
FABÁCEAS PALEÓGENAS DA REGIÃO SUDESTE DE MINAS … · distribuída por muitos biomas terrestres,...
20
343 Rev. bras. paleontol. 17(3):343-362, Setembro/Dezembro 2014 © 2014 by the Sociedade Brasileira de Paleontologia doi: 10.4072/rbp.2014.3.06 INTRODUÇÃO A família Fabaceae é monofilética com base em dados morfológicos e moleculares e constituída de três subfamílias, conforme Polhill & Raven (1981): Mimosoideae, Caesalpinoideae e Faboideae (= Papilionoideae). Também chamada de “Leguminosae A.L. de Jussieu”, tem aproximadamente 730 gêneros envolvendo mais de 19.000 espécies. É a terceira das maiores famílias botânicas modernas, depois das Orchideaceae e Asteraceae; amplamente distribuída por muitos biomas terrestres, desde desertos de altas latitudes e altitudes (regiões árticas ou alpinas) até florestas tropicais pluviais ou sazonalmente secas, de regiões equatoriais, todavia são especialmente abundantes nas regiões tropicais, subtropicais e temperadas do mundo sob climas sazonais, semi-áridos e áridos (Lewis et al., 2005; Simpson, 2006; Wojciechowski et al., 2006). São plantas de hábito variado: na forma arbórea (de 4 a 30 m de altura), nas matas FABÁCEAS PALEÓGENAS DA REGIÃO SUDESTE DE MINAS GERAIS, FORMAÇÃO ENTRE-CÓRREGOS, BACIA DE AIURUOCA, BRASIL MARY ELIZABETH CERRUTI BERNARDES-DE-OLIVEIRA MAG/CEPPE/UnG; Programa de Pós-Graduação em Geoquímica e Geotectônica, IGc/USP - Laboratório de Paleobotânica e Palinologia, Rua do Lago, 562, Cidade Universitária, 05508-900, São Paulo, SP, Brasil. [email protected] MARIA JUDITE GARCIA Laboratório de Paleobotânica e Palinologia, IGc/USP. [email protected] MARIA CRISTINA DE CASTRO-FERNANDES & KAROLINE GONÇALVES PEREIRA Mestrado em Análise Geoambiental, CEPPE/UnG, Praça Tereza Cristina n.1, 07023-070, Guarulhos, SP, Brasil. [email protected]; [email protected] ABSTRACT – PALEOGENE FABACEAE OF THE SOUTHEAST REGION OF MINAS GERAIS, ENTRE-CÓRREGOS FORMATION, AIURUOCA BASIN, BRAZIL. The family Fabaceae, one of the most abundant and diversified of the recent eudicots, has a fossil record in the taphoflora of the Entre-Córregos Formation (Eocene-Oligocene). This formation is constituted by papiraceous shales deposited under euxinic lacustrine conditions of the Aiuruoca taphrogenic basin, in the southeastern area of the Minas Gerais State. This basin is inserted in the Cenozoic Rifting System of the Southeast Brazil. The Fabaceae record presents itself like impressions of detached and attached folioles of the macrophylls of a new species of Caesalpinia (Subfamily Caesalpinoideae), two new species and one form identified only in generic level of Machaerium (Subfamily Faboideae) and as affine pollen grains. Considering their modern climatic, habitat and habits preferences it is possible to infer that they are components of a humid climate Neotropical flora, being member of the upper arborescent stratum of the association (15 to 20 m high), in the submountain area of the Atlantic Forest. In Brazil, these genera, were only known from Paleogene record from the Brazilian Southeast (Fonseca, Aiuruoca, Taubaté, Tanque and São Paulo basins). The total palynological association suggests an Eocene-Oligocene age. Key words: Paleogene Caesalpinoideae and Faboideae, Aiuruoca Basin, Brazil. RESUMO – A família Fabaceae, uma das mais abundantes e diversificadas dentre as eudicotiledôneas atuais, tem registro fossilífero na tafoflora da Formação Entre-Córregos (Eoceno-Oligoceno). Essa formação é constituída de folhelhos papiráceos depositados sob condições lacustres redutoras, na bacia tafrogênica de Aiuruoca, sudeste do Estado de Minas Gerais, que, por sua vez, provavelmente é coetânea ao sistema de rifteamento cenozoico do Sudeste brasileiro. Esse registro apresenta- se como impressões de folíolos destacados e em conexão de macrófilos de uma espécie nova de Caesalpinia (Subfamília Caesalpinoideae), de duas espécies novas e uma forma identificada apenas em nível genérico de Machaerium (Subfamília Faboideae) e na forma de grãos de pólen afins. Considerando-se suas preferências climáticas, de habitat e de hábito atuais, pode-se inferir tratar-se de elementos integrantes de uma flora neotropical de clima quente e úmido, fazendo parte do estrato arbóreo superior da associação (15 a 20 m de altura), em áreas submontanas da Floresta Atlântica. Trata-se de gêneros, até o momento, com registro paleógeno apenas no Sudeste brasileiro (bacias de Fonseca, Aiuruoca, Taubaté, Tanque e São Paulo). A associação palinológica total sugere idade eoceno-oligocena. Palavras-chave: Caesalpinoideae e Faboideae paleógenas, bacia de Aiuruoca, Brasil.
Transcript of FABÁCEAS PALEÓGENAS DA REGIÃO SUDESTE DE MINAS … · distribuída por muitos biomas terrestres,...

343
Rev. bras. paleontol. 17(3):343-362, Setembro/Dezembro 2014© 2014 by the Sociedade Brasileira de Paleontologiadoi: 10.4072/rbp.2014.3.06
INTRODUÇÃO
A família Fabaceae é monofilética com base em dados morfológicos e moleculares e constituída de três subfamílias, conforme Polhill & Raven (1981): Mimosoideae, Caesalpinoideae e Faboideae (= Papilionoideae). Também chamada de “Leguminosae A.L. de Jussieu”, tem aproximadamente 730 gêneros envolvendo mais de 19.000 espécies. É a terceira das maiores famílias botânicas
modernas, depois das Orchideaceae e Asteraceae; amplamente distribuída por muitos biomas terrestres, desde desertos de altas latitudes e altitudes (regiões árticas ou alpinas) até fl orestas tropicais pluviais ou sazonalmente secas, de regiões equatoriais, todavia são especialmente abundantes nas regiões tropicais, subtropicais e temperadas do mundo sob climas sazonais, semi-áridos e áridos (Lewis et al., 2005; Simpson, 2006; Wojciechowski et al., 2006). São plantas de hábito variado: na forma arbórea (de 4 a 30 m de altura), nas matas
FABÁCEAS PALEÓGENAS DA REGIÃO SUDESTE DE MINAS GERAIS, FORMAÇÃO ENTRE-CÓRREGOS, BACIA DE AIURUOCA, BRASIL
MARY ELIZABETH CERRUTI BERNARDES-DE-OLIVEIRAMAG/CEPPE/UnG; Programa de Pós-Graduação em Geoquímica e Geotectônica, IGc/USP - Laboratório de Paleobotânica
e Palinologia, Rua do Lago, 562, Cidade Universitária, 05508-900, São Paulo, SP, Brasil. [email protected]
MARIA JUDITE GARCIALaboratório de Paleobotânica e Palinologia, IGc/USP. [email protected]
MARIA CRISTINA DE CASTRO-FERNANDES & KAROLINE GONÇALVES PEREIRAMestrado em Análise Geoambiental, CEPPE/UnG, Praça Tereza Cristina n.1, 07023-070, Guarulhos, SP, Brasil.
[email protected]; [email protected]
ABSTRACT – PALEOGENE FABACEAE OF THE SOUTHEAST REGION OF MINAS GERAIS, ENTRE-CÓRREGOS FORMATION, AIURUOCA BASIN, BRAZIL. The family Fabaceae, one of the most abundant and diversifi ed of the recent eudicots, has a fossil record in the taphofl ora of the Entre-Córregos Formation (Eocene-Oligocene). This formation is constituted by papiraceous shales deposited under euxinic lacustrine conditions of the Aiuruoca taphrogenic basin, in the southeastern area of the Minas Gerais State. This basin is inserted in the Cenozoic Rifting System of the Southeast Brazil. The Fabaceae record presents itself like impressions of detached and attached folioles of the macrophylls of a new species of Caesalpinia (Subfamily Caesalpinoideae), two new species and one form identifi ed only in generic level of Machaerium (Subfamily Faboideae) and as affi ne pollen grains. Considering their modern climatic, habitat and habits preferences it is possible to infer that they are components of a humid climate Neotropical fl ora, being member of the upper arborescent stratum of the association (15 to 20 m high), in the submountain area of the Atlantic Forest. In Brazil, these genera, were only known from Paleogene record from the Brazilian Southeast (Fonseca, Aiuruoca, Taubaté, Tanque and São Paulo basins). The total palynological association suggests an Eocene-Oligocene age.
Key words: Paleogene Caesalpinoideae and Faboideae, Aiuruoca Basin, Brazil.
RESUMO – A família Fabaceae, uma das mais abundantes e diversifi cadas dentre as eudicotiledôneas atuais, tem registro fossilífero na tafofl ora da Formação Entre-Córregos (Eoceno-Oligoceno). Essa formação é constituída de folhelhos papiráceos depositados sob condições lacustres redutoras, na bacia tafrogênica de Aiuruoca, sudeste do Estado de Minas Gerais, que, por sua vez, provavelmente é coetânea ao sistema de rifteamento cenozoico do Sudeste brasileiro. Esse registro apresenta-se como impressões de folíolos destacados e em conexão de macrófi los de uma espécie nova de Caesalpinia (Subfamília Caesalpinoideae), de duas espécies novas e uma forma identifi cada apenas em nível genérico de Machaerium (Subfamília Faboideae) e na forma de grãos de pólen afi ns. Considerando-se suas preferências climáticas, de habitat e de hábito atuais, pode-se inferir tratar-se de elementos integrantes de uma fl ora neotropical de clima quente e úmido, fazendo parte do estrato arbóreo superior da associação (15 a 20 m de altura), em áreas submontanas da Floresta Atlântica. Trata-se de gêneros, até o momento, com registro paleógeno apenas no Sudeste brasileiro (bacias de Fonseca, Aiuruoca, Taubaté, Tanque e São Paulo). A associação palinológica total sugere idade eoceno-oligocena.
Palavras-chave: Caesalpinoideae e Faboideae paleógenas, bacia de Aiuruoca, Brasil.

344 REVISTA BRASILEIRA DE PALEONTOLOGIA, 17(3), 2014
tropicais (onde podem estar entre as árvores mais altas), assim como arbustos, ervas anuais ou perenes, trepadeiras e ainda como umas poucas aquáticas. Sua principal característica é apresentar fruto do tipo legume ou vagem. Seu sucesso pode ser atribuído ao desenvolvimento de relações simbiontes com bactérias fi xadoras de nitrogênio, viventes em seus nódulos radiculares, que as tornam capazes de converter o nitrogênio do ar em sais nitrogenados, conferindo-lhes a possibilidade de colonizar áreas com solos pobres (Marchiori, 1995).
As folhas dessa família são, geralmente, compostas (paripinadas ou imparipinadas), com pulvínos na base do pecíolo, folíolos glabros, normalmente assimétricos ou de base assimétrica, com possível face inferior ferruginosa e textura cartácea a coriácea, margem inteira, venação primária pinada, veias secundárias broquidódromas; presença de estípulas. Tendem a perder os folíolos ou a sofrer fusão deles (Hickey & Wolfe, 1975; Simpson, 2006).
Atualmente, ocorrem no Brasil cerca de 180 gêneros e 1.500 espécies; alguns biomas brasileiros são considerados centros de diversidade para o grupo (Marchiori, 1997).
A família diversifi cou-se em suas subfamílias, possivelmente durante o Neocretáceo (74 a 79 Ma), com base em dados de seqüência molecular, segundo Lavin et al. (2005). Entretanto, muitos consideram que o clado tronco originou-se no Neopaleoceno, a cerca de 1 a 2,5 Ma antes do clado coroa, atingindo uma rápida diversifi cação das fabáceas entre 50 e 40 Ma (Wojciechowski et al., 2004). Os autores Wing et al. (2009) atribuíram a fabáceas cerca de cinco a sete morfotipos foliares e sete tipos de frutos e sementes aladas, encontrados na Formação Cerrejón (Paleoceno superior da Colômbia). Estes foram considerados como os registros paleógenos mais antigos (~58 Ma) da fl ora neotropical pluvial ou ombrófi la densa. Os primeiros macrofósseis, seguramente, identifi cados como Fabaceae eram flores e frutos de mimosoídeas e cesalpinoídeas do sudeste norte-americano e do sul da Inglaterra (Herendeen & Crane, 1992; Herendeen et al.,1992) datados como paleoceno-eocenos. Muller (1981) ressaltou que Tricolporopollenites ludensis Gruas-Cavagnetto (Aubry et al., 1977; Gruas-Cavagnetto, 1977) do Eoceno superior da França tem afi nidade com grãos de pólen de Faboideae. Entretanto, estudos mais recentes têm sugerido, com base em evidências fósseis e fi logenética, que as fabáceas, originalmente, evoluíram como espécies herbáceas em regiões temperadas, áridas e/ou semi-áridas ao longo da área costeira do mar de Tethys, durante o Paleógeno (Schrire et al., 2005a,b). Outros autores consideraram que a África ou as Américas não podem ser descartadas como áreas de origem da família (Wojciechowski et al., 2006).
Achados de fabáceas sulamericanos paleocenos recentemente encontrados, em fl oras argentinas da Patagônia (Formação Salamanca, ~61,7 Ma) por Iglesias et al. (2007) e por Brea et al. (2008) e na fl ora colombiana de Cerrejón (~58 Ma) por Wing et al. (2009), estão entre os mais antigos da família e revelaram uma diversidade de formas e grande abundância, rapidamente alcançada no Paleoceno do continente como já haviam proposto Lavin et al. (2005).
No material fitofossilífero eoceno-oligoceno (~40 a 30 Ma) de Aiuruoca (Estado de Minas Gerais, sudeste do
Brasil), estão, claramente, representadas as subfamílias Caesalpinoideae e Faboideae através de fragmentos de folhas compostas, folíolos destacados e grãos de pólen. Isto signifi ca que as duas subfamílias já estariam presentes no Brasil não muito depois de seus registros mundiais mais antigos, ou seja, ainda dentro do Paleógeno. Esse fato será mencionado com mais detalhes no subtítulo sobre distribuição geográfi ca e estratigráfi ca das formas estudadas.
ÁREA DE ESTUDO
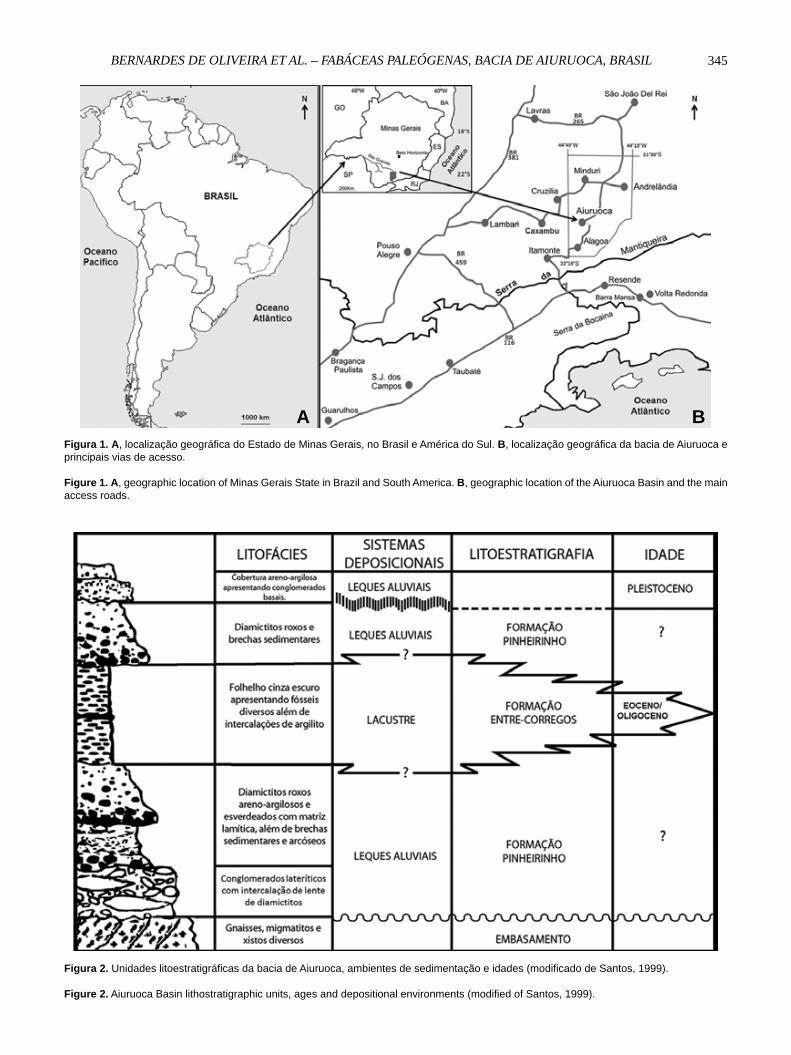
A bacia sedimentar de Aiuruoca localiza-se no Estado de Minas Gerais, nas proximidades das fronteiras deste com os estados de São Paulo e Rio de Janeiro, abrangendo parte da Serra da Mantiqueira e do Planalto do Alto Rio Grande, no sudeste do Brasil (Figura 1).
A bacia compreende duas unidades litoestratigráfi cas informais, interdigitadas que repousam sobre o embasamento cristalino: formações Pinheirinho e Entre-Córregos, conforme defi nido por Santos (1999). A Formação Entre-Córregos é constituída por folhelhos papiráceos, cinza escuro e arenitos fi nos, argilosos, cinza claro, depositados em ambiente lacustre redutor. Informações geográfi cas e geológicas mais detalhadas podem ser vistas em Franco-Delgado & Bernardes-de-Oliveira (2004) e sumarizadas na Figura 2.
Em afl oramento de folhelhos papiráceos da Formação Entre-Córregos, situado à margem esquerda do córrego Entre-Córregos, ocorre uma tafofl ora de idade eocena-oligocena, datada com base em palinologia por Garcia et al. (2000). Essa assembléia tafofl orística é composta de macrofósseis foliares e caulinares, parcialmente identifi cados e descritos em Franco-Delgado (2002) e Franco-Delgado & Bernardes-de-Oliveira (2004), cujos gêneros foram listados na Tabela 1. Os gêneros ocorrentes na bacia de Aiuruoca foram registrados em outras localidades eocenas ou oligocenas brasileiras, mas estão ausentes, até o momento, no registro paleoceno de todo o país.
MATERIAL E MÉTODOS
O afl oramento, de onde os fi tofósseis procedem, pode ser descrito, da base para o topo, da seguinte forma: uma camada de 1,42 m de espessura de folhelhos papiráceos argilosos, cinza médio, contendo pipídeos, alguns fósseis vegetais com predominância de caules e pequena quantidade de folhas e frutos, insetos e peixes; sobreposta por uma camada de 0,10 m de um ritmito constituído por lâminas de folhelho alternadas com lâminas de arenito fi no, argiloso, micáceo, rico em fi tofósseis, de onde provêem as amostras estudadas; recobre-a uma camada de 0,24 m de folhelho papiráceo, micáceo, cinza escuro, pobre em fi tofósseis, muito alterado por intromissões de raízes atuais, e, fi nalmente, se sobrepõe um solo orgânico argiloso, escuro e permeado por raízes atuais, de 0,60 m de espessura, conforme Bedani (2002) e Franco-Delgado & Bernardes-de-Oliveira (2004).
No Laboratório de Palinologia e Paleobotânica da UnG, após preparação mecânica, os fi tofósseis foram observados e fotografados em estereomicroscópio C. Zeiss, modelo-
345BERNARDES DE OLIVEIRA ET AL. – FABÁCEAS PALEÓGENAS, BACIA DE AIURUOCA, BRASIL
Figura 1. A, localização geográfica do Estado de Minas Gerais, no Brasil e América do Sul. B, localização geográfica da bacia de Aiuruoca e principais vias de acesso.
Figure 1. A, geographic location of Minas Gerais State in Brazil and South America. B, geographic location of the Aiuruoca Basin and the main access roads.
Figura 2. Unidades litoestratigráficas da bacia de Aiuruoca, ambientes de sedimentação e idades (modificado de Santos, 1999).
Figure 2. Aiuruoca Basin lithostratigraphic units, ages and depositional environments (modified of Santos, 1999).
A B

346 REVISTA BRASILEIRA DE PALEONTOLOGIA, 17(3), 2014
Tabela 1. Outras ocorrências paleógenas brasileiras dos gêneros da tafoflora de Entre-Córregos. Localidades: 1, Fonseca, Minas Gerais; 2, Gandarela, Minas Gerais; 3, Aiuruoca, Minas Gerais; 4, Mina Nossa Senhora da Guia, Tremembé, São Paulo; 5, Fazenda Santa Fé, Tremembé, São Paulo; 6, km 11 da Rodovia Quiririm/Campos do Jordão, Taubaté, São Paulo; 7, Mina Itaquaréia I, Itaquaquecetuba, São Paulo.
Table 1. Other Brazilian Paleogene occurrences of the Entre-Córregos taphoflora genera. Locations: 1, Fonseca, Minas Gerais; 2, Gandarela, Minas Gerais; 3, Aiuruoca Basin, Minas Gerais; 4, Mina Nossa Senhora da Guia, Tremembé, São Paulo; 5, Fazenda Santa Fé, Tremembé, São Paulo; 6, km 11 of Quiririm route/Campos do Jordão, Taubaté, São Paulo; 7, Mine Itaquaréia I, Itaquaquecetuba, São Paulo.
TÁXONS ÉPOCAS / LOCALIDADES
EOCENO/ OLIGOCENO OLIGOCENO
ORDEM FAMÍLIA SUBFAMÍLIA GÊNEROS 1 2 3 4 5 6 7
LAURALES Lauraceae Nectandra X
MAGNOLIALES Annonaceae Annona X X
MALVALES Malvaceae
Apeiba X
Luehea X X X X
Campomanesia X
MYRTALES Myrtaceae
Eugenia X
Myrciaria X X
Psidium X X X
FABALES FabaceaeCaesalpinoideae Caesalpinia X X
Faboideae Machaerium X X X
MALPIGHIALES Passifloraceae Passiflora X
SAPINDALES Sapindaceae Sapindus X X X
Stemi SV6, visando sua análise, descrição e a documentação gráfi ca do padrão de venação e dos detalhes morfológicos. As fotomacrografi as foram obtidas no Laboratório de Fotografi a da UnG com auxílio do Prof. Elcio Macias de Melo.
As dimensões lineares e angulares do limbo e venação foram mensuradas através da utilização do Programa Axiovision Zeiss e da utilização de paquímetro digital Mitotoyo, sobre desenhos obtidos com câmara clara. No cálculo da área foliar, bem como na classifi cação dos espécimes, segundo sua área, e para descrição da venação e morfobiometria foliares foram utilizados os métodos e designações propostos no Manual de Arquitetura Foliar (Ellis et al., 2009).
Na sistemática angiospérmica foram respeitadas as relações fi logenéticas propostas pelo APG III (2009) e Watson & Dallwitz (1992) e, para táxons de categoria inferior a ordem, foi seguida a classifi cação apresentada por Simpson (2006) e Souza & Lorenzi (2008).
Na identifi cação taxonômica, foram feitas comparações com a chave de Hickey & Wolfe (1975) até ordem ou família e com espécimes fósseis e atuais (da literatura) e da fl ora atual (herborizados). O material atual herborizado foi clarifi cado, colorido e montado em lâmina, conforme procedimento proposto por Foster (1952) e, recentemente, descrito por Dutra & Stranz (2002). A preparação química e montagem de lâminas foram feitas no Laboratório de Paleobotânica e Palinologia do IGc/USP.
Tanto o material fóssil como o material de comparação clarificado encontram-se depositados no Laboratório de
Palinologia e Paleobotânica “Prof. Dr. Murilo Rodolfo de Lima” da UnG: o material fóssil, sob os números UnG/3T- 239 – UnG/ 3T- 258 e o material atual, em laminário, sob os números UnG 04, 44, 51b, 95, 100, 135, 136.
Na extração dos palinomorfos seguiram-se os métodos convencionais descritos em Uesugui (1979), com modificações e os seguintes procedimentos: (i) foram utilizadas 20 g da amostra de folhelho, fragmentado; (ii) ácido fl uorídrico-PA, para eliminar a sílica; ácido clorídrico-PA, para eliminar os silicofl uoretos que se formaram no processo anterior; (iii) hidróxido de potássio a 10%, para eliminar a matéria orgânica amorfa e fi toclastos. Foram realizadas sempre três lavagens com água destilada, entre cada uma das etapas dos reagentes. As lâminas foram montadas com Entellan.
A análise dos palinomorfos foi efetuada ao microscópio óptico Olympus BX 51 com câmara digital acoplada e a identifi cação baseou-se na literatura nacional e internacional disponível para o Paleógeno, Neógeno e Quaternário, além de consulta a actuopalinotecas de referência.
SISTEMÁTICA PALEOBOTÂNICA
ANGIOSPERMAE Clado FABIDEAS APG III, 2009
Ordem FABALES Bromhead, 1838Família FABACEAE Lindley, 1836 ss.
(=LEGUMINOSAE)

347BERNARDES DE OLIVEIRA ET AL. – FABÁCEAS PALEÓGENAS, BACIA DE AIURUOCA, BRASIL
Os folíolos ora estudados, isolados ou em conexão a fragmentos de raque, ostentam dentre os elementos apontados como típicos da família: lâminas ou bases assimétricas, margem inteira, peciólulos com pulvínulos infl ados, venação broquidódroma ou craspedódroma, dentre os apontados por Herendeen et al. (1992).
Subfamília CAESALPINOIDEAE De Candolle, 1825
Caesalpinia L., 1753
São fabáceas espinhosas pantropicais, englobando mais de 100 espécies, desde porte arbóreo até herbáceo. Entre as mais notáveis estão: pau-brasil (Caesalpinia echinata Lam.), pau-ferro [Caesalpinia leiostachya (Benth.) Ducke] e sibipiruna (Caesalpinia peltophoroides Benth.). As folhas são compostas, bipinadas geralmente alternas, imparipinadas, com número variável de pares de pinas (dois a nove pares) e uma terminal; cada pina porta de cinco a 27 pares de folíolos, normalmente oblongos ou elípticos ou trapezoidais, de tamanho variado; ápice arredondado ou retuso; base assimétrica.
Espécie tipo. Caesalpinia brasiliensis L., 1753.
Caesalpinia veraechinataformis sp. nov.(Figuras 3A-B,D)
2002 Caesalpinia cf. C. echinata Lam. Franco-Delgado, Dissertação de Mestrado, UnG, est. XV, fi gs. 1 e 4, fi g. text. 10, p. 79-81.2009 Caesalpinia cf. C. echinataformis Berry. Veiga, Dissertação Mestrado, UnG, est. IV, fi gs. A,C, fi g. tex. 6.8, p. 79-81.
Holótipo. UnG/3T - 239 (ex-VAI 19), Figura 3B.Parátipos. UnG/ 3T - 240 a UnG/3T-244 (ex-VAI 27, 50, 18, 21 e s/nº preliminar, respectivamente) Figuras 3A (UnG/3T-240) e D (UnG/3T-241).Estratigrafi a. Formação Entre-Córregos, bacia de Aiuruoca, Estado de Minas Gerais.Idade. Eoceno-Oligoceno.Procedência. Do ritmito constituído de lâminas de folhelho alternadas com lâminas de arenito fi no, argiloso, micáceo, do afl oramento da margem esquerda do córrego Entre-Córregos, entre Aiuruoca e Minduri, Minas Gerais.Etimologia. veraechinataformis refere-se à grande semelhança de forma entre estes folíolos dispersos e aqueles da espécie atual Caesalpinia echinata.Diagnose. Impressões de folíolos isolados de proporção laminar 2:1, com heterofoliolia marcante, de textura subcoriácea, elípticos a oblongos, inequilaterais, assimétricos, principalmente na base cuneada ou truncada; ápice arredondado ou retuso; margem inteira; venação pinada, veia mediana bem marcada que se afi na para o ápice; veias secundárias broquidódromas festonadas de calibre grosso, veias intersecundárias fracas formadas por veias terciárias coalescentes, do tipo percorrente alternada, sinuosas de
direção admedial, com início nos loops marginais, terminando próximo à veia primária, com a qual fazem ângulo obtuso, em ramifi cações mais fi nas, abertas ou fechadas em aréolas, com vênulas internas em pequenos ramos livres. Glândulas distribuídas pela lâmina foliolar.Diagnosis. Isolate folioles impressions of laminar ratio 2:1, marked heterophilly of leafl ets, subcoriaceous texture, elliptical to oblong, unequilateral, asymmetric, mostly in the cuneate or truncated base; rounded or retuse apex; entire rim; pinnate venation, midrib sharp clear tapering toward apex; festooned brochidodromous secondary veins of thick caliber, weak intersecondary veins as coalescent tertiary veins, of alternate percurrent category and sinuous admedially direction, from the marginal loops arriving near the midrib, with which they are at obtuse angle, with thinner ramifi cations, open or closed forming areoles, with two or more branched freely ending ultimate veins. Distributed glands aroud laminar surface of the leafl ets.Descrição. Cerca de seis impressões de folíolos isolados, elípticos a ovado-estreitos (2:1); assimétricos; ápice arredondado ou retuso; base cuneada ou truncada e assimétrica; margem inteira; textura subcoriácea, com glândulas distribuídas pela lâmina, pecíolo não preservado (Figuras 3A-B, D). Medidas: 10 a 15 mm de comprimento, por 4 a 7 mm de largura. Venação pinada broquidódroma festonada. A veia primária é de calibre resistente (4%) e curso suavemente sinuoso. As veias secundárias, do tipo broquidódromo festonado, divergem em ângulos agudos moderados (entre 45º e 75º) que são mais agudos na área proximal e distal (45º a 54º). Têm calibre moderado e curso entre reto a ligeiramente curvo. No semilimbo maior, os ângulos formados pelos “loops” das veias secundárias com as suprajacentes são agudos (48º a 62º), no semilimbo menor, esses ângulos são retos a obtusos (90º a 110º). Veias intersecundárias fracas formadas por veias terciárias coalescentes, do tipo percorrente alternada, sinuosas de direção admedial, de ângulo obtuso em relação à primária.Comparação e discussão. Todas essas características são diagnósticas de folíolos do gênero Caesalpinia, conforme mencionado anteriormente, e muito afins às da espécie atual C. echinata (= pau-brasil). Observando os folíolos das folhas compostas de C. echinata, verifica-se uma acentuada heterofoliolia entre aqueles de posição mais distal, mediana e proximal da mesma pina, os quais exibem bases assimétricas mais cuneadas ou nitidamente truncadas e ápices arredondados ou retusos. Portanto, os folíolos isolados, com formas ligeiramente diferentes, aqui descritos, poderiam ser procedentes de posições distintas até de uma mesma pina (ver Figura 4).
Na comparação, por exemplo, de folíolos atuais clarifi cados da espécie Caesalpinia echinata (lâminas UnG-51b, 135 e UnG-100, na Figura 3C), com o espécime fóssil UnG/ 3T-239 (Figura 3B), torna-se evidente a semelhança de características arquiteturais foliolares.
Apesar da grande semelhança entre o folíolo fóssil e o folíolo atual de Caesalpinia echinata, por se tratar de forma muito antiga (eoceno-oligocena) e detectada apenas por uns poucos

348 REVISTA BRASILEIRA DE PALEONTOLOGIA, 17(3), 2014
Figura 3. A-B,D, Caesalpinia veraechinataformis sp. nov. C, Caesalpinia echinata. A, UnG/3T-243, folíolo isolado com detalhe de venação; B, UnG/3T-239, holótipo, folíolo isolado com detalhe de venação broquidódroma; C, UnG-100-255a, lâmina; D, UnG/3T-241, outro folíolo isolado. Escalas: A = 0,5 mm; B,D = 1 mm; C = 2 mm.
Figure 3. A-B,D, Caesalpinia veraechinataformis sp. nov. C, Caesalpinia echinata. A, UnG/3T-243, isolated leaflet with detail of the venation, B, UnG/3T-239, holotype, isolated leaflet with detail of the brochidodromous venation; C, UnG-100-255a, slide; D, UnG/3T-241, another isolated leaflet. Scale bars: A = 0,5 mm; B,D = 1 mm; C = 2 mm.
folíolos dispersos enquanto, na forma atual, a folha composta é bipinada e os folíolos são subopostos ou subalternos, opta-se pelo não uso do mesmo epíteto específi co atual.
A forma eocena Caesalpinia echinataformis Berry (Berry, 1935) encontrada na Formação Fonseca, no sudeste de Minas Gerais, também mencionada por Mello et al. (2002, fi gura 4b), foi aproximada dos gêneros Senna e Cassia por Fanton et al. (2009). Os espécimes aqui observados são distintos daquele apresentado por Berry (1935), Mello et al.
(2002) e Fanton et al. (2009), verifi cando-se uma diferença quanto à forma e à proporção foliolar e, a impossibilidade de comprovação do arranjo foliolar na raque. Caesalpinia cf. C. echinataformis Berry, encontrada por Veiga (2009, estampa IV, fi guras A,C e fi gura-texto 6.8) na Formação Tremembé, tem todas as características, incluindo-se a proporção foliolar (comprimento: largura), compatíveis, com a forma Caesalpinia veraechinataformis sp. nov., excetuando-se o tamanho (ligeiramente maior no espécime estudado), caráter
A B
C D

349BERNARDES DE OLIVEIRA ET AL. – FABÁCEAS PALEÓGENAS, BACIA DE AIURUOCA, BRASIL
sem importância dada a sua variação numa mesma folha. Isto permite colocar as formas procedentes da Formação Tremembé na sinonímia da espécie ora descrita para a Formação Entre-Córregos.
Não há uma quantidade de dados diagnósticos que justifi que a comprovação da mesma designação específi ca, mas dada sua similaridade de arquitetura foliolar com a espécie atual Caesalpinia echinata, opta-se pela designação Caesalpinia veraechinataformis sp. nov. para esses espécimes foliolares da Formação Entre-Córregos.Distribuição geográfi ca e ecologia da espécie atual afi m. Caesalpinia echinata (popularmente designada pau-brasil) é uma forma de porte arbóreo, de 8 m até 30 m de altura, semidecídua, heliófi ta ou esciófi ta, de folhas compostas bipinadas. Ocorre, preferencialmente, em terrenos secos, sendo típica do interior da floresta primária densa. Há evidências da distribuição geográfi ca da espécie endêmica à costa oriental atlântica brasileira, na Floresta Pluvial Atlântica, desde o Rio Grande do Norte até o Rio de Janeiro ou mesmo desde o Ceará. Porém, não se sabe ao certo sua frequência ao longo dessa área, devido à drástica redução das populações pela exploração antropogênica predatória (Lorenzi, 2002; Corrêa, 2003).Distribuição estratigráfi ca sulamericana. De acordo com Berry (1935) e Ferrusquía-Villafranca & González-Guzmán (2005), o gênero Caesalpinia aparece no registro fóssil desde o início do Cretáceo Superior do sudoeste da América do Norte (México) alcançando a Europa, durante o Eoceno. Na bacia de Fonseca (Formação Fonseca, de idade eoeocena a mesoeocena, Minas Gerais), o gênero foi registrado por Berry (1935) na forma de um espécime de folha composta, identifi cada pelo epíteto echinataformis Berry 1935. A criação
do epíteto específi co foi justifi cada pelo fato do espécime apresentar disposição oposta dos folíolos, diferentemente da suboposta a subalternada de C. echinata. Referiram-se a este registro Wey et al. (1999) e Mello et al. (2002). Entretanto, Fanton et al. (2009) discordaram dessa identificação e sugeriram nova afi nidade com a tribo Cassieae (gêneros Senna e Cassia).
Considerando a proposta de Fanton et al. (2009) em relação aos espécimes da bacia de Fonseca, os folíolos isolados de Caesalpinia veraechinataformis sp. nov. ocorrentes tanto na bacia de Aiuruoca, Minas Gerais, como na de Taubaté, São Paulo, tornam-se os mais antigos registros de Caesalpinia em território brasileiro.
Subfamília FABOIDEAE (= PAPILIONOIDEAE De Candolle, 1825)
Tribo DALBERGIEAE
Machaerium Persoon, 1807
Gênero tipicamente neotropical, que se estende desde o sul do México até a Argentina. Sua maior diversidade está no Brasil (cerca de 120 das 130 espécies existentes), com formas escandentes predominantes na hiléia amazônica e arbóreas, no centro-sul. Na maioria dos biomas brasileiros apresenta grande riqueza específi ca (Sartori & Tozzi, 1998; Mendonça-Filho et al., 2007), enquanto apenas uma espécie ocorre na costa oeste da África (Rudd, 1977).
Conforme Bastos (1987), o gênero Machaerium apresenta folhas compostas imparipinadas, alternas, pecioladas, de dimensões variadas. Seus folíolos são alternos, em geral peciolulados, raramente sésseis, de textura cartácea a coriácea, de contorno oblongo a linear-oblongo, de ápice arredondado a retuso ou de contorno elíptico-lanceolado, de ápice acuminado. No geral, com folíolo apical distinto dos demais e com face adaxial glabra e abaxial pilosa ou glabra. Veia mediana, às vezes, ultrapassa o ápice do limbo, constituindo um mucron ou arista. Venação secundária craspedódroma ou camptódroma e veias terciárias livres ou formando aréolas com veias até quinta ordem de forma escorpioide.
Espécie-tipo. Machaerium ferrugineum (Willd.) Pers., sinônimo de M. quinata (Aubl.) Sandwith (Mendonça-Filho et al., 2007).
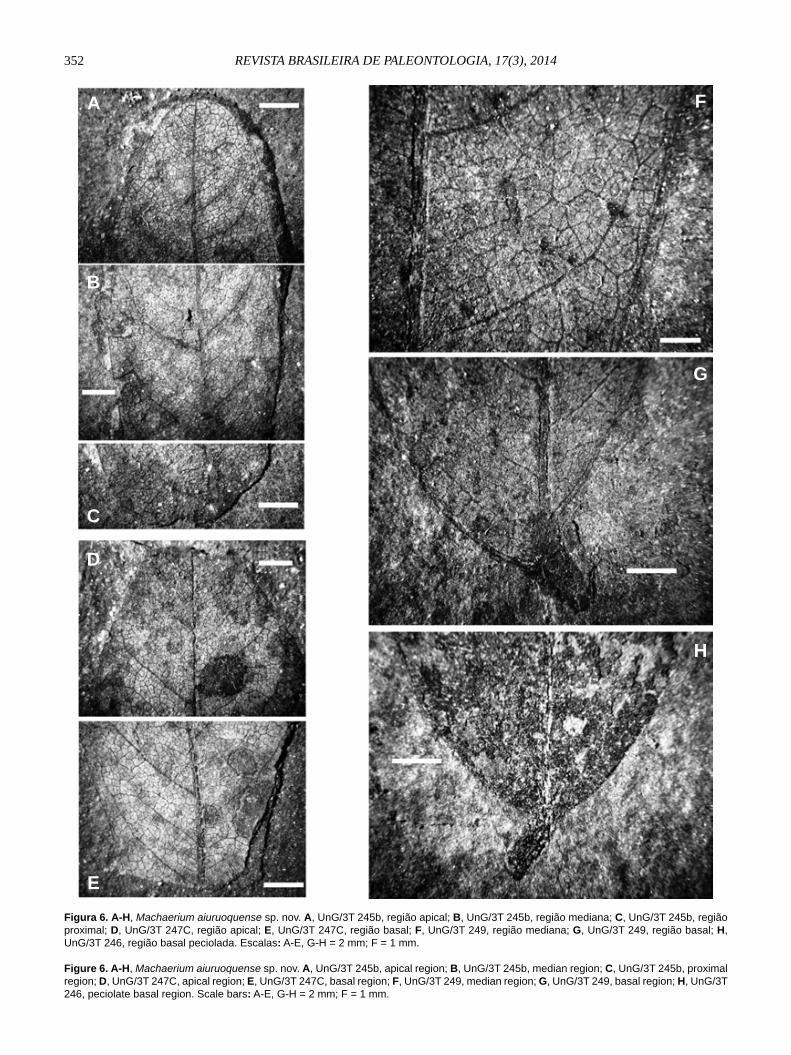
Machaerium aiuruoquense sp. nov.(Figuras 5A-E; 6A-H)
Holótipo. UnG/3T – 245 a,b ( Figuras 5C e E, Figuras 6A-C).Parátipos. UnG/3T- 246-249, 251 (Figuras 5A-B,D; 6D-H).Estratigrafi a. Formação Entre-Córregos, bacia de Aiuruoca, Minas Gerais.Idade. Eoceno-Oligoceno.Procedência. Do ritmito constituído de lâminas de folhelho alternadas com lâminas de arenito fi no, argiloso, micáceo, do afl oramento da margem esquerda do córrego Entre-Córregos, entre Aiuruoca e Minduri, Estado de Minas Gerais.
Figura 4. Folha composta de Caesalpinia echinata exibindo folíolos de diferentes formas e tamanhos ao longo das pinas, foto de Mario Franco (agosto/2011). Extraída de http://plantas-ornamentais.blogspot.com.br/2011/08/pau-brasil-caesalpinia-echinata-lam.html, em 2 de maio de 2014.
Figure 4. Compound leaf of Caesalpinia echinata showing leaflets of different shapes and sizes along the lenght of the pinnes, picture by Mario Franco (August/2011). From http://plantas-ornamentais.blogspot.com.br/2011/08/pau-brasil-caesalpinia-echinata-lam.html, May 2nd, 2014.

350 REVISTA BRASILEIRA DE PALEONTOLOGIA, 17(3), 2014
Etimologia. aiuruoquense, proveniente de Aiuruoca (Minas Gerais).Diagnose. Folíolos isolados, oblongos, de proporção 2,3:1 a 3:1; ápice arredondado ou retuso, base convexa ligeiramente assimétrica, margem inteira, textura cartácea, com pontos de inserção de pelos distribuídos pela lâmina foliolar; veia primária pinada de calibre médio e curso pouco encurvado; 8 ou 9 pares de veias secundárias em disposição suboposta a alterna, com ângulo quase reto; fechando em arco com secundária sub ou suprajacente; veias terciárias reticuladas ao acaso; veias quaternárias delimitando aréolas pentagonais a triangulares; venação de quinta ordem ramifi cando-se uma ou duas vezes; venação marginal fi mbriada.Diagnosis. Isolated leafl ets, oblong, rate length by width equal 2,3:1 to 3:1; rounded or retuse apex, convex, slender asymmetrical base, entire margin, papyraceous texture, with insertion points of hair dispersed for all laminar surface, pinnate venation of median caliber and slightly curved course; 8 or 9 pairs of secondary veins in sub-opposite to alternate disposition, with almost orthogonal angle jointing to supra or subjacent secondary veins; tertiary veins random reticulate; quaternary veins forming pentagonal to triangular areoles; fi fth order venation ramifying once or twice; fi mbriate marginal venation. Descrição. Impressões de face abaxial (UnG/3T-245b, Figuras 5E; 6A-C) e face adaxial (UnG /3T-245 a, Figura 5C) de folíolos isolados, assimétricos, geralmente elípticos (2,3:1); de ápice arredondado ou retuso, base convexa ligeiramente assimétrica e peciólulo pulvinado e encurvado, margem inteira, textura papirácea ou cartácea, com pontos distribuídos pela lâmina foliar; venação broquidódroma fraca. Veia primária de calibre resistente (2,6%) e de curso curvo. Nove pares de veias secundárias, em disposição suboposta a alterna, divergindo da primária em ângulo agudo moderado (45º - 70º). De calibre moderado e curso uniformemente em arco, unindo-se à secundária suprajacente em ângulos variáveis, com predomínio de, mais ou menos, ângulos retos (Figuras 5C,E). Veias intersecundárias de padrão variável: (Figuras 6E-G) simples (fechando em arco com as secundárias subjacentes ou suprajacentes) ou composto (formadas por terciárias coalescentes). Veias terciárias reticuladas ao acaso e oblíquas em relação à primária. Veias quaternárias emergem a ângulos variáveis, delimitando aréolas perfeitas pentagonais, quadrangulares ou triangulares. Venação de quinta ordem, de menor calibre, ramifi ca-se uma ou duas vezes. Venação marginal fi mbriada (Figuras 6A-B). Inúmeras estruturas tipo base de pelo distribuem-se pela lâmina foliolar.Comparação e discussão. Os espécimes foram identifi cados como pertencentes ao gênero Machaerium, dada a combinação de alguns critérios morfológicos foliolares observados em formas atuais e fósseis: (i) pela morfologia (folíolo assimétrico, de contorno elíptico a oblongo, base convexa, ligeiramente assimétrica, margem inteira, presença de peciólulos pulvinados, etc.); (ii) pela venação primária pinada de calibre consistente, secundária broquidódroma fraca, nove a dez pares de veias secundárias subopostas a alternas, com ângulo de divergência agudo moderado, de curso
uniformemente curvado, unindo-se à suprajacente em ângulo variável próximo a reto, esparsas veias intersecundárias; veias terciárias percorrentes bifurcadas, de percurso variado, quase sempre oposto; venação marginal fi mbriada (sensu Hickey, 1979), com percurso exatamente na borda da lâmina foliar, etc.; (iii) tratam-se de folíolos, aparentemente glabros, mas com abundantes poros ou pontos observados em sua superfície laminar que poderiam estar relacionados a pontos de inserção de pelos, como os observados em folíolos atuais do gênero Machaerium, onde fi níssimos pelos (só visíveis sob grande ampliação) recobrem toda a lâmina.
A espécie fóssil Machaerium nervosum Duarte & Rezende-Martins (1983), ocorrente na Formação Tremembé, Paleógeno da bacia de Taubaté, distingue-se desses espécimes pelas seguintes características: simetria do folíolo, venação secundária sinuosa, curvando-se repentinamente no terço distal, para formar ângulos agudos com a suprajacente, bem como no tamanho foliolar e na sua proporção, embora concorde no contorno oblongo, na forma arredondada da base, no ápice retuso e na venação secundária broquidódroma (Tabela 2).
A espécie fóssil Machaerium piranii Fittipaldi (1990), ocorrente na Formação Itaquaquecetuba (Eoceno-Oligoceno), bacia de São Paulo, assemelha-se aos espécimes estudados nas seguintes características: presença de estruturas de pontos ou poros dispersos pelo limbo e pela venação quaternária e quinquenária de padrão semelhante, bem como, pelas aréolas de desenvolvimento perfeito. Entretanto, difere desses espécimes por apresentar: tamanho foliolar maior e proporção menor, forma elíptica, simetria foliolar, ápice emarginado, veia primária retilínea, veias secundárias de curvatura abrupta, próximo à margem ao unir-se à suprajacente (Tabela 2).
Espécimes foliolares clarifi cados das seguintes espécies atuais foram utilizados na comparação: Machaerium hirtum (Vell.) Stellfeld (lâmina UnG-04); M. nyctitans (Vell.) Benth (lâmina UnG-44, Figura 5F); M. villosum Vog (lâminas UnG-95 e UnG-136), procedentes do Herbário da Prefeitura Municipal de São Paulo. Também foram utilizados na comparação espécimes foliolares da espécie M. scleroxylon Tul., conforme características fornecidas por Mendonça Filho et al. (2007) (Tabela 2).
Os espécimes fósseis estudados diferem dos folíolos da espécie M. hirtum por seu tamanho maior, pela proporção foliolar menor, resultante de uma largura proporcionalmente maior, pela pilosidade mais esparsa, em ambas as faces foliolares e pela venação secundária broquidódroma arqueada em vez de craspedódroma (Tabela 2).
A distinção de Machaerium villosum, conforme dados de Sartori & Tozzi (1998), evidencia-se pelo tamanho e proporção foliolares muito menores nos espécimes fósseis, na forma foliolar tendendo para lanceolada a obovada, de ápice reto, naquela espécie, e oblonga de ápice arredondado ou retuso, nas formas fósseis (Tabela 2).
Assemelham-se a Machaerium scleroxylon pela forma basal arredondada, por apresentar ambas as faces pilosas e venação broquidódroma e distinguem-se pelo tamanho foliolar menor, mas com proporção foliolar tendendo a ser igual ou menor (Tabela 2).

351BERNARDES DE OLIVEIRA ET AL. – FABÁCEAS PALEÓGENAS, BACIA DE AIURUOCA, BRASIL
Figura 5. A-E, Machaerium aiuruoquense sp. nov. A, UnG/3T-249; B, UnG/3T-247; C, UnG /3T-245a, holótipo; D, UnG/3T-248; E, UnG /3T-245b, holótipo, folíolos isolados com detalhe de venação. F, Machaerium nyctitans, lâmina UnG-44. Escalas: A, D = 4 mm; B = 5 mm; C = 6 mm; E = 9 mm; F = 3 mm.
Figure 5. A-E, Machaerium aiuruoquense sp. nov. A, UnG/3T-249; B, UnG/3T-247; C, UnG /3T-245a holotype; D, UnG/3T-248; E, UnG /3T-245b, holotype, isolated leaflets with detail of the venation. F, Machaerium nyctitans, slide UnG-44. Scale bars: A, D = 4 mm; B = 5 mm; C = 6 mm; E = 9 mm; F = 3 mm.
A B C
D E F
352 REVISTA BRASILEIRA DE PALEONTOLOGIA, 17(3), 2014
Figura 6. A-H, Machaerium aiuruoquense sp. nov. A, UnG/3T 245b, região apical; B, UnG/3T 245b, região mediana; C, UnG/3T 245b, região proximal; D, UnG/3T 247C, região apical; E, UnG/3T 247C, região basal; F, UnG/3T 249, região mediana; G, UnG/3T 249, região basal; H, UnG/3T 246, região basal peciolada. Escalas: A-E, G-H = 2 mm; F = 1 mm.
Figure 6. A-H, Machaerium aiuruoquense sp. nov. A, UnG/3T 245b, apical region; B, UnG/3T 245b, median region; C, UnG/3T 245b, proximal region; D, UnG/3T 247C, apical region; E, UnG/3T 247C, basal region; F, UnG/3T 249, median region; G, UnG/3T 249, basal region; H, UnG/3T 246, peciolate basal region. Scale bars: A-E, G-H = 2 mm; F = 1 mm.
A
B
C
D
E
H
G
F
353BERNARDES DE OLIVEIRA ET AL. – FABÁCEAS PALEÓGENAS, BACIA DE AIURUOCA, BRASIL
Tabela 2. Comparação de Machaerium aiuruoquense sp. nov. com espécies atuais e fósseis de M. hirtum, M. nyctitans e M. villosum. Referências: 1, Sartori & Tozzi (1998); 2, Duarte & Rezende-Martins (1983); 3, Fittipaldi (1990); 4, neste trabalho.
Table 2. Comparison of Machaerium aiuruoquense sp. nov. with current and fossil species: M. hirtum, M. nyctitans and M. villosum. References: 1, Sartori & Tozzi (1998); 2, Duarte & Rezende-Martins (1983); 3, Fittipaldi (1990); 4, this study.
Espécies atuais e fósseis
Tamanho foliar (mm)
Proporção foliar Forma foliolar Forma basal Forma apical Face adaxial Face abaxial Venação
Secundária
M. scleroxylon (1)
20-28 x
6-10
2,8:1 -
3,5:1oblonga arredondada
simétricaretuso
mucronadoesparso-
pubescente vilosa broquidódroma
M. nyctitans (1)
20-34 x
6-10
2,8: 1-
3,5:1oblonga
arredon dada ou subcordada
pouco assimétrica
arredondada e retuso,
mucronada
pubérula a glabrescente
serícea no limbo e
tomentosa na veia mediana
broquidódroma
M. hirtum (1)
8-22 x
2-6
3,5:1 -
4:1
oblongo-estreita convexa obtuso, retuso
e mucronado - serícea craspedódroma
M. villosum (1)
56-94 x
17-22
3:1 –
4:1
lanceolada ou obovada arredondada agudo esparso-vilosa
canescente-vilosa
principalmente sobre a nervura
principal
broquidódroma
M. nervosum(2)
21x
7,82,7:1 oblonga
simétricaarredondada
simétrica retuso - - broquidódromasinuosa
M. piranii (3)
41-44x
23-241,8:1 elíptica
simétricaarredondada
simétrica emarginada base de pelos base de pelosbroquidódroma
de curva abrupta
M. aiuroquensesp. nov.(4)
23-43 x
10-14
2,3:1-
3:1oblonga
arredondada pouco
assimétricaretuso base de pelos base de pelos broquidódroma
fraca, arqueada
Comparam-se a Machaerium nyctitans na forma foliolar, no ápice, na base, margem fi mbriada e proporção foliolar (2,8:1), mas diferem na venação intersecundária menos pronunciada, com menor número de pares de veias secundárias e no tamanho visivelmente menor.
Dentre essas espécies atuais aquelas com maior afi nidade morfológica são: Machaerium nyctitans, (Figura 5F, lâmina UnG-44) e M. scleroxylon, embora ambas apresentem também algumas feições distintas.
Por se tratarem de seis folíolos desconectados e, por conseguinte sem possibilidade de verifi car sua disposição na folha, prefere-se mantê-los em espécie fóssil distinta, designada como Machaerium aiuruoquense sp. nov.Distribuição geográfi ca e ecologia das espécies atuais afi ns. No Brasil, o gênero é encontrado, atualmente, na Floresta Pluvial Atlântica e no Cerrado desde o Piauí e Ceará até o Paraná (Lorenzi, 2002).
A espécie afi m atual Machaerium nyctitans, de nome popular jacarandá-bico-de-pato, restringe sua ocorrência desde o sul dos estados da Bahia, Espírito Santo, Rio de Janeiro, São Paulo e Minas Gerais até o Rio Grande do Sul em fl orestas ombrófi las densa e mista (Mata Atlântica e de Araucária), em comunidade mesófi la semidecídua, restingas e caatingas. Ocorre também na Argentina, na área de San Pedro e San Tomé (Mendonça-Filho et al., 2007).
A outra espécie afi m, Machaerium scleroxylon, de nome popular caviúna, ocorre na região dos estados da Bahia, Goiás,
Minas Gerais, São Paulo e Paraná, bem como na Bolívia e Paraguai. Trata-se de elemento da fl oresta mesófi la, fl oresta costeira, mata seca, cerradão, em regiões calcáreas, e em áreas periodicamente inundáveis, com solo argiloso ou arenoso (Mendonça-Filho et al., 2007).Distribuição estratigráfi ca sulamericana. O documentário do gênero Machaerium tem sido feito a partir do Eoceno-Oligoceno, no SE do Brasil. Está presente em sedimentos eocenos-oligocenos da bacia de São Paulo (Formação Itaquaquecetuba); nos eocenos-oligocenos da bacia de Aiuruoca e nos oligocenos da bacia de Taubaté (Formação Tremembé).
Machaerium paleogenum sp. nov.(Figura 7)
Holótipo. UnG/3T- 250a,b (Figura 7).Estratigrafi a. Formação Entre-Córregos, bacia de Aiuruoca, Minas Gerais.Idade. Eoceno-Oligoceno.Procedência. Do ritmito constituído por lâminas de folhelho alternadas com lâminas de arenito fi no, argiloso, micáceo, do afl oramento da margem esquerda do córrego Entre-Córregos, entre Aiuruoca e Minduri, Estado de Minas Gerais.Etimologia. paleogenum referente ao período geológico Paleógeno.Diagnose. Folha composta imparipinada (?), com pelo menos quatro pares de folíolos (8-9 folíolos), de disposição oposta

354 REVISTA BRASILEIRA DE PALEONTOLOGIA, 17(3), 2014
a suboposta, de forma oblonga a elíptica larga, de ápice arredondado ou retuso, de base convexa ou arredondada, assimétrica, com peciólulos pulvinados, venação secundária broquidódroma, textura cartácea ou papirácea. Medidas folíolares 42-67 mm de comprimento por 21-28 mm de largura (proporção: 2-2,5:1); os menores situados na base.Diagnosis. Imparipinnate compound leaf (?), with at least four pairs of leafl ets (8-9 leafl ets), opposite to subopposite disposition, oblong to elliptical wide shape, rounded or retuse apex, asymmetrical, rounded or convex base, pulvinate petiolule, secondary brochidodromous venation, chartaceous to papyraceous texture. Leafl ets measures: 42-67 mm of length for 21-28 mm of width (rate: 2-2,5:1); the size increases from the base toward apex. Descrição. Impressão e contra-impressão de fragmento de folha composta, provavelmente, imparipinada, com pelo menos nove folíolos, dispostos em quatro pares opostos a subopostos. Seus folíolos possuem peciólulos pulvinados de 2 a 4 mm de comprimento por 1,5 mm de diâmetro. A fronde tem, aproximadamente, 100 mm de largura máxima com raque de mais de 60 mm de comprimento por 1,5 mm de largura. Cada folíolo tem forma oblonga a elíptica larga, margem inteira (ciliada?), base convexa ou arredondada, assimétrica, ápice arredondado ou retuso. Medidas: 42 a 67 mm de comprimento por 21 a 28 mm de largura, em proporção variável de 2:1 a 2,5:1. A venação do folíolo é pinada, broquidódroma fraca, chegando muito próximo à margem, antes de encurvar-se para cima ao encontro da secundária suprajacente; intersecundárias presentes e textura possivelmente cartácea ou papirácea (Figura 7).Comparação e discussão. Devido à sua preservação em folíolos conectados e forma foliolar oblonga a elíptica larga, com tamanho médio a grande, essa fronde pode ser comparada com as seguintes espécies atuais, consideradas mais similares: Machaerium acutifolium Benth., M. amplum Benth., M. nyctitans, M. oblongifolium Vog. ex Benth. e M. villosum (espécimes clarifi cados, lâminas nº UnG-95 e UnG-136) conforme apresentado em Sartori & Tozzi (1998), Lorenzi (2002) e Mendonça-Filho et al. (2007).
Fazendo um levantamento das características morfográfi cas da forma estudada e dessas espécies atuais, chega-se à Tabela 3, onde duas espécies ressaltam nas similaridades: pelo número de pares de folíolos (~ 4), forma oblonga a elíptica e tamanho médio foliolar, disposição oposta a suboposta, o ápice retuso, a forma de base arredondada assimétrica. Estas são: M. amplum e M. oblongifolium. As características apontadas para ambas as espécies em Sartori & Tozzi (1998) foram utilizadas na comparação (Tabela 3).
A forma de Aiuruoca distingue-se de Machaerium amplum por apresentar tamanho ligeiramente maior, ápice arredondado, base assimétrica e venação broquidódroma em vez de craspedódroma. Distingue-se de M. oblongifolium por apresentar maior número de pares de folíolos, predominantemente, opostos a subopostos, de forma oblonga ou elíptica larga, ápice arredondado ou retuso, enquanto M. oblongifolium tende mais para folíolos alternos, de forma oblonga a obovada e ápice convexo ou retuso.
Consideradas essas diferenças morfológicas além da idade, julga-se conveniente, ainda que com base em apenas um espécime, dar-lhe novo epíteto específi co: Machaerium paleogenum sp. nov. Distribuição geográfi ca e ecologia da espécie atual afi m. A espécie afi m atual Machaerium oblongifolium é uma planta escandente ou árvore de cerca de 6 m de altura, típica de mata de restinga e de encosta, que ocorre nos estados de Minas Gerais, Rio de Janeiro e, em São Paulo, desde o litoral até o noroeste.
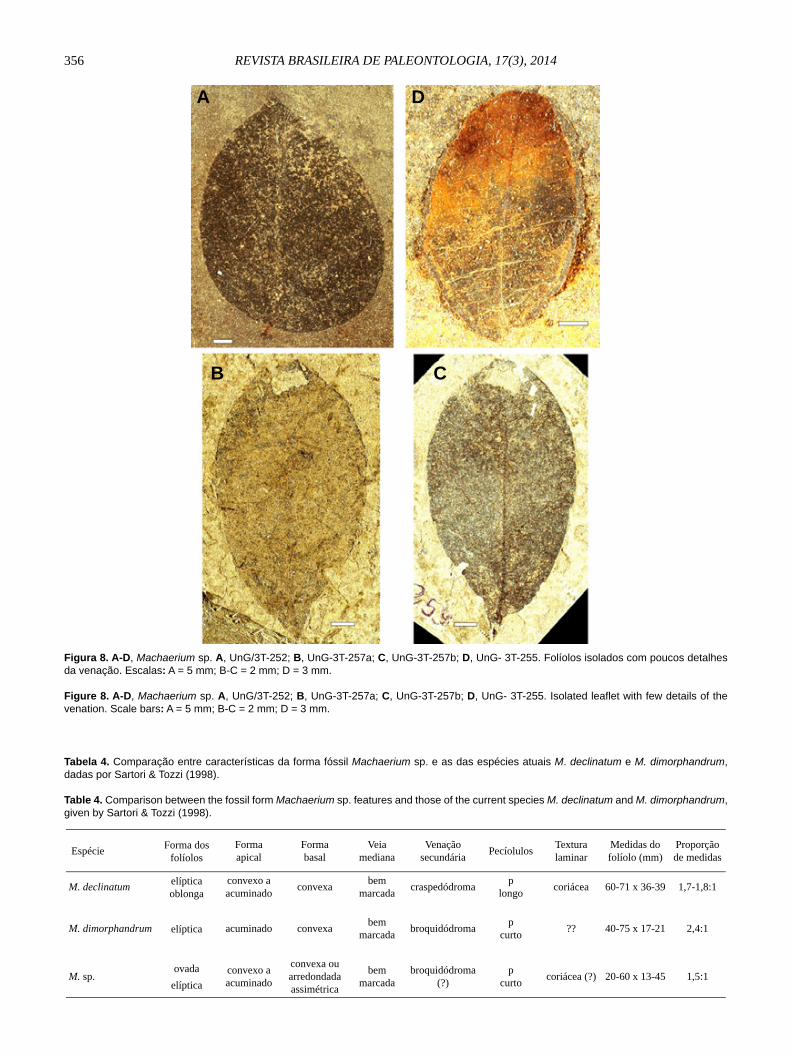
Machaerium sp.(Figuras 8A-D)
Espécimes estudados. UnG/3T- 252 a UnG/3T- 258.Estratigrafi a. Formação Entre-Córregos, bacia de Aiuruoca, Minas Gerais.Idade. Eoceno-Oligoceno.Procedência. Do ritmito constituído por lâminas de folhelho alternadas com lâminas de arenito fi no, argiloso, micáceo, do afl oramento da margem esquerda do córrego Entre-Córregos, entre Aiuruoca e Minduri, Estado de Minas Gerais.Descrição. Seis folíolos assimétricos, de forma ovada a elíptica, de ápice convexo a acuminado, base convexa ou arredondada larga, fortemente assimétrica, veia mediana bem marcada, venação secundária sugere ser broquidódroma, peciólulos pulvinados curtos (medindo 2-4 mm de comprimento por 1-3 mm de largura). Limbo é sugestivo de textura coriácea, não permitindo visualizar detalhes da venação secundária
Figura 7. Machaerium paleogenum sp. nov., UnG/3T 250a,b, holótipo, folha composta imparipinada. Escala = 10 mm.
Figure 7. Machaerium paleogenum sp. nov., UnG/3T 250a,b, holotype, imparipinnate compound leaf. Scale bar = 10 mm.
A

355BERNARDES DE OLIVEIRA ET AL. – FABÁCEAS PALEÓGENAS, BACIA DE AIURUOCA, BRASIL
e de ordem mais alta. Medidas do folíolo: 20-60 mm de comprimento por 13-45 mm de largura, numa proporção de 1,5:1 (Figura 8).Comparação. Folíolos isolados que se assemelham, por seu aspecto muito assimétrico e tamanho variado, embora de proporção foliolar constante, com os folíolos de duas espécies de Machaerium: M. declinatum (Vell.) Stellfeld e M. dimorphandrum Hoehne. A espécie M. declinatum, conforme apresentada por Sartori & Tozzi (1998), caracteriza-se por apresentar forma foliolar elíptico-larga, com ápice convexo a acuminado e base convexa, veia mediana bem marcada e secundária craspedódroma, peciólulos pulvinados longos, textura coriácea, medida foliolar de 60-71 mm x 36-39 mm, numa proporção de 1,7-1,8:1. Os espécimes fósseis distinguem-se dos daquela espécie atual pela forma foliolar mais ovada elíptica, forma basal mais arredondada assimétrica, venação possivelmente broquidódroma, peciólulos pulvinados curtos, medidas e proporções foliolares menores (Tabela 4).
Machaerium dimorphandrum, segundo Sartori & Tozzi (1998), caracteriza-se por apresentar folíolos sempre elípticos de ápices acuminados e base convexa, com veia mediana bem marcada, venação secundária broquidódroma, peciólulo pulvinado curto e medidas de 40-75 mm x 17-32 mm, numa proporção aproximada de 2,4:1. Comparando os espécimes fósseis com essa espécie, verifica-se que se distinguem
dela pela forma foliolar mais ovada-elíptica, ápice convexo a acuminado, base convexa a arredondada assimétrica e dimensões e proporções foliolares menores. Pelas feições distintas dessas duas espécies que lhe são ainda as mais semelhantes e, considerando tratar-se de folíolos isolados, onde não foi possível um exame e comparação mais acurados da venação, é recomendável mantê-los como Machaerium sp..Distribuição geográfi ca e ecologia da espécie atual afi m. Machaerium dimorphandrum é uma planta escandente, às vezes, com gavinhas aculeadas, ramos glabros, lenticelados. Ocorre em mata de encosta. Presença constatada na grande São Paulo, litoral norte e entre os limites geográfi cos dos estados do Rio de Janeiro e São Paulo. Aparecem ainda, na Bahia, Minas Gerais e Paraná (Sartori & Tozzi, 1998).
SISTEMÁTICA PALINOLÓGICA
A análise dos palinomorfos revelou grande diversidade e quantidade de todos os grupos, desde fungos a angiospermas, neste último as leguminosas apresentam-se com elevada frequência, conforme Garcia et al. (2000). Além da literatura paleo e actuopalinológica, foram consultadas actuopalinotecas de referência, no intuito de verifi car a morfologia dos grãos de pólen correspondentes aos táxons paleobotânicos (Caesalpinia e Machaerium) reconhecidos por seus aspectos foliolares.
Tabela 3. Comparação entre Machaerium paleogenum sp. nov. e algumas espécies afins atuais e fósseis de Machaerium. Referências: 1, Sartori & Tozzi (1998); 2, Duarte & Rezende-Martins (1983); 3, Fittipaldi (1990); 4, neste trabalho.
Table 3. Comparison among Machaerium paleogenum sp. nov. and some affinis current and fossil species of Machaerium. Referências: 1, Sartori & Tozzi (1998); 2, Duarte & Rezende-Martins (1983); 3, Fittipaldi (1990); 4, this study.
Espécies atuais e fósseis
Nº de pares de folíolos
Tamanho foliar (mm)
Proporção foliar
Disp. de folíolos na raque Forma foliolar Forma basal Forma apical Venação 2
M. acutifolium (1) +5 45-90 x 15-30 3:1 alterna/
suboposta
ovado-lanceolada/ oblongo-
lanceolada
arredondada ou raro cordada
reto/acuminado
broquidódroma encurvada
M. amplum(1) 4-7 27-45 x 11-17 2,5:1 oposta/
suboposta oblonga/ eliptico arredondada convexo e retuso craspedódroma
M. nyctitans(1) 3-4 20-34 x 6-10 3,5:1 alterna oblonga
arredondada ou subcordada
pouco assimétrica
arredondada e retuso,
mucronadabroquidódroma
M. oblongifolium (2) 2-3 36-58 x 20-30
>terminais 2:1 alterna/ suboposta
oblonga/ obovada
arredondadasimétrica
obtusoretuso broquidódroma
M. villosum(3) 5-12 56-94 x 17-22 3:1 x 4,5:1 alterna/
suboposta
oblonga-lanceolada ou
obovadaarredondada reta broquidódroma
M. paleogenumsp. nov.(4)
+4 42-67 x 21-28>terminais 2:1 x 2,5:1 oposta/suboposta oblonga-
elíptica largaarredondada assimétrica
Arredondada ou retusa broquidódroma
356 REVISTA BRASILEIRA DE PALEONTOLOGIA, 17(3), 2014
Figura 8. A-D, Machaerium sp. A, UnG/3T-252; B, UnG-3T-257a; C, UnG-3T-257b; D, UnG- 3T-255. Folíolos isolados com poucos detalhes da venação. Escalas: A = 5 mm; B-C = 2 mm; D = 3 mm.
Figure 8. A-D, Machaerium sp. A, UnG/3T-252; B, UnG-3T-257a; C, UnG-3T-257b; D, UnG- 3T-255. Isolated leaflet with few details of the venation. Scale bars: A = 5 mm; B-C = 2 mm; D = 3 mm.
Tabela 4. Comparação entre características da forma fóssil Machaerium sp. e as das espécies atuais M. declinatum e M. dimorphandrum, dadas por Sartori & Tozzi (1998).
Table 4. Comparison between the fossil form Machaerium sp. features and those of the current species M. declinatum and M. dimorphandrum, given by Sartori & Tozzi (1998).
Espécie Forma dos folíolos
Forma apical
Forma basal
Veia mediana
Venação secundária Pecíolulos Textura
laminarMedidas do folíolo (mm)
Proporção de medidas
M. declinatum elíptica oblonga
convexo a acuminado convexa bem
marcada craspedódroma plongo coriácea 60-71 x 36-39 1,7-1,8:1
M. dimorphandrum elíptica acuminado convexa bem marcada broquidódroma p
curto ?? 40-75 x 17-21 2,4:1
M. sp.ovada
elípticaconvexo a acuminado
convexa ou arredondada assimétrica
bem marcada
broquidódroma (?)
p curto coriácea (?) 20-60 x 13-45 1,5:1
A
B C
D

357BERNARDES DE OLIVEIRA ET AL. – FABÁCEAS PALEÓGENAS, BACIA DE AIURUOCA, BRASIL
Classe TRICOLPORATAE Iversen & Troels-Smith, 1950
Margocolporites Ramanujam, 1966
Espécie-tipo. Margocolporites tsudakai Ramanujam, 1966.
Margocolporites vanwijhei Germeraad, Hopping & Muller, 1968
(Figura 9A)
Descrição. Grãos de pólen mônades, simetria radial, isopolares, oblatos-esferoidais, âmbito circular, 3-brevicolporados. Endoabertura lalongada. Exina reticulada, retículos menores nos pólos, na porção externa das margens dos colpos, e intectada na porção interna. Muros columelados, margens fi namente baculadas, bordas retas e pontiagudas, sem conexão nos pólos. Apocolpo e mesocolpos delimitados pela sombra de um degrau, devido à mudança brusca na redução da espessura da sexina, que aumenta no contorno do colpo, formando uma suave depressão com margem microrreticulada.Dimensões. Diâmetro polar: 37-42 μm; espessura da exina 1,3-2,1 μm; intercolpos 8,3-10,5 μm; diâmetro do lúmen 0,4-1,8 μm; profundidade dos colpos 8-11 μm. Comparação. A descrição concorda com a de Corrêa (2003), onde os grãos de pólen recentes de Caesalpinia echinata foram descritos como médios a grandes, suboblatos a oblato-esferoidais, âmbito circular, 3-brevicolporados, colpo estreito, nem sempre visível, com margem bem larga, microrreticulada, formada por um afi namento da sexina nessa região, endoabertura lalongada, exina reticulada e heterorreticulada, com retículos menores nos pólos; muros simples columelados.
Portanto, Margocolporites vanwijhei, considerado afi m ao gênero Caesalpinia, ocorre com representação regular, nos sedimentos da Formação Entre-Córregos. Sua presença ali, além de corroborar no registro do gênero Caesalpinia, também é sugestiva de idade eocena/oligocena.
Germeraad et al. (1968) apresentaram esta espécie para o Caribe (Venezuela, Colômbia e Trinidad) do Eoceno médio ao Pleistoceno; para a Nigéria e Bornéo, do Eoceno médio ao Pleistoceno, e sugeriram afi nidade com os gêneros atuais Adipera, Brasilettia, Caesalpinia,
Haematoxylon, Mezoneuron, Poincianella e, especial-mente, com as espécies Caesalpinia bonduc (L.) Roxb. e C. coriaria (Jacq.) Willd.
No presente trabalho, comparou-se Margocolporites com grãos de pólen de diversas espécies de Caesalpinia que apresentam morfologia similar. A comparação do grão de pólen de C. echinata, com os de outras espécies do gênero, já foi realizada por diversos autores, como sumarizado em Corrêa (2003). Os grãos de pólen daquela espécie apresentam aberturas complexas e foram interpretados de diversas maneiras: 3-aperturados (Erdtman, 1952); 3-colporados margeados por uma região mais fi na da sexina-pseudocolpo (Nair & Sharma, 1962); 3-margocolporados (Tsukada, 1963); e 3 aperturados compostos por um colpo curto e rodeado por margens complexas (Senesse, 1988).
Os morfotipos afi ns a Margocolporites apresentam uma distribuição geográfi ca neotropical muito ampla, como pode ser visto na Tabela 5.
No Brasil, o morfotipo fóssil Margocolporites vanwijhei apresenta distribuição temporal a partir do Eoceno superior (Regali et al., 1974a,b; Lima & Amador, 1985; Maizatto, 2001; Garcia et al., 2008; Santos et al., 2010).
Muller et al. (1987), com base em estudos palinológicos desenvolvidos em sedimentos do Cretáceo ao recente no Brasil, Colômbia, Guiana, Suriname, Trinidad e Venezuela, demonstraram que o primeiro registro desta espécie no reino Neotropical ocorreu no Eoceno médio da Venezuela. Na Colômbia, Jaramillo & Dilcher (2001) mostraram que Margocolporites vanwijhei ocorre no Eoceno inferior, base da Formação Mirador.
Para o Eoceno superior da Nigéria, grãos de pólen de reticulado grosseiro, como o de Margocolporites, foram descritos como Margocolporites foveolatus e M. umuahiaensis por Jan du Chêne et al. (1978).
Na Nova Zelândia, Pocknall (1989) assinalou a distribuição de M. vanwijhei do Eoceno superior ao Mioceno inferior. Na Índia, a ocorrência mais antiga é Paleocaesalpiniaceaepites eocenica descrita por Baksi (1962) e Margocolporites complexum descrito por Sah & Dutta (1966), ambos do Eoceno inferior de Assam. Com base nesses dados, Muller (1981) ressaltou que os grãos de pólen do tipo Caesalpinia são encontrados desde o Eoeoceno e apontou a Índia como o berço das Caesalpinoideae por apresentar as ocorrências mais antigas, até então, e de maior diversidade de grãos de pólen do tipo Caesalpinia.
Marchiori (1997) constatou que as cesalpinoídeas são abundantes no reino Paleotropical, principalmente, na África. No Brasil, são mais numerosas no centro-norte e mais escassas ao sul.
A distribuição dessa subfamília sugere que, no Neocretáceo, época de seu surgimento, a dispersão entre a África e América do Sul era recíproca e muito mais fácil devido à menor distância entre os dois continentes (principalmente entre a região nordestina brasileira e a região ocidental africana), aliada às numerosas ilhas vulcânicas que existiam entre os
Tabela 5. Distribuição geográfica das espécies atuais que apresentam morfotipos polínicos similares aos da espécie Margocolporites vanwijhei.
Table 5. Geographic distribution of the current species bearing similar pollen morphotypes to those of the species Margocolporites vanwijhei.
ESPÉCIES DISTRIBUIÇÃO GEOGRÁFICA
Caesalpinia bonduc sul da América do Norte, Antilhas e Hawaí
C. pulcherrima centro-americana
C. coriaria Caribe, México, América Central e Norte da América do Sul
C. gilliesii Uruguai e Argentina
C. paraguariensis Mato Grosso (Brasil), Paraguai, Argentina
C. echinata no Brasil, do Rio Grande do Norte ao Rio de Janeiro.

358 REVISTA BRASILEIRA DE PALEONTOLOGIA, 17(3), 2014
mesmos. A especiação das Caesalpinoideae sulamericanas e africanas ter-se-ia processado com o paulatino distanciamento geográfi co entre os dois continentes, após o Paleoceno. Entre a África e o sudeste asiático ocorreram ligações similares até o fi nal do Neógeno, quando uma crescente aridez e distanciamento geográfi co sobrevieram.
A presença do grão de pólen Margocolporites vanwijhei Germeraad, Hopping & Muller, 1968 (Figura 9A), detectada nas análises palinológicas (Garcia et al., 2000 e neste trabalho) dos sedimentos da Formação Entre-Córregos, em associação com macrofósseis do gênero Caesalpinia corrobora sua afi nidade botânica.
Psilatricolporites (Van der Hammen, 1956) Pirce, 1961
Espécie-tipo. Psilatricolporites inornatus (Van der Hammen, 1956) Pirce, 1961.
Psilatricolporites sp.(Figura 9B)
Figura 9. A-B, grãos de pólen; A, Margocolporites vanwijhei; B, Psilatricolporites sp. Escalas = 20 μm.
Figure 9. A-B, pollen grains; A, Margocolporites vanwijhei; B, Psilatricolporites sp. Scale bars = 20 μm.
Descrição. Grãos de pólen mônades, pequenos, subprolados a esferoidais, tricolporados, com exina espessa e superfície psilada. Dimensões. Diâmetro polar: +/- 22 μm; diâmetro equatorial: 20 μm. Discussão. Os grãos de Psilatricolporites estão bem representados nos sedimentos da Formação Entre-Córregos; foram observados e comparados com grãos de diversas espécies de Machaerium em actuopalinotecas de referência, dentre as quais se destacam M. aculeatum Raddi, M. paraguaiense Hassl., M. stipitatum (DC.) Vogel. No entanto, estas espécies variam na espessura da exina, na presença/ausência de microrretículo na superfície e de colpos operculados, neste último caso, se a exina for muito fi na, podem ou não ser preservados, características essas já referidas por Barth (1964).
CONSIDERAÇÕES FINAIS
Um bioma de diferentes estratos, com plantas heliófi tas a esciófi tas, semelhante ao da Mata Atlântica atual, isto é, de mata tropical pluvial de encosta, delineia-se quando: as respectivas espécies atuais afi ns dos táxons de fabáceas detectados na paleofl ora de Entre-Córregos são consideradas quanto aos seus hábitos, habitats, condições climáticas e biomas, conforme dados de Sartori & Tozzi (1998), Lorenzi (2002) e Mendonça-Filho et al. (2007) (Tabela 6).
As Fábaceas registradas nessa tafofl ora são afi ns a espécies de árvores de 5 a 30 m de altura e arbustos escandentes (menores de 2 m até 6 m de altura) sugerindo pelo menos dois estratos. Considerando-se que Caesalpinia echinata, Machaerium dimorphandrum, M. oblongifolium e M. nyctitans são as espécies mais semelhantes ou afi ns, suas características sugerem que havia fl orestas, do tipo encosta ou fl oresta costeira Pluvial Atlântica, na área de Aiuruoca. Interessante é notar que essas características também foram sugeridas por Franco-Delgado & Bernardes-de-Oliveira (2004), com base em anonáceas (Annona sp.) e lauráceas (Nectandra sp.) identifi cadas na mesma tafofl ora de Entre-Córregos. Deve-se ressaltar que esse tipo de vegetação corresponderia a um bioma de clima mais quente e úmido típico para o Paleógeno, ocorrendo na região sudeste mineira, provavelmente, quando ainda não existia a barreira das serras costeiras.
Considerando a possibilidade de: (i) o paleoclima eoceno-oligoceno ser mais quente e mais úmido que o atual; (ii) o tectonismo formador da bacia de Aiuruoca, do Rift Continental do Sudeste e do relevo da Serra do Mar e da Serra da Mantiqueira estar em franco desenvolvimento, permitindo que a umidade marinha adentrasse mais o continente, até a área do alto rio Grande, em Minas Gerais; as condições paleoclimáticas sugeridas por essas fabáceas e outros elementos da paleofl ora Aiuruoca, o ambiente deposicional e as evidências litológicas poderiam ser confi rmados.
Muitas outras formas de fabáceas, dentre as quais mimosoídeas (evidenciadas por seus grãos de pólen e prováveis macrofósseis), parecem estar presentes na tafofl ora
A
B

359BERNARDES DE OLIVEIRA ET AL. – FABÁCEAS PALEÓGENAS, BACIA DE AIURUOCA, BRASIL
ora em análise, aguardando identifi cação e denunciando para o Paleógeno regional uma provável precoce presença das três subfamílias: Caesalpinoideae, Mimosoideae e Faboideae.
Acredita-se que a família das Fabaceae (Leguminosae) tenha evoluído durante o Neocretáceo, com base em registros fósseis das três subfamílias, conforme Polhill & Raven (1981). O documentário mais antigo conhecido para fabáceas foi registrado na forma de lenho fóssil por Giraud & Lejal-Nicol (1989) sob a designação Cassinium dongolense, do Nubiano (Cretáceo médio a superior) do norte do Sudão. A partir do Eoceno, o documentário fossilífero fabáceo torna-se comum na forma de lenhos, folhas, folíolos, fl ores, frutos e grãos de pólen em muitas regiões do globo: Formação Claiborne da América do Norte (Eoceno), registrado por Herendeen & Dilcher (1990). Calvillo-Canadell & Cevallos-Ferriz (2002, 2005) têm registrado numerosas fabáceas do Eoceno e Oligoceno do México.
Wikströim et al. (2001) estimaram uma idade de 74 a 70 Ma, para o aparecimento das fabáceas, com base na análise de genes. Formas duvidosas, atribuídas à família, têm sido registradas desde o Maastrichtiano, no Canadá, Colúmbia e Sibéria como grãos de pólen (Sindora-like) e de lenho (Cassia) da subfamília Caesalpinoideae por Polhill & Raven (1981). Há ainda registro, razoavelmente, seguro mencionado, mas ainda não descrito, de fósseis de leguminosas no Neopaleoceno (~ 56 Ma) por Wing et al. (2009) para a Formação Cerrejón
Tabela 6. Hábitos e habitats de espécies atuais afins às espécies fósseis de fabáceas da ta foflora de Aiuruoca, Minas Gerais. Referências e abreviações: 1, Lorenzi (2002); 2, Mendonça Filho et al. (2007); 3, Sartori & Tozzi (1998); 4, Mendonça Filho et al. (2007); 5, Polido & Sartori (2007); AM, Amazonas; BA, Bahia; CE, Ceará; DF, Distrito Federal; GO, Goiás; MA, Maranhão; MG, Minas Gerais; MS, Mato Grosso do Sul; MT, Mato Grosso; PR, Paraná; RJ, Rio de Janeiro; RR, Roraima; RS, Rio Grande do Sul; SP, São Paulo.
Table 6. Habits and habitats of current species affinis to the Fabaceae fossil species from the Aiuruoca taphoflora, Minas Gerais. References and abbreviations: 1, Lorenzi (2002); 2, Mendonça Filho et al. (2007); 3, Sartori & Tozzi (1998); 4, Mendonça Filho et al. (2007); 5, Polido & Sartori (2007); AM, Amazonas; BA, Bahia; CE, Ceará; DF, Distrito Federal; GO, Goiás; MA, Maranhão; MG, Minas Gerais; MS, Mato Grosso do Sul; MT, Mato Grosso; PR, Paraná; RJ, Rio de Janeiro; RR, Roraima; RS, Rio Grande do Sul; SP, São Paulo.
Táxons da tafoflora de Aiuruoca Táxons atuais afins Hábito Habitat Localização geográfica
Caesalpinia veraechinataformissp.nov.
C. echinata(1)
Árvore de 8 a 30 m de alt., acúleos cônicos, semi-decídua, helióficta
ou esciófita.
Endêmica da Floresta pluvial Atlântica, terrenos secos, Floresta primária densa
CE até RS. Frequente no sul da BA
Machaerium aiuruoquensesp. nov.
M. scleroxylon(2) (3)
Árvore de 6 a 30 m de alt. acúleos lineares a triangulares nos rebrotos
Floresta mesófila, semidecídua, Mata costeira, seca, cerradão em regiões calcárias, em áreas periodicamente
inundáveis, em solo arenoso ou argiloso
Bolívia, Paraguai, BA, GO, MG, PR, SP
M. nyctitans(2) (3)
Árvore de 5 a 25 m de alt., tronco reto 6-57 cm de diâm., aculeado, ramos com acúleos piramidais
Florestas costeiras, Mata de Araucária, Mata mesófila semi-decídua
Argentina, BA, MG, RJ, PR, SP
Machaeriumpaleogenumsp. nov.
M. amplum(2) (4)
Arbusto escandente, até 2 m alt., caules e ramos sulcados, espinhos
recurvados na base foliar e da inflore scência
Exclusivamente na Savana (Cerrado), borda de Mata mesófila semidecídua
AM, CE, DF, GO, MA, MG, SP, RJ, RO, MT, MS
M. oblongifolium(3)
Arbusto escandente ou árvore de 6 m com acúleos
Mata de restinga e Mata de encosta, predominantemente, no litoral SP, MG, RJ
Machaerium sp.M. dimorphandrum(2) (3)
Arbusto escandente com gavinhas aculeadas, ramos glabros,
lenticeladosMata de encosta
Grande SP, litoral norte SP, limites SP/RJ, BA, MG, PR
(Península Guajiva, NE de Colômbia) e por Herendeen & Wing (2001) para o NW de Wyoming (EUA) e Mimosoideae, no Eopaleoceno da Patagônia (Formação Salamanca) por Brea et al. (2008).
De fato, a ocorrência de diversas assembléias contendo táxons das três subfamílias em muitas localidades datadas do Eoceno médio a superior sugere que uma diversifi cação intensa do grupo ocorreu nesse tempo. Na América do Sul e no Brasil, em particular, há registro das três subfamílias desde esse tempo, nas diferentes bacias do sudeste, confi rmando a diversifi cação paleógena intensa do grupo observada por Herendeen et al. (1992).
O registro aqui feito na Formação Entre-Córregos, na forma de grãos de pólen e de folíolos destacados e em conexão foliar, vem confi rmar esse cenário. Infelizmente, permanece incógnita a área berço das Fabáceas.
AGRADECIMENTOS
As autoras desejam externar seus agradecimentos a I.A.S. Mendes pelo apoio dado nas atividades de informática, revisão e versão para o inglês. A E.M. Melo, pelo apoio na obtenção de fotomacrografi as e na montagem de estampas, a A. Hoelzel pela confecção de esquemas e mapas, bem como a F.C. Casado, pela confecção de fotomicrografi as. Ao CNPq, pelas bolsas de produtividade em pesquisa, processo 304978/2013-2

360 REVISTA BRASILEIRA DE PALEONTOLOGIA, 17(3), 2014
a M.E.C.B-O. e processo 306609/2012-6 a M.J.G. A terceira autora agradece também à Fundação de Amparo à Pesquisa do Estado de São Paulo/FAPESP, que lhe concedeu bolsa de mestrado através do processo 2008/02234-0, possibilitando-lhe o desenvolvimento de pesquisa sobre o tema desse manuscrito. As autoras ainda agradecem, especialmente, às revisoras D. Pons da Université Pierre et Marie Curie, Paris, França, e L.M. Anzótegui da Universidad Nacional de Nordeste, Corrientes, Argentina, pelas cuidadosas revisões e sugestões construtivas que só vieram a aprimorar o texto.
REFERÊNCIAS
APG III (The Angiosperm Phylogeny Group III). 2009. An update of the Angiosperm Phylogeny Group classifi cation for the orders and families of fl owering plants: APG III. Botanical Journal of the Linnean Society, 161:105-121. doi:10.1111/j.1095-8339.2009.00996.x
Aubry, M.-P.; Blondeau, A.; Cavelier, C.; Damotte, R.; Gruas-Cavagnetto, C.L.; Calvez, Y.; Odin, G.; Pomerol, C.; Renard, M.; Riveline, J. & Tourenq, J. 1977. Le Paléogène dans le sondage du Mont Pagnotte, AGBP, 14:3-50.
Baksi, S.K. 1962. Palynological investigation of Simsang River Tertiaries, South Shillong Front, Assam. Bulletin of the Geological, Mining and Metallurgical Society of India, 26:1-22.
Barth, O.M. 1964. Catálogo sistemático dos pólens das plantas arbóreas do Brasil meridional: V- Leguminosae, Papilionatae. Memórias do Instituto Oswaldo Cruz, 62:95-123.
Bastos, M.N.C. 1987. Contribuição ao estudo sistemático de algumas espécies do gênero Machaerium Persoon (Leguminosae-Papilionoideae) ocorrentes na Amazônia brasileira. Boletim do Museu Paraense Emílio Goeldi, Serie Botânica, 3:182-278.
Bedani, E.F. 2002. Ocorrência de anfíbios (Anuros) na Bacia Terciária de Aiuruoca, Minas Gerais, Brasil. Centro de Pós-Graduação, Universidade de Guarulhos, Dissertação de Mestrado, 77 p.
Berry, E.W. 1935. Tertiary plants from Brazil. Proceedings of the American Philosophical Society, 75:565-590.
Brea, M.; Zamuner, A.; Matheos, S.; Iglesias, A. & Zucol, A. 2008. Fossil wood of the Mimosoideae from the early Paleocene of Patagonia. Alcheringa, 32:427-441. doi:10.1080/03115510802417695
Calvillo-Canadell, L. & Cevallos-Ferriz, S.R.S. 2002. Bauhcis moranii gen. et sp. nov. (Cercideae, Caesalpinieae), an Oligocene plant from Tepexi de Rodríguez, Puebla, Mex., with leaf architecture similar to Bauhinia and Cercis. Review of Palaeobotany and Palynology, 122:171-184. doi:10.1016/s0034-6667(02)00135-5
Calvillo-Canadell, L. & Cevallos-Ferriz, S.R.S. 2005. Diverse assemblage of Eocene and Oligocene Leguminosae from Mexico. International Journal of Plant Sciences, 166:671-692. doi:10.1086/430096
Corrêa, A.M.S. 2003. Morfologia polínica de Caesalpinia echinata Lam (Leguminosae-Caesalpinioideae). Revista Brasileira de Botânica, 26:355-359.
De Candolle, A.P. 1825. Leguminosae. In: A.P. de Candolle (ed.) Prodromus systematis naturalis regni vegetabili, Treuttel & Würtz, p. 93-423.
Duarte, L. & Mandarim-de-Lacerda, A.F. 1989. Flora cenozóica do Brasil: Formação Tremembé, Bacia de Taubaté, São Paulo. III Frutos (Phytolaccaceae e Leguminosae). In: CONGRESSO
BRASILEIRO DE PALEONTOLOGIA, 11, 1989. Boletim de Resumos, Curitiba, p. 395-410.
Duarte, L. & Rezende-Martins, A.F. 1983. Contribuição ao conhecimento da fl ora cenozóica do Brasil: jazigo Vargem Grande do Sul, SP. Série Taubaté. Anais da Academia Brasileira de Ciências, 55:109-121.
Dutra, T. & Stranz, A. 2002. Clarifi cação e diafanização de folhas. In: T.L. Dutra (ed.) Técnicas e procedimentos de trabalho com fósseis e formas modernas comparativas, Unisinos, p. 27-29.
Ellis, B.; Daly, D.C.; Hickey, L.J.; Johnson, K.R.; Mitchell, J.D.; Wilf, P. & Wing, S.L. 2009. Manual of Leaf Architecture. Ithaca, Cornell University Press, 190 p.
Erdtman, 1952. Pollen morphology and plant taxonomy - Angiosperms. Stockholm, Almqvist & Wiksell, 539 p.
Fanton, J.C.; Ricardi-Branco, F.; García, R.J.F. & Moreira, A.S. 2009. Arquitectura y anatomia foliar de Leguminosae eocénica, Cuenca de Fonseca, sureste del Brasil: In: SIMPOSIO ARGENTINO DE PALEOBOTÁNICA Y PALINOLOGÍA, 14, 2009. Libro de Resúmenes, Mar del Plata, p. 20.
Ferrusquía-Villafranca, I. & González-Guzmán, L.I. 2005. Northern Mexico’s Landscapes, Part II: The Biotic Setting across Time. In: J.L. Cartron; G. Ceballos & R.S. Felger (eds.) Biodiversity, Ecosystems, and Conservation in Northern Mexico, Oxford University Press, p. 39-51.
Fittipaldi, F.C. 1990. Vegetais Fósseis da Formação Itaquaquecetuba: Cenozóico, Bacia de São Paulo. Programa de Pós-Graduação em Geologia, Universidade de São Paulo, Tese de Doutorado, 146 p.
Foster, A.S. 1952. Foliar venation in angiosperms from an ontogenetic viewpoint. American Journal of Botany, 39:752-766. doi:10.2307/2438624
Franco-Delgado, S.G. 2002. Tafofl ora Paleógena da Formação Entre-Córregos, Bacia de Aiuruoca, Estado de Minas Gerais, Brasil. Centro de Pós-Graduação, Universidade de Guarulhos, Dissertação de Mestrado, 163 p.
Franco-Delgado, S.G. & Bernardes-De-Oliveira, M.E.C. 2004. Annonaceae e Lauraceae da Formação Entre-Córregos (Paleógeno) na Bacia de Aiuruoca: implicações paleoclimáticas. Revista Brasileira de Paleontologia, 7:117-126. doi:10.4072/rbp.2004.2.04
Garcia, M.J.; Bistrichi, C.A.; Saad, A.R.; Campanha, V.A. & Oliveira, P.E. 2008. Stratigraphy and palaeoenvironments of the Tanque Basin, southeastern Brazil. Revista Brasileira de Paleontologia, 11:147-168. doi:10.4072/rbp.2008.3.02
Garcia, M.J.; Santos, M. & Hasui, Y. 2000. Palinologia da parte afl orante da Formação Entre-Córregos, Bacia de Aiuruoca, Terciário de Minas Gerais, Brasil. Revista Universidade Guarulhos, 5:259.
Germeraad, J.J.; Hopping, G.A.Y. & Muller, J. 1968. Palynology of Tertiary sediments from tropical areas. Review of Palaeobotany and Palynology, 6:189-348. doi:10.1016/0034-6667(68)90051-1
Giraud, B. & Lejal-Nicol, A. 1989. Cassinium dongolense n. sp. bois fossile de Caesalpiniaceae du Nubien du Soudan septentrional. Review of Palaeobotany and Palynology, 59:37-50. doi:10.1016/0034-6667(89)90004-3
Gruas-Cavagnetto, C. 1977. Etude palynologique de l’Eocène du Bassin Anglo-Parisien. Université Pierre et Marie Curie, Thèse d’Etat, 288 p.
Herendeen, P.S. & Crane, P.R. 1992. Early Caesalpinoid fruits from the Paleogene of southern England. In: P.S. Herendeen & D.L. Dilcher (eds.) Advances in Legume Systematics. The Fossil Record, Royal Botanic Gardens, p. 57-68.

361BERNARDES DE OLIVEIRA ET AL. – FABÁCEAS PALEÓGENAS, BACIA DE AIURUOCA, BRASIL
Herendeen, P.S.; Crepet, W.L. & Dilcher, D.L. 1992. The fossil history of the Leguminosae: phylogenetic and biogeographic implications. In: P.S. Herendeen & D.L. Dilcher (eds.) Advances in Legume Systematics. The Fossil Record, Royal Botanic Gardens, p. 303-316.
Herendeen, P.S. & Dilcher D.L. 1990. Dilotropis (Leguminosae, Papilionoideae) from the Middle Eocene of southeastern North America. Systematic Botany, 15:526-533. doi:10.2307/2419152
Herendeen, P.S. & Wing, S. 2001. Papilionoid legumes fruits and leaves from the Paleocene of northwestern Wyoming. In: ANNUAL MEETING OF THE BOTANY, 3, 2001. Abstracts, Albuquerque, Botanical Society of America, p. 65.
Hickey, L.J. 1979. A revised classifi cation of the architecture of dicotyledonous leaves. In: C.R. Metcalfe & L.W. Chalk (eds.) Anatomy of the Dicotyledons, Clarendon Press, p. 25-39.
Hickey, L.J. & Wolfe, J.A. 1975. The bases of angiosperm phylogeny: vegetative morphology. Annals Missouri Botanical Garden, 62:538-589.
Iglesias, A.; Wiff, P.; Johnson, K.R.; Zamuner, A.B.; Cúneo, M.R.; Matheos, S.D.E. & Singer, B.S. 2007. A Paleocene lowland macroflora from Patagonia reveals significantly greater richness than North American analogs. Geology, 35:947-950. doi: 10.1130/G23889A.1
Iversen, J. & Troels-Smith, J. 1950. Pollenmorfologiske defi nitioner og typer. Denmarks Geologiske Undersøgelse, 3:1-52.
Jan Du Chêne, J.; Onyike, M.S. & Sowunmi, M.A. 1978. Some new Eocene pollen of the Ogwashi Asamba Formation, Southeastern, Nigeria. Revista Española de Micropaleontología, 10:285-322.
Jaramillo, C. & Dilcher, D.L. 2001. Middle Paleogene Palynology of Central Colombia, South America: a Study of Pollen and Spores from Tropical Latitudes. Palaeontographica Abteilung B, 285:87-213.
Joly, A.B. 1998. Botânica - Introdução à taxonomia vegetal. 12th ed. São Paulo, Editora Nacional, 777 p.
Lavin, M.; Herendeen, P.S. & Wojciechowski, M.F. 2005. Evolutionary rates analysis of Leguminosae implicates a rapid diversification of lineages during the Tertiary. Systematic Biology, 54:575-594. doi:10.1080/10635150590947131
Lewis, G.P.; Schrire, B.; Mackinder B. & Lock, M. 2005. Legumes of the world. Kew, Royal Botanical Gardens, 577 p.
Lima, M.R. & Amador, E.S. 1985. Análise palinológica dos sedimentos da Formação Resende, Terciário do Estado do Rio de Janeiro, Brasil. In: Coletânea de trabalhos paleontológicos, Brasília, Departamento Nacional de Produção Mineral, p. 371-378 (Série Geológica 27).
Lorenzi, H. 2002. Árvores brasileiras. Manual de identifi cação e cultivo de plantas arbóreas do Brasil. Nova Odessa, Instituto Plantarum de Estudos da Flora, 352 p.
Maizatto, J.R. 2001. Análise bioestratigráfi ca, paleoecológica e sedimentológica das bacias terciárias do Gandarela e Fonseca, Quadrilátero Ferrífero, Minas Gerais, com base nos aspectos palinológicos e sedimentares. Programa de Pós-graduação em Evolução Crustal e Recursos Naturais, Universidade Federal de Ouro Preto, Tese de Doutorado, 333 p.
Marchiori, J.N.C. 1995. Elementos de Dendrologia. Santa Maria, UFSM, 163 p.
Marchiori, J.N.C. 1997. Dendrologia das Angiospermas: Leguminosas. Santa Maria, UFSM, 199 p.
Mello, C.L.; Sant’Anna, L.G. & Bergqvist, L.P. 2002. Fonseca, MG - Vegetais fósseis do Terciário brasileiro. In: C. Schobbenhaus; D.A. Campos; E.T. Queiroz; M. Winge & M.L.C. Berbert-Born (eds.) Sítios Geológicos e Paleontológicos do Brasil,
DNPM/CPRM – Comissão Brasileira de Sítios Geológicos e Paleobiológicos, p. 73-79.
Mendonça-Filho, C.V.; Tozzi, A.M.G.A & Martins, E.R.F. 2007. Revisão taxonômica de Machaerium sect. oblonga (Benth.) Taub. (Leguminosae, Papilionoideae, Dalbergieae). Rodriguésia, 58:283-312.
Muller, J. 1981. Fossil pollen records of extant angiosperms. Botany Review, 47:1-142. doi:10.1007/BF02860537
Muller, J.; Di Giacomo, E. & van Erve, A.W. 1987. A palynological zonation for the Cretaceous, Tertiary, and Quaternary of northern South America. American Association of Stratigraphic Palynologists, 19:1-77.
Nair, P.K.K. & Sharma, M. 1962. Pollen grains of Indian plants - IV Leguminosae (Part-I). Bulletin of the National Botanic Gardens, 65-37.
Persoon, C.H. 1807. Synopsis plantarum seu enchiridium botanicum. Paris, Parisis Lutetiorum, 360 p.
Pocknall, D.T. 1989. Late Eocene to Early Miocene vegetation and climate history of New Zealand. Journal of the Royal Society of New Zealand, 19:1-18.
Polhill, R.M. 1981. Dalbergieae Bronn ex DC (1825). In: R.M. Polhill & P.H. Raven (eds.) Advances in Legume systematics, Royal Botanic Gardens, p. 233-242.
Polhill, R.M. & Raven, P.H. 1981. Advances in Legume Systematics. Knew, Royal Botanic Gardens, 1050 p.
Polido, C.A. & Sartori, A.L.B. 2011. Machaerium (Legumenosae, Papilionoideae, Dalbergieae), nos estados de Mato Grosso e Mato Grosso do Sul, Brasil. Rodriguésia, 62:107-122.
Ramanujam C.G.K. 1966. Palynology of the Miocene lignite from South Arcot District, Madras, India. Pollen et Spores, 8:149-203.
Regali, M.S.P.; Uesugui, N. & Santos, A.S. 1974a. Palinologia dos sedimentos meso-cenozóicos do Brasil. Boletim Técnico Petrobrás, 17:177-191.
Regali, M.S.P.; Uesugui, N. & Santos, A.S. 1974b. Palinologia dos sedimentos meso-cenozóicos do Brasil. Boletim Técnico Petrobrás, 17:263-301.
Rudd, V.E. 1977. The genus Machaerium (Leguminosae) in Mexico. Boletin de la Sociedad Botánica de Mexico, 37:119-146.
Sah, S.C.D & Dutta, S.K. 1966. Palynostratigraphy of the sedimentary formations of Assam: 1. Stratigraphical position of the Cherra Formation. The Palaeobotanist, 15:72-86.
Santos, M. 1999. Serra da Mantiqueira e Planalto do Alto Rio Grande: a bacia terciária de Aiuruoca e evolução morfotectônica. Programa de Pós-Graduação em Geologia Sedimentar, Universidade Estadual Paulista, Tese de Doutorado, 134 p.
Santos, D.B.; Garcia, M.J.; Saad, A.R. & Bistrichi, C.A. 2010. Palinoestratígrafi a da Formação Itaquaquecetuba, Bacia de São Paulo, Brasil. Revista Brasileira de Paleontologia, 13:205-220. doi:10.4072/rbp.2010.3.05
Sartori, A.L.B. & Tozzi, A.M.G.A. 1998. As espécies de Machaerium Pers. (Leguminosae - Papilionoideae - Dalbergieae) ocorrentes no estado de São Paulo. Revista Brasileira de Botânica, 21:211-246.
Senesse, S. 1988. Second type pollinique dans le genre Caesalpinia. Nouvelles donnés sur l’évolution dans la tribu des Caesalpinieae. Bulletin du Muséum National D’H istoire Naturelle, 10:77-88.
Schrire, B.D.; Lavin, M. & Lewis, G.P. 2005. Global distribution patterns of the Leguminosae: insights from recent phylogenies. In: I. Friis & H. Balslev (eds.) Plant diversity and complexity patterns: local, regional and global dimensions, Biologiske Skrifter, p. 375-422.
Schrire, B.D.; Lewis, G.P. & Lavin, M. 2005. Biogeography of the Leguminosae. In: G. Lewis; G. Schrire & B. Mackinder (eds.) Legumes of the world, Royal Botanic Gardens, p. 21-54.

362 REVISTA BRASILEIRA DE PALEONTOLOGIA, 17(3), 2014
Simpson, M.G. 2006. Plant Systematics. 1st ed. San Diego, Elsevier Academic Press, 590 p.
Souza, V.C. & Lorenzi, H. 2008. Botânica sistemática: guia ilustrado para identificação das famílias de angiospermas da flora brasileira. 2ª ed. Nova Odessa, Instituto Plantarum de Estudos da Flora, 704 p.
Tsukada, M. 1963. Pollen morphology and identification I. Eucaesalpinieae. Pollen et Spores, 5:239-284.
Uesugui, N. 1979. Palinologia: técnicas de tratamento de amostras. Boletim Técnico da Petrobras, 22:229-240.
Veiga, E.C. 2009. Tafofl ora paleógena da Formação Tremembé, ocorrência em afloramento do km 11 da rodovia SP-123, Município de Taubaté, Bacia sedimentar de Taubaté, Brasil. Programa de Pós-Graduação em Análise Geoambiental, Universidade de Guarulhos, Dissertação de Mestrado, 162 p.
Watson, L. & Dallwitz, M.J. 1992. The families of fl owering plants. Available athttp://delta-intkey.com; accessed on 31/07/2014.
Wey, A.S.; Barros, M.A.; Dino, R.; Bergqvist, L.P.; Mello, C.L. & Silva. R.C.B. 1999. Resultados preliminares de novos achados na bacia de Fonseca, MG (Paleógeno): macro e microfósseis vegetais. In: CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 16, 1999. Boletim de Resumos, Crato, SBP, p. 126-127.
Wikström, N.; Savolainen, V. & Chase, M.W. 2001. Evolution of the angiosperms: calibrating the family tree. Proceedings of the Royal Society B, 268:2211-2220. doi:10.1098/rspb.2001.1782
Wing, L.; Herrera, F.; Jaramillo, C.A.; Gómes-Navarro, C.; Wilf, P. & Labandeira, C.C. 2009. Late Paleocene fossils from the Cerrejón Formation, Colombia, are the earliest record of Neotropical rainforest. Proceedings of the National Academy of Sciences of the United States of America, 106:18627-18632. doi:10.1073/pnas.0905130106
Wojciechowski, M.F.; Johanna, M. & Bruce, J. 2006. Fabaceae. Available at http://tolweb.org/Fabaceae/21093/2006.06.14; accessed on 18/06/2010.
Wojciechowski, M.F.; Lavin, M. & Sanderson, M.J. 2004. A phylogeny of legumes (Leguminosae) based on analysis of the plastid mat gene resolves many well-supported subclades within the family. American Journal of Botany, 91:1846-1862. doi:10.3732/ajb.91.11.1846
Received in June, 2013; accepted in August, 2014.


















