ao armazenamento das informações se torna equivalente a 200 listas telefônicas com mil páginas cada uma. O GenBank (USA) e o DNA Database (Japão) já dispõem de informações de mapeamento e sequenciamento de mais de 2500 diferentes organismos. Mapas físico e de ligação foram divulgados (com resolução elevada) nos anos de 1993 e 1994 por cientistas franceses e americanos. Tais mapas facilitarão a clonagem de genes humanos, como aqueles envolvidos com as doenças, a obesidade, entre outras.
Especificamente Arabidopsis thaliana, que hoje se constitui no organismo experimental para isolamento e clonagem de genes de plantas, está totalmente sequenciado (Theologis et al., 2000). Esta planta contém aproximadamente 125 milhões de pares de bases, portanto, o seu genoma é relativamente pequeno, se comparado com o das outras espécies vegetais, o que facilita o seu estudo. O número de genes é estimado em 26.000.
O sequenciamento do genoma de Arabidopsis foi feito por muitos cientistas e estudantes em laboratórios ao redor do mundo, vinculado ao Arabidopsis Genome Initiative
O desenvolvimento de densos mapas de ligação genética e o sequenciamento de parte do genoma de outras plantas cultivadas facilitará a identificação e isolamento de importantes genes. O genoma do milho, soja, arroz e outras plantas também já estão praticamente sequenciados.
Professores e estudantes do Laboratório de Fisiologia do Desenvolvimento e Genética Vegetal fizeram parte do Consórcio Genopar que sequenciou o microrganismo Herbaspirillum seropedicae Cepa Z78 (Pedrosa et al., 2011). Esta bactéria que tem aproximadamente 5.513.887 de pares de bases, está envolvida no processo de fixação do nitrogênio atmosférico em algumas espécies de gramíneas, como a cana-de-açúcar.

11
Figura 1.5. Fotografias de plantas de arabidopsis na natura eou em alboratório.
IlustraçãoFonte: http://pt.wikipedia.org/wiki/Ficheiro:Arabidopsis_thaliana_habito.jpg
Introns e exons
Foi descoberto nos anos 70 a presença de seqüências presentes no DNA mas não no RNA mensageiro, produto da transcrição do DNA. Tais seqüências foram denominadas de introns (intervening sequences) e estão intercaladas com os exons (expressed sequences), que são as regiões codificadoras dos genes. A remoção dos introns é feita por enzimas e faz parte do processamento que sofre o pré-mRNA antes de sair do núcleo (Figura 1.6). A presença de introns ou sequências intervenientes sugere uma maior oportunidade para recombinações e maior acúmulo de mutações. Introns são comuns nos eucariotos e raramente encontrados nos procariotos. Quando é o intron que faz o processamento, ele se regenera no final do processo. Neste caso, o intron seria uma enzima, proporcionando ao RNA a função de catálise. Nas bactérias ainda não foram detectados introns. Uma das hipóteses é de que as bactérias perderam os introns durante a evolução. Neste caso os introns teriam se originado no início da vida. Outra hipótese admite que os introns surgiram com os eucariotos. Na realidade, ainda não se sabe exatamente como os introns surgiram, nem tampouco se apareceram logo no início da vida ou surgiram mais recentemente.
Embora tenham características similares, os introns são muito diversos quanto ao tamanho, processamento e funções. Certos introns, em especial os do chamado grupo I, comuns em genomas de organelas celulares (como a mitocôndria) e em alguns genes do núcleo (como o rRNA), apresentam características especiais. Eles próprios realizam sua remoção do pré-mRNA (autocatálise) e ligam os exons, fenômeno denominado self-splicing.
Alguns introns desse grupo são elementos móveis (transposons), capazes de se transferir em cruzamentos genéticos para alelos que não os continham pelo processo denominado homing, iniciado com o corte do DNA por uma endonuclease, enzima codificada pelo próprio intron. Outros introns do grupo I codificam cofatores protéicos, como

12
as maturases. São poucos os casos conhecidos em que um mesmo produto desse tipo de intron realiza ambas as funções -- de endonuclease e de maturase.
Além do processamento, distintas combinações de introns podem ocorrer, resultando na fromação de diferentes peptideos a partir de um mesmo gene.
Já são conhecidos casos de transferência de introns do grupo I entre indivíduos da mesma espécie (transferência vertical). Nesse caso, um intron passa de um alelo para outro que não o continha. Também já foi constatado que introns desse grupo presentes no genoma das mitocôndrias podem passar de uma espécie para outra (transferência horizontal, ou lateral), mas dentro do mesmo filo.
A transferência lateral, entre organismos que não se acasalam sexualmente, foi objeto de profundo estudo de Yangrae Cho e colaboradores, publicado em novembro de 1998. O estudo envolveu um intron do grupo I do genoma mitocondrial de plantas vasculares, bastante conhecido e localizado no gene cox1 da erva Peperomia polybotrya, que teria sido adquirido de um fungo, por transferência lateral. Analisando o DNA de 335 plantas de diferentes gêneros, os autores verificaram que esse intron está amplamente disseminado nos genes cox1 das angiospermas.
O intron estudado está presente em 48 gêneros diferentes, a partir de 32 eventos independentes de transferência lateral. A conclusão sobre as transferências baseia-se em três pontos principais: a presença constante do gene cox1 e esporádica do intron, a incongruência entre as filogenias (histórias evolucionárias) das espécies e dos introns e a co-conversão (Co-conversão é quando parte das extremidades de um segmento de DNA – 3 a 18 pb -, após o processo de recombinação/reparo, é convertida à sequência do DNA doador ou invasor. Assim, o DNA da espécie recipiente é parcialmente degradado e uma nova sequência é sintetizada com base no molde do DNA da espécie doadora. Desta forma, a conversão deixa um rastro, pois a sequência original é alterada.) das seqüências próximas do local de inserção do intron. O primeiro ponto indica que o gene cox1 se disseminou com alta freqüência e manteve-se nas espécies que o receberam, enquanto o intron foi perdido
Figura 1.6: Representação esquemática da estrutura de um gene eucarioto, contendo exons e introns. No rocesso de splicing do RNA, os introns são retirados, ao mesmo tempo que um cap e uma cauda de adeninas são adicionados ao mRNA.

13
na maioria dos casos. O segundo demonstra que a transferência independe do grau de parentesco entre as diferentes plantas. E o último -- a divergência genética das regiões que flanqueiam a inserção do intron -- revela que a transferência se dá via recombinação/reparo e é catalisada por uma endonuclease. Esse processo, conhecido como homing, é exatamente o que esse tipo de intron promove.
Os resultados geram várias preocupações. Entre as dúvidas principais estão a causa da extraordinária invasão desse intron, os passos do processo de transferência em nível celular e o caminho evolutivo da dispersão do intron do grupo I do gene cox1 entre as angiospermas. Entre as implicações, a mais importante está ligada à freqüência com que o DNA é transferido de uma espécie a outra. A transmissão planta a planta requer acasalamento sexual ou a ajuda de vetores (vírus, bactérias, insetos e outros). A questão é bastante atual, já que muitas plantas transgênicas estão sendo liberadas para cultivo.
O trabalho de Cho e colegas (1998) demonstra claramente que a transferência horizontal ocorre e é mais freqüente do que se imagina. Isso torna imperativo estudar o fluxo gênico entre plantas transgênicas e espécies afins, antes de sua liberação para cultivo, para testar a possibilidade de uma irradiação de genes, que podem ser desejáveis em uma espécie, mas completamente indesejáveis em outras. A probabilidade desta irradiação aumenta com o aumento do cultivo destas plantas, principalmente no sistema de monocultura. Num dado momento, um mesmo gene poderá estar presente em milhões de plantas, aumentando o risco da transferência horizontal.
1.2.2-Ácido ribonucleico - RNA
Apesar de ser também um ácido nucleíco, os RNAs têm muitas diferenças em relação ao DNA. Em primeiro lugar, todos os RNAs são formados por apenas uma fita. Entretanto, pode apresentar uma configuração denominada de secundária, quando ocorre o pareamento entre bases complementares. Ao invés de desoxirribose como no DNA, o açúcar do RNA é uma ribose (uma oxidrila a mais em relação a desoxirribose do DNA). A terceira principal diferença é a presença de uracil (U) ao invés de timina (T). Podem ocorrer pelo menos quatro tipos de RNA: mRNA (1-3%), rRNA (>90%), tRNA (1-2%) e sRNA (?%), denominados de mensageiro, ribossomal, transportador e small RNAs, respectivamente. Cada um deles desempenha funções específicas. Dentro do último grupo, são incluídos um grande grupo de RNAs, muitos dos quais ainda sem função conhecida. Outros estão envolvidos na regulação gênica.
Além das funções de mensageiro entre o DNA e os ribossomos, formador dos ribossomos, e transportador de aminoácidos, os RNAs podem ainda desempenhar a função de catálise e de regulação gênica. A função de catálise (até então exclusividade das proteínas) foi descoberta na década passada e os RNAs que têm esta habilidade, as ribozimas, realizam a separação do RNA transcrito em várias partes, fenômeno que se chama de splicing. O autoprocessamento do RNA não é idêntico à catálise enzimática executada pelas proteínas. Numa reação enzimática, a proteína se envolve, mas é liberada intacta ao final do processo. No caso do autoprocessamento, o pré-RNA se processa a si próprio, sem a presença de enzimas, mas não se regenera no fim do processo. Portanto, o pré-RNA não é uma enzima, mas tem a propriedade de catálise. Além disso, foi verificado experimentalmente que o RNA tem a capacidade de retirar bases de um segmento de RNA e adicioná-las em outro, demonstrando a capacidade de sintetizar algo semelhante a si próprio.
mRNA
Resultam da transcrição de um gene. São os RNA mensageiros (mRNA), aqueles que serão decodificados pelos ribossomos e contém informações para a produção de uma proteína. O tamanho dos mRNAs é variável, dependendo do número de bases contidas no gene transcrito. Como contém uma mensagem, diz-se que existe uma colinearidade entre

14
as bases do mRNA e a sequência de aminoácidos da proteína resultante de sua decodificação. O tempo de vida de um mRNA é muito pequeno. Na maioria dos procariotos a meia vida de um mRNA não ultrapassa 2 minutos. Já nos eucariotos, alguns mRNAs duram algumas horas.
rRNA
O RNA ribossomal (rRNA) também é resultante da transcrição de genes de uma região do DNA, neste caso denominada de rDNA. O produto da transcrição não é decodificada, pois os próprios RNAs produzidos juntamente com proteínas vão formar os ribossomos e executar a função específica, que é a produção de proteínas. Participam da formação do ribossomo de um procarioto três rRNAs: o 5S rRNA com 120 nucleotídeos, o 16S rRNA com 1542 nucleotídeos e o 23S rRNA com 2904 nucleotídeos. Nos eucariotos, estes rRNAs são um pouco maiores e designados de o 5S rRNA, o 18S rRNA e o 28S rRNA. Entretanto, nem todos os eucariotos têm os rRNAs do mesmo tamanho.
tRNA
Denominada de adaptadores por Francis Crick, o tRNA (RNA transportador) é um RNA que tem a função específica de transportar os aminoácidos até o ribossomo durante a síntese de uma proteína. São moléculas relativamente pequenas, contendo de 73 a 93 nucleotídeos. Dos ácidos nucleicos conhecidos, o tRNA é o único que apresenta algumas bases que não A, C, G e T. Numa célula existem pelo menos tantos tRNAs quanto são os aminoácidos, e estes estão ligados ao tRNA na extremidade 3'OH. A estrutura tridimensional de um tRNA assemelha-se a uma folha de trevo, contendo numa das alças (loop ou hairpin) o anticodon, que é uma seqüência de três bases.
Outros RNAs
Além dos RNA acima mencionados, existem outros RNAs, muitos deles transcritos e que permanecem no núcleo da célula sem função aparente. Os ncRNAs (non-coding RNA) são pequenas moléculas de RNA que não codificam proteínas funcionais. Os small nuclear RNAs (snRNAs) estão envolvidos no processo de splicing de mRNA (retirda dos introns ejunçao dos exons de um gene). Já os small nucleolar RNAs (snoRNAs) que estão envolvidos no processo de splicing de rRNA. Outras formas de RNA também são conhecidas, como o circRNA (circular RNA), mas as suas funções reguladoras e vias metabólicas ainda não estão totalmente elucidadas.
Os ncRNAs regulatórios estão envolvidos em diversos processos biológicos. Estes ncRNAs podem ser divididos, de acordo com o seu comprimento, em pequenos e longos ncRNAs (lncRNAs). Os ncRNAs pequenos possuem comprimento menor que 200 nucleotídeos e incluem microRNAs (miRNAs), short interfering RNA (siRNA), trans-acting RNA (tasiRNA) e piwi-interacting RNA (piRNA), enquanto os lncRNAs possuem comprimento maior que 200 nt, variando até 100 kb. Os miRNAs fazem parte de processos biológicos cruciais, como resposta à estresse biológico, desenvolvimento e comportamento celular miRNAs constituem uma família de elementos reguladores de expressão gênica, com tamanho variando de 18 a 26 nt, e que controlam diversos processos celulares em organismos eucarióticos. (Ramesh et al., 2014) Em animais, a maioria dos miRNAs são processados de longos transcritos em forma de hairpin através de consecutivas ações de enzimas membros da família da RNA III, DROSHA e DICER, enquanto em plantas somente a enzima DICER é responsável pelo processamento de miRNAs. A maioria das plantas possuem mais de 100 genes de miRNAs (chamados de MIR), localizados quase que exclusivamente em regiões intergênicas do genoma. Já siRNAs e piRNAs são produzidos por vias diferentes às dos miRNAs, estão envolvidos no silenciamento gênico de transposons e sequências repetitivas.

15
Os siRNAs são gerados de RNAs de dupla fita que por sua vez tem distontas origens, tais como RNAs transcritos de regiões invertidas, pares de transcritos em cis-antisenso, pela ação de RNA-dependent RNA polymerases (RDRs) que convertem RNA de fita simples em RNA dupla fita (dsRNA), entre outros. O dsRNA é clivado em siRNAs curtos (21 – 24 nt) pela ação de proteínas DCLs, as quais definem o tamanho do siRNA de acordo com a sua atividade catalítica. De forma similar aos miRNAs, os siRNAs também são incorporados em complexos RISC, podendo interferir na regulação gênica a níveis pós-transcricionais ou transcricionais (por meio de vias RNA-directed DNA Methylation- RdDM).
A interferência mediada por RNAi é um mecanismo natural que ocorre nos organismos eucariotos e exerce o papel na eliminação de RNAs mensageiros anômalos e na defesa do organismo contra parasitas moleculares como transposons e vírus. A interferência de RNA (ou RNAi) leva ao silenciamento estável de genes específicos e é herdável. Estes siRNAs podem destruir os RNAs menssageiros. Também podem ser amplificados. A presença pode ser devido ao possível combate de invasores (ex: vírus) e outras sequências (ex: transposons). O RNAi (miRNAS ou siRNAS) podem ser transmitidos para células-filhas ou outras células mais distantes.
1.2.3-Ácido peptídeo nucléico (PNA)
Esta nova molécula, criada em 1991 em laboratório, têm as quatro bases nitrogenadas do DNA ou RNA ligadas ao esqueleto de uma proteína. Este novo composto sintético além de ser mais estável nas células que o DNA e o RNA, se liga naturalmente a estes com uma intensidade 50 a 100 vezes mais forte que os próprios ácidos nucleicos naturais o fazem entre si. Quando se liga ao DNA, forma uma estrutura de três fitas. Isto permite o uso destas moléculas na terapia gênica, pois pode provocar a indisponibilidade daquela região genômica ser acessada por enzimas e proteínas. Neste caso, poderia ser utilizado um PNA para se ligar a um gene defeituoso que, então, deixaria de expressar uma proteína defeituosa. Os PNAs podem procurar e se ligar a outra fita com seqüência complementar de bases, estratégia similar ao antisenso.
O PNA é construído ligando-se cada base nitrogenada a um peptídeo ao invés de um açúcar e um grupo fosfato. Como a cadeia de peptídeos tem carga elétrica neutra, os PNAs apresentam uma grande capacidade de ligação, eliminando a repulsão criada pela carga elétrica negativa devido a presença dos grupos fosfatos presentes no DNA e RNA. Além disso, os PNAs podem atacar genes invadindo a dupla hélice, algo que DNA e RNA não conseguem. Mais ainda, a química de peptídeos é simples e mais barata que sintetizar ácidos nucleicos.
Este produto da biotecnologia poderá ser aplicado na saúde humana. O principal argumento da utilização dos PNAs em diagnóstico decorre do fato da grande afinidade com o alvo; quanto maior a afinidade, maior é a possibilidade de ligação com seqüências específicas e consequentemente, a sua marcação. Mas como a molécula é artificial, ainda não se conhece ainda a sua toxicidade.
1.2.4-Ácidos nucleicos e a origem da vida
Como é capaz de armazenar o código genético em alguns vírus, tem a função catalítica e de regulação gênica, o RNA passou a ser admitido (hipótese) como a provável molécula que poderia ter originado a vida a partir do 'caldo primitivo'. Esta teoria tem recebido contribuições científicas por uma grande quantidade de cientistas do mundo inteiro. Duplicando RNAs semelhantes como os RNAs ribossomais e participando da produção das proteínas, o RNA é um forte candidato a ser a estrutura do primeiro ser vivo na face da Terra. A função catalítica, entendida aqui como sendo a capacidade de quebrar e ligar outros RNAs, já foi comprovada. Há também resultados de pesquisa que atribuem ao RNA a capacidade de editoração, um sistema simplificado do sistema de reparo do DNA. Os vírus que possuem RNA como material genético necessitam da enzima transcriptase reversa para

16
produzir DNA e então se replicarem. Quando se provar que o RNA tem ou teve capacidade de autoduplicação, será dado um passo importante favorável a hipótese do 'Mundo do RNA'. Nenhuma outra molécula teria a capacidade e a versatilidade de desempenhar tantas funções como o RNA no 'caldo primitivo'. Outra hipótese considera uma molécula mais simples, precursora do RNA, composta de um ácido nucléico ligado a peptídeos (denominada de PNA).
Alguns cientistas não concordam com estas hipóteses por considerarem que as moléculas de RNA são muito complexas para ter tido origem no ambiente primitivo terrestre, onde só havia água, gás carbônico, nitrogênio e radiação ultravioleta. Além disso, no 'caldo primitivo' deveriam existir substâncias muitos tóxicas. Em contrapartida, admitem que sob as condições primitivas, a estrutura cristalográfica dos minerais seria capaz de reduzir dióxido de carbono para formar aldeídos e a partir destes se formariam açúcares e moléculas orgânicas essenciais. A transferência de elétrons de uma molécula outra poderia ter contribuído para as transformações metabólicas. Recentemente, cientistas obtiveram moléculas de RNA mais complexas quando utilizaram uma mistura de pequenas moléculas de RNA sob condições de altas temperaturas, situação que deve ter ocorrido na época do surgimento da vida.
Outra possibilidade da origem da vida seria via metabólitos secundários. Tais metabólitos, considerados secundários no atual estágio evolutivo, teriam sido relevantes no período pré-biótico como integrantes do metabolismo primário responsável pela síntese dos ácidos núcleicos e tradução e replicação.
De qualquer forma, a hipótese de maior consenso é a de que o RNA teria sido o primeiro material genético sobre o qual a evolução agiu, resultando numa quantidade enorme de formas de vida que se conhecem atualmente.
2-REPLICAÇÃO (Replication)
O DNA funciona como um modelo para a síntese de novas fitas de DNA de maneira semiconservativa, ou seja, cada uma das duas moléculas filhas tem uma fita da molécula mãe e outra recém sintetizada. A replicação ocorre bidirecionalmente a partir de uma (procariotos) ou várias (eucariotos) origens. A replicação é precisa (alta fidelidade), ou seja, a maioria dos erros é corrigida. Cabe a replicação o desafio maior de perpetuar, com alta fidelidade, um genoma e ao mesmo tempo permitir erros que originam a variabilidade necessária para a evolução.
A origem de replicação é uma região do DNA que contém uma seqüência de bases específica. Nas bactérias só existe uma destas seqüências. A rigor, a replicação completa do cromossomo de uma bactéria depende da iniciação nesta seqüência. Neste caso, é dito que as bactérias têm apenas um replicon. Replicon é a unidade de DNA no qual a replicação ocorre a partir de uma origem. Já os eucariotos, por terem genomas bem maiores que as bactérias e mais de um cromossomo, têm várias origens de replicação. Nas leveduras (ex: Saccharomyces cerevisiae) existem pelo menos umas 500 origens de replicação, denominadas de ARS (Autonomously Replicating Sequences); ou seja, 500 replicons. Na Drosophila melanogaster existem cerca de 3.500 replicons. Já na Vicia faba estima-se a presença de pelo menos 35.000 replicons. As origens de replicação dos eucariotos são ativadas em diferentes tempos durante o período de replicação do ciclo celular (fase S da mitose). Estas origens de replicação estão espaçadas em média de 50 a 100 kb. A velocidade de replicação em Escherichia coli, a bactéria residente no intestino de todas as pessoas, chega alcançar 50.000 bases por minuto. Nos eucariotos, o movimento do garfo de replicação é pelo menos 10 vezes mais lento.
Os vírus apresentam um modo de replicação específico denominado de círculo rolando (rolling circle). Uma vez iniciada a replicação, o genoma circular vai sendo replicado indefinidamente. Posteriormente uma enzima produzida pelo próprio genoma viral, corta a

17
longa cadeia produzida em partes iguais, cada uma contendo uma cópia do genoma do vírus, a ser subseqüentemente encapsulada.
Mais de 20 enzimas atuam diretamente no processo de replicação das bactérias. As principais proteínas envolvidas e sua função na replicação são apresentadas abaixo: toposisomerases - desenovelam o DNA helicases - separam as duas fitas Single strand binding proteins (SSB) - protegem o DNA na forma de fita simples Primase - adiciona os primers ou iniciadores DNA polimerase III - polimeriza, i.é., adiciona os dNTP no sentido 5'-3' DNA polimerase I - substitui os iniciadores de RNA por bases do DNA; também tem a função de reparo ligase - une os dNTP de dois fragmentos.
Nos procariotos, além destas duas polimerases, existe uma terceira, a DNA polimerase II, cuja função ainda é desconhecida. Das três, somente a DNA Pol I apresenta a função de edição, ou seja, de correção dos possíveis erros de replicação. A DNA Pol I é formada por várias subunidades. O agrupamento de algumas delas forma o que se conhece por fragmento Klenow, utilizado para replicação do DNA in vitro. Este fragmento não tem a habilidade de edição como a enzima completa, pode ser comprado de vários fornecedores e é usado em laboratórios. A DNA Pol III é formada por sete subunidades ou polipetídeos.
Nos eucariotos também existem três polimerases. Duas delas atuam no núcleo,
sendo que a DNA Pol teria a mesma função que a DNA pol III dos procariotos. A DNA
Pol ß teria a função de reparo. A terceira polimerase (DNA Pol ) é específica para a replicação do genoma das mitocôndrias.
A replicação dos genomas dos retrovírus, que são codificados por RNA, é feita pela transcritpase reversa (RT), o que pode produzir inúmeros variantes. O conhecimento da natureza molecular destes vírus permite a criação de estratégias para combatê-los. Moléculas ribozimas de RNA foram engenheiradas e podem ser introduzidas nos hospedeiros para procurar e destruir o genoma do HIV, cortando-os em dois.
O avanço no conhecimento científico sobre a replicação foi de fundamental importância no desenvolvimento da reação da polimerização em cadeia (PCR), uma das técnicas moleculares mais utilizadas no momento.
3-TRANSCRIÇÃO (Transcription)
Transcrição é o processo pelo qual uma região do DNA é transcrita resultando num RNA. Existem dois grandes grupos de RNAs: (i) os RNA mensageiros (mRNA), aqueles que serão decodificados pelos ribossomos e contém informações para a produção de uma proteína e (ii) o outro grupo de RNAs, formado pela transcrição de determinadas regiões genômicas e que permanecem como RNA para executar uma função específica. Entre eles estão o transportador (tRNA), o ribossomal (rRNA) que juntamente com proteínas forma os ribossomos e outros RNAs (snRNA, hnRNA, etc.) com função na regulação gênica ou desconhecida. A região (segmento) do DNA transcrita é a parte estrutural do gene.
A transcrição nos procariotos é feita pela RNA polimerase. Numa E. coli podem existir até 3.000 cópias dela. Esta enzima usa o DNA como molde e sintetiza uma cadeia de nucleotídeos de RNA complementar ao molde. Aparentemente não há conferência do produto transcrito. Se no DNA estão A, C, G e T, vai aparecer no mRNA U, G, C e A, respectivamente. A exemplo da replicação, a transcrição ocorre na direção 5'-3'.
Seis peptídeos ou sub-unidades fazem parte da RNA pol (ß'ß2). A rigor o fator tem a habilidade de reconhecer o promotor, que é a região 5', situada imediatamente anterior ao início da parte codificadora (ou estrutural) do gene. Posteriormente, juntam-se ao

18
fator s os demais peptídeos quando então a RNA Pol inicia o processo de transcrição. Vários fatores de transcrição (pequenos polipeptídeos), os TFs, atuam no início, durante a elongação e no término da transcrição.
O fator (é de fundamental importância. Quando um vírus entra numa célula hospedeira, um fator (do vírus é transcrito e agora os outros cinco peptídeos da RNA Pol ficam a disposição do fator ( do vírus, que reconhece tão somente os genes do vírus. Desta forma, em pouco tempo os vírus conseguem expressar seus genes no hospedeiro e se replicando a uma velocidade impressionante, atingem milhões de cópias. Afetam drasticamente o organismo hospedeiro porque também reprimem a produção de proteínas deste.
O promotor das bactérias é formado por duas seqüências localizadas nas posições -10 e -35 (região 5') da base codificante +1 do gene. Nestas regiões, normalmente são encontradas as seqüências (consenso) TATAAT (denominada de TATA box ou Pribnow box) e TTGACA (CAAT box), respectivamente. Nos eucariotos, a região regulatória dos genes é bem mais complexa. Em alguns casos, podem ser encontrados vários elementos que controlam ou afetam a transcrição. Entre eles estão o promotor, o enhancer e elementos como o GLE, o MRE, etc. Os enhancers são seqüências de DNA que estão muito distantes dos genes e são compostas de seqüências muitas vezes repetidas. Os elementos são sequências de DNA, que são alvos de ligação para proteínas especificas, que constituem o que se chama de fatores de transcrição (TF). Os fatores de transcrição podem aumentar dramaticamente a taxa de transcrição de um gene nos organismos eucariotos. Além do promotor, outras regiões podem acelerar a taxa de transcrição como os enhancers e os terminadores. Os terminadores são seqüências que a RNA Pol identifica como o fim da região de DNA codificadora ou de um gene.
Existem algumas diferenças entre eucariotos e procariotos com relação a transcrição. Em primeiro lugar existem três RNA polimerases ao invés de uma. A RNA Pol I só transcreve o rDNA (sequência de DNA que codifica o rRNA). A RNA pol II transcreve os genes que codificam para proteínas, produzindo então mRNAs. Os demais RNAs (tRNA, snRNA e a 5 S rRNA) são transcritos pelo RNA pol III. Nos procariotos, os ribossomos identificam os mRNAs porque estes apresentam uma seqüência denominada de Shine-Dalgarno que é incluída antes das bases codificadoras, complementar a uma região do componente 16 S rRNA. Por sua vez os mRNAs dos eucariotos apresentam uma estrutura denominada de quepe (Cap) resultante de uma ligação 5'-5' entre duas guaninas ou entre G e A. Após a transcrição, ao mRNA é adicionado uma longa cauda de adeninas, o que se convencionou denominar de poli-A. Esta característica dos eucariotos permite a separação dos mRNAs dos demais RNAs, o que normalmente pode ser feito em laboratório. Nos procariotos, a cauda de adenina é bem reduzida. Uma quarta diferença entre procariotos e eucariotos relaciona-se com o processamento do pré-mRNA nas células eucariotas. Nestas, após a transcrição, são removidos os introns do pré-mRNA. Só então, este RNA se desloca para o citoplasma e recebe a denominação de mRNA.
4-TRADUÇÃO (Translation)
Tradução é o processo de decodificação do mRNA nos ribossomos resultando na formação de um peptídeo. Na maioria dos casos as proteínas são formadas por apenas um peptídeo. Para a produção de um peptídeo in vitro são necessários o mRNA, os ribossomos, os tRNAs, os amino ácidos, fatores da tradução e energia.
Os ribossomos dos procariotos são formados por duas subunidades: a grande, chamada de 50 S, é constituída por dois rRNAs, o 23 S rRNA e o 5 S rRNA, e por 34 proteínas; a pequena, chamada de 30 S, é constituída pela unidade 16 S rRNA e por 21 proteínas. Dependendo da fase, uma bactéria pode ter aproximadamente 5.000 ribossomos, o que representa 25% da massa celular.

19
Os tRNAs são os RNAs transportadores, também chamados de adaptadores, que transportam os amino ácidos do meio até os ribossomos para serem incorporados à cadeia peptídica. Uma enzima, é encarregada de carregar o amino ácido específico na extremidade 3'OH do tRNA, com base no seu anticodon. Existem mais de 20 tRNAs, pois na maioria dos casos, mais de um codon codifica para um mesmo amino ácido.
O processo de tradução (5'-3') inicia quando a sub-unidade pequena do ribossomo reconhece a seqüência líder do mRNA. Em seguida o primeiro codon (um conjunto de 3 bases) é lido e geralmente codifica para metionina. Um tRNA traz o amino ácido correspondente ao codon lido. Sucessivamente os codons vão sendo lidos e os amino ácidos correspondentes incorporados ao peptídeo nascente pela enzima peptidil transferase. A velocidade da tradução chega a 40 amino ácidos por segundo. Qualquer um dos codons de terminação UAG, UAA ou UGA, significa o fim do peptídeo, cuja interpretação é feita pelos ribossomos. Nos procariotos, algumas mensagens são policistrônicas.
Nos procariotos a tradução é simultânea à transcrição. Mais ainda, um mesmo mRNA pode ser traduzido por dezenas de ribossomos enfileirados, o que resulta num número elevado de cópias repetidas de uma proteína a partir de uma única molécula mensageira.
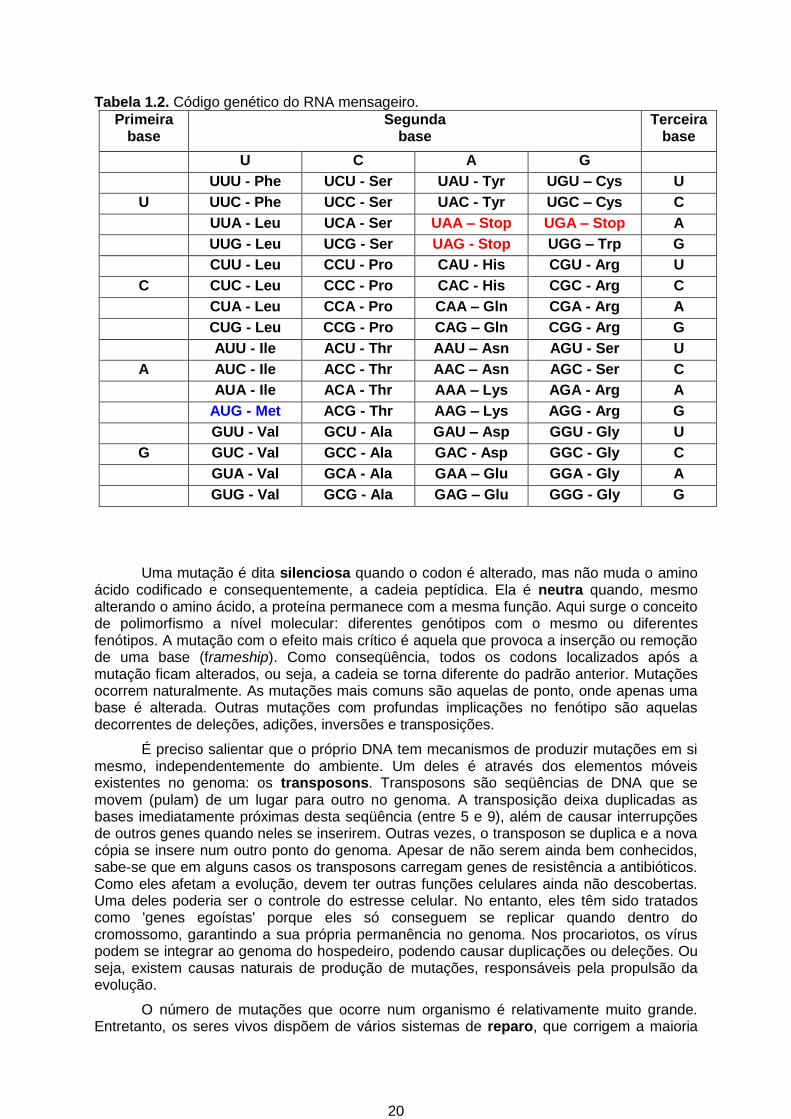
O código genético (Tabela 1.2) está estruturado em codons (trincas), cada um com três bases. A probabilidade de associar três bases independentemente da ordem e natureza é de 64. Três codons são de terminação. Os outros 61 codificam os 20 amino ácidos. Consequentemente, um mesmo amino ácido pode ser codificado por mais de um codon. As principais características do código genético são:
- estruturado em trinca de bases
- não há sobreposição (uma base pertence a um e somente um codon)
- universal (reforça a teoria da origem única da vida); somente poucas diferenças com o código genético das mitocôndrias
- degenerativo (mais de um codon codificam para um mesmo amino ácido)
- o primeiro codon (das proteínas) é AUG ou GUG
- há diferença ou preferência de uso de diferentes codons de um mesmo amino ácido
- a hipótese de Wobble permite a não ocorrência dos 61 tRNAs.
O conhecimento do funcionamento desta fábrica permitiu a compreensão da ação dos antibióticos e o desenvolvimento de remédios para várias doenças. Geralmente os antibióticos se ligam ao rRNA ou às proteínas dos ribossomos, impedindo ou a leitura do mRNA, ou o emparelhamento do tRNA com o ribossomo ou impedindo outra atividade nos ribossomos. Como os ribossomos dos procariotos são diferentes dos eucariotos, um antibiótico pode afetar o funcionamento da sub-unidade pequena (30 S) de uma bactéria, sem contudo interferir no ribossomo da célula eucariota hospedeira, cujas sub-unidades tem rRNAs de diferentes tamanhos e seqüência.
5-MUTAÇÃO E REPARO
Mutação é uma modificação no DNA. Mutante é o fenótipo resultante da mutação. As mutações são causadas por erros de replicação do DNA e alterações do DNA por deleção, duplicação ou rearranjamentos causados por vírus, transposons, ação enzimática ou processos físicos e químicos. A taxa média de mutação que ocorre naturalmente atinge 1x10-7. Agentes químicos e físicos (radiações) são utilizados em laboratório para aumentar esta taxa.
20
Tabela 1.2. Código genético do RNA mensageiro.
Primeira base
Segunda base
Terceira base
U C A G
UUU - Phe UCU - Ser UAU - Tyr UGU – Cys U
U UUC - Phe UCC - Ser UAC - Tyr UGC – Cys C
UUA - Leu UCA - Ser UAA – Stop UGA – Stop A
UUG - Leu UCG - Ser UAG - Stop UGG – Trp G
CUU - Leu CCU - Pro CAU - His CGU - Arg U
C CUC - Leu CCC - Pro CAC - His CGC - Arg C
CUA - Leu CCA - Pro CAA – Gln CGA - Arg A
CUG - Leu CCG - Pro CAG – Gln CGG - Arg G
AUU - Ile ACU - Thr AAU – Asn AGU - Ser U
A AUC - Ile ACC - Thr AAC – Asn AGC - Ser C
AUA - Ile ACA - Thr AAA – Lys AGA - Arg A
AUG - Met ACG - Thr AAG – Lys AGG - Arg G
GUU - Val GCU - Ala GAU – Asp GGU - Gly U
G GUC - Val GCC - Ala GAC - Asp GGC - Gly C
GUA - Val GCA - Ala GAA – Glu GGA - Gly A
GUG - Val GCG - Ala GAG – Glu GGG - Gly G
Uma mutação é dita silenciosa quando o codon é alterado, mas não muda o amino ácido codificado e consequentemente, a cadeia peptídica. Ela é neutra quando, mesmo alterando o amino ácido, a proteína permanece com a mesma função. Aqui surge o conceito de polimorfismo a nível molecular: diferentes genótipos com o mesmo ou diferentes fenótipos. A mutação com o efeito mais crítico é aquela que provoca a inserção ou remoção de uma base (frameship). Como conseqüência, todos os codons localizados após a mutação ficam alterados, ou seja, a cadeia se torna diferente do padrão anterior. Mutações ocorrem naturalmente. As mutações mais comuns são aquelas de ponto, onde apenas uma base é alterada. Outras mutações com profundas implicações no fenótipo são aquelas decorrentes de deleções, adições, inversões e transposições.
É preciso salientar que o próprio DNA tem mecanismos de produzir mutações em si mesmo, independentemente do ambiente. Um deles é através dos elementos móveis existentes no genoma: os transposons. Transposons são seqüências de DNA que se movem (pulam) de um lugar para outro no genoma. A transposição deixa duplicadas as bases imediatamente próximas desta seqüência (entre 5 e 9), além de causar interrupções de outros genes quando neles se inserirem. Outras vezes, o transposon se duplica e a nova cópia se insere num outro ponto do genoma. Apesar de não serem ainda bem conhecidos, sabe-se que em alguns casos os transposons carregam genes de resistência a antibióticos. Como eles afetam a evolução, devem ter outras funções celulares ainda não descobertas. Uma deles poderia ser o controle do estresse celular. No entanto, eles têm sido tratados como 'genes egoístas' porque eles só conseguem se replicar quando dentro do cromossomo, garantindo a sua própria permanência no genoma. Nos procariotos, os vírus podem se integrar ao genoma do hospedeiro, podendo causar duplicações ou deleções. Ou seja, existem causas naturais de produção de mutações, responsáveis pela propulsão da evolução.
O número de mutações que ocorre num organismo é relativamente muito grande. Entretanto, os seres vivos dispõem de vários sistemas de reparo, que corrigem a maioria

21
dos erros ocorridos. Outros erros, quando não corrigidos, podem causar enormes problemas tanto na sobrevivência como na reprodução do organismo. Neste caso atua a seleção natural, ou eliminado este indivíduo ou fazendo com que ele deixe um menor número de descendentes. O acúmulo de mutações em diferentes populações pode provocar, a longo prazo (prazo em termos de evolução), a diminuição da freqüência de cruzamentos com o conseqüente início da especiação, processo que pode culminar com a origem de uma nova espécie.
Ao nível de laboratório, os agentes químicos mais utilizados para induzir mutações são: etil metil sulfanato (EMS), ácido nitroso, etil metano e alguns agrotóxicos ou defensivos. A ação dos agentes químicos normalmente produz alteração de uma base qualquer. Exemplo: substituição de A por T. Muitos vegetais contêm substâncias que causam mutações na espécie humana. Ex: nas frutas e legumes são encontradas as psoraleínas (o limão contém quantidades elevadas), que também dimerizam duas timinas, se ocorrem lado a lado. Entre os agentes físicos, os mais usados são as radiações (UV, gama, etc.). Os agentes físicos geralmente causam quebras e rearranjos de cromossomos. Especificamente a radiação UV causa a dimerização de duas timinas se estiverem lado a lado. Durante a replicação, a DNA Pol não consegue ler este dímero, o que provoca a inserção de duas bases quaisquer no lugar das timinas, se não houver reparo. Muitos problemas de pele são causados pela radiação UV. É por isto que existe tanta preocupação com a diminuição da camada de ozônio, pois este atua como uma barreira aos raios UV.
A mutagênese direcionada permite a alteração de uma ou mais bases de uma seqüência de DNA qualquer. Inicialmente a seqüência de interesse é inserida num vetor, como o vírus M13 que é de fita simples. Posteriormente é feito um primer (iniciador) num sintetizador de oligonucleotídeos. Este primer é complementar a um certo segmento da seqüência de interesse, mas contendo uma base diferente. Posteriormente, o restante da molécula é duplicado. Resultado: a nova seqüência difere da original por uma base apenas. Esta seqüência pode ser avaliada in vitro ou in vivo. Pela técnica da recombinação homóloga, esta seqüência mutante pode substituir a seqüência normal de um organismo. Desta forma, é avaliado o efeito de uma mutação in vivo.
Foi desenvolvido por Ames, um teste para avaliar a capacidade mutagênica dos produtos químicos utilizados, com base no tipo de mutação que os produtos provocam. Tais produtos químicos são classificados quanto ao potencial de causar danos nas pessoas, dependendo do tipo de mutação e a freqüência que são causadas. Este teste associa a capacidade de ação mutagênica com a capacidade de causar câncer, pois estas duas estão estreitamente relacionadas. Outros tipos de testes também são utilizados para confirmar a periculosidade do produto. Com base nestes testes, a fabricação e a comercialização de muitos produtos químicos já foram proibidas.
Exemplo de uma mutação que causa a anemia falciforme (Figura 1.7).
Seqüência normal: atggtgcacctgactcctgtggagaagtctgccgttactgccctgtggggcaaggtgaacgtggatgaagttggtggtgaggccc tgggcaggttggtatcaaggttacaagacaggtttaaggagaccaatagaaactgggcatgt Seqüência contendo a mutação: atggtgcacctgactcctgaggagaagtctgccgttactgccctgtggggcaaggtgaacgtggatgaagttggtggtgaggccctgggcaggttggtatcaaggttacaagacaggtttaaggagaccaatagaaactgggcatgt O códon gtg na sequência normal codifica para o ácido glutâmico, que é substituído pela valina na cadeia proteína, em razão da muatção de t para a, formando um novo codon gag. O individuo portador desta mutação apresenta a anemia falciforme e, simultaneamente, resistência a malária, ilustrada na Figura 6 abaixo

22
Figura 1.7. Glóbulos em forma de meia lua, caracterizando a anemia falciforme.
6-METILAÇÃO
Uma fração das citosinas no DNA de muitos organismos torna-se metilada (5mC) após a replicação. Esta metilação não tem distribuição ao acaso. Algumas seqüências como as denominadas ilhas de CpG em animais, são raramente ou não metiladas. Enquanto algumas seqüências são metiladas em certas condições, como aquelas herdadas da mãe e não do pai, outras são sempre metiladas em todos os tecidos.
Nas plantas e fungos as ilhas CpG são freqüentemente metiladas pelas metilases, embora há evidência de uma substancial quantidade delas não metiladas. Em fungos, a metilação atinge 1,5% das Citosinas e não ocorre somente de forma simétrica.
Tanto o controle da metilação quanto sua função nos eucariotos, ainda não são suficientemente compreendidos. A metilação tem sido correlacionada com redução na atividade gênica, havendo evidências de inibição da expressão de vários genes. Em ratos, a redução da metilação do DNA em 70%, resultante da mutação no gene metiltransferase do DNA, leva a morte os indivíduos na embriogênese. A hipótese levantada admite que as regiões com bases metiladas dificilmente são transcritas. Neste caso, a morte dos ratos poderia ter sido provocada pela falta de proteínas e/ou RNAs. A metilação também é requerida para o comportamento normal dos cromossomos em Neurospora crassa. Sua necessidade foi comprovada, mas sua função ainda não está totalmente esclarecida.
7-REGULAÇÃO GÊNICA
Na definição de Jacob e Monod (1961), gene é uma seqüência de DNA que codifica para um produto difusível. A região regulatória do gene é uma seqüência de DNA que não é convertida em outra forma (como a região codificadora) e que só funciona in situ. Além disso, existem genes estruturais e genes reguladores de outros genes.
O princípio básico da regulação gênica é a interação entre proteínas regulatórias e certas regiões (seqüências) do DNA. Assim, nos procariotos a regulação gênica é chamada de negativa se um gene não se expressa caso o repressor, que é uma proteína, liga-se ao DNA na região do promotor do gene (Figura 1.8). Para que o gene possa ser transcrito, há a necessidade de remover a proteína repressora. Isto é possível, pela presença do indutor, para o qual a proteína repressora tem muito mais afinidade que pela região do DNA responsável pela regulação do gene. O indutor então tem um efeito inativador sobre o repressor. Este tipo de regulação gênica é o mais comum nos genes de organismos procariotos. No controle dito positivo, o mais frequente nos eucariotos, o gene é ativado pela presença de um ativador. Em outras palavras, no controle negativo, a interação proteína-DNA desliga o gene, enquanto no controle positivo, a interação liga o gene.

23
O controle negativo é bastante comum nas bactérias, onde a maioria dos genes estaria ligada (on) até que os repressores os desligariam (off). Já o sistema positivo é mais comum nos eucariotos, onde os genes estariam desligados até que os ativadores os ligariam.
A rigor, existem cinco pontos de controle na regulação de um gene eucarioto: 1) na ativação de gene estrutural, 2) no início da transcrição, 3) no processamento da transcrição, 4) no transporte para o citoplasma e 5) na tradução do mRNA. Na ativação de um gene estrutural, um gene é regulado por uma seqüência no promotor e/ou no enhancer, as quais são reconhecidas por proteínas específicas. Esta proteína funciona como um fator de transcrição necessário para o início da transcrição através da RNA Pol. Proteína ativa só é disponível sob condições quando o gene é para ser expresso. In vitro é possível modular a regulação nos diversos pontos de controle. In vivo, a adição de determinados genes permitem o controle de um ou mais pontos de controle.
Nos eucariotos ainda não se conhece profundamente a regulação gênica. Entretanto, vários mecanismos já foram amplamente estudados. Em primeiro lugar, um grande número de genes é ativado em determinados tecidos e órgãos e não em outros. Os genes denominados de Homeobox são os responsáveis por este controle. Já nas primeiras divisões celulares do zigoto formado, os genes Homeobox se encarregam de marcar quais os genes que poderão e quais os genes que não poderão ser expressos num determinado tecido ou órgão. Outros genes dependem de um complexo sistema de eventos: sinal ambiental (temperatura, umidade, etc.) faz com que uma substância seja produzida e/ou movida para as células. Este sinal químico seria recebido por um receptor na célula, cujo complexo tem habilidade para penetrar no núcleo da célula e ativar um conjunto de genes de forma coordenada.
8-EPIGENÉTICA
Uma das incógnitas da diferenciação no desenvolvimento de organismos é como o sistema de expressão gênica e de herança produzem diferentes tecidos ou órgãos a se as células têm a mesma informação genética. Os avanços científicos culminaram com o desenvolvimento de um segundo código genético, a epigenética. Epigenética é o estudo de mudanças herdáveis na expressão e função dos genes que não podem ser explicado por alterações na sequência de DNA (exemplo na Figura 1.9). As mudanças epigenéticas são baseadas num conjunto de processos moleculares que podem ativar, reduzir ou eliminar completamente a atividade de um determinado gene:
• (i) metilação da citosina no DNA,
Figura 1.8: Modelo de funcionamento do operon lac em bactérias. O repressor impede a transcrição dos genes Z, Y e A, que é ativada na presença de β-galactosídio.

24
• (ii) remodelamento da estrutura da cromatina por modificação química, em particular acetilação ou metilação de histonas e
• (iii) processos regulatórios mediados por moléculas pequenas de RNA (ex: siRNA).
Sistemas de herança epigenéticos - Com poucas exceções, as diferenças entre células especializadas são epigenéticas e não genéticas. Estas células não só mantém o fenótipo por longos períodos, mas também o transmitem para as células-filhas. Esta transmissão de informação é denominada de sistemas de herança epigenéticos. Seria a segunda dimensão da herança e da evolução. A evolução é possível a partir da variação epigenética herdável até mesmo quando não existe nenhuma variação genética.
No livro EVOLUÇÃO EM QUATRO DIMENSÕES - DNA, comportamento e a história da vida, de autoria de Eva Jablonka e Marion J. Lamb, lançado no Brasil em 2010, são discutidas as quatro "dimensões" na evolução - quatro sistemas de herança que desempenham um papel na evolução: a genética, a epigenética (ou transmissão de características celulares, alheia ao DNA), a comportamental e a simbólica (transmissão por meio da linguagem e de outras formas de comunicação simbólica). Em todos esses sistemas ocorre alguma herança de caracteres adquiridos.
Figura 1.9. Relações hipotéticas entre a variação genética, epigenética e fenotípica em populações naturais. São mostrados dois genes para cada um dos dois indivíduos de duas populações. As barras horizontais são o DNA, com diferenças na seqüência de DNA indicado por diferentes tons de cinza. modificações epigenéticas em um determinado gene são indicados por triângulos negros. Variação epigenética natural pode ser encontrada dentro (A1 x B1) ou entre (A2/B2 vs C2/D2) populações. Variação epigenética pode ser independente (A1 x B1) ou confundida com (C1 x D1) a variação genética. Algumas variações epigenéticas em populações naturais podem resultar da plasticidade fenotípica, podendo ser não hereditárias, ou seja, não persistirá em um ambiente comum (C2 versus D2). Se a variação epigenética independente persistir em um ambiente comum (como no A1/B1), esta é uma evidência para a herança epigenética. Esta variação epigenética hereditária é traduzida em diferenças fenotípicas e de valor adaptativo (como ilustrado acima), é ecológica e evolutivamente relevante (Fonte: Bossdorf et al., 2008).

25
PARTE 2 - MARCADORES GENÉTICOS
1-INTRODUÇÃO Marcador genético é uma característica que é capaz de detectar diferenças (polimorfismos genéticos) entre dois ou mais indivíduos ou organismos. Entre suas propriedades um marcador genético deve: (i) ser capaz de diferenciar os progenitores e (ii) ser reproduzido com precisão na progênie.
Do ponto de vista molecular, um marcador genético (ou loco marcador) serve para identificar um local ou uma região de um cromossomo. Um marcador genético ideal deve apresentar uma série de atributos: (i) alto nível de polimorfismo (ii) estabilidade em diferentes ambientes (iii) detectar grande número de locos não ligados (iv) herança mendeliana simples
Entretanto, a simplicidade e os baixos custos do método são fatores determinantes no uso de forma rotineira de um marcador molecular. Aqui será apresentada uma descrição resumida dos principais tipos de marcadores genéticos bem como suas principais aplicações no melhoramento de plantas.
Todo e qualquer fenótipo molecular proveniente de um gene expresso, como no caso de isoenzimas , ou de um segmento específico de DNA (correspondendo a regiões expressas ou não do genoma) é chamado de marcador molecular.
2-MARCADORES MORFOLÓGICOS
Até os meados da década de 60, os marcadores utilizados em estudos de genética e melhoramento eram controlados por genes associados a caracteres morfológicos, Em geral, características fenotípicas de variação discreta são utilizadas como marcadores morfológicos desde os tempos de Mendel, como fenótipos de fácil identificação visual (Ex.: nanismo, deficiência clorofítica, cor de pétala ou morfologia foliar). Um número variável de marcadores morfológicos existe para as diferentes espécies de plantas, contudo insuficientes para mapeamento genético ou outras aplicações. Além disso, esses marcadores freqüentemente são afetados pela ação gênica de dominância, efeito ambiental, pleiotropia e epistasia. O reduzido número e a natureza dos marcadores morfológicos restringiram os estudos dos caracteres quantitativos (QTs) às espécies onde havia sido alcançada uma caracterização genética substancial. Sax (1923) verificou em feijão que as diferenças nas médias do peso de grãos estavam associadas a cor das sementes. Foi a primeira tentativa de caracterização individual dos locos (QTL) envolvidos na expressão de um caráter quantitativo (QT) com auxílio de marcadores morfológicos.
Marcadores morfológicos apresentam a desvantagem de serem somente identificados em sua maioria, na planta inteira ou adulta demandando de bastante tempo e esforço na parte de campo.
3-MARCADOR DE PROTEÍNAS DE SEMENTES
As proteínas das sementes podem ser classificadas de acordo com a sua solubilidade em quatro diferentes grupos. Numerosos métodos têm sido utilizados in vitro para caracterizar as proteínas de sementes. Polipeptídeos variantes que apresentam distintos pesos moleculares podem ser separados em gel de poliacrilamida (Figura 2.1) através do processo de eletroforese (ver Quadro 2.1 e Figura 2.2)). A eletroforese de duas dimensões (SDS-PAGE) tem habilidade de separar proteínas pelo ponto isoelétrico (carga) e pelo peso molecular (tamanho). Diferentes variantes aparecem como distintas bandas

26
num gel. Embora o número de variantes de uma proteína (polimorfismo) seja relativamente alto, o número de proteínas de sementes que podem ser analisados é baixo. Apesar da base genética complexa (normalmente são famílias de genes) a interpretação é relativamente simples (Observar foto abaixo, Guimarães et al., 2002).
QUADRO 2.1: ELETROFORESE O termo eletroforese (eletro: carga elétrica; forese: deriva) foi criado por Michaelis em 1909, para descrever migração de colóides sob a influência de um campo elétrico. Seu princípio é simples: moléculas de carga negativa migram para o pólo positivo, e moléculas com carga positiva migram para o pólo negativo. A eletroforese visa a separação de moléculas em função de suas cargas elétricas, de seus pesos moleculares e de suas conformações, em suportes porosos (géis) e soluções - tampões (estabilizam o pH do meio e permitem o fluxo de corrente elétrica). Ou seja, na prática a eletroforese consiste da extração de amostras, seja de proteínas, RNA ou DNA obtido de um tecido e da migração destas num gel (amido, agarose, acrilamida) submetido a uma corrente elétrica contínua. O sentido e a velocidade de migração são determinados pelo tamanho e carga das proteínas. Por exemplo, quanto maior a carga elétrica de uma proteína, mais rápido a sua migração no gel em direção ao eletrodo de carga contrária, como é observado na figura 1. A passagem de corrente elétrica através de uma solução-tampão segue a Lei de Ohm:
V = R. I onde, V = voltagem R= resistência I = amperagem
A eletroforese pode ser conduzida ora sob voltagem, ora sob amperagem (corrente) ou, então, wattagem (potência) constantes reguladas pela fonte elétrica. É bom observar que para cada tipo de marcador a ser utilizado diferencia grandemente na corrente elétrica a ser utilizada. A eletroforese pode ser desenvolvida em suportes como papel-filtro, sílica gel, membranas de acetato de celulose e géis de agarose, de amido ou de poliacrilamida. Para enzimas, géis de amido e poliacrilamida oferecem melhor separação do que outros suportes. Para marcadores DNA os mais utilizados são géis de agarose e poliacrilamida.
Figura 2.1: Perfil eletroforético de proteínas extraídas pelo calor em sementes de cafeeiros nos estágios de desenvolvimento verde (A), verde-cana (B) e cereja (C), com diferentes tratamentos de secagem.

27
Quadro 1: Continuação
A B
Figura 2.2b: Exemplos de aparatos de eletroforese. A) Cuba de eletroforese horizontal submersa para gel de agarose. B) Cuba de eletroforese vertical para gel de acrilamida.
Figura 2.2a: Princípios gerais do sistema de eletroforese.

28
4-ISOENZIMAS
Na década de 1960, um novo tipo de marcador genético foi desenvolvido: as isoenzimas, então denominados de marcadores bioquímicos. Isoenzimas foram definidas como diferentes formas moleculares (variantes) de uma mesma enzima, apresentando função idêntica ou similar, presente num mesmo indivíduo (Markert & Moller, 1959). É o resultado da presença de mais de um gene codificando cada uma das enzimas.
As vantagens sobre os marcadores morfológicos são a insensibilidade à pleiotropria e à epistasia, além de sua natureza co-dominante (possibilita a identificação de indivíduos homozigotos e heterozigotos). Desde a sua resolução pelos métodos histoquímicos, a principal aplicação das isoenzimas é nos estudos de diversidade genética e evolução,o que têm sido extremamente importantes para as investigações sobre variação intraespecífica, genética de populações, também na evolução e mapeamento genético, já realizadas em centenas de espécies. Apesar de estar sendo utilizada em vários programas de melhoramento, o reduzido número de sistemas enzimáticos polimórficos impõe limitações variáveis dependendo do objetivo do estudo ou atividade.
Comumente muitas enzimas existem em múltiplas formas moleculares, mas apresentando a mesma especificidade. O princípio básico da técnica reside no uso de eletroforese em gel de amido ou poliacrilamida e na visualização do produto enzimático por métodos histoquímicos (Hunter e Market, 1957). As distintas bandas observadas no gel, representam diferentes formas moleculares que apresentam diferentes propriedades de mobilidade eletroforética. Subsequentemente, a posição de uma enzima no gel de amido pode ser verificada pela sua atividade que é detectada por um sistema de revelação colorimétrica. Este sistema inclui reagentes específicos para revelar uma determinada enzima. A conseqüência é o aparecimento de uma ou mais bandas no gel. Portanto, as distintas formas de uma mesma enzima, as isoenzimas, codificadas por diferentes alelos, podem ser detectadas em diferentes regiões do gel, caso apresentem diferentes mobilidades eletroforéticas. Com esta técnica o estudo da variabilidade genética de populações de uma dada espécie será baseada na variação observada nas isoenzimas. Cada banda revelada no gel se constitui num marcador genético, já que por marcador genético entende-se a constituição genotípica de um loco num determinado indivíduo. As isoenzimas começaram a ser utilizadas como marcadores genéticos somente a partir de 1966 (Lewontin & Hubby, 1966).
4.1-Vantagens das isoenzimas em relação aos marcadores morfológicos: a) determinação genotípica dos locos em qualquer parte da planta, b) ocorrência de um número razoável de alelos e ausência de alelos nulos; c) ausência de efeitos deletérios associados com alelos isoenzímicos, d) herança Mendeliana simples com codominância entre alelos na maioria dos locos, e) ausência de efeitos epistáticos, pleiotrópicos e ambientais.
4.2-Aplicabilidade das isoenzimas:
A propriedade mais expressiva é a base genética simples envolvida na expressão destas enzimas (Soltis & Soltis, 1989), o que torna a identificação de polimorfismos rápida e simples (Brewer, 1970). A maioria das enzimas já reveladas em gel de amido tem mais de uma isoenzima. Como consequência, uma grande quantidade de sistemas isoenzimáticos são potencialmente informativos. A eletroforese de enzimas tem proporcionado dados úteis na abordagem de questões importantes em sistemática e evolução de plantas. Do ponto de vista da variação intraespecífica, as isoenzimas têm contribuído para o estudo da organização da variabilidade genética e a identificação de raças (Singh et al., 1991).
Além da caracterização da diversidade genética de populações naturais e genótipos cultivados, as isoenzimas têm sido utilizadas com bastante freqüência em outros estudos.

29
Ligação genética entre sistemas enzimáticos ou destes com outros locos tem aumentado a resolução de mapas genéticos em várias espécies. As isoenzimas também têm sido utilizadas na identificação de genes que controlam caracteres quantitativos em feijão, milho, soja e tomate.
4.3-Base genética dos marcadores isoenzimáticos
A premissa básica de se utilizar dados enzimáticos é que diferenças na mobilidade de isoenzimas em um campo elétrico são resultantes de diferenças nas seqüências de DNA que codificam tais enzimas. Assim, se os padrões de bandas de dois indivíduos diferem, assume-se que estas diferenças possuem base genética e sejam herdáveis. O controle genético de isoenzimas ocorre através de vários genes, que podem ser alelos de um mesmo loco, ou estar situados em diferentes locos.
Isoenzimas codificadas por genes alélicos são também chamados de aloenzimas. A expressão das isoenzimas é co-dominante, isto é, em um indivíduo diplóide ambos os alelos de um loco são expresso e visualizados, ou seja, discrimina o heterozigoto do homozigoto.
5-RFLPs (Restriction Fragment Length Polymorphism)
As variações nos nucleotídeos do DNA devido à mutação, deleção, inserção e inversão, podem ser detectadas se ocorrerem num sítio de corte das enzimas de restrição. Se o DNA de plantas diferindo num ou vários desses nucleotídeos forem expostos a essas enzimas, fragmentos de diferentes tamanhos, portanto polimórficos, são gerados e podem ser identificados e clonados. Tais fragmentos são denominados de RFLPs ('Restriction Fragment Length Polymorphims'; polimórfismo no comprimento de fragmentos restrição) e foram desenvolvidos por Botstein et al. (1980). Os polimorfismos de comprimento de fragmentos de restrição ou polimorfismo de tamanho de fragmento são locos no DNA que podem ser identificados e mapeados. Os RFLPs têm sido suficientemente numerosos na maioria dos cruzamentos e têm permitido uma cobertura adequada do genoma, proporcionando a construção de densos mapas genéticos de ligação, que possibilitam a realização de análises genéticas e moleculares e várias aplicações no melhoramento de plantas, como clonagem de genes e mapeamento de QTLs (Nodari et al., 1993). O elevado custo e o tempo necessário na geração destes marcadores restringem drasticamente seu uso de forma frequente, principalmente em países como o Brasil.
A obtenção de RFLPs envolve várias etapas. Em primeiro lugar é preciso extrair e purificar o DNA de um indivíduo. Após, este DNA deve ser digerido (cortado) por enzimas de restrição (ER) que são capazes de reconhecer um pequena seqüência de pares de bases (pb) e então cortar o DNA neste sítio de reconhecimento ou clivagem. Entretanto, a maioria das plantas contém mais de um bilhão de pb. Como consequência, a digestão do DNA de uma planta com apenas uma ER produz milhares de fragmentos que variam em comprimento de acordo com a distribuição dos sítios de clivagem. Tal quantidade impossibilita a análise de todos de uma só vez.
A terceira etapa do processo consiste em separar esta mistura de fragmentos de diferentes comprimentos pela eletroforese em gel de agarose. A migração dos fragmentos de DNA num gel é dependente do seu tamanho, migrando mais rapidamente, os menores. Subsequentemente, os fragmentos de DNA na condição de fita simples (após tratamento com hidróxido de sódio), são transferidos para uma membrana de nylon ou celulose (carregada positivamente), técnica que é denominada de Southern blot, e que proporciona um suporte sólido para o DNA que passa a ser imobilizado neste suporte. Agora é possível analisar individualmente cada um destes fragmentos.
A próxima etapa do RFLP é a hibridização do DNA destas plantas já imobilizados em membranas com uma sonda radioativa de DNA (que pode ser um fragmento de DNA da própria planta, um clone) complementar ao fragmento de interesse. Para que haja

30
hibridização, há a necessidade que pelo menos parte da sonda seja complementar ao fragmento de interesse. Existem alternativas de marcação de sondas que não a radioativa.
A última etapa, a autoradiografia, consiste da exposição da membrana hibridizada com a sonda radioativa a um filme de Raio X, que é queimado somente onde houve as hibridizações. A sonda sendo radioativa, emite radiação que pode ser detectada por filmes de Raio X. Já que a sonda só hibridiza com fragmentos complementares, a precisão é elevadíssima. Portanto, as cópias únicas (genes) normalmente aparecem uma vez só no genoma, e, portanto apenas uma banda pode ser detectada nos indivíduos homozigotos. Assim, a associação enzima de restrição e sonda identifica um loco RFLP, que tem herança mendeliana.
Admitindo-se que duas plantas diferem em um sítio de reconhecimento, apresentarão fragmentos de diferentes comprimentos, com relação a uma sonda complementar. Tais fragmentos localizam-se em diferentes posições na membrana. Consequentemente apresentarão bandas ocupando diferentes posições no filme, indicando a existência do polimorfismo ao nível de DNA, portanto genotípico. Os fragmentos de diferentes tamanhos são denominados de alelos, e apresentam herança mendeliana. A principal característica da técnica do RFLP é a sua habilidade em detectar tais diferenças.
As seqüências genômicas de duas plantas de uma mesma espécie são muito parecidas. Entretanto, as plantas sofrem freqüentes alterações ao nível de DNA: mutações simples, rearranjamentos e recombinação; as quais podem ocasionalmente alterar a seqüência ou substituir bases nitrogenadas em um ou mais sítios de reconhecimento de uma determinada ER. Numa população, estas variações podem ocorrer numa planta e não em outra. Tais diferenças (que normalmente são denominadas de variação genética) produzem fragmentos de DNA de diferentes tamanhos (polimorfismo de comprimento de fragmento) quando o DNA é exposto a estas enzimas.
Para o desenvolvimento das sondas, o DNA de uma planta precisa ser digerido por uma ER ou quebrado mecanicamente e os fragmentos inseridos em um vetor (geralmente plasmídeo), uma espécie de carregador. Este plasmídeo recombinante pode ser amplificado ilimitadamente, após sua inclusão numa bactéria ou mesmo in vitro. A denominação de sonda ocorre quando uma certa quantidade amplificada deste DNA é marcada com radioisótopos, ou ligada a reagentes que posteriormente podem ser coloridos, portanto identificáveis. As sondas desta forma são utilizadas para detectar seqüências complementares a elas.
Os RFLPs mais informativos são aqueles cuja seqüência ocorre somente uma vez no genoma, denominados de cópia única. Desta forma, os RFLPs são específicos. Como as isoenzimas, os RFLPs nucleares exibem codominância. Pleiotropia e epistasia que afetam a resolução dos marcadores morfológicos, não têm o menor efeito sobre os RFLPs. Além disso, os RFLPs apresentam alta estabilidade. O DNA a ser analisado pode ser extraído de qualquer parte da planta. Outra característica fundamental é a de que a herdabilidade deste tipo de marcadores é virtualmente 1. Isto possibilita a realização da seleção indireta, cuja teoria foi desenvolvida há bastante tempo, mas sua implementação não existiu por falta de marcadores com as características dos RFLPs. Por sua segura informação genotípica e ocorrência em grande número, estes marcadores possibilitam o desenvolvimento de mapas genéticos de ligação altamente saturados. Estes são a ferramenta básica para estudos de genética, evolução e melhoramento de plantas.
6-MINISSATÉLITES
Os minissatélites ou locos VNTR ('Variable Number of Tandem Repeats') são regiões dispersas no genoma que contém um número variável de seqüências repetidas e enfileiradas (tandem) de DNA que têm um núcleo comum de 10 a 15 pares de bases (Jeffreys et al., 1985). Podem ser analisados tanto através de RFLPs ou PCR (reação em cadeia da polimerase, Quadro 2). Muitos dos minissatélites são altamente polimórficos,

31
produzindo um grande número de bandas. Por estarem espalhadas por todo o genoma e apresentarem um número variável de repetições em diferentes indivíduos em relação a uma mesma região cromossômica (loco), os minissatélites simultaneamente proporcionam um conjunto de marcadores genéticos que se constitui no que tem sido denominado de impressões digitais de DNA (DNA fingerprinting), conseqüentemente, indivíduo-específicos.

32
QUADRO 2.2: A REAÇÃO EM CADEIA DA POLIMERASE (PCR) Esta reação foi concebida em 1983 por Kary Mullis (Prêmio Nobel em 1993),
publicada em 1985, mas utilizada de forma rotineira a partir de 1988 (Saiki et al., 1988). Esse método tem a habilidade de amplificar um fragmento de DNA, normalmente de até 4000pb, mas até 30 kb em condições especiais, com a utilização de DNA polimerases de alta eficiência e fidelidade. Para amplificar, o primer (ou iniciador) utilizado, que é um oligonucleotídeo de aproximadamente 10 nucleotídeos, precisa anelar com sequências complementares e invertidas com relação às duas fitas que foram previamente separadas
pelo aumento da temperatura (92-94C). O anelamento entre os primers e as seqüências
complementares é efetuado a uma temperatura de 45 a 55C. Uma Taq DNA polimerase estende (ou sintetiza) as cadeias originadas pelos primers, cuja temperatura ótima de
catálise é de 72C. Existem máquinas programáveis de PCR, os termocicladores, capazes de modificar a temperatura rapidamente. Na realidade cada ciclo da PCR é composto de três
etapas: a separação das fitas ou desnaturação (92-94C), o anelamento ou
hibridização do primers com o DNA (45 a 55C) e a extensão ou polimerização da
cadeia (72C). Os tempos utilizados em cada fase são aproximadamente de 1 min, 1 min e 2 min, respectivamente. A rigor, uma vez atingida as temperaturas de cada fase, são necessários poucos segundos para que a reação ocorra. E as máquinas de PCR têm a capacidade de alterar a temperatura de forma rápida e repetir o ciclo tantas vezes quantas ordenadas. O número de fragmentos amplificados duplica a cada ciclo. Sucessivos ciclos de separação, anelamento e de síntese produzem milhões de fragmentos virtualmente idênticos, em apenas algumas horas. Os produtos da PCR podem ser facilmente visualizados num gel de agarose. Esta visualização é possível com auxílio do brometo de etideo, que quando presente no gel se interpõe entre as duas fitas do DNA e se torna avermelhado com absorção da luz ultravioleta. A técnica da PCR tem dezenas de aplicações. A amplificação de fragmento(s) a partir de primers arbitrários (sequência de bases completamente casualizadas) foi denominada de RAPD. Em plantas, os RAPDs têm facilitado a realização de estudos em genética e melhoramento, até então, considerados inexequíveis com as técnicas tradicionais. Uma diferença entre duas plantas ao nível de DNA que ocorra na região de anelamento do primer é identificada pela ausência da referida banda em uma delas e presença da banda na outra. No caso de indivíduos heterozigotos, estes produzem as mesmas bandas que os homozigotos. De fato, os marcadores RAPDs são dominantes. Combinando DNA de plantas segregantes com uma grande quantidade de sondas, é possível a identificação de dezenas, centenas e mesmo milhares de RFLPs e/ou RAPDs. Quanto mais próximas as diferenças no DNA, maior será o grau de co-segregação entre elas. A análise da segregação destes alelos permite o estabelecimento da relação da ordem e da distância entre eles nos cromossomos, o que pode ser visualizado num mapa genético de ligação.
Na área da saúde, a técnica da PCR está sendo utilizada intensamente (Vosberg, 1989). Uma das aplicações é na diagnose de doenças causada por vírus como Hepatite, AIDS, etc. Nestes casos, utilizam-se os primers que anelam a regiões específicas do DNA ou RNA do vírus causador da doença. Portanto, primers com sequência conhecida e pré-estabelecida. Se houver amplificação de uma banda a partir do DNA de uma pessoa, é porque existe DNA ou RNA do vírus nas células humanas. Este diagnóstico, rápido e confiável, já está sendo feito em várias cidades brasileiras. Existe um esforço integrado entre a Secretaria da Saúde e a UFSC no desenvolvimento deste sistema aqui em Florianópolis.

33
7-RAPDs (Randomly Amplified Polymorphic DNA)
Na década de 80, surgiu um novo tipo de marcador molecular denominado de RAPDs ('Randomly Amplified Polymorphic DNA'; DNA polimórfico amplificado ao acaso; Welsh & McClelland, 1990; Williams et al., 1990). O uso da reação da polimerização em cadeia (PCR) proporciona a amplificação de um segmento de DNA, delimitado por dois iniciadores (ou primers), comumente com 10 pares de bases, que são complementares a dois sítios de nucleotídeos: um em cada fita do DNA, posicionados inversamente a uma distância geralmente não superior a 4kb. Os produtos resultantes da amplificação podem ser visualizados como bandas em géis de agarose ou poliacrilamida. Diferenças ao nível do DNA são inferidas pela presença ou ausência de um determinado fragmento amplificado (banda no gel). Em relação aos RFLPs, os RAPDs são mais baratos, requerem pouco tempo e não necessitam de radioisótopos. Nos últimos anos, alguns mapas desenvolvidos com RFLPs e isoenzimas, se tornaram altamente saturados com RAPDs, como em soja, tomate, milho, feijão, ervilha, amendoim, Arabidopsis e em muitas outras espécies domesticadas ou não. Outros mapas foram desenvolvidos somente com marcadores RAPDs.
Quadro 2.2: continuação O mais fascinante, entretanto, é a amplificação de DNA de espécies extintas fossilizadas ou conservadas na forma de múmia, o que é denominado de DNA ancestral (ancient DNA). Atualmente é possível amplificar segmentos de DNA extraído de ossos e outros tecidos macios, o que tem permitido conhecer seqüências de DNA de vários mamíferos fósseis. Outra maneira de conhecer o DNA dos fósseis ou espécies extintas seria a de decodificar o DNA extraído de insetos sugadores, embebidos em amber a milhões de anos atrás. Amber é a designação dada à resina solidificada de árvores antigas e tem a capacidade de proteção contra água e o ar. Tais insetos podem carregar nas estruturas que usam para sugar ou no aparelho digestivo, o sangue de animais. Estas descobertas auxiliaram a realização do filme Jurassic Park. Em maio de 1995, do interior de uma abelha envolta de amber e que teria vivido há 20-25 milhões de anos atrás, foi isolada uma bactéria que está se reproduzindo normalmente e de cujo DNA, foram amplificados vários fragmentos via PCR. A sequência destes fragmentos mostrou grande similaridade com o DNA da bactéria Bacillus.
Figura 2.3: Padrão de bandas polimórficas (indicadas por setas) e monomórficas de marcador RAPD em Araucaria angustifolia. As bandas são separadas em gel de agarose e visualizadas sob luz ultra-violeta após coloração com brometo de etídeo. (Fonte: Stefenon et al., 2004).

34
O princípio dos RAPDs está igualmente baseado na identificação de diferenças ao nível do DNA. Entretanto a metodologia é totalmente diferente daquela dos RFLPs e minissatélites e se baseia na PCR. Uma desvantagem dos RAPDs é sua natureza dominante (incapacidade de discriminar entre homozigotos e heterozigotos) e também o desconhecimento da localização das marcas no genoma.. As bandas observadas no gel após a eletroforese são codificadas como presentes ou ausentes (1 e 0, respectivamente) em cada indivíduo.
8-MICROSSATÉLITES
Entre as diversas sequências repetidas em tandem, algumas são simples, formadas por um ou poucos nucleotídeos. Tais repetições curtas em tandem são denominadas de microssatélites. Microssatélites, também chamados STR ('Short Tandem Repeat'), SSRP ('Simple Sequence Repeat Polymorphisms') ou STMS ('Sequence Tagged Microsatellite Sites') são sequências repetidas de um, dois, três ou quatro nucleotídeos e que estão espalhadas pelo genoma de um indivíduo. São altamente polimórficos em plantas, animais e microorganismos. Em plantas seria mais fácil utilizar microssatélites GA (ou CT) e GT (ou CA), pois os AT, embora frequentes, causam problemas. Assim, cada região genômica que contenha um determinado número de repetições de uma destas sequências constitui-se num loco genético, altamente variável entre indivíduos e multialélico, portanto, altamente informativo (Ferreira e Grattapaglia, 1995).
Comparativamente aos RFLPs, os microssatélites proporcionam 3 a 4 vezes mais polimorfismo ou informação. Entretanto, para o uso rotineiro dos microssatélites, há a necessidade de primeiro amplificar uma região, posteriormente sequenciá-la e em terceiro lugar, sintetizar os iniciadores específicos para cada loco. Uma vez feito isto, o loco marcador pode ser utilizado indefinidamente naquela espécie. Desta forma, existe um custo elevado e trabalho no início, mas o custo subsequente é baixo e a simplicidade a posteriori, é muito grande. O mapeamento genético e a caracterização varietal para fins de proteção e de germoplasma para fins de conservação de várias espécies está sendo feito com o uso dos marcadores microssatélites. Seu uso está associado principalmente à caracterização varietal para fins de proteção e de conservação germoplasma. O alto polimosfismo e a natureza co-dominante dos marcadores microssatélites permitem sua utilização em estudos de genética populacional e evolução de espécies selvagens, como na caracterização de estrutura genética intra-populacional (Stefenon et al., 2008a) e reconstrução da história demográfica (Stefenon et al., 2008b) do pinheiro brasileiro.

35
Figura 2.4. Exemplo de marcador microssatélite, que detecta diferenças entre dois parentais e na progênie.
9-AFLPs (Amplified Fragment Length Polymorphism)
Os polimorfismos de comprimento de fragmentos amplificados (AFLPs; Zabeau, 1993) é resultante do uso combinado de enzimas de restrição e da reação da polimerização em cadeia. Suas principais características são a alta especificidade e resolução e poder de amostragem. Nos protocolos dos AFLPs constam pelo menos sete etapas importantes: 1) digestão do DNA, 2) ligação dos adaptadores, 3) primeira amplificação, 4) segunda amplificação, 5) preparo do gel, 6) a corrida do gel e 7) o processamento do gel (Figura 2.5).
O DNA e digerido por duas enzimas de restrição, uma que corta sítios de seis pares (corte raro) de base (geralmente a EcoRI) e a outra que corta seqüências de 4 pares de bases (corte freuente)(geralmente a MseI). Este processo de clivagem gera milhões de fragmentos de distintos tamanhos. O DNA utilizado deve ser de alta qualidade. De preferência utilizar um protocolo ou etapa que inclua fenol. A qualidade (quantidade, integridade e ausência de contaminantes) do DNA é a base de todo o processo.
O processo de ligação dos adaptadores envolve o uso de ligases que permite que os fragmentos de DNA que foram cortados se liguem a pequenos oligonucleotídeos de DNA de seqüência conhecida.
Subseqüentemente é feita a primeira amplificação, que consiste na amplificação dos fragmentos agora ligados aos adaptadores através da reação da polimerização em cadeia com o uso de iniciadores, complementares aos adaptadores com uma extra base a mais na extremidade 3‟. Isto é importante, pois somente 25% dos fragmentos serão amplificados (aqueles com a base complementar ao nucleotídeo final da extremidade 3‟ do iniciador), caso contrário todos os fragmentos cortados seriam amplificados e a resolução no gel seria virtualmente impossível. Neste ponto do protocolo é importante verificar se a reação foi bem feita. Para tanto deve-se rodar um gel com parte da reação de amplificação. Dependendo do resultado se continua ou não o processo.
A segunda amplificação é feita com uma pequena amostra da primeira amplificação. Neste caso são utilizados iniciadores que são compostos de todas as bases dos primers da primeira amplificação, mais duas a três bases na extremidade 3‟, dependendo do nível de polimorfismo da espécie ou da população. Caso isto não seja conhecido, há a necessidade de experimentar diferentes combinações de iniciadores. Para os laboratórios que usam radioisótopos, neste quarto passo também é feita simultaneamente a marcação radioativa dos produtos da PCR, para posterior detecção em

36
filme de raio X. Na realidade se marca só um dos iniciadores porque o sinal é suficiente para detecção.
O preparo do gel (geralmente de poli-acrilamida) é uma etapa delicada. A completa limpeza do material, os tipos de molduras, a maneira de colocar as soluções nos moldes, a qualidade dos reagentes, etc., afetam a qualidade do gel. Qualquer defeito no gel pode causar a perda de reação completa. Existem diferentes aparatos para corrida. Nos diferentes laboratórios, há diferentes equipamentos. Todos com suas vantagens e desvantagens.
A corrida do gel envolve o carregamento e a corrida propriamente dita. O carregamento das amostras é um passo crucial. Os cuidados vão desde a limpeza das cavidades no gel, o uso adequado das pipetas, a precisão na liberação das amostras e o acompanhamento na fase inicial da corrida. Como o gel é submetido a alta voltagem, há a
necessidade de acompanhar a temperatura que não pode ultrapassar a 55C, sob pena de desnaturar o sistema e danificar os instrumentos.
A fase final consiste no processamento do gel. Existem basicamente três formas de visualização das bandas. A primeira delas é com nitrato de prata. A segunda envolve a utilização de radioisótopos e a terceira utiliza terminações coloridas. De maneira geral, a maioria dos laboratórios usa o fósforo y-33P radioativo, por vários motivos. Em primeiro lugar, a nitidez dos géis é bastante alta com radioatividade. Em segundo lugar, o filme é um documento importante. O uso dos radioisótopos gamas como o 33P possibilita o seu manuseio sem grandes riscos para as pessoas, uma vez que este tipo de radiação não vai além de alguns centímetros. Outra vantagem deste radioisótopo é que a sua meia vida é maior que a do 32P. A principal desvantagem é que o aparecimento de sinal no filme requer um tempo maior que os outros isótopos. Entretanto, já existem câmaras intensificadoras de sinal, mas cujo preço é muito alto. A outra maneira consiste na utilização de kits comerciais com moléculas fluorescentes (fluoróforos ou dyes) com terminadores coloridos (dyes) e utilizar sequenciadores automáticos. Desta forma, evita-se a radioatividade. A última forma de visualizar as bandas é através da coloração do gel de poli-acrilamida com nitrato de prata. Entre as principais vantagens estão a ausência de radioatividade e o baixo custo. Entretanto, a resolução não é tão boa quanto os outros dois métodos. Empresas químicas já estão anunciando o desenvolvimento de dyes para a utilização direta em géis. Desta forma, por colorimetria será possível visualizar bandas, no futuro, diretamente no gel sem qualquer outro tratamento. Contudo, não sabe-se ainda o preço que custarão tais kits.
A reação de digestão do DNA permite a obtenção de fragmentos grandes, pequenos e uma combinação de grandes de pequenos, respectivamente. Com isto, um grande número de fragmentos podem ser amplificados e resolvidos num só gel. Desta forma, esta estratégia permite que sejam analisadas num único gel o maior número de marcadores comparativamente às outras metodologias. Embora robusto e de alta reproducibilidade, os marcadores AFLPs são dominantes não se distinguindo heterozigotos de um dos homozigotos. As principais restrições deste grupo de marcadores referem-se a necessidade do uso de radioisótopos, da alta qualidade do DNA e da proteção por patente desta tecnologia.
Marcadores AFLP têm sido utilizados na construção de mapas genéticos, estudos de filogenia (Stefenon et al., 2006), genética populacional (Stefenon et al., 2007) e identificação de variação somaclonal em clones de plantas micropropagadas (Steinmacher et al., 2007).

37
10-SCARs (Sequence characterized amplified RAPD)
As etapas principais no desenvolvimento de um SCAR são: 1) identificação de um iniciador que confere polimorfismo a dois bulks de DNA com fenótipos contrastantes, 2) o isolamento e a clonagem do fragmento amplificado em um vetor (plasmideo), 3) sequenciamento do fragmento isolado, 4) desenho dos iniciadores de tamanho maior que os decâmeros e 5) o teste final (Paran e Michelmore, 1993).
Para a identificação de um iniciador que confere polimorfismo a dois bulks contrastantes, é necessário a extração de DNA de plantas da geração F2. Posteriormente, estas plantas F2 ou a sua progênie (F2:3) são testadas com relação a uma característica, resistência a uma raça de uma doença por exemplo. Desta forma, as plantas F2 são agrupadas em duas classes fenotípicas ou alternativamente se for utilizado as plantas F2:3 em três classes fenotípicas. Misturando-se quantidades equimolares de DNA de seis plantas de mesmo fenótipo (ex: resistência), pode-se dizer que os seis genótipos têm uma seqüência de DNA em comum, que é em relação ao gene que confere o referido fenótipo e talvez um conjunto adicional de pares de bases. Da mesma forma se constrói o outro bulk, com base no fenótipo contrastante (susceptibilidade). Desta forma, os dois bulks só são diferentes, genotipicamente com relação a característica analisada. Testando iniciadores que amplificam seqüências arbitrárias de DNA, por pura chance, é possível encontrar iniciadores de 10 pares de bases (decâmeros) capazes de amplificar o DNA de um bulk e não o do outro. Quando se testa este iniciador em todos os DNAs das demais plantas F2 e a seqüência realmente está ligada, ou seja, quando todas (ou a maioria) das plantas resistentes apresentam a banda e todas ou uma minoria das plantas susceptíveis não
Figura 2.5: Etapas da geração de marcadores AFLP.

38
apresentam a banda, conclui-se que o segmento amplificado está ligado ao gene de interesse. Pela quantidade de recombinação entre o local do anelamento do iniciador e o fenótipo das plantas pode-se estimar a distância entre o marcador e o gene de interesse. O Ideal é que o marcador deve estar o mais próximo possível do gene, para que possa ser utilizado como critério de seleção.
O segundo passo é o isolamento e a clonagem do fragmento amplificado em um vetor, geralmente um plasmídeo. Posteriormente, os plasmídeos contendo os fragmentos de DNA desejados são utilizados para transformar bactérias. Das colônias transformadas é preciso separar as que contêm o fragmento daquelas que não contém o fragmento de DNA desejado. Posteriormente deve se crescer as colônias selecionadas e extrair o DNA do plasmídeo. Como o DNA vai para sequenciamento, há a necessidade de alta pureza. Existem vários métodos e kits comerciais disponíveis para clonar este fragmento. O melhor é purificação em kits com colunas de sílica. O melhor seria a purificação com cloreto de césio, mas o método é trabalhoso. Após a obtenção do DNA plasmidial, deve verificar se os plamídios contêm o fragmento desejado. Então digere-se com uma enzima de restrição capaz de cortar o plamídio em sítios que flanqueiam o inserto. Corre-se um gel e verificam-se quais os plamídios com insertos.
O terceiro passo é o sequenciamento do fragmento isolado. O sequenciamento é necessário para se conhecer a sequência do fragmento, ou seja, as bases que estão entre os iniciadores. De posse da sequência, se desenham os iniciadores (quarto passo) com comprimento variável entre 16 e 24 pares de bases. A idéia de um iniciador mais comprido surgiu de cálculos feitos sobre o comprimento mínimo de um iniciador capaz de amplificar uma seqüência única num genoma da maioria das plantas. Desta forma, espera-se a presença de uma única banda com o uso dos referidos iniciadores. Existem critérios que são levados em consideração no desenho de iniciadores: a inclusão do decâmero que originou a banda, uma percentagem mínima de 50% de C e G, tamanho mínimo que proporciona uma temperatura de anelamento maior que 56 C, a terminação em C ou G e a possibilidade de formação de diemres de inicaidores e estruturas secundárias (hairpin ou loopback). Existem programas de computador que auxiliam a tomada de decisão, já que proporcionam valiosas informações comparativas a respeito de diferentes iniciadores que são gerados quando é fornecida ao programa uma determinada seqüência de bases.
Finalmente, de posse nos iniciadores, se fazem os testes incluindo-se tanto os bulks como também um certo número de amostras da população F2 e de outras plantas da mesma espécie.
11-SNPs (Single Nucleotide Polymorphism)
Diferenças em um único nucleotídeo em um ponto particular do genoma são chamadas polimorfismo de simples nucleotídeo (single nucleotide polymorphism ou SNP). Esse tipo de polimorfismo ocorre aproximadamente uma vez a cada 1000 bases no genoma humano. SNPs são caracterizados através do sequenciamento de fragmentos de DNA, sendo detectados principalmente por PCR qualitativa (qPCR). Nas quatro seqüências hipotéticas abaixo existem dois SNPs, um na seqüência 3 e outro na seqüência 4.
SEQUÊNCIA CONCENSO: A C T T T G A C C A A A T T G
SEQUÊNCIA 2: A C T T T G A C C A A A T T G
SEQUÊNCIA 3: A C T T T G A C C C A A T T G
SEQUÊNCIA 4: A C T T T G A G C A A A T T G
Em alguns casos, a base mutada pode vir a deslocar a fase de leitura no momento da tradução do mRNA para peptídeos, assim produzindo um peptídeo diferente do tipo

39
selvagem (normal). Este peptídeo mutado pode não exercer sua determinada função, tornando ineficiente ou interrompendo determinado processo metabólico.
12-ESCOLHA DO MARCADOR
A escolha do marcador a ser utilizado depende de diversos fatores, como o tipo de estudo, as facilidades laboratoriais e os custos envolvidos. As características mais importantes a serem considerandas quando se comparam marcadores para um determinado estudo são a capacidade multiplex (número de locos aplificados em uma única reação), o número de alelos por locos) e a proporção de locos polimórficos. A natureza dominante ou co-dominante do marcador também é crucial para alguns estudos (Tabela 3).
13-APLICAÇÕES DOS MARCADORES MOLECULARES
Especificamente no melhoramento de plantas os marcadores moleculares têm muitas aplicações. Em primeiro lugar, o desenvolvimento de mapas de ligação, altamente saturados com marcadores. Estes mapas servem de base para o mapeamento de outras características de importância agronômica, principalmente as de natureza quantitativa e governadas por muitos genes. Desta forma é possível verificar as associações (ligações genéticas) entre os marcadores moleculares e os genes que afetam um caráter quantitativo.
Quando isto está estabelecido, o critério de seleção agora pode ser um ou vários marcadores (bandas) e não mais o fenótipo, já que selecionando-se um marcador, teoricamente seleciona-se os genes próximos a este. Assim é possível se fazer uma seleção genotípica ao invés de seleção fenotípica, que é muito menos eficiente. A seleção indireta faz sentido mesmo para um caráter qualitativo, quando este é muito caro ou difícil para ser avaliado, como é o caso de resistência a nematóides ou produção de uma determinada proteína ou substância de interesse industrial ou farmacológico.
Os marcadores moleculares ainda têm outras utilidades como a identificação de germoplasma, a identificação de variedades, o controle de qualidade na produção de sementes híbridas, a caracterização genética de populações, o monitoramento nos retrocruzamentos e auxílio na identificação e clonagem de genes, entre outras.
* Construção de mapas genéticos - Em primeiro lugar o grande volume de marcadores disponíveis possibilita o desenvolvimento de densos mapas de ligação, uma ferramenta tanto para pesquisa básica quanto aplicada. Os marcadores de DNA segregam em proporções mendelianas e não interferem na segregação de outros genes. Quando em grande quantidade segregando num cruzamento, é possível a construção de um mapa genético de ligação, cuja densidade depende da quantidade de marcadores. Mapas genéticos de alta densidade eram praticamente utopia numa fase anterior ao desenvolvimento desses marcadores. Nos últimos anos foram construídos mapas genéticos de ligação das principais espécies vegetais cultivadas, de animais domesticados e de espécies utilizadas como modelo em laboratório.
Além de mapas, os marcadores facilitam o mapeamento de genes específicos. cDNA é uma molécula de DNA sintetizada a partir do mRNA. Portanto, o cDNA seria um gene (DNA) sem os introns. Quando o cDNA é obtido de um mRNA de um gene conhecido, sabe-se a função deste cDNA.
* Caracterização da variabilidade genética - Entre 1966 e 1984 (18 anos) a eletroforese foi utilizada em mais de 1000 espécies, para estudos de genética e evolução. De maneira geral, foram avaliados em média de 23 locos em mais de 200 indivíduos. Uma vez caracterizado o germoplasma disponível, o melhorista pode escolher genotipicamente os progenitores para um cruzamento tanto com o objetivo de maximizar a segregação de genes de importância agronômica como restringir esta segregação a poucos genes. Além da escolha dos progenitores, será possível identificar os recombinantes desejados.

40
* Monitoramento - Monitorar a recuperação do genoma do pai doador nos retrocruzamentos (intra e interespecíficos) através de marcadores específicos pode diminuir o tempo e a quantidade de trabalho necessários para a introgressão de um ou poucos genes. A avaliação genotípica através de marcadores moleculares de 120 linhagens BCF6 de tomate, provenientes do cruzamento entre L. pennellii e L. esculentum e retrocruzadas para o L. esculentum, foi verificado que 21 delas cobrem 95% do genoma da espécie L. pennellii.
* "Fingerprinting" - Fingerprinting ou a caracterização genética de um genótipo é outra aplicação dos marcadores moleculares. Isoladamente os mini ou microssatélites ou em conjunto com outros marcadores moleculares, podem ser utilizados para caracterizar e distinguir uma variedade de outra. Para a diferenciação varietal três requisitos básicos são essenciais: 1º) distinção - diferentes genótipos devem apresentar distintos padrões de bandas; 2º) uniformidade - o mesmo padrão de bandas deve ser obtido se o procedimento for repetido e 3º) estabilidade - o padrão de bandas não se altera mesmo que o genótipo for cultivado em diferentes ambientes. Dependendo da legislação brasileira de proteção às cultivares e regras de patenteamento a ser definida, as impressões digitais de DNA ('fingerprinting') poderão ter grande utilidade.
* Mapeamento de QTLs - A maioria das características relacionadas com os processos de crescimento em plantas dependem da expressão de muitos genes. Historicamente, a biometria possibilitava a análise em massa desses genes, sem a caracterização da contribuição individual de cada um dos componentes do sistema. Com o advento dos mapas genéticos de ligação, altamente saturados, foram criadas as condições para o estudo individualizado dos QTL (Quantitative Trait Loci), pois tais mapas proporcionam marcadores moleculares em todas as regiões do genomas, em alguns casos espaçados apenas de menos de 2 cM.
Neste caso, a progênie oriunda do cruzamento entre plantas que diferem para um QT (Quantitative trait), são agrupadas com base num marcador molecular e então estimada a média e variância da característica fenotípica das plantas de cada classe. Uma diferença significativa entre as médias das classes, indica a relação entre o marcador e a característica, mais especificamente, uma ligação entre o marcador de DNA e um dos alelos que afeta este caráter.
Vários QTL relacionados com as características do fruto em tomate (Paterson et a., 1988, 1991) e com as interações entre bactéria e feijão comum (Nodari et al., 1993). No primeiro caso, foram identificados seis QTL afetando o tamanho do fruto e explicando 58% da variação fenotípica do caráter. Alguns desses QTLs demonstraram efeito sobre o caráter em dois ou mais ambientes e outros em apenas um só ambiente.
Cinco QTLs associados com a tolerância a baixo teor de fósforo foram identificados em milho com auxílio de um mapa de RFLP. Todos os cinco QTLs apresentaram efeitos apenas aditivos. Entretanto, uma interação entre dois QTLs foi significativa. Alelos que contribuem para a tolerância foram detectados em ambos os progenitores.
O mapeamento de QTLs proporciona a identificação não só de alelos envolvidos na expressão do caráter, mas o que é mais importante, as possíveis interações entre os QTLs, proporcionado ao melhorista informações que podem ser úteis na escolha dos progenitores para a realização dos cruzamentos. Proporciona ainda condições para o desenvolvimento de estoques genéticos com diferentes composições genéticas. Tais combinações permitirão a comprovação dos efeitos individuais dos QTLs, anteriormente estimados.
Existem programas que permitem determinar as distâncias genéticas entre marcadores como é o caso do Fstat (Goudet, 1995), GDA (Lewis e Zaykin) e outros que facilitam a construção de mapas como o MAPMAKER (Lander et al., 1987).
* Seleção assistida por marcadores (MAS) - A prática da seleção indireta para caracteres de baixa herdabilidade poderá ser intensamente explorada desde que os genes de interesse estejam fortemente ligados a marcadores moleculares. A seleção indireta e

41
genotípica (marcador molecular como critério de seleção), possibilita ainda a seleção de alelos com efeitos positivos provenientes dos dois ou mais progenitores envolvidos na geração da população segregante. A ligação entre o alelo Aps1 da fosfatase ácida e o gene Mi (distância de ± 1cM) que codifica a resistência ao nematóide, tem possibilitado a seleção de plantas de tomate resistentes em populações segregantes através da eletroforese desde 1974, quando foi iniciado por Charles Rick. O alelo Aps1 que está ligado do gene Mi que causa resistência ao nematóide em L. esculentum foi transferido do L. peruvianum através do sistema por retrocruzamento (mais de 30 retrocruzamentos para o L. esculentum). Um segundo exemplo relaciona-se com a incorporação de três genes de resistência à ferrugem em feijão realizada por James Kelly, da Universidade de Michigan, utilizando marcadores RAPDs, altamente ligados aos 3 principais genes de resistência.
O procedimento 'Bulked Segregant Analysis' (BSA) (Michelmore et al., 1991) em conjugação com a PCR é uma alternativa eficiente de mapear genes específicos e selecionar indiretamente genótipos desejados.
* Clonagem de genes - Em sétimo lugar, os marcadores auxiliam na clonagem e transferência de genes de interesse agronômico. Entre os mais freqüentemente citados encontram-se os genes de resistência a pragas e doenças. Entretanto, outros genes podem causar profundo impacto nos produtos finais das plantas. Trata-se dos genes que podem proporcionar às plantas o uso de rotas metabólicas alternativas, resultando em produtos novos ou modificados, em muitos casos de alto valor econômico.
Os genes já caracterizados pela genética clássica,têm seu fenótipo conhecido, mas normalmente seu produto é desconhecido. Um marcador de DNA que está próximo de um desses genes, pode ser o ponto de partida para o sua identificação e clonagem. Uma das alternativas é pela técnica denominada de 'caminhar no cromossomo’ (chromosome walking). Esta técnica compreende o isolamento de vários clones com sobreposição parcial. O marcador de DNA é utilizado inicialmente como sonda para identificar um desses clones. Pela sub-divisão desse clone identificado, é possível a identificação de um segundo clone, adjacente ao primeiro, e é similar a este na região de sobreposição. Este segundo clone é então utilizado como sonda para identificar um terceiro clone e assim por diante. Esta 'caminhada' pode eventualmente atingir o gene de interesse, que estaria contido num dos clones.
Assim, vários genes foram isolados com auxílio deste 'caminhar no cromossomo'. Entretanto esta técnica é difícil, cara e demorada. Ainda apresenta alguns problemas como seqüências repetidas de DNA que podem estar em um grande número de clones, impossibilitando a 'caminhada' na direção exata do gene de interesse. O outro problema, refere-se a grande distância entre um marcador e o gene de interesse.
Avanços como a possibilidade de clonar fragmentos de grande tamanho (YAC; Yeast Artificial Chromosome e BAC; Bacterial Artificial Chromosome) e de separar grandes moléculas de DNA (PFGE; Pulse Field Gel Electrophoresis) facilitarão a clonagem de um gene a partir de um marcador molecular
* Estudos de crescimento e desenvolvimento das plantas - O crescimento e o desenvolvimento das plantas estão sob o controle de muitos genes. Vários desses genes já foram identificados, inicialmente através da genética clássica e mais recentemente com auxílio da genética molecular. Exemplos: fitocromo e genes que afetam o padrão de cor das plantas. O gene Phs responsável pela produção da faseolina como a principal proteína de reserva das sementes de feijão foi mapeado com auxílio de marcadores moleculares (Nodari et al., 1993). O gene nts (nodulação tolerante ao nitrato) foi mapeado com auxílio de marcadores moleculares numa população F2 (10cM).
* Modificações na organização do genoma - Existem amplas evidências do surgimento de variantes durante a regeneração a partir de cultura de tecidos. Variação somaclonal que ocorre ao nível do DNA, tanto nos sítios de reconhecimento de uma enzima de restrição ou na região de anelamento de um primer podem ser detectadas via RFLP, AFLP ou RAPD, respectivamente. Variação no número de cópias também pode ser

42
detectadas pela intensidade de hibridização, via RFLP. Os RFLPs também têm potencial para detectar variação fenotípica decorrente de alterações no padrão de metilação, já verificado em milho. Variação somaclonal em milho foi atribuída a variação ocorrida ao nível do DNA.
Além disso, os marcadores moleculares são extremamente úteis na diagnose de doenças, sexo, oncogenes, etc. Neste caso, os marcadores com base na PCR são os mais adequados, considerando-se rapidez, distinção, custos e praticidade e reproducibilidade em outros laboratórios.

Tabela 2.1 - Análise comparativa entre os marcadores moleculares
Atributos
Isoenzimas
Proteínas de sementes
RFLPs
RAPDs
Microssatélites
AFLPs
Nível de Polimorfismo baixo alto baixo-alto baixo-alto muito alto muito alto
Estabilidade ambiental
moderada alta alta alta alta alta
Número de locos moderado (<50) baixo (<10) alto alto alto alto
Expressão genética co-dominate co-dominante co-dominante dominante co-dominante dominante
Número de alelos por loco
2-5 multialélico multialélico 2 multialélico 2
Distribuição no genoma
regiões de cópia única
regiões de cópia única
várias ao acaso ao acaso ao acaso
Acessibilidade tecnológica
baixa muito baixa média média média média
Aplicabilidade no melhoramento
rápido, baixo custo
rápido, baixo custo
lento, custo médio
rápido, baixo custo
lento, custo alto rápido, custo baixo
Identificação de genótipos
baixa baixa média muito alta muito alta muito alta
Avaliação de germoplasma
média baixa alta alta alta muito alta
Mapeamento genético
baixa muito baixa alta alta muito alta alta
Mapeamento de regiões específicas
baixa inadequado média muito alta média muito alta
Mapeamento comparativo
baixa inadequado muito alta baixa alta baixa
Genética de Autógamas
baixa baixa média alta muito alta muito alta
Genética de Alógamas
média baixa média alta muito alta muito alta
Análise Filogenética média baixa muito alta média alta média
Adaptado de Gepts (1993) e Ferreira & Grattapaglia (1995).

44
PARTE 3 - ORGANISMOS GENETICAMENTE MODIFICADOS
1. INTRODUÇÃO
Organismos transgênicos (ou Organismos Geneticamente Modificados - OGM) são organismos (plantas, animais ou microrganismos) que têm inserido em seu genoma, uma sequência de DNA manipulado em laboratório por técnicas moleculares ou biotecnológicas. O DNA inserido pode ser da mesma, de outra espécie ou ainda sintético. Tais técnicas, desenvolvidas nos últimos 20 anos, possibilitam o corte e a ligação de fragmentos de DNA de uma forma altamente precisa. Particularmente, seqüências de DNA (genes) podem ser removidas de um organismo, ligadas a seqüências regulatórias e inseridas em outros organismos. A fonte destes genes pode ser qualquer organismo vivo (microorganismo, planta, animal) e o organismo receptor, nesse caso específico, uma variedade de uma espécie de planta cultivada.
As plantas, animais e microrganismos transgênicos possibilitam tanto (i) estudar questões biológicas fundamentais a nível molecular como também (ii) materializar aplicações da biologia celular e molecular, como por exemplo ingroduzir em plantas uma nova característica (ex: resistência a herbicidas). Em tese, a tecnologia tem potencial para modificar o código genético dequalquer espécie visando a prouçõ de uma nova proteína, outro produto ou mesmo alterar a regulação de um ou mais genes.
A expressão tecnologia do DNA Recombinante surgiu em 1973 quando moléculas DNA de diferentes espécies foram recombinadas in vitro. Basicamente, trata-se do uso de dois grupos de enzimas: as de restrição (do tipo II) que são capazes de reconhecer uma pequena seqüência de pares de bases e então cortar o DNA neste sítio de reconhecimento ou as de corte, conhecidas como ligases, que são enzimas capazes de ligar dois fragmentos de DNA.
Para obter uma planta transgênica a tecnologia desenvolvida após 1973 consiste de construir um vetor com as sequencias de DNA recombinante desejáveis (cortar e colar) que pudesse transferir tais sequencias a um hospedeiro. O primeiro vetor foi utilizar um plasmideeo evinserir nele tais sequências recombinantes (transgene). Plasmideo é uma molécula de DNAcircular, de tamanho pequeno, que habitam em bactérias, e que são facilmente manipouladas em laboratório. O primeiro plasmídeo modificado in vitro (Cohen et al, 1973) foi construído a partir do corte de DNA com enzimas de restrição e a ligação de fragmentos específicos (transgene) com as enzimas ligases. Surge então o que se convencionou denominar de „tecnologia do DNA recombinante‟, posteriormente denominada de „engenharia genética‟. Posterrmente, o trsngene deste plasmidio em contacto com células de um hospedeiro, pode transferir tal DNA recombinate para células do hospedeiro e integrar-se em seu genoma, técnica denominada de transformação genética. É uma tentativa de se fazer in vitro o que ocorre na natureza: a recombinação de fragmentos de DNA. Contudo, na natureza, dificilmente o DNA de uma espécie pode ser cortado e ligado ao DNA de outra espécie.
No entanto, 25 anos antes do clássico trabalho de Watson e Crick sobre a estrutura do DNA e 55 anos antes da tecnologia do DNA recombinate, já havia sido descoberta a transformação genética. Em 1928 Frederick Griffith conseguiu transformar uma cepa de Streptococcus pneumoniae atenuada e não encapsulada (denominada na época de pneumococcus Tipo II) em uma cepa, agora virulenta e com capacidade de encapsulamento (Tipo III). Para tal, Griffith inoculou simultaneamente em um rato uma pequena quantidade de uma cultura viva de pneumococcus Tipo II (R) e uma grande quantidade de uma cultura Tipo III (S), morta pelo calor (Susuki et al., 1986). Não só o rato morreu como as células recuperadas foram igualmente virulentas em inoculações subseqüentes. O fato de que o Tipo

45
II (R) se tornou virulento foi considerado uma prova da aquisição desta característica a partir do outro tipo. Este fenômeno foi chamado na época de „transformação‟.
De lá para cá, o avanço no conhecimento da genética, bioquímica e fisiologia de procariotos possibilitou ao ser humano a manipulação do DNA, a molécula cuja função é carregar a informação genética que é lida pela maquinaria celular durante o desenvolvimento de um organismo ou vírus. A manipulação do DNA e de sistemas celulares possibilitou o desenvolvimento de vários produtos e processos, permitindo ao homem reprogramar a vida dos seres vivos e de vírus (Nodari e Guerra, 2003).
A introdução de uma molécula de DNA recombinante numa planta se constitui na transformação de plantas. Para tal, utiliza-se de um vetor para que a construção genética feita em laboratório seja inserida no genoma da planta. As técnicas de engenharia genética possibilitam a transferência de genes por via não sexual.
2. TECNOLOGIA DO DNA RECOMBINANTE
A construção de uma molécula de DNA recombinante depende da ação de enzimas de restrição e enzimas ligases. As enzimas de restrição são nucleases capazes de clivar (cortar) o DNA em sítios específicos, ou seja, a partir de seqüências nucleotídicas específicas reconhecidas por estas enzimas. A utilização de enzimas de restrição (tesouras químicas) é de extremo interesse pois a partir da ação delas existe a possibilidade de se clivar seqüências de interesse, ou seja, genes e elementos regulatórios de interesse para a transformação genética de organismos. Portanto, quando se separam os fragmentos de DNA de interesse com enzimas de restrição, falta a ação das enzimas ligases para fazer a junção destes fragmentos. Como o DNA e uma molécula universal para todos os organismos vivos, as ligases são capazes de unir DNA de origens diferentes (diferentes espécies) ou mesmo sintética (seqüência nucleotídica sintetizada em laboratório).
O transgene é essa sequência de interesse, construída a partir da utilização de enzimas de restrição e ligases, que inclui não apenas o gene de interesse em si, mas também todas as sequências que o regula, como promotores, terminadores e genes repórteres ou de seleção (Figura 3.1).
Figura 3.1. Ilustração de um transgene.
A tecnologia do DNA recombinante tem interesse especial quando este transgene está inseridoema um plasmídeo, que tem função de vetor. Um vetor pode ser descrito como um veículo utilizado na construção do transgene ou mesmo na transformação de plantas e outros organismos.

46
3. GENES MARCADORES E GENES REPÓRTERES PARA SELEÇÃO
Os genes marcadores são utilizados para possibilitar a discriminação entre células transformadas e não transformadas e, consequentemente, a seleção das primeiras. Tais genes são introduzidos para facilitar o trabalho de identificação das mesmas, pois são uma minoria em relação ao total de células submetidas a transformação.
Os genes marcadores são geralmente genes de resistência a antibióticos. Assim, no momento da regeneração das plantas a partir de uma célula, a adição de antibiótico ao meio, permitirá apenas o crescimento daquelas células transformadas que expresses a referida proteína.
Os genes marcadores (e suas respectivas proteínas) mais utilizados são: gene neo, isolado do transposon Tn5 de Escherichia coli, codifica para neomicina fosfotransferase (NPTII), que confere resistência a kanamicina, e o gene hpt, também isolado de Escherichia coli, codifica para higromicina fosfotransferase (HPT).
Genes de resistência a herbicidas também estão sendo utilizados. Dentre eles, destacam-se: gene bar, isolado de Streptomyces hygroscopicus, codifica para fosfinotricina acetiltransferase (PAT) que induz a resistência a herbicidas a base de fosfinotricina; gene aroA, isolado de Salmonella typhimurium, que induz a resistência a herbicidas a base de glifosato e o gene csr1, que induz a resistência a herbicidas a base de imidazolidonas e sulfonilureas.
Genes repórteres codificam para proteínas que são facilmente detectáveis. Dentre os genes reportes, os mais utilizados são: gene uidA, extraído de Escherichia coli, codifica para a β–glucuronidase (GUS), detectada por métodos histoquímicos; gene gfp, extraído da medusa Aequorea victoria, codifica par a proteína fluorescente verde (GFP); gene luc, isolado do vagalume Photinus pyralis, codifica para a luciferase.
4. MÉTODOS DE TRANSFORMAÇÃO DE PLANTAS
A transformação de plantas consiste na introdução de um fragmento de ácido nucléico em um genoma. Existem duas estratégias para transformar plantas: direta e indireta. A estratégia indireta é aquela que utiliza um vetor como a Agrobacterium tumefaciens (o método mais usado para a obtenção de plantas transgênicas) ou A. rhizogenes como veículo de entrega do DNA à planta. Métodos químicos e físicos possibilitam a transformação direta de genomas. Dentre eles, destacam-se: métodos físicos como a biobalística (ou aceleração/bombardeamento de partículas), eletroporação e microinjeção; além dos métodos químicos, como polietilenoglicol-PEG.
Agrobacterium tumefaciens - Pertencente ao grupo das bactérias gram-negativas, tipo bacilo aeróbico, A. tumefaciens causa em algumas plantas uma doença chamada de galha-da-coroa, uma espécie de tumor. Este tumor é causado por genes bacterianos, que naturalmente são transferidos pela bactéria e inseridos no genoma nuclear da planta hospedeira. O segmento de DNA transferido à planta é denominado de T-DNA, que faz parte do plasmídeo bacteriano, chamado de plamídeo Ti (tamanho variável de 120 a 250 kb). O processo de transferência ocorre após a infecção, que tem início após a liberação de determinados compostos pela planta. Imediatamente vários genes da região vir do plasmídeo são expressos, os causadores da virulência, O T-DNA transferido está contido entre duas sequências terminais de 25 pares de bases, denominadas de extremidades esquerda e direita. A extremidade direita é imprescindível para a transferência. As demais sequências que naturalmente são transferidas às plantas não são necessárias ao processo em si de transferência. Desta forma, um plasmídeo pode ser engenheirado, com a substituição de todas as bases, exceção àquelas que compõem as extremidades, por genes de interesse. Assim, a A. tumefaciens se encarrega de transferir e inserir no genoma nuclear das plantas

47
uma construção quimérica contendo genes de interesse. O método é bastante eficiente, entretanto, esta bactéria não consegue infectar um grande número de espécies vegetais, o que limita bastante seu uso, como no caso das monocotiledôneas em geral. A primeira planta transformada com Agrobacterium tumafasciens foi em 1983 e só 11 anos mais tarde, a primeira variedade transgênica foi liberada para cultivo, o tomate longa vida (Flavr Savr).
A similaridade entre os métodos diretos de transformação de plantas consiste na capacidade de romper a parece celular e do envelope nuclear. Estes métodos são mais adequados do que os indiretos para transformação de pólen, embrião e meristemas (Brasileiro e Dusi, 1999). Serão descritos, agora, brevemente alguns métodos.
Biobalistica ou aceleração de partículas/ bombardeamento - É um método que utiliza microprojéteis em alta velocidade envoltos por DNA, com objetivo de superar a parede celular pela força, na esperança que algumas moléculas de DNA atinjam o núcleo e se integrem ao genoma nuclear. Os microprojéteis são constituídos principalmente de partículas esféricas de ouro ou tungstênio, de 1 mm de diâmetro. O DNA adere facilmente e fortemente a estas partículas, pois tais metais são carregados positivamente. Geralmente os equipamentos utilizam o gás hélio, eletricidade ou propulsão a ar e alta pressão na aceleração das partículas. Esta estratégia é empregada em plantas que normalmente não conseguem ser infectadas por A. tumefaciens. Por utilizar a força bruta para penetrar no núcleo da célula, esta estratégia pode a rigor ser utilizada em qualquer tecido e planta. A obtenção de uma planta transformada depende da regeneração de uma célula transformada.
Eletroporação - Método que consiste em submeter protoplastos misturados com DNA a uma descarga elétrica controlada opor um curto espaço de tempo. Esta descarga cria poros na membrana nuclear, facilitando a entrada de DNA no núcleo. Nesta solução de protoplastos, células sem a parede celular (núcleos com citoplasma) também estão presentes plasmídeos contendo genes de interesse. Com a criação de poros pela descarga elétrica, um ou mais plasmídeos podem penetrar no núcleo e se integrarem no genoma da célula. A obtenção de uma planta transformada também depende da regeneração de uma célula transformada.
Químicos – Existem várias substâncias químicas que facilitam a entrada no núcleo de construções quiméricas bem como a sua integração no genoma de células de plantas. O polietilenoglicol (PEG), é um policátion, é um dos mais utilizados, mas é de baixa eficácia. O PEG também é utilizado conjuntamente com outras estratégias. Polivinil álcool (PVA) também é utilizado.
Lipossomas – Neste método o DNA é envolto pelos lipossomas, que são vesículas fosfolipídicas, que são misturadas com protoplastos previamente tratados com PEG. De eficência muito baixo, é pouco utilizado.
Microinjeção - Tubos microcapilares (microsseringas) são utilizados para injetar o DNA no núcleo das células, sem causar danos severos. Este método é mais comum em animais. O uso de agulhas permite ultrapassar a parede celular e também o envelope nuclear. Outros métodos incluem o uso de fibras (de Silicon Carbide) ou laser, para perfurar a parece celular. Neste processo, são misturados os plasmídeos contendo os genes de interesse com fibras de silicon carbide e as células a serem transformadas. Sob agitação, as fibras de silicon carbide conseguem abrir poros nas células vegetais, o que permite a entrada de DNA. Alternativamente, microrraios laser podem perfurar a parede celular.
Também a embebição de uma solução de DNA com sementes e tubo polínico podem levar a transformação de células.
4.1-CONCEITO DE OGM OU TRANSGÊNICO
A transformação genética de plantas consiste na inserção no seu genoma de uma ou mais seqüências, geralmente isoladas de mais de uma espécie, especialmente arranjadas, de forma a garantir a expressão gênica de um ou mais genes de interesse. Neste contexto, o prefixo „trans‟ era plenamente justificado, pois exprimia a idéia de além de, neste caso,

48
significando o rompimento da barreira da espécie. Com o estabelecimento de normas gerais de biossegurança é que se começou a utilizar a expressão Organismo Geneticamente Modificado - OGM. Em tese, a expressão Organismo Geneticamente Modificado causa certa confusão, porque alguns cientistas dizem que todos os organismos são geneticamente modificados. Entretanto a Lei de Biossegruança no Brasil, define claramente o que são OGMs.
Quando se utiliza a transgenia, uma nova sequência gênica é introduzida, geralmente não nativa daquela espécie. Em muitos casos, a sequência inserida é formada por partes de diferentes genes de diferentes espécies ou sequências semi-sintéticas. O conjunto destas seqüências é chamada de quimera. Assim, a Soja RR transgênica resistente ao Round-up, herbicida à base de glifosato, contém material genético de pelo menos quatro diferentes organismos: promotor do vírus-do-mosaico-da-couve-flor (CaMV), peptídeo sinal da petúnia, gene EPSPS da Agrobacterium CP4 e a sequência 3‟ (NOS) da Agrobacterium tumefasciens.
Do ponto de vista legal, no Brasil, OGM é o organismo cujo material genético (ADN/ARN) tenha sido modificado por qualquer técnica de engenharia genética. A Lei 8.974, de 5/01/95, definiu ainda engenharia genética como a atividade de manipulação de moléculas ADN/ARN recombinantes. Esta lei foi subsituida pela nova lei de biossegurança (Lei n° 11.105, de 24/03/2005), que manteve a mesma definição de OGM. Pela legislação brasileira, então, qualquer planta que tenha seqüência(s) de DNA ou RNA engenheiradas (neste texto ADN e DNA serão utilizados como sinônimos, assim como ARN e RNA), deve ser considerada OGM, e está, portanto, submetida aos efeitos da referida lei, mesmo porque ela também regulamenta os produtos obtidos pelo processo do DNA recombinante. No presente trabalho, OGM será utilizado como sinônimo de transgênico, embora não haja concordância absoluta a respeito desta sinonímia.
No artigo 3 da Lei 11.105, a definição de um OGM (Organismo geneticamente modificado) é feita por meio de três incicos: III – moléculas de ADN/ARNrecombinante: as moléculas manipuladas fora das células vivas mediante a modificação de segmentos de ADN/ARN natural ou sintético e que possam multiplicar-se em uma célula viva, ou ainda as moléculas de ADN/ARN resultantes dessa multiplicação; consideram-se também os segmentos de ADN/ARN sintéticos equivalentes aos de ADN/ARN natural; IV – engenharia genética: atividade de produção e manipulação de moléculas de ADN/ARN recombinante; V –– OGM: organismo cujo material genético – ADN/ARN tenha sido modificado por qualquer técnica de engenharia genética.
Desta forma, pode-se definir plantas transgênicas (ou OGM) como plantas que têm inserido em seu genoma, uma ou mais seqüências de DNA manipulado em laboratório por técnicas de DNA recombinante ou engenharia genética. Alternativamente, plantas transgênicas poderiam ser definidas como organismos que tiveram seu material genético alterado por métodos que não aqueles naturais, considerando-se como métodos naturais em plantas o acasalamento sexual e a recombinação genética. A indução à mutagênese é uma outra maneira de alteração genética de uma planta. Neste caso, o genótipo do indivíduo é alterado também diretamente in vivo. Um exemplo disto é a exposição de sementes a agentes químicos, como o metil sulfonato, ou físicos, como raios de cobalto ou X, na esperança de que alguma modificação benéfica ocorra no genótipo previamente escolhido. No sentido conceitual de modificação não-natural, a transgenia equivaleria à mutagênese, pois também provoca uma alteração genética não oriunda de cruzamento. Também há similaridade entre ambas quanto à aleatoriedade no loco onde ocorrerá a modificação, o que impossibilita, com o que se conhece hoje, antecipar o que vai acontecer.
Contudo, existem várias diferenças entre ambas. Enquanto na mutagênese as modificações podem ser de substituição de uma base por outra, deleção ou duplicação de uma ou mais bases e rearranjos diversos. Na transgenia, as seqüências externas introduzidas são adicionadas no todo ou em parte do genoma hospedeiro.

49
Esta diferença é crucial, pois na tecnologia está embutida a possibilidade da aplicação de leis de propriedade industrial que permite o patenteamento das seqüências engenheiradas, bem como do processo de transgenia. Esta possibilidade baseia-se naquilo que é adicionado, uma vez que é conhecido, engenheirado e patenteado. O mesmo não ocorreu com a técnica da mutagênese de plantas, embora uma cultivar desenvolvida com esta estratégia possa ser protegida por leis de proteção intelectual.
Outra técnica desenvolvida para terapia genética na espécie humana, a quimeroplastia, foi adaptada para plantas (Beetham et al., 1999; Zhu et al., 1999). Ela possibilita a substituição ou a adição de uma base, em uma seqüência conhecida. Neste caso a diferença em relação à transgenia clássica é a utilização de oligonucleotídeos quiméricos. Seu alcance, contudo, é menor, restringindo-se a alterar ou adicionar uma ou poucas bases.
Com o objetivo de confundir a opinião pública, freqüentemente é dito por cientistas que “o homem vem produzindo transgênicos há milênios com a seleção artificial de plantas”. Como é possível perceber pela definição de OGM na legislação, os agricultores que domesticaram as plantas cultivadas ou os melhoristas não conseguiram alterar um genótipo in vivo. Selecionavam sim, as novas combinações (progênies), oriundas da recombinação genética da geração anterior. É preciso não esquecer que o processo evolutivo é composto de forças que criam ou amplificam a variabilidade genética e outras que afetam o destino desta variação, como bem destacou Charles Darwin, em sua obra A origem das espécies (1859). O efeito conjunto das mutações, aqui incluídas todas as modificações de DNA em condições naturais, e das recombinações entre mutantes, promove o surgimento de uma ampla gama de associações alélicas (Allard, 1960; Fehr, 1987), cujo destino é então dependente das diversas forças evolutivas como seleção, migração e deriva. Os primeiros agricultores selecionaram estas novas associações alélicas que melhor se adaptavam a sua maneira de cultivar em cada situação. Assim, não cabe aqui falar de transgenia, mas sim de processo evolutivo.
4.2- CARACTERIZAÇÃO DE UM EVENTO
Um evento transgênico é caracterizado quando uma seqüência específica de DNA recombinante, ou transgene, é inserida num genoma hospedeiro. Ou seja, cada planta transgênica liberada para uso comercial num pais é considerada um evento diferente. Se o mesmo transgene é inserido em mais de um genoma hospedeiro, se consideram eventos diferentes. Isto porque como não existe controle sobre o local da inserção, o número de cópias a integridade das cópias inseridas, e a composição deste organismo geneticamente modificado pode ser diferente.
5-DIFERENÇAS ENTRE OS MÉTODOS DE MELHORAMENTO CONVENCIONAIS E BIOTECNOLÓGICOS
Os agricultores, assim como os melhoristas, utilizam os princípios da diversidade genética quando fazem cruzamentos, e da segregação quando selecionam plantas ou animais considerados superiores. O melhoramento genético pode ser considerado uma forma de biotecnologia, empregada há milênios para diversos propósitos, incluindo a introdução novas variedades de plantas no ambiente. De fato, o melhoramento envolve a manipulação genética, mas não envolve as técnicas da engenharia genética conforme ficaram conhecidas desde 1973.
Por meio dos métodos de melhoramento, agora também chamados de convencionais, tradicionais ou clássicos, novas combinações genéticas são geradas por meio de cruzamentos sexuais entre plantas que apresentam as características consideradas como desejadas. Cruzamentos são feitos entre plantas da mesma espécie e, ocasionalmente, quando a variação genética desejada não existe dentro da espécie, alelos ou genes são transferidos ou substituídos de outras espécies do mesmo gênero. Juntamente com os genes desejados,

50
outros segmentos de DNA do genótipo doador, podem também ser transferidos ou substituídos e podem expressar características indesejáveis. Desta forma, a amplitude do estoque genético (gene-pool) para o melhoramento é determinada pela compatibilidade sexual de uma espécie e espécies aparentadas. Técnicas radicais como resgate de embrião e o cultivo de embriões têm contribuído para aumentar o gene-pool, mas de forma muito limitada. Quando se utilizam métodos de melhoramento, os cruzamentos sexuais possibilitam a substituição de alelos via recombinação homóloga e não a adição de uma quimera como na transgenia.
Das metodologias utilizadas pelo melhoramento de plantas, a introgressão de genes, feita por retrocruzamentos sucessivos do F1
para o genótipo recorrente, é a que mais se
assemelha à transgenia, em termos de obtenção de uma nova associação alélica. Contudo, existem muitas diferenças entre ambas, que estão explicitadas na Tabela 3.1.
Na transgenia, seqüências de DNA (genes) podem ser removidas de um organismo, modificadas ou não, ligadas a outras seqüências, incluindo as regulatórias, e inseridas em outros organismos. A fonte destes genes pode ser qualquer organismo vivo (microorganismo, planta, animal) ou vírus.
Uma das principais implicações da transgenia é o rompimento da barreira sexual. Desta forma, a transformação genética possibilita uma alternativa de introdução de genes em plantas. A rigor, isto implica que, teoricamente, qualquer gene, natural ou sintético, pode ser introduzido numa espécie vegetal. Assim, o pool gênico de uma espécie se torna extraordinariamente grande. As oportunidades para o melhoramento aumentam drasticamente, pois além dos recombinantes produzidos naturalmente pela meiose, é possível obter recombinantes não convencionais. Desta forma, problemas de difícil solução ou mesmo a expressão de características em outros organismos poderiam ser adequadamente resolvidos.
Tabela 3.1. Comparação entre o método do retrocruzamento e a transgenia.
Retrocruzamento Transgenia
Objetivo Alterar ou introduzir uma característica
Alterar ou introduzir uma característica
Natureza Substituição de alelos Introdução de seqüências novas (quimera)
Tempo 3 a 6 anos Variável
Tecnologia Simples Sofisticada
Pool gênico Limitado Ilimitado
Custo Baixo Elevado
Resultados Previsíveis Limitados
Imprevisíveis Ilimitados
Efeitos adversos Raros Ex: alelos indesejáveis
Freqüentes Ex.: genes marcadores, promotores e outras seqüências filogeneticamente bem distintas; efeitos pleiotrópicos
Distribuição dos benefícios
Instituições públicas e privadas, pequenos agricultores, consumidores.
Grandes empresas, grandes agricultores, melhoristas
Fonte: Nodari e Guerra (2001)
Neste cenário, e considerando-se o ponto de vista científico, duas limitações restringem o uso de genes via transgenia: a criatividade e o julgamento inadequado do valor

51
de um gene, desde que há disponibilidade de tecnologias de isolamento e transformação de uma dada espécie. Esta última limitação refere-se a situações em que o pesquisador não consegue perceber ou não tem informações sobre a utilidade de um gene num programa de melhoramento de uma espécie.
Além dessas limitações, já estão sendo adicionadas outras, como: a real necessidade de um determinado OGM (comparação com alternativas) e a magnitude das implicações que ele possa apresentar se cultivado e ou consumido em larga escala.
A transgenia introduz novos genes exóticos, e muitas vezes de sequências rearranjadas e diferentes das originais, criando recombinações não naturais cujas localizações no genoma do organismo são imprevisíveis. Ou seja, a tecnologia ainda não permite o controle do local da inserção, nem da integridade da sequência inserida. Isto pode resultar em efeitos imprevisíveis no metabolismo, fisiologia e bioquímica do organismo receptor. O relatório do Governo da Noruega, divulgado em 1999, denominado Too early maybe too late: ecological risks associated with the use of naked DNA as a biological tool for research, production and therapy, concluiu que qualquer OGM deve sofrer avaliação de impacto ambiental antes de ser liberado. Este relatório refuta a ideia de que a transgenia em plantas é similar ao melhoramento genético convencional (Traavik, 1999).
O desenvolvimento de OGMs pode ser denominado de „tecnologia‟? Tradicionalmente uma tecnologia está associada com (i) previsibilidade, (ii) controle e (iii) reproducibilidade. Contudo, o atual estágio das tecnologias utilizadas na obtenção de OGMs pode ser caracterizado como (i) sem previsibilidade; (ii) sem controle dos sítios de inserção; (iii) sem controle do destino do transgene ou partes dele; (iv) sem controle nas mudanças de expressão gênica; (v) sem controle dos transgenes no ecossistema e (vi) de difícil reproducibilidade.
Ou seja, ainda não existe tecnologia disponível para a inserção da construção quimérica num loco específico do genoma da espécie recipiente. Um exemplo disto é o fato de que duas sequências de DNA (72 e 250 pb) derivadas da transformação original foram inesperadamente encontradas na Soja RR (The Scientist 14[15]:20, Jul. 24, 2000). Elas estão separadas do transgene que condiciona a resistência ao herbicida Roundup. Posteriormente, outros artigos demonstraram que em todos os eventos de transformação genética o transgene inserido não está como desenhado e construído. Isto demonstra que a modificação genética é inerentemente imprevisível.
Tampouco construído os resultados das transformações são previsíveis, sendo que algumas dão o resultado esperado, outras não. Também não é possível controlar a expressão gênica do gene inserido. Um exemplo disso é que diferentes variedades de milho com o mesmo gene de Bt produzem diferentes quantidades de toxina nos diferentes órgãos estudados e comparados. Outro aspecto importante é que não se consegue controlar o transgene inserido, uma vez que ele pode se disseminar para outras espécies e causar poluição genética, e como tal enormes e irreparáveis danos. Exemplo disto foi a contaminação de várias plantações de milho nos Estados Unidos provocada pelo cultivo de uma variedade transgênica, StarLink, que causou enormes prejuízos aos agricultores, aos consumidores e à empresa.
6-APLICAÇÕES
PLANTAS
Como aproximadamente 90% das calorias provem de plantas, é no reino vegetal que existe um grande potencial de oportunidades para as diversas biotecnologias, incluindo-se a transgenia, especialmente na produção de alimentos e energia. Contudo, na área da saúde são esperados investimentos financeiros elevados e o desenvolvimento de muitos produtos, muitos deles, de aplicação praticamente pessoal.

52
O aumento da resistência de plantas a pragas e moléstias pela ação de produtos naturais com auxílio da Tecnologia do DNA Recombinante é a oportunidade importante. No entanto, a maioria dos genes inseridos em plantas inclui aqueles que conferem resistência a herbicidas e a poucos insetos, ou ambos. Outros genes, como por exemplo controlando o teor de proteínas e óleos em plantas estão sendo testados. A partir de 1994, foram identificados, clonados e sequenciados vários genes de resistência a doenças. O conhecimento pleno destes genes possibilitará um melhor entendimento de como ocorrem as reações de resistência ou susceptibilidade de plantas à fungos, bactérias e vírus, bem como desenhar estratégias apropriadas de melhoramento e seleção de plantas resistentes.
Oportunidades agrícolas projetas por parte da comunidade cientitifca incluem ainda genes que conferem tolerância a estresses climáticos (altas temperaturas e seca) e de solo (baixos teores de nutrientes e altos teores de elementos tóxicos). Embora, não se saiba ao certo o mecanismo de tolerância, novas abordagens para a manipulação genética visando a tolerância aos estresses estão sendo desenvolvidas há muito tem, mas ainda sem êxito.
Uma segunda área de grande atividade da tecnologia do DNA Recomninante é relacionada com o aumento do valor de certas espécies agrícolas pode ser alcançado através de modificações genéticas que alteram a quantidade ou composição de compostos de reservas não protéicos, os quais podem substituir inclusive certos produtos derivados do petróleo. Estes novos produtos ainda são promessas.
As plantas também poderão se tornar fábricas de produtos ou substâncias, já que, na maioria dos países, a produção de uma substância em cultura de células ou em determinados microrganimos tem inúmeras restrições. Exemplo disto são os testes em andamento para a produção de produtos como o hormônio do crescimento humano em milho, vacinas, anticoagulantes entre outros.
Vacinas comestíveis produzidas por plantas, advogam alguns cientistas, é um sistema bastante apelativo, pois apresenta inúmeras vantagens sobre as formas convencionais: armazenamento em condições menos sofisticadas, simplicidade de aplicação, custos reduzidos, fácil produção e diminuição dos riscos de transmissão de outras doenças com equipamentos e materiais contaminados. Contudo, uma questão ainda pendente é a segurança e a eficiência destas vacinas produzidas por plantas. Outra preocupação relaciona-se com a quantidade da fruta ou alimento a ser ingerido, bem como o controle da produção dos mesmos. Embora o assunto é complexo e polêmico, vários laboratórios em muitos países estão desenvolvendo este tipo de vacinas utilizando estratégias diferentes. Embora muitas promessas foram feitas desde 1995, ainda (estamos em 2016) não existe nenhuma vacina contida em tecidos de plantas (ex: folhas) ou em frutos (ex: banana).
Outra aplicação relacionada com a manipulação dos metabólitos secundários é a produção de polímeros biodegradáveis. Tais polímeros são na realidade uma mistura de amido e polietileno. Quando o amido é o maior componente, temos os plásticos complexos, já em comercialização como Novon e Fertec. (Novon - 80% amido mais etileno-acetato de vinil ou co-polímero etileno-ácido acrílico; Fertec - 50% amido e polímeros). Os filmes são resultantes de misturas com baixos teores de amido.
Do ponto de vista alimentar, novas promessas continuam sendo anunciadas. São as chamadas segunda e terceira ondas, cujas aplicações da engenharia genética estão relacionadas com o aumento da qualidade dos produtos alimentícios. Como exemplo menciona-se que estão sendo desenvolvidos OGMs com alto teor de aminoácidos, proteínas ou alta qualidade do óleo e plantas que produzem altas quantidades de vitaminas, como experimentalmente já obtido em cenoura e arroz. Tais alimentos são chamados de nutracêuticos.
Um programa resultando do esforço mundial de proponentes da transgenia com apoio financeiro de empresários (ex: Fundação Bill Gates) denomina-se biofortificação. O programa visa o desenvolvimento de variedades com altos teores de um ou mais nutrientes. Esta visão reducionista é uma estratégia que visa resolver problemas nutricionais em várias partes do

53
mundo. Esta deficiência nutricional é resultante de vários fatores: falta de acesso a alimentação, desenvolvimento de novas variedades com menores teores de nutrientes ou vitaminas, estreitamento da base alimentar No Brasil, este programa é gerenciado pela EMBRAPA e recebe o nome de biofortificação. Neste contexto, muitas propostas sugerem o uso da transgenia ou para inserir genes ou para modificar rotas metabolócias visando a maior produção de um ou outro nutriente.
ANIMAIS
A primeira leva de animais transgênicos foi destinada a produzir substâncias para uso na saúde humana ou para fornecer órgãos para transplante, também para a espécie humana. Dentre as proteínas humanas produzidas em animais transgênicos destaca-se o fator de coagulação, necessário no tratamento da hemofilia, a eritropoietina, que é utilizada para estimular a medula óssea quando deprimida por outras drogas e a alfa-1 antitripsina, utilizada no tratamento de enfisema pulmonar.
A transgênia em animais engloba aqueles que tiveram transgenes adicionados (transgênicos por adição), genes modificados (knockin) ou genes retirados (knockout). A primeira linhagem de camundongos transgênicos em 1982, foi produzida por Palmiter, da Universidade da Pensilvânia e colaboradores. Cinco anos depois, foi desenvolvido o primeiro rato para produzir um medicamento de uso humano tPA (Tissue Plasminogen Activator) no tratamento de sangue.
Os possíveis benefícios mais diretos e biotecnológicos do uso de animais transgênicos podem ser divididos em pelo menos três grupos: agricultura, medicina e indústria. Segundo Pereira (2008) na agricultura, "a transgênia pode permitir a criação de animais de grande porte com características comercialmente interessantes, cuja produção por técnicas clássicas de cruzamentos e seleção são extremamente demoradas. Assim, existem vacas transgênicas que produzem mais leite, ou leite com menos lactose ou colesterol, porcos e gado transgênicos com mais carne e ovelhas transgênicas que produzem mais lã. Além disso, há um grande esforço no sentido de se produzir animais resistentes a doenças, como a gripe suína ou a febre aftosa em bovinos. Porém, isso dependerá da identificação de genes responsáveis pela resistência a essas doenças".
A mesma autora menciona ainda que a transgenia em animais de grande porte vem sendo utilizada para a produção de fármacos. Produtos como insulina, hormônio de crescimento e fator de coagulação podem ser obtidos do leite de vacas, cabras ou ovelhas transgênicas.
Além disso, os animais transformados com genes humanos destinados à produção de órgãos para xenotransplantes, ou seja, o transplante de órgãos animais para o ser humano, como é o caso de suínos, estão sendo alvo de inúmeras discussões, não só do ponto de vista ético, mas também biológico. Os porcos transgênicos imuno-compatíveis com o ser humano foram produzidos pela técnica de nocaute (knockout), e os animais produzidos não expressam uma proteína imunogênica em seres humanos. Em relação a saúde humana, os riscos dos xenotransplantes estão basicamente centralizados na disseminação de vírus ou outras entidades (micoplasmas e partículas infecciosas) que também podem causar doenças ou injúrias à saúde humana. Do ponto de vista ético e religioso, é pertinente uma discussão mais ampla com os diversos segmentos da sociedade, uma vez que este assunto é extremamente polêmico. No entanto, é importante ressaltar que, se por um lado o xenotransplante resolveria a questão da disponibilidade de órgãos para transplantes, ele cria uma outra questão séria de biossegurança, criando o risco de transmissão de patógenos suínos para o ser humano (Pereira, 2008).
A aplicação da transgenia na indústria transforma os animais em bio-reatores através da transgenia. Um exemplo é a cabra transgênica que produz em seu leite uma proteína da teia de aranha. A purificação em grande escala desses polímeros a partir do leite permite a

54
criação de um material leve e flexível com enorme resistência, que poderá ser usado em aplicações militares (coletes e uniformes a prova de bala) e médicas (fio de sutura), entre outras (Pereira, 2008).
Peixes transgênicos já estão prestes a chegar à mesa do consumidor americano. A liberação de salmão transgênico já foi aprovado pela FDA, a agência que regula a entrada de alimentos e medicamentos no mercado americano. No entando, a mesma agência suspendeu a entrada e criação do freferiudo Salmão em território americano, o que causou estranheza. Quando isto acontecer de fato, será a primeira vez que um animal transgênico estará disponível para consumo humano. A diferença entre os salmões naturais e os transgênicos é que nestes foi inserido um gene que acelera seu crescimento, isolado de outro peixe, a lampréia. Os genes introduzidos estimulam a produção contínua de hormônios de crescimento.
Mais recentemente, galinhas transgênicas foram desenvolvidas para render mais carne como é o caso da Terminator Chicken – da empresa AviGenics. A mesma empresa engenheirou galinhas com genes humanos para produzir medicamentos. Em ambas, a empresa inseriu também uma seqüência de DNA que considera segredo e que possibilita ser detectada, visando a rastreabilidade para fins comerciais, ou seja, impedir que alguém use as galinhas sem pagar pela tecnologia. Animais de outras espécies também já foram modificados via transgenia como vacas, ovelhas e ratos.
Um dos métodos mais utilizados na produção de animais transgênicos é a microinjeção pronuclear, que consiste na injeção de uma solução de DNA, contendo o transgene de interesse, no pronúcleo de um óvulo recém-fertilizado. Esta metodologia faz com que várias cópias do transgene injetado se integrem em tandem em um sítio aleatório no genoma e sejam transmitidas de forma mendeliana (Pereira, 2008).
Segundo esta autora, apesar de ser uma importante ferramenta de pesquisa, esse método apresenta algumas limitações. Por causa do sítio aleatório de integração do transgene, este poderá não estar sob o controle de todos os elementos em cis (no mesmo cromossomo) que controlam a expressão do gene endógeno. Assim, a expressão temporal e espacial do transgene não seguirá o padrão de expressão do gene endógeno. Além disso, no que diz respeito a modelos de doenças genéticas, a introdução de um terceiro alelo, o transgene mutante, cria uma situação artificial no que diz respeito à proporção entre os transcritos normais e mutantes. Enquanto uma pessoa com uma doença genética dominante possui um alelo normal e um mutado, o camundongo transgênico possuirá os dois alelos endógenos normais e diversas cópias do alelo mutante (transgene). Esta proporção pode ser crítica em doenças suscetíveis a efeitos de dosagem gênica.
No entanto, a maior aplicação da tecnologia do DNA Recombinante em animais foi o desenvolvimento de milhares de ratos transgencicos portadores de transgenes que causam alguma deficiência, anomalia ou mesmo doença,. Tais ratos, são utilziados como cobaias para aprofucndar os estudos sobre tais deficiências edoenças bem como a cura das mesmas.
Enquanto os cientistas buscam formas de justificar e convencer a opinião pública sobre a utilização de animais em experimentação animal, movimentos sociais cada vez mais intensos e numerosos são completamente contra a utilização de animais em experimentos científicos. A questão, segundo Nádia Farage, professora do Departamento de Antropologia da Unicamp, não é discutir formas de usar estes animais e sim não usá-los.
MICRORGANISMOS E PRODUTOS DERIVADOS
Com relação aos microorganismos (especialmente bactérias e fungos) existe grande potencial para obtenção de produtos industrializados, como por exemplo para a medicina humana, pois podem ser produzidos aminoácidos e vitaminas nestes microrganismos. Bactérias geneticamente transformadas podem ser usadas para produzir muitas proteinas importantes, hormônios de crescimento humano (hGH), interferons e vacinas (como contra a

55
Hepatite B) para imunização contra viroses. O uso dos microorganismos também se estende para a fermentação Láctea e alcoólica e a degradação de poluentes. Nestes casos, os microrganismos são transgênicos, produzem produtos que posteriormente são purificados e utilziados, diretamente por humanos ou como reagentes de processos fermentativos ou de transformaçao (ex: queijo). Desta forma, os OGMs não são consumidos diretamente pelos humanos, mas sim seus produtos derivados.
O primeiro produto comercial decorrente do uso da tecnologia do DNA Recombinante foi a insulina, comercializada a partir de 1982 nos Estados Unidos, justamente a partir de uma microorganismo transgênico. O gene humano responsável pela insulina foi isolado na espécie humana e introduzido na bactéria Escherichia coli, que passou a produzir e excretar este produto. Após a purificação, a insulina produzida em laboratório passou a substituir a insulina extraída de pâncreas de animais, uma vez que proporciona menos riscos aos diabéticos, que dependem deste medicamento. No Brasil, a insulina também já vem sendo produzida com microrganismos transgênicos. Cabe destacar que o produto não é transgênico, uma vez que é a expressão do próprio gene humano, mas somente o organismo que o produz.
Outro aspecto importante, é que estes produtos destinados à saúde humana oriundos de microrganismos transgênicos passam pelos mesmos testes que passam os medicamentos convencionais. Sendo assim, a expectativa é de que estes produtos apresentam mais riscos relacionados a contaminações do que propriamente decorrentes do uso per se da tecnologia do DNA Recombinante.
TERAPIA GENÉTICA
A terapia genética ou gênica é uma forma de tentativa de tratamento para doenças, geralmente hereditárias, que consiste na inserção de um transgene funcional dentro da célula humana a fim de conferir uma nova função ou melhorar os efeitos de um gene anormal ou que não esteja funcionando normalmente. Na espécie humana, a terapia gênica se constitui também numa das áreas de pesquisa.
Trata-se de uma estratégia que visa disseminar no corpo humano ou num órgão específico, um transgene que contém um gene normal para que o mesmo possa expressar seu produto adequadamente, naqueles casos onde um ser humano é portador de defeito genético. Dependendo do alvo, as técnicas de terapia genética são de dois tipos: a germinativa, na qual ocorre a introdução do material genético nos espermatozóides ou óvulos (células germinativas), e a somática, que compreende as demais células nas quais é introduzido o material genético.
São dois os principais fatores limitantes: O primeiro é o vetor que deve levar os transgenes até o tecido ou órgão onde a terapia deve ocorrer. O segundo fator, é expressar o transgene somente no tecido ou órgão alvo e não nas demais células do organismo.
Os elementos que auxiliam o transporte e expressão destes transgenes são previamente modificados in vitro de forma a garantir sua inocuidade como elementos transportadores de sequências gênicas. Mesmo os retrovírus, modificados in vitro para carrear genes codificadores de proteínas de amplo interesse médico, como a expressão de adenosina deaminase - ADA, cuja ausência impede a maturação dos linfócitos e, conseqüentemente, leva à ausência de qualquer resposta imunológica, espera-se não causarem doenças nos pacientes que estão recebendo este tipo de vírus transgênico como carreador de um gene de interesse. Na década de 1990, o panorama das fases dos protocolos clínicos aprovados em curso foi bastante desfavorável. Nos Estados Unidos, neste período, menos de 1% dos protocolos clínicos (apenas quatro até o momento) avançaram pelas fases I e II que testam a segurança e a eficácia do método, respectivamente (Nardi, 2002).
Várias experiências resultaram em mortes de pacientes ou de aparecimento de doenças como a leucemia, após o tratamento com terapia genética. A terapia genética, que

56
causou duas mortes entre 1998 e 2006, é acusada de causar uma terceira morte em 2007 (Science, 2007). A possível causar seria o vetor popular usado neste experimento, o vírus adeno-associado (AAV). No entanto, permaneceram dúvidas quão bem o paciente foi informado e como ela foi selecionada para a terapia genética. O estudo envolveu a injeção nas articulações de AAV que carregava um transgene que codificava para uma proteína que inibe uma citocina pró-inflamatória. A paciente recebeu uma injeção inicial em seu joelho direito em 26 de Fevereiro de 2007 e uma segunda em 02 de julho do mesmo ano. Após a segunda injeção, ela desenvolveu sintomas semelhantes aos da gripe. Dez dias depois, ela foi internada no hospital e faleceu lá depois de falência múltipla dos órgãos, em 24 de Julho.
O tratamento de terapia genética feito em 2005 para severe combined immune deficiency (SCID) ligada ao cromossomo X, no Great Ormond Street Hospital / Instituto da Criança Saúde em Londres, causou em uma criança uma forma de câncer (Nature, 2002). Os cientistas que conduziram a terapia lamentaram informar que uma das 10 crianças tratadas desenvolveu uma leucemia de células T após a utilização da terapia gênica.
7-EVOLUÇÃO DO DESENVOLVIMENTO E CULTIVO DAS PLANTAS TRANSGÊNICAS
Nos Estados Unidos os testes de campo iniciaram em 1987 e o primeiro cultivo comercial só ocorreu em 1994 com a liberação do tomate FLAVR SAVR, que apresenta a característica de retardar a maturação. A inserção do gene da poligalacturonase (do próprio tomate) no sentido anti-senso retarda a o acúmulo desta enzima em quantidades suficientes para a degradação das paredes celulares, causando um atraso na maturação.
Não há uma estatística oficial da área cultivada com transgênicos no mundo. Assim, utilizam-se dados de uma organização mantida pelas empresas interessadas. A área plantada com plantas transgênicas saltou de pouco mais de 1,7 milhões de hectares em 1996 para 43 milhões de hectares em 2000 (Tabela 2). Embora o número de países que plantaram transgênicos no ano de 2000 era 12, os três países responsáveis por 98% da produção mundial de grãos transgênicos são os Estados Unidos, a Argentina e o Canadá (Tabela 2). A área total com transgênicos em 2000 foi de 43 milhões de ha. Em 2013 a área cultivada com plantas transgênicas alcançou 175 milhões de ha.
Estima-se que a safra de 2013 as variedades transgênicas alcançaram 79% da área de soja cultivada no mundo, para o algodão 70%; para o milho 32%, e finalmente para a canola, 24%. Embora cultivado em 27 paises, 10 deles (Estados Unidos, Brasil, Argentina, India, Canadá, China, Paraguay, África do Sul, Pakistão e Uruguai) produzem praticamente 98% de toda a aprodução transgência no mundo. Atuamente pouco mais de 10% da área cultivada no planeta são com variedasdes transgênicas. Mas, 171 países do mundo (85%) não cultivam transgênicos. Portanto, o cultivo destas variedades ainda é um fenômeno restrito. Extra oficialmente sabe-se que na China existem dezenas de cultivares transgênicas em cultivo, carregando diferentes características. Contudo, as cifras oficiais são desconhecidas.
Dentre as características introduzidas nestas variedades mais cultivadas destacam-se resistência a herbicidas, plantas inseticidas (produtoras de toxinas danosas a insetos) ou ambas. Projetos de alteração na composição nutricional estão em andamento. Um exemplo disto é o arroz dourado, assim chamado porque foi introduzido numa variedade de arroz um gene que deverá produzir vitamina A. A produção em grandes quantidades da pró-vitamina A no arroz ainda não está garantida, razão pela qual, uma pessoa deveria ingerir quantidades elevadas de arroz (estimativas riam de 1,9 a 4,3 kg/dia) para satisfazer as necessidades diárias deste componente alimentar.
Este quadro não se alterou muito nos últimos anos, pois os dois principais genes são os de resistência a herbicidas ou de produção de toxinas mortais a insetos. Estas cifras sugerem que a tecnologia não se alastrou como se esperava, nem tampouco alcançou a maioria dos paises ou das espécies.
57
Tabela 3.2. Principais países produtores de plantas transgênicas. Área (milhões de ha)/Ano
País 1966 2000 2005 2011 2014
Estados Unidos 1,7 30,0 (70%) 49,8 (55%) 69,0 (43%) 70,1
Brasil 9,4 (10%) 30,3 (19%) 40,3
Argentina 9,0 (21%) 17,1 (19%) 23,7 (18%) 24,4
Índia 1,3 (1,4%) 10,6 (6,7%) 11,0
Canadá 3,0 (7%) 5,8 (6,4%) 10,4 (6,5 %) 10,8
China 3,3 (3,7%) 3,9 (2,4%) 4,2
Paraguai 1,8 (2%) 2,8 (1,8%) 3,6
Pakistão 2,6 (1,6%) 2,8
África do Sul 0,1 0,5 2,3 (1,4%) 2,9
Uruguai 0,3 1,3 (0,8%) 1,5
Bolivia 0,9 1,0
Filipinas 0,1 0,6 0,8
Austrália 0,1 0,3 0,7 0,6
Burkina Faso 0,3 0,5
Myanmar 0,6 0,3
Espanha 0,1 0,1 0,1
México 0,1 0,2 0,1
Colombia 0,1
Sudão 0,1
Outros 8 paises <0,1
Total 1,7 43,0 90 160 181.5
Fonte: ISAAA, 2015 (https://isaaa.org/resources/publications/pocketk/16/default.asp)
Na área total cultivada estão incluídas cultuvares de soja, algodão, milho e canola, cuja percentagem de transgênicos do total para dada uma das espécies é de 82, 68, 30 e 25, respectivamente. As principais razões pela baixa proporção d evraiedades transgênicas de milho e canola em frelaçao ao total é devido a pressão dos consumidores e o alto custo das mesmas.
No ano de 2015 houve uma diminuiçãoo na área cultivada de variedades transgênicas comparativamente ao ano de 2014.
Na Europa existe uma grande controvérsia a respeito de plantas transgênicas que também apresentam genes de resistência a antibióticos estão sendo proibidas para cultivo. No inicio, vários eventos foram aprovados. Posteriroemnete devido a pressão dos consumidores, durante vários anos, nenhuma nova variedade transgênica foi aprovada para plantio ou consumo na Europa.
Alguns eventos que foram autorizados foram depois proibidos como os milhos Bt176 e GA21 x MON810 e as canolas MS1, RF1, MS1xRF1; MS1, RF2, MS1xRF2 e TOPAS19/2. Há países em que é proibido o cultivo (ex: Hungria, Noruega, Itália, Irlanda, outros) ou variedades são proibidas (ex: MOn 810 na França).
E o Brasil ?

58
No Brasil a Soja Roundup Ready (Soja RR), da Monsanto, foi aprovada pela CTNBio (em setembro de 1998), registrada no o Ministério da Agricultura e Abastecimento (junho de 1999), mas como o cultivo e consumo foram suspensos por decisão judicial até que sejam feitos os estudos de impacto ambiental e relatório de impactos no meio ambiente (EIARIMA) e cumpridas outras exigências como elaboração de normas de fiscalização e rotulagem, ela só veio a ser cultivada legalmente a partir de 2003. Para tal, o Governo Federal, desreipeitando as leis ambientais, por meio de Medidas Provisórias, posteriormente aprovadas pelo Congresso Nacional aprovou a colheita da safra ilegal de 2002/2003 e o plantio e colheita da safra 2003/2004. Por fim, a nova Lei de Biossegurança (Lei n° 11.105, de 24 de março de 2005) incluiu artigos que aprovaram o cultivo e o consumo da Soja RR. Mesmo assim, o processo judicial não está concluído (Ferment et al., 2010). É possível que na safra 2009/2010 o cultivo com soja RR alcançou os 50% da área total de soja cultivada no país. Contudo, não há cifras oficiais a este respeito.
Os testes com plantas transgênicas no Brasil anteriormente ao cultivo comercial, em geral, foram destinados para avaliar a performance agronômica ou para melhoramento genético. Pouquíssimos foram destinados a avaliação de risco. O agravante é que foram aprovados testes em grandes áreas experimentais, as vezes em propriedades de agricultores inexperientes no trato com plantas transgênicas, eu ficaram à margem de qualquer fiscalização. Variedades transgênicas de poucas espécies têm sido utilizadas na experimentação no Brasil. Elas se restringem às lavouras de algodão, cana-de-açúcar, fumo, batata, arroz, eucalipto, mamão, milho e soja. As empresas notadamente estão apostando em
três espécies: milho, soja e algodão, mas de fato concentram-se em duas: milho e soja.
Até 2015 foram aprovadas variedades transgênicas listadas na Tabela 3.3.
Tabela 3.3. Eventos de transformação genética em plantas aprovadas pela CTNBio. Espécie Evento de
transformação genética
Transgene(s) Ano Resitência a
Soja GTS 40-3-2 cp4 epsps 2005 Glifosato
Algodão 531 Cry1Ac 2005 Alguns insetos
Milho T25 ou LL25 Pat 2008 Glufosinato de Amônio
Milho MON 810 Cry1Ab 2008 Alguns insetos
Milho BT 11 Cry1Ab, pat 2008 Alguns insetos/ Glufosinato de amônio
Algodão LL25 Pat 2008 Glufosinato de Amônio
Milho NK 603 Mepsps 2008 Glifosato
Milho GA 21 mepsps 2008 Glifosato
Algodão MON 1445 cp4 epsps 2008 Glifosato
Milho TC 1507 Cry1F 2008 Alguns insetos
Algodão 281-24-236/3006-210-23
Cry1F, Cry1Ac 2009 Alguns insetos
Algodão MON 15985 Cry1Ac, Cry2Ab 2009 Alguns insetos
Milho BT11 x GA21 Cry1Ab, mepsps 2009 Alguns insetos/ Glifosato
Milho MON 810 x NK603 Cry1Ab, mepsps 2009 Alguns insetos/ Glifosato
Milho MIR 162 Vip3A 2009 Alguns insetos
Algodão MON 15985 cry1Ac, cry2Ab2, nptII, aad e uidA
2009 Alguns insetos
Algodão MON 531 x MON 1445
Cry1Ac, cp4 epsps 2009 Alguns insetos/ Glifosato
Milho MON 89034 Cry1A.105, Cry2Ab 2009 Alguns insetos
Milho TC 1507 x NK603 Cry1F, mepsps 2009 Alguns insetos/Glifosato
SojaCV127 BPS-CV127-9 Csr1-2 2009 Imidazolinonas
Soja A5547-127 Pat 2010 Glufosinato de amônio
Soja A2704-12 Pat 2010 Glufosinato de amônio
Soja Soja MON 87701 x Cry1Ac e cp4 epsps 2010 Alguns insetos/ Glifosato

59
MON 89788
Milho TC1507 x MON810 Cry1F, Cry1Ab 2010 Alguns insetos
Feijão Embrapa 5.1 Rep e AHAS 2011 BGMV e herbicidas imidazolinonas
Milho MON 89034 x MON 88017
Cry1A.105, Cry2Ab 2011 Alguns insetos
Milho TC1507 x MON810 Cry1F, PAT e Cry1Ab 2011 Alguns insetos e tolerante ao herbicida glufosinato de amônio
Milho TC1507 x MON810 x NK603
Cry1F, PAT, Cry1Ab e cp4 epsps
2012 Alguns insetos e tolerante aos herbicidas glufosinato de amônio e glifosato
Algodão Eventos GHB 614 x T304-40 x GHB119
2mepsps, bar, cry1Ab e cry2Ae
2012 tolerante a herbicidas e resistente a insetos
Algodão GHB614 x LLCotton25
2mepsps e bar 2012 tolerância aos herbicidas glifosato e glufosinato de amônio
Algodão MON 15985 x MON 88913
cry1Ac e cry2Ab2 2012 Alguns insetos e tolerante ao glifosato
Milho TC1507 × DAS-59122-7
Cry1F/pat e Cry34Ab1/Cry35Ab1/pa
2013 Alguns insetos e tolerante aos herbicidas glufosinato de amônio
Milho Bt11xMIR162xMIR604xGA21
Cry1Ab, PAT, VIP3Aa20, mcry3A e mEPSPS
2014 Alguns insetos e tolerante aos herbicidas glufosinato de amônio e glifosato
Milho MIR604 mcry3A 2014 Alguns insetos e tolerante aos herbicidas glufosinato de amônio e glifosato
Milho DAS-40278-9 aad-1v3 2015 tolerante aos herbicidas 2,4-D
Milho NK603 x T25 CP4-EPSPS e PAT 2015 tolerante aos herbicidas glufosinato de amônio e glifosato
Milho TC1507 x MON810 x MIR162 x NK603
cry1F, cry1Ab, PAT, VIP3Aa20 e CP4-EPSPS
2015 Alguns insetos e tolerante aos herbicidas glufosinato de amônio e glifosato
Milho TC1507xMIR162xNK603
cry1F, PAT, VIP3Aa20 e CP4- EPSPS
2015 Alguns insetos e tolerante aos herbicidas glufosinato de amônio e glifosato
Milho TC1507xMIR162 cry1F, PAT e VIP3Aa20
2015 Alguns insetos e tolerante aos herbicidas glufosinato de amônio
Milho MIR162xNK603 VIP3Aa20 e CP4-EPSPS
2015 Alguns insetos e tolerante aos herbicidas glifosato
Milho MON810xMIR162 Cry1Ab eVIP3Aa20 2015 Alguns insetos
Milho TC1507 x MON810 x MIR162
Cry1F, pat, VIP3Aa20 e cry1Ab
2015 Alguns insetos e tolerante aos herbicidas glufosinato de amônio
Milho DAS-40278-9xNK60 AAD-1 e epsps 2015 tolerante aos herbicidas 2,4-D e glifosato
Milho MilhoBt11xMIR162xMIR604xTC1 507x5307xGA21
eCry3.1Ab, cry1Ab, Vip3Aa20, cry3A, cry1F, pat e mepsps
2015 Alguns insetos e tolerante aos herbicidas glufosinato de amônio e glifosato
Milho SPT 32138
zm-aa1, ms45, dsred2(ALT1)
2015 Restauração de fertilidade para producao de sementes
Eucalipto H421 cell1 2015 aumento volumetrico de madeira
Soja DAS-68416-4 aad12 e pat 2015 tolerante aos herbicidas

60
glufosinato de amônio e 2,4-D
Soja FG72 hppd e 2mepsps 2015 tolerante aos herbicidas HPPD e glifosato
Soja DAS44406-6 aad-12, v1, pat e 2mepsps
2015 tolerante aos herbicidas glufosinato de amônio, 2,4-D e glifosato
Soja FG72 x A55547-127 hppd, 2mepsps e pat 2015 tolerante aos herbicidas, HPPD, 2,4-D e glifosato
Milho MON89034xTC1507xNK603xDAS 40278-9
Cry1A.105, Cry2Ab2, Cry1F, PAT, CP4-EPSPS e aad-1
2016 Alguns insetos e tolerante aos herbicidas glufosinato de amônio, 2,4-D e glifosato
Fonte: CTNBio (http://ctnbio.mcti.gov.br/liberacao-comercial/-/document_library_display/SqhWdohU4BvU/view/1684467?_110_INSTANCE_SqhWdohU4BvU_redirect=http%3A%2F%2Fctnbio.mcti.gov.br%2Fliberacao-comercial%2F-%2Fdocument_library_display%2FSqhWdohU4BvU%2Fview%2F614405%3F_110_INSTANCE_SqhWdohU4BvU_redirect%3Dhttp%253A%252F%252Fctnbio.mcti.gov.br%252Fliberacao-comercial%253Fp_p_id%253D110_INSTANCE_SqhWdohU4BvU%2526p_p_lifecycle%253D0%2526p_p_state%253Dnormal%2526p_p_mode%253Dview%2526p_p_col_id%253Dcolumn-2%2526p_p_col_count%253D3#/liberacao-comercial/consultar-processo)
Atualmente, a única restrição legal que existe é de transgênicos que contenham tecnologias genéticas de restrição de uso, também denominadas de GURTs. Alguns tipos de GURTs são conhecidos como “Terminator” pelo fato que as plantas produzem os grãos com o embrião defeituoso. Isto impede a sua germinação e, assim, o agricultor é obrigado a comprar sementes, que são patenteadas, todos os anos.
Das decisões da CTNBio cabe recurso ao Conselho Nacional de Biosegurança (CNBS), órgão formado pro 11 Ministros. Até hoje, houve apenas recursos contra a decisão de aprovação para fiuns comerciais de três eventos transgegênicos. Mas o CNBS decidiu não dar provimento aos recursos do IBAMA e da ANVISA contra a decisão da CTNBio de liberar os seguintes milhos transgênicos:
Evento T25 ou milho LL 25, da Bayer, contendo uma versão sintética do gene pat isolado de Streptomyces viridochromogenes, raça Tü 494, que codifica para a síntese da enzima fosfinotricina – N – acetiltransferase (PAT), enzima esta que catalisa a conversão de L-fosfinotricina, inativando o ingrediente ativo Glufosinato de Amônio e, deste modo, conferindo à planta a resistência ao referido herbicida.
Evento MON 810 ou milho Yeldgard da Monsanto, que contém o gene cry1Ab, proveniente de Bacillus thuringiensis subsp. kurstaki, que codifica a proteína Cry1Ab com efeito tóxico sobre os insetos da ordem Lepidoptera lagarta-do-cartucho, lagarta-da-espiga e lagarta-do-colmo;
Evento Bt 11 da Syngenta, contendo os genes (i) cryIA(b) que expressa uma forma truncada da toxina; (ii) o gene pat que codifica a enzima fosfinotricina-N-acetil transferase que confere resistência ao herbicida glufosinato de amônia (L-Fosfinotricina, PPT - Phosphinothricin), obtido da bactéria de solo Streptomyces viridochromogenes.
Dentre as razões aprresentadas, cabe destacar:
ausência de normas efetivas de monitoramento e de coexistência;
não realização de estudos suficientes para assegurar que não haverá danos aos meio ambiente, notadamente em ecossistemas brasileiros;
apresentação de estudos inconclusivos ou sem sustentação científica (baixa qualidade dos dados aportados);
afirmativas sem comprovação cientifica;

61
não apresentações de dados ou informações solicitadas;
não inclusão de dados da literatura cientifica obtidos por pesquisadores independentes;
insuficiência dos dados apresentados pelos proponentes para garantir a segurança alimentar;
Insuficiência de dados sobre a caracterização do produto de expressão gênica – toxinas de Bt;
Insuficiência de dados que garantam a ausência de efeitos epistáticos e pleiotrópicos resultante do evento de inserção do transgene;
Insuficiência de estudos toxicológicos para comprovar a segurança dos milhos transgênicos acima referidos para o consumo humano.
Além de plantas genetocamente modificadas a CTNBio aprovou para uso comercial outros organismos geneticamente modificados como vacinas, microrganismos e outros. As vacinas recombinantes aprovadas são: Vaxxitek MD/IBD contra a doença de Marek e Gumboro (Merial Saúde Animal Ltda), ProteqFlu e ProteqFlu TE que possui como agentes imunogênicos os poxvírus recombinantes da bouba de canário, carregando o gene da proteína HA do vírus da influenza equina, cepas vCP1533 e vCP2241(Merial Saúde Animal Ltda), INNOVAX® ND para aves contra a Doença de Marek e a Newcastle (Intervet do Brasil Veterinária Ltda), contra a bouba aviária, laringotraqueíte aviária e encefalomielite aviária VECTORMUNE FP-LT+AE (Ceva Saúde Animal Ltda), VECTORMUNE FP-LT, contra a bouba aviária e laringotraqueíte aviária (Ceva Saúde Animal Ltda), VECTORMUNE HVT-IBD contra a Doença de Marek e Doença de Gumboro (Ceva Saúde Animal Ltda), VECTORMUNE® HVT-NDV contra a Doença de Marek e Doença de Newcastle (Ceva Saúde Animal Ltda), VECTORMUNE FP-MG contra a Bouba aviária e Mycoplasma gallisepticum (Ceva Saúde Animal Ltda), VECTORMUNE FP-MG+AE contra a Bouba aviária, Mycoplasma gallisepticum e Encefalomielite Aviária (Ceva Saúde Animal Ltda), INNOVAX® ILT contra a doença de Marek e a Laringotraquíte infecciosa das aves (Intervet do Brasil Veterinária Ltda), Poulvac ST contra Salmonella typhimurium (Fort Dodge Saúde Animal), Poulvac E. coli contra Escherichia coli (Fort Dodge Saúde Animal), Suvaxyn PCV2 One Dose contra Circovirose Suína (Fort Dodge Saúde Animal), Ingelvac Circoflex contra Circovirose Suína (Boehringer Ingelheim do Brasil Química e Farmacêutica Ltda),
Os microrganismos geneticamente modificados aprovados são: linhagem RN1016 de Levedura Saccharomyces cerevisiae para produção de etanol (Bio Celere Agroindustrial Ltda.), Prototheca moriformis para a produção de triglicerídeos e bioprodutos (Solazyme Brasil Óleos Renováveis e Bioprodutos Ltda), levedura (Saccharomyces cerevisiae) para produção de farneseno pela cepa Y5056 (Amyris Brasil SA) e levedura (Saccharomyces cerevisiae) geneticamente modificada para produção de farneseno cepa Y1979 (Amyris Brasil SA), para produção industrial da enzima Fitase e para obtenção da enzima Achromobacter lyticus protease (Novozymes).
Além disso foi autorizado a comercialização de 203 produtos derivados (enzimas) de diferentes organismos geneticamente modificados (Uniscience do Brasil).
8-LIMITAÇÕES Uma das principais limitações da modificação de plantas é a dificuldade de identificar e
isolar genes úteis. A maioria dos genes inseridos em plantas é proveniente de bactérias e vírus porque o reduzido genoma desses organismos facilita a identificação e clonagem de genes. Intensivos estudos em vários laboratórios estão sendo feitos para sequenciar genomas, com diversas finalidades.
Outro fator limitante da transgenia é a necessidade de obtenção de uma planta adulta a partir de uma célula transformada. A regeneração não ocorre em todas as espécies. Nestes casos, a transformação é feita em tecidos cotiledonares. Embora existam muitos métodos de

62
transformação de plantas, algumas espécies são bastante recalcitrantes. Em geral, pode-se transformar a maioria das dicotiledôneas com Agrobacterium tumefasciens. O mesmo não se pode dizer das monocotiledôneas. Para este grupo de plantas utiliza-se um dos métodos diretos. Contudo, para cada espécie ou tecido a ser transformado, há a necessidade de testes sobre o método e o protocolo de regeneração das células ou tecidos transformados.
Embora há precisão no isolamento do gene, não há possibilidade de controlar a integração do inserto no genoma. O local da inserção da construção quimérica pode ser qualquer ponto do cromossomo. Como consequência, poderá ocorrer a interrupção da expressão gênica de um gene da planta se o inserto se integrar no referido loco. Ou ainda, a inserção do gene transferido poderá ocorrer numa região rica em heterocromatina, onde a expressão gênica poderá ser reduzida ou insignificante. Além disso, uma vez inserido, a nova sequência poderá ser alvo de metilação e a consequente inativação em termos de transcrição. Outras vezes, o gene pode ser silenciado ou ocorrer a interferência de outro gene ou inserção (Brasileiro e Dusi, 1999).
Como o número de cópias inseridas é variável, muitas plantas são descartadas por possuírem um número elevado de cópias. Nenhum método é controlável a ponto de possibilitar apenas uma inserção.
Além disso, a inserção do trnsgene no genoma do hospedeiro provoca rearranjamentos tanto no transgene como o genoma do hospedeiro.
Existem vários casos, onde o gene isolado de uma espécie não se expressa adequadamente em outra, em geral devido a diferença na preferência de uso de códons pelas diferentes espécies. Tem-se então, os genes semi-sintéticos. O uso de genes semi-sintéticos é cada vez mais freqüente. Um exemplo é o uso de um gene do Bt (δ-endotoxina) que foi sintetizado in vitro a partir do molde natural e que proporciona resistência a lagarta Heliotis em milho. Testes com plantas transgênicas (com estes genes, parcialmente sintetizados in vitro) já foram concluídos e variedades comerciais já estão sendo cultivadas em vários países.
Após 20 anos do cultivo com plantas transgênicas o quadro que caracteriza a transgenia é a absoluta falta de controle. Em primeiro lugar, as técnicas de modificação transgênica são adequadas para a introdução, mas provocam rearranjamentos porque o DNA transgênico exógeno transferido nas plantas elícita uma resposta, a qual ativa nucleases e enzimas de reparação de DNA (Travik e Heinemann, 2007). Praticamente, em todas as plantas transgênicas em cultivo, o transgene inserido é diferente daquale contido no vetor de transformação. As vezes o transgene inserido é menor, outras vezes há inserção de mais de uma cópia ou pedaços do transgene, e assim por diante. Um exemplo emblemático é o rearranjamento que ocorreu no milho transgênico GA21. Neste, ocorreu a inserção de seis cópias do transgene:
• duas cópias idênticas ao transgene contido no vetor de transformação (cópias 3 e 4); • cópia do transgene com mutação Citonina no lugar de Guanina (cópia 2); • cópia do transgene com mutação Citonina no lugar de Guanina, além de uma
deleção de 696 pb no promotor na região 5‟ (cópia 1); • cópia incompleta, contendo as primeiras 288 (ou 291) pb ou faltando 1050 (ou 1047)
pb do gene mepsps, além de não possuir a terminação NOS (cópia 5) e • cópia contendo o promotor e o primeiro exon truncado da actina do arroz (cópia 6).
Desta forma, a transformação genética de plantas é uma tecnologia não precisa.
9- BIOSSEGURANÇA - REGULAMENTAÇÃO
Segiundo o dicionário Aurélio, Biossegurança. [De bi(o)- + segurança] S. f. Méd. O conjunto de estudos e procedimentos que visam a evitar ou controlar os eventuais problemas suscitados por pesquisas biológicas e/ou por suas aplicações (Dicionário Aurélio (2000). Editora Nova Fronteira, página 302-303).

63
Do ponto de vista técnico, Biossegurança é o conjunto de ações voltadas para a prevenção, minimização ou eliminação dos riscos inerentes as atividades de pesquisa, produção, ensino, desenvolvimento tecnológico e prestação de serviços, riscos que podem comprometer a saúde do Homem, dos animais, das plantas, do meio ambiente Valle e Teixeira (1996).
Na visão da FAO, biossegurança significa o uso sadio e sustentável em termos de meio ambiente de produtos biotecnológicos e aplicações para a saúde humana, biodiversidade e sustentabilidade ambiental, como suporte ao aumento da segurança alimentar global. Desta forma, normas adequadas de biossegurança, análise de riscos de produtos biotecnológicos, mecanismos e instrumentos de monitoramento e rastreabilidade são necessários para assegurar que não haverá danos à saúde humana e efeitos danosos ao meio ambiente.
Em outubro de 1991 a 'European Community' emitiu um documento, o qual inclui os procedimentos para o manuseio dos testes e liberação de organismos transgênicos. Cada Estado membro foi obrigado a estabelecer sua regulamentação (ou legislação) em harmonia com as diretrizes emitidas pela EEC.
A União Européia decidiu, desde 1999, rever as diretrizes de liberação de transgênicos. Contudo, vários países já decretaram ou estão em fase de adotar uma moratória comercial, até que novos estudos sobre biossegurança dos produtos transgênicos indiquem riscos aceitáveis para a saúde humana e ao meio ambiente. Como resultado disto, não houve nenhuma nova liberação para plantio comercial desde junho de 1999. Contudo, as pressões das grandes empresas começam a surtir efeitos e já há indícios de que o processo de liberação de novas variedades transgênicas seja retomado, embora, a contrariedade dos Ministros de Meio Ambiente.
O Brasil aprovou sua primeira norma de biossegurança em 1995. A lei que trata do assunto, Lei n° 8.974 (DOU de 6/1/95), foi votada pelo Congresso Nacional em dezembro de 1994 e sancionada pelo Presidente da Republica em 05 de janeiro de 1995. A lei estabelecia normas de segurança e mecanismos de fiscalização no uso das técnicas de engenharia genética na construção, cultivo, manipulação, transporte, comercialização, consumo, liberação e descarte de Organismos Geneticamente Modificados (OGMs), visando proteger a vida e a saúde do homem, dos animais e das plantas, bem como o meio ambiente. O aspecto mais relevante da lei brasileira diz respeito que o que está sob regulamentação é o produto oriundo da engenharia genética, ou seja, a lei regulamente o produto se oriundo de um processo específico.
No dia seguinte, muitas criticas foram feitas à referida lei (inclusive a do prof Silvio Valle da Fiocruz), em particular porque o Presidente da República, quando sancionou a lei, vetou dois artigos e outros dispositivos. Um dos artigos vetados criava a CTNBio - Comissão Técnica NAcional de Biossegurança. Outro dispositivo vetado dava poder aos Órgãos de Registro e Fiscalização (IBAMA, ANVISA e MAPA) para emitir autorizações sobre o uso de OGMs.
Em 2005, foi aprovada a nova Lei de Biossegurança, a Lei n° 11.105, de 24 de março de 2005. Regulamenta os incisos II, IV e V do § 1º do art. 225 da Constituição Federal, estabelece normas de segurança e mecanismos de fiscalização de atividades que envolvam organismos geneticamente modificados – OGM e seus derivados, cria o Conselho Nacional de Biossegurança – CNBS, reestrutura a Comissão Técnica Nacional de Biossegurança – CTNBio, dispõe sobre a Política Nacional de Biossegurança – PNB, revoga a Lei nº 8.974, de 5 de janeiro de 1995, e a Medida Provisória nº 2.191-9, de 23 de agosto de 2001, e os arts. 5º, 6º, 7º, 8º, 9º, 10 e 16 da Lei nº 10.814, de 15 de dezembro de 2003, e dá outras providências.
O fato mais relevante foi a inclusão do Principio da Precaução no Artigo 1º da lei, embora ele não é observado pela CTNBio:

64
Esta Lei estabelece normas de segurança e mecanismos de fiscalização sobre a construção, o cultivo, a produção, a manipulação, o transporte, a transferência, a importação, a exportação, o armazenamento, a pesquisa, a comercialização, o consumo, a liberação no meio ambiente e o descarte de organismos geneticamente modificados – OGM e seus derivados, tendo como diretrizes o estímulo ao avanço científico na área de biossegurança e biotecnologia, a proteção à vida e à saúde humana, animal e vegetal, e a observância do princípio da precaução para a proteção do meio ambiente.
A lei traz ainda artigos sobre definições, proibição, composição e atributos do Conselho Nacional de Biossegurança (CNBS) e da Comissão Técnica Nacional de Biossegurança (CTNBio), além das atribuições dos órgãos e entidades de registro e fiscalização.
Outra importante inclusão foi o principio da publicidade. Na gestão das informações de biossegurança, há que ser observada a transparência. Da mesma forma, a Legislação, atos administrativos; processos em andamento; decisões da CTNBio, do CNBS e dos órgãos de registro e fiscalização; atas das reuniões e outras informações consideradas não sigilosas, bem como os votos fundamentados de cada membro deverão ser tornados públicos. Entretanto, este dispositivo ainda não foi implementado.
Poucos países da América do Sul têm legislação referente aos testes e a comercialização de produtos oriundos da engenharia genética. Na Argentina não existe uma lei de Biossegurança semelhante a do Brasil. Apenas um decreto. No Paraguai, uma portaria do governo criou uma comissão de biossegurança que tem também representantes da universidade e de organizações não governamentais. Em um de seus primeiros atos, a Comissão de Biossegurança não autorizou a introdução da Soja RR da Monsanto no Paraguai.
Nos Estados Unidos também não existe uma lei específica. Basicamente as leis já existentes foram emendadas para tratarem também dos produtos transgênicos. Como neste país, o processo de transgenia não é considerado relevante, pois é considerado similar ao melhoramento genético de plantas. Se um produto transgênico é considerado equivalente a um não transgênico, os testes exigidos são de comum acordo entre as agencias governamentais e as empresas, estando os consumidores totalmente fora das decisões. O sistema após análise desregulamenta o produto. O processo de concessão de autorização é baseado no fenótipo da planta, na segurança ambiental, utilização do produto e risco do produto.
Uma pergunta frequente tem sido: a liberação destas plantas nos EUA foi precedida por testes rigorosos e análises rigorosas das agências americanas Food and Drug Administration - FDA, Environmental Protection Agency - EPA e United States Department of Agriculture - USDA?
Basicamente, a agência americana encarregada da alimentação, a FDA, não aprova, nem autoriza, simplesmente afirma o que o proponente do OGM menciona. Abaixo está um exemplo. Pontos mencionados na carta para o evento MON 810 (milho Bt), datada de 26 de Setembro de 1996, enviada pela FDA a Monsanto:
“Monsanto submeteu um resumo da avaliação do milho contendo o evento de transformação MON 810 em 6 de Junho de 1996”;
“Baseado na avaliação de segurança e nutricional que vocês conduziram, é nosso entendimento que a Monsanto concluiu que os produtos derivados desta nova variedade de milho não é materialmente diferente em composição, segurança, e outros parametros relevantes de outros milhos que estão no mercado, e que o milho geneticamente modificado não levanta questões que requerem uma revisão antes da comercialização e sua aprovação por parte da FDA.” www.cfsan.fda.gov/~acrobat2/bnfL034.pdf.

65
Estas duas sentenças acima podem ser encontradas em todas decisões da FDA como respostas às consultas sobre desregulamentação de plantas transgênicas. Portanto, ao contrário do que é afirmado com frequência, a FDA não requer avaliação de risco na fase de pré-comercialização e não emite sua própria opinião a respeito da segurança da variedade transgênica.
As plantas transgênicas, aprovadas para o cultivo comercial nos Estados Unidos, tiveram sua liberação baseada no princípio da equivalência substancial. Assim, a soja RR foi considerada “equivalente” a sua antecedente natural, a soja convencional, porque não difere dela nos aspectos cor, textura, teor de óleo, composição e teor de aminoácidos essenciais e de nenhuma outra qualidade bioquímica. Desta forma, não foram submetidas à rotulagem pela agência americana encarregada de sua liberação, a FDA.
Este conceito de equivalência substancial tem sido alvo de críticas, entre outras, porque a falta de critérios mais rigorosos pode ser útil à indústria, mas é inaceitável do ponto de vista do consumidor e da saúde pública (Millstone et al., 1999). Há dificuldades práticas no conceito de equivalência entre plantas engenheiradas e naturais, ou obtidas por técnicas convencionais de melhoramento genético. Equivalência significa dispor de igual valor ou outro atributo, normalmente expresso em unidades ou parâmetros: um grama do produto Y equivale a X calorias. Equivalência se refere sempre a quantidade ou algo mensurável a que corresponde um sentido tecnicamente comparável (Momma, 1999). A rigor, em termos de genoma, elas não são equivalentes nem iguais. Só seriam iguais se uma fosse originária da outra por multiplicação vegetativa ou micropropagação. A construção genética inserida na planta contém elementos bastante distintos daqueles naturais encontrados nas plantas, que proporcionam novos produtos gênicos e que podem desencadear efeitos pleiotrópicos substanciais, para que sejam considerados desprezíveis.
Por este critério, a vaca louca seria equivalente, em termos de segurança, a vaca sadia, já que a diferença entre ambas é apenas da conformação espacial de uma proreina.
Uma das criticas se originou da análise da documentação que foi utilizada pela FDA para considerar a Soja RR substancialmente equivalente a soja convencional. Segundo Barbara Keeler que fez a análise, existem diferenças significativas entre soja não transgênica e Soja RR: em 3 dos seis macronutrientes; em um ácido graxo; 29% menos de choline; mais (27%) de inibidor de tripsina, um potente alergênico. Para chegar a conclusão de que ambas variedades eram equivalentes, não foram aplicados testes estatísticos nas comparações. Além disso, em um dos 3 experimentos feitos em Porto Rico foi omitido da publicação no Journal of Nutrition, mas os dados foram submetidos ao FDA. Estes revelaram que a Soja RR apresentou menor nível de proteína e de fenilalanina; o inibidor de tripsina foi 18% maior nas tortas tostadas a base de Soja RR que nos controles e as lectinas apareceram em dobro. Neste caso, por esta análise a soja convencional e a Soja RR não seriam equivalentes.
Quando se utiliza a equivalência substancial, nenhum teste é requerido para excluir a presença de toxinas prejudiciais, carcinogênicas e mutagênicas. Este critério da equivalência substancial é equivocado, carece de base científica e deveria ser abandonado em favor de testes biológicos, toxicológicos e imunológicos mais aprofundados e eficazes (Guerra e Nodari, 1999). Com base nesta equivalência, o FDA exige apenas testes de curta duração com animais e testes bioquímicos para avaliar, entre outros, a alergenicidade. Esta insuficiência de dados, que não consegue subsidiar cientificamente a análise da segurança alimentar, está sendo questionada não só pela população em geral, mas também por grande parte da comunidade científica e agora (outubro de 2000) pelos governos, como é o caso da Itália.
Como o transgene é, na verdade, uma nova característica – em geral desconhecida – introduzida num genoma cultivado que vem sendo lapidado pelas seleções natural e artificial, ainda não há experiência acumulada, nem conhecimento suficiente para tratar adequadamente este assunto. Contudo, a comunidade científica e os agricultores já têm experiência acumulada com os agroquímicos ou agrotóxicos que foram liberados, após a

66
Segunda Guerra Mundial para uso, sem a realização de testes adequados de biossegurança. Só posteriormente, parte dos efeitos nefastos causados por eles se tornaria conhecido. Foi preciso a morte e a dor de inúmeras pessoas contaminadas para que as restrições de uso aumentassem. Até hoje não houve reparação alguma por partes das empresas fabricantes destes produtos às vitimas intoxicadas ou mortas (Nodari e Guerra, 2001).
A equivalência substancial é utilizada também pelo Canadá e Argentina. Nestes países a rotulagem não é obrigatória. A rigor, todos os processos de solicitação de liberações comerciais já provadas pela CTNBio foram tqmbém julgados pela equivalência substancial, contrariando a norma legal brasileira. Nem mesmo os princípios e a metodologia estabelecidos no Anexo III do protocolo de Cartagena sobre Biossegurança têm sido seguido. De um lado as empresas não fazem os estudos recomendados, de outro lado a CTNBio não exige. Assim, nem a comunidade cientifica dispõe de informações técnico-científicas a respeito dos riscos. Isto contribui para um debate na sociedade, vazio de informações científicas e técnicas.
Protocolo de Cartagena sobre Biossegurança
Os países membros da Convenção sobre Diversidade Biológica (CDB) em janeiro de 2000. Os dois principais pontos são: (i) o princípio da precaução deve ser adotado em caso de dúvida ou falta de conhecimento científico e (ii) os produtos transgênicos devem ser rotulados (art. 18a). O referido protocolo tem cerca de 40 artigos e trata basicamente da movimentação de transgênicos entre países, com atribuição de responsabilidades em caso de danos. Garante ainda, que o país importador recuse o produto caso não esteja acompanhado de estudo de risco adequado. Um terceiro aspecto, explicitado no artigo 15 e anexo II, impõe que a análise de risco seja conduzida cientificamente pelo exportador. Na ausência desta análise, os importadores podem se negar a receber os produtos.
Já foram realizadas seis reuniões (denominadas de MOP), nas quais foram tomadas decisões consensuadas sobre vários temas, sendo o mais polêmico os requisitos em termos de informação sobre o OGM que deve acompanhar o documento fiscal nos carregamentos de OGM em movimentos transfronteiriços.
Até o final de 2015, 170 países haviam ratificado o Protocolo de Cartagena sobre Biossegurança, incluindo o Brasil. Mas não ratificaram o Protocolo, Estados Unidos e Argentina, por exemplo.
Situação em Santa Catarina
A primeira é a Lei Promulgada Nº 11.403, de 10/05/2000, que dispõe sobre pesquisas, testes, experiências ou atividades nas áreas de Biotecnologia e Engenharia Genética e adota outras providências.
Seu Art. 1º diz “As empresas nacionais ou estrangeiras, que desenvolverem no Estado de Santa Catarina pesquisas, testes, experiências e outras atividades nas áreas da biotecnologia e engenharia genética, envolvendo Organismos Geneticamente Modificados (OGMs), bem como os produtos advindos desta tecnologia, deverão notificar o Poder Executivo na forma disposta nesta Lei.” Já o Art. 3º proíbe a comercialização em todo o Estado de Santa Catarina dos produtos advindos da tecnologia.
A segunda é a Lei Promulgada 11.643, de 4/06/2000, que cria o Conselho Técnico Catarinense de Biossegurança – CTCBio – e adota outras providências. Seu Art. 1º diz: “ Fica criado o Conselho Técnico Catarinense de Biossegurança – CTCBio –, órgão normativo-jurisdicional, consultivo e de assessoramento vinculado diretamente ao Poder Executivo, com a finalidade de deliberar sobre matéria relacionada a sua área de competência.”

67
Ambas as leis embora vigentes ainda não foram totalmente implementadas até esta data. O CTCBio chegou a ser criado, mas poucas vezes se reuniu ou foi demandado.
Decorrente da mobilização da sociedade civil e da sensibilidade dos governantes e dos legisladores na época foi possível aprovar uma lei de consenso no estado de Santa Catarina, a Lei nº 12.128, de 15 de janeiro de 2002. No Art. 1º consta:
No Estado de Santa Catarina, durante cinco anos, a partir da publicação desta Lei, fica vedado o plantio e cultivo para fins industriais e comerciais de organismos geneticamente modificados - OGM - e seus derivados, que tenham como finalidade a alimentação humana ou animal. § 1º Decorrido o prazo de cinco anos, a Assembléia Legislativa verificará a necessidade ou não da prorrogação do período de moratória.
Embora previsto, a lei não só não foi cumprida como também o plebiscitoo que deveria ter ocorrido não foi realizado.
Esta lei também contém outros dispositivos de interesse dos consumidores. Em seu Art. 2º diz:
Os produtos alimentícios que contenham ou provenham de organismos geneticamente modificados e seus derivados somente serão industrializados e/ou disponibilizados em estabelecimentos comerciais, no Estado de Santa Catarina, caso expressem no recipiente, embalagem e rótulo, a informação de que no seu processo produtivo utilizaram-se técnicas transgênicas.
Com base neste dispositivo a rotulagem é obrigatória, independente da quantidade de OGM nos alimentos. Questionado na justiça pela empresa NESTLÉ, o judiciário manteve o entendimento de que em Santa Catarina osalimentos que são OGMs ou feito a partir destes, indpendente do percentual, devem ser rotulados.
Este movimento da sociedade civil organizada foi muito ativo no período de 2000 a 2004, estando atualmente sem atividades. Disso resultou o avanço do poder político ligao aos interesses dos proponentes de atividades com os transgênicos. Exemplo disso foi a revogação do inciso IV do art. 8º da Lei nº 12.128, de 15 de janeiro de 2002, pela Lei nº 14.675, de 13 de abril de 2009, que criou o polêmico Código Ambiental de Santa Catarina, ainda sob ação de inconstitucionalidade no Supremo Tribunal Federal.
Dizia o inciso IV do art. 8º da Lei nº 12.128:
IV - realização do Estudo e do Relatório de Impacto Ambiental EIA/RIMA relativo às atividades desenvolvidas, devidamente aprovado.
Ou seja, uma conquista da sociedade civil foi descumprida pela Assembléia Legislativa que deixou de exigir estudos nos ecossistemas catarinenses para avaliar ou não seus impactos.
10-FISCALIZAÇÃO
Apesar da Legislação Brasileira de Biossegurança ter sido promulgada desde 1995 e da CTNBio ter sido implantada em 1996 e re-implantada em 2005, a operacionalização da fiscalização dos produtos transgênicos nas Unidade da Federação tem enfrentado várias dificuldades. A fiscalização tanto de experimentos quanto de área plantada clandestina não está sendo feita a contento.
Mesmo com a nova lei em 2005, pouco ou nada mudou. Fatos comprovados por jornalistas e mesmo pela fiscalização comprovaram a existência de algodão e milho transgênicos antes de terem sido liberados no país. Assim, a fiscalização é praticamente ineficiente para proteger o país de cultivos ilegais e de contaminação por transgenes.

68
A manchete de capa do jornal Folha de São Paulo de 10/05/2009 dizia: Brasil não tem controle sobre milho transgênico.
Figura 3.2. O agricultor Ademir Ferronato em sua plantação de milho convencional, no Paraná; ele teme contaminação por lavoura transgênica (Fonte: FSP, 10/05/2009).
11-ANÁLISE DE RISCO
As Biotecnologias têm sido utilizadas por milênios para diversos propósitos, incluindo as fermentações para produção de alimentos e bebidas e a seleção de novas variedades de plantas ou animais. Na última metade do século passado, novas biotecnologias foram desenvolvidas, dentre as quais merecem destaque a micropropagação, a fusão de protoplastos, os marcadores moleculares, a clonagem de animais, DNA recombinante e a transgenia. Conseqüentemente, a precisão e o poder de manipulação dos organismos vivos aumentoaram consideravelmente com o avanço da genética molecular. De todas elas, o que causa maior apreensão é a transgenia, não em si pela tecnologia, mas pelas implicações que seus produtos podem apresentar à saúde humana e ao meio ambiente.
Se um transgênico é diferente de uma variedade comum e o transgene nele inserido pode apresentar um determinado risco, há a necessidade da avaliação do risco, tanto para a saúde humana como para o meio ambiente. A razão disto está no fato de que os genes transferidos de fora do gene-pool de uma espécie produzem produtos com os quais temos pouca ou nenhuma experiência. Não se conhecem as implicações que podem ser provocadas pela introdução desses genes em plantas. Desta forma, há um consenso entre os pesquisadores que a sociedade precisa desenvolver regras para o desenvolvimento, testes e comércio de OGMs.
Embora a transformação genética transfira somente sequências curtas de DNA, comparativamente ao genoma de uma variedade, o fenótipo resultante, que inclui a característica transgênica, é possivelmente acompanhado de mudanças nas características e pode produzir um organismo novo em termos de relações ecológicas (Wolfenbarger e Phifer, 2000). Segundo estes autores, os ecossistemas são complexos e nem todo o risco associado com a liberação de um OGM pode ser identificado e considerado. Os testes a serem realizados, os protocolos mais apropriados, os termos de referência, os instrumentos mais adequados ainda são pouco conhecidos e estão sendo discutidos e desenvolvidos.
Risco pode ser definido como uma medida dos efeitos de uma ocorrência em termos de sua probabilidade e da magnitude de suas conseqüências. Assim, a avaliação de risco („risk assessment‟) como sendo o processo com base científica que consiste na identificação e caracterização dos perigos, da avaliação da exposição e da caracterização dos efeitos dos

69
riscos. Por perigo entende-se a propriedade de uma substância ou processo que cause dano. Ou seja, dano é a materialização do perigo. Então, se o potencial de dano é elevado, mesmo uma baixa probabilidade pode significar um risco inaceitável.
A avaliação de segurança deve ser baseada nos riscos potenciais impostos pelo produto obtido (Fontes et al., 1996). Assim, a avaliação deve levar em consideração as características do doador, do recipiente, ou quando apropriado, do organismo parental. Devem ainda ser avaliadas as características e a utilização pretendida do OGM, incluindo a escala e a freqüência das introduções e considerações ambientais e de saúde.
O manejo dos riscos deve levar em conta as alternativas decorrentes da avaliação de riscos e, se necessário, a seleção e implementação de opções de controle apropriadas, incluindo normas regulatórias. Os danos podem ser diretos ou indiretos, intencionais ou involuntários, imediatos ou não. Segundo o Dr. Chris Glidon, espera-se, ao final do processo, eliminar ou reduzir o risco que possa causar um dano de fato. A diretriz maior é a de que o produto deve ser seguro e sadio para a espécie humana e para o meio ambiente. Portanto, o impacto de um transgene no ambiente e na saúde humana deve ser criteriosamente avaliado (Glidon, 1999).
Pode-se também definer Risco como sendo a medida dos efeitos (injúrias, ambientais, econômicos) de uma ocorrência em termos de probabilidade e da magnitude de suas conseqüências. Neste caso, um OGM poderia ser POTENCIALMENTE PERIGOSO, em razão de apresentar, como propriedade, uma substância ou processo que causa dano (injúria ou perda). Assim, DANO seria a manifestação de uma substância ou processo perigoso. Tais danos podem se diretos ou indiretos, imediato ou longo prazo, naturais ou tecnológicos e intencionais ou imprevisíveis.
Em tese, os riscos não estão relacionados ao que os cientistas sabem, mas ao que eles não sabem (Caruso, 2006). Desta forma, riscos estão associados a incertezas. Neste mesmo sentido “é no contexto da incerteza que viceja a esperança, o juízo e a valoração da subjetividade, capaz de concretizar o inusitado”, segundo Lieber e Romano-Lieber (2003).
Já em 1989, pelo menos 15 anos antes da liberaçao no meio ambiente da primeira planta transgenica, o tomate Flavr Svr, Tiedje et al. (1989) anteciparam os sete principais riscos ambientais:
• criação de novas pragas e plantas daninhas;
• um aumento das pragas já existentes por meio da recombinação gênica entre a planta transgênica e outras espécies filogeneticamente relacionadas;
• a produção de substâncias que são ou poderiam ser tóxicas a organismos não-alvos;
• o efeito disruptivo em comunidades bióticas e o desperdício de valiosos recursos genéticos, seguido de contaminação de espécies nativas com características originadas de parentes distantes ou de espécies não relacionadas e efeitos adversos em processos dos ecossistemas;
• origem de substâncias secundárias tóxicas após a degradação incompleta de químicos perigosos;
• efeito adverso nos processos ecológicos;
• extravagância de recursos biológicos valorosos. Praticamente todos os efeitos adversos previstos ocorreram com os OGMs liberados. Portanto, não é correto dizer que os mesmo são imprevistos, pois os efeitos adversos ou os danos foram alertados por parte da própria comunidade cientifica.
A Resolução Normativa Nº 2 da CTNBio, de 27 de novembro de 2006, dispõe sobre a classificação de riscos de Organismos Geneticamente Modificados (OGM) e os níveis de biossegurança a serem aplicados nas atividades e projetos com OGM e seus derivados em contenção. Para efeitos desta Resolução Normativa (Art. 3º), considera-se Risco "possibilidade de promoção de evento negativo, cientificamente fundamentada, para a saúde

70
humana e animal, os vegetais, outros organismos e o meio ambiente, decorrente de processos ou situações envolvendo OGM e seus derivados".
A norma definiu ainda as classes de risco que os transgenicos devem ser enquadrados em seu Art. 8º - As classes de risco dos OGM serão assim definidas:
I – Classe de Risco 1 (baixo risco individual e baixo risco para a coletividade): O OGM que contém seqüências de ADN/ARN de organismo doador e receptor que não causem agravos à saúde humana e animal e efeitos adversos aos vegetais e ao meio ambiente;
II – Classe de Risco 2 (moderado risco individual e baixo risco para a coletividade): O OGM que contém seqüências de ADN/ARN de organismo doador ou receptor com moderado risco de agravo à saúde humana e animal, que tenha baixo risco de disseminação e de causar efeitos adversos aos vegetais e ao meio ambiente;
III – Classe de Risco 3 (alto risco individual e risco moderado para a coletividade): O OGM que contém seqüências de ADN/ARN de organismo doador ou receptor, com alto risco de agravo à saúde humana e animal, que tenha baixo ou moderado risco de disseminação e de causar efeitos adversos aos vegetais e ao meio ambiente;
IV – Classe de Risco 4 (alto risco individual e alto risco para a coletividade): O OGM que contém seqüências de ADN/ARN de organismo doador ou receptor com alto risco de agravo à saúde humana e animal, que tenha elevado risco de disseminação e de causar efeitos adversos aos vegetais e ao meio ambiente.
Mas foi só em março de 2008 que a CTNBio elaborou e aprovou a RESOLUÇÃO NORMATIVA Nº 05, que dispõe sobre normas para liberação comercial de Organismos Geneticamente Modificados e seus derivados. Cabe destacar que nesta norma estão explicitados os procedimentos e os tópicos que os proponentes devem avaliar para solicitar a liberação de um transgênico no país. Dentre os dispositivos desta norma cabe destacar:
Art. 6º. Para efeitos desta Resolução Normativa considera-se: I – avaliação de risco: combinação de procedimentos ou métodos, por meio dos quais se avaliam, caso a caso, os potenciais efeitos da liberação comercial do OGM e seus derivados sobre o ambiente e a saúde humana e animal.
Art. 19. A avaliação de risco, conforme definida no art. 4º, inciso I, desta Resolução Normativa, deverá identificar e avaliar os efeitos adversos potenciais do OGM e seus derivados na saúde humana e animal, no ambiente e nos vegetais, mantendo a transparência, o método científico e o princípio da precaução.
Riscos à saúde humana e animal
A maioria das plantas transgênicas desta primeira geração de OGMs contém genes de resistência a antibióticos, cuja função é possibilitar a seleção das células transformadas. O que os genes de resistência a antibióticos tem a ver com a saúde humana? Nos últimos 20 anos, mais de 30 novas doenças ocorreram na espécie humana (AIDS, ebola e hepatites, entre outras). Além disso, houve o ressurgimento de doenças como a tuberculose, malária, cólera e difteria com muito mais agressividade por parte dos microrganismos patogênicos. Paralelamente, houve um decréscimo na eficiência dos antibióticos. Nos anos 40, um antibiótico tinha uma vida útil de 15 anos. Nos anos 80, a vida útil passou para cinco anos, ou seja, três vezes menos. Os estudos comprovam de que tanto a recombinação como a transferência horizontal entre bactérias acelerara a disseminação de regiões genômicas destes organismos causadores de doenças, bem como a disseminação de genes de resistência a antibióticos (Ho et al., 1998). É bem conhecido o exemplo da estreptomicina em suínos. Após um ano de aplicação aos animais (1983), genes de resistência a estreptomicina estavam presentes nos plasmídeos de bactérias que viviam na garganta e estômago dos

71
suínos. Um ano mais tarde, bactérias humanas dos familiares que lidavam com estes animais também apresentaram resistência a estreptomicina. Esta é uma prova inequívoca de transferência lateral de genes entre bactérias. Em 1990, este antibiótico foi retirado de circulação.
Embora a frequência de transformação e, consequentemente, a transferência horizontal em bactérias é extremamente baixa, os genes de resistência a antibióticos inseridos em plantas transgênicas, poderão ser transferidos para bactérias humanas, o que se constitui num risco a ser considerado. Tem sido sugerido o desenvolvimento de OGMs sem genes de resistência a antibióticos para evitar os riscos acima mencionados. Cabe então o aperfeiçoamento do sistema de seleção tanto via desenvolvimento de outras formas de seleção ou utilização de outros genes.
Um segundo tipo de risco relaciona-se com as reações adversas dos alimentos OGMs ingeridos, que podem ser agrupadas em duas categorias: alergênicos e intolerantes. Neste grupo estão os alimentos que causam hipersensibilidade ou alergia. No segundo grupo estão as alterações fisiológicas, como reações metabólicas anormais, toxicidade, reações farmacológicas e idiossincráticas (Finardi, 1999).
Os resultados dos primeiros experimentos sobre os efeitos da inclusão de derivados de OGM na ração animal feitos por pesquisadores independentes começam a ser analisados. Segundo o jornal britânico The Guardian, de 04/11/2000, os pesquisadores Steve Kestin e Toby Knowles, da University of Bristol, verificaram que a mortalidade de frangos alimentados com milho transgênico foi praticamente o dobro (7,14%) comparativamente à mortalidade de frangos tratados com milho convencional (3,57%). Os cientistas questionaram ainda os métodos e conclusões dos estudos da Aventis submetidos para análise das autoridades britânicas visando à liberação do milho transgênico. Contudo, estes resultados ainda devem ser validados cientificamente, pois este tipo de experimento deve ser efetuado para diferentes combinações de nutrientes, raças e condições climáticas.
É ilustrativo o caso do milho transgênico StarLink (da Aventis CropScience) um tipo de Bt que contém o gene rCry9C, aprovado pela Environmental Protection Agency (EPA). Dos Estado Unidos, para alimentação animal mas não para consumo humano. Este milho contém uma proteína (Cry9C) que pode causar reações alérgicas em humanos, uma vez que ela não foi quebrada imediatamente nos testes de digestão. Tanto grãos quanto subprodutos foram misturados com grãos não-transgênicos, conforme análise de produtos alimentícios de consumo humano. Além disso, houve também a contaminação de colheitas que deveriam ser não-transgênicas devido à disseminação do pólen.
Para os transgenes que produzem toxinas recombinantes (rCry) a partir da bactéria Bacillus thuringiensis (Bt) transferido para o milho, Sagstad et al. (2007) constatou que o salmão alimentado MON 810 tinha uma grande proporção de granulócitos, monócitos, crescimento somático, com uma menor proporção de linfócitos, as mudanças nas atividades das proteínas de estresse e alterações nas populações de leucócitos associada a resposta imune.
Do ponto de vista de efeitos pleiotrópicos, Zolla et al. (2008) verificaram diferenças na expressão de proteínas entre o evento Mon810 (que contém o transgene rCry1Ab) e o seu isogênico não GM. Os autores constatam alterações estatisticamente significativas em 43 proteínas. Dentre elas, uma nova proteína expressada no milho GM corresponde a gama zeina (50 kDa), uma proteína alergênica conhecida.
Recentemente, Andreassen et al. (2015), constataram que a exposição a rCry1Ab purificada resultou em reação específica IgG1 anti-Cry1Ab e produção de IgE, indicando imunogenicidade inerente e alergenicidade. Ratos expostos a extratos de folhas de ambos MON810 e milho não modificado demonstraram influxo de linfócitos e eosinófilos no lavado broncoalveolar, bem como no aumento da liberação de citocinas em células de nódulos linfáticos do mediastino. Os autores também concluíram que os resultados indicaram que a exposição das vias respiratórias às proteínas rCry1Ab pode ser uma via de relevância prática.

72
Então faz sentido saber se uma nova variedade transgênica intensifica ou não a alergia. No caso da Soja RR, os testes realizados não foram suficientes para discriminar as possíveis variações nas 16 proteínas alergênicas desta espécie. Os testes revelaram que houve um aumento (26,7%) do inibidor de tripsina, também alergênico e antinutricional (Padgette et al., 1996), além de uma maior reatividade de uma banda relativa a uma proteína alergênica. Segundo a análise feita por uma pesquisadora independente, Barbara Keeler, a documentação que a empresa forneceu a FDA demonstra que em um dos experimentos também o teor de lectina, que é alergênico, produzido pela Soja RR foi maior (o dobro) que na convencional. O desafio neste caso é sabe quais os tipos de ensaios que fornecem os dados mais inequívocos sobre alergenicidade.
Estudos em ratos alimentados com soja RR (Malatesta et al., 2002) na Itália, mostrou, por exemplo, a formação de micronúcleos, núcleos de forma irregular e também um grande número de poros nucleares, sugerindo uma alta taxa metabólica. A publicação deste estudo custou o emprego da pesquisadora.
Em um estudo recente de toxicidade a longo prazo de um herbicida e de milho geneticamente modificado tolerante Roundup, os resultados demonstram claramente que os níveis mais baixos de formulações comerciais de herbicidas de glifosato, em concentrações bastante abaixo dos limites de segurança estabelecidos oficialmente, induzem graves distúrbios hepáticos e renais, dependentes de hormonio (Seralini et al., 2012). A relevância deste estudo baseia-se: (i) os tumores cancerígenos aparecem aos 4 meses em ratos machos e aos 7 meses e ratas femeas.Todavia, as agências exígem estudos de apenas 3 meses; (ii) as alterações bioquímicas e às falhas fisiológicas são mais graves em ratos alimentados com milho transgênico ou com Glifosato que quando alimentados com milho convencional. Assim, o critério da equivalência substancial não tem sustentação científica. (iii) As alterações bioquímicas e as falhas fisiológicas elevaram a probabilidade de desenvolvimento de tumores em ratas. Em razão da grande pressão de cientistas favoráveis a transgenia e das grandes empresas de biotecnologia a revista decidiu retirar o referido artigo com base no argumento de que o estudo era inconclusivo. Esta atitude foi a primeira na história da Ciência. Posteriomente, o artigo foi publicado pela Revista Environmental Sciences Europe.
Dentre os muitos avanços científicos cabe destar dois estudos. O primeiro refere-se a constatação feita por Agapito-Tenfen et al. (2013) de que há proteínas expressas a mais ou a menos e também novas proteínas expressas no milho transgênico (MON 810, que carrega o transgene rCry1Ab) comparativamente à variedade não transgênica. Neste estudo, foram constados, entre outros, que houve (i) uma nítida redução de níveis de transcrição para todos os três transgenes empilhados (EPSPS, Cry1A.105 e cry2Ab2) e (ii) alteração na expressão de 17 proteínas, sendo cinco presentes em um ou dois dos genomas estudados. No entanto, as alterações nos níveis de expressão dos transgenes de um evento empilhado podem afetar a sua segurança e utilidade. Assim, não há dados suficientes sobre a correlação entre a acumulação de mRNA e os níveis de proteínas transgênicas, o que impede de fazer inferências sobre os possíveis riscos deste tipo de evento.
Em soja, os estudos feitos po Bohn et al. (2013) demonstraram que a soja RR não é equivalente a soja convencional, pois examinando mais de 30 variedades cultivadas em sistema orgânico, convencional e transgênicas, os teores de elementos da composição centesimal das sojas foram estatisticamente diferentes. Além disso, no graãos das variedades transgênicas foram encontrados entre 10 e 20 ppm de glifosato ou AMPA, comparativamente a zero nas variedades cultivadas convencionalmente ou no sistema orgânico.
Em 2013 houve a comprovação de que tanto toxinas de Bt produzidas em plantas transgênicas quanto resíduos do glifosato aplicado sobre plantas estavam presentes em fetos de mulheres grávidas (Aris e Leblanc, 2011). Isto significa que esta tecnologia já está provocando exposição de pessoas não nascidas a substâncias tóxicas. Embora o assunto é de grande relevância, as autoridades governamentais estão ignorando completamente as consequências desta exposição em humanos intrauterinos.

73
Existe ainda uma série de outros riscos à saúde humana que devem ser analisados com protocolos adequados. Um deles é o efeito tóxico que um alimento transgênico pode causar à saúde humana. Riscos ao meio ambiente
A avaliação de risco ambiental é a avaliação sistemática dos riscos associados à saúde e à segurança humana e ambiental. Os procedimentos devem incluir a identificação dos perigos e a estimativa de suas magnitudes e freqüências de ocorrência, bem como das alternativas ao OGM. Como os riscos associados a uma variedade transgênica dependem das interações complexas decorrentes da modificação genética, da história natural dos organismos envolvidos e das propriedades do ecossistema no qual o OGM é liberado (Peterson et al., 2000; Wolfenbarger e Phifer, 2000), estes procedimentos devem ser aplicados em escala ampla, em termos espaciais e sociais (ver Figura 3.2).
O conhecimento dos riscos também é indispensável porque possibilita a elaboração de planos de seu gerenciamento. O manejo dos riscos é um processo que envolve a análise das alternativas decorrentes dos resultados alcançados com a avaliação destes. Quando requerido, o manejo seleciona e implementa opções apropriadas de controle, incluindo normas reguladoras (Glidon, 1999). Assim, o manejo de riscos deve também fazer parte do estudo de impacto ambiental para fins de licenciamento de atividades com plantas transgênicas.
Na ausência de efeitos pleiotrópicos, os efeitos diretos do transgene numa planta seriam razoavelmente previsíveis. Quando os biólogos moleculares dizem que foram feitos estudos e não foram detectados efeitos adversos, eles normalmente estão se referindo à primeira das várias células possíveis de serem analisadas (Figura 3.2). Existem também estudos de parcela (segunda célula da Figura 3.2), associados predominantemente à performance agronômica do OGM, e que, a rigor, não podem ser tomados como estudos de impactos e riscos ambientais. Não há estudos científicos relacionados a todas as células relevantes desta matriz. Existem sim, relatos científicos de estudos isolados com algumas espécies e que serão apresentados mais adiante.
A complexidade da avaliação é decorrente do fato de que os riscos e os benefícios associados a uma cultura específica mudam e tornam-se mais difíceis de serem avaliados na medida que a área de cultivo aumenta e outros aspectos são considerados. Impactos indiretos nos ecossistemas são muito mais difíceis de investigar, monitorar e, portanto, predizer (Peterson et al., 2000). Segundo estes autores, esta é uma das origens da controvérsia estabelecida entre os ambientalistas e os biólogos moleculares. Enquanto os primeiros referem-se aos impactos sociais e nos ecossistemas, os últimos fazem menção aos testes feitos com uma ou poucas plantas em laboratório ou em casa de vegetação.
A complexidade também é decorrente do fato de que inúmeros trabalhos científicos demonstraram que o padrão de variação fenotípica, sua base genética e a seleção natural sobre eles variam em diferentes condições ambientais (Griffiths et al.,2015; Ackerly et al., 2000). O problema da biologia é que, em contraste com outros ramos do mundo físico, nos quais poucas grandes forças dominam os fenômenos, o organismo vivo é resultante de um grande número de caminhos fracos causais determinantes, fazendo com que seja extremamente difícil proporcionar explanações completas (Lewontin, 2000). Em seu recente texto, o autor afirma ainda que um organismo vivo num momento qualquer de sua vida é a conseqüência única da história do desenvolvimento que resulta de interações e determinações de forças internas e externas.

74
Figura 3.2. Efeitos diretos e indiretos de variedades transgênicas (OGM) e as interações
complexas que fazem parte da avaliação de risco ambiental (Adaptado de Peterson et al., 2000).
Entre os riscos ambientais, a poluição genética, por meio da transferência vertical e da transferência horizontal, é a ameaça considerada mais importante. Em decorrência disto, espécies que adquirirem certos transgenes poderão alterar seu valor adaptativo e, conseqüentemente, a dinâmica de suas populações e de outras espécies as quais interagem estará desafiada. Contudo, outros riscos são possíveis como efeitos danosos em espécies não-alvo (aves, minhocas, peixes, entre outros), contaminação de solo e água, cujas dimensões também são impossíveis de prever antes dos estudos a serem realizados (Nodari e Guerra, 2000a).
Nestes 20 anos de cultivo de plantas transgênicas já foram observados danos aos organismos não alvo, contaminação de variedades crioulas, aumento no uso de agrotóxicos, aumento no número de plantas resistentes a herbicidas, aparecimento de novas pragas, entre outros.
Contaminaçao genetica
Refere-se ao acasalamento entre indivíduos sexualmente compatíveis, geralmente da mesma espécie e, raramente, de espécies afins. O acasalamento é uma via para o fluxo gênico, entre plantas da mesma espécie, como entre plantas de diferentes espécies. Assim, de longa data têm sido observados cruzamentos entre indivíduos de populações em estado incipiente de especiação ou de espécies aparentadas. Exemplos disso são os cruzamentos entre o arroz cultivado e o arroz perene, milho e teosinto, um de seus possíveis ancestrais (Doebley, 1990), beterraba cultivada e beterraba não domesticada e entre espécies cultivadas e inços do gênero das abóboras (Wilson, 1990).
Os impactos ecológicos da transferência de pólen, um mecanismo reprodutivo pelo qual a introgressão pode ocorrer, dependem da capacidade dos híbridos em sobreviver e reproduzir. Taxas de sobrevivência ou de reprodução indicam a oportunidade da introgressão de transgenes em populações naturais, dependendo do fluxo gênico subseqüente e da pressão de seleção (Wolfenbarger e Phifer, 2000). Estes autores relataram 11 casos de formação de híbridos entre variedades transgênicas e plantas aparentadas e/ou daninhas. Para se tornar uma ameaça, como uma planta invasiva, os híbridos precisam ser viáveis e competitivos, além de férteis quando dependem da reprodução sexual para propagação. Com base no se conhece hoje, nem todos os híbridos vão atingir a última fase.
Os poucos estudos associados à introgressão de transgenes e suas conseqüências ecológicas em populações naturais ainda não permitem fazer previsões confiáveis. Contudo, a experiência anterior com plantas de lavoura sugere que os efeitos negativos são possíveis. Para doze das treze espécies de maior importância econômica mundial, a hibridização com parentes selvagens contribuiu para a evolução de algumas espécies de ervas daninhas. Em alguns casos, os elevados níveis de introgressão a partir de parentes cultivados ou

75
introduzidos eliminaram a diversidade genética e contribuíram para sua extinção (Ellstrand et al., 1999).
Quando são viáveis e havendo fertilidade, mesmo baixa, a sobrevivência dos híbridos interespecíficos se torna possível, e estes podem cruzar com plantas de qualquer uma das duas espécies parentais. Caracteriza-se, então, o processo de introgressão de genes de uma espécie para outra. No caso do cruzamento entre canola transgênica e a mostarda silvestre, o número de sementes da segunda geração do híbrido foi dez vezes maior do que o F
1.
Algumas plantas descendentes do cruzamento produziram 10 mil sementes e o gene de resistência ao herbicida ainda permanecia numa grande quantidade de plantas. Isto demonstra que a transferência de genes que condicionam resistência a herbicidas pode ocorrer com maior intensidade e facilidade do que se imaginava antes desta descoberta (Chèvre et al., 1998).
Uma vez dentro de populações silvestres, os transgenes poderão tornar estas plantas mais invasivas e, portanto, potencialmente perigosas para a agricultura ou a biodiversidade (Fontes et al., 1996). Mas também pode ocorrer, segundo as autoras, que a presença do transgene diminua a adaptação natural, o que tornaria a população vulnerável à extinção. No caso de transferência de outras características para outras espécies afins, praticamente nada pode ser antecipado, devido à ausência de dados. Contudo, se o valor adaptativo de um híbrido interespecífico for aumentado com a presença deste gene transferido, é factível que tal gene se mantenha via introgressão.
O número de contaminações de variedades crioulas ou mesmo convencionais por transgenes aumenta todo o ano. Um conjunto de organizações da sociedade civil vem acompanhando e registrando estas contaminações (www.gmcontaminationregister.org). Entre 1997 e 2006 ocorreram 107 contaminações genéticas; 24 cultivos ilegais e 8 efeitos colaterais agrícolas negativos. Destes 144 casos comprovados, envolveram 44 países, sendo a media de 14,2 ao ano. O mais espantoso é que 35% ocorreram com milho, que é um alimento nobre.
A liberaçao da soja RR no Brasil prejudica quem nada ter a ver com isso: os produtores orgânicos, por exemplo. Abaixo está o relato de um entre centenas de casos já comprovados.
Dedicado ao cultivo de produtos orgânicos, sem agrotóxicos e com sementes naturais, por mais de 30 anos, o agricultor Max Enro Dockhorn, de 73 anos, desistiu, no ano passado, da lavoura de soja que mantinha em uma área de 70 ha no município gaúcho de Três Passos. "Na safra de 2005 para 2006 perdi metade da minha produção orgânica. No momento de vender, testes identificaram proteína transgênica na minha soja", conta Dockhorn, desapontado com os meses de dedicação à lavoura. Além da perda de valor, que superava os 10 reais por saca, ele teve de pagar royalties por ter sido acusado de usar sementes transgênicas. “...bastou que, ao redor de minha propriedade, outros produtores usassem sementes transgênicas para haver a contaminação". Os riscos da omissão, Revista Carta Capital, p.22-29, 18/07/2007
Diante disso começaram as preocupações com a coexistência. A coexistência significa a possibilidade efetiva, para os agricultores, de escolherem entre o modo de produção convencional ou biológico, ou ainda a produção de culturas GM, no respeito das obrigações legais em matéria de rotulagem ou de normas de pureza. A rigor, é impossivel ocorrer a coexistência sem contaminação.
A CTNBio baixou a Resoluçao Normativa n° 4, de 16 de agosto de 2007 e publicada no DOU, n° 163 de 23/08/2007, p.19. Nela esta estabelecido que “Para permitir a coexistência, a distância entre uma lavoura comercial de milho geneticamente modificado e outra de milho não geneticamente modificado, localizada em área vizinha, deve ser igual ou superior a 100 (cem) metros ou, alternativamente, 20 (vinte) metros, desde que acrescida de bordadura com, no mínimo, 10 (dez) fileiras de plantas de milho convencional de porte e ciclo vegetativo similar ao milho geneticamente modificado” (art. 2).

76
Ironicamente ou intrigantemente, no mesmo dia a CTNBio aprovou o evento MON810, milho transgenico, por meio do Parecer Técnico nº 1.100/2007, de 16 de agosto de 2007. Nele, está escrito que “Comparando-se as concentrações a 1 m da cultura fonte sob ventos baixos a moderados estimou-se que, aproximadamente, 2% de pólen são anotados a 60 m, 1,1% a 200 m e 0,75-0,5% a 500 m de distância. Ou seja, a RN n° 4 é totalmente ineficiente para garantir a coexistência sem contaminaçao caso o que está contido no próprio parecer da CTNBio, o pollen do milho deve se disseminar pelo menos a 500 m de distância.
Não só o cultivo de variedades melhoradas não-transgênicas, mas a agrodiversidade, que pode ser definida como a diversidade de espécies agrícolas, composta de variedades crioulas mantidas pelos agricultores, também pode ser ameaçada pelo cultivo dos transgênicos. Na análise dos riscos está sendo ignorada uma realidade fundamental: o pólen de milho pode ser carregado pelo vento até 9,6 km. Segundo o professor Walter Fehr, melhorista da Iowa State University, "não é somente o que você faz. É também o que seu vizinho faz", ressaltando que “agricultura é vizinhança”, quando se trata de identificação, segregação e rotulagem de cultivos transgênicos. Com esta mobilidade do pólen, uma simples lavoura de transgênicos pode contaminar várias outras não-transgênicas, numa área relativamente grande.
Em diversos municípios do Sul do Brasil, estão sendo organizadas anualmente Feiras de Sementes. Na segunda edição de uma delas, realizada em 15 de julho de 2000 em Porto União (PR), 49 representantes de comunidades situadas em 13 municípios expuseram amostras de 41 variedades crioulas de milho e 46 de feijão, para citar apenas duas das 51 espécies identificadas na referida feira. Surpreendentemente, formas de teosinte também são mantidas pelos agricultores daquela região. Assim como esta, uma ampla diversidade de espécies e formas dentro de espécies é exposta ano a ano nestas feiras de sementes. Ensaios com variedades crioulas feitas por técnicos da Emater/RS, em David Canabarro, revelaram que seu potencial chegou a mais de seis toneladas por hectare (Dados não publicados). Além do rendimento, estas variedades crioulas contêm uma ampla gama de características, com alta variabilidade genética, estando continuamente submetidas ao processo evolutivo e gerando, anualmente, novas recombinações.
Estudos recentes feitos pro professores e estudantes do Programa de Pós-graduação em Recursos Genéticos Vegetais detectou tanta diversidade genética no Oeste de Santa Ctarina, que a região pode ser enquadrada como um micro centro de diversidade genética de milho (Costa et al., 2016). A equipe de estudos encontrou 1513 variedades crioulas, sendo 1078 de oipoca e 337 de milho. Além disso, tipos ancestrais do milho, os tesosintes, também foram encontrados na região.
Esta agrodiversidade deve ser considerada nas avaliações de riscos ambientais. O mínimo que se pode fazer é informar aos agricultores o que poderá acontecer com seus materiais, caso transgênicos sejam cultivados nas proximidades e levar em consideração a opinião deles. Toda esta rica diversidade está ameaçada em razão dos interesses de uns poucos. Transferência horizontal ou lateral (TH)
Quando existe transferência de genes entre espécies filogeneticamente diferentes, na ausência do acasalamento sexual, configura-se a transferência lateral ou transferência horizontal. Neste caso, o material genético é transmitido de uma espécie para outra, provavelmente com auxílio de vetores (plasmídios, transposons e vírus). Elementos similares a transposons são veículos para cortar e ligar DNA genômico de um organismo noutro. Vírus também poderiam ser responsáveis pela transmissão de genes entre eucariotos. Na verdade, os mecanismos de transferência lateral são pouco estudados e, portanto, praticamente desconhecidos.

77
Diversos casos de absorção de DNA por parte de células eucariotas foram também registrados (Tappeser et al., 1999). Num deles, foi demonstrado que o DNA fornecido na alimentação de ratos não só não era totalmente destruído no trato gastrointestinal, mas também poderia alcançar a corrente sangüínea e temporariamente ser detectado nos leucócitos ou células do fígado. Outros exemplos de detecção de DNA de eucariotos em bactérias e animais, como DNA de milho transgênico em bactérias de intestino de abelhas ou DNA de milho transgênico em vários órgãos de galinhas, estão sendo noticiados pela imprensa, mas necessitam aparecer em publicações científicas ou serem validados cientificamente. A transferência horizontal é bem mais conhecida em bactérias, sendo os eventos menos comuns em animais e no homem comparativamente a plantas e microrganismos.
Experimentalmente, Nielsen et al. (2000) verificaram que o DNA de beterraba transgênica pode ser transferido para Acinetobacter sp. Strain BD413, uma bactéria de solo. Neste caso, a TH ocorreu de um extrato celular para plasmídeos de bactérias. Casos de transferência via recombinação homóloga são mais freqüentes do que se imaginava (Nielsen et al., 1998).
Uma pergunta comumente feita relaciona-se com as conseqüências da introdução em plantas de genes (intactos ou modificados) originados de vírus patogênicos. Trocas de material genético também podem ocorrer entre plantas e vírus. A primeira evidência experimental sobre a recombinação entre uma planta transgênica contendo genes virais e um vírus foi obtida por Greene e Allison, em 1994, embora este tipo de recombinação já fosse conhecido desde os anos 80. A introdução de genes que codificam a capa protéica originada de vírus patogênicos, ou outras seqüências virais, é utilizada para conferir às plantas resistência aos próprios vírus doadores. É difícil estabelecer as conseqüências, caso este gene seja transferido para outras plantas. Contudo, um vírus poderá infectar um planta transgênica que tem a proteína do encapsulamento de outro vírus. Neste caso ocorrerá uma transencapsidação, cujas conseqüências são totalmente desconhecidas.
Recentemente também, um estudo com arroz transgênico, conduzido no John Innes Institute, da Inglaterra, corroborou a evidência de que o promotor do vírus do mosaico-da-couve-flor (CaMV), que também está presente na maioria das plantas transgênicas e nas suas progênies, é um sítio de alta freqüência de recombinação gênica. Recombinação gênica é a troca de material genético entre duas moléculas de DNA, altamente similares geneticamente, que pode resultar numa terceira molécula diferente das duas parentais, e, portanto, um variante. O mais intrigante, entretanto, é que os autores verificaram que a maioria dos eventos era do tipo de recombinação “ilegítima” ou não-homóloga e não requeriam uma similaridade substancial na seqüência de bases. Tais eventos podiam ocorrer mesmo na ausência de genes virais (Kohli et al., 1999). Além disso, a seqüência de bases do promotor do CaMV, usado em várias plantas transgênicas, como a soja e o milho, é similar a regiões de vírus patogênicos à espécie humana. Desta forma, não se pode descartar a possibilidade de recombinações entre o transgene e outros vírus, resultando em novas combinações genéticas, cujas propriedades não são conhecidas, mas que necessitam ser estudadas antes do cultivo em larga escala de plantas que contêm estas seqüências. A priori, não se pode descartar, então, que a inserçãoinserção de seqüências virais em plantas poderá tornar os vírus mais promíscuos e com isto provocar mais doenças em plantas.
Embora não se conheça a magnitude da contribuição da engenharia genética para a transferência horizontal, é possível levantar a hipótese de que o cultivo em larga escala de plantas transgênicas deve favorecer a TH. Geralmente, as plantas transgênicas contêm elementos mediadores da transformação in vitro, ou parte deles, e também da TH, como plasmídeos, transposons e vírus. Os vetores utilizados para a obtenção de plantas transgênicas freqüentemente apresentam na construção quimérica origem de replicação, seqüências de transferência, promotores fortes e genes de resistência a antibióticos. Todos estes elementos facilitam a recombinação e a transferência de genes. Plasmídeos e vírus quiméricos estão sujeitos a instabilidades estruturais, o que facilita também a recombinação

78
(Ho et al., 1998). Na natureza, a poluição com metais pesados pode se constituir em fator benéfico para a transferência de genes. Como parte das seqüências introduzidas são homólogas a muitos procariotos, a transferência de material genético para eles via recombinação é factível. Dependendo das seqüências introduzidas na planta transgênica, haverá uma maior ou menor probabilidade de favorecimento para a TH.
Outro aspecto importante está relacionado com a freqüência de ocorrência da TH.
Embora, algumas estimativas sejam baixas, como 2x10-17
, o número de cópias em cultivo poderá ser muito alto. O fato de que uma planta pode conter mais de dois trilhões de células, e um hectare de soja mais de 300 mil plantas, permite supor a probabilidade da existência de
mais de 1,2 x 10-18
de cópias por hectare, de um transgene. Considerando o cultivo em pelo menos cinco milhões de hectares, não é difícil concluir que uma ou mais recombinações podem de fato ocorrer, mesmo porque, a probabilidade de sua ocorrência, embora baixa, é finita, ou seja, tem um valor que é influenciado por vários fatores.
São duas, então, as principais implicações da TH. A primeira refere-se à maior probabilidade de transferência horizontal de genes a partir de plantas transgênicas comparativamente às variedades tradicionais. A segunda refere-se ao fato de que os genes com potencial de disseminação podem dar vantagem seletiva aos organismos receptores, o que poderá alterar dramaticamente a dinâmica das populações e a paisagem. Como ainda não é possível determinar a probabilidade de um evento de TH ocorrer, bem como suas conseqüências, torna-se praticamente impossível fazer qualquer previsão realística na ausência de novos estudos.
Ameaças diretas aos componentes da biodiversidade
As ameaças aos componentes da biodiversidade são múltiplas, pois, em um ecossistema devem ser considerados não somente os organismos vivos, mas também os processos ecológicos.
Um trabalho que causou grande impacto na comunidade científica avaliou o efeito do pólen de milho transgênico em lagartas da borboleta monarca (Danaus plexippus). A taxa de mortalidade destas lagartas atingiu 44% quando se adicionaram ao seu alimento natural folhas de Asclepias curassavica, pólen de uma variedade de milho transgênico, que contém um gene de Bacillus thuringiensis (Bt) que codifica para uma toxina, que é tóxica a vários insetos. Entretanto, todas as lagartas que receberam pólen de milho não-transgênico ou nenhum pólen, sobreviveram (Losey et al., 1999). O trabalho recebeu críticas metodológicas, porém, um ano depois, resultados semelhantes foram obtidos em experimentos no campo. Neste caso, o pólen das variedades de milho transgênicas KnockOut (evento 176) e YieldGard (Bt 11), ambos da Novartis Seeds, também provocou mortalidade (Hansen Jesse e Olbrycki, 2001).
Também se conhece pouco sobre as possíveis alterações na associação entre plantas e fungos micorrízicos. O primeiro estudo sobre os exudatos na rizosfera de plantas transgênicas foi publicado recentemente (Saxena et al., 1999). Nesse trabalho observou-se que as toxinas inseticidas Bt podem permanecer ativas no solo, onde se ligam a argila e ácidos húmicos. Mesmo ligadas a estes componentes do solo, as toxinas mantêm suas propriedades inseticidas e são protegidas contra a degradação por microrganismos porque estão ligadas às partículas do solo, onde podem persistir por pelo menos 234 dias. Quais são as implicações destes fatos?
Nas regiões de ocorrência natural de alta diversidade genética de uma espécie ou espécies afins, como é o caso de algodão, milho ou amendoim no Brasil, o cultivo de plantas transgênicas destas espécies merece análise mais rigorosa. No México, por exemplo, ainda não foi liberado o cultivo comercial de milho transgênico, devido à existência de extensas áreas com populações ancestrais e parentes silvestres da espécie. O Brasil é ainda berço de várias espécies cultivadas ou apresenta regiões com alta variabilidade genética nas

79
populações crioulas ainda em cultivo, situação esta que requer muita cautela. Como avaliar adequadamente este tipo de risco é sem dúvida um grande desafio.
A determinação de riscos de plantas transgênicas que contêm inseticidas é complexa. Não se conhece ainda profundamente o efeito destas sobre insetos ou outros organismos benéficos. Tampouco, os poucos estudos sobre pássaros ou outros animais que se alimentam de insetos que se alimentam de plantas transgênicas não proporcionam um conhecimento amplo do assunto.
Já há considerável literatura cientifica sobre os efeitos adversos do produto expresso do transgene rCry1Ab em organismos não alvos. Alguns exemplos são aqui resumidos. Hilbeck et al. (1998) estudaram a toxicidade da toxina rCry1Ab em um agente de controle biológico Chrysoperla carnea. O estudo demonstrou que 57% das larvas de C. carnea morreram quando alimentadas com uma dieta contendo toxina Cry1Ab. Em um estudo com camundongos alimentados com milho GM MON863, observou-se que eles mostraram-se fracos, com sinais de toxicidade hepatorrenal, com 24-40% de aumento dos triglicerídeos nas fêmeas e diminuição de 31-35% da excreção de sódio e fósforo na urina em machos (Seralini et al., 2007). Abelhas (Apis mellifera) expostas a diferentes concentrações da proteína rCry1Ab (3 e 5000 ppb) mostraram comportamento alterado depois de expostas a maior concentração de proteína rCry1Ab e levavam mais tempo para absorver o xarope contaminado com a proteína rCry1Ab (Ramirez-Romero et al., 2008). Um estudo ecotoxicológico com Daphnia magna alimentada com milho MON810, que contém uma versão do transgene rCry1Ab, demonstrou que esta sofre uma redução significativa do valor adaptativo (fitness) comparado com as alimentadas com o milho isogênico. A mortalidade dela foi maior, a proporção de fêmeas que alcançaram a maturação sexual foi menor e a proporção de ovos foi menor em comparação a D. magna alimentada com milho isogênico não GM (Bøhn et al., 2008). Bøhn e colaboradores (2010) realizaram outro estudo com a mesma espécie alimentada com milho MON810. Os autores observaram que nas populações de D. magna alimentadas com ração contendo milho GM a taxa de crescimento, sobrevivência e a fecundação nos primeiros estágios de vida foram desfavorecidas. Rosi-Marshall et al. (2007) detectaram uma diminuição na taxa de crescimento ou mesmo mortalidade de espécies anfíbias aquáticas. Desta forma, neste momento seria recomendável que a empresa proponente realizasse estudos similares com os transgenes envolvidos nesta proposta nos distintos backgrounds genéticos. Rosi-Marshall et al. (2007) detectaram uma diminuição na taxa de crescimento ou mesmo mortalidade de espécies aquáticas. Desta forma, neste momento seria recomendável que a empresa proponente realizasse estudos similares com os transgenes envolvidos nesta proposta nos distintos backgrounds genéticos.
Em resumo, existe ameaça a diversidade biológica decorre da liberação de um OGM devido as propriedades do transgene ou de sua transferência e expressão em outras espécies. A adição de um novo genótipo numa comunidade de plantas pode proporcionar vários efeitos indesejáveis: deslocamento ou eliminação de espécies não domesticadas, exposição de espécies a novos patógenos ou agentes tóxicos, poluição do pool gênico, erosão da diversidade genética e interrupção da reciclagem de nutrientes e energia.
Riscos socioeconômicos, com ênfase na agricultura
Dentre eles, os mais relevantes seriam o aumento da população de pragas e microrganismos resistentes e/ou patogênicos, o aumento ou promoção de plantas espontâneas (indevidamente denominadas de plantas daninhas) resistentes a herbicidas, a contaminação de variedades crioulas mantidas pelos agricultores, a contaminação de produtos naturais como o mel, a diminuição da diversidade em cultivo com o aumento da vulnerabilidade genética, a dependência dos agricultores a poucas empresas produtoras de sementes, igual ou menor produtividade e os altos preços das novas variedades.
Um fato é inquestionável: os insetos que são susceptíveis a toxinas de Bt por ocasião do lançamentp das variedades transgênicas, serão no futuro resistentes a uma ou mais

80
toxinas de Bt. Resta saber em quanto tempo. Se houver uma grande área plantada com variedades transgênicas resistentes a um inseto, somente os resistentes sobreviverão, gerando progênies recombinantes, que eventualmente apresentarão maior nível de resistência à toxina. Após vários ciclos de recombinação, deverão aparecer insetos resistentes ao gene Bt. No caso de esta resistência ser condicionada por genes dominantes, a velocidade do aumento da freqüência dos alelos de resistência é extraordinariamente maior, comparativamente àquela observada para alelos recessivos. Já existem mais de 500 insetos resistentes a inseticidas. Porque então as toxinas recombinantes de Bt também não iriam selecionar insetos resistentes? Mas esta foi a promessa, sem base cientitica. Veja abaixo.
Com o cultivo em larga escla de plantas produtoras de toxinas, cria-se uma superpraga, como já ocorreu com o uso de agrotóxicos. O fato de que a resistência da lagartas de lepidópteras às formulações comerciais de Bt (ex: Dipel) seja controlada por um gene parcialmente dominante (Huang et al., 1999) indica que o sistema de refúgio só será efetivo por poucos anos, porque a maioria da progênie dos insetos será resistente à toxina e, portanto, atacará as variedades Bt. Conforme já era esperado do ponto de vista cientifíco, o aumento de frequência de insetos pragas resistentes as toxinas de Bt vem aumentando proporcionalmente ao aumento da área em cultivo das variedades transgênicas que produzem estas toxinas. Em levantamento feito e publicado plea Revista Nature Biotechnology, Tabashnik et al. (2013) já relataram a ocorrência de cinco espécies de insetos praga resistentes as toxinas BT.
O que de fato acontece com a frequência dos insetos resistentes, alvos e não-alvos, nas condições nas condições de cultivos de plantas transgências que produzem toxinas recombinantes já está constatado pelos estudos científicos. A resistência a campo às toxinas Bt em cultivos GM foi relatada pela primeira vez em 2006 para S. frugiperda em Puerto Rico (ex: Storer et al., 2010). Muitos outros casos de resistência a campo foram confirmados posteriormente (ex: Huang et al., 2011; Gassmann et al., 2011; Kruger et al., 2011). As causas da resistência foram atribuídas principalmente à falta de cumprimento dos produtores que não seguem estritamente as exigências para o plantio de áreas de refúgio com variedades não-GM. Entretanto, segundo os autores dos quatro estudos acima referidos, a dose de toxina poderia ter sido muito baixa ou variável para matar consistentemente insetos resistentes heterozigotos. Por exemplo, estudos anteriores revelaram que a variação sazonal e espacial do teor de toxinas rCry em algodão GM tem sido frequentemente ligada a características das plantas e às condições ambientais (ex: Showalter et al., 2009). Neste mesmo ano, outro estudo demonstrou em milho Bt, que concentrações da toxina Cry diminuíram à medida que as plantas avançaram no estádio de crescimento, mas as mudanças sazonais na concentração de toxina são variáveis entre toxinas e cultivares (Nguyen et al., 2009). As razões para a redução sazonal da concentração da proteína rCry permanecem obscuras, mas podem estar relacionadas com a instabilidade do mRNA, a declínio da atividade do promotor, a redução do metabolismo do nitrogênio, a menor produção global de proteínas, e as interações de toxina (ex: Chen et al., 2005; Olsen et al., 2005). Neste ano de 2015, um novo estudo (Reisig e Reay-Jones, 2015) concluiu que não houve redução em peso larval, número de insetos ao entrar no estádio pupal, peso de pupas, tempo para eclosão e número de pupas capaz de eclodir com sucesso para a vida adulta de Helicoverpa zea no híbrido expressando rCry1Ab em comparação com um hibrido isogênico. Como as toxinas de rCry1Ab vem afetando estes insetos desde 1996, H. zea pode estar desenvolvendo resistência a rCry1Ab no milho, embora esses resultados não são abrangentes, pois os dado são limitados ao período de amostragem (que foi de dois anos), tamanho e geografia da área cultivada. Os autores também verificaram que os impactos negativos sobre o crescimento e desenvolvimento larval foi maior no milho híbrido com empilhado comparativamente aos eventos simples.
O aumento dos custos de produção já é uma realidade em vários países. Na China, por exemplo, a estratégia de refúgio para algodão Bt transgênico não foi empregada pelos agricultores. Como resultado, as pragas secundárias se tornaram importantes e o custo com inseticidas aumentou a tal ponto de que a rentabilidade das tecnologias convencional ou

81
transgênica se equivalem cinco anos após sua implementação (Wang et al., 2006). O aumento do uso dos agrotóxicos nos cultivos transgênicos foi decorrente da alteração do status de algumas pragas que eram secundárias e passaram a ser primárias e predominantes.
Sobre a rápida evolução de pragas secundárias tornarem-se primárias na China, a empresa atribui o fato à inexistência de um programa de manejo de insetos. Segundo o mesmo artigo, em certas regiões da China, o custo com inseticidas em lavouras de algodão Bollgard aumentou a tal ponto que a rentabilidade das tecnologias convencional ou transgênica se equivaleram cinco anos após sua implementação.
Uma prática sugerida pelos proponentes da tecnologia é o uso de refúgio, que consiste no plantio de uma bordaura que varia de 10 a 20% da área cultivada com variedades não transgênicas e, portanto, variedades susceptíveis a algumas pragas, o que permitiria o acasalamento entre insetos susceptíveis e resistentes. Uma das premissas para que o sistema seja duradouro, é que a resistência dos insetos à toxina Bt deve ser recessiva. Em caso contrário, rapidamente os alelos de resistência serão prevalentes. Com o aumento rápido da frequência de insetos resistentes ao Bt, o uso atual de formulações comerciais à base de Bt em lavouras orgânicas fica comprometido, como também a produção de produtos com este tipo de inseticida, considerado muito menos tóxico que os demais.
O cultivo de plantas transgênicas também podem provocar o aparecimento de novas pragas. Exemplo disso, foi o cultivo sucessivo de uma ou poucas variedades em grandes áreas levou a uma grande epidemia da Helicoverpa armigera nos plantios de variedades transgênicas de algodão e soja, notadamente nos estados do nordeste. Este espécie não era considerada praga nem da soja nem do algodão anteriormente ao cultivo de variedades transgênicas.
A transgênia também pode levar ao aumento de pragas de solo. Na cultivar transgênica de algodoeiro, Paymaster 1560 BG, resistente ao glifosato, observou-se um aumento na susceptibilidade ao nematóide-das-galhas (Meloidogyne incognita Kofoid e White), quando comparado com o parental não-transgênico Paymaster 1560 (Colyer et al., 2000). Outro estudo nos Estados Unidos, indicou que as variedades transgênicas que carregam a toxina rCry3Bb1 ou a mCry3A foram atacadas pela Diabrotica virgifera, a larva ocidental do milho (Western Corn Rootworm)(Grassman et al., 2013). Os resultados destes trabalhos também indicam a necessidade de estudos sobre a reação de plantas transgênicas às pragas e doenças antes da liberação para cultivo.
A dinâmica das populações de microrganismos de solo também poderá ser afetada pelo cultivo de plantas transgênicas. O uso de glifosato combinado ou não com outros herbicidas nas doses recomendadas sobre o cultivo de Soja RR apresentou maior incidência de fusarium nas raízes uma semana após a aplicação, comparativamente à soja não-transgênica que não recebeu (Kremer et al, 2000). Os testes que foram realizados no campo no período 1997-2000 revelaram que a freqüência de fusarium nas raízes aumentou de 0,5 a 5 vezes entre a segunda e a quarta semana após a aplicação dos herbicidas. O fusarium causa a síndrome da morte repentina (SDS) em soja.
Outro impacto negativo é o aumento de plantas resistentes aos herbicidas. Desde o inicio do cultivo de variedades transgênicas tolerantes a herbicidas o número de espécies ou populações de plantas resistentes resistentes ao glifosato tem aumentado dramaticamente, o que também tem provocado um aumento no uso do referido herbicida. Na pagina da internet da Weed Science (www.weedscience.org) já foram registrados estudos que confirmaram dezenas de populações de plantas pertencentes a mais de 25 espécies que são comumente denominadas de plantas daninhas e já resistentes a um ou mais herbicidas (ex: glifosato). Isto requer o uso de outros herbicidas que por sua vez causam danos ambientais significativos nos elementos da biodiversidade e de processos ecológicos.
O processo submetido pela empresa proponente da Soja RR, menciona que a liberação da variedade não era prov;avel que plantas daninhas resistentes a herbicidas a base de glifosato se tornasse um problema. Entretanto, nenhum estudo foi aportado. Al‟;em disso,

82
já na época da decisão da liberação comercial da soja RR, a CTNBio contatou que dentre as mais de 100 plantas resistentes a herbicidas, três delas são plantas daninhas resistentes a formulações comerciais à base de glifosato: poaia-branca (Richardia brasiliensis), trapoeraba (Commelina virginica) e erva-quente (Spermacoce latifolia) (CTNBio, 1998).
Mais do que isso, a própria soja RR se tornou uma planta invasora, porque os grãos que ficam na susperficie do solo após a colheita germinam e são resistentes ao glifosato.Isto está obrigando os agricultores a utilziar outros herbicidas igualmente ou mais tóxiucos que o glufosato. O fato de empresas produtoras do herbicida 2,4-D terem solicitado registro para uso deste produto visando pó controle da soja RR como planta invasoa é a demonstração do fato.
A soja RR também comçou a perder a competitividade para a soja convencional ainda em 2008 no Brasil. Pela primeira vez, os produtores de soja convencional tiveram mais rentabilidade do que os de soja transgênica. A Confederação Nacional da Agricultura (CNA) revelou que, este ano de 2008, a comercialização da saca de soja convencional deverá render ao produtor R$ 0,27 a mais do que a da convencional no Mato Grosso. A explicação para a inversão é o aumento de 46,2% no preço do glifosato, principal herbicida utilizado na cultura. No Brasil, a Monsanto é praticamente a única empresa a comercializar o glifosato, com cerca de 90% do mercado. Apesar do aumento do custo, vamos insistir na produção do transgênico. Não podemos perder este mercado, porque há oportunidades para os dois produtos - disse Fábio de Sá Meirelles, da CNA. Com o aumento do preço, o custo de produção ficou extremamente alto - disse Meirelles. O preço do litro do glifosato no Mato Grosso passou de de R$ 8,00 para R$ 11,63 o litro na safra 2007/2008, o que gerou uma acréscimo de 23% nas despesas em lavouras transgênicas e de 14,3% nas convencionais. O resultado foi um aumento de 7,5% no custo operacional da soja geneticamente modificada e de 3,8% no da convencional. (Cláudia Dianni Viviane Monteiro - Jornal do Brasil 19/12/2007).
Posteriormente, a análise dos custos de produção realizados pela EMBRAPA no Mato Grosso do Sul para a safra 2010/2011 também indicou que a soja convencional tem menores custos de produção que a soja transgênica. As estimativas de custo consideram dois sistemas de produção, sendo um com soja convencional e outro com soja transgênica (RR). Os custos de produção da soja convencional foram estimados em R$ 1.187,60 e os da soja transgênica (RR) em R$ 1.219,86. O custo de produção na soja transgênica é maior, tendo em vista que a semente transgênica é mais cara que a convencional e também porque sobre ela incide o pagamento da taxa tecnológica, que era de R$ 0,30 por quilograma em 2010 (Richetti, 2010).
Além de herbicidas causarem aumento na frequência de plantas resistentes aos mesmo, em razão do grande aumento de seu uso provicado pela expansãoo do cultivo de variedades transgênicas, certas plantas transgênicas tem transferido, por meio de cruzamentos, genes de resistência a herbicidas para outras plantas da mesma espécie ou de espécies afins (exemplos na Tabela 3.4). Um dos exemplos mais emblemáticos é a contaminação de brassicas nativas ou naturalizadas nos Estados Unidos. Genes de variedades transgênicas foram encontradas em 45% das amostras. Aproximadamente 80% das amostras contaminadas com um transgene, 41% era com o transgene cp4 epsps, 39% com o gene pat, que proporcionam resistência ao Roundup e ao Glufosinato de amônio., respectivamente (Schafer et al., 2011). Além disso, 0,7% apresentaram os dois transgenes. Assim, este impacto tem consequências de difícil mensuraçãoo, mas certamente impacta no aumento dos custos da agricultura.
Tabela 3.4. Exemplos selecionados de transferência de genes de resistência a herbicida de
plantas transgênicas para suas plantas daninhas. Cultura Planta espontânea (‘daninha’) Herbicida Autor
Canola Mostarda silvestre Basta Chèvre et al., 1998; Schafer et al., 2011
Trigo Aegilops cylindrica Roundup Steven et al.,1998
Sorgo „Johnson grass‟ Roundup Arriola e Ellstrand, 1998

83
Beterraba Beterraba não domesticada Roundup New Scientist, 21/10/2000
Agrostis stolonifera A. canina, A. capillaris, A. castellana, A. Gigantea e A. Pallens.
Roundup Wipff e Fricker, 2000; Snow, 21012
Arroz Arroz vernelho Glufosinato de amônio
Busconi et al., 2013
O artigo More "Funny" Honey, publicado no FOEE Biotech Mailout, aborda aquestão da perda de status do mel como alimento sadio e natural, como resultado da poluição causada pelos OGM. Análises efetuadas no mel indicaram a presença de pólen de canola transgênica tolerante a um herbicida. Este mel, coletado na Inglaterra em 1999 e analisado no Austrian Federal Laboratory em Vienna revelou a presença de DNA do gene de resistência ao mesmo herbicida. Os apicultores do Canadá também estão tendo problemas com a comercailização do mel, pois análises feitas na Europa detectaram contaminação com pólen de canola de variedades transgênicas. Agora, diante das novas normas da Europa, os apicultores se sentem sem o menor poder de reação e os preços do mel (contaminado) despencaram.
O comportamento das abelhas foi afetado quando exposto a alta concentração de proteína rCry1Ab, sendo que as abelhas levaram mais tempo para absorver o xarope contaminado com a toxina Bt. Além disso, as abelhas expostas a 5000 ppb de Cry1Ab tiveram sua aprendizagem perturbada. As abelhas continuaram a responder a um odor condicionado, mesmo na ausência de uma recompensa do alimento (Ramirez-Romero et al., 2008). Os resultados deste trabalho indicam que as plantações transgênicas expressando a proteína rCry1Ab podem impactar a eficiência das abelhas no forrageamento. Anteriormente, trabalhos efetuados em abelhas com inibidores de proteases demonstraram efeitos adversos quando abelhas foram alimentadas com açúcar contendo os referidos inibidores (Pham-Delégue M.-H., 1997).
No estudo conduzido por Rótolo et al. (2015) foram avaliados 12 sistemas de produção de grãos da Argentina, USA, Itália,Brasil e México, caracterizados por diferentes praticas agrícolas e intensidades em uso de recursos. Os resultados mostraram uma clara insustentabilidade de ambos padrões de cultivo a base de híbridos convencionais e a base de OGM. Sua fragilidade interna deriva de sua dependência de alta intensidade de insumos, o uso recursos não renováveis, de modo que o desempenho de seus indicadores a base de emergia não diferem substancialmente entre si. Os sistemas baseados em OGM não demonstraram melhor desempenho que o sistema padrão que usa sementes hibridas convencionais em termos de comercialização, ambiental e termodinâmico. Comparado à agricultura de subsistência, ambos os padrões de híbridos convencionais ou GM estão longe de serem sustentáveis, quando se utiliza baseado indicadores ambientais. Além disso, os autores constataram que quando todos os custos de produção e inflação são apropriadamente levados em conta, os sistemas que usam OGMs não confirmam maior rentabilidade econômica reivindicada, nem mesmo quando o comércio internacional é analisado. Segundo os autores. os resultados sugerem que as soluções para atividades agrícolas sustentáveis não virão da intensificação de ferramentas de alta tecnologia e uso de recursos, mas, ao invés, dependerão do melhor equilíbrio de no uso dos recursos locais renováveis e recursos importados não renováveis e de tecnologias apropriadas.
As alternativas às plantas transgênicas
As principais demandas dos mais de seis milhões de pequenos agricultores familiares no Brasil, os quais, historicamente, ainda produzem a maior parte dos alimentos que chega à mesa dos consumidores, não estão associadas à necessidade das plantas transgênicas, mas, sim, à necessidade de políticas publicas como a agrícola e a agrária que visem à

84
sustentabilidade e à rentabilidade de suas atividades. Assim, a necessidade e a urgência das plantas transgênicas para a agricultura brasileira é uma falsa questão. É importante mencionar que as plantas transgênicas desenvolvidas até o presente momento não atendem às necessidades da pequena propriedade familiar, ainda preponderante no país. As evidências científicas da utilização de plantas transgênicas com características de resistências a herbicidas (por exemplo, RR) ou portadoras de biocidas (por exemplo, Bt) na produção de commodities agrícolas nas grandes propriedades revelam o aumento na freqüência de plantas invasoras e insetos resistentes aos transgenes, implicando a vida curta dessas tecnologias. Isto gerará demandas de novas tecnologias (variedades transgênicas e/ou agrotóxicos), o que aumentará o grau de dependência dos agricultores. A avaliação de risco deve necessariamente conter informações sobre outras alternativas que poderiam ser utilizadas, bem como um comparativo entre os riscos das diversas soluções.
Assim, é preciso avaliar simultaneamente alternativas sustentáveis do ponto de vista agrícola e ambiental. Uma delas seria a agrodiversidade, termo empregado para definir a diversidade genética (intra-específica) e a diversidade de espécies (interespecífica) em cultivo nas propriedades agrícolas. Recentemente, pesquisadores chineses demonstraram que a heterogeneidade das culturas é uma alternativa possível à vulnerabilidade das monoculturas às doenças. Observou-se que variedades de arroz susceptíveis à doença bruzone, cultivadas em mistura com variedades resistentes a esta doença, apresentaram 89% de acréscimo na produtividade e uma redução de 94% de severidade dessa moléstia comparativamente à monocultura (Zhu et al., 2000). O sucesso dessa técnica, que é a simples mistura de diferentes variedades, foi tão significativo que, no segundo ano, não foi necessária a aplicação de fungicidas. Os resultados mostraram que a diversificação intra-específica das culturas proporciona um ambiente adequado para o controle de doenças que pode ser efetivo em grandes áreas, podendo contribuir para a sustentabilidade da produção agrícola.
Dentre os vários sistemas agrícolas sustentáveis ou alternativos, a agroecologia surge com muitas qualificações: não uso de insumos químicos, ambientalmente sustentável, uso de grande diversidade genética em cultivo em geral sócio-econômico associada, vizinhança, e com produtos alimentícios de alta qualidade biológica. No âmbito da agroecologia e não da agricultura industrial ou química, há inúmeras oportunidades para a C&T desenvolver pesquisas participativas, contextualizadas, que podem empoderar tanto a agricultura familiar, quanto as comunidades tradicionais no aperfeiçoamento dos processos e princípios agroecológicos utilizados. Assim, muitas das externalidades negativas atualmente inadmissíveis poderiam ser evitadas (Nodari e Guerra, 2015).
O país que detém a maior diversidade de espécies vegetais certamente deve ter um número de espécies comestíveis e agricultáveis capaz de proporcionar diferentes dietas balanceadas para as diferentes populações, respeitando-se sua cultura e suas necessidades. Vitamina A ou caroteno, por exemplo, são encontrados em dezenas de espécies comestíveis. O fato é que as plantas transgênicas estão sendo consideradas como a única maneira de aumentar a competitividade. Mas análises comparativas com outras matrizes de produção agrícola ainda não foram feitas.
12-PRINCIPIO DA PRECAUÇÃO
É importante ter em mente que a engenharia genética opera com base na manipulação do DNA de organismos vivos. Esta intervenção ocorre em âmbito muito mais complexo do que qualquer outra tecnologia já anteriormente aplicada. Esta tecnologia é aplicada em um nível de funcionamento da natureza a respeito do qual nossa base de conhecimento científico é ainda insuficiente (Griffiths, 1999).
Embora tenha havido avanços no conhecimento científico sobre os riscos associados ao cultivo de plantas transgênicas, o desenvolvimento da tecnologia de OGM ainda se baseia em processos do tipo “tentativa e erro”, portanto, imprecisos e pouco científicos. Assim, os

85
cientistas têm poucas condições de prever o comportamento do novo gene no organismo hospedeiro, sendo inadequado caracterizar-se a transgenia como science-based technology. Em suma, a engenharia genética encontra-se em seu estágio básico de pesquisa e ciência, sendo ainda prematura a liberação comercial de plantas transgênicas (Guerra e Nodari, 1999).
Desta forma, assume importância a adoção do Princípio da Precaução, estabelecido em acordos internacionais, como um princípio ético que afirma que a responsabilidade pelas futuras gerações e pelo meio ambiente deve ser combinada com as necessidades antropocêntricas do presente. Adotado no preâmbulo da CDB-, o Princípio da Precaução destaca “que quando exista ameaça de sensível redução ou perda de diversidade biológica, a falta de plena certeza científica não deve ser usada como razão para postergar medidas para evitar ou minimizar essa ameaça”. Assim, a adoção do Princípio da Precaução, se constitui em alternativa concreta a ser adotada diante de tantas incertezas científicas. Desta associação respeitosa e funcional do homem com a natureza, surgem as ações antecipatórias para proteger a saúde das pessoas e dos ecossistemas. Este princípio deve guiar as atividades humanas, mas incorpora outros atributos, como justiça, equidade, respeito, senso comum e prevenção (Raffensperger e Tikner, 1999). Também, este princípio admite que a adoção de cautela poderia evitar conseqüências danosas que, eventualmente, um OGM possa apresentar como resultado de sua liberação apressada ao meio ambiente.
As avaliações, ainda iniciais, dos impactos ambientais potenciais, podem permitir uma decisão balanceada entre os possíveis benefícios e a extensão e irreversibilidade dos danos e riscos. É importante que a toxicidade ambiental relativa seja incorporada na análise das mudanças de padrões de uso e quantidade de pesticidas, e que os impactos das culturas tolerantes a herbicidas na conservação do solo sejam quantificados. Por outro lado, devem ser tomadas medidas que possam prevenir a transferência de genes para populações selvagens, bem como reduzir a evolução da resistência aos transgenes.
Como concluem Wolfenbarger e Phifer (2000), tanto os riscos quanto os benefícios dos OGM podem variar temporal e espacialmente e devem ser analisados caso a caso. A elucidação destes riscos e benefícios dos OGM envolve a necessidade de estudos comparativos com outros sistemas e práticas agrícolas, tais como a agricultura orgânica. Nossa capacidade de predizer os impactos ecológicos de espécies introduzidas, incluindo OGM, é imprecisa e os dados empregados para avaliar impactos ecológicos potenciais apresentam limitações. Esta inabilidade de predizer acuradamente as conseqüências ecológicas, especialmente no longo prazo, aumentam a incerteza associada à avaliação de riscos, exigindo modificações nas estratégias de manejo destes riscos.
O intrigante neste momento de crise no uso das biotecnologias ditas modernas é que muitos dos riscos potenciais previamente anunciados estão de fato ocorrendo. Em 1989, Tiedje e colegas, e Pimentel e colegas mencionaram que os principais riscos potenciais dos OGM ao meio ambiente seriam: criação de novas pragas e plantas daninhas e um aumento das pragas já existentes por meio da recombinação gênica entre a planta transgênica e outras espécies filogeneticamente relacionadas; a produção de substâncias que são ou poderiam ser tóxicas a organismos não-alvos; o efeito disruptivo em comunidades bióticas e o desperdício de valiosos recursos genéticos, seguido de contaminação de espécies nativas com características originadas de parentes distantes ou de espécies não relacionadas e efeitos adversos em processos dos ecossistemas e origem de substâncias secundárias tóxicas após a degradação incompleta de químicos perigosos. Trabalhos publicados confirmaram os dois primeiros. Quanto aos dois últimos há a necessidade de estudos.
Princípio da precaução é inseparável da posição ética mais geral, segundo a qual é irresponsável participar do tipo de pesquisa que leva a inovações tecnocientíficas, a não ser que pesquisas rigorosas e sistemáticas, de dimensões comparáveis, sobre as consequências (riscos) ecológicas e sociais em longo prazo de sua implementação sejam efetuadas. É imprescindível levar em conta as condições socioeconômicas das implementações planejadas; a não ser que pesquisas adequadas, localizadas num espaço de alternativas bem

86
escolhido e pertinente para a avaliação do valor social geral (benefícios) das implementações, seja conduzida (LACEY, 2005 e 2009).
O Principio da Precaução está estabelecido no artigo 1 da nova lei de biossegurança. Portanto, é obrigação de todos os brasileiros observarem.
13. ROTULAGEM
A rotulagem dos alimentos está prevista no Código de Defesa do Consumidor (Lei n° 8.078, de 11/09/90 – art. 6°, III e art. 8°). Trata-se então de uma norma, que garante ao cidadão ser informado sobre um produto, o que lhe permite o direito de escolha. Além disso, a rotulagem permite a rastreabilidade, pois em casos de efeitos na saúde humana, os produtos rotulados seriam facilmente identificados e recolhidos.
No Brasil, a fiscalização sobre a rotulagem está a cargo da Vigilância Sanitária. Contudo, a decisão e mesmo o conteúdo e outras características do rótulo, está no âmbito do Ministério da Justiça.
O Decreto nº 4.680, de 24 de abril de 2003, regulamenta o direito à informação, assegurado pela Lei no 8.078, de 11 de setembro de 1990, quanto aos alimentos e ingredientes alimentares destinados ao consumo humano ou animal que contenham, ou seja, produzidos a partir de organismos geneticamente modificados, sem prejuízo do cumprimento das demais normas aplicáveis. Na comercialização de alimentos e ingredientes alimentares destinados ao consumo humano ou animal que contenham, ou seja, produzidos a partir de organismos geneticamente modificados, com presença acima do limite de um por cento do produto, o consumidor deverá ser informado da natureza transgênica desse produto. Tanto nos produtos embalados como nos vendidos a granel ou in natura, o rótulo da embalagem ou do recipiente em que estão contidos deverá constar, em destaque, no painel principal e em conjunto com o símbolo a ser definido mediante ato do Ministério da Justiça, uma das seguintes expressões, dependendo do caso: "(nome do produto) transgênico", "contém (nome do ingrediente ou ingredientes) transgênico(s)" ou "produto produzido a partir de (nome do produto) transgênico". O consumidor deverá ser informado sobre a espécie doadora do gene no local reservado para a identificação dos ingredientes. A informação também deverá constar do documento fiscal, de modo que essa formação acompanhe o produto ou ingrediente em todas as etapas da cadeia produtiva.
A rotulagem consitui-se em:
NECESSIDADE DE SABER - A rotulagem plena é um requisito fundamental e imprescindível para se estabelecer uma efetiva vigilância dos alimentos contendo OGMs e seus derivados.
DIREITO DE SABER – Direito previsto no Código de Defesa do Consumidor. O tipo de gene inserido, os aspectos religiosos e os valores culturais e pessoais devem ser considerados.
CONVENIÊNCIA DE SABER – É um requisito que considera o quantitativo de ADN e de proteína recombinante no produto final (Ex: acima de 1%).
Em termos de saúde pública, tudo tem que ser rotulado, não importa quanto tem dentro da embalagem.
A nível internacional existe um Grupo de Trabalho de Rotulagem que foi encarregado de preparar uma versão preliminar a ser discutida na reunião do Codex Alimentarius. Tomando-se em consideração o que houve na Conferência de Partes da CDB, pode ser que ainda no ano de 2000, a reunião do Codex também aprove as normas internacionais de rotulagem dos alimentos transgênicos ou que contenham ingredientes de OGMs.

87
14- NOVA TECNOLOGIA CRISPR
Adicionar, remover ou alterar sequências de DNA tem sido essencial para realizar estudos que buscam a compreensão genética de características fenotípicas. Depois da Tecnologia Recombinante, desenvolvida há pouco mais de 40 anos, surge com a promessa de eficiência e facilidade de uso sem precedentes, a tecnologia de edição de DNA” denominada de CRISPR (clustered regularly interspersed short palindromic repeats), porque é baseada em repetições palindrômicas curtas intercaladas regularmente agrupadas). Se associada a proteína Cas9 (que é uma nuclease) o sistema é considerado pelos seus proponentes como completamente revolucionário em termos de engenheirar os genomas. Este padrão de sequências CRISPR aparece em mais de 40% das bactérias e em 90 % dos micróbios pertencentes a Archaea (The CRISPR Craze. Science, v. 341, 23 August 2013, p.833-836).
Trata-se de processos naturais que incluem de corte da dupla fita do DNA por enzimas e seus posterior reparo com possibilidade de inserir, deletar ou mesmo altera sequências de DNA. O sistema baseia-se na CRISPR bacterianas e de Archaeas, que é parte do sistema imune adaptativo para expurgar ou mesmo degradar o DNA virai ou plasmidial invasor, que se baseia na atividade conjunta da endonuclease CRISPR e da proteína Cas, com a especificidade da sequência dirigida por RNAs CRISPR (crRNAs).
In vivo, quando CRISPR entra em ação em resposta a um fago invasor, as bactérias transcrevem os espaçadores e o DNA palindrómico formando uma molécula de RNA longa que a célula, em seguida, corta em RNAs curtos derivados de espaçadoras, denominados de crRNAs. Um segmento adicional de RNA, chamado tracrRNA, atua com Cas9 para produzir o crRNA. Juntos, Cas9, tracrRNA e crRNA de alguma forma atacam o DNA exógeno (invasor) que coincide (matches) com o crRNA.
A proteína Cas9 é uma nuclease, enzima especializada em cortar DNA, nos dois sítios (locais) ativos de corte, um sitio para cada fita da dupla hélice do DNA. Como é possível desativar a habilidade da Cas9 de cortar em um ou nos dois sítios de corte, sem interferir na capacidade complexa de reconhecer o seu DNA alvo, permitiu que o sistema CRISPR/Cas9 pudesse ser desenhado para reconhecer alvos e alterar o genoma de outros organismos.
Assim, in vitro, o sistema foi adapatado da seguinte maneira. O sistema CRISPR tipo II de Streptococcus pyogenes é composto por três genes, incluindo um que codifica para a nuclease Cas9 e dois genes de RNA não-codificantes: trans-activação crRNA (tracrRNA) e precursor crRNA (pre-crRNA). O pre-crRNA programável, que contém sequências de nuclease guia (espaçadores) interespaçadas por repetições diretas idênticos, é processado para se tornar crRNA em combinação com tracrRNA. Os dois genes de RNA podem ser substituídos por um gene de RNA recombinante de guia única (gRNA), contendo um gancho (hairpin) desenhado que imita o complexo crRNA- tracrRNA. A especificidade da ligação da Cas9 com o DNA alvo é determinada por ambos o emparelhamento de bases entre gRNA-DNA e um domínio proto-espaçador adjacente (PAM, sequência: NGG) imediatamente a jusante (downstream) da região alvo. Ambos os domínios da nuclease Cas9 (HNH e RuvC-like) cortarão uma fita de DNA da cadeia dupla no mesmo local (três nucleotídeos [distante do PAM), resultando em uma quebra (corte) na dupla fita. O sistema Cas 9/CRISPR tem sido aproveitado para realizar a edição eficaz do genoma de uma variedade de organismos, incluindo bactérias, leveduras, plantas e animais, bem como linhas de células humanas. Mais importante, o uso desta tecnologia de endonuclease guiada por RNA, múltiplas mutações de genes e a sua transmissão nas linhas germinativas foram alcançados (Xing et al., 2014).
Com a tecnologia CRISPR/Cas9, os cientistas envolvidos com o desenvolvimenyto da mesma, admitem que podem criar modelos de ratos que simulam doenças humanas muito mais rapidamente do que antes, como também estudar genes individuais muito mais rápido e, por fim, facilmente alterar múltiplos genes nas células de uma só vez para estudar suas interações.

88
Atualmente, várias empresas comercializam kit CRISPR/Cas9 para diferentes genes como alvo de alteração ou sequencias alvos para inserção de transgenes. Alternativamente, é possível fazer o próprio sistema em laboratório.
Como a tecnologia CRISPR / Cas9 funciona: a) Uma molécula de RNA é introduzido numa célula. Ele serve como um guia para localizar um segmento (sequência) específico de DNA que contém um gene o pesquisador quer editar. Quando a molécula de RNA encontra o gene correto, o RNA se liga na sequência de DNA do gene; b. Cas9, uma enzima bacteriana ligada ao RNA guia (gRNA), corta a sequência de DNA no local desejado do genoma; c. Uma vez que o genoma é quebrado (cortado), a célula irá tentar reparar o corte, que pode desativar ou nocautear um gene especifico. Ou inserir um novo segmento (sequência) de DNA no corte, essencialmente colando um gene no local desejado e alterar o genoma; d. Mesmo pequenas mudanças no genoma normalmente fará com que ele pare de funcionar; isto seria útil quando se tenta evitar a expressão de uma característica indesejada.

89
PARTE 4 – DIREITOS DE PROPRIEDADE INTELECTUAL
1-DIREITOS DE PROTEÇÃO E PATENTES
A proteção à propriedade intelectual incide sobre criações do intelecto humano, não abrangendo a descoberta de algo preexistente. Assim a patente é a expressão legal do privilégio temporário (exploração comercial) concedido pelo Estado à pessoa física ou jurídica, pela criação de algo novo. Para ser patenteável o invento deve ser descrito de tal maneira que possa ser reproduzido por qualquer pessoa que tem competência na arte. Além disso a inovação deve ter uso prático definido.
Para obter a patente, a invenção deve ser também novidade. Uma criação mecânica é nova quando ainda não foi divulgada publicamente. No caso de microrganismos, mesmo que identificados recentemente, existiu previamente em estado natural e então não seria novidade. A maioria dos pedidos de patentes em biotecnologia se constituem em descobertas em não em invenções, e então não seriam patenteáveis. Um invento não pode ser óbvio: deve expressar solução inovadora, em relação ao estado da arte - distinto de descoberta, referente a algo desconhecido, porém preexistente (Schneider, 1993). Neste caso, tanto as enzimas quanto os genes utilizados em plantas transgênicas preexistiam na natureza assim como os princípios ativos de organismos vivos usados na industrialização de produtos diversos.
O Congresso Nacional aprovou a lei nº 9.279 de 14 de maio de 1996 (DOU de 15/05/96), que regula direitos e obrigações relativos à propriedade industrial. Apesar dos quatro anos de tramitação, a discussão deste complexo projeto na comunidade científica e mesmo na sociedade ocorreu de forma tímida, infreqüente e superficial Sua aprovação ocorreu num ambiente de divergência de opiniões e pressões políticas e econômicas, as mais diversas. Entre as características da lei, merecem destaque:
- A sua complexidade: a lei possui 243 artigos e é complexa do ponto de vista técnico
- Ao detentor de patentes são conferidos amplos direitos e praticamente nenhum dever;
- Uma vez concedida a patente, se cria o monopólio. A lei, então restringe a soberania com relação a proteção de determinados setores da economia nacional;
- É patenteável a invenção que atenda aos requisitos de novidade, atividade inventiva e aplicação industrial;
-Não se considera invenção nem modelo de utilidade: o todo ou parte de seres vivos naturais e materiais biológicos, encontrados na natureza, ou ainda que dela isolados, inclusive o genoma ou germoplasma de qualquer ser vivo natural e os processos biológicos naturais;
- Não são patenteáveis: o todo ou parte de seres vivos, exceto microorganismos transgênicos que atendam aos três requisitos de patenteabilidade - novidade, atividade inventiva e aplicação industrial e que não sejam mera descoberta... Neste caso, microorganismos transgênicos são organismos, exceto o todo ou parte de plantas e animais, que expressem, mediante intervenção humana direta em sua composição genética, uma característica normalmente não alcançável pela espécie em condições naturais;
O patenteamento de genes poder vir a conflitar com o registro de uma nova cultivar, nos países onde é adotado o esquema de Direito de Proteção de Cultivares. Caso o gene seja isolado por um laboratório mas inserido em plantas por uma outra instituição, o direito de comercializar e cultivar a nova variedade provavelmente deverá reembolsar duas novas operações até agora não feitas no Brasil. Como conseqüência, o custo da semente deverá aumentar significativamente, dependendo do gene transferido. Nos países da Europa, a companhia de melhoramento pagará os 'royalties' pelo uso do gene nos seu programa e

90
cobrará 'royalties' sobre o uso e comércio de eventuais OGMs que desenvolver com tal gene. O tipo de acordo entre as partes ainda está sendo estudado. A regulamentação nos países europeus vai ainda prever a transferência de genes inseridos de PTs para outras cultivares.
Muitas patentes 'amplas' (ex: qualquer método de modificação de genes de Bacillus thuringiensis) tem sido concedidas nos Estados Unidos. Em decorrência disso, está havendo uma série de ações na Justiça de várias empresas contra a empresa detentora da patente.
As patentes na área da transgênia estão causando muitos embates juridicos e éticos. Nos EUA, Michael Hansen, especialista da Associação dos Consumidores, de forma consciente e crítica à possibilidade de patentear sementes transgênicas, indaga-se com a seguinte incongruência: ao mesmo tempo em que dizem que não há necessidade de testar as plantas transgênicas, pelo fato de serem similares às suas homólogas convencionais, solicitam patentes, sob a alegação de que os OGMs representam uma criação nova.
"A possibilidade de patenteamento de tecnologias transgênicas inseridas em variedades permite não só a cobrança de royalties, o que aumenta os custos de produção, mas também o privilégio de disponibilizar ou não um recurso genético aos agricultores, o que ameaça a soberania alimentar. O fato de o governo garantir patentes em tecnologias transgênicas inseridas em variedades, além de garantir a continuidade do apoio ao modelo agrícola industrial, com todas as suas consequências adversas, gera um paradoxo ético profundo. Ao permitir que uma variedade seja indiretamente patenteada, o país garante proteção a uma tecnologia transgênica, legaliza uma apropriação indébita, que é a associação do transgene com outros 30 a 40 mil genes, fruto não só do trabalho dos melhoristas passados, mas também de inúmeras gerações de agricultores que mantiveram e selecionaram tipos mais adaptados, a partir dos quais, os melhoristas desenvolveram novas variedades. Assim, num passe de mágica, este patrimônio dos povos foi apropriado por empresas transnacionais, sem nenhuma compensação. Ao contrário, os usuários ainda tem que pagar royalties" (Nodari e Guerra, 2009).
2-LEI DE PROTEÇÃO DAS CULTIVARES
Um dos primeiros países a adotar a proteção de cultivares foi os Estados Unidos em 1930, com o “Plant Patent Act”. Esta medida garantia ao melhorista o direito de propagar as mudas de variedades protegidas por um período de 17 anos. A justificativa utilizada para a implantação da medida foi incentivar o investimento em pesquisas com plantas de propagação vegetativa. Somente 40 anos mais tarde os Estados Unidos implantaram o sistema de proteção de cultivares com propagação sexuada, o “Plant Variety Protection Act”.
O desenvolvimento de novas cultivares e de outras tecnologias agrícolas provocou um grande impacto na agricultura mundial. Concomitantemente a isto ocorreu uma grande mobilização para estabelecer sistemas de proteção nos países industrializados. No ano de 1961, em Paris, ocorreu a primeira convenção internacional que resultou na criação da União Internacional para a Proteção de Obtenções Vegetais (UPOV). A UPOV é um organismo internacional, que estabelece os direitos de melhorista ou de propriedade intelectual sobre as variedades melhoradas. Posteriormente esta convenção foi revisada em 1972, 1978 e 1991. A adesão a uma das duas últimas convenções (1978 ou 1991) requer que o país tenha estabelecido uma legislação própria e compatível com as diretrizes estabelecidas. Além disso, a Organização Mundial de Propriedade Industrial (WIPO ou OMPI) determinou que os países membros que não tivessem estabelecido legislação sobre o assunto não poderiam aderir à Convenção de 78, estando automaticamente incluídos na Convenção de 1991.
O Brasil, que agora tem sua Lei de Proteção de Cultivares (Lei n 9456 de 25/04/97), solicitou adesão a Convenção de 1978, a qual tem a preferência da maioria dos países, uma vez que este é o sistema de proteção mais adequado para o desenvolvimento agrícola mundial. Atualmente, já assinaram esta convenção mais de 20 países, entre os quais Canadá, Estados Unidos, países da Europa, Argentina, Uruguai e Chile. Especialistas do mundo inteiro

91
tem sido unânimes em afirmar que a convenção de 1991 satisfaz preferencialmente as grandes empresas produtoras de sementes em detrimento do interesse social. Por isto mesmo, poucos países aderiram a esta última convenção.
Embora em alguns países exista o direito de patente sobre variedades, o acordo TRIPS permitiu aos estados membros o direito de excluir da patenteabilidade as cultivares de plantas e as raças de animais. O Brasil utilizou esta prerrogativa. A nova lei de propriedade
industrial (Lei n 9.279), também chamada de Lei de Patentes, aprovada em maio de 1996, prevê em seu art. 18 que as variedades vegetais não são patenteáveis. Com a lei 9456, as cultivares melhoradas passaram a ser protegidas pelos „direitos de melhorista‟. A diferença entre o sistema de patentes e o de direitos de melhorista, está basicamente restrita aos efeitos da proteção. Ou seja, a proteção não é tão severa com os pesquisadores, agricultores e consumidores, como é o caso das patentes. Nos países onde as patentes de cultivares são permitidas, a proteção abrange até a fase de industrialização do produto primário.
Além desta lei, existem outros instrumentos que afetam o uso de recursos genéticos vegetais como a Convenção da Biodiversidade Biológica (de 5/6/1992) e a Lei de Acessos,
que ora tramita no Senado Federal (PLS n 306, de 1995).
Principais aspectos da Lei de Proteção de Cultivares
Em consonância com a legislação disponível, o órgão a quem compete a proteção das cultivares é o Serviço Nacional de Proteção de Cultivares (SNPC), vinculado ao Ministério
da Agricultura e Abastecimento. A lei n 9456 não especifica claramente a estrutura nem as atribuições deste órgão, o que foi feito recentemente através do MAA.
Para o registro de uma determinada cultivar no SNPC, a mesma deve ter nome próprio e apresentar as caraterísticas de distinguibilidade, homogeneidade e estabilidade (simbolicamente abreviadas por DHE). Portanto, a variedade a ser protegida não poderá ser idêntica a uma já registrada no país ou em países com os quais o Brasil tem tratados. No caso de cultivares de autofecundação ou híbridos, a cultivar também deve apresentar a característica de homogeneidade, ou seja não poderá apresentar misturas. Finalmente, a cultivar tem que ser estável, ou seja manter suas características através das gerações.
A Lei de Proteção de Cultivares protege pelo período de 18 anos as videiras, plantas frutíferas, florestais e ornamentais e por 15 anos, as demais espécies. A ata vigente da UPOV é a de 1978, pela qual os Estados membros devem aplicar a Convenção para um mínimo de
24 espécies ou gêneros num prazo de 8 anos, após a entrada em vigor lei. Em seu artigo 4, a lei prevê a inclusão das mesmas gradativa. Assim, num primeiro momento a Lei abrangerá 5 espécies, às quais serão acrescidas de mais 5 após 3 anos da regulamentação da lei. Outras 14 espécies serão incorporadas até o oitavo ano após a regulamentação. Quando protegida, o detentor do registro, chamado de titular, detém os direitos de melhorista. Ou seja, o produtor de sementes (ou mudas) que quer utilizar a cultivar em lavoura comercial de produção de sementes (ou mudas) deverá ter licença do titular, a ser obtida mediante acordo. Por ocasião da compra de semente (ou muda) de cultivar protegida para o primeiro plantio de lavoura comercial, o agricultor estará pagando os royalties referente a proteção no preço final do produto.
A lei ainda prevê salvaguardas que permitem a interferência do Ministério da Agricultura na multiplicação e comercialização das cultivares protegidas. A primeira delas é a licença compulsória que permite a exploração de uma cultivar protegida sem a autorização de seu titular. Nos casos de emergência nacional ou abuso do poder econômico, uma cultivar protegida poderá ser tornar de uso público restrito. Entretanto, em ambos os casos, o titular terá assegurado a remuneração referente a exploração e o assunto terá especificidade em regulamento posterior.

92
Principais Implicações da Lei
Do ponto de vista do produtor, a lei também é flexível ao lhe permitir utilizar como semente para a safra seguinte, material colhido no ano anterior, com exceção da cana-de-açúcar. Para os pequenos produtores, a lei permite além do uso da própria semente, a troca de material protegido com outros pequenos agricultores sem ferir a legislação. Para tanto, o interessado deve atender o que está previsto nas normas do INCRA para seu enquadramento como pequeno produtor rural.
No âmbito do Mercosul a existência de um mercado livre, num curto prazo de tempo, implica na necessidade de compatibilização das legislações dos Estados membros, que hoje apresentam diferenças marcantes. Dos países membros do Mercosul, agora só o Paraguai não tem legislação própria. Atualmente variedades desenvolvidas no Brasil estão sendo cultivadas nos diversos países da América Latina e vice-versa, sem nenhum pagamento de royaties. Por certo, esta situação deverá ser outra após esta lei.
Do ponto de vista técnico, a questão mais polêmica é a possibilidade de proteção de cultivar essencialmente derivada. O problema é estabelecer as diferenças mínimas entre uma cultivar essencialmente derivada e a cultivar ancestral protegida. Estas diferenças mínimas são difíceis e onerosas de serem estabelecidas. A própria lei no seu artigo 3 (incisos III e IX), não determina com precisão qual é a margem mínima que separa ambas, ao remeter para órgão competente o estabelecimento dos critérios de diferenciação.
Embora a lei de patentes proíba o patenteamento de plantas e animais, ela permite o patenteamento de processos, inclusive os biotecnológicos. Neste caso haveria a possibilidade de uma planta transgênica ser duplamente protegida, pela lei de cultivares e pela lei de patentes. No Brasil, esta têm sido a forma preferida por empresas do setor para tentar obter o patenteamento de plantas transgênicas. Este aspecto vêm gerando controvérsias em vários países, inclusive no âmbito da Comunidade Européia uma vez que alguns países membros aceitam a dupla proteção Guerra e Nodari, 1997).
3- IMPLICAÇÕES DAS NORMAS DE PROPRIEDADE INTELECTUAL SOBRE TRANSGÊNICOS
A complexidade que envolve o tema da propriedade intelectual de organismos geneticamente modificados e seus derivados, disciutido no artigo de Pellanda e Nodari (2010) é aqui apresentado de forma resumida.
As patentes dos transgênicos - além de envolver uma tecnologia incerta quanto aos impactos causados pela produção e consumo em longo prazo – produz reflexos negativos sob vários aspectos, tais como: econômico, cultural, ambiental e no direito do consumidor.
A partir dos efeitos da patenteabilidade dos transgênicos desenvolvidos nessa pesquisa, é possível concluir que:
a) as patentes de sementes transgênicas trazem prejuízos econômicos, principalmente aos agricultores, os quais ficam reféns dessa tecnologia, por meio da obrigatoriedade do pagamento de royalties principalmente a grandes multinacionais do ramo da transgenia e a proibição de reutilização de sementes sem o pagamento dessa taxa de uso, onerando pequenas e grandes plantações;
b) as patentes servem como incentivo ao investimento de empresas e agricultores em grãos produzidos em larga escala, ou seja, contribui ao avanço da monocultura, resultando na perda de conhecimentos tradicionais e na erosão genética da diversidade genética anteriormente em cultivo;
c) as patentes de sementes garantem que seus detentores recebam royalties pelo uso de sua tecnologia, os quais podem ser cobrados inclusive de agricultores que a produzem de forma involuntária em razão da polinização cruzada ou mistura de sementes;

93
d) a disseminação do pólen ou das proteínas expressas pelas plantações transgênicas resulta em efeitos adversos a organismos não alvo, como a perda da biodiversidade, produzindo reflexos negativos ao meio ambiente;
e) perde-se em segurança alimentar, uma vez que a população poderá consumir com maior frequência grãos que, em sua maioria, são transgênicos, sendo tal fato desconhecido pela maioria dos consumidores.
Por todas essas razões, verifica-se que o sistema de patente e proteção de cultivares estabelece o monopólio sobre certas variedades, além da apropriação indevida de recursos naturais. Entretanto, esses recursos provindos da natureza - bem de uso comum de todos, porém objeto dessa apropriação, aliados ao conhecimento tradicional, desenvolvido de forma milenar e reconhecidos internacionalmente - devem ser protegidos em prol da preservação do meio ambiente e da manutenção das culturas agrícolas e tradições das populações. Assim, a legislação nacional deveria assegurar de forma eficaz a proteção ao conhecimento tradicional, a participação na repartição dos benefícios e a participação nas tomadas de decisões. Porém, se a vontade política existir para tal, haverá a necessidade de mudanças na legislação vigente sobre direitos de propriedade intelectual, tanto em termos de proteção industrial quanto de cultivares.
4- BIODIVERSIDADE, BIOTECNOLOGIAS E AGRICULTURA
A biodiversidade no seu conceito mais amplo compreende todas as formas de vida, ecossistemas e processos ecológicos, reconhecendo hierarquias nos níveis genético, taxonômico e do ecossistema. A magnitude da biodiversidade brasileira não é conhecida com precisão tal a sua complexidade. A estimativa é de que no território brasileiro existam mais de 2 milhões de espécies distintas de plantas, animais e microorganismos. O Brasil é o país com a maior diversidade genética vegetal do mundo, contando com mais de 55.000 espécies catalogadas de um total estimado entre 350.000 e 550.000. Cerca de 2/3 destas espécies se encontram nos trópicos, estimando-se que o Brasil detenha cerca de 75% de todas as espécies existentes nas grandes florestas. Apenas 8% das espécies vegetais tem sido estudadas em termos de compostos fitoterápicos bioativos e apenas 1.100 espécies de plantas foram exaustivamente estudadas em suas propriedades medicinais (Guerra e Nodari, 1996).
O potencial de utilização sustentável da biodiversidade é fruto da disponibilidade de matéria-prima, tecnologia e mercado. Por exemplo, um parente silvestre do trigo originário da Turquia proporcionou genes para a resistência a doenças, que transferidos para variedades comerciais de trigo resultam num ganho anual de US$ 50 milhões, somente nos EUA. Uma variedade de cevada da Etiópia forneceu um gene de resistência a vírus que transferido para variedades em cultivo na Califórnia, proporciona uma economia de US$ de 160 milhões. Outro exemplo elucidativo é o de Catharantus roseus, originário de Madagascar. As vendas pela Eli Lilly das drogas anti-leucêmicas vincristina e vinblastina, derivadas desta planta, atingem valores anuais de US$ 200 milhões.
Apesar da riqueza da nossa biodiversidade vegetal, a maior parte das atividades econômicas baseia-se em espécies exóticas: cana-de-açúcar originada de Nova Guiné, café da Etiópia, arroz das Filipinas, soja da China, cacau do México, citros da China, trigo da Asia Menor, eucaliptos da Austrália, pinheiros da América Central e gramíneas forrageiras da África, entre outras.
Afirma-se que desde o início da agricultura, em torno de 90% de todas as variedades vegetais desenvolveram-se pelas "forças da natureza"; 9,9% por meio dos esforços da humanidade até o início deste século e apenas 0,1% pela utilização de métodos modernos de melhoramento genético. Apesar de não ser possível precisar com segurança, as chamadas variedades híbridas modernas, geradas principalmente nos países com pesquisa mais avançada, respondem por uma grande parte da produção agrícola mundial e a

94
expansão de grandes áreas de monocultura com estas variedades poderia colocar em risco o total da diversidade genética. Afirma-se também que as sementes são um reflexo do código genético da sociedade que as desenvolvem, produzindo réplicas dos sistemas agrícolas destas sociedades e colocando novamente em cena a divisão entre um Hemisfério Norte rico em tecnologia mas pobre em recursos genéticos e um Hemisfério Sul pobre em tecnologia mas riquissimo em diversidade biológica. Estima-se que um gene potencialmente útil do Sul pode representar negócios de US$ 1 bilhão no Norte e que o germoplasma do Sul contribua com valores estimados em US$ 66 bilhões por ano na economia dos EUA, prevendo-se o advento da revolução do gene com genes patenteados pelas grandes corporações transnacionais, associando os recursos genéticos como estratégia central para controle do suprimento mundial de alimentos.
Revolução Verde, Biodiversidade e Biotecnologias
A emergência das biotecnologias na produção agrícola mundial vem ocorrendo em um contexto de esgotamento de um modelo de exploração agrícola baseado na chamada "revolução verde". Estas tecnologias fundamentadas no uso intensivo de energia e insumos não beneficiaram todas as culturas e todos os agricultores, especialmente os pequenos produtores. De uma maneira geral estas técnicas visavam uma adequação do ambiente à variedade melhorada. Os programas de melhoramento vegetal baseados na utilização racional da biodiversidade e orientados a uma agricultura sustentável consistem em um processo de ajuste de uma determinada variedade a um determinado ambiente.
A chamada revolução verde caracterizou-se por alguns equívocos merecedores de reflexão. O primeiro diz respeito ao fato de que os geneticistas foram solicitados a criar variedades altamente produtivas em condições de abundância de fertilizantes e água e apesar do êxito inicial, essas variedades demonstraram suscetibilidade a pragas e doenças, necessitando-se agregar mais um componente oneroso ao sistema de produção, os pesticidas. O segundo relaciona-se à excessiva sub-estimação dos desgastes ambientais causados por concentrações excessivas de fertilizantes e pesticidas que acabaram por contaminar mananciais de água implicando em riscos para a população. O terceiro diz respeito à ameaça a diversidade genética em consequência da disseminação em escala global de poucas variedades.
O fluxo relativamente livre de materiais e informações entre pesquisadores agrícolas em diferentes países do mundo é essencial para reduzir as disparidades na capacidade de pesquisa destes países. Uma das maiores diferenças entre o sistema de pesquisa durante a revolução verde e aquele que emerge das biotecnologias é que, enquanto o primeiro caracterizou-se pela predominância do domínio público nos investimentos e resultados da pesquisa e pelo fluxo relativamente livre de informações, o segundo vem se caracterizando pelo domínio privado de investimentos e pelas restrições no fluxo de informações.
Biotecnologias e Agricultura
Nos anos 90, os setores da agroindústria, florestal e pesqueiro respondem por 40%, 4% e 1% do PIB brasileiro, respectivamente. Produtos da biodiversidade respondem por 31% das exportações brasileiras, especialmente através do café, soja e laranja. A biomassa vegetal através do álcool da cana-de-açúcar, da lenha e do carvão derivados de florestas nativas e plantadas, responde por 17% da matriz energética nacional.
A obtenção de plantas transgênicas depende basicamente da possibilidade de identificar, isolar, clonar, transferir e integrar características importantes, sendo que, em última análise, o sucesso das técnicas de engenharia genética baseia-se na expressão adequada do gene inserido. O escasso conhecimento sobre estes genes é o principal entrave para a aplicação destas biotecnologias avançadas na agricultura brasileira e uma vez eliminados os entraves relacionados com a lei de patentes e de biossegurança, os produtos a serem ofertados no mercado serão as variedades transgênicas resistentes a herbicidas e as que contém genes de Bacillus thuringensis para resistência a insetos. Outro entrave é a grande dificuldade na resolução e manipulação de caracteres quantitativos, os

95
de maior importância do ponto de vista econômico. Estas abordagens são estratégicas para a ampliação de mercado de grandes empresas do setor no Hemisfério Norte. Não por acaso, no ano de 1994, nos EUA, foram realizados 1.500 testes de campo com plantas transgênicas, 28% dos quais sobre resistência a herbicidas e 23% sobres resistência a insetos.
Dado o avanço das biotecnologias na agricultura mundial cabe uma apreciação de sua pertinência no modelo agrícola brasileiro. Por biotecnologias pertinentes entende-se aquelas tecnologias que contribuem ao desenvolvimento sustentado por serem tecnicamente factíveis dentro do nível de desenvolvimento técnico-científico do país, por trazerem benefícios mensuráveis aos destinatários, por serem ambientalmente seguras e por serem socioeconomica e culturalmente aceitáveis. Desta maneira cabe questionar quais as biotecnologias pertinentes ao atual estágio de desenvolvimento da agricultura brasileira. Das chamadas biotecnologias avançadas ênfase poderia ser dada às modificações dos constituintes dos produtos agrícolas, visando o aumento de sua qualidade, como por exemplo a alteração da biossíntese de carboidrato e proteínas de reserva. Técnicas de engenharia genética podem ser aplicadas para a produção de tipos específicos de amido ou alterar outros carboidrato como celulose e pectina. Genes que regulam a produção de amilose e batatinha já foram clonados, sugerindo que a produção deste composto pode ser manipulada. A engenharia genética também poderá contribuir para minimizar os efeitos do estresse abiótico sobre as cultura agrícolas. Plantas submetidas a condições limitantes de seca, temperaturas e salinidade acumulam compostos de baixo peso molecular e a inserção de genes originados de bactérias permite o acúmulo compostos de alto peso molecular elicitando mecanismos de tolerância nestas plantas.
Já as chamadas biotecnologias intermediárias apresentam um potencial maior de aplicação a curto prazo na agricultura brasileira. Entre elas cabe citar o desenvolvimento de variedades com capacidade de fixação biológica do nitrogênio e de biofertilizantes como fungos micorrízicos. Técnicas associadas à produção de bioinseticidas já são rotineiramente empregadas na agricultura brasileira como é o caso da produção do fungo entomopatógeno Beauveria bassiana. Com isto pode-se diminuir os custos de produção bem como eliminar os impactos negativos dos pesticidas sobre o ambiente e à saúde humana. Entre as biotecnologias intermediárias observa-se um grande potencial para a utilização dos marcadores moleculares no mapeamento genético. Uma das principais aplicações destes mapas genéticos relaciona-se com a seleção assistida por marcadores (MAS). Esta metodologia se baseia na escolha do marcador molecular como critério de seleção na expectativa de selecionar-se de forma indireta os alelos de interesse a ele ligados.
É nas técnicas de cultura de tecidos vegetais ou de micropropagação que se observa o maior impacto das biotecnologias hoje no Brasil, principalmente no que tange à espécies ornamentais, frutíferas e florestais. A propagação clonal massal de variedades melhoradas e isentas de patógenos vem sendo empregada rotineiramente nos setores mais avançados destas áreas no Brasil. No estado do RS, o emprego de variedades de moranguinho originadas da cultura de meristemas a partir da metade da década de 80, foi o ponto de partida para a melhoria do sistema de produção desta cultura permitindo que a produtividade média passasse de 3,6 para 40 t/ha. Hoje em todo o Brasil empregam-se mudas provindas desta técnica relativamente simples e de baixo custo. Impacto similar vem ocorrendo com a cultura da batatinha, cuja produtividade média elevou-se de 10,7 t/ha em 1980 para 15,2 t/ha em 1995. Este aumento de produtividade foi atribuído principalmente ao plantio de batatas-semente certificadas, livres de vírus, produzidas pela Embrapa. Em Santa Catarina, nos laboratórios da EPAGRI foram desenvolvidos protocolos para a micropropagação de mudas de bananeira livres de nematóides e da broca da bananeira, reduzindo drasticamente a necessidade de aplicação de pesticidas de alto impacto ambiental, humano e com possíveis efeitos residuais no fruto. Paralelamente a isto, instalou-se um laboratório de produção do fungo entomopatógeno Beauveria bassiana, permitindo o controle biológico do moleque da bananeira.

96
Os exemplos anteriores mostram que, paradoxalmente, as biotecnologias intermediárias são as que vem tendo maior aplicação no atual estágio de desenvolvimento agrícola do país. Esta constatação também é valida para diversos países da América Latina e do Caribe. Na Costa Rica, Honduras, Colômbia e em Cuba, a maior parte das mudas de abacaxizeiros e bananeiras são produzidas por técnicas de micropropropagação. Em laboratórios da Costa Rica, Honduras e Cuba, técnicas biotecnológicas relativamente simples como a seleção de linhagens celulares resistentes permitiram a obtenção de variedades de bananeiras resistentes à moléstia fúngica sigatoka-negra (Mychosphaerella fijiensis) (Izquierdo, 1995). Nos bananais de Cuba estima-se um gastos de US$ 700,00/ha para o controle desta moléstia. Em países da Ásia um programa da FAO intitulado "Do laboratório ao campo: biotecnologia agrícolas para pequenos produtores" identificou e recomendou as biotecnologias que deveriam estar disponíveis e seu custos passíveis de serem absorvidos pelos pequenos produtores. Estas biotecnologias incluem a cultura de tecidos para a micropropagação de variedades sadias de raízes e tubérculos, frutíferas e ornamentais, inoculantes derivados de bactérias, fungos e algas, bioinseticidas, produção de fungos comestíveis. Este projeto vem revelando ótimos resultados nos países de sua abrangência: Bangladesh, India, Indonésia, Nepal, Filipinas, Sri Lanka, Tailândia e Vietname.

97
PARTE 5 - BIOÉTICA EM RELAÇÃO AOS TRANSGÊNICOS
1-INTRODUÇÃO
A expressão ética é resultante da fusão de duas palavras gregas: ethos - modo de ser ou caráter; mos ou mores - costume ou costumes. Refere-se à avaliação normativa das ações e do caráter de indivíduos e grupos sociais. Usada alternativamente com moralidade para se referir às obrigações e deveres que governam a ação individual. "A ética é a teoria ou ciência do comportamento moral dos homens em sociedade" (Vazquez, 1980). O estudo da ética é a revisão crítica sobre valores. Para tal há necessidade de liberdade e ausência de preconceitos.
A BIOÉTICA é um neologismo: bios e ethos - modo de ser (ética) da vida. Trata das questões éticas da medicina, da saúde pública e das ciências da vida. A bioética pergunta-se sobre a legitimidade dos projetos de efeitos biotecnológicos.
Em 1948 o Código de Nuremberg foi estabelecido e contém normas para a pesquisa com seres humanos. Estabelece ainda a responsabilidade individual do pesquisador. Posteriormente, em 1964 houve um aperfeiçoamento do mesmo com a Declaração de Helsinque e suas versões seguintes com as revisões de 1975 (Japão), 1983 e 1989 (Venezuela).
A primeira Conferência de Biossegurança foi realizada em Asilomar no ano de 1975. Foram estabelecidas recomendações para manuseio, contenção e armazenamento de produtos perigosos bem como protocolos laboratoriais e os procedimentos associados aos diversos tipos de riscos. Também foi declarada uma moratória voluntária com relação às pesquisas na espécie humana (Science, 188:991-994, 1975) porque as previsões dos impactos eram impossíveis de serem adequadamente conhecidas. O maior saldo foi o respeito do público pelo gesto de precaução dos cientistas.
Em 1992 foi realizada a Convenção sobre Diversidade Biológica (CDB) no Rio de Janeiro, a qual contemplou a necessidade de um protocolo de internacional de biossegurança visando proteger a saúde e o meio ambiente.
Uma segunda conferência, 25 anos depois, ou seja, no ano 2000, foi realizada em Asilomar. Nesta conferência, foram enfatizados o estreitamento do investimento privado e o avanço da ciência; a ampliação e o fortalecimento das leis de proteção, notadamente a de patentes; a pressa na comercialização dos produtos e serviços da biotecnologia; a omissão de resultados; a falta de precaução e o rompimento de valores éticos. Em decorrência, os cientistas começaram a perder a credibilidade da sociedade e uma reação aos produtos das biotecnologias, em especial os transgênicos.
O paradigma bioético atual refere-se ao padrão de reflexão e argumentação sobre os valores e suas justificativas a respeito da vigência de competência biotecnocientífica em reprograma a vida de qualquer ser vivo. O próprio conceito de doença está sendo alterado, pois ele poderá não mais se restringir a um conjunto de sinais e sintomas, mas estender-se a predisposições genéticas para a manifestação de futuras sintomatologias.
Como um interesse é um interesse, seja lá de quem for esse interesse (Singer, 1984), a discussão deve ocorrer num ambiente de liberdade e sem preconceitos. Desta forma estaria garantido o pluralismo do espaço público bem como o politeísmo e a tolerância, características do espaço privado.
Também é importante no debate identificar os conflitos de interesse, pois do debate participam tanto pesquisadores independentes quantos aqueles ligados direta ou indiretamente aos proponentes das tecnologias.

98
2. PERCEPÇÃO PÚBLICA
Embora a existência de discussão permanente na sociedade, a questão das plantas e animais transgênicos é quase que desconhecido da maioria da população brasileira. Também para a maioria das pessoas que têm um diploma de ensino superior, estes tópicos são indecifráveis. É preciso então desenvolver ações junto a população no sentido de desconstruir esta novidade. Para tal, a mídia bem como os cientistas têm um papel preponderante, se engajados num processo educativo, sem paixões ou crenças. Há a necessidade do envolvimento de pessoas que têm conhecimento sobre o assunto de participarem sem preconceitos ou interesses além daquele de desconstruir este assunto complexo.
Inúmeras ONGs estão envolvidas na discussão desta questão. Dias globais de ação contra a Biotecnologia foram organizados. Contudo, nem tudo o que é dito ou escrito tem base científica ou técnica. Um posicionamento pessoal com base em crenças pode levar o processo ao descrédito.
Vários países realizaram pesquisas de opinião publica. Existem diferenças bastante expressivas entre as populações de diferentes países com relação a aceitabilidade de produtos transgênicos. Enquanto na Áustria, Luxemburgo e mesmo Inglaterra a maioria da população rejeita, os japoneses manifestam-se favoráveis ao consumo destes produtos. Mas querem que o produto seja sadio e seguro. Tanto em 1998, como em 2005, a Suíça realizou plebiscito para decidir se o país deveria banir ou não os produtos transgênicos em seu território. Em ambos plebiscitos, os suíços decidiram moratória pelo não uso de OGMs.
No cenário internacional, o International Rice Research Institute (IRRI), que co-patrocina o arroz dourado (transgênico para produzir vitamina A), fez uma pesquisa de opinião pública agora em 2001 perguntando: você comeria arroz que foi geneticamente modificado? Dos 1815 entrevistados, 76,97% responderam que não. Mesmo assim, o institruto vem tentando testar e comercializar este OGM, ainda sem sucesso.
A questão da transgenia causa profunda perplexidade nas pessoas por vários motivos. Em primeiro lugar, a passagem da doença da vaca louca do alimento para as vacas e destas para as pessoas ocorreu de fato, embora cientistas e políticos afirmaram que isto não iria ocorrer. Um outro aspecto é que a técnica é muito poderosa e isto assusta as pessoas. O homem pode reprogramar o código genético e as pessoas não têm idéia das consequências disto. Um outro aspecto está relacionado com o tipo de produtos. Os primeiros transgenes diminuem a qualidade dos alimentos. Os consumidores querem algo melhor.
As pessoas reagem de maneira diferente. A maioria boicota as compras. Outras praticam atos de sabotagem nas áreas cultivadas com variedades transgênicas. Tanto na Irlanda quanto na Inglaterra, dezenas de propriedades tiveram suas lavouras destruídas ou altamente danificadas por grupos contrários a biotecnologia.
No Brasil, os debates públicos sobre a transgenia e suas conseqüências desde 1998 vêm possibilitando o conhecimento da questão pela sociedade. Mas o fato é que, a maioria da população ainda não está suficientemente informada, nem mesmo tem conhecimento suficiente para entender e opinar a respeito de plantas transgênicas. Daí a responsabilidade inadiável do poder público, das universidades e dos técnicos de prestar este tipo de serviço à população brasileira.
Os consumidores brasileiros, na sua grande maioria, também mencionaram, 16 anos atrás, que não queriam consumir alimentos transgênicos conforme pesquisas efetuadas no mês de julho de 2000 pelos jornais O Globo (72%), Correio Brasiliense (70%) e Gazeta Mercantil (60%). Embora outras oesquisas foram feitos pelo IBOPE, não há informaçãoo sobre o que pensam os brasileiros nesta década sobre os OGMs.
3.OS INTERESSES ECONÔMICOS DA TRANSGÊNIA

99
Na maior parte dos casos de liberação de plantas transgênicas predominou o interesse comercial destas grandes empresas. Isto pode ser comprovado pelas investidas frequentes do governo americano junto aos países europeus e Japão. Para citar apenas um exemplo, os EUA atacaram a Comissão Européia que havia decidido pela rotulagem dos produtos transgênicos, em junho de 1997, argumentando que isto contrariava o livre comércio. Na época Dan Glickman, Secretário da Agricultura, disse que os Estados Unidos “não tolerariam” a segregação de produtos geneticamente modificados dos tradicionais. A resposta americana pode ser exemplificada pela atitude da companhia Monsanto que misturou os grãos transgênicos com não transgênicos, obrigando os europeus a comprarem apenas o bulk com a mistura.
Um episódio que ocorreu no ano de 2000, cujas conseqüências ainda não findaram, ilustra vários tipos de conflitos. O maior fiasco da biotecnologia como já é considerado, trata-se do StarLink. StarLink, um tipo de Bt que contém o gene Cry9C, foi aprovada nos EEUU para alimentação animal mas não para consumo humano, pois contém uma proteína que pode causar reações alérgicas em humanos, uma vez que a proteína Cry9C não é quebrada imediatamente nos testes de digestão. A empresa produtora desta variedade (Aventis), distribuiu as sementes sem nenhuma ressalva. Assim, houve contaminação de lavouras vizinhas com pólen desta variedade. Também, os milhos foram colhidos e misturados com os demais. Resíduos desta proteína foram detectados em produtos alimentícios e bebidas, tanto nos Estados Unidos quanto em outros países. Conseqüências: alergia detectada em 7 de 54 pessoas suspeitas, necessidade de recolher não só o milho colhido mas também os produtos já processados que poderiam contem a farinha contaminada com este milho, indenização dos supermercados, indenização dos compradores no país e no exterior, indenização de agricultores que tiveram sua lavoura contaminada pelo pólen do StarLink. Estima-se um gasto entre 100 milhões e 1 bilhão de dólares, o custo da operação.
Dentre as várias lições, duas são relevantes: 1) não foi possível localizar 12% da produção desta variedade, o que demonstra que uma vez liberado no ambiente, dificilmente existirá controle sobre um OGM; 2) as empresas não estão preocupadas com a saúde das pessoas nem com os agricultores, mas em vender seus produtos. Um outro tipo de conflito comercial poderá ocorrer entre agricultores, basicamente devido a contaminações pelos transgênicos. Ocorrendo cruzamentos entre plantas transgênicas e não transgênicas espécie, poderá criar conflitos entre produtores que utilizam transgênicos e produtores de alimentos chamados orgânicos, que são considerados de alta qualidade biológica. Como será resolvido este impasse? Um caso nos Estados Unidos implicou no prejuízo de US$170.000 a um produtor cuja produção orgânica foi contaminada por milho transgênico Bt. Na Inglaterra e outros países existem muitas ações tramitando na justiça, sobre esta questão, que ainda não tem solução fácil. O que acontecerá no Brasil? Pergunta ainda sem resposta.
Pólen de plantas transgênicas estão sendo coletados pelas abelhas e espalhados no mel. Já em 1999, Friends of the Earth, uma ONG, descobriu pólen de canola tolerante a herbicida (Arventis, ex AgrEvo) em abelhas cujas colméias estavam localizadas a 4 km de distância do experimento de OGM mais próximo. Implicação da poluição genética: o mel está perdendo o status de alimento sadio e natural. Os apicultores estão sendo forçados a se retirar das áreas próximas dos testes com OGMs: danos aos produtores de frutas e hortaliças. Carregamentos de mel orgânico enviados do Brasil a Europa retornam porque estavam contaminados por polen de plantas transgênicas. Os danos são tanto para os apicultores como para os produtores de frutas, cujas consequências poderão ser muito sérias. A questão da responsabilidade sobre a poluição ainda não está resolvida na Inglaterra.
“Uma dessas suposições é que, se alguém investe em uma inovação tecnocientífica, está autorizado a recuperar os custos do investimento e gerar um lucro. Assim, no processo de implementação da tecnologia, os riscos (para a saúde, para o ambiente etc.) tornam-se secundários, não por se ignorar os riscos conhecidos, mas por não se aceitar o ônus de chegar a antecipar teoricamente os possíveis riscos e comprová-los e certamente para não respaldar o ônus dos custos de avaliá-los [...]” (Lacey, 2006).

100
Um dos impactos menos discutidos no âmbito da transgenia em plantas refere-se à dependência tecnológica dos agricultores ao grande complexo industrial-genético, expressão utilizada por Berlan e Lewontin (1999) para designar as grandes empresas transnacionais do setor biotecnológico, que nos últimos 20 anos passaram a atuar de forma agressiva na apropriação dos recursos genéticos.
Em seu artigo publicado no Le Monde Diplomatique (janeiro de 1999), os referidos autores apresentam e discutem quatro argumentos sobre a apropriação dos recursos genéticos vegetais por parte deste complexo genético-industrial, cuja síntese pode ser assim descrita:
1) A riqueza das variedades agrícolas foi criada por agricultores de todo o mundo, em especial aqueles do terceiro mundo. A domesticação e a seleção feita por agricultores por milhares de anos gerou uma herança biológica que beneficiou as nações industrializadas. A agricultura norte-americana, por exemplo, foi construída em cima desses recursos, livremente importados do resto do mundo. Não é justo que poucas companhias agora se apropriem dessa herança biológica universal.
2) O aumento (sem precedentes) nas colheitas do mundo industrializado, assim como do terceiro mundo, pode ser atribuído ao livre movimento de conhecimento, aos recursos genéticos e à pesquisa pública. As colheitas aumentaram cinco vezes em duas gerações, depois de serem necessárias 15 gerações anteriores para esta colheita dobrar. Na década de 70, quase todos os híbridos norte-americanos de milho resultaram do cruzamento de duas linhagens, originadas de programas de melhoramento de universidade públicas.
3) A experiência mostra que o custo de privatizar o „progresso genético‟ é e será exorbitante. Estudos feitos na França pelo Institut National de la Recherche Agronomique (INRA), mostram que o custo adicional das sementes de trigo híbrido equivale a US$ 500 milhões (orçamento do INRA) para um ganho genético que poderia ser mais facilmente obtido usando-se variedades crioulas produzidas pelos agricultores.
4) Desistir dos direitos sobre essa herança significa liberar o complexo genético-industrial para direcionar o progresso tecnológico unicamente para os lucros. Da forma como a questão vem sendo conduzida pelas grandes empresas, não há uma demanda social para OGMs. O termo é somente uma cortina de fumaça para as demandas desse complexo genético-industrial.
Como é possível perceber, são muitas as implicações dessa tecnologia e estas precisam ser profundamente avaliadas, explicitadas e discutidas, pois é do interesse de toda a sociedade a percepção clara dos seus possíveis riscos e benefícios.
4. A RELAÇÃO DA COMUNIDADE CIENTÍFICA COM O GOVERNO
Para ilustrar a fealçao entre parte da comunidade coientifica e o governo o episódio da vaca louca é muito ilustrativo. A investigação que ocorreu na Inglaterra visando elucidar o veredicto final da comissão especialmente formada para aconselhar uma decisão do governo a respeito da „vaca louca‟, trouxe a tona, uma discussão a respeito da relação entre cientistas e governo.
Em sua edição de 5 de agosto deste ano, a Revista Nature, além de considerar o assunto em seu editorial, informa na página 490, que os membros do “Spongiform Encephalopathy Advisory Committee (SEAC) foram pressionados por representantes governamentais no sentido de endossar um parecer sobre a segurança da carne. Membros da referida comissão declararam que foram procurados por membros de órgãos governamentais que solicitaram-lhes a aprovação de um texto que a carne bovina era segura. Segundo o Presidente desta comissão, os membros se sentiram inconfortáveis e apreensivos em ter que aprovar uma nota tão curta.
É intrigante o fato de que as versões do parecer circulou por diversas autoridades inglesas para comentários. A temeridade da reação pública expressada por autoridades

101
governamentais fez com que a comissão retirasse frases do parecer final tipo “nenhum cientista diria que não haveria risco em comer carne bovina”.
A abdicação de se basear em dados puramente científicos por parte de membros da comunidade científica quando convocada para aconselhar o governo, como está sendo constatado neste episódio, se constitui num perigo para a população.
O balanço feito em maio de 2001 indicou que mais de 100 pessoas já morreram na Inglaterra e França, e que a doença já atingiu vários países europeus, tanto no gado quanto na espécie humana. O fato de que carne e gado europeu foram importados por outros países nos últimos anos, se constitui numa ameaça, pois os agentes infecciosos desta doença, os prions, podem ter sido disseminados.
Será que com a aprovação dos OGMs ocorre o mesmo que ocorreu com relatório sobre a „vaca louca‟ ?
Esta relação entre cientistas membros de comissões governamentais e governo deve ser melhor definida. O recado vem da própria população, que já não acredita mais nas decisões governamentais sobre questões que envolvem riscos à saúde e ao ambiente. Este fato não é exclusividade da Inglaterra. A polêmica em torno das implicações dos alimentos transgênicos é um exemplo notório em vários países.
No Brasil, a aprovação para liberação comercial da soja transgênica pela Comissão Técnica Nacional de Biossegurança (CTNBio) também foi tomada de forma apressada? Algum paralelo com a Inglaterra? A decisão se deu sem os dados dos estudos de impacto ambiental nos diversos ecossistemas brasileiros e do efeito do herbicida a base de glifosate, que será aplicado na referida soja, na saúde humana e meio ambiente.
É legitimo (para a transgênia) inferir com base na ausência de evidência contra uma teoria ou hipótese que é aceitável de acordo com a imparcialidade? (Lacey, 2005). Segundo Traavik et al. (1999), este é um paradoxo científico, pois a ausência de evidência jamais deve ser tomada como evidência da ausência.
Os diversos interesses econômicos, o grande número de cientistas envolvidos e a grande quantidade de estudos apontando, ao mesmo tempo, riscos e benefícios alimentam tal polêmica. Como assinala Lacey, a falta de clareza que envolve essas questões [relacionadas aos OGMs] revela que estão em jogo valores, interesses e modos de vida fundamentalmente opostos (Lacey, 2006). As consequências das decisões, ou indecisões, políticas sobre biossegurança dos transgênicos afetam diretamente a sociedade. Mais do que objeto biológico, os transgênicos também são objetos socioeconômicos, uma vez que configuram-se em mercadoria com necessidade de propriedade intelectual (Lacey, 2007).
Neste momento, cabe uma reflexão acompanhada de um conjunto de ações, a respeito do comportamento e das relações entre cientistas e governo. Para evitar tais tipos de episódios como o da vaca louca, há a necessidade de uma definição clara do papel destas comissões, a forma de escolha, bem como transparência nos trabalhos das mesmas.
É possível rejeitar o princípio da precaução quando a população corre risco? Devem os interesses maiores da população não podem ser sobrepostos por interesses econômicos imediatos?
5. A NECESSIDADE DE UM DEBATE PÚBLICO COM A SOCIEDADE
A ampla gama de implicações que este tema dos OGM engendra, ultrapassa hoje os limites da ciência. As questões éticas, sociais, econômicas e políticas não podem estar dissociadas do tema e do eixo das discussões. Parte da sociedade comunga a percepção de que este assunto está sendo conduzido de forma inadequada, como demonstram protestos de grupos de pressão e ONG. Esta percepção encontra respaldo nos episódios recentes da

102
doença da vaca louca, entre outros. Portanto, o diálogo deve ser social e extrapolar as paredes dos laboratórios científicos e gabinetes governamentais.
Por fim, também é preciso avaliar os impactos sobre o domínio no acesso e uso dos recursos genéticos. Afirma-se, com freqüência, que o insumo mais importante para o novo milênio é o conhecimento. As tecnologias decorrentes deste conhecimento poderão acentuar assimetrias nas relações econômicas e sociais entre as nações mais desenvolvidas e menos desenvolvidas, caso não forem estabelecidos mecanismos compensatórios e regulatórios. Não se pode admitir que interesses econômicos de uma minoria se sobreponham aos interesses maiores da sociedade.
Contudo, os recursos genéticos não terão papel menos importante que o conhecimento. Biotecnologias sem diversidade são mero exercício acadêmico, como afirma um documento da FAO (1999). Desta forma, é imperiosa a manutenção da diversidade bem como é fundamental tomar as medidas para evitar as ameaças à sua erosão genética.
As sociedades secularizadas e complexas estão dispostas a renunciar aos benefícios da biotecnociência? O fato é que existem muitas biotecnologias e há a necessidade de avaliar individualmente a aplicação de cada uma delas nos mais diversos aspectos.
É importante ter em mente que a engenharia genética opera com base na manipulação do DNA de organismos vivos. Esta intervenção ocorre em um nível muito mais complexo do que qualquer outra tecnologia já anteriormente aplicada. Esta tecnologia é aplicada em um nível de funcionamento da natureza a respeito do qual nossa base de conhecimento científico é ainda insuficiente, porque deliberadamente os proponentes da tecnologia não procuram avaliar todos os impactos, porque teriam dificuldades de lançar seus produtos.
Depois de quase 30 anos de desenvolvimento a tecnologia de OGM ainda se baseia em processos do tipo “tentativa e erro”, portanto imprecisos e pouco científicos. Assim, os cientistas têm poucas condições de prever o comportamento do novo gene no organismo hospedeiro, sendo inadequado chamar-se esta tecnologia de „science-based‟. Em suma, a engenharia genética encontra-se em seu estágio básico de pesquisa e ciência, sendo prematura a liberação comercial de plantas transgênicas.
6-IMPLICAÇÕES DA CLONAGEM DE ANIMAIS E DE HUMANOS
A Dolly popularizou a questão e gerou problemas e dúvidas. A Polly, que é uma ovelha com genes humanos, não recebeu atenção da mídia. A discussão sobre os xenotransplantes (transplante de órgãos de animais para seres humanos) ainda é muito polêmica.
A clonagem em animais é mais recente que em plantas. Nos anos 60 foi obtida a clonagem em sapos e 10 anos depois, em ratos. Em 1994 nasce Astrid, a porca transgênica com genes humanos, que produzem uma proteína de membrana, capaz de diminuir ou mesmo eliminar os riscos de rejeição de transplantes de órgãos do porco para seres humanos. ("Porco-irmão"). Em 1997 a clonagem alcançou outros animais (ovelhas, vacas e macacas).
A Clonagem humana vai acontecer num curto espaço de tempo? Embora não seja possível responder esta questão de modo conclusivo, existem fatos relacionados ao assunto que merecem reflexão:
• gêmeos são clones;
• a clonagem animal pressionará a clonagem humana;
• demandas individuais (ex: em São Paulo pai que perde filho em acidente quer um clone; mãe doa óvulo para filha gerar neto; mulheres podem gerar filho sem fecundação);
• a terapia gênica com células somáticas é quase uma realidade;

103
• fertilização in vitro (ou bebês de provetas) - Em 1780 na Inglaterra foi feita a primeira tentativa de inseminação artificial com o esperma do marido. Mais tarde, em 1884 foi feita a inseminação com esperma de um doador. Em 1978, o primeiro bebê de proveta. No Brasil, nasce em 1984 o primeiro bebê (uma menina que hoje tem 17 anos. Os bebês de provetas, uma realidade nos anos 1980 da realização de uma idéia surgida duzentos anos antes.
• doação de órgãos - crianças são geradas para doação de medula a irmãos;
• clonagem de embriões humanos não utilizados para reprodução (por serem defeituosos) até o estádio de 32 células (Science, 262:652-653, 1993);
• os xenotransplantes (transplantes de órgãos de animais para humanos);
• recomposição de órgãos humanos via cultura de tecidos (clonagem);
• bebês com material genético de duas mulheres (impropriamente denominados de geneticamente modificados) – criança gerada com a fertilização por um espermatozóide de um óvulo contendo genoma nuclear da mãe e mitocôndrias de uma doadora.
Entretanto tanto a engenharia genética em célula germinal humana, zigoto humano e embrião humano como a clonagem humana estão proibidos no Brasil, pelo Art. 6o da Lei da Biossegurança (Lei n° 11.105).
Em1996, a Resolução 196/96 de 16/10/96 Cria Conselho Nacional de Ética de Pesquisa e Comitês de Ética de Pesquisa Institucional (CEPI) com pelo menos 6 membros. Os CEPI passaram (i) a ser co-responsável pelas decisões e a ter as funções de (ii) consultoria e
(iii) educação. Cada comitê deve ser registrado no Ministério da Saúde. Na UFSC foi criado o Comitê de Ética em Pesquisa com Seres Humanos (CEPSH-UFSC), que é um órgão colegiado interdisciplinar, deliberativo, consultivo e educativo, vinculado à Universidade Federal de Santa Catarina, mas independente na tomada de decisões, criado para defender os interesses dos sujeitos da pesquisa em sua integridade e dignidade e para contribuir no desenvolvimento da pesquisa dentro de padrões éticos. Mais tarde, em 1999, a UFSC institui a Comissão Interna de Biossegurança (CIBio) composta de 5 membros.
Porque a discussão continua?
1) É que anteriormente não havia massa crítica para a discussão. No início do século havia em torno de 8 mil cientistas e químicos na Europa. Nos anos 80 este número cresceu para mais de 5 milhões, com a inclusão dos engenheiros. Portanto, a ciência e a tecnologia são consideradas dois dos principais componentes da cultura contemporânea.
2) O potencial das tecnologias pode reprogramar o código genético, e conseqüentemente a vida dos organismos.
3) A existência de vários conflitos decorrentes de diferentes interesses.
4) A geração da Dolly popularizou a questão, mas provocou problemas e dúvidas.
5) A percepção pública, após o episódio da „vaca louca‟;
6) Os possíveis riscos associados aos alimentos transgênicos.
7- TERAPIA GENÉTICA COM VETORES RECOMBINANTES NA ESPÉCIE HUMANA
Terapia genética ou gênica - diz respeito à possibilidade de corrigir defeitos ou prejuízos para a qualidade de vida saudável de indivíduos e populações. Na visão dos defensores da tecnologia, a terapia genética deve ser considerada como qualquer outra terapia, e não usá-la significaria infringir os próprios princípios da beneficência e de não-

104
maleficência que imperam desde Hipócrates. Não haveria, portanto, nenhuma objeção moralmente relevante contra o uso da terapia genética, desde que seja também respeitados o princípio da autonomia do consumidor e o princípio da justiça (ou de eqüidade).
Contudo, há objeções de cunho religioso e de cunho naturalista, que não consideram adequadamente o ponto de vista segundo o qual a natureza humana é algo dinâmico, suscetível de ser remoldado pela própria competência biotecnocientífica em rápida expansão. Há, contudo, uma objeção mais pertinente que refere-se a terapia genética aplicada a linha germinal. Neste caso, a alegação é que as conseqüências a médio e longo prazos são amplamente desconhecidas. Há também outras objeções. Ainda não se conhecem os efeitos colaterais da terapia gênica na espécie humama.
Cientistas como Richard Lewontin, diz em seu livro a Triple Helix (2000), que a expressão fenotípica é resultante da ontogenia de um indivíduo. Ou seja, depende da composição genética, dos eventos ao acaso durante a ontogenia e do ambiente. No atual estágio de conhecimento, pouco ou nada adiantará a disponibilidade de uma seqüência de 3 bilhões de pares de bases.
O primeiro teste de terapia genético foi feito em 1999, por cientistas da Penn University em Jesse Gelsinger, um jovem de 18 anos, com deficiência em uma enzima. O gene para controlar o metabolismo da amônia (gene para ornithine-transcarbamylase - OTC) foi introduzido no Vetor adenovirus. Este virus GM foi aplicaddo em Jesse. O gene invadiu não só o fígado (alvo), mas muitos outros órgãos, o que causou uma resposta sistêmica inflamatória. A temperatura do paciente alcançou 104,5 F. A dose do vetor foi de 38 trilhões de partículas virais - somente 1% alcançaram as células do fígado que era o alvo. Os cientistas da Penn University não compreenderam ainda o que aconteceu com Jesse Gelsinger, que veio a falacer em 24h após o inicio do tratamento (Science, 1999).
Três anos mais tarde, outra tentativa. Um paciente francês de terapia gênica
desenvolveu uma forma de câncer. Este fato iniciou um debete global de como avaliar os riscos que estão expostos os pacientes de terapia genética. (Nature, 420, p.116-118, 14/11/2002).
Em outra tentativa, a pessoa sofria de artritis reumática. Recebeu duas doses do vetor - Vírus AAV (adeno-associated virus) que continha o gene que codifica para uma proteína que inibe o fator de tumor necrótico (TNF-a), uma citoquina pró-inflamatória. Esta paciente também faleceu. A investigação sobre a morte de uma mulher de 36 anos num experimento de terapia gênica revelou uma complexa tragédia, mas não conseguiu ser claramente conclusivo se o experimento deveria ser condenado ou responsabilizado. Ela morreu 22 dias após tomar a segunda dose do “gene curador” (Science, v.317, 21/09/2007, p.1665).
Outras tentativas ainda foram feitas, até o momento sem grande sucesso.
8. O QUE SE ESPERA DOS PROFISSIONAIS DA BIOLOGIA E DA AGRONOMIA? 1) que se informe com base no acesso ao conhecimento cientifico de pesquisas independentes; 2) uma atitude crítica e imparcial face aos riscos e às potencialidades; 3) uma atitude eticamente responsável, engajada em acompanhar individual e publicamente os atos da biotecnociência e em praticar tanto uma "sabedoria prudencial" quanto uma prevenção eficaz; 4) obediência as normas legais e precaucionárias.
Como as nações e os grupos internacionais movem-se na direção do desenvolvimento ou evolução das normas de biossegurança, é essencial que existam métodos científicos para avaliar os riscos associados com as introduções na agricultura (Barton et al., 1997). O estado de valores do pesquisador é tão importante para a qualidade da ciência que produz quanto sua titulação, competência metodológica e capacitação técnica (Azevedo, 1995).

105
A bioética deve identificar racionalmente e responsavelmente as implicações sociais e culturais das descobertas nas ciências da vida concernentes a saúde, agricultura, alimentos, ambiente e estratégias de desenvolvimento. As aplicações da biotecnologia não podem ser restritas a um território. Então a bioética, inevitavelmente, tem uma dimensão internacional, o que não quer dizer que a dimensão nacional deve ser relegada (Kutukdjian, 1997).
É dever dos cientistas atuar como debatedores, decodificadores e facilitadores deste debate abrangente e polêmico, atual e de extrema importância para o país. Análises com bases em dados científicos evitam a promiscuidade dos debates e permitem a distinção entre ciência e crença.
9. CONLUSÕES
Não deve ses admitido que questões da mais alta relevância como a vida, a saúde e a morte do homem e de componentes ambientais, sejam decididas por pequenos grupos de cientistas, na presença de incertezas e interesses econômicos de uma minoria em detrimento dos interesses maiores da sociedade..

106
BIBLIOGRAFIA Ackerly, D.D.; Dudley, S.A.; Sultan, S.E.; Schmitt, J.; Colleman, J.S.; Linder, C. R.; Sandquist, D.R.; Geber, M.A.;
Evans, A.S.; Dawson, T.E.; Lechowicz, M.J. The evolution of plant ecophysiological traits: recent advances and future directions. BioScience, v.50, n.11, p.979-995, 2000.
Agapito-Tenfen, S.A.; Guerra, M.P.; Wikmar, O-G.; Nodari, R.O. Comparative proteomic analysis of genetically modified maize grown under different agroecosystems conditions in Brazil. Proteome Science, 11:46, 2013.
Agapito-Tenfen, S. ; Vilperte, V.; Benevenuto, R.; Rover, C.; Traavik, T.; Nodari, R. Effect of stacking insecticidal cry and herbicide tolerance epsps transgenes on transgenic maize proteome. BMC Plant Biology (Online), v. 14,
p. 346, 2014. Allard, R.W. Princípios do melhoramento genético das plantas. São Paulo: Blucher-USAID, 1960. 381p. Andreassen, M.; Rocca, E.; Bøhn, T.; Wikmark,O-G,; van den Berg, J.; Løvik, M.; Traavik, T.; Nygaard, U.C.
Humoral and cellular immune responses in mice after airway administration of Bacillus thuringiensis Cry1Ab
and MON810 cry1Ab-transgenic maize, Food and Agricultural Immunology, 26:4, 521-537, 2015. DOI: 10.1080/09540105.2014.988128
Antoniou, M.; Habib, M.E.M.; Howard, C.V.; Jennings, R.C.; Leifert, C.; Nodari, R.O.; Robinson, C.J.; Fagan, J. Teratogenic Effects of Glyphosate-Based Herbicides: Divergence of Regulatory Decisions from Scientific Evidence. Journal of Environmental and Analitycal Toxicololgy, S4:006. 2012.
Aris,A.; Leblanc, S. Maternal and fetal exposure to pesticides associated to genetically modified foods in Eastern Townships of Quebec, Canada. Reprod Toxicol.; v.31, n.4, p.528-533, 2011.
Arriola, P.E. and Ellstrand, N.C. Crop-to-weed flow in the genus Sorghum (Poaceae): spontaneous interspecific hybridization between johnsongrass, Sorghum halapense, and crop sorghum (S. bicolor). Am. J. Botany, 83: 1153-1160,1998.
Azevedo, E.S. Bioética é tão importante quanto aptidão científica. Folha de São Paulo, 8/01/95, p.6-12. 1995. Barton, J.; Crandon, J.; Kennedy, D.; Miller, H. A model protocol to assess the risk of agricultural introductions.
Nature Biotechnology, 15:845-848, 1997.
Beetham, P.R.; Kipp, P.B.; Sawycky, X.L.; Artzen, C.J.; May, G.D. A tool for functional plant genomics: Chimeric RNA/DNA oligonucleotides cause in vivo gene-specific mutations. Proceedings of National Academy of Sciences, v.96, p.8.874-8.878, 1999.
Beiguelman, B. Genética, Ética e Estado. Brazilian Journal of Genetics, 20:525-530, 1997. Bergelson, J., Purrington, C.B. e Wichmann, G. Promiscuity in transgenic plants Nature, 395:25,1998. Berlan L.P.; Lewontin R.C. Menace of the Genetic-industrial complex. Le Monde Diplomatique, Janeiro de 1999, pp.
8-9. Bøhn, T.; Primicerio, R.; Hessen, D.O.; Traavik, T. Reduced Fitness of Daphnia magna Fed a Bt-Transgenic Maize
Variety. Archives of Environmental Contamination and Toxicology, v. 55, p. 584–592. 2008. Bøhn, T.; Traavik, T.; Primicerio, R. Demographic responses of Daphnia magna fed transgenic Bt-maize.
Ecotoxicology, v.19, n.2, p.419-430, 2010. Bøhn, T.; Cuhra,M.; Traavik, T.; Sanden, M.; Fagan, J.; Primicerio, R. Compositional differences in soybeans on the
market: Glyphosate accumulates in Roundup Ready GM soybeans. Food Chemistry, 153: 207–215, 2014. Bossdorf, O.; Richards, C.L.; Pigliucci, M. Epigenetics for ecologists. Ecology Letters, V. 11, n. 2, p. 106–115, 2008. Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.V. Construction of a genetic linkage map in man using restriction
fragment length polymorphisms. Am J Hum Genet 32:314-331. 1980.
Brasileiro, A.C.M. e Dusi, D.M.A. Transfortmação genética de plantas. In: Torres, A.C.; Caldas, L.S.; Buso, J.A. Cultura de Tecidos e Transformação genética de plantas. Brasília, Embrapa/CBAB, p.679-735. 1999.
Brewer, G. J. Introduction to Isozyme Techniques. London and New York: Academic Press, 1970, 186p. British Medical Association. The impact of genetic modification on agriculture, food and health. Londres, BMA, 18p.
1999. Busconi, M.; Baldi, G.; Lorenzoni, C.; Fogher, C.. Gene flow from transgenic rice to red rice (Oryza sativa L.) in the
field. Plant Biology 16 (2014) 22–27 © 2013. doi:10.1111/plb.1202 Caruso, D. Intervention. San Francisco, Hybrid Vigor Press, 2006, 252p. Catarino, R.; Ceddia, G, Areal, F.J.; Park, J. The impact of secondary pests on Bacillus thuringiensis (Bt)
crops. Plant Biotechnology Journal, 13, p. 601–612, 2015. doi: 10.1111/pbi.12363 Chen, D., Ye, G., Yang, C., Chena, Y., Wu, Y. Effect of introducing Bacillus thuringiensis gene on nitrogen
metabolism in cotton. Field Crop Res, 92:1–9. 2005. Chèvre, A-M., Baranger, F.E.A., Renard, M. Gene Flow from transgenic crops, Nature, 389:924, 1998. Cho, Y., Qiu, Y-L., Kuhlman, P., Palmer, J.D. Explosive invasion of plan mitochondria by a group I intron.
Proceedings of National Academy of Sciences, 95:14.244-14.249. 1998. CIPRIANO, J., CARRASCO, J., ARBÓS, M. (2006). La Impossible Coexistencia. 04 de abril de 2006. Assemblea
Pangesa y GREENPEACE. http://www.greenpeace.org/raw/content/espana/reports/copy-of-la-imposible-coexisten.pdf Acesso em 20 de novembro de 2006.
Cohen, N.C.; Chang, C.Y.; Boyer, H.W.; Helling, R.B. Construction of biologically functional bacterial plasmids in vitro. Proc. Nat. Acad. Sci USA, 70(11):3240-3244. 1973.
Colyer, P.D.; Kirkpatrick, T.L.; Caldwell, W.D.; Vernon, P.R. Root-Knot Nematode reproduction and root galling severity on related conventional and transgenic cotton cultivars. The Journal of Cotton Science, v.4, p.232-236, 2000.
Costa, F.L.; Silva, N.C.A.; Ogliari, J.B. Maize diversity in southern Brazil: indication of a microcenter of Zea mays L. Genet Resour Crop Evol DOI 10.1007/s10722-016-0391-2

107
Cox, C. Glyphosate. J. of Pesticide Reform, 18(3):3-16. 1998. Crow, J. F. Basic concepts in population, quantitative, and evolutionary genetics. New York: W.H. Freeman and
Company, 1986. 273p. CTNBio. Legislação. Comunicados. Processos. [on-line] URL: http:/www.mct.gov.br/ctnbio. 1998 e 1999. CTNBio. Comunicado n.º 54 , de 29 de setembro de 1998. Publicado no D.O.U. nº 188 de 01de outubro de 1998,
Seção 03, p.56. Darwin, C. On the origin of species. New York: Athenaeum, 1967. 502p. A facsimile of the first edition,
Londres,1859. Doebley, J. Molecular evidence for gene flow among Zea species. BioScience, 40:443-448. 1990. Emberlin, Jean. The dispersal of maize pollen Zea mays – A report based on evidence available from publications
and internet sites. National Pollen Research Unit, University College, Worcester WR2 6 AJ, United Kingdom. 1999
Ellstrand, N.C.; Prentice, H.C.; Hancok, J.F. Gene flow and introgression from domesticated plants into their wild relatives. Annu. Rev. Ecol. Syst. v.30, p.539-563, 1999.
EMBRAPA .Compatibilidade de Agrotóxicos com Rhizobium spp. e a simbiose das leguminosas. Seropédica, CNPBS, 1986.
FAO. Biotechnology. Roma, Food and Agriculture Organization. 12p. [on-line] URL: http:/www.fao.org/unfao/bodies/COAG. 1999.
Fehr, W.R. Principles of Cultivar Development. London: Macmillan Publ., Vol. 1 e 2. 1987. FERREIRA, M.E.; GRATTAPAGLIA, D. Introdução ao uso de marcadores moleculares em análise genética. 3.ed.
Brasília: EMBRAPA-CENARGEN, 1998. p.220. (EMBRAPA-CENARGEN. Documentos, 20). Finardi Filho F. Plantas transgênicas e a segurança alimentar. In: 50 Reunião Anual da SBPC, Porto Alegre, RS,
1999. Fontes, E.; Santos, I.K.M.; Olas, C.A. A biossegurança de Plantas Cultivadas transgênicas. In: Teixeira, P. e Valle,
S. ((Orgs.). Biossegurança. Uma abordagem multidisciplinar. Rio de Janeiro, Fundação Osvaldo Cruz, 1996. p.313-327.
Ganz, P.R., A.K. Dudani, E.S. Tackaberry, R. Sardana, C. Sauder, X. Cheng and I. Altosaar. Expression of Human Blood Proteins in Transgenic Plants: The Cytokine GM-CSF as a Model Protein. In Owen, M.R.L.; Pen, J. (eds.).Transgenic Plants: A Production System for Industrial and Pharmaceutical Proteins. New York, John Wiley and Sons, pp.281-297.1996.
Garcia, M. A. E Altieri, M.A. (2005). Transgenic Crops: Implications for Biodiversity and Sustainable Agriculture. Bulletin of Science, Technology & Society, Vol. 25, No. 4, 335-353.
Gassmann, A.J., Petzold-Maxwell, J.L., Keweshan, R.S., Dunbar, M.W. Field-evolved resistance to Bt maize by western corn rootworm. PLoS One, 6:e22629. 2011.
Gassmann, A.J., Petzold-Maxwell, J.L., Clifton, E.H., Hoffmann, A.M., Ingber, D.A., Keweshan, R.S., Dunbar, M.W. and Ryan S. Keweshan . Field-evolved resistance by western corn rootworm to multiple Bacillus thuringiensis toxins in transgenic maize. vol. 111 no. 14, 5141–5146, 2013. doi: 10.1073/pnas.1317179111
Gepts P. The use of molecular and biochemical markers in crop evolution studies. Evol Biol 27: 51-94. 1993. Gliddon, C. Memorandum: Select Committee on European Communities Second Report – Written Evidence, [on-
line] URL: http:/www.publications.parliament.uk/pa/Id199899?Idselect/Ideucom/11/11we22.htm. 1999. Goudet, J. Fstat version 1.2: a computer program to calculate Fstatistics. Journal of Heredity. 86(6): 485-486. 1995.
Programa disponível em http://www2.unil.ch/popgen/softwares/fstat.htm Greene, A.E., Allison, R.F. Recombination between viral RNA and transgenic plant transcripts. Science, 263:1423-
1425. 1994. Griffiths, M. The millennium choice - genetic engineering ou natural law? 1999.
http:/www.bt.internet.com/~nlpvessex. 1999. Griffiths, M. The millennium choice - genetic engineering or natural law? [on-line] URL:
http:/www.bt.internet.com/~nlpvessex. 1999. Guerra, M.P.; Nodari, R.O. Plantas transgênicas: os desafios da comunidade científica. O Biológico, São Paulo,
v.61, n.2, p.107-112. 1999. Hansen Jesse, L.C.; Olbrycki, J.J. Field deposition of Bt transgenic corn pollen: lethal effects on the monarch
butterfly. Oecologia, v.125, n.2, p.241. 2001. Hardell, H. e Eriksson, M. A Case-Control Study of Non-Hodgkin Lymphoma and Exposure to Pesticides. Cancer,
85 (6): 1999. Hilbeck, A.; Moar, W.J.; Pusztai-Carey, M.; Filipini, A.; Bigler, F. Toxicity of Baccilus thuringiensis Cry1Ab toxin to
the predator Chrysoperla carnea (Neoroptera: Chrysopidae). Environmental Entomology, 27(5):1255-1263. 1998.
Ho, M-W., Traavik, T., Olsvik, O., Tappeser, B., Howard, C.V., von Weizsacker, C., McGavin, G.C. Gene Technology and gene ecology of infectious diseases, Microbial Ecology in Health and Disease, 10:33-59, 1998.
Huang, F.; Buschman, L.L.; Higgins, R.A.; McGaughey, W.H. Inheritance of resistance to Bacillus thuringiensis toxin (Dipel ES) in the Europian Corn Borer. Science, 284:965-967. 1999.
HUNTER, R. L.; MARKET, C. L. Histochemical demonstration of enzymes separated by zone electrophoresis in starch gels. Science. v. 125, p. 1294-1295, 1957.
Jablonka, E.; Lamb, M.J. EVOLUÇÃO EM QUATRO DIMENSÕES - DNA, comportamento e a história da vida, S~ao Paulo: Companhia das Letras, 512p, 2010. Com ilustrações de Anna Zeligowski.

108
Jacob, F.; Monod, J. "Genetic regulatory mechanisms in the synthesis of proteins". Journal of Molecular Biology 3: 318–356. 1961. doi:10.1016/S0022-2836(61)80072-7
Jeffrey, A.J.; Wilson, V.; Thein, S.L. Hypervariable minisatellite regions in human DNA. Nature, 314:67-73. 1985. Kohli, A. et al. Molecular characterization of transforming plasmid rearrangement in transgenic rice reveals a
recombination hotspot in the CaMV promoter and confirms the predominance of microhomology mediated recombination. The Plant Journal, 17(6):591-601. 1999.
Kremer, R.J.; Donald, P.A.; Keaster, A.J.; Minor, H.C. Herbicide Impact on Fusarium spp. and Soybean Cyst Nematode in Glyphosate-Tolerant Soybean. [on-line] URL: http://www.asa-cssa-sssa.org/cgi-bin/abstract_database_search.cgi?objective=Kremer. 2000.
Kruger, M., Van Rensburg, J., Van den Berg, J. Resistance to Bt maize in Busseola fusca (Lepidoptera: Noctuidae) from Vaalharts. S Afr Environ Entomol, 40:477–483. 2011.
Kutukdjian, G.B. The needs for bioethics is universal. Biotechnology and Development Monitor, 31:24, 1997. Lacey, H. A controvérsia sobre os transgênicos: Questões científicas e éticas. São Paulo: Idéias & letras. 2006. Lacey, H. Há alternativas ao uso dos transgênicos. Novos Estudos: CEBRAP, São Paulo. v.78, p. 31-39. jul. 2007. LACEY, H. Values and objectivity in science: the current controversy about transgenic crops. Lanham, MD:
Lexington Books, 2005. Lacey, H.. Crescimento econômico, meio ambiente e sustentabilidade. Mercado ético: sua plataforma global para
sustentabilidade. Disponível em: <http://mercadoetico.terra.com.br/?p=11519&preview=true>. Acesso em: 6 dez. 2009.
Lander Es; Green P; Abrahamson J; Barlow A; Daly Mj; Lincoln Se; Newburg L. MAPMAKER: an interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics, v. 1,
p.174-181, 1987. Lewin, B. Genes V. New York, Oxford University Press, 1272p.1994. Lewis, P.O.; Zaykin, D. Genetic data analysis: computer program for the analysis of allelic data. Version 1. 0 (d15).
Free program distributed by the authors over the internet from the GDA. Programa disponível em: http://www.eeb.uconn.edu/people/plewis/software.php (GDA)
Lewontin, R. The triple helix. Cambridge, Harvard University Press, 136p. 2000. Lewontin, R. It ain‟t necessarily so – The dream of the human genome and other illusions. New York: New York
Review Books, 2000. 330p. LEWONTIN, R.C.; HUBBY, J. L. A molecular approach to the study of genic heterozygosity in natural populations.
11. Amount of variation and degree of heterozygosity in natural populations of Drosophzla pseudoobscura. Genetics, 54: 595-609, 1966.
LOSEY, J.E.; RAYOR, L.S.; CARTER, M.E. Transgenic pollen harms monarch larvae. Nature, v.399, p.214, 1999. Malatesta, M., Caporaloni, C., Gavaudan, S., Rocchi, M.B., Serafini, S., Tiberi, C., Gazzanelli, G. Ultrastructural
morphometrical and immunocytochemical analyses of hepatocyte nuclei from mice fed on genetically modified soybean. Cell Struct Funct. 27(4):173-80. 2002
MARKET, C.; MOLLER, F. Multiple forms of enzymes: Tissue, autogene and species specific patterns. Proceedings of the National Academy of Sciences, Washington, v.45, p.753-763. 1959.
Michelmore RW, Paran I, Kesseli RV. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations.Proceedings of the National Academy of Sciences.88(21):9828-9832. 1991
Lieber, RR. & Romano-Lieber NS. Risco, incerteza e as possibilidades de ação na saúde ambiental. Rev. Bras. Epidemiol., 6(2):121-34, 2003.
Losey, J.E,; Rayor, L.S.; Carter, M.E. Transgenic pollen harms monarch larvae. Nature 399:214, 1999.). Mason H.S., Lam D.M.K., Arntzen C.J . Expression of hepatitis B surface antigen in transgenic plants. Proc. Natl.
Acad. Sci. USA 89:11745-11749. 1992. Millstone, E.; Brunner, E.; Mayer, S. Beyond „Substantial equivalence‟. Nature, 401:525-526, 1999. Momma, A. N. Rotulagem de Plantas Transgênicas e o Agronegócio. Revista de Direito Ambiental, v.16, p.153-162,
1999. (Depoimento à Câmara dos Deputados em 13 de abril de 1999, Brasilia, DF.). More "Funny" Honey. FOEE Biotech Mailout, v.6, n.5, p.6, 31/07/2000. Nardi, N.B.; Teixeira, L.A.K.; Silva, E.F.A. Terapia gênica. Ciência e Saude Coletiva, vol.7, n.1, p.109-116, 2002. Nature. A tragic setback. Nature, vol. 420, n.6912, p.116-118. 2002. New Scientist. Section: This Week, p. 6, 21 Octu.10/2000. Nguyen, H.T., Jehle, J.A. Expression of Cry3Bb1 in transgenic corn MON88017. J Agric Food Chem, 57:9990–
9996. 2009. Nielsen, K., Bones, A.M., Smalla, K., van Elsas, J.D. Horizontal gene transfer from transgenic plants to terrestrial
bacteria – a rare event ? Fems Microbiiology Reviews, v.22, n.2, p.79-103, 1998. Nielsen, K.; van Elsas, J.D.; Smalla, K. Transformation of Acinetobacter sp. Starin BD413 with transgenic plant DNA
in soil microcosms and effetcts of kamycin on selection of transformants. Applied and Environmental Microbiology, 66(3):1237-1242. 2000.
NODARI, R. O., TSAI, S. M., GUZMAN, P., GILBERTSON, R., GEPTS, P. Towards an integrated linkage map of common bean. Iii. Mapping factors controlling host-bacterium interactions. Genetics. , v.134, p.341 - 350, 1993.
Nodari, R.O.; Guerra, M.P. Biossegurança de plantas transgênicas. In: GÖRGENS, F.S.A. (Org.). Riscos dos transgênicos. Petrópolis: Vozes, 2000a. p.39-60.
Nodari, R.O.; Guerra, M.P. Implicações dos transgênicos na sustentabilidade ambiental e agrícola. História, Ciências, Saúde – Manguinhos, Rio de Janeiro, v.7, n.2, p.481-491, 2000b.

109
Nodari, R.O.; Guerra, M.P. Avaliação de riscos ambientais de plantas transgênicas. Cadernos de Ciência &Tecnologia, Brasília, v.18,n.1, p.61-116, 2001.
Nodari. R.O.; Guerra. M.P. Da transformação em bactérias (1928) às plantas transgênicas. Ciência e Ambiente, Santa Maria, v. 26, n. 1, p. 49-65, 2003.
Nodari,R.O.; Guerra, M.G. A agroecologia: suas estratégias de pesquisa e sua relação dialética com os valores da sustentabilidade, justiça social e o bem estar humano. Estudos Avançados, v.29, p.183 - 207, 2015.
Olsen, K.M., Daly, J.C., Holt, H.E., Finnegan, E.J. Season-long variation in expression of Cry1Ac gene and efficacy of Bacillus thuringiensis toxin in transgenic cotton against Helicoverpa armigera (Lepidoptera: Noctuidae). J Econ Entomol, 98:1007–1017. 2005.
Padgette, S.R., Taylor, N.B., Nida, D.L., Bailey, M.R., MacDonald, J., Holden., L.R., Fuchs, R.L. The composition of glyphosate-tolerant soybean seeds is equivalent to that of conventional soybeans. Journal of Nutrition, v.126, p.702-716, 1996.
Palmiter, R.D., Brinster, R.L., Hammer, R.E., Trumbauer, M.E., Rosenfeld, M.G., Birnberg, N.C., Evans, R.M. Dramatic growth of mice that develop from eggs microinjected with metallothionein-growth hormone fusion genes. Nature, vol. 300, n. 5893, p. 611-615, 1982.
Paran, I; Michelmore,R.W. Development of reliable PCR-based markers linked to downy mildew resistance genes in lettuce. Theoretical Applied Genetics, 85:985-993. 1993.
Parmenter D.L., Boothe J.G. and Moloney M.M. Production and purification of recombinant hirudin from plant seeds. In: Owen, M.R.L.; Pen, J. (eds.).Transgenic Plants: A Production System for Industrial and Pharmaceutical Proteins. John Wiley & Sons. p.p. 261-280. 1996.
Pedrosa, Fábio O., Monteiro, Rose Adele, Wassem, Roseli, Cruz, Leonardo M., Ayub, Ricardo A., Colauto, Nelson B., Fernandez, Maria Aparecida, Fungaro, Maria Helena P., Grisard, Edmundo C., Hungria, Mariangela, Madeira, Humberto M. F., Nodari, Rubens O., Osaku, Clarice A., Petzl-Erler, Maria Luiza, Terenzi, Hernán, Vieira, Luiz G. E., Steffens, Maria Berenice R., Weiss, Vinicius A., Pereira, Luiz F. P., Almeida, Marina I. M., Alves, Lysangela R., Marin, Anelis, Araujo, Luiza Maria, Balsanelli, Eduardo, Baura, Valter A., Chubatsu, Leda S., Faoro, Helisson, Favetti, Augusto, Friedermann, Geraldo, Glienke, Chirlei, Karp, Susan, Kava-Cordeiro, Vanessa, Raittz, Roberto T., Ramos, Humberto J. O., Ribeiro, Enilze Maria S. F., Rigo, Liu Un, Rocha, Saul N., Schwab, Stefan, Silva, Anilda G., Souza, Eliel M. Genome of Herbaspirillum seropedicae Strain SmR1, a Specialized Diazotrophic Endophyte of Tropical Grasses. PLOS Genetics (Online). , v.7, p.e1002064, 2011. doi:10.1371/journal.pgen.1002064
PELLANDA, P. S. P., NODARI, R. O. A propriedade intelectual dos transgênicos no Brasi In: A efetivação do direito de propriedade para o desenvolvimento sustentável: relatos e proposições.1 ed. Florianópolis : Fundação Boiteux, 2010, v.1, p. 381-412.
Pereira, L.V. Animais transgênicos – nova fronteira do saber. Ciência e Cultura, vol. 60, n.2, p.40-42, 2008. Peterson, G.; Cunningham, S.; Deutsch, L.; Erickson, J.; Quinlan, A.; Raez-Luna, E.; Tinch, R.; Troel, M.;
Woodbury, P.; Zens, S. The risks and benefits of genetically modified crops: a multidisciplinary perspective. Conservation Ecology, v.4, n.1, p.13 [on-line] URL: http:/www.consecol.org/vol4/iss1/art13. 2000.
Pham-Delégue M. H. Risk assessment of transgenic oilseed rape on the honeybee. Paris, INRA, Laboratoire de neurobiologie comparée des invertebrés, 1997. p. 1-3,
Pimentel, D.; Hunter, M.S.; Lagro, J.A.; Efroymson, R.A.; Landers, J.C.; Mervis, F.T.; Mccarthy, C.A.; Boyd, A.E. Benefits and risks of genetic engineering in agriculture. BioScience, v.39, n.9, p.606-614, 1989.
Raffensperger, C.; Tikckner, J. Protecting public health & the environment: implementing the precautionary principle. Washington: Island Press, 1999, 385p.
RAMESH, S.V.; RATNAPARKHE, M.B.; KUMAWAT, G.; GUPTA, G.K.; HUSAIN, S.M. Plant miRNAome and antiviral resistance: a retrospective view and prospective challenges. Virus Genes, v.48(1), 1-14, 2014.
Ramirez-Romero, R.; Desneux, N.; Decpurtye, A.; Chaffiol, A.; Pham-Délègue, M. Does Cry1Ab protein affect learning performances of the honey bee Apis mellifera L. (Hymenoptera, Apidae)? Ecotoxicology and Environmental Safety, v. 70, p. 327-333, 2008.
Reisig, D.D.; Reay-Jones, F.P.F. Inhibition of Helicoverpa zea (Lepidoptera: Noctuidae) growth by transgenic corn expressing Bt toxins and development of resistance to Cry1Ab. Environ. Entomol. 1–11. 2015. DOI: 10.1093/ee/nvv076
Richetti, A. Estimativa do Custo de Produção de Soja no Sistema Plantio Direto, Safra 2010/2011, para Mato Grosso do Sul. Comunicado Técnico 160. Dourado, MS: Embrapa, 2010, 9p.
Rosi-Marshall, E. J.; Tank, J.L.; Royer, T.V.; R. Whiles, M.; Evans-White, M.; Chambers, C.; A. Griffiths, N.; Pokelsek, J.; Stephen, M.L. (2007) Toxins in transgenic crop byproducts may affect headwater stream ecosystems. PNAS, v. 104, n.41, p.16204–16208, 2007.
Rótolo, G.C.; Francis, C.; Craviottoa, R.M.; Ulgiatid, S. Environmental assessment of maize production alternatives: Traditional, intensive and GMO-based cropping patterns. Ecological Indicators , 57: 48–60. 2015. doi:10.1016/j.ecolind.2015.03.036
Sagstad, A.; Sanden, M., Haugland, Ø., Hansen, A.C., Olsvik, P.A., Hemre, G.I. Evaluation of stress- and immune-response biomarkers in Atlantic salmon, Salmo salar L., fed different levels of genetically modified maize (Bt maize), compared with its near-isogenic parental line and a commercial suprex maize. Journal of Fish Diseases 30(4):201-12. 2007
Sax, K. The association of size differences with seed-coat pattern and pigmentation in Phaseolus vulgaris. Genetics 8:552–560. 1923.
Saxena, D; Flores,S.; Stotzky, G. Insecticidal toxin in root exudates from Bt corn. Nature, v.402, p.480, 1999. Science. Microbial management. Science, Washington, v.284, p.1301-1307, 1999.

110
Science. GeneTerapy: Questions Remain on Cause of Death in Arthritis Trial. Science, vol.317, n.5845, p. 1665. 2007
Science 1999. CLINICAL TRIALS: Gene Therapy Death Prompts Review of Adenovirus Vector. Science, 286:2244-2245, 1999.
Schafer MG, Ross AA, Londo JP, Burdick CA, Lee EH, Travers SE, et al. (2011) The Establishment of Genetically Engineered Canola Populations in the U.S. PLoS ONE 6(10): e25736. doi:10.1371/journal.pone.0025736
Schmidt, E.E.; Taylor, D.S.; Prigge, J.R.; Barnett, S.; Capecchi, M.R. Illegitimate Cre-dependent chromosome rearrangements in transgenic mouse spermatids. PNAS, v.97, p.13.702-13.707, 2000.
Seralini, G.E., Cellier, D., Spiroux de Vendomois, J. New. New analysis of a rat feeding study with a genetically modified maize reveals signs of hepatorenal toxicity. Archives Environmental Contamination and Toxicology, v.52, p.596-602, 2007.
Séralini, G-E.; Clair, E.; Mesnage, R.; Gress, S.; Defarge, N.; Malatesta, M.; Hennequin, D.; de Vendômois, J.S. Long term toxicity of a roundup herbicide and a roundup-tolerant genetically modified maize. Food and Chemical Toxicology, 50:4221-4231, 2012.
Showalter, A.M., Heuberger, S., Tabashnik, B.E., Carrière, Y., Coates, B. A primer for using transgenic insecticidal cotton in developing countries. J Insect Sci, 9:22. 2009. SNOW, A.A.. Illegal gene flow from transgenic creeping bentgrass: the saga continues. Molecular Ecology (2012) 21, 4663–4664.
Singer, P. Ética Prática. São Paulo: Martins Fontes, 1984. SINGH, S. P., NODARI, R. O., GEPTS, P. Genetic Diversity in Cultivated Common Bean: I. Allozymes. Crop
Science, v.31, p.19 - 23, 1991. Soltis, D.E.; Soltis, P.S.; Dudley, T.R. Isozymes in Plant Biology. Portland: Dioscorides Press, 1989, 268p. Stefenon VM, Nodari RO, Reis MS (2003) Padronização de protocolo AFLP e sua capacidade informative para
análise da diversidade genética em Araucaria angustifolia. Scientia Forestalis 64:163-171 Stefenon VM, Nodari RO, Guerra MP (2004) Genética e conservação de Araucaria angustifolia: III. Protocolo de
extração de DNA e capacidade informativa de marcadores RAPD para análise da diversidae genética em populações naturais. Biotemas 17:47-63
Stefenon VM, Gailing O, Finkeldey R (2006) Searching natural populations of Araucaria. angustifolia: conservation strategies for forest genetic resources in southern Brazil. In: Bohnes J and Paar E (eds) Forstliche Genressources als Produktionsfaktor. Proceedings of the 26
th Tagung der Arbeitsgemeinschaft Forstgenetik
und Forstpflanzenzüchtung. Hessen-Forst Forsteinrichtung, Information, Versuchwesen. Stefenon VM, Gailing O, Finkeldey R (2007a) Genetic structure of Araucaria angustifolia (Araucariaceae) in Brazil:
Implications for the in situ conservation of genetic resources. Plant Biology 9:516-525 Stefenon VM, Gailing , Finkeldey R (2007b) Recovery and conservation of Araucaria forest in Brazil through
plantation‟s establishment: a genetic point of view. In: Proceedings of the Conference on International Agricultural Research for Development, Tropentag 2007. http://www.tropentag2007/abstract/full/160.pdf cited 20 may 2008
Stefenon VM, Gailing O, Finkeldey R (2008a) The role of gene flow in shaping genetic structures of the subtropical conifer species Araucaria angustifolia. Plant Biology 10:356-364
Stefenon VM, Behling H, Gailing O, Finkeldey R (2008b) Evidences of delayed size recovery in Araucaria angustifolia populations after post-glacial colonization of highlands in Southeastern Brazil. Anais da Academia Brasileira de Ciências. 80:433-443.
Stefenon VM, Gailing O, Finkeldey R (2008c) Genetic structure of plantations and the conservation of genetic resources of Brazilian pine (Araucaria angustifolia). Forest Ecology and Management. 255:2718-2725
Steven, S., Zemetra, R., Francis, Y.L., Jones, S.S. Production of herbicide-resistant jointed goatgrass (Aegilops cylindrica) X wheat (Triticum aestivum) hybrids in the field by natural hybridization. [on line]. Washington, 1999. . [on-line] URL: http:/www.nalusda.gov/ttic/tektran.
Storer, N.P., Babcock, J.M., Schlenz, M., Meade, T., Thompson, G.D., Bing, J.W., Huckaba, R.M. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico.J Econ Entomol, 103:1031–1038. 2010.
Syvanen, M. Horizontal ene transfer: evidence and possible consequences. Annu Rev Genet, 28:237-261. 1994 Tabashnik, B.E.; Brévault, T.; Carrière, Y. Insect resistance to Bt crops: lessons from the first billion acres. Nature
Biotechnology, 31: 510–521, 2013. Tappeser, B.; Jäger, M.; Eckelkamp, C. Survival, persistence, transfer: An update on current knowledge on GMs
and the fate of ther recombinant DNA. Penang: TWN, 1999, 44p. Theologis, A., Ecker, J.R., Palm,C.J., Federspiel, N.A.,et.al. and Davis,R.W. Sequence and analysis of chromosome
1 of the plant Arabidopsis thaliana. Nature, 408(6814):816-20. 2000. Tiedje, J.M.; Colwell, R.K.; Grossman, Y. L.; Hodson, R.E.; Lenski, R.E; Mack, R.N.; Regal, P.J. The planned
introduction of genetically engineered organisms – Ecological considerations and recommendations. Ecology, v.70, n.2, p.298-315. 1989.
Traavik, T. Too early may be too late. Research Report for DN 1999-1. Ecological risks associated with the use of naked DNA as biological tool for research, production and therapy. Trondheim, Norway, 1999. 106p.
Traavik, T.; Heinemann, J. Genetic Engineering and Omitted Health Research: Still No Answers to Ageing Questions. Third World Network, 36p. 2007. (ISBN: 978-983-2729-76-1).
Vale, S.; Teixeira, P. (org). Biossegurança:uma abordagem multidisciplinar. Rio de Janeiro: Ed. FIOCRUZ, 1996. Vazquez, A.S. Ética. 4 ed. São Paulo: Civilização Brasileira, 247p. 1980.

111
Vazquez-Padrón, R.I.; Moreno-Fierro, L.; Neri-Bazán, L.; Riva, G. A.; López-Revilla, R, Intragastric and intraperitoneal administration of Cry1Ac protoxin from Bacillus thuringiensis induces systemic and mucosal antibody response in mice. Life Sciences 64(21):1897-1912, 1999.
Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; Lee, T.V.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kulper, M.; Zabeau, M. AFLP: a new technique for DNA fingerprinting. Nucleic Acids Research, v.23, n.1, p.4407-4414,
1995. Wallace, B. The search for the gene. Ithaca, Cornell University Press, 224p. 1992. Walsh, L.O.; McCormick, C.; Martin, C.; Stocco, D.M.. Roundup Inhibits Steroidogenesis by Disrupting
Steroidogenic Acute Regulatory (StAR) Protein Expression. Environ Health Perspect 108:769-776 (2000). [Online 12 July 2000]
Wang, S. et al. Tarnishing Silver Bullets: Bt Technology Adoption, Bounded Rationality and the Outbreak of Secondary Pest Infestations in China. Cornell University. Selected Paper prepared for presentation at the American Agricultural Economics, Association Annual Meeting Long Beach, CA, July 22-26, 2006.
Welsh, J.; Mclelland, M. Fingerprinting genomes using PCR with arbitrary primers. Nuleic Acids Res, 18:7213-7218. 1990.
Williams, J.G.K.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res, 18:6531-6535. 1990.
Wilson, H.D. Gene Flow in squash species. BioScience, 40:449-455. 1990. Wipff, J.K.; Fricker, C.R. Determining gene flow of transgenic creeping bentgrass and gene transfer to other
bentgrass species. Diversity, 16(1,2):36-39. 2000. Wolfenbarger, L.L.; Phifer, P.R. The ecological risks and benefits of genetically engineered plants. Science, v.290,
p.2088-2093, 2000. Xing, H-L.; Dong, L.; Wang, Z-P.; Zhang, H-Y.; Han, C-Y.; Liu, B.; Wang, X-C.; Chen, Q-J. A CRISPR/Cas9 toolkit
for multiplex genome editing in plants. BMC Plant Biology, 14:327. 2014 Zhu, T.; Mettenburg, K.; Peterson, D.J.; Tagliani, L.; Baszczynski, C.L. Targeted manipulation of maize genes in vivo
using chimeric RNA/DNA oligonucleotides. Proceedings of National Academy of Sciences, v.96, p.8868-8873. 1999.
Zhu, Y.; Chen, H.; Fan, J.; Wang, Y. Li, Y.; Chen, J.; Fan, J.X.; Yang, S.; Hu, L.; Leung, H.; Mew, T.W.; Teng, P.S.; Wang, Z.; Mundt, C.C. Genetic diversity and disease control in rice. Nature, v.406, p.718-722, 2000.
Zolla, L., Rinalducci, S., Antonioli, P., Righetti, P.G. Proteomics as a complementary tool for identifying unintended side effects occurring in transgenic maize seeds as a result of genetic modifications. Journal of Proteome
Research 7: 1850--‐1861. 2008.


















