
EXTRAÇÃO LÍQUIDO-LÍQUIDO DE ~-XILOSIDASE POR...
121
FACULDADE DE ENGENHARIA QUÍMICA DE LORENA DEPARTAMENTO DE BIOTECNOLOGIA Dissertação de Mestrado EXTRAÇÃO LÍQUIDO-LÍQUIDO DE ~-XILOSIDASE POR MICELAS REVERSAS . -·.,.... -- Francislene Andréia Hasmann Lorena - SP - Brasil Fevereiro de 2000
Transcript of EXTRAÇÃO LÍQUIDO-LÍQUIDO DE ~-XILOSIDASE POR...

FACULDADE DE ENGENHARIA QUÍMICA DE LORENA
DEPARTAMENTO DE BIOTECNOLOGIA
Dissertação de Mestrado
EXTRAÇÃO LÍQUIDO-LÍQUIDO DE ~-XILOSIDASE POR MICELAS REVERSAS
. -·.,.... --
Francislene Andréia Hasmann
Lorena - SP - Brasil Fevereiro de 2000

FACULDADE DE ENGENHARIA QUÍMICA DE LORENA.
DEPARTAMENTO DE BIOTECNOLOGIA
PÓS-GRADUAÇÃO EM BIOTECNOLOGIA INDUSTRIAL
EXTRAÇÃO LÍQUIDO-LÍQUIDO DE (3-XILOSIDASE POR MICELAS REVERSAS
Dissertação de mestrado apresentada como parte das exigências para obtenção do título de Mestre em Biotecnologia Industrial
Banca Examinadora:
Dr. Adalberto Pessoa Junior (presidente)
Dr. Elias Tambourghi
Ora. Inês Conceição Roberto
Estudante:
Francislene Andréia Hasmann
Lorena - SP - Brasil Fevereiro de 2000

FACULDADE DE ENGENHARIA QUÍMICA DE LORENA
DEPARTAMENTO DE BIOTECNOLOGIA
PÓS-GRADUAÇÃO EM BIOTECNOLOGIA INDUSTRIAL
EXTRAÇÃO LÍQUIDO-LÍQUIDO DE J3-XILOSIDASE POR MICELAS REVERSAS
Este exemplar corresponde a versão final da dissertação de mestrado aprovada pela banca examinadora
Lorena - SP - Brasil Fevereiro de 2000

RESUMO
Extração líquido-líquido de f3-xilosidase por micelas reversas. Francislene Andréia Hasmann. Dissertação de Mestrado. Programa de Pós-Graduação em Biotecnologia Industrial, Departamento de Biotecnologia, Faculdade de Engenharia Química de Lorena. Orientador: Dr. Adalberto Pessoa Jr. (Faculdade de Ciências Farmacêuticas - USP - CP 66083, CEP 05315-970, São Paulo/SP, Brasil). Banca examinadora:Dr. Elias 8. Tambourgi e Ora. Inês Conceição Roberto. Fevereiro de 2000.
O objetivo deste trabalho foi estudar a recuperação da enzima !3-xilosidase, produzida pelo fungo Penicillium janthinellum, através da técnica de extração líquido- líquido por micelas reversas. Para tanto utilizaram-se três sistemas micelares distintos formados a partir de BDBAC (agente tensoativo catiônico), AOT (agente tensoativo aniônico) e CTAB (agente tensoativo catiônico).
Foram utilizados planejamentos estatísiticos (distintos para cada sistema micelar) com o intuito de se realizar uma triagem das variáveis significativas a cada processo. A metodologia de superfície de resposta foi empregada para quantifificação dos níveis das variáveis significativas. O trabalho resultou em dois modelos matemáticos otimizados; um representando o processo de extração da enzima f3-xilosidase por micelas reversas de BDBAC e o outro modelo matemático para o agente tensoativo CTAB. No entanto, nas condições averiguadas, as extrações da enzima utilizando sistema de micelas reversas formadas a partir do agente tensoativo aniônico AOT resultaram em valores de recuperação iguais a zero.
Rendimentos em atividade da ordem de 43% foram previstos e observados experimentalmente para ambos os modelos obtidos. Observou-se ainda um aumento na atividade específica da enzima de 2 e 5,34 vezes, para o processo de extração por micelas reversas de BDBAC e CTAB, respectivamente.

ABSTRACT
This a study on the recovery of J3-xylosidase enzyme produced by Penicillium
janthinellum fungus, through liquid-liquid extraction by reversed micelles. Three
distinct micelar systems formed from BDBAC (cationic surfactant), AOT (anionic
surfactant) and CTAB (cationic surfactant) were employed. Statistical designs (one for
each micelar system) were used to select the most significant variables for each
process. Surface responce methodology was used to quantify the leveis of the
significant variables. The work resulted in two optimized mathematical models: one
for extraction by BDBAC reversed micelles and one for extraction by CT AB reversed
micelles. However, in the conditions tested, the extractions using the system formed
from AOT resulted in recovery values equal to zero. Activity yield of about 43% were
predicted and experimentally observed for both models. lncreases of 2 and 5.34
times in the specific enzyme activity were observed for the extractions by BDBAC and
CTAB reversed micelles, respectivelly.

Aos meus pais/ irmãs e scombos. Ao meu mando Luiz Cláudio.

AGRADECIMENTOS
Ao Prof. Dr. Adalberto Pessoa Jr. não somente pela orientação dedicada
durante a realização deste trabalho, mas também pelos constantes incentivos e
amizade.
À Prof. Ora. Inês Conceição Roberto, pela amizade, orientação e freqüente
colaboração no desenvolvimento deste trabalho.
Ao querido e sempre disponível, o amigo e professor Dr. Arnaldo Márcio R.
Prata, pela interminável paciência e valiosa colaboração.
Ao Paulinho e Rita, pela amizade e auxílios prestados dentro e fora do
laboratório.
Á Maria Eunice pela inestimável participação.
Aos demais funcionários e professores do Departamento de Biotecnologia.
Aos alunos que com seu companheirismo, amizade e principalmente, festas,
tornaram o desenvolvimento deste trabalho ainda mais prazeroso.
Ao Departamento de Biotecnologia da Faculdade de Engenharia Química de
Lorena, pelo apoio durante o desenvolvimento deste trabalho.
À FAPESP e CAPES, pelo apoio financeiro.
ii

CONTEÚDO
LISTA DE FIGURAS v LISTA DE TABELAS vii
LISTA DE ABREVIATURAS x
1 - INTRODUÇÃO 1
2 - REVISÃO BIBLIOGRÁFICA 3
2.1 - PROCESSOS DE RECUPERAÇÃO DE BIOPRODUTOS-("DOWNSTREAM PROCESSING") ............................................................................................................................ 3 2.2 - PRECIPITAÇÃO 5
2.2.1 - Precipitação por Adição Fracionada de Etanol 6 2.3 - EXTRAÇÃO LÍQUIDO-LÍQUIDO POR MICELAS REVERSAS 7
2.3.1 - Micelas 7 2.3.2 - Tipos de Solventes 8 2.3.3 - Tipos de Co-solventes 8 2.3.4 - Tipos de Tensoativos 9 2.3.5 - Parâmetro Wo······················································································ 10 2.3.6 - Composição das Micelas Reversas 11 2.3.7 - Sistemas Micelares Reversos 12
2.4 - EXTRAÇÃO DE PROTEÍNAS EM SISTEMAS DE MICELAS REVERSAS 14 2.4.1 - Mecanismo de Formação de Mice las Reversas 16 2.4.2 - Modelos de Absorção de Biomoléculas 17 2.4.3 - Fatores que Afetam a Extração de Proteínas 18
2.4.3. 1 - pH 18 2.4.3.2 - Força iônica e tipo de íon 20 2.4.3.3 - Tipo de proteína 22 2.4.3.4 - Temperatura 22 2.4.3.5 - Tensoativo 23 2.4.3.6 - Tamanho das mice/as 26 2.4. 3. 7 - Influência da velocidade de agitação 27
2.5 - UTILIZAÇÃO DE ENZIMAS XILANOLÍTICAS 27 2.5.1 - f3-xilosidase 29
2.5.1.1 - pH ótimo 30 2.5.1.2 - Temperatura ótima 30
3. OBJETIVOS 31
4 - MATERIAIS E MÉTODOS 33
4.1 - MICRORGANISMO 33 4.2 - MEIO DE MANUTENÇÃO DO FUNGO 33 4.3 - HIDROLISADO HEMICELULÓSICO DE BAGAÇO DE CANA-DE-AÇÚCAR 34 4.4 - PRODUÇÃO DA ENZIMA f3-XILOSIDASE 34
iii

·"········"··-·;-··---..._ »: e: nci1rnnc:l,. .... ,
f ~ '(.,. ., !,:,, -, /» o> ! '?) '\'.'.::.-\ f; . ., - - . . - ·-··· ~:.- \
, ._.. ( t·· r~ t ! \ &..: '
4.5 -AVALIAÇÃO DA ATIVIDADE ENZIMÁTICA \t ·~· .. ·: .. ~ ~ ~i4 • \(.V '
4.5.1 - Xilanases \~ ';' .!/ 34 4.5.2 - f3-xilosidase >:-~./~.a.1:.~_:::r. 35
4.6 - PRECIPITAÇÃO PELA ADIÇÃO FRACIONADA DE ETANOL --:-:·:·.::~~ 36 4. 7 - DETERMINAÇÃO DOS PARÂMETROS CINÉTICOS 36 4.8 - DETERMINAÇÃO DA ESTABILIDADE À ESTOCAGEM DA ENZIMA f3-XILOSIDASE 37 4.9 - EXTRAÇÃO LÍQUIDO-LÍQUIDO POR MICELAS REVERSAS 37 4.10 - DELINEAMENTO EXPERIMENTAL. 39 4.11 - DETERMINAÇÃO DO RAIO MICELAR - W0 43 4.11 - ENSAIOS ELETROFORÉTICOS 44
4.11. 1 - Preparo das Amostras 44 4.11.1.1 - Diálise 44 4.11.1.2 - Liofilização 44
4. 11.2 - Condução dos Ensaios eletroforéticos 45 4.12 - DOSAGEM DO TEOR TOTAL DE PROTEÍNAS - MÉTODO DE LOWRY 4 7 4.13 -ANÁLISE DOS RESULTADOS 47
5 - RESULTADOS E DISCUSSÕES 49
5. 1 - A ENZIMA f3-XILOSIDASE 49 5.1. 1 - Estabilidade 49 5.1.2 - Parâmetros Cinéticos 51
5.2 - EXTRAÇÃO DA ENZIMA f3-XILOSIDASE POR SISTEMA DE MICELAS REVERSAS FORMADO A PARTIR DO AGENTE TENSOATIVO CATIÔNICO BDBAC 54
5.2.1 - Otimização do Processo de Extração por Micelas Reversas de BDBAC em lsooctano e Hexanol 65 5.2.2 - Determinação do Raio Micelar 70 5.2.3 - Eletroforese 71
5.3 - EXTRAÇÃO DA ENZIMA f3-XILOSIDASE POR SISTEMA DE MICELAS REVERSAS FORMADO A PARTIR DO AGENTE TENSOATIVOANIÔNICOAOT 74 5.4 - EXTRAÇÃO DA ENZIMA f3-XILOSIDASE POR SISTEMA DE MICELAS REVERSAS FORMADO A PARTIR DO AGENTE TENSOATIVO CATIÔNICO CTAB 76
5.4. 1 - Otimização do Processo de Extração por Micelas Reversas de CTAB em lsooctano, Hexanol e Butanol 86 5.4.2 - Determinação do Raio Micelar 93
5.5 - EXTRAÇÃO DA ENZIMA f3-XILOSIDASE POR MICELAS REVERSAS DE BDBAC E CTAB, DIRETAMENTE DO MEIO FERMENTADO 94
6 - CONCLUSÕES 97
7 - REFERÊNCIAS BIBLIOGRÁFICAS 99
iv

LISTA DE FIGURAS
Figura 2.1 Produção Nacional de Enzimas versus Importações (Adaptado de ENZITEC, 1999). 4
Figura 2.2 Esquema de Micela, Micela Reversa Vazia e Micela Reversa Contendo Proteína em seu Interior. 7
Figura 2.3 Composição de uma Micela Reversa (Adaptado de SEOUD, 1999). 12
Figura 2.4 Estruturas que Formam o Sistema Genérico/Tensoativo/Solvente/ H20 (Adaptado de SEOUD 1999). 13
Figura 2.5 Diagramas de Fases dos Agentes Tensoativos CTAB (I) e AOT (II). 13
Figura 2.6 Representação Esquemática do Processo de Extração em Sistemas Micelares Reversos. 15
Figura 2.7 Mecanismo de Formação de Micelas Reversas (Adaptado de DUNGAN, 1991). 17
Figura 2.8 Modelos de Proteínas Englobadas por Micelas Reversas (Adaptado de LUISI, 1988). 18
Figura 2.9 Evolução da Formação de Estruturas no Sistema de Micelas Reversas (Adaptado de SEOUD et ai., 1999). 25
Figura 4.1 Representação Esquemática da Etapa de Extração de uma Biomolécula por Micelas Reversas. 38
Figura 4.2 Representação Esquemática da Etapa de Reextração de uma Biomolécula por Micelas Reversas. 38
Figura 5.1 Atividade da í3-xilosidase após Precipitação Fracionada com Etanol em Função da Concentração de Substrato (pNpX) para os Valores de pH 3,0; 6,0; 7,0 e 9,0. 51
Figura 5.2 Curvas Correspondentes ao Inverso dos Valores das Atividades (1/v) de í3-xilosidase em Função do Inverso das Concentrações do Substrato (1/s). 52
Figura 5.3a Recuperação da í3-xilosidase por Micelas Reversas em Função da Temperatura e do pH, usando BDBAC O, 15M, Condutividade Elétrica de 7,0mS/cm, 7,5% de Hexanol. 58
Figura 5.3b Recuperação da í3-xilosidase por Micelas Reversas em Função da Temperatura e da Concentração do BDBAC, usando pH6,0, Condutividade Elétrica de 7,0mS/cm, 7,5% de Hexanol. 58
Figura 5.3c Recuperação da í3-xilosidase por Micelas Reversas em Função da Concentração de BDBAC e do pH, temperatura de 26ºC, 7,0mS/cm, 7,5% de Hexanol. 59
V

Figura 5.3d Recuperação da '3-xilosidase por Micelas Reversas em Função da Condutividade Elétrica e da Concentração de Hexanol, usando BDBAC O, 15M, 7,0mS/cm, 7,5% de Hexanol. 59
Figura 5.4 Extração de uma Proteína por Micelas Reversas Considerando a Existência de Forças Hidrofóbicas de Interação (Adaptado de LUISI, et ai., 1988). 61
Figura 5.5 Superfície de Resposta Descrita pelo Modelo da Equação 5.2, que Representa a Extração em Condições Otimizadas de '3- xilosidase por Micelas Reversas do Agente T ensoativo Catiônico BDBAC em lsooctano e Hexanol. 69
Figura 5.6 Eletroforese em Condições Desnaturantes (SDS-PAGE) das Etapas do Processo de Extração da Enzima '3-xilosidase por Micelas Reversas Pré Purificada por Precipitação Fracionada com Etanol: Linha 1 e 2 - Marcadores de Massa Molar (205, 116, 97, 84, 66, 55, 45, 36, 29, 24, 20 e 6,5 kDa), Linha 3 - Precipitado Solubilizado, Linha 4 - F AI e Linha 5 - F AII. 72
Figura 5. 7a Recuperação da p-xilosidase por Micelas Reversas em função do pH e da Concentração de CTAB, usando, Condutividade Elétrica de 7,0mS/cm, 7.5% de Hexanol e 15% de butanol. 80
Figura 5.7b Recuperação da '3-xilosidase por Micelas Reversas em Função do pH e da Concentração de Butanol, usando CTAB O, 15M, 7,0mS/cm e 7,5% de Hexanol. 80
Figura 5.7c Recuperação da '3-xilosidase por Micelas Reversas em Função da Condutividade Elétrica e da Concentração de CTAB, usando pH 5,5, 7,0 mS/cm e 7,5% de Hexanol. 81
Figura 5.7d Recuperação da '3-xilosidase por Micelas Reversas em Função da Concentração de CTAB e da Concentração de Butanol, usando pH 5,5, 7,0mS/cm e 7,5% de Hexanol.
81
Figura 5.7e Recuperação da '3-xilosidase por Micelas Reversas em Função da Concentração de Hexanol e da Concentração de Butanol, usando pH 5,5, 7,0mS/cm e CTAB O, 15M. 82
Figura 5.8 Esboço do Planejamento em Estrela Ortogonal. 87
Figura 5.9 Superfície de Resposta Descrita pelo Modelo da Equação 5.4, que Representa a Extração em Condições Otimizadas de 13- xilosidase por Micelas Reversas do Agente T ensoativo Catiônico CT AB em lsooctano, Hexanol e Butanol. 92
Figura 5.10 Efeito do Número de Etapas de DSP no Rendimento Final do Bioproduto. 94
vi

LISTA DE TABELAS
Tabela 4.1 Valores das Variáveis Empregadas nos Planejamentos para 39 Recuperação da 13-Xilosidase por Micelas Reversas
Tabela 4.2 Matriz do Planejamento Fatorial Fracionário 2s-1 com 01 Repetição Utilizada para Avaliação da Influência do pH, da Condutividade Elétrica, da Concentração de BDBAC, da 40 Concentração de Hexano! e da Temperatura na Recuperação da 13-xilosidase por Micelas Reversas do Agente Tensoativo Catiônico BDBAC
Tabela 4.3 Matriz do Planejamento Fatorial Completo 24 com 03 Repetições no Ponto Central Utilizada para Avaliação da Influência do pH, Condutividade Elétrica, Concentração de AOT e Temperatura na 41 Recuperação da 13-Xilosidase por Micelas Reversas do Agente Tensoativo Aniônico AOT
Tabela 4.4 Matriz do Planejamento Fatorial Fracionário 25•1 com 03 Repetições no Ponto Central Utilizada para Avaliação da Influência do pH, da Condutividade Elétrica, da Concentração de 42 CTAB, da Concentração de Hexanol e da Concentração de Butano! na Recuperação da 13-xilosidase por Micelas Reversas do Agente T ensoativo Catiônico CT AB
Tabela 4.5 Seqüência Usada para Desenvolvimento do Processo Otimizado 46 de Coloração de Géis SDS-PAGE por Prata
Tabela 5.1 Atividade da 13-xilosidase em Diferentes Valores de pH, em 50 Função do Tempo, a 4°C
' --
Tabela 5.2 Atividade da 13-xilosidase em Diferentes Valores de pH, em Função do Tempo, a -4ºC 50
Tabela 5.3 ~(app) e Vm da 13-xilosidase Obtidos após Precipitação Fracionada com Etanol em Função do Inverso das 53 Concentrações do Substrato para Diferentes Valores de pH
Tabela 5.4 Planejamento Fatorial Fracionário 2s-1 com 1 Repetição Utilizado para Avaliação da Influência do pH, condutividade elétrica, concentração de hexanol e Temperatura na Recuperação da 54 13-xilosidase por Micelas Reversas de Agente Tensoativo Catiônico BDBAC
Tabela 5.5 Estimativa dos Efeitos, Erros-padrão e teste t (Student) para os Parâmetros Estudados na Recuperação da 13-xilosidase por 56 Micelas Reversas de BDBAC
vii

Tabela 5.6 Parâmetros da Análise de Variância (ANOVA) dos Fatores Significativos ao Processo de Extração da Enzima í3-xilosidase 63 por Micelas Reversas do Agente Tensoativo BDBAC
Tabela 5. 7 Análise de Variância de Regressão para o Modelo Representativo do Processo de Extração de í3-xilosidase por 64 Micelas Reversas de BDBAC, na Região em Estudo
Tabela 5.8 Matriz do Planejamento Fatorial Completo 22 Com Face Centrada e 03 Repetições no Ponto Central Utilizada para Avaliação da Influência do Binômio pH e Temperatura na 66 Recuperação da f3-xilosidase por Micelas Reversas de Agente T ensoativo Catiônico BDBAC
Tabela 5.9 Variáveis, Coeficientes, Teste t (Student), Valores de p do Planejamento Completo com Face Centrada 22 com 03 Repetições no Ponto Central para Extração de f3-xilosidase por 68 Micelas Reversas de BDBAC
Tabela 5.10 Parâmetros da Análise de Variância (ANOVA) dos Fatores Significativos ao Processo de Extração de í3-xilosidase por 69 Micelas Reversas de BDBAC
Tabela 5.11 Aumento de Pureza para Enzima í3-xilosidase Extraída por 70 Micelas Reversas de Agente Tensoativo Catiônico BDBAC
Tabela 5.12 Wo e Rm Determinados na FAI E FAII Obtidas após Extração por Micelas Reversas de BDBAC a 0,2M, 1 O % de Hexanol, Condutividade Elétrica de 4 mS/cm, Temperatura de 24ºC e pH 71
3,3
Tabela 5.13 Planejamento Fatorial Completo 24 com 2 Repetições no Ponto Central Utilizado para Avaliação da Influência do pH, Condutividade Elétrica, Concentração de AOT e Temperatura na 74 Recuperação da í3-xilosidase por Micelas Reversas de Agente T ensoativo Aniôníco AOT
Tabela 5.14 Planejamento Fatorial Fracionário 25-1 para Estudo da Influência do pH, Concentração de CTAB, Condutividade Elétrica, Concentração de Hexano!, Concentração de Butano!, na 76 Recuperação de f3-xilosidase por Micelas Reversas do Agente Tensoativo Catiônico CTAB
Tabela 5.15 Estimativa dos Efeitos, Erros-padrão e teste t (Student) para os parâmetros Estudados na Recuperação da í3-xilosidase por 78 Micelas Reversas de CTAB
Tabela 5.16 Parâmetros de Análise de Variância para os Efeitos 83 Significativos ao Processo de Extração de f3-xilosidase por
t -- .,._
. ---
viii

Micelas Reversas de CTAB
Tabela 5.17 Análise de Variância de Regressão para o Modelo Representativo do Processo de Extração de 13-xilosidase por 85 Micelas Reversas de CTAB, na Região em Estudo
Tabela 5.18 Níveis Codificados E Valores Utilizados Na Realização Dos Ensaios Do Passo Ascendente no Processo de Extração de 87 13-xilosidase por Micelas Reversas de Agente Tensoativo Catiônico CTAB
Tabela 5.19 Planejamento Ortogonal Completo 22 com 03 Repetições no Ponto Central para Estudo da Influência da Concentração de CT AB e da Concentração de Butanol, na Recuperação de 88 j3-xilosidase por Micelas Reversas do Agente Tensoativo Catiônico CTAB
Tabela 5.20 Parâmetros de Análise de Variância para os Efeitos Significativos ao Processo de Extração de 13-xilosidase por 90 Micelas Reversas de CTAB
Tabela 5.21 Variáveis, Coeficientes, Teste t (Student), Valores de p do Planejamento Completo Ortogonal 22 com 03 Repetições no 90 Ponto Central para Extração de 13-xilosidase por Micelas Reversas de CTAB
Tabela 5.22 Parâmetros da Análise de Variância (ANOVA) dos Fatores Significativos ao Processo de Extração de 13-xilosidase por 91 Micelas Reversas de BDBAC
Tabela 5.23 Aumento de Pureza para Enzima f3-xilosidase Extraída por 92 Micelas Reversas de Agente Tensoativo Catiônico CTAB
Tabela 5.24 W0 e Rm Determinados na FAI E FAII Obtidas após Extração por Micelas Reversas de CTAB a 0,26M, com 10% de Hexanol e 29,4% de butanol, em pH 8,0, Condutividade Elétrica de 93 4 mS/cm, Temperatura de 26ºC
Tabela 5.25 Comparação entre a Porcentagem de Recuperação da Extração por Micelas Reversas de BDBAC e CTAB de j3-xilosidase Sem 95 Precipitação e Com Precipitação por Etanol
ix

Abreviaturas
DSP
CTAB
BDBAC
AOT
TOMAC
DDAB
CPB
NaDEHP
Wo Nag
Rm
pi CMC
ROH
DNS
pNpX
KM Vm FAI FAII FMI
FMII
BSA
%R
ANOVA
LISTA DE NOMENCLATURAS
Definições
"Downstream Processing"; recuperação de produtos biotecnológicos
brometo de cetiltrimetil amônio
cloreto de benzil dodecil bis(hidroxietil) amônio
bis(2-etilhexil) sulfosuccinato de sódio
cloreto de trioctilmetil amônio
brometo de didodecil dimetil amônio
brometo de cetil piridínio
bis(2-etilhexil) fosfato de sódio
"Water in oil"; quantidade de água presente no interior da micela
Número de moléculas de tensoativo agregadas na micela
Raio micelar
Ponto isoelétrico
Concentração micelar crítica, concentração mínima de tensoativo necessária para a formação de micelas Solvente qualquer
Ácido 3,5 dinitrosalicílico
p-nitrofenil-f3-D-xilopiranosídeo
Constante de Michaelis e Menten
Velocidade máxima de reação enzimática
Fase aquosa I
Fase aquosa II
Fase micelar I
Fase micelar II
"Sovine serum albumine"; albumina de soro bovino
Porcentagem de enzima recuperada
Análise de Variância
X

Introdução
1 - INTRODUÇÃO
Existem inúmeras alternativas para extração seletiva de enzimas dentre as
quais podemos citar: precipitação, extração líquido-líquido em sistemas de duas
fases aquosas e extração líquido-líquido por micelas reversas.
Este trabalho tem como objetivo a recuperação da enzima 13- xilosidase,
utilizando a técnica de extração por micelas reversas, esta enzima pertence ao
complexo de xilanases e possui vasta aplicação no setor industrial, como na
clarificação de sucos e vinhos e na filtração de cerveja. A aplicação mais promissora
de xilanases é na etapa de branqueamento de polpas de celulose, na indústria de
papel e celulose, como alternativa para redução no uso de agentes clorados, no
intuito de se diminuir a agressão ao ambiente causada por tais agentes. Consiste de
uma alternativa auxiliar para remoção da lignina residual que confere à polpa uma
cor escura (CHRISTOV & PRIOR, 1996; MILAGRES & PRADES, 1994;VIIKARI et ai.,
1991).
4 -· -
Assim, o principal objetivo do desenvolvimento de novos processos de
separação é a possibilidade de recuperação e purificação de enzimas a custos
viáveis.
Apesar do interesse de vários grupos de pesquisa em desenvolver e aplicar a
técnica de extração por micelas reversas para separação e purificação de enzimas,

Introdução
sabe-se que o processo pode ser modificado por um grande número de variáveis que
alteram a solubilização das proteínas.
As variáveis mais importantes são aquelas que regulam as interações entre as
proteínas e os outros componentes formadores das micelas reversas. São o pH, a
força iônica do meio e outras variáveis que definem a forma e tamanho das micelas
reversas, tais como temperatura, concentração do agente tensoativo e agitação.
Assim, devido ao grande número de variáveis importantes para o processo de
extração por micelas reversas, o uso da ferramenta estatística, através de
planejamentos experimentais, torna-se de suma importância para identificação da
significância de cada uma das variáveis ao processo estudado.
Na presente dissertação, foram estudadas as seguintes variáveis que atuam
sobre o rendimento da extração líquido-líquido de j3-xilosidase por micelas reversas:
pH, concentração do agente tensoativo, condutividade elétrica (força iônica),
concentração de co-solvente e temperatura.
2

Revisão Bibliográfica
2 • REVISÃO BIBLIOGRÁFICA
2.1 - PROCESSOS DE RECUPERAÇÃO DE BIOPRODUTOS ("OOWNSTREAM PROCESSING")
O objetivo do "Downstream Processing" (DSP) é recuperar o produto de um
biorreator obtendo-o no nível desejado de pureza e com a formulação correta. Para
isso, diferentes tipos de operações são envolvidas. Com exceção do rompimento
celular, da secagem e da formulação (a qual inclui imobilização), processos de
biosseparação formam o centro do "Downstream Processing" (KRIJGSMAN, 1991 ).
DSP é particularmente importante em biotecnologia onde as formas finais dos
produtos são, usualmente, muito diferentes do estado em que são obtidos no
biorreator. Como exemplo, pode ser citado um típico processo de fermentação que
fornece uma mistura de massa celular com outros componentes do meio nutriente. O
produto desejado pode estar dentro da célula, como constituinte de uma mistura
muito complexa, no meio aquoso diluído, ou ainda distribuído entre os dois. Em
qualquer caso, sua recuperação, concentração e purificação necessita de várias
operações, as quais muitas vezes são limitadas por aspectos econômicos (SCOPES,
1994). Muitas operações padrões nos laboratórios tornam-se impraticáveis, ou
dispendiosas, em processos industriais. Além disso, bioprodutos são freqüentemente
compostos sensíveis, cujas estruturas ativas só podem ser preservadas sob
condições limitadas de pH, temperatura e força iônica. Em função destas restrições
não existem técnicas gerais, ou mesmo uma seqüência de operações que possa ser
3

Revisão Bibliogrãfica
recomendada para recuperação e purificação de qualquer produto biotecnológico.
Cada caso requer o estudo aprofundado para definição das operações unitárias
ideais (GLATZ, 1990; SCAWEN et ai., 1985).
Mesmo sendo onerosas as operações necessárias para purificar diversos
produtos biotecnológicos, há uma tendência crescente da demanda desses produtos
na próxima década. Pesquisas recentes apontam um aumento médio anual de 12%
nas vendas de produtos biotecnológicos nos Estados Unidos, partindo de US$1 O, 1
bilhões no ano de 1996, para uma previsão de US$32,4 bilhões no ano de 2006. As
enzimas fazem parte de um segmento de expressiva participação no contexto geral
de produtos biotecnológicos, o qual deve saltar de US$275 milhões em 1996 para
US$1,6 bilhão no ano de 2006, correspondendo a uma alta nas vendas de 19% ao
ano durante este período (SHAMEL & KEOUGH, 1995).
A Figura 2.1 ilustra claramente a dependência do mercado interno brasileiro
de enzimas importadas.
MERCADO DE ENZIMAS NO BRASIL (US$ J rf)
«oco flTílTITnTITTTr,rr--r-r-- 30000
20000
10000
º~~~~~~~~~,!!~ 19 lmpo,tação -:-- • Exportação
....
Figura 2.1 - Produção Nacional de Enzimas versus Importações (Adaptado de ENZITEC, 1999)
Para satisfazer a demanda destes produtos, várias são as técnicas de DSP
empregadas, dentre as quais podem ser citadas a precipitação por sais, a
precipitação por polietilenoglicol, a precipitação por solventes orgânicos,
ultrafiltração, a cromatografia, a extração em sistemas de duas fases aquosas e por
4

Revisão Bibliogrãfica
micelas reversas, entre outras (PESSOA JR & VITOLO, 1998; SILVA et ai., 1998;
CORTEZ et ai., 1998; COSTA et ai., 1998; ASENJO, 1990).
A extração líquido-líquido usando micelas reversas é potencialmente atrativa
em processos biotecnológicos, pois com a manipulação de alguns fatores que regem
o processo, é possível não somente a extração de proteínas, mas também a sua
separação e concentração, utilizando-se de uma única etapa de DSP (RAHAMAN &
HA TTON, 1988).
2.2 - PRECIPITAÇÃO
A precipitação é uma operação unitária amplamente utilizada em recuperação
e purificação de biomoléculas. Com amplo consenso, a precipitação pode ser
definida como um processo no qual a relação entre um soluto e um solvente é
alterada, sendo que no geral, a solubilidade é reduzida pela adição de um reagente
que modifica o balanço de energia entre eles (GARCIA, 1993).
Segundo GARCIA (1993), a precipitação é um processo através do qual uma
proteína no estado coloidal é desestabilizada, o que promove a agregação de suas
moléculas. Isto requer a adição de um agente apropriado e agitação, fatores que
modificam o balanço de atração e repulsão entre as partículas.
Para o desenvolvimento de um processo de precipitação existem fatores que
devem ser considerados: seleção do método de precipitação, seleção e dosagem do
agente precipitante para que desestabilize a solução e a dinâmica da precipitação.
Os métodos de precipitação incluem: precipitação pela adição de um sal
neutro ("salting-out"), pelo controle do pH no ponto isoelétrico, pela adição de
polímeros hidrofílicos não iônicos, pela adição de íons metálicos polivalentes e
finalmente pela adição de solvente orgânico miscível. Este último tem se mostrado
efetivo na precipitação da j3-xilosidase (PESSOA JR & VITOLO, 1999; CORTEZ, 1998).
5

Revisão Bibliogrãfica
2.2.1- Precipitação por Adição Fracionada de Etanol
A precipitação fracionada é uma importante operação para recuperação e
purificação de proteínas (tanto para escala laboratorial quanto industrial). Tem como
objetivo separar seletivamente uma proteína de interesse de uma mistura de
proteínas e contaminantes não protéicos (lipídeos e ácidos nucléicos, por exemplo)
(CLARKSON et ai. 1996).
Esta técnica é usualmente executada em dois "cortes": no primeiro, são
removidas proteínas de baixa solubilidade através da fase precipitada, sendo que no
geral, apenas pequena parcela da proteína de interesse é arrastada pela
precipitação. No segundo corte, recupera-se a proteína de interesse através da fase
precipitada. No entanto, proteínas de alta solubilidade consideradas contaminantes
(sem interesse), podem permanecer em solução, sendo necessário desta forma, a
otimização destes dois pontos de corte, de modo a se obter um nível aceitável de
purificação sem prejuízos para o rendimento do processo (SCOPES, 1994; CORTEZ &
PESSOA, 1999).
CORTEZ (1998), partindo de um caldo enzimático contendo o complexo
xilanolítico, observou que com a adição de uma quantidade de etanol equivalente a
20% (v/v) do meio total, grande parcela das proteínas, analisadas pelo método de
BRADFORD (1976), sofriam precipitação, e com a fração equivalente a 60% (v/v) de
etanol, promovia-se a precipitação da 13-xilosidase, originando um meio onde
praticamente só esta enzima está presente em quantidades apreciáveis.
A precipitação fracionada causa uma pequena perda de rendimento em
função da precipitação de parte da proteína de interesse juntamente com aquelas
consideradas contaminantes, no entanto, o aumento de pureza compensa esta perda
(CORTEZ, 1998).
6

Revisão Bibliográfica
2.3 - EXTRAÇÃO LÍQUIDO-LÍQUIDO POR MICELAS REVERSAS
2.3.1 - Mice/as
O termo "micela" foi utilizado pela primeira vez por McBain, em 1913, para
descrever os agregados de eletrólitos anfifílicos (substâncias que possuem uma
parte hidrofílica e outra hidrofóbica) em soluções aquosas. As micelas reversas são
agregados de moléculas de agentes tensoativos contidos em um solvente orgânico
apoiar.
A técnica de extração utilizando micelas reversas baseia-se na propriedade
que os agentes tensoativos (moléculas que possuem em sua estrutura uma
extremidade hidrofóbica e outra hidrofílica) têm de se auto-organizar quando em
solução aquosa. A extremidade apoiar da molécula de tensoativo procura um
ambiente apoiar. Sendo o único ambiente apoiar existente no meio a própria
extremidade apoiar da outra molécula, formam-se estruturas esféricas e elipsoidais,
que são chamadas micelas. Porém quando em meio apoiar estas micelas
apresentam orientação inversa em relação à formação da estrutura micelar em água,
sendo esta formação denominada micela reversa. Ela contém água em seu núcleo
interno e fica dispersa num meio com solvente orgânico (KREI & HUSTEDT, 1989). A
Figura 2.2 mostra a diferença entre os dois tipos de micelas.
Mi cela Micelas Reversas
- .. - ... - ...... - -- .,,
~-~ --,= -,~~ ' -
. . - - - ,,. - - - - .. - . _- \ - ..::::- . ..,_
~ Meio Aquoso Biomolécula
0 Meio Orgânico
Figura 2.2 - Esquema de Micela, Micela Reversa Vazia e Micela Reversa Contendo Proteína em seu Interior.
7

Revisão Bibliogrãfica
Os sistemas micelares representados na Figura 2.2 apresentam formas
aproximadamente esféricas, no entanto, micelas reversas podem ser elípticas; o
diâmetro varia em torno de 15 a 20 A, porém pode chegar até a 200 A (KREI &
HUSTEDT, 1989).
Tanto a forma e o tamanho, quanto a transferência de enzimas presentes
numa fase aquosa para a pseudofase micelar dependem da natureza do solvente e
do co-solvente, do tipo de tensoativo, da temperatura, da força iônica e do pH, dentre
outros (RODRIGUES et ai., 1999; ZAMARRO et ai., 1996; KADAM, 1986).
2.3.2- Tipos de Solventes
Para que haja a formação das micelas reversas é necessário que o solvente e
a água sejam imiscíveis. Dos diversos solventes que podem ser usados nos
sistemas de micelas reversas, os mais comuns são hidrocarbonetos alifáticos, como
o isooctano, o n-octano, o n-heptano e o ciclo hexano (RODRIGUES et ai., 1999;
CHANG & CHEN, 1996; LUISI et ai., 1988).
O tipo de solvente pode influenciar no tamanho da micela reversa. Sistemas
com hexano, isooctano, octano e querosene formam micelas reversas com diâmetros
maiores que sistemas com ciclo hexano, xileno e clorofórmio (CHANG & CHEN, 1996;
KADAM, 1986). A diminuição da concentração do solvente, ocasiona a redução na
transferência de proteínas para o interior da micela reversa, prejudicando o processo
de extração (KREI & HUSTEDT, 1992).
2.3.3 - Tipos de Co-solventes
Os co-solventes são utilizados como auxiliares em sistemas de micelas
reversas de tensoativos catiônicos para aumentar o seu tamanho, pois micelas
formadas por estas substâncias são muito pequenas. O co-solvente assemelha-se a
uma solução tampão que provoca uma forte repulsão entre as interações íon-íon dos
núcleos do tensoativo, permitindo a formação do núcleo interno da micela reversa
(LUISI et ai., 1988). Geralmente utilizam-se, como co-solventes, álcoois de cadeias
8

Revisão Bibliográfica
não muito curtas como hexanol, n-butanol e octanol (KREI & HUSTEDT, 1992; CHANG
& CHEN, 1996; PIRES et ai., 1996).
Os co-solventes também podem ter influência sobre as estruturas das micelas
reversas, como é o caso da água solubilizada na micela reversa de TOMAC que
sofre variação em função da quantidade de co-solvente do meio (CHANG & CHEN,
1996). Também a extração de a-amilase, utilizando Aliquat 336 mostrou-se sensível
às variações no tipo de co-solvente utilizado no sistema (HILHORST et ai., 1995).
2.3.4 - Tipos de Tensoativos
Os agentes tensoativos (ou surfactantes) são moléculas anfifílicas com um
núcleo polar (parte hidrofílica) e uma cauda hidrofóbica. Eles são classificados de
acordo com a carga de sua parte polar em neutros, aniônicos e catiônicos
(MEYER, 1992; KREI & HUSTEDT, 1989). De acordo com a literatura, o agente
tensoativo aniônico AOT, tem sido amplamente utilizado (PATEL et ai., 1996;
REGALADO et ai., 1996; ZAMARRO et ai., 1996; HUANG & LEE, 1994; HENTSCH et
ai., 1992; AIRES-BARROS et ai., 1991) e mostra-se adequado para separação de
proteínas de elevado ponto isoelétrico. Para proteínas de baixo ponto isoelétrico, são
recomendados agentes tensoativos catiônicos como o BDBAC. No entanto, tais
tensoativos formam micelas reversas pequenas, tornando-se necessária a adição de
um co-solvente, ou ainda de um outro tensoativo não iônico, para obtenção de
micelas maiores (HILHORST et ai., 1995).
Apesar da existência de inúmeros tensoativos diferentes, os mais utilizados
para a formação de micelas reversas são:
a) Catiônicos:
brometo de cetiltrimetil amônio - CTAB;
cloreto de benzi! dodecil bis(hidroxietil) amônio - BDBAC;
cloreto de trioctilmetil amônio - TOMAC;
brometo de cetil piridínio - CPB;
brometo de didodecil dimetil amônio - DDAB.
9

Revisão Bibliográfica
b) Aniônico:
bis(2-etilhexil) sulfosuccinato de sódio - AOT;
bis(2-etilhexil) fosfato de sódio - NaDEHP.
e) Neutros:
lecitina de soja;
lecitina de clara de ovo;
Rewopal HV5;
Tergitol NP-4.
2.3.5 - Parâmetro W0
A quantidade de água nas micelas reversas, definida pelo parâmetro Wo
("water in oil"), pode ser obtida pela Equação 2.1, que exprime a relação entre a
concentração molar de água e de tensoativo (LUISI et ai., 1988). Este é um parâmetro
de suma importância no sistema, pois determina a estrutura e o tamanho das micelas
reversas, e também o número de moléculas de tensoativo por micela reversa
(CASTRO & CABRAL, 1988; AIRES-BARROS, 1991 ). Assumindo a hipótese de que as
micelas reversas sejam esféricas e que existe uma distribuição monodispersa dos
raios dos núcleos aquosos, o raio micelar (Rm) pode ser equacionado pela Equação
2.2. É possível ainda, equacionar-se o número de moléculas de tensoativo que forma
cada micela, que é o número de agregação (Nag). Tal parâmetro pode ser calculado
utilizando-se a Equação 2.3. No entanto esse valor pode variar com a presença da
proteína no interior da micela (L YE et ai., 1995).
- -_.,..,
Wo = [H20]/[tens.]
Rm (A)= 1,64 Wo
Nag = 0,611 Wo
Equação 2.1
Equação 2.2
Equação2.3
10

Revisão Bibliográfica
O tamanho das micelas reversas pode ser determinado também através de
métodos experimentais como ressonância magnética e técnicas de dispersão de luz
(SEOUD et ai., 1999).
2.3.6- Composição das Mice/as Reversas
A micela reversa é formada por três regiões: uma primeira região formada
pelas caudas hidrofóbicas das moléculas do tensoativo, que fica em contato direto
com o solvente apoiar, a segunda região é a periferia micelar onde as moléculas de
água estão fortemente ligadas aos núcleos polares do tensoativo e, por fim, a
terceira região, o centro micelar, formado pela água contida no interior da micela que
está ligada ao tensoativo ou água livre (AIRES-BARROS, 1991). A Figura 2.3 ilustra a
composição de uma micela reversa.
A água que fica na periferia micelar possui propriedades físico-químicas
diferentes da água em solução, principalmente em micelas de diâmetro pequeno,
uma vez que suas moléculas se ligam fortemente ao tensoativo, alterando a estrutura
e viscosidade.
Normalmente, a cada molécula de tensoativo aniônico, encontra-se de 4 a 1 o moléculas de água ligadas ao seu núcleo polar. Em tensoativos catiônicos, esse
número se reduz para uma molécula de água (AIRES-BARROS, 1991 ).
Por outro lado, a água do centro micelar tem propriedades semelhantes às da
água, pois essa está livre e portanto não teve sua estrutura modificada; tal
característica é importante visto que a água do centro micelar entra em contato com
a substância de interesse (LUISI et ai., 1988; KADAM, 1986).
11

Revisão Bibliogrãfica
____ cauda Hidrofóbica
Figura 2.3 - Composição de uma Micela Reversa (Adaptado de SEOUD et ai., 1999).
2.3. 7 - Sistemas Mice/ares Reversos
No preparo de sistemas contendo tensoativo, solvente e água é necessário
definir as proporções entre os componentes de modo que ocorra a formação de
micelas reversas. Para tanto pode-se utilizar diagramas de fases específicos para
cada sistema. O diagrama de fases para os sistemas de micelas reversas são, no
geral, do tipo triangular, ou seja ternários, semelhantemente aos diagramas
tradicionais, onde cada componente ocupa um vértice do triângulo (no caso,
tensoativo, solvente e água). Os diagramas são compostos de diferentes regiões
(Figura 2.4): regiões de cristais líquidos hexagonais e lamelares, solução micelar
aquosa e solução micelar reversa (SEOUD et ai., 1999).
12

Revisão Bibliográfica
Tensoativo
Mi cela Aquosa
Solvente Cristal Líquido Hexagonal
Figura 2.4 - Estruturas que Formam o Sistema GenéricoTensoativo/Solvente/ H20 (Adaptado de SEOUD et ai., 1999).
A Figura 2.5 mostra o diagrama de fases para CTAB e AOT (TAMAMUSHI &
WATANABE, 1980), os quais são de grande importância na determinação das
proporções entre as substâncias que constituem as microemulsões utilizadas.
Através do diagrama de fases, é possível se identificar quantas e quais fases estão
presentes. A determinação de tais diagramas é difícil devido à existência de regiões
muito distintas e são portanto, dificilmente encontrados na literatura.
HEXANO!.
•
(I} (II)
Figura 2.5 - Diagramas de Fases dos Agentes Tensoativos CTAB (I) e AOT (li).
13

Revisão Bibliogrãfica
Após a determinação dos componentes responsáveis pela formação dos
sistemas micelares, deve-se definir as condições de extração, tais como: pH, força
iônica e temperatura. Existem três maneiras de se preparar os sistemas de micelas
reversas, sendo que a primeira delas consiste da adição de pequena quantidade de
solução aquosa contendo a proteína à solução contendo o solvente orgânico e o
tensoativo (microemulsão). Após a agitação, formar-se-á uma solução límpida de
micelas reversas contendo proteína. Na segunda (aplicável a proteínas de baixa
solubilidade ou insolúveis em água) usa-se a proteína na forma de pó, injetando-a
diretamente na microemulsão. Na terceira, denominada transferência de fases,
mistura-se volumes iguais de solução aquosa contendo a proteína e de
microemulsão, separa-se as fases por centrifugação. (RODRIGUES, et ai., 1999;
PESSOA JR, 1995; LUISI et ai. 1988).
2.4 - EXTRAÇÃO DE PROTEÍNAS EM SISTEMAS DE MICELAS REVERSAS
' - . ·-~ A extração de uma proteína de um caldo fermentado, visando sua separação
e purificação é feita em duas etapas. A primeira etapa, denominada extração, visa
transferir a proteína, que está na fase aquosa para a fase micelar; a segunda etapa
consiste da reextração da proteína para uma nova fase aquosa. A Figura 2.6 ilustra o
processo de extração descrito.
14

Revisão Bibliogrãfica
Figura 2.6 - Representação Esquemática do Processo de Extração em Sistemas Micelares Reversos.
A escolha do tensoativo (catiônico ou aniônico) é fundamental neste processo
de separação e purificação de proteínas. Para que o tensoativo exerça atração sobre
a proteína, é necessário que tenham cargas líquidas totais opostas. No que se refere
à proteína, a carga líquida total é determinada pelo pH da fase aquosa. Assim,
tensoativos aniônicos são indicados para processos de separação em que a proteína
desejada possua pi elevado, desta forma, a diferença entre o pi e pH, também será
alta (LUISI et ai., 1988).
Outro fator que merece atenção é a força iônica da fase aquosa. Esta não
deve ser elevada o suficiente para prejudicar a eletroneutralidade necessária para
que ocorram as interações eletrostáticas entre a proteína e o tensoativo (HATTON,
1987).
Assim, para que se obtenha condições, as mais favoráveis possíveis, para
extração de proteínas por micelas reversas, deve-se usar baixa força iônica, pH<pl
para tensoativos aniônicos e pH>pl para tensoativos catiônicos. No entanto, para a
etapa de reextração, a situação se inverte, uma vez que se deseja que a proteína
seja "expulsa" da micela. Portanto usa-se: alta força iônica, pH>pl para tensoativos
aniônicos e pH<pl para tensoativos catiônicos (PYLE, 1994).
15

Revisão Bibliográfica
2.4.1 - Mecanismo de Formação de Mice/as Reversas
O mecanismo de formação das micelas reversas não está totalmente
elucidado. No entanto, a teoria mais aceita descreve a formação da micela reversa
como um mecanismo cooperativo entre proteína e tensoativo: a interface entre as
duas fases (aquosa e micelar) se deforma em torno da proteína, formando a micela e
transferindo-a para a fase orgânica, conforme ilustrado na Figura 2.7 (DUNGAN, et ai.,
1991 ). Um dos fatores que causam a deformação da interface seria a interação
eletrostática provocada pela ação da força iônica e pH.
A Equação 2.4, da força eletrostática, auxilia no entendimento do mecanismo
de formação de micelas reversas:
Fe = 1t - % i, E2 Equação 2.4
Onde: Fe = força eletrostática 1t = pressão osmótica i, = constante dielétrica E2 = campo elétrico
Quanto mais distante a proteína estiver da interface micelar, maior será a
diferença de potencial entre elas, uma vez que a carga da proteína é oposta à do
tensoativo. Assim, será maior também o segundo termo da equação (% i, E2),
aumentando a força de atração e fazendo com que a proteína se aproxime da
interface (Figura 2. 7a). Devido à aproximação entre proteína e tensoativo, a
concentração de cargas opostas aumenta e com isso ocorre a elevação da pressão
osmótica do núcleo aquoso, que age em sentido contrário ao da força eletrostática
(Figura 2.7b). Este fenômeno faz com que haja ruptura da interface e,
consequentemente, deslocamento da micela reversa formada em direção à fase
orgânica (Figura 2. 7c). Desse modo, a superfície interfacial é atraída para junto da
proteína, fechando a micela reversa (Figura 2. 7d).
A tensão superficial juntamente com a força eletrostática, tendem a resistir às
deformações das camadas superficiais e portanto, a interface deforma até que a
tensão superficial e a força eletrostática fiquem equilibradas (DUNGAN et ai., 1991).
16

Revisão Bibliográfica
localizadas em contato com a cauda do tensoativo e até mesmo com o
solvente);
),,> Situação V - várias micelas reversas englobando diversas proteínas.
Figura 2.8 - Modelos de Proteínas Englobadas por Micelas Reversas (Adaptado de LUISI et ai., 1988).
2.4.3 - Fatores que Afetam a Extração de Proteínas
Existem alguns fatores que afetam a transferência da proteína da fase aquosa
para a pseudofase micelar e também sua reextração. Dentre eles, destacam-se: pH,
força iônica, tipo de íon, tipo da proteína, temperatura, tipo e concentração do
tensoativo, tamanho da micela e velocidade de agitação. Cada um dos fatores serão
discutidos a seguir.
2.4.3.1-pH
Esta variável determina a rede de cargas da proteína, influenciando o estado
de ionização dos grupos eletricamente carregados em sua superfície. A solubilização
de proteínas é favorecida por interações eletrostáticas entre os núcleos do tensoativo
e a superfície da proteína, que ocorrem quando a carga líquida total da proteína é
18

e,:,::. =· ~ ~ ~ ~ ~
Fase ~ Aquosa~ ~ e.::::
Fase Aquosa
Revisão Bibliográfica
Fase Aquosa -a
-e Fase
Aquosa
.,;<:;. ~ ~ «s»: ~~~
5~·.··~ -d e,:::; ~ · Micela· ~ . . ~ · Reversa .,;<:;. ~
Fase Orgânica Proteína
~ Tensoativo
-b
Figura 2. 7 - Mecanismo de Formação de Micelas Reversas (Adaptado de DUNGAN et ai., 1991 ).
2.4.2 - Modelos de Absorção de Biomoléculas
A localização da proteína no interior da micela reversa depende da natureza
da proteína e do tipo de tensoativo. Existem inúmeros possíveis modelos que visam
ilustrar a absorção de proteínas pelas micelas reversas. As diferentes situações são
descritas abaixo e ilustradas na Figura 2.8 (LUISI et ai., 1988):
),> Situação I - proteínas hidrofílicas estariam localizadas no centro
micelar;
),> Situação II - molécula de proteína ativa, pode interagir fortemente
com a parede da micela reversa;
),> Situação III - proteínas hidrofílicas localizadas no centro micelar
solubilizadas juntamente com outras proteínas contaminantes;
),> Situação IV - grande número de micelas reversas solubilizando uma
proteína com caráter dominantemente hidrofóbico (proteínas
17

Revisão Bibliográfica
oposta à carga do núcleo do tensoativo. Assim, quando se usa agente tensoativo
aniônico, em processos de extração líquido-líquido por micelas reversas, deve-se
trabalhar com valores de pH inferiores ao do ponto isoelétrico, pois desta forma a
proteína apresentará carga líquida positiva, condição que favorece a atração
eletrostática entre a proteína e o agente tensoativo.
De modo oposto, para tensoativos catiônicos, a transferência da proteína seria
favorecida em valores de pH acima do ponto isoelétrico da proteína. Segundo
ASENJO (1990), o pH de uma solução também afeta as características de
solubilização das proteínas, principalmente porque modifica a distribuição de cargas
na sua superfície, podendo ocorrer repulsão eletrostática e inibição da transferência
de proteínas para o interior da micela reversa (MEYER,1992).
No entanto, tal comportamento pode não ser observado. Por exemplo, PIRES
et ai. (1996), em trabalhos de extração de a-quimiotripsina (que possui pI = 8,3) e
pepsina (pi < 1, 1) utilizando em ambos os casos TOMAC (tensoativo catiônico),
observaram perfis de pH idênticos para ambas as proteínas. A explicação para tal
fato consiste na existência de íons em solução aquosa que exerceriam maior
influência que o pH (PIRES et ai., 1996, CASTRO & CABRAL, 1988, DEKKER et ai.,
1989).
Via de regra, para proteínas de elevada massa molar, a diferença entre o pH
da fase aquosa e o seu pi dever suficiente para obter uma transferência de massa
satisfatória. No entanto para proteínas de massa molar menores, não existe esta
necessidade, uma vez que serão solubilizadas mesmo quando o valor do pH estiver
próximo ao do pi da enzima. O pH influi na transferência da proteína em função do
sua massa molar pois, para proteínas de massas molares elevadas, são necessárias
micelas reversas maiores que a micela vazia, tornando o processo energeticamente
desfavorável. Para viabilizá-lo, deve-se favorecer as interações eletrostáticas ( entre
proteína e tensoativo) através do aumento da densidade de cargas da proteína, o
que ocorre com o aumento da diferença entre o pH e o pi. Assim, para proteínas
menores, o tamanho necessário da micela também é menor, portanto não há a
necessidade de uma diferença grande entre o pH e o pi.
19

Revisão Bibliogrãfica
No entanto, deve-se dar atenção especial ao valor de pH empregado, uma vez
que valores muito elevados podem levar à desnaturação da proteína. ANDREWS et
ai. (1994), verificaram que uma ou duas unidades de pH abaixo do pi é suficiente
para transferência de praticamente 100% da proteína. Porém em alguns casos,
podem existir intervalos de pH em que ocorre redução na solubilização da proteína
para o interior da micela, voltando a subir em seguida. Como exemplo, pode-se citar
BRANDANI et ai. (1993) que durante a extração de a-amilase (pi = 5,5) utilizando o
sistema TOMAC/isooctano/octanol, não observaram a solubilização da enzima em
valores de pH inferiores a 9. No entanto, no intervalo de pH entre 9,3 e 9,9 houve a
solubilização da a-amilase, provavelmente devido ao aumento de cargas negativas
na superfície da enzima. No intervalo de 9,9 a 10,5 foi observado um decréscimo na
solubilização causado, provavelmente, pela grande quantidade de íons OH- em
solução, o que ocasionou a redução da interação eletrostática entre a carga da
proteína e o núcleo do tensoativo. Ao aumentarem ainda mais o valor do pH, entre
11, 5 e 12,4, a transferência da a-amilase para as micelas reversas voltou a
aumentar.
2. 4. 3. 2 - Força iônica e tipo de íon
A transferência das proteínas para as micelas reversas depende das
interações eletrostáticas entre a proteína e o núcleo do tensoativo. Portanto, a força
iônica e o tipo de íon da fase aquosa são muito importantes, devido às cargas
presentes na solução (PYLE, 1994).
A força iônica, que pode ser expressa em termos de condutividade elétrica,
controla a solubilização de proteínas no sistema micelar, pois o alcance da força
eletrostática da micela depende da força iônica do meio. O aumento da concentração
de íons no meio ocasiona uma redução na eficiência de transferência de proteínas
pelas micela, seguida de um enfraquecimento das trocas eletrostáticas entre as
proteínas e as porções polares dos tensoativos, e conseqüente diminuição da
solubilidade das proteínas na micela. Também as enzimas de maior massa molar
20

Revisão Bibliográfica
presentes na micela tornam-se menos solúveis em decorrência do aumento da força
iônica (KREI & HUSTEDT, 1989; HA TTON, 1987).
ANDREWS et ai. (1994) verificaram que a força iônica pode afetar o tamanho
da micela, pois os grupamentos polares dos agentes tensoativos repelem-se, e com
isto micelas maiores são formadas quando submetidas à baixa força iônica. Este fato
está relacionado com a teoria de Debye (HATTON, 1987), segundo a qual a
capacidade de aproximação da proteína e dos grupamentos polares do tensoativo é
inversamente proporcional à raiz quadrada da força iônica do meio. Desta forma, à medida que a força iônica da solução que contém a proteína aumenta, as interações
entre a proteína e o agente tensoativo diminuem, devido à redução do efeito
repulsivo entre os grupos polares do tensoativo. Esse fato leva a um decréscimo do
transporte de água ou outras moléculas e, consequentemente, à formação de menos
e menores micelas reversas.
O tipo de íon pode afetar significativamente a transferência de proteínas para
o interior das micelas reversas. Em estudos visando analisar o efeito do tipo de íon
sobre a extração da ribonuclease A e da taumatina, ANDREWS & HA YWOOD
(1994), verificaram que na presença de íons maiores, como o K+. a solubilização da
proteína é diminuída, devido provavelmente à formação de um campo eletrostático
que reduz as interações entre a proteína e o tensoativo (o chamado "screening'). No
entanto, com íons menores como Na+. este efeito não é verificado. Ainda, ao
utilizarem MgCl2, verificaram um aumento na transferência da enzima, confirmando a
influência do tamanho do íon sobre a extração da enzima, pois o raio atômico do
Mg2+ é inferior ao dos dois cátions anteriores.
Também na etapa de reextração foi analisada a influência do tipo de íon
empregado. MARCOZZI et ai. (1991) verificaram que a adição de CaC'2 ao tampão
fosfato 20mM pH 7,5, torna a reextração de a-quimiotripsina mais eficiente que
quando usados os sais: NaCI, KCI e LiCI.
21

Revisão Bibliográfica
2.4.3.3 - Tipo de proteína
Outro fator de suma importância nos processos de extração em sistemas de
micelas reversas é a natureza da proteína, devido principalmente às interações entre
ela e o tensoativo. As diferentes interações entre eles podem sofrer alterações em
função do tipo de proteína, afetando até mormente a transferência da proteína que a
força iônica (CASTRO & CABRAL, 1988).
As interações podem até permitir a solubilização de proteínas maiores que o
espaço interno das micelas reversas, não sendo encontrados dados literários que
permitam definir o tamanho máximo da molécula a ser encapsulada por elas. O
caráter hidrofílico das proteínas de alta massa molar é muito relevante na formação
das micelas reversas, pois favorecem a formação de uma camada de tensoativo ao
redor da proteína devido à solubilização com a camada de água, possibilitando a
transferência de proteínas maiores, mesmo que sua carga líquida total seja igual a
do tensoativo (CASTRO & CABRAL, 1988).
Por outro lado, a proteína pode apresentar caráter hidrofóbico. Neste caso, as
interações predominantes são as hidrofóbicas. Normalmente, tais proteínas não
apresentam um comportamento tradicional, podendo ser extraídas em pH
desfavorável ou em força iônica elevada (PIRES et ai., 1996). Como por exemplo
pode-se citar a tau matina, que apresenta 100% de transferência pelo sistema
AOT/isooctano mesmo em valores de pH acima do pi e força iônica entre 0,3 e 0,8 M
(ANDREWS et ai., 1994).
Com relação a reextração de proteínas hidrofóbicas, podem surgir problemas
com a recuperação, pois o processo pode tornar-se irreversível. Nesse caso, o uso
de um álcool polar surge como alternativa para auxiliar no rompimento das
interações hidrofóbicas entre a proteína e o tensoativo (PIRES et ai., 1996).
2.4.3.4 - Temperatura
A temperatura também é um fator de importância na absorção de proteínas
pela micela, pois dela depende a capacidade máxima de absorção da micela. Dentro
de uma determinada faixa de temperatura, as micelas reversas são formações
22

Revisão Bibliogrãfica
estáveis, o que favorece a absorção de moléculas. Segundo LUISI et ai. (1988), a
presença de enzimas no interior da micela a temperaturas mais elevadas vai
depender mais da sua termoestabilidade do que da capacidade de absorção da
micela. A temperaturas inferiores a uma faixa ótima, as micelas encolhem e perdem
sua capacidade de absorção. Temperaturas elevadas afetam o coeficiente de
partição do agente tensoativo, de modo que, praticamente, este não permaneça mais
na fase orgânica e só seja encontrado na fase polar (KREI & HUSTEDT, 1989).
Também, segundo estes autores, a capacidade máxima da micela é dependente da
temperatura, ou seja, existe uma faixa de temperatura na qual esta apresenta-se
estável, favorecendo portanto a absorção de proteínas.
PESSOA JR & VITOLO (1998), em trabalhos de extração da enzima inulinase,
também constataram que o tamanho da micela reversa é bastante dependente da
temperatura e, na faixa de 5 a 45°C, as micelas reversas, em solventes orgânicos,
são formações estáveis e podem favorecer a absorção desta enzima.
2.4.3.5 - Tensoativo
O tipo de tensoativo influencia na transferência da proteína para o interior das
micelas reversas devido, principalmente às interações eletrostáticas entre o seu
núcleo e a proteína. Influi também no tamanho das micelas reversas e na energia
necessária para o encapsulamento de uma proteína de maior tamanho quando existe
uma força de atração ou repulsão elevada entre os núcleos do tensoativo (AIRES-
BARROS, 1991). Estes autores em estudos de extração de amiloglicosidase
utilizando o sistema micelar TOMAC/isooctano/1-octanol alcançaram resultados de
recuperação que variaram entre 20 e 30% após a reextração da enzima. No entanto,
quando o sistema foi substituído por CTAB/isooctano/hexanol/butanol, os resultados
chegaram a 90%.
Outro fator que afeta a recuperação de proteínas é a adição de um tensoativo
não iônico em um tensoativo iônico. Um exemplo é a partição da a-amilase onde foi
adicionado um tensoativo neutro ao TOMAC em isooctano (DEKKER et ai., 1989). Um
aumento na transferência da enzima foi observado por estes autores, com o aumento
23

Revisão Bibliográfica
na proporção do tensoativo neutro adicionado. Tal comportamento é verificado
devido à influência deste tensoativo na densidade de cargas das micelas reversas,
aumenta-se a concentração de tensoativo no meio sem que se promova aumento de
cargas, ocorrendo ainda mudanças no tamanho das micelas reversas pela adição
das moléculas do tensoativo em sua estrutura.
Também a concentração do tensoativo é um fator importante no uso de
sistemas de micelas reversas, por influenciar a formação destas. Existe uma
concentração mínima de tensoativo necessária para formar as micelas reversas,
chamada concentração micelar crítica (CMC) e que depende de fatores como:
temperatura, pressão, solvente e estrutura química do tensoativo. Os valores de
CMC podem ser obtidos através das propriedades do sistema, tais como: tensão
superficial, turbidez e pressão osmótica (SEOUD et ai., 1999). No entanto, devido não
apresentarem precisão, são normalmente expressos através de intervalos (CASTRO
& CABRAL, 1988; KADAM, 1986). A Figura 2.9, ilustra a evolução do sistema formado
por tensoativo e solventes (ROH), levando à formação de sistemas de micelas
reversas.
24

Revisão Bibliogrãfica
Mice la Reversa
Micela
Cristal Líquido Reverso
Tensoativo
Tensoativo
Cristal Líquido Lamelar
Tensoativo
> Cristal Líquido Cúbico
Cristal Líquido Hexagonal
Figura 2.9 - Evolução da Formação de Estruturas no Sistema de Micelas Reversas (Adaptado de SEOUD et ai., 1999).
25

Revisão Bibliográfica
O aumento da concentração de tensoativo provoca o aumento no tamanho
das micelas, aumentando a transferência de proteínas para o interior destas. Estudos
de extração de a-amilase tiveram os resultados de recuperação melhorados com o
aumento da concentração de tensoativo (TOMAC) de 0,01 para 0,04 M. Porém uma
outra hipótese é levantada de que o aumento da concentração de tensoativo altera o
número e não o tamanho das micelas existentes (BRANDANI et ai., 1993; AIRES-
BARROS, 1991 ). Existe no entanto, um limite para a concentração de tensoativo no
sistema; concentrações muito elevadas dificultam a etapa de reextração da proteína,
pois fortes interações eletrostáticas entre a proteína e o tensoativo provocam
redução nos valores de recuperação.
2.4.3.6- Tamanho das mice/as
O parâmetro que define o tamanho das micelas reversas e as propriedades
físico-químicas da água solubilizada no seu interior é o Wo ("water in oil"), dado este,
importante para o estudo da biocatálise em micelas reversas. Esta variável que
exprime a relação entre a concentração molar de água e a concentração de --
tensoativo (LUISI et ai., 1988) é de suma importância no sistema, pois determina a
estrutura, o tamanho das micelas reversas e também o número de moléculas de
tensoativo por micela reversa (CASTRO & CABRAL, 1988; AIRES-BARROS, 1991 ).
Mudanças no valor de Wo podem influenciar a partição da proteína, uma vez
que a quantidade de água no interior das micelas reversas interfere nas interações
eletrostáticas entre proteína e tensoativo. Além disso, afeta a estabilidade da
proteína quando a quantidade de água é pequena, pois as propriedades e
densidades de cargas desta água sofrerão alterações, podendo ocasionar
modificações na proteína e influindo em seu desempenho (KRIEGER, 1995; KADAM,
1986).
O valor do Wo pode ser maior, quanto maior for a concentração de proteínas,
pois a quantidade de proteína extraída é proporcional à quantidade de água
solubilizada. Se o valor de Wo diminui, a fração de micelas reversas menores que a
proteína aumenta, o que minimiza a solubilização das moléculas de proteína. Com o
26

Revisão Bibliográfica
aumento de W0 essa fração aumenta, e com isso o sistema é capaz de armazenar
proteínas maiores (ISHIKAWA et ai., 1992).
2.4.3. 7 - Influência da velocidade de agitação
A velocidade de agitação pode afetar a separação da proteína, pois interfere
no transporte de massa e nas resistências existentes na interface entre as fases
aquosa e orgânica. Quando a proteína está se movimentando da fase aquosa para a
micelar, ela encontra a resistência difusional, associada ao transporte do soluto de
dentro da solução para a interface e vice-versa. Com o aumento da velocidade de
agitação entre as duas fases, ocorre a diminuição drástica da resistência ao
englobamento da proteína devido à redução da camada limite entre as duas regiões.
Quando o valor do pH e a concentração de sais são elevados, a partição da proteína
independe da velocidade de agitação. Nessas condições a força iônica predomina
sobre a resistência na interface (DUNGAN et ai., 1991 ).
2.5 - ENZIMAS XILANOLÍTICAS
As xilanases têm como substratos naturais as hemiceluloses, geralmente
localizada na lamela média das plantas em toda a extensão da fibra, intimamente
associadas à celulose (SUNNA & ANTRANIKIAN, 1997; DEKKER, 1985; WOODWARD,
1984; JANES, 1969). A forma estrutural de hemicelulose mais comumente encontrada
na parede celular de plantas terrestres é a xilana, representando mais de 30% em
peso seco (JOSELEAU et ai., 1992). As xilanas são compostas de resíduos de ligações
13(1-4)-D-xilopiranosídicas, sendo que a maioria se apresenta na forma de
heteropolissacarídeos, contendo diferentes grupos substituintes nas cadeias centrais
e laterais. Os substituintes mais comuns encontrados na cadeia central das xilanas
são os grupos acetil, arabinosil e glucuronosil. Homoxilanas, por outro lado,
consistem exclusivamente de resíduos de xilosil e não são muito comuns na
natureza, mas foram isoladas de gramíneas e talos de tabaco (PULS & POUNTANEN,
1989; BIELY, 1985).
27

Revisão Bibliográfica
A poluição do meio ambiente tornou-se assunto de grande interesse público
em todo o mundo, pois tanto países desenvolvidos quanto aqueles em
desenvolvimento vêm sendo afetados por problemas ambientais resultantes do
crescimento econômico acelerado, em conseqüência da exploração dos recursos
naturais. É inegável a urgência de se adotar medidas de prevenção e controle da
poluição, a fim de se evitar danos cada vez maiores e, possivelmente irreparáveis.
Tais medidas freqüentemente são definidas em termos da melhor tecnologia
disponível, cuja determinação depende de dois fatores essenciais: informação
disponível e condições econômicas. Dentro deste contexto, as enzimas xilanolíticas
exercem papel de destaque.
2.5.1 - p.xilosidase
As p-xilosidases (E.C. 3.2.1.3. 7) são responsáveis pela hidrólise de xilobiose
liberada pela ação das endo-xilanases sobre a xilana, produzindo xilose. A xilose é o
principal açúcar obtido da hidrólise ácida ou enzimática da xilana e é fermentescível
à etanol por várias linhagens de leveduras (JEFFRIES, 1983). A atividade de P- xilosidase gera D-xilose de oligossacarídeos de cadeias curtas e xilobiose.
Desempenha um papel crucial na liberação do produto final de inibição da atividade
das endo-xilanases por xilobiose (RIELLY, 1981). A importância das p-xilosidases no
auxílio à hidrólise da xilana tem sido mostrada como sendo análoga a das celobioses
na hidrólise da celulose (DEKKER & RICHARDS, 1976).
De forma geral, enzimas degradadoras de xilana produzidas por fungos, têm
recebido maior atenção que as produzidas por bactérias, sendo que inúmeras p-
xilosidase extracelulares fúngicas têm sido parcialmente ou totalmente
caracterizadas (CORTEZ, 1997; MILAGRES, 1994; UZIIE, et a/., 1985; MATSUO & YASUI,
1984), desta forma, o desenvolvimento do presente trabalho, pôde ser alicerçado em
informações sobre a enzima, tornando o estudo da técnica de recuperação mais
confiável.
29

Revisão Bibliográfica
2.5.1.1 - pH ótimo
As J3-xilosidases podem apresentar uma ampla faixa de pH ótimo de atividade,
dependendo da origem. De forma geral apresentam atividade dentro da faixa de pH
de 3,0 a 7,0; embora a J3-xilosidase de uma linhagem termofílica da bactéria
Thermonospora sp tenha sido descrita por exibir uma faixa de pH ótimo de atividade
que vai de 5,0 à 9,0 (RISTROPH & HUMPHREY, 1985) e a J3-xilosidase da levedura
Aureobasidium pul/ulans (DOBBERSTEIN & EMEIS, 1978) tenha apresentado uma
faixa mais ampla de atividade (2,0 à 9,0). As í3-xilosidases de Neurospora crassa
(DESHPANDE et ai., 1994), Trichoderma reesei (PULS & POUNTANEN, 1989},
Emericella nidulans (MATSUO & YASUI, 1984) possuem pH ótimo de atividade
próximo de 4,0.
2.5.1.2 - Temperatura ótima
As í3-xilosidases podem ter temperatura ótima de atividade na faixa que vai de
25°C a 80°C (RUTTERSMITH & DANIEL, 1993; KERSTERS-HILDERSON et ei., 1969).
º Linhagens de Thermonospora têm atraído atenção considerável como fonte de
enzimas altamente termoestáveis para sacarificação de lignocelulose em geral
(BACHMANN & McCARTHY, 1989).
-: -· - -_..,.,
30

Objetivos
3. OBJETIVOS
O presente trabalho de dissertação teve como objetivo principal o estudo da
técnica de extração de enzimas por micelas reversas. Para tanto foram utilizados três
tensoativos no preparo dos sistemas micelares: os agentes tensoativos catiônicos
BDBAC e CTAB e o agente tensoativo aniônico AOT. A enzima alvo, (3-xilosidase, foi
produzida pelo fungo Penicillium janthinellum (através de processo fermentativo, sob
condições estabelecidas anteriormente ao presente trabalho) em hidrolisado
hemicelulósico de bagaço de cana-de-açúcar. Os seguintes objetivos específicos
foram cumpridos durante a execução deste projeto:
a) estudar da estabilidade da f3-xilosidase frente às variações de temperatura
(-4° e 4°C) e pH (3,0 a 9,0), determinando desta forma os limites que
poderiam ser usados nos ensaios de extração;
b) determinar os fatores (pH, temperatura, condutividade elétrica,
concentração e tipo do agente tensoativo, concentração de co-solvente)
que favorecem a extração da f3-xilosidase da fase aquosa para a fase
micelar, com o auxílio de planejamento estatístico específico para cada
tensoativo testado;
c) determinar os parâmetros que se mostrarem efeito significativo no processo
de extração líquido-líquido por micela reversa da f3-xilosidase, para os três
31

Objetivos
agentes tensoativos testados (BDBAC, AOT e CTAB) e avaliar dos
rendimentos obtidos;
d) otimizar o processo, através da determinação do modelo estatístico que
descreve o processo, teste do ponto ótimo e construção de superfícies de
resposta;
e) extrair da enzima í3-xilosidase diretamente do meio fermentado, sem pré
purificação por etanol, utilizando-se para isto as condições otimizadas do
planejamento que apresentou melhores valores de recuperação para
enzima.
32

Materiais & Métodos
4 · MATERIAIS E MÉTODOS
4.1 - MICRORGANISMO
O microrganismo utilizado no presente trabalho foi o fungo
Penicilliumjanthinellum CRC 87M-115, isolado de madeira em decomposição em
Lorena-SP por MILAGRES (1988).
4.2 - MEIO DE MANUTENÇÃO DO FUNGO
O meio de cultura para obtenção de esporos de P. janthinellum foi o mesmo
usado por MILAGRES (1988): 2% de glicose, 0,25% de extrato de levedura, 2% (v/v)
de solução mineral (VOGEL, 1956) e 2% de ágar-ágar.
Preparada a mistura, a mesma foi dividida em porções de 5 ml e colocada em
tubos de ensaio devidamente arrolhados os quais foram autoclavados por 15 minutos
a 112ºC e deixados em superfície inclinada (cerca de 20°). Após resfriamento, os
tubos foram inoculados com esporos de P. janthinellum de um tubo matriz e
incubados em estufa termostatizada a 30 ºC.
33

Materiais & Métodos
4.3 - HIDROLISADO HEMICELULÔSICO DE BAGAÇO DE CANA-DE-AÇÚCAR
Para produção do hidrolisado foi utilizado bagaço de cana-de-açúcar
proveniente da Usina Nova América (Tarumã-SP). O procedimento de hidrólise foi
realizado conforme descrito por MILAGRES & LACIS (1991).
Foram utilizados 46 mg de H2S04 p.a. por grama de matéria seca, com uma
relação de matéria seca: solução ácida de 1: 1 O. O bagaço foi seco a 80 ºC até ser
atingida a umidade de 10%, passando em seguida por um moinho com peneira de 20
"mesh" (aproximadamente 1,27 mm de diâmetro). Posteriormente foi tratado com
ácido (20 mL de ácido sulfúrico p.a. e 8 L de água para 800 g de bagaço) e
autoclavado a 121 ºC por 45 minutos. A fração líquida, foi recuperada por filtração
em papel de filtro qualitativo (Klabin) e sendo o pH elevado para 5,5 com NaOH 1 N.
4.4 - PRODUÇÃO DA ENZIMA l3-XILOSIDASE
A partir do hidrolisado hemicelulósico, foi preparado um meio para o cultivo de
P. janthinellum, adicionando-se ao mesmo solução mineral de Vogel (2% v/v) e
extrato de levedura (O, 1%). A obtenção do complexo xilanolítico (no qual está
presente a í3-Xilosidase), se deu através do cultivo de P. Janthinellum em meio à
base de hidrolisado hemicelulósico, obtido como descrito no item 4.3. Os frascos
utilizados no processo fermentativo (Erlenmeyers de 125 mL contendo 25 mL de
meio) foram autoclavados por 15 minutos a 112ºC e posteriormente inoculados com
1 mL de solução de esporos em água autoclavada nas mesmas condições (105
esporos/mL) e incubados a 30°C sob agitação orbital de 60 rpm, por 5 dias.
4.5 - AVALIAÇÃO DA ATIVIDADE ENZIMÁTICA
4.5.1 -Xilanases
A atividade de xilanase extracelular foi determinada pela quantidade de
açúcares redutores liberados a partir de xilana, de acordo com o método de BAILEY
34

Materiais & Métodos
et ai. (1992). Os açúcares redutores totais (ART) foram dosados pelo método do
ácido 3, 5-dinitrosalicílico (DNS) (MILLER, 1959).
Uma mistura de reação contendo 900 µL de xilana e 100 µL do meio
fermentado devidamente diluído em tampão acetato de sódio (50 mM, pH 5,5) foi
incubada a 50 ºC por 5 min. A reação foi interrompida pela adição de 1,5 ml de ácido
3,5-dinitrosalicílico (DNS) e a mistura foi aquecida em banho de água fervente por
5 min. A absorbância foi medida em espectrofotômetro, a 570 nm, e comparada com
uma curva padrão. O branco para leitura foi preparado segundo a mesma
metodologia, sendo que o meio fermentado foi adicionado ao final, após os demais
reagentes.
Uma unidade de atividade enzimática foi definida como a quantidade de
enzima necessária para liberar 1 µmolde ART, expressos como xilose, por minuto, a
50°C.
4.5.2 - {J-xilosidase
A atividade da f3-Xilosidase foi determinada através da quantidade de
p-nitrofenol liberada após a ação da enzima numa solução de p-nitrofenil-f3-D-
xilopiranosídeo (pNpX) em água (KUMAR & RAMÓN, 1996).
Uma mistura de reação contendo 250 µL de solução 2 mM de pNpX em água
e 250 µL do meio fermentado, devidamente diluído em tampão acetato de sódio (50
mM, pH 5,5), foi incubada a SOºC por 30 min. A reação foi interrompida pela adição
de 1 ml de solução de carbonato de sódio 2 M. A absorbância deste meio foi medida
em espectrofotômetro, a 400 nm, e comparada com uma curva padrão. O branco foi
preparado segundo a mesma metodologia, sendo que o meio fermentado foi
adicionado ao final, após os demais reagentes.
Uma unidade de atividade de 13-Xilosidase foi definida como a quantidade de
enzima necessária para liberar 1 µmol de p-nitrofenol/min. ml, nas condições
descritas anteriormente.
35

Materiais & Métodos
4.6 - PRECIPITAÇÃO PELA ADIÇÃO FRACIONADA DE ETANOL
A precipitação das proteínas do meio fermentado foi realizada segundo as
condições estabelecidas por CORTEZ & PESSOA JR (1998). O meio, previamente
tamponado (na proporção de 9 partes de meio para 1 de tampão acetato 1 M, pH
4,0), foi colocado em um becker de 25 ml o qual, juntamente com um tubo de ensaio
contendo etanol, foi levado a um banho termostatizado (Marca Heto, modelo CB 15-
25) de água e monoetilenoglicol à temperatura de -4 ºC, e deixado para a
estabilização da temperatura. Este procedimento evita que a temperatura ultrapasse
4 ºC quando da adição de etanol ao meio fermentado. Através de bomba peristáltica,
em volume de etanol correspondente a 20% (v/v) do meio foi transferido lentamente
para o becker contendo o meio fermentado. A temperatura foi conferida
constantemente e a agitação executada manualmente com bastão de vidro. Após a
adição completa do etanol, a mistura foi mantida em repouso sob refrigeração (a -
4 ºC) por 15 minutos, e então, centrifugada a 2000xg por 15 min, a uma temperatura
de o ºC (± 2 ºC). Após a centrifugação, o sobrenadante foi levado novamente ao
banho termostatizado, nas mesmas condições e, também através de bomba
peristáltica, um volume de etanol correspondente a 60% (v/v) do meio foi adicionado
lentamente a este. Nesta etapa também se monitorou a temperatura e se agitou a
mistura. Novamente após a adição do etanol, a mistura foi mantida em repouso sob
refrigeração (-4 ºC) por 15 minutos, e centrifugada nas mesmas condições da
primeira etapa. Após a centrifugação o precipitado (contendo í3-Xilosidase) foi
separado do sobrenadante e solubilizado em tampão adequado, conforme o pH a ser
estudado.
4.7 - DETERMINAÇÃO DOS PARÂMETROS CINÉTICOS
A determinação dos parâmetros cinéticos aparentes: kM (app) (constante de
Michaelis/Menten) e Vm (velocidade máxima da reação enzimática) foi feita através
da medida da atividade enzimática (item 4.5.2) da í3-xilosidase precipitada com
etanol (item 4.6), utilizando-se as seguintes concentrações de substrato (pNpX):
0,25; 0,5; 1,0; 1,5; 2,0; 2,5; 3;0; 4,0; 6,0; 8,0; 10,0; 15,0 e 20,0 mM. Os valores de
36

Materiais & Métodos
kM (app) e Vm foram determinados pelo método de Lineweaver-Burk (LEHNINGER,
1976).
4.8 - DETERMINAÇÃO DA ESTABILIDADE À ESTOCAGEM DA ENZIMA J3-XILOSIDASE
A determinação da estabilidade da enzima !3-xilosidase em função do tempo,
foi feita para diferentes valores de pH nas temperaturas de estocagem de -4 ºC e
4 ºC. Foi feito o acompanhamento da atividade enzimática, após a precipitação com
etanol e solubilização em tampão de pH desejado (item 4.6): acetato de sódio para
pH de 3,0, fosfato de potássio para pH 6,0 e 7,0 e Tris-HCI para pH 9,0.
4.9 - EXTRAÇÃO LÍQUIDO-LÍQUIDO POR MICELAS REVERSAS
A enzima J3-xilosidase foi extraída do meio fermentado previamente
precipitado por etanol, por micelas reversas de BDBAC, AOT e CTAB. O processo foi
conduzido em duas etapas. como mostrado nas Figuras 4.1 e 4.2. O pH da fase
aquosa foi definido pelo tampão (de pH apropriado) utilizado na solubilização da
amostra após a precipitação. Nesta etapa, denominada extração, a solução aquosa
contendo !3-xilosidase foi misturada a microemulsão em partes iguais (5ml). Após
agitação em agitador tipo vórtice por 1 min, a mistura foi centrifugada (Centrifuga
Jouan Mod.1812, Saint Herblain) a 1677xg por 1 O min para separação das fases
(FMI e FAI). Para a segunda etapa, a fase superior do tubo (FMI) foi retirada e
utilizada nos ensaios de reextração. Tal etapa foi realizada misturando-se 2,0 mL da
pseudofase micelar (FMI) com 2,0 mL de tampão acetato de sódio 1,0M pH 5,5
adicionado de NaCI (de modo a se obter solução final 1,0 M do mesmo), com a
finalidade de transferir (reextrair) a enzima da micela para a nova fase aquosa (FAII).
Na segunda etapa, a amostra foi novamente agitada e centrifugada nas condições
anteriormente citadas. Os resultados da extração foram expressos em termos da
atividade total recuperada (%) da enzima, tendo como referencial a quantidade de
proteínas do precipitado solubilizado no valor de pH estudado.
37
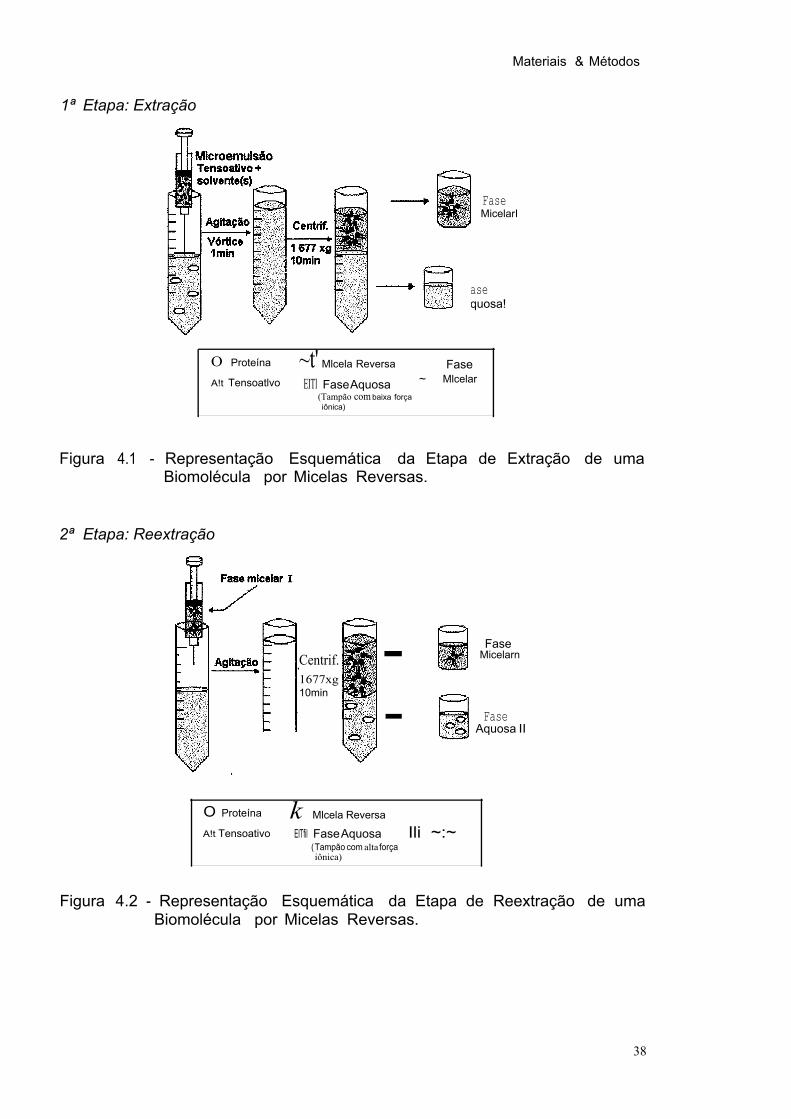
Materiais & Métodos
1ª Etapa: Extração
Fase MicelarI
Fase Aquosa!
O Proteína
A!t Tensoatlvo
~t' Mlcela Reversa
EITl Fase Aquosa (Tampão com baixa força iônica)
Fase ~ Mlcelar
Figura 4.1 - Representação Esquemática da Etapa de Extração de uma Biomolécula por Micelas Reversas.
2ª Etapa: Reextração
Vórtice 1mln
Centrif. 1677xg 10min
- Fase Micelarn
- Fase Aquosa II
~-./-..: --Tampão+ NaCI
O Proteína
A!t Tensoativo
k Mlcela Reversa
EITfil Fase Aquosa Ili ~:~ ( Tampão com alta força iônica)
Figura 4.2 - Representação Esquemática da Etapa de Reextração de uma Biomolécula por Micelas Reversas.
38

Materiais & Métodos
4.10 - DELINEAMENTO EXPERIMENTAL
Foram utilizados planejamentos estatísticos específicos (BOX et ai., 1978),
para cada tensoativo estudado.
As variáveis estudadas no planejamento foram codificadas em -1 , O e +1 e
seus valores reais constam da Tabela 4.1. Os ensaios foram realizados em ordem
aleatória, de forma a minimizar a ocorrência de erros sistemáticos.
Tabela 4.1 - Valores das Variáveis Empregados nos Planejamentos para Recuperação da 13-Xilosidase por Micelas Reversas
Tensoativo Usado Variável Estudada Valores -1 o +1
pH 3,0 6,0 9,0 Condut. Elétrica (mS/cm) 4,0 7,0 10,0
BDBAC Concentração de BDBAC (M) O, 1 O, 15 0,2 Concentração de Hexanol (%) 5,0 7,5 10,0 Temperatura (ºC) 4 15 26
.. - -_.,.,
pH 3,0 6,0 9,0
AOT Condut. Elétrica (mS/cm) 4,0 7,0 10,0 Concentração de AOT (M) O, 1 0, 15 0,2 Temperatura (ºC) 4 15 26
pH 3,0 5,5 8,0 Concentração de CT AB (M) O, 1 O, 15 0,2
CTAB Condut. Elétrica (mS/cm) 4,0 7,0 10,0 Concentração de Hexanol (%) 5,0 7,5 10,0 Concentração de Butanol (%) 10 15 20
Os planejamentos utilizados para verificar a influência das variáveis pH,
condutividade elétrica, concentração de tensoativo, concentração de co-solvente e
temperatura, sobre a recuperação da enzima 13-Xilosidase, foram os seguintes:
39

Materiais & Métodos
a) Para o agente tensoativo BDBAC: utilizou-se um planejamento
fatorial fracionário 25-1 com repetição, conforme mostrado na
Tabela 4.2;
b) Para o agente tensoativo AOT utilizou-se um planejamento fatorial
completo 24 com 4 repetições no ponto central, conforme mostrado na
Tabela 4.3;
c) Para o agente tensoativo CTAB: utilizou-se um planejamento
fatorial fracionário 25-1 com
Tabela 4.4;
repetição, conforme mostrado na
Tabela 4.2 - Matriz do Planejamento Fatorial Fracionãrio 25-1 com 01 Repetição Utilizada para Avaliação da Influência do pH, da Condutividade Elétrica, da Concentração de BDBAC, da Concentração de Hexanol e da Temperatura na Recuperação da (3-xilosidase por Micelas Reversas do Agente Tensoativo Catiônico BDBAC
Ensaio Variãveis Codificadas
n. pH Condutividade BDBAC Hexanol Temperatura Elétrica (mS/cm) (M) (%) (ºC)
1 -1 -1 -1 -1 +1 2 +1 -1 -1 -1 -1 3 -1 +1 -1 -1 -1 4 +1 +1 -1 -1 +1 5 -1 -1 +1 -1 -1 6 +1 -1 +1 -1 +1 7 -1 +1 +1 -1 +1 8 +1 +1 +1 -1 -1 9 -1 -1 -1 +1 -1 10 +1 -1 -1 +1 +1 11 -1 +1 -1 +1 +1 12 +1 +1 -1 +1 -1 13 -1 -1 +1 +1 +1 14 +1 -1 +1 +1 -1 15 -1 +1 +1 +1 -1 16 +1 +1 +1 +1 +1
40

Materiais & Métodos
Tabela 4.3 - Matriz do Planejamento Fatorial Completo 24 com 03 Repetições no Ponto Central Utilizada para Avaliação da Influência do pH, Condutividade Elétrica, Concentração de AOT e Temperatura na Recuperação da 13-Xilosidase por Micelas Reversas do Agente Tensoativo Aniônico AOT
Ensaio Variáveis Codificadas
n. pH Condutividade AOT Temperatura Elétrica (mS/cm) (M) (ºC)
1 -1 -1 -1 -1 2 +1 -1 -1 -1 3 -1 +1 -1 -1 4 +1 +1 -1 -1 5 -1 -1 +1 -1 6 +1 -1 +1 -1 7 -1 +1 +1 -1 8 +1 +1 +1 -1 9 -1 -1 -1 +1 10 +1 -1 -1 +1 11 -1 +1 -1 +1 12 +1 +1 -1 +1 13 -1 -1 +1 +1 14 +1 -1 +1 +1 15 -1 +1 +1 +1 16 +1 +1 +1 +1 17 o o o o 18 o o o o 19 o o o o
41

Materiais & Métodos
Tabela 4.4 - Matriz do Planejamento Fatorial Fracionário 25•1 com 03 Repetições no Ponto Central Utilizada para Avaliação da Influência do pH, da Condutividade Elétrica, da Concentração de CT AB, da Concentração de Hexanol e da Concentração de Butanol na Recuperação da f3-xilosidase por Micelas Reversas do Agente Tensoativo Catiônico CTAB
Ensaio Variáveis Codificadas
n. pH CTAB Condutividade Hexanol Butanol (M) Elétrica (mS/cm) (%) (%)
1 -1 -1 -1 -1 +1 2 +1 -1 -1 -1 -1 3 -1 +1 -1 -1 -1 4 +1 +1 -1 -1 +1 5 -1 -1 +1 -1 -1 6 +1 -1 +1 -1 +1 7 -1 +1 +1 -1 +1 8 +1 +1 +1 -1 -1 9 -1 -1 -1 +1 -1 10 +1 -1 -1 +1 +1 11 -1 +1 -1 +1 +1 12 +1 +1 -1 +1 -1 13 -1 -1 +1 +1 +1 14 +1 -1 +1 +1 -1 15 -1 +1 +1 +1 -1 16 +1 +1 +1 +1 +1 17 o o o o o 18 o o o o o 19 o o o o o
42

Materiais & Métodos
Os dados experimentais foram analisados estatisticamente, de acordo com o
planejamento estabelecido, para verificar o efeito dos fatores em estudo. A análise
dos dados foi feita com o auxílio de "software" específico para este fim, utilizando
como variável resposta a porcentagem de recuperação da enzima 13-Xilosidase.
Os modelos foram determinados por regressão linear pelo método dos
mínimos quadrados, representados pela Equação 4.1:
Equação 4.1
onde: Y; = variável resposta (recuperação da enzima),
bo, b, b;;, bij os coeficientes da regressão;
X; e Xj as variáveis independentes estudadas.
4.11 - DETERMINAÇÃO DO RAIO MICELAR - W0
A determinação do valor de Wo (conteúdo de água) da fase orgânica foi feita
por titulação, através da reação de Karl-Fisher (WIELAND, 1987), utilizando-se um
titulador automático Metler DL 18. A reação ocorreu em metanol (isento de água),
utilizando uma solução de dióxido de enxofre, iodo e piridina (reagente de Karl-
Fisher) para titular a água do sistema. Esta reação pode ser representada por:
A detecção do ponto final da titulação baseia-se nas diferenças de
condutividade entre a espécie r e 12. Após a titulação de toda a água presente no
sistema, ocorre a acumulo de li, provocando uma queda de tensão entre os
terminais do eletrodo.
43

Materiais & Métodos
4.11 • ENSAIOS ELETROFORÉTICOS
Os ensaios eletroforéticos foram realizados no equipamento Phast System
(PHARMACIA®) do Laboratório de Enzimologia Industrial, do Departamento de
Tecnologia Bioquímico-Farmacêutica da Faculdade de Ciências Farmacêuticas da
USP.
Para a eliminação de prováveis problemas decorrentes do uso de amostras
com elevada concentração salina e interferência de peptídeos de baixa massa molar
nos ensaios eletroforéticos, bem como aumentar a concentração de proteínas, as
amostras que foram utilizadas na realização das eletroforeses passaram por um
processo de tratamento que consistiu de duas etapas: diálise e liofilização.
4.11.1- Preparo das Amostras
4.11.1.1- Diálise
Com o intuito de se eliminar os efeitos causados pela elevada concentração
de sais e peptídeos de baixa massa molar nos ensaios eletroforéticos, optou-se por
dialisar as amostras que seriam posteriormente utilizadas nestes ensaios.
Utilizou-se para realização da diálise, membranas de 12k Da de corte
(SIGMA), pois em trabalhos anteriores, não se identificou a presença de proteínas de
massa molar inferior a 14k Da (RODRIGUES, 1997). Desta forma eliminou-se os
íons presentes, bem como prováveis peptídeos, sem no entanto provocar perdas de
proteínas.
As amostras colocadas nos "sacos" de diálise imersos em frasco (becker) com
água bidestilada, foram deixadas em ambiente de temperatura controlada a -4 ºC
(câmara fria) sob agitação (agitador magnético), por 48 horas.
4. 11. 1. 2 - Liofilização
As diferentes frações previamente dialisadas (precipitado solubilizado, fases
aquosas I e II) passaram por um processo de liofilização com o intuito de se
44

Materiais & Métodos
aumentar a concentração de proteínas nas frações para posterior utilização nos
ensaios eletroforéticos.
Para a condução do processo de liofilização, volumes conhecidos das frações,
foram levados ao "biofreezer" a uma temperatura de -70 ºC (Forma Scientific lnc. -
Ohio/USA) até completo congelamento. A seguir, as amostras foram liofilizadas em
liofilizador (L YOLAB-G) por 48 horas ou até completa sublimação da porção líquida;
obtendo-se assim, a máxima concentração possível das amostras.
4.11.2- Condução dos Ensaios eletroforéticos
As eletroforeses foram realizadas no aparelho Phast System®
(PHARMACIA®) (WEBER et ai. ,1972). Utilizou-se gel de poliacrilamida a 12,5%,
fornecido pela PHARMACIA®, sendo a corrida eletroforética conduzida a 15ºC,
utilizando uma mistura contendo tampão tricina 0,2 M e tampão tris/HCI 0,2 M com
pH 8, 1 e SOS 0,55%.
A coloração dos géis foi feita automaticamente em uma câmara acoplada ao
equipamento de corrida eletroforética. Na Tabela 4.5, encontra-se as soluções, o
tempo de exposição dos géis às diferentes soluções e à temperatura, para coloração
por prata.
45

Materiais & Métodos
Tabela 4.5 - Seqüência usada para desenvolvimento do processo otimizado de coloração de géis SDS-page por Prata
TCG* Temperatura Função da Solução Etapa Solução
(min) (ºC) sobre os géis
1 10% etanol, 5% 2,0 50 Lavagem inicial dos géis ácido Acético
2 10% etanol, 5% 4,0 50 Lavagem inicial dos géis ácido Acético
3 5% glutaraldeído 6,0 50 Aumentar a sensiblidade das proteínas
4 10% etanol, 5% 3,0 50 Lavagem dos resíduos ácido Acético da etapa anterior
5 10% etanol, 5% 5,0 50 Lavagem dos resíduos ácido Acético da etapa anterior
6 Água deionizada 2,0 50 Lavagem dos resíduos da etapa anterior
7 Água deionizada 2,0 50 Lavagem dos resíduos da etapa anterior
8 0,4% nitrato de 6,5 40 Coloração das bandas prata nos géis
9 Água deíonizada 0,5 30 Lavagem dos resíduos da coloração
10 Água deionizada 0,5 30 Lavagem dos resíduos da coloração
11 Reveladora 0,5 30 Revelação dos géis
12 Reveladora 4,0 30 Revelação dos géis
13 Redutora 2,0 30 Diminuição de rastros deixados por peptídeos
14 10% glicerol 5,0 50 Preservação/interrupção
* tempo de contato da solução com o gel
Para o preparo da solução reveladora do gel, adicionou-se o conteúdo de uma
ampola (PHARMACIA®) contendo solução de formaldeído 2% a 150mL de carbonato
de sódio a 2,5 %. A solução redutora, por sua vez, foi preparada pela dissolução de
1,6 g de tiossulfato de sódio e 3,7 g de Trisma base em 100 mL de água deionizada.
46

Materiais & Métodos
4.12 - DOSAGEM DO TEOR TOTAL DE PROTEÍNAS- MÉTODO DE LOWRY
O teor de proteínas foi determinado de acordo com o método de LOWRY
(1959). Foi utilizado como padrão a albumina de soro bovino (BSA -"bovine serum
albumine"). Visando a identificação de prováveis interferências na determinação do
teor de proteínas no meio, foi realizado um ensaio em branco para proteínas, sendo
adicionado ao meio reacional, em substituição à amostra contendo enzima, uma
solução tampão com características de pH e de condutividade elétrica idênticas a da
amostra estudada.
4.13 - ANÁLISE DOS RESULTADOS
Para avaliação dos processos de extração foram analisados os seguintes
parâmetros: Ae, AP, np, 11A , descritas pelas equações abaixo:
a) Aumento de atividade específica da enzima: obtido através da
relação entre as atividades específicas da enzima antes (Ae2) e
após a purificação (Ae1). Define-se atividade específica (Ae) como:
Ae = AJP [U/mg] Equação 4.3
onde: A=atividade da enzima expressa em [U];
P=proteínas totais expressa em [mg], ambas relativas à mesma fase
analisada.
b) Define-se portanto aumento de pureza (AP):
Equação 4.4
onde: Ae=atlvídade específica na fase extraída - FAII [U/mg];
Ae2=atividade específica da amostra antes da purificação [U/mg].
47

Materiais & Métodos
e) Rendimento expresso em proteínas totais ( np ): que será definido por:
np = P2 / P1 X 100 Equação 4.5
onde:
P2=proteínas totais da fase extraída [mg];
Pr=protelnas totais antes da purificação [mg].
As massas P2 e P1 correspondem à concentração de proteína multiplicada
pelo volume da fase considerada.
d) Rendimento expresso em atividade ou Recuperação (R) definido
por:
R = A2 I A1 X 100 Equação 4.6
onde: A2=atividade na fase extraída [U];
Ai=atividade da amostra antes da purificação [U].
As atividades A1 e A2 correspondem à atividade enzimática multiplicada pelo
volume da fase considerada.
48

Resultados & Discussões
5 - RESULTADOS E DISCUSSÕES
5.1 - A ENZIMA (3-XILOSIDASE
5. 1. 1 - Estabilidade
O estudo da estabilidade é importante para caracterização da enzima
(3-Xilosidase produzida pelo P. janthinellum assim como para o conhecimento de
suas limitações de manuseio durante a realização dos estudos de extração líquido-
líquido. Os resultados deste estudo são mostrados na Tabela 5.1 e 5.2:
Para ambas as temperaturas estudadas (4 ºC e -4 ºC}, a enzima, quando
solubilizada em pH ácido (3,0 ou 6,0) e neutro (7,0), apresentou grande estabilidade.
Sua atividade catalítica não variou significativamente, sendo que as oscilações
observadas, podem ser atribuídas a deficiências na homogeneização das amostra
analisadas. No entanto, quando a (3-Xilosidase foi solubilizada em pH 9,0, em ambas
as temperaturas, observou-se uma nítida redução de sua atividade catalítica: a partir
do 3° dia a 4 ºC e do 29° dia a -4 ºC. Tais resultados estão de acordo com os
encontrados na literatura (RODINOVA et ai., 1983; PULS et ai., 1995), que mostram
ter esta enzima melhor estabilidade e atividade catalítica em valores de pH ácido.
49

Resultados & Discussões
Tabela 5.1 - Atividade da p-xilosidase em diferentes valores de pH, em função do tempo, a 4ºC
Atividade da 13-Xilosidase [U/ml]
Tempo (dias) pH o 1 2 3 6 29 52 96
3,0 0,15 0,20 0,20 O, 16 0,19 O, 15 O, 12 O, 16
6,0 0,24 0,29 0,21 0,26 0,26 0,32 0,17 0,20
7,0 O, 11 O, 12 O, 12 O, 10 O, 11 O, 10 0,08 O, 12
9,0 0,15 O, 13 O, 10 0,05 0,03 o o o
Tabela 5.2 - Atividade da J3-xilosidase em diferentes valores de pH, em função do tempo a -4°C
Atividade da p-Xilosidase [U/ml]
pH Tempo (dias) ..
o 1 2 3 6 29 52 96 - - ...,. _____ ........_ __ ........_.,,......_. ___ . ---
3,0 O, 15 O, 14 0,24 0,25 0,20 0, 17 0, 12 0, 17 6,0 0,24 0,16 0,37 0,28 0,31 0,26 0, 17 0,27
7,0 O, 11 0,14 0,17 O, 18 0,14 0,10 0,08 0, 12
9,0 O, 15 0,14 0,35 0,33 0,29 0,08 0,05 0,03
RODINOVA et ai. (1983), em estudos da estabilidade desta enzima, para faixa
de pH de 4,0 a 6,0, observaram que a mesma é estável por 8 horas quando mantida
à temperatura ambiente. Essa faixa de pH muito se assemelha à avaliada neste
trabalho de dissertação, sendo que provavelmente, a temperatura exerça grande
influência sobre a atividade catalítica da enzima, visto que, apesar da similaridade da
faixa de pH estudado, após 96 dias, a enzima J3-xilosidase teve sua atividade
catalítica reduzida somente quando solubilizada e estocada em pH 9,0. Neste caso a
enzima perdeu a atividade em ambas as temperaturas estudadas.
50
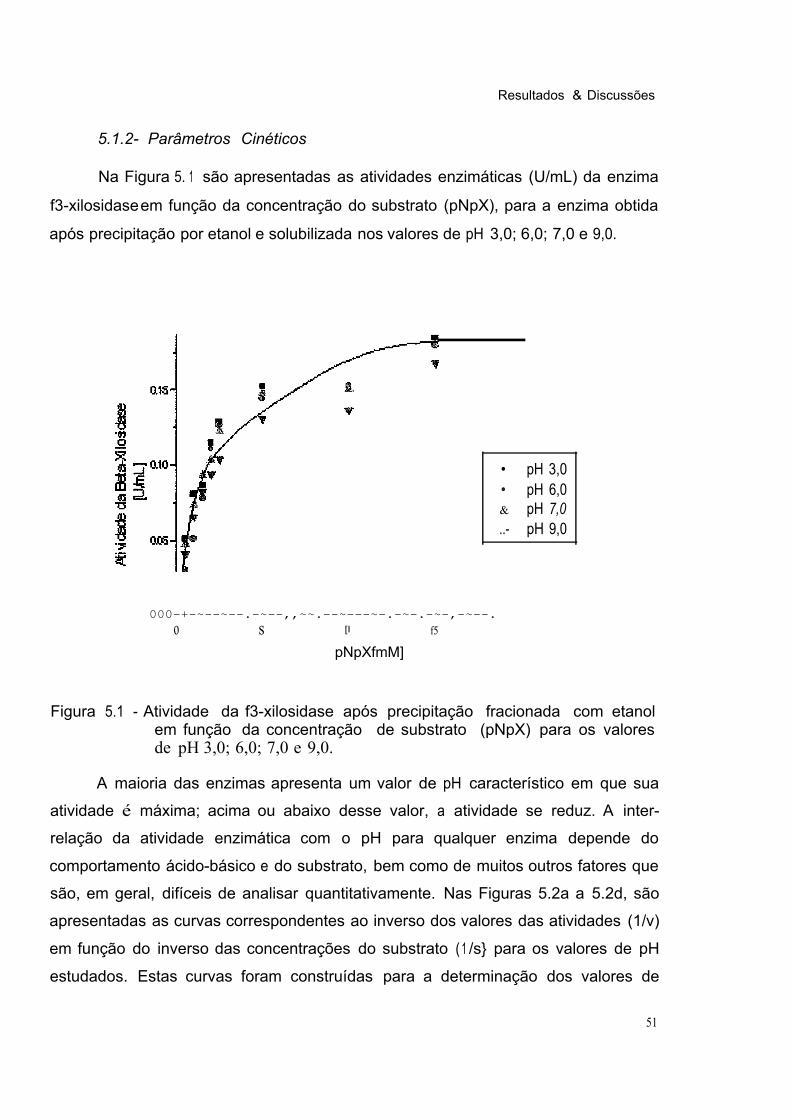
Resultados & Discussões
5.1.2- Parâmetros Cinéticos
Na Figura 5. 1 são apresentadas as atividades enzimáticas (U/mL) da enzima
f3-xilosidase em função da concentração do substrato (pNpX), para a enzima obtida
após precipitação por etanol e solubilizada nos valores de pH 3,0; 6,0; 7,0 e 9,0.
• pH 3,0 • pH 6,0 & pH 7,0 ..- pH 9,0
000-+-~--~--.-~--,,~~.--~---~-.-~-.-~-,-~--. o s [I
pNpXfmM] f5
Figura 5.1 - Atividade da f3-xilosidase após precipitação fracionada com etanol em função da concentração de substrato (pNpX) para os valores de pH 3,0; 6,0; 7,0 e 9,0.
A maioria das enzimas apresenta um valor de pH característico em que sua
atividade é máxima; acima ou abaixo desse valor, a atividade se reduz. A inter-
relação da atividade enzimática com o pH para qualquer enzima depende do
comportamento ácido-básico e do substrato, bem como de muitos outros fatores que
são, em geral, difíceis de analisar quantitativamente. Nas Figuras 5.2a a 5.2d, são
apresentadas as curvas correspondentes ao inverso dos valores das atividades (1/v)
em função do inverso das concentrações do substrato ( 1 /s} para os valores de pH
estudados. Estas curvas foram construídas para a determinação dos valores de
51
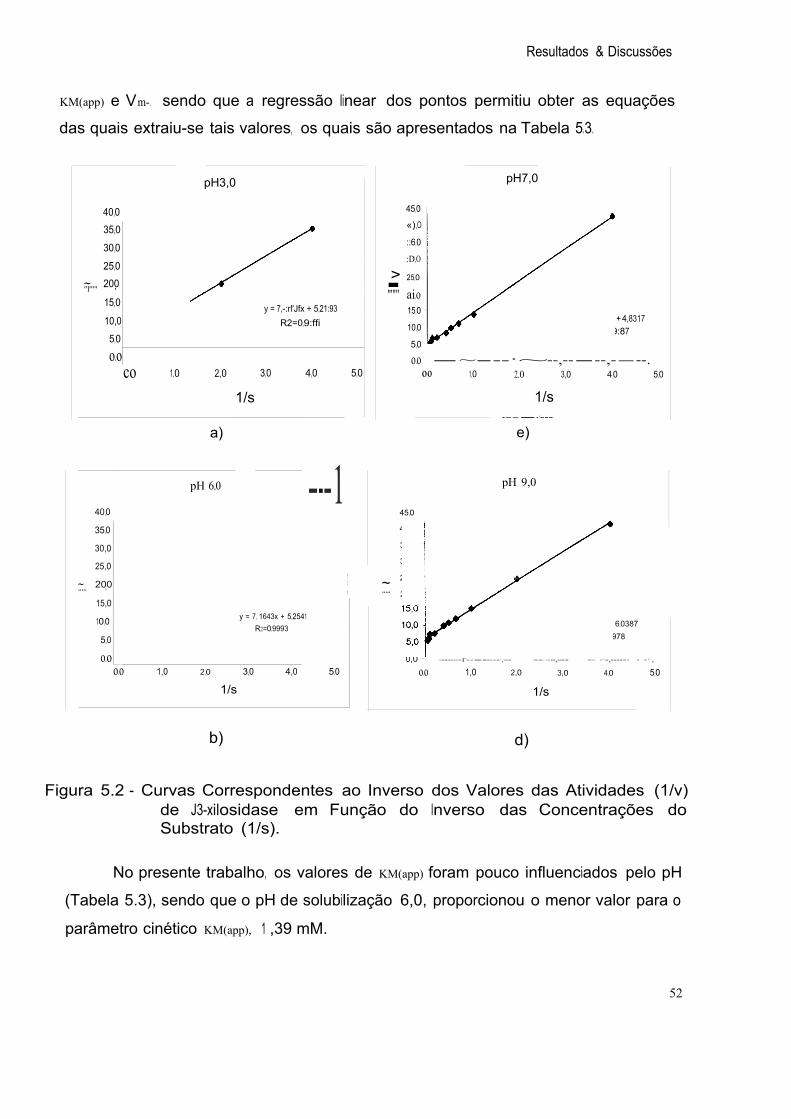
Resultados & Discussões
KM(app) e V m-. sendo que a regressão linear dos pontos permitiu obter as equações
das quais extraiu-se tais valores, os quais são apresentados na Tabela 5.3.
5,0
pH3,0
40,0 35,0
30,0
25,0 ~ 200 "l'"" •
15,0 y = 7,-:rl'Jfx + 5,21:93 10,0 R2=0,9:ffi
5,0
o.o co 1,0 2,0 3,0 4,0 5,0
1/s
a)
pH 6,0 ---1 40,0
35,0
30,0
1 25,0
~ 200 1 ..... . 1
15,0
10,0 y = 7, 1643x + 5,2541 R2=0,9993
5,0
o.o o.o 1,0 2,0 3,0 4,0 5,0
1/s
b)
pH7,0
45,0
«),O
::6,0
:D,O
> 25,0· - "'"" ai.o 15,0 ·
10,0
5,0
o.o -~---·~--,-----,----.
y = 9,:JlBc + 4,8317 R'=0,9:87
5,0
º·º 1,0 2.0 3,0 4,0
Figura 5.2 - Curvas Correspondentes ao Inverso dos Valores das Atividades (1/v) de J3-xilosidase em Função do Inverso das Concentrações do Substrato (1/s).
1/s -----·---
No presente trabalho, os valores de KM(app) foram pouco influenciados pelo pH
(Tabela 5.3), sendo que o pH de solubilização 6,0, proporcionou o menor valor para o
parâmetro cinético KM(app), 1 ,39 mM.
e)
pH 9,0
45,0
40,0
35,0
30,0
~ 25,0 ..... 20,0
y = 8,862x + 6,0387 R2 = 0,9978
0,0 · ----------r---------·,·---- ··--··--·--,----· ·-- -··--,-----·-· ·-- ··-··,
o.o 1,0 2,0 3,0 4,0
1/s
d)
52

Resultados & Discussões
Tabela 5.3 - KM(app) e Vm da 13-xilosidase obtidos após precipitação fracionada com etanol em função do inverso das concentrações do substrato para diferentes valores de pH
pH v; (U/ml) KM(app) (mM)
3,0 O, 19 1,42
6,0 O, 19 1,39
7,0 0,22 2,01
9,0 O, 17 1,47
RISTROPH & HUMPHREY (1985) calcularam a velocidade de reação da
í3-Xilosidase do caldo fermentado produzido por Thermonospora crassa e obtiveram
um KM de 0,82 mM e um Vm de 0,0085 U/ml, utilizando pNpX como substrato nas
concentrações de 0,049 a 8,9 mM. KUMAR & RAMÓN (1996), utilizando a mesma
metodologia de medida de atividade citada no presente trabalho, encontraram para a
í3-Xilosidase pura de Aspergillus nidulans, valores de 1, 1 mM e 2,56 U/ml para KM e
V m, respectivamente.
Comparando os resultados obtidos no presente trabalho (para o pH 6,0) com
aqueles dos autores citados, verifica-se que KUMAR & RAMÓN (1996) obtiveram um
KM 21 % menor, e que RISTROPH & HUMPHREY (1985) um v-, 95 % menor para
um KM 59 % menor. Estes autores, citam ainda, valores de KM de 0,22, 1,43 e 3,01
respectivamente, para Aspergillus, Bacillus e Malbranchea. Desta forma, o KM da
í3-Xilosidase encontrado no presente trabalho, é um valor condizente com a literatura,
pois este parâmetro apresenta significativa variação (para a enzima í3-Xilosidase),
quando analisados diferentes microrganismos produtores.
É importante ressaltar, que os valores de KM(app) e de V m obtidos neste
trabalho provavelmente seriam diferentes daqueles da enzima pura, pois as
determinações foram feitas com a í3-Xilosidase parcialmente purificada, portanto são
valores aparentes (app). Entretanto, são valores com boa aproximação das reais
condições de aplicação industrial, pois enzimas xilanolíticas normalmente não são
utilizadas na forma pura (CORTEZ, 1998).
53

Resultados & Discussões
5.2 - EXTRAÇÃO DA ENZIMA (3-XILOSIDASE POR SISTEMA DE MICELAS REVERSAS FORMADO A PARTIR DO AGENTE TENSOATIVO CATIÔNICO BDBAC
Na Tabela 5.4 estão apresentados os resultados da extração, em termos de
porcentagem de recuperação (Ro/o) da enzima (3-xilosidase por micelas reversas do
agente tensoativo BDBAC, obtidos nos ensaios realizados conforme o planejamento
fatorial 25-1.
Tabela 5.4 - Planejamento Fatorial Fracionário 25•1 com 1 Repetição Utilizado para Avaliação da Influência do pH, condutividade elétrica, concentração de hexanol e Temperatura na Recuperação da (3-xilosidase por Micelas Reversas de Agente Tensoativo Catiônico BDBAC
Ensaio nº.
pH Condutividade Elétrica (mS/cm)
BDBAC (M)
Hexanol (%)
Temp. (ºC)
y (%)
1 3,0 4,0 2 9,0 3 3,0 4 9,0 5 3,0 6 9,0 7 3,0 8 9,0 9 3,0 10 9,0 11 3,0 12 9,0 13 3,0 14 9,0 15 3,0 16 9,0
4,0 10 10 4,0 4,0 10 10 4,0 4,0 10 10 4,0 4,0 10 10
0,1 0,1 0,1 0,1 0,2 0,2 0,2 0,2 0,1 O, 1 O, 1 0,1 0,2 0,2 0,2 0,2
5,0 5,0 5,0 5,0 5,0 5,0 5,0 5,0 10 10 10 10 10 10 10 10
26 4 4 26 4 26 26 4 4 26 26 4 26 4 4 26
19,9 18,7 0,1 0,3 2,0 3,7 0,2 0,2 0,0 0,0 0,0 0,0 30,4 42,5 6,1 6,2 1,1 0,8 0,4 0,4 2,7 3,3 5,5 8,5 44,2 43,2 2,6 2,7 0,0 0,0 1,4 0,5
19,3 0,2 2,9 0,2 0,0 0,0 36,4 6,1 1,0 0,4 3,0 7,0 43,7 2,6 0,0 0,9
54

Resultados & Discussões
Com base nos resultados obtidos, pode-se observar que em todos os ensaios
realizados em pH 9,0, independente da combinação das outras variáveis, a
recuperação da enzima foi muito baixa (< 8%). Tal fato pode ser explicado
principalmente devido ao pH ótimo de atividade catalítica da enzima se encontrar na
faixa ácida (DESHPANDE et ai., 1994). Provavelmente nesta condição, ou seja em
pH 9,0, tenha ocorrido perda de atividade da enzima e consequentemente redução
na porcentagem de recuperação.
Baixa recuperação (<8%) para a enzima foi também encontrada em todos os
ensaios realizados no menor nível de temperatura (4ºC). Estes resultados
possivelmente se devem ao pequeno diâmetro das micelas reversas formadas
quando da utilização de tal temperatura, diminuindo assim sua capacidade de
absorção e englobamento da enzima. Segundo KREI & HUSTEDT (1989) a
capacidade máxima da micela é dependente da temperatura, ou seja, existe uma
faixa de temperatura na qual estas apresentam-se como formações estáveis,
favorecendo portanto a absorção de proteínas. PESSOA JR & VITOLO (1998), em
trabalhos de extração da enzima inulinase, também constataram que o tamanho da
micela reversa é bastante dependente da temperatura e, na faixa de 5 a 45ºC, as
micelas reversas em solventes orgânicos são formações estáveis e podem favorecer
a absorção desta enzima. Os resultados acima indicam que as variáveis pH e
temperatura influem na recuperação das biomoléculas em estudo.
Os maiores valores de recuperação para 13-Xilosidase (19,3, 36,4 e 43,7 %)
foram obtidos nos ensaios 01, 07 e 13, respectivamente (Tabela 5.4). Comparando-
se o ensaio 1 com o ensaio 7 (em que os níveis para o pH, temperatura e hexano!
foram mantidos constantes) verifica-se que o aumento da condutividade elétrica de 4
para 1 O mS/cm e da concentração de BDBAC de O, 1 para 0,2 M promoveu um
aumento na recuperação da enzima de 88,52 %. Este aumento foi ainda maior
(126, 13%) quando foram mantidos pH, condutividade elétrica e temperatura, e
aumentou-se a concentração de BDBAC de O, 1 para 0,2 M e a concentração de
hexanol de 5 para 10% (ensaios 1 e 13, respectivamente). Estes resultados sugerem
que diferentes combinações das variáveis interferem nos resultados de recuperação,
o que dificulta uma análise direta dos efeitos destas variáveis. Desta forma, os
55

Resultados & Discussões
efeitos principais das variáveis pH, condutividade elétrica, concentração de BDBAC,
concentração de hexanol e temperatura, bem como os efeitos interativos entre elas,
podem ser melhor avaliadas empregando-se a análise estatística.
As estimativas dos efeitos, erros-padrão e o teste t ( de Student) para a
recuperação da 13-xilosidase estão representados na Tabela 5.5. Os resultados da
análise estatística confirmam os efeitos sugeridos pela análise direta dos dados, e
também a existência de efeitos de interação entre as variáveis.
Tabela 5.5 - Estimativa dos Efeitos, Erros-padrão e teste t (Student) para os Parâmetros Estudados na Recuperação da 13-xilosidase por Micelas Reversas de BDBAC
Efeitos Estimativas Erros-padrão t
Média 7,73 +/-0,397 19,49 Fator A ( pH) -11, 11 +/-0,793 14,00*
Fator B (Condutividade) -1,35 +/-0,793 1,70
Fator C (Cone. BDBAC) 6,99 +/-0,793 8,81*
Fator D (Cone. Hexanol) -0,82 +/-0,793 1,03
Fator E (Temperatura) 10,52 +/-0,793 13,26*
AB 4,09 +/-0,793 5, 16*
AC -6,51 +/-0,793 8,21*
AD 1,92 +/-0,793 2,42*
AE -14, 13 +/-0,793 17,82*
BC 0,63 +/-0,793 0,78
BD -7,87 +/-0,793 9,92*
BE -4,39 +/-0,793 5,53*
CD 1,99 +/-0,793 2,50*
CE 7,55 +/-0,793 9,52*
DE -1, 17 +/-0,793 1,48 Valor de t para 95% de confiança com 16 GL (t = 2, 12043) * Significativo ao nível de 95% de Confiança
Como pode ser visto na Tabela 5.5, os maiores efeitos principais observados
para a recuperação da enzima foram os das variáveis pH e temperatura, que
56

Resultados & Discussões
apresentaram estimativas de -11, 11 e + 1 O, 52, respectivamente. Observa-se ainda
efeito interativo, ao nível de 95% de significância, entre estas variáveis, o qual
apresenta uma estimativa de valor elevado (-14, 13). O fato desta interação
apresentar sinal negativo indica que os resultados de recuperação são melhorados
com a condução dos ensaios em menores valores de pH e temperaturas mais
elevadas, nas respectivas faixas estudadas.
Nas Figuras 5.3a a 5.3d estão representadas as superfícies de resposta das
interações mais significativas. A superfície mostrada na Figura 5.3a, que representa
a interação entre as variáveis pH e temperatura, permite visualizar o comportamento
anteriormente citado.
A concentração de BDBAC também apresentou efeito principal significativo
(estimativa de 6,99) na recuperação da enzima, sugerindo que o aumento da
concentração de BDBAC promove um aumento no valor da variável resposta. Foi
constatada ainda a existência de interações significativas entre BDBAC e todas as
outras variáveis avaliadas, impossibilitando desta forma uma análise isolada do efeito
do mesmo. As Figuras 5.3b e 5.3c representam as superfícies de resposta referentes
as interações da concentração de BDBAC (C) com a temperatura (E) e com o pH (A).
Nota-se que, para estas duas interações houve aumento na recuperação da enzima
com o aumento da concentração de BDBAC, sendo que a recuperação foi maior com
o aumento da temperatura (Figuras 5.3b) e com a diminuição do pH (Figura 5.3c).
57

Resultados & Discussões
Figura 5.3a - Recuperação da f3-xilosidase por micelas reversas em função da temperatura e do pH, usando BDBAC O, 15M, condutividade elétrica de 7 ,OmS/cm, 7 ,5% de hexanol.
Figura 5.3b - Recuperação da f3-xilosidase por micelas reversas em função da temperatura e da concentração do BDBAC, usando pH 6,0, condutividade elétrica de 7,0mS/cm, 7,5% de hexanol.
58

Resultados & Discussões
Figura 5.3c - Recuperação da í3-xilosidase por micelas reversas em função da concentração de BDBAC e do pH, usando temperatura de15°C, 7,0mS/cm, 7,5% de hexanol.
- ·~
Figura 5.3d - Recuperação da í3-xilosidase por micelas reversas em função da condutividade elétrica e da concentração de hexanol, usando BDBAC O, 15M, pH 6,0, 7,5% de hexanol.
59

Resultados & Discussões
Por se tratar de uma molécula grande, da ordem de 100 kDa
(SULISTYO et ai., 1995; CORTEZ & PESSOA JR, 1999), a enzima 13-xilosidase
necessita de micelas reversas de raios maiores para englobá-la. Desta maneira o
uso do nível superior para a variável concentração de BDBAC se justifica, pois as
micelas podem apresentar variações de tamanho, dependendo, entre outros fatores,
da concentração do agente tensoativo e da temperatura (KREI & HUSTEDT, 1989;
PESSOA JR & VITOLO, 1998). Tal fato é previsível, visto que a micela é uma
organização de moléculas de um tensoativo que se orientam de modo a formar
micelas em soluções polares e micelas reversas e soluções apoiares. Seu tamanho
portanto, depende do número de moléculas de tensoativo. De fato, a temperatura
também influi diretamente sobre a estabilidade micelar, ou seja, a diminuição da
temperatura promove o encolhimento da micela e a diminuição da sua capacidade de
absorção. Por outro lado, temperaturas elevadas ocasionam o desaparecimento da
pseudofase micelar, deixando de existir na forma orgânica. Embora haja uma faixa
ideal de temperatura para formação de micelas reversas estáveis, no presente
trabalho (Figura 5.3b) ficou demonstrado que a recuperação foi dependente de
ambos os fatores, ou seja, houve o aumento da recuperação com o aumento da
concentração de BDBAC e da temperatura.
Para extração de biomoléculas por micelas reversas empregando tensoativo
catiônico, espera-se obter melhores resultados quando da utilização de valores de
pH superiores ao pI da biomolécula estudada, devido ao favorecimento das
interações eletrostáticas entre as enzimas e o agente tensoativo. Entretanto, no
presente trabalho, este comportamento não foi observado, visto que os melhores
rendimentos de extração ocorreram em valores de pH inferiores ao pI da
13-Xilosidase, que, conforme citado na literatura (PULS et ai., 1995; SUL YSTIO et
ai., 1995) está na faixa de 4,9 a 6,0. Pela Figura 5.3c pode-se constatar este fato,
uma vez que a maior recuperação ocorreu no nível -1 para o pH, que corresponde
ao valor 3,0. Esta mudança da faixa de pH considerada ótima para extração pode ser
explicada pela existência de interações hidrofóbicas entre a enzima e o agente
tensoativo, conforme modelo proposto por LUISI et ai. (1988), ilustrado na Figura 5.4.
60

Resultados & Discussões
Enzima Fase Orgânica Fase Aquosa
Figura 5.4 - Extração de uma Proteína por Micelas Reversas Considerando a Existência de Forças Hidrofóbicas de Interação (Adaptado de LUISI, et ai., 1988).
Outras importantes interações podem ser observadas como entre pH e
condutividade elétrica (AB), pH e concentração de hexano! (AD), condutividade
elétrica e concentração de hexano! (BD) e concentração de BDBAC e concentração
de hexano! (CD). Dentre estas, a que apresentou maior efeito foi a interação BD
(-7,87). A Figura 5.3d mostra a superfície de resposta para esta interação, sendo que
a recuperação aumenta quando se usam os níveis +1 da concentração de hexano! e
-1 de condutividade elétrica, e para o inverso, isto é -1 e +1, caracterizando, desta
forma, uma interação antagônica entre estes fatores.
ANDREWS et ai. (1994) verificaram que a força iônica pode afetar o tamanho
da micela, pois os grupamentos polares dos agentes tensoativos repelem-se e, com
isto, micelas maiores são formadas quando submetidas à baixa força iônica. Desta
forma, à medida que a força iônica da solução que contém a proteína aumenta, as
interações entre a proteína e o agente tensoativo diminuem, devido à redução do
efeito repulsivo dos grupos polares do tensoativo. Isto leva a um decréscimo do
transporte de água ou outras moléculas e, consequentemente, à formação de menos
e menores micelas reversas.
61

Resultados & Discussões
A interação antagônica observada no presente trabalho entre o co-solvente e
a condutividade elétrica não foi ainda elucidada pela literatura. Mas, segundo KREI &
HUSTEDT (1992), os co-solventes são adicionados aos sistemas de micelas
reversas de tensoativos catiônicos para aumentar o tamanho das mesmas. Eles se
assemelham a soluções tampão, que provocam repulsão entre as interações íon-íon
dos núcleos do tensoativo, permitindo a formação do núcleo interno da micela
reversa. Desta forma, a opção pelo uso da força iônica em seu nível inferior, isto é
4 mS/cm, implica na utilização do co-solvente no nível superior (10%), ou o inverso,
sem que haja prejuízo dos rendimentos.
Após a seleção das variáveis que apresentam efeitos significativos sobre o
processo estudado, conduziu-se uma análise de variância (ANOVA) para verificação
da possibilidade de ajuste de um modelo linear aos dados experimentais, o que foi
confirmado pelo elevado coeficiente de correlação múltiplo, ou seja R2 = 0,98. Os
parâmetros desta análise encontram-se na Tabela 5.6.
62

Resultados & Discussões
Tabela 5.6 - Parâmetros da Análise de Variância (ANOVA) dos Fatores Significativos ao Processo de Extração da Enzima P-xilosidase por Micelas Reversas do Agente Tensoativo BDBAC
Fonte de SQ GL QM Fator - f Valor - P Variação Fator A 987,01 1 987,01 189,74 0,0000* Fator B 14,47 1 14,47 2,78 0, 1136*
Fator e 390,88 1 390,88 75,14 0,0000*
Fator D 5,33 1 5,33 1,02 0,3256
Fator E 885,36 1 885,36 170, 19 0,0000*
AB 133,91 1 133,91 25,74 0,0001*
AC 339, 17 1 339, 17 65,20 0,0000*
AD 29,49 1 29,49 5,67 0,0292*
AE 1598, 10 1 1598, 10 307,21 0,0000*
BD 494,87 1 494,87 95,13 0,0000*
BE 153,91 1 153,91 29,59 0,0000*
CD 31,52 1 31,52 6,06 0,0248* .. - ._,..,
CE 456, 17 1 456, 17 87,69 0,0000*
Bloco 6,27 1 6,27 1,2 0,2877
Erro Total 94,71 18 5,03
Total 5614,90 31 R2 = 0,98 GL=graus de liberdade, SQ=soma quadrática, MQ=média quadrática * Significativos a 95%.
A validade do modelo linear foi verificada pela análise de variância de
regressão do modelo (Tabela 5.7). Através do teste F, ou seja, comparando-se o
valor F do modelo (80,7) com o valor de F tabelado (2,3), fica claro que o modelo é
altamente significativo. Além disso, ele não apresenta falta de ajuste, isto é, a
diferença entre os valores obtidos experimentalmente e os valores previstos pela
equação do modelo pode ser explicada pelo erro experimental.
63

Resultados & Discussões
Tabela 5.7 -Análise de variância de regressão para o modelo representativo do processo de extração de J3-xilosidase por micelas reversas de BDBAC, na região em estudo
Causa de SQ GL MQ F
Variação
Modelo 5520,20 13 424,63 80,71
Erro 94,70 18 5,26
Total 5614,90 31 R =0,98 GL=graus de liberdade, SQ=soma quadrática, MQ=média quadrática
Desta forma, podemos descrever um modelo matemático para representar o
processo, o qual pode ser expresso pela equação 5.1, onde Y representa
porcentagem de recuperação da J3-xilosidase, A o pH, B a condutividade elétrica,
C a concentração de BDBAC, D a concentração de hexano! e E a temperatura.
Y=7,73 • 5,55A • 0,678 + 3,49C • 0,410 + 5,26E + 2,05A8 • 3,26AC
+ 0,96AO • 7,07AE • 3,9380 • 2,198E + 0,99CO + 0,78CE Equação 5.1
A equação 5.1 prevê valores de recuperação para a J3-xilosidase de 40%, para
as seguintes condições: pH 3,0, condutividade elétrica 4,0 mS/cm, concentração de
BDBAC 0,2M, concentração de hexano! 10% e temperatura 26ºC. Nestas condições,
realizou-se um novo ensaio visando a confirmação do modelo e obteve-se 39,5% de
recuperação. Este resultado demonstra que o modelo se adequou perfeitamente aos
resultados experimentais, pois nenhuma diferença significativa entre o valor previsto
e o experimental foi observada.
64

Resultados & Discussões
5.2.1 - Otimização do Processo de Extração por Mice/as Reversas de BDBAC em lsooctano e Hexano/
Os resultados obtidos anteriormente mostraram que dentre os 05 fatores
inicialmente estudados para esse sistema (pH, condutividade elétrica, concentração
de BDBAC, concentração de hexanol e temperatura), apresentaram significância em
ordem decrescente os seguintes fatores: pH>temperatura>concentração de BDBAC.
Os maiores efeitos foram causados pela influência negativa do pH e positiva da
temperatura. Isto equivale a dizer que os valores para a variável resposta, são
aumentados quando o fator pH é utilizado no seu nível inferior (3,0) e temperatura no
superior (26 ºC).
Desta forma, com o objetivo de se otimizar o processo de extração de
j3-xilosidase por micelas reversas do agente tensoativo catiônico BDBAC, estudou-se
a influência do binômio pH e temperatura sobre o processo.
Buscando-se obter um modelo otimizado que pudesse ser usado para o
sistema de micelas reversas formados pelo tensoativo BDBAC em isooctano e
hexano!, empregou-se um planejamento fatorial completo 22 com face centrada e 03
repetições no ponto central, sendo estudadas as variáveis pH e temperatura. A
Tabela 5.8 mostra os níveis utilizados nos ensaios para as duas variáveis estudadas
e os valores obtidos para a variável resposta, porcentagem de recuperação da
enzima.
Os níveis dos fatores usados na otimização foram codificados em -1, O, +1,
correspondendo respectivamente ao nível superior, ponto central e nível inferior.
65

Resultados & Discussões
Tabela 5.8 - Matriz do Planejamento Fatorial Completo 22 Com Face Centrada e 03 Repetições no Ponto Central Utilizada para Avaliação da Influência do Binômio pH e Temperatura na Recuperação da (3-xilosidase por Micelas Reversas de Agente Tensoativo Catiônico BDBAC
Variáveis Valores Ensaio Codificadas Empregados Recuperação
Nº pH Temperatura pH Temperatura (%) 1 -1 -1 2 22 o 2 +1 -1 4 22 24
3 -1 +1 2 30 o 4 +1 +1 4 30 o 5 -1 o 2 26 o 6 +1 o 4 26 41
7 o -1 3 22 49
8 o +1 3 30 o 9 o o 3 26 38 ..
10 o o 3 26 40
11 o o 3 26 31
Os dados experimentais foram analisados estatisticamente, de acordo com o
planejamento preestabelecido, para verificar o efeito dos fatores em estudo. A
análise dos dados foi feita com o auxílio de software específico para este fim,
utilizando como variável resposta a porcentagem de recuperação da enzima
(3-Xilosidase.
Como pode ser visualizado na Tabela 5.8, independentemente da temperatura
utilizada, em valores de pH 2,0, os resultados de recuperação obtidos foram iguais a
zero. Isto se deve provavelmente à desnaturação da enzima. Os mesmos resultados
nulos foram observados quando empregada no processo de extração a temperatura
de 30ºC, independente do pH utilizado no ensaio. Tais resultados nulos de extração
podem ter sido ocasionados pela inexistência de fase micelar a 30 ºC, quando da
utilização deste sistema. Porém, a existência ou não de fase micelar, somente
66

Resultados & Discussões
poderia ser elucidada pelo estudo do diagrama de fases do sistema formado por
BDBAC/hexanol/isooctano/água. No entanto, tal sistema não encontra-se disponível
na literatura.
Por outro lado, as extrações realizadas nas condições definidas pelo ponto
central (pH 3,0 e 26ºC), a recuperação da enzima J3-xilosidase alcançou maiores
valores (da ordem de 40%). Isto sugere a existência de efeitos quadráticos para
ambas as variáveis avaliadas e que o ponto máximo de recuperação encontra-se na
região estudada.
Tensoativos catiônicos, tais como o BDBAC, normalmente favorecem o
transporte de enzimas para o interior micela quando essas possuem carga elétrica
negativa, o que é alcançado com o uso de valores de pH superiores ao pi da enzima.
O ponto isoelétrico da J3-xilosidase de diferentes microorganismos encontra-se entre
4,9 e 6,0, segundo dados encontrados na literatura (DESHPAND et ai. 1994;
SULYSTIO et ai., 1995) e portanto é esperado que com o uso de agentes tensoativos
catiônicos a enzima migre para o interior das micelas reversas quando o pH do meio
é superior ao seu pi.
Todavia, neste trabalho, a situação é oposta ou seja, maiores valores de
recuperação da enzima foram obtidos em pH inferior ao pI da enzima, sugerindo que
a transferência da enzima para a fase micelar seja governada por interações
hidrofóbicas entre a proteina e o tensoativo (LUISI et ai. 1988).
Os parâmetros de um modelo de segunda ordem (quadrático) usados para
estimar a porcentagem da enzima recuperada em função do pH e da temperatura
foram obtidos através da análise de regressões múltiplas, mostrada na Tabela 5.9.
- --
67

Resultados & Discussões
Tabela 5.9 - Variáveis, Coeficientes, Teste t (Student), Valores de p do Planejamento Completo com Face Centrada 22 com 03 Repetições no Ponto Central para Extração de (3-xilosidase por Micelas Reversas de BDBAC
Variáveis Coeficientes t p
Média 36,89 5,74 <0,005 pH (A) 10,83 2,12 0,08
Temperatura (B) -12, 17 -2,38 0,06 pH x Temperatura (AB) -6 -0,96 0,38
pH x pH (A2) -17,24 -2, 18 0,08 Temperatura x Temperatura (82) -13,24 -1,68 0, 15
A significância estatística do modelo quadrático (Tabela 5.1 O) foi avaliada pelo
teste F, e revelou que esta regressão é estatisticamente significativa a um nível de
10% de probabilidade. O modelo não apresenta falta de ajuste e o coeficiente de
determinação (R2 = 0,81) indica que 81% das variações na recuperação da enzima
por este sistema podem ser explicadas pelo modelo. Assim, um modelo matemático
que represente a recuperação de (3-xilosidase por micelas reversas de BDBAC (em
isooctano e hexanol) pode ser expresso pela Equação 5.2:
Y = 36,89 + 10,83A-12,178-6,00AB • 17,24A2 -13,2482 Equação 5.2
onde: Y=recuperação da enzima(%), A e B valores codificados de pH e temperatura,
respectivamente.
68
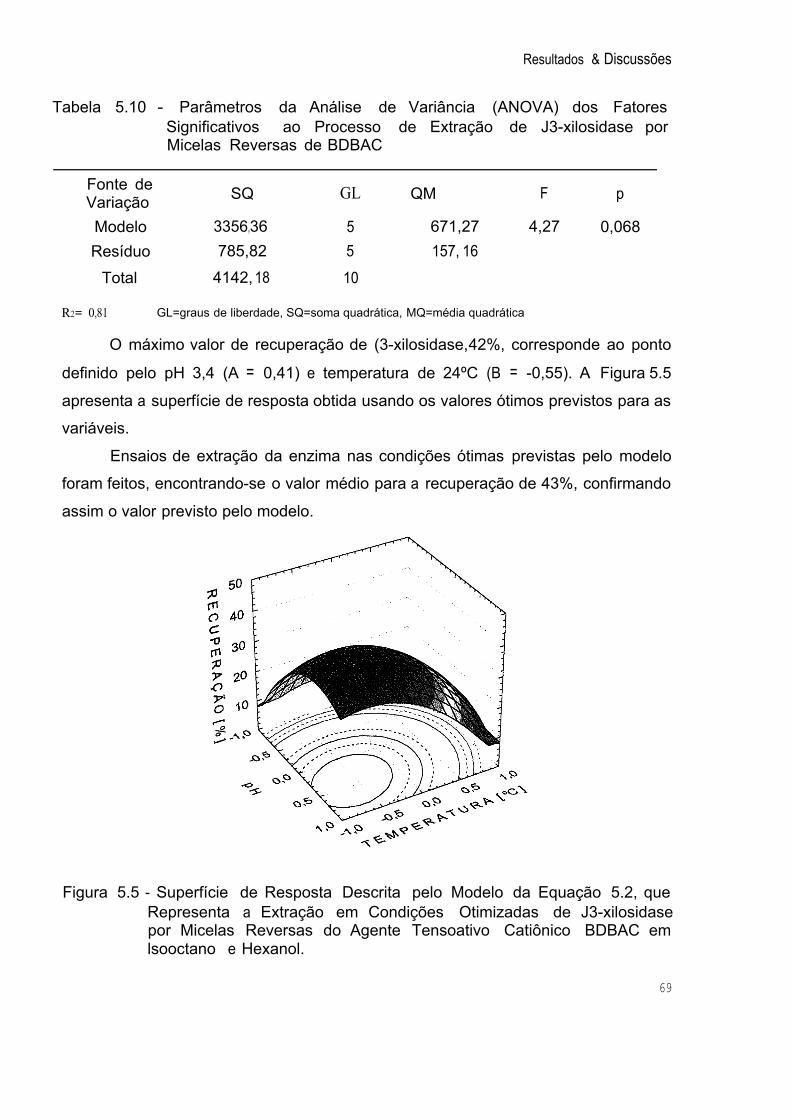
Resultados & Discussões
Tabela 5.10 - Parâmetros da Análise de Variância (ANOVA) dos Fatores Significativos ao Processo de Extração de J3-xilosidase por Micelas Reversas de BDBAC
Fonte de SQ GL QM F Variação p
Modelo 3356,36 5 671,27 4,27 0,068 Resíduo 785,82 5 157, 16
Total 4142, 18 10
R2= 0,81 GL=graus de liberdade, SQ=soma quadrática, MQ=média quadrática
O máximo valor de recuperação de (3-xilosidase, 42%, corresponde ao ponto
definido pelo pH 3,4 (A = 0,41) e temperatura de 24ºC (B = -0,55). A Figura 5.5
apresenta a superfície de resposta obtida usando os valores ótimos previstos para as
variáveis.
Ensaios de extração da enzima nas condições ótimas previstas pelo modelo
foram feitos, encontrando-se o valor médio para a recuperação de 43%, confirmando
assim o valor previsto pelo modelo.
Figura 5.5 - Superfície de Resposta Descrita pelo Modelo da Equação 5.2, que Representa a Extração em Condições Otimizadas de J3-xilosidase por Micelas Reversas do Agente Tensoativo Catiônico BDBAC em lsooctano e Hexanol.
69

Resultados & Discussões
A extração da enzima J3-xilosidase por micelas reversas de BDBAC, nas
condições otimizadas previstas pelo modelo obtido: pH 3,4, temperatura de 24ºC,
condutividade elétrica 4mS/cm, concentração de tensoativo 0,2M e concentração de
hexano! de 10%, proporcionou um aumento de pureza de 2 vezes para a enzima
extraída para FAII (em relação ao precipitado solubilizado), como pode ser visto na
Tabela 5.11.
Tabela 5.11 - Aumento de Pureza para Enzima J3-xilosidase
Atividade Específica U/mg
Ae (após a extração) Ae ( antes da extração)
5,98 3,0
Ap 2 Ae = atividade específica; Ap = aumento de pureza
Apesar do aumento de pureza ser modesto, os resultados mostram mais uma
vez, ser possível a utilização da extração por micelas reversas, para separação de
enzimas de interesse. No entanto, os valores obtidos no presente são similares a
outros encontrados na literatura para purificação de enzimas por micelas reversas
(PESSOA JR & VITOLO, 1997; GIOVENCO & VERHEGGEN, 1987).
5.2.2 - Determinação do Raio Mice/ar
Foi determinado o raio das micelas reversas no ponto de ótimo do
planejamento, utilizando microemulsão com concentração de BDBAC de 0,2 M, 10 %
de hexano!, condutividade elétrica de 4 mS/cm, temperatura de 24°C e pH 3,3.
O raio micelar obtido (Tabela 5.12) foi superior a 5nm, valor indicado por KREI
& HUSTEDT (1992) para que uma proteína de 100 kDa possa ser englobada. Assim,
como a J3-Xilosidase possui massa molar nesta ordem de grandeza, o raio micelar
não constituiu um obstáculo à extração. Pode-se verificar, também, na Tabela 5.12
que o raio micelar na fase micelar II (FMII) é menor (1,8 nm) que 5 nm e indica que a
70

Resultados & Discussões
micela reversa teve seu diâmetro reduzido para um valor que não permite que a
enzima p-xilosidase esteja ainda englobada.
Tabela 5.12 - Wo e Rm Determinados na FAI E FAII Obtidas após Extração por Micelas Reversas de BDBAC a 0,2M, 10 % de Hexanol, Condutividade Elétrica de 4 mS/cm, Temperatura de 24°C e pH 3,4
Fase da Extração Rm (nm)
FMI
FMII
84,19
11,04
13,80
1,80
Apesar do raio micelar ser favorável à absorção da p-xilosidase, verificou-se
que a extração da enzima foi regida por forças hidrofóbicas, pois mesmo com o uso
de agente tensoativo catiônico, os melhores resultados de extração foram
conseguidos nos ensaios onde valores de pH eram inferiores ao pi da enzima.
Portanto provavelmente a proteína não estava localizada no interior da micela
reversa, e sim, ligada às suas partes hidrofóbicas, conforme proposto por LUISI et ai.
(1988).
- ·-
5.2.3 - Eletroforese
Foi realizada eletroforese em condições desnaturantes (SDS-PAGE) para
identificação das proteínas presentes no precipitado solubilizado, na FAI e na FAII.
Para realização do processo de extração e obtenção das amostras que foram
analisadas, utilizaram-se as condições otimizadas obtidas, ou seja, microemulsão
com concentração de BDBAC de 0,2M, 1 O % de hexano!, condutividade elétrica de 4
mS/cm, temperatura de 24ºC e pH 3,3.
Na Figura 5.6, as linhas 1 e 2 correspondem aos marcadores de massa molar,
as linhas 3, 4 e 5, o precipitado solubilizado, a FAI e a FAII, respectivamente.
71

Resultados & Discussões
205.000
116.000 -110kDa 91.UOO
-1001dla
84.000 66.000
55.lJOO 45.000 36.000
29.000 24JJOO -20kDa 20.000 .
6..500
1 2 3 4 5
Figura 5.6 - Eletroforese em Condições Desnaturantes (505-PAGE) das Etapas do Processo de Extração da Enzima f3-Xilosidase por Micelas Reversas pré Purificada por Precipitação Fracionada com Etanol: Linha 1 e 2 - Marcadores de Massa Molar (205, 116, 97, 84, 66, 55, 45, 36, 29, 24, 20 e 6,5 kDa), Linha 3 - Precipitado Solubilizado, Linha 4 - FAI e Linha 5 - FAil.
A linha 3 da eletroforese, cuja amostra foi obtida após precipitação
francionada com etanol a 20 e 60%, permite verificar a presença de proteínas com
bandas nítidas na faixa de 100, 11 O kDa e 20 kDa. Semelhante aos resultados
obtidos por CORTEZ & PESSOA ( 1999), as bandas obtidas de 100 e 11 O kDa
correspondem à !3-xilosidase. VAN-PEIJ-N-N-M-E et ai. ( 1997), cultivando
Aspergíllus níger em arabinoxilana, obtiveram e purificaram uma !3-xilosidase de
massa molar de 110 kDa identificada por SDS-PAGE. A banda obtida a 20 kDa,
provavelmente se deva a uma endoxilanase, cuja identificação foi feita por
RODRIGUES (1997) utilizando como técnica de purificação a extração por micelas
reversas.
72

Resultados & Discussões
Nas linhas 4 e 5, correspondente a FAI e FAII respectivamente, observa-se
somente as bandas de 100 e 11 O kDa, constatando assim que somente a enzima
13-xilosidase está presente em ambas as fases em quantidade apreciáveis.
Assim, conclui-se que a extração por micelas reversas nas condições
utilizadas, mostrou-se eficiente na separação da enzima de interesse.
73
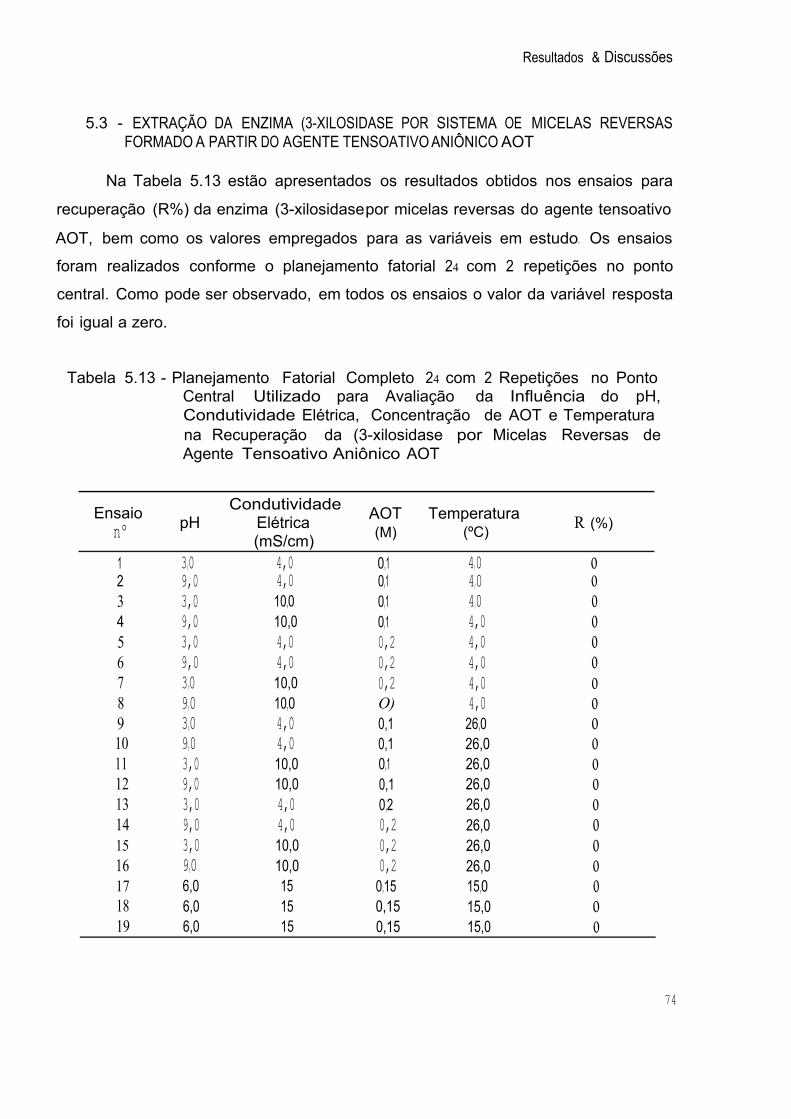
Resultados & Discussões
5.3 - EXTRAÇÃO DA ENZIMA (3-XILOSIDASE POR SISTEMA OE MICELAS REVERSAS FORMADO A PARTIR DO AGENTE TENSOATIVO ANIÔNICO AOT
Na Tabela 5.13 estão apresentados os resultados obtidos nos ensaios para
recuperação (R%) da enzima (3-xilosidase por micelas reversas do agente tensoativo
AOT, bem como os valores empregados para as variáveis em estudo. Os ensaios
foram realizados conforme o planejamento fatorial 24 com 2 repetições no ponto
central. Como pode ser observado, em todos os ensaios o valor da variável resposta
foi igual a zero.
Tabela 5.13 - Planejamento Fatorial Completo 24 com 2 Repetições no Ponto Central Utilizado para Avaliação da Influência do pH, Condutividade Elétrica, Concentração de AOT e Temperatura na Recuperação da (3-xilosidase por Micelas Reversas de Agente Tensoativo Aniônico AOT
Ensaio nº pH
Condutividade Elétrica (mS/cm)
AOT (M)
Temperatura (ºC) R (%)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
3,0 9,0 3,0 9,0 3,0 9,0 3,0 9,0 3,0 9,0 3,0 9,0 3,0 9,0 3,0 9,0 6,0 6,0 6,0
4,0 4,0 10,0 10,0 4,0 4,0 10,0 10,0 4,0 4,0 10,0 10,0 4,0 4,0 10,0 10,0 15 15 15
0,1 0,1 0,1 0,1 0,2 0,2 0,2 O) 0,1 0,1 0,1 0,1 0,2 0,2 0,2 0,2
0,15 0,15 0,15
4,0 4,0 4,0 4,0 4,0 4,0 4,0 4,0 26,0 26,0 26,0 26,0 26,0 26,0 26,0 26,0 15,0 15,0 15,0
o o o o o o o o o o o o o o o o o o o
74

Resultados & Discussões
5.4 - EXTRAÇÃO DA ENZIMA !3-XILOSIDASE POR SISTEMA DE MICELAS REVERSAS FORMADO A PARTIR DO AGENTE TENSOATIVO CATIÔNICO CTAB
Para o estudo da influência das variáveis pH, concentração de CTAB,
condutividade elétrica, concentração de hexanol e concentração de butanol, sobre a
extração de !3-xilosidase utilizou-se um planejamento fatorial fracionário 25-1 com 03
repetições no ponto central. Esse planejamento foi proposto para identificar os
fatores significativos para o sistema de extração sem que o número de ensaios fosse
demasiadamente grande. A matriz da Tabela 5.14 mostra os níveis utilizados para
cada variável nos diferentes ensaios, bem como os resultados de recuperação por
micelas reversas da enzima f3-xilosidase obtidos.
Tabela 5.14 - Planejamento Fatorial Fracionário 25-1 para Estudo da Influência do pH, Concentração de CT AB, Condutividade Elétrica, Concentração de Hexanol, Concentração de Butanol, na Recuperação de 13-xilosidase por Micelas Reversas do Agente Tensoativo Catiônico CTAB
Ensaio nº pH CTAB (M)
Condutividade Elétrica (mS/cm)
Hexanol Butanol Recuperação (%) (%) (%)
1 3,0 2 8,0 3 3,0 4 8,0 5 3,0 6 8,0 7 3,0 8 8,0 9 3,0 10 8,0 11 3,0 12 8,0 13 3,0 14 8,0 15 3,0 16 8,0 17 5,5 18 5,5 19 5,5
0,1 0,1 0,2 0,2 0,1 O, 1 0,2 0,2 0,1 O, 1 0,2 0,2 0,1 0,1 0,2 0,2 0,15 0,15 0,15
4,0 4,0 4,0 4,0 10,0 10,0 10,0 10,0 4,0 4,0 4,0 4,0 10,0 10,0 10,0 10,0 7,0 7,0 7,0
5,0 5,0 5,0 5,0 5,0 5,0 5,0 5,0 10,0 10,0 10,0 10,0 10,0 10,0 10,0 10,0 7,5 7,5 7,5
20 10 10 20 10 20 20 10 10 20 20 10 20 10 10 20 15 15 15
o.o 0,6 0,0
59,3 7,0 18,7 1,7 0,0 7,3 17,5 2,9
32,3 1,5
18,8 0,0 35,3 5,1 4,5 9,5
76

Resultados & Discussões
RODRIGUES (1997), obteve 9% de rendimento em atividade de xilanases, em
condições semelhantes de extração (temperatura, pH, concentração de AOT).
Porém, é importante ressaltar que as xilanases têm sua atividade enzimática
determinada pelo método descrito por MILLER (1959), que mede a quantidade de
açúcares redutores totais liberados após reação de hidrólise. Sendo as xilanases um
complexo formado por diversas enzimas hidrolíticas, este método não possui grande
especificidade e portanto todas enzimas do complexo apresentam resposta quando
de sua utilização. Essa diferença de metodologia empregada por RODRIGUES
(1997) em relação ao presente trabalho de dissertação, pode explicar as diferenças
encontradas nos resultados. Além disso, outras possibilidades de ter-se obtido
resultados nulos no presente trabalho, é a baixa concentração de f3-xilosidase e o
uso de um método de determinação de atividade específico para esta enzima
(KUMAR & RÁMON, 1996), o qual pode ter sido influenciado pelos componentes do
meio de extração por micelas reversas como o solvente isooctano ou o tensoativo
AOT.
75

Resultados & Discussões
Observa-se nos resultados obtidos, que na maioria dos ensaios realizados em
pH 8,0, a recuperação da enzima foi superior a 15%, chegando a 59,3 % no ensaio
de número 04. Por outro lado, verifica-se que em todos os ensaios conduzidos em
pH 3,0, independente do nível das outras variáveis, os resultados de recuperação da
enzima obtidos foram baixos (inferiores a 8%). Tais resultados diferem daqueles
obtidos anteriormente com o uso do agente tensoativo também catiônico BDBAC, em
que os melhores resultados de extração foram obtidos com o uso de pH 3,0. Essa
diferença nos resultados se deve, principalmente, às forças que atuam no sistema ou
seja, no presente caso, a extração por micelas reversas do agente tensoativo CT AB
é muito provavelmente governada por atrações eletrostáticas. Esses resultados
foram análogos aos obtidos por SILVA et ai. (1998) em estudos de extração de
p-xilosidase e RODRIGUES et ai. (1999) na extração de xilanases, ambas enzimas
produzidas por P. janthínellum e extraídas por micelas reversas de BDBAC.
Nos ensaios 04 e 12, foram obtidos valores de recuperação de 59,3 e 32,3%
respectivamente. Em ambos os casos utilizou-se o pH 8,0, concentração de CTAB
de 0,2 M e condutividade elétrica de 4mS/cm. Verifica-se ainda que o aumento da
concentração de butano! de 1 O para 20% e a diminuição da concentração de hexano!
de 10 para 5% ocasionou um aumento no valor da recuperação da enzima de 84%.
Observou-se, também, aumento na recuperação (cerca de 10%), nos ensaios 12 e
16 quando aumentou-se a concentração de butano! de 1 O para 20% e a
condutividade elétrica do meio de 4 para 1 O mS/cm, mantendo-se os mesmos
valores de pH, concentração de CTAB e concentração de hexano!. Em ambos os
casos, a recuperação foi beneficiada com o aumento da concentração de butano! no
sistema. Uma vez que o papel desta substância no sistema é a de um segundo co-
solvente, tal comportamento era esperado, visto que os co-solventes são
freqüentemente adicionados aos sistemas de micelas reversas formadas por agentes
tensoativos catiônicos para aumentar o tamanho das micelas reversas (KREI &
HUSTEDT, 1992).
Conclui-se, portanto, existirem diferentes combinações de variáveis
interferindo no processo de extração de p-xilosidase por micelas reversas de CTAB.
Em função disso, conclui-se que é recomendável a utilização de um planejamento
77
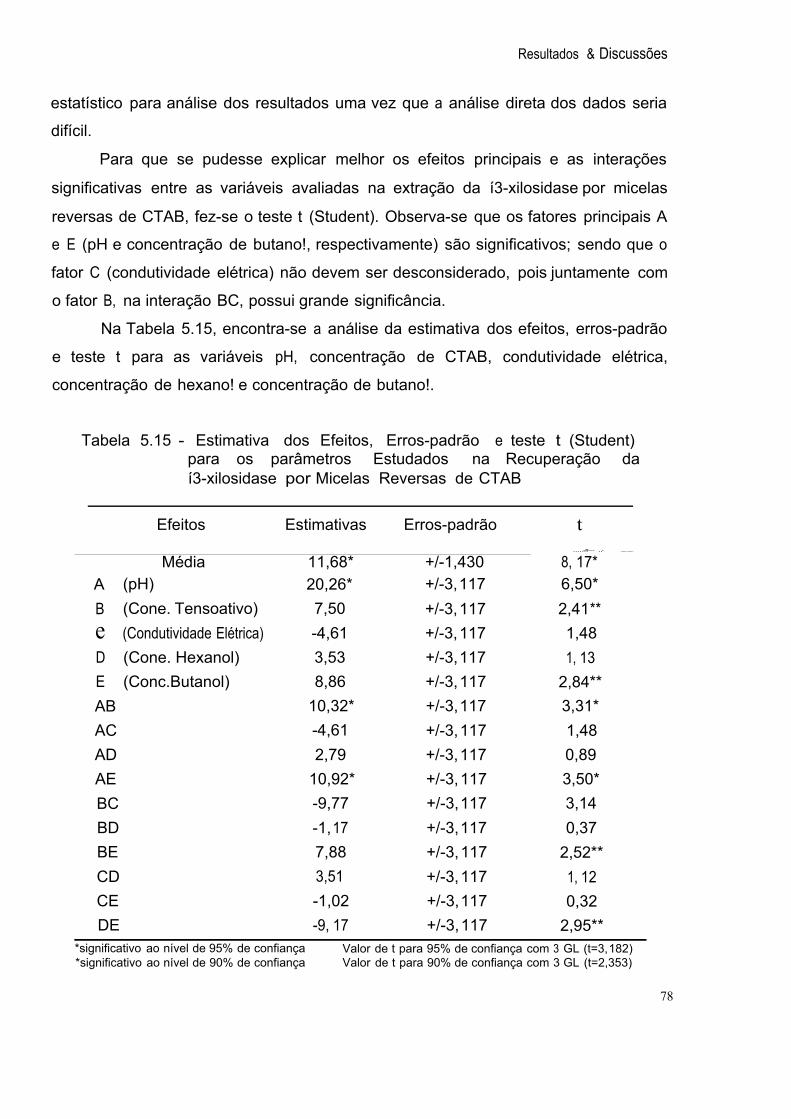
Resultados & Discussões
estatístico para análise dos resultados uma vez que a análise direta dos dados seria
difícil.
Para que se pudesse explicar melhor os efeitos principais e as interações
significativas entre as variáveis avaliadas na extração da í3-xilosidase por micelas
reversas de CTAB, fez-se o teste t (Student). Observa-se que os fatores principais A
e E (pH e concentração de butano!, respectivamente) são significativos; sendo que o
fator C (condutividade elétrica) não devem ser desconsiderado, pois juntamente com
o fator B, na interação BC, possui grande significância.
Na Tabela 5.15, encontra-se a análise da estimativa dos efeitos, erros-padrão
e teste t para as variáveis pH, concentração de CTAB, condutividade elétrica,
concentração de hexano! e concentração de butano!.
Tabela 5.15 - Estimativa dos Efeitos, Erros-padrão e teste t (Student) para os parâmetros Estudados na Recuperação da í3-xilosidase por Micelas Reversas de CTAB
Efeitos Estimativas Erros-padrão t ....... -=--- .. - ...........
Média 11,68* +/-1,430 8, 17* A (pH) 20,26* +/-3, 117 6,50* B (Cone. Tensoativo) 7,50 +/-3, 117 2,41 ** e (Condutividade Elétrica) -4,61 +/-3, 117 1,48 D (Cone. Hexanol) 3,53 +/-3, 117 1, 13 E (Conc.Butanol) 8,86 +/-3, 117 2,84** AB 10,32* +/-3, 117 3,31* AC -4,61 +/-3, 117 1,48 AD 2,79 +/-3, 117 0,89 AE 10,92* +/-3, 117 3,50* BC -9,77 +/-3, 117 3,14 BD -1, 17 +/-3, 117 0,37 BE 7,88 +/-3, 117 2,52** CD 3,51 +/-3, 117 1, 12 CE -1,02 +/-3, 117 0,32 DE -9, 17 +/-3, 117 2,95**
*significativo ao nível de 95% de confiança Valor de t para 95% de confiança com 3 GL (t=3, 182) *significativo ao nível de 90% de confiança Valor de t para 90% de confiança com 3 GL (t=2,353)
78

Resultados & Discussões
Como pode ser visto na Tabela 5.15, os maiores efeitos principais observados
para a recuperação da enzima foram os das variáveis pH e concentração de butano!,
que apresentaram estimativas de (+20,26) e (+8,86), respectivamente. Observa-se
ainda efeito interativo, ao nível de 90% de significância, entre as variáveis AB com
estimativas de (+10,32) e BC de (-9,77). A interação AB, por apresentar sinal
positivo, significa que os valores de recuperação da enzima 13-xilosidase por micelas
reversas de CTAB, serão melhorados com a condução do processo em maiores
valores de pH e concentração de tensoativo, ou seja pH 8,0 e 0,2 M,
respectivamente. Este fato confirma a hipótese da atuação da atração eletrostática
entre as moléculas do tensoativo e a proteína, como sendo a força motriz para
transferência da enzima para o interior das micelas (LUISI, et el., 1989). Neste caso
a utilização da concentração de CTAB em seu valor máximo de 0,2 M, foi necessária
para obtenção de micelas maiores e em maior número, capazes de englobar maior
quantidade de enzima. Com relação à interação de sinal negativo, BC, significa que
os valores da variável resposta, recuperação, são melhorados quando se utilizar nos
ensaios os maiores valores para concentração de tensoativo (análogo a interação
AB) e menores para condutividade elétrica, ou seja 4 mS/cm.
Assim, uma vez identificadas as variáveis que influenciam significativamente o
processo de extração de !3-xilosidase por micelas reversas de CT AB, foram
elaboradas as superfícies de resposta com as interações significativas (5.7a a 5.7e).
79
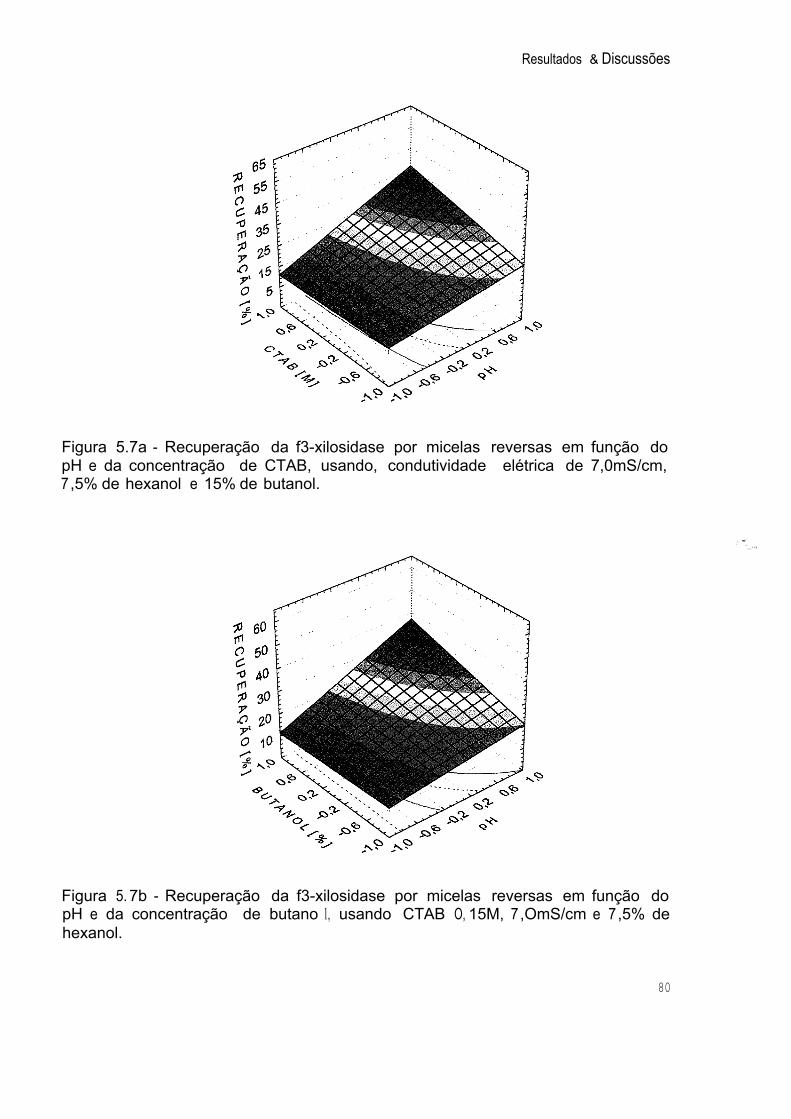
Resultados & Discussões
Figura 5.7a - Recuperação da f3-xilosidase por micelas reversas em função do pH e da concentração de CTAB, usando, condutividade elétrica de 7,0mS/cm, 7 ,5% de hexanol e 15% de butanol.
,' -· . ~_..,
Figura 5. 7b - Recuperação da f3-xilosidase por micelas reversas em função do pH e da concentração de butano 1, usando CT AB O, 15M, 7 ,OmS/cm e 7 ,5% de hexanol.
80

Resultados & Discussões
Figura 5.7c - Recuperação da f3-xilosidase por micelas reversas em função da condutividade elétrica e da concentração de CT AB, usando pH 5,5, 7 ,O mS/cm e 7 ,5% de hexano 1.
Figura 5.7d - Recuperação da f3-xilosidase por micelas reversas em função da concentração de CT AB e da concentração de butano 1, usando pH 5,5, 7,0mS/cm e 7,5% de hexanol.
81

Resultados & Discussões
Figura 5. 7e - Recuperação da J3-xilosidase por micelas reversas em função da concentração de hexanol e da concentração de butanol, usando pH 5,5, 7,0mS/cm e CTAB 0,15M.
Após seleção dos fatores e interações significativos, num intervalo de
confiança superior a 90%, conduziu-se a análise de variância do modelo (ANOVA).
Na Tabela 5. 16, encontram-se os parâmetros analisados para as variáveis e
interações significativas para o processo estudado.
82

Resultados & Discussões
Tabela 5.16 - Parâmetros de Análise de Variância para os Efeitos Significativos ao Processo de Extração de [3-xilosidase por Micelas Reversas de CT AB
Efeitos SQ GL MQ F p
A (pH) 1642,48 1 1642,48 218,49 0,004*
B (Cone. Tensoativo) 225,53 1 225,53 30,00 0,032*
E (Conc.Butanol) 313,91 1 313,91 41,76 0,023*
AB 426,32 1 426,32 56,71 0,017*
AE 477,31 1 477,31 63,50 0,015*
BC 382, 11 1 382, 11 50,83 0,019*
BE 248,30 1 248,30 33,03 0,029*
DE 336,26 1 336,26 44,73 0,021 *
Falta de Ajuste 411,98 8 51,50 6,85 0,133
Erro Puro 15,04 2 7,52
Total 4479,23 18 *significativo ao nível de 95% de confiança
GL=graus de liberdade, SQ=soma quadrática, MQ=média quadrática R2 = 0,905
As superfícies de resposta das interações significativas entre as variáveis
avaliadas no planejamento experimental para extração de í3-xilosidase por sistema
de micelas reversas de CT AB permitem visualizar que os valores de recuperação
são aumentados quando se utiliza o nível superior para CTAB (+1),
independentemente das outras variáveis. Na Figura 5. 7a, na interação da variável
concentração de CTAB com a variável pH os melhores valores de recuperação são
obtidos com uso destas variáveis em seu nível superior (+1). Tal fato evidencia mais
uma vez, que as forças agindo no processo de extração são governadas por
interações eletrostáticas (pH de extração>pI da enzima) e que também a enzima í3-
xilosidase é extraída da fase aquosa para o interior da micela reversa, com o auxílio
de micelas reversas que possuem tamanho e quantidade suficiente para englobar a
enzima. Desta forma, se justifica o uso de concentração superior de tensoativo.
83

Resultados & Discussões
A concentração de butanol (E) também apresentou efeito principal significativo
(estimativa de 8,86) sobre a recuperação da enzima, sugerindo que o aumento de
sua concentração promove um aumento no valor da variável resposta. Foi
constatada ainda a existência de interações significativas da variável butanol com pH
e CTAB (Figura 5.7b e 5.7d), novamente sugerindo melhora nos valores de
recuperação da enzima estudada com o uso de pH e concentração de butanol em
seus níveis superiores 8,0 e 20%, respectivamente. Micelas reversas formadas a
partir de tensoativos catiônicos, no geral, apresentam pequenos diâmetros, sendo
necessário o uso de co-solventes que auxiliam a formação do núcleo interno destas
micelas reversa. Eles agem como soluções salinas, provocando uma repulsão entre
as partes hidrofóbicas do tensoativo e aumentando o tamanho das micelas reversas.
No caso a p-xilosidase que é uma enzima com massa molar da ordem de 1 OOkDa,
necessita-se de micelas reversas de raios maiores (aproximadamente 5nm) (KREI &
HUSTEDT, 1992; KREI & HUSTEDT, 1989).
A Figura 5. 7c apresenta a superfície de resposta referente à interação entre
condutividade elétrica (C) e da concentração de CTAB. Verifica-se um aumento na
variável resposta com o aumento da concentração de CTAB e com a diminuição da
condutividade elétrica. Estes resultados estão de acordo com a literatura, pois
elevada condutividade, ou seja, elevadas concentrações salinas, poderiam provocar
o rompimento das formações micelares e ainda dificultar o processo de reextração
da enzima. (PESSOA JR & VITOLO, 1999; KREI & HUSTEDT, 1992).
Com relação à interação entre a concentração dos co-solventes, hexano! e
butanol (DE), observa-se, na Figura 5.7e, a existência de efeito antagônico entre
eles. Ou seja, para que seja favorecida a extração da enzima p-xilosidase por
micelas reversas de CT AB nas condições averiguadas, deve-se optar pelo uso de
um dos co-solventes em seu nível superior (+1) e do outro em seu nível inferior (-1).
No entanto, nota-se que, para esta interação, houve um aumento maior na
recuperação da enzima com o aumento da concentração de butanol. Assim, o uso da
variável concentração de butanol em seu nível superior (20%), em detrimento do uso
do hexanol, o qual deve ser usado em seu nível inferior (5%), aumentariam o valor
84

Resultados & Discussões
da recuperação da enzima em cerca de 45% (a recuperação passaria de 20% para
29%).
A validade do modelo linear foi verificada pela análise de variância de
regressão para o modelo. Na Tabela 5.17 é apresentado o teste F para o modelo.
Tabela 5.17 - Análise de variância de regressão para o modelo representativo do processo de extração de P-xilosidase por micelas reversas de CTAB, na região em estudo
Causa de SQ GL MQ F
Variação
Modelo 4052,22 8 506,53 11,86
Erro 427,02 10 42,70
Total 4479,24 18
R = 0,905 GL=graus de liberdade, SQ=soma quadrática, MQ=média quadrática
Analisando a Tabela 5.17 verifica-se que todos os fatores são significativos a
5% de probabilidade. Através do teste F, ou seja, comparando-se o valor de F do
modelo (F=11,86) com o valor de Ftabelacto (Ftab=3,35) fica evidente a significância do
modelo.
Com base nos resultados discutidos até o momento, foi possível obter uma
equação para o modelo linear, tendo a finalidade de descrever a extração da enzima
p-xilosidase por micelas reversas de CTAB, para região em estudo, (Equação 5.3):
Y = 11,69 + 10,13A + 3,758 + 4,43E + 5,16AB + 5,46AE • 4,89BC - 3,94BE - 4,58DE
Onde: Y = recuperação da enzima (%)
A=pH;
B = CTAB;
C = Condutividade elétrica;
D= Concentração de hexano!;
E = Concentração de butanol.
Equação 5.3
85

Resultados & Discussões
Para validação do modelo, o qual prevê valores de recuperação de 44%,
foram realizados experimentos no ponto indicado pela análise estatística: pH 8,0,
concentração de CTAB 0,2 M, condutividade elétrica 4mS/cm, concentração de
hexanol de 5% e concentração de butanol 20%. O resultado médio alcançado foi de
38, 7 % de recuperação para enzima !3-xilosidase, mostrando a adequação do modelo
aos resultados experimentais.
5.4.1 • Otimização do Processo de Extração por Mice/as Reversas de CTAB em lsooctano, Hexano/ e Butano/
Tendo em vista as constatações feitas anteriormente, o passo seguinte para
buscar a otimização do processo de extração de 13-xilosidase por micelas reversas de
CTAB foi a busca de uma região ótima obtida com o uso da metodologia de
superfície de resposta. Esta metodologia prevê o deslocamento da região analisada
através do uso de um planejamento na região vizinha.
Neste caso, para buscar-se a otimização do processo de recuperação da
enzima 13-xilosidase por micelas reversas de CTAB, propôs-se primeiramente um
passo ascendente, com o intuito de se definir a nova região a ser estudada.
Devido às limitações da enzima quanto ao pH de solubilização, tal variável foi
fixada em seu nível superior (pH 8,0).
Desta forma, foram alterados os níveis e valores, segundo um passo
ascendente os fatores: concentração de CT AB e concentração de Butanol. Na
Tabela 5.18 encontram-se os novos valores utilizados e os rendimentos de
recuperação obtidos.
86

Resultados & Discussões
Tabela 5.18 - Níveis Codificados e Valores Utilizados na Realização dos Ensaios do Passo Ascendente no Processo de Extração de f3-xilosidase por Micelas Reversas de CTAB
Valores Recuperação(%)
CTAB (M) Butanol (%)
0,2
0,225
0,25
0,30
20,9
23,8
26,8
32,7
38,6
31,8
45,74
18,6
Após a definição da nova região optou-se por utilizar um planejamento em
estrela, uma vez que este é mais abrangente que o de face centrada, podendo ser
portanto, mais eficiente na procura da resposta otimizada.
Assim, um planejamento ortogonal 22 completo com 03 repetições no ponto
central (a=1,41) foi utilizado. A distribuição ortogonal resultante deste planejamento,
consiste da agregação ao planejamento já existente um outro, idêntico, porém com
inclinação de 45° em relação à orientação de partida (Figura 5.8).
+21'2
+1
o
-1 -21'2
-·· r_·_-_._._._. : ~----------.-.i ,,r ~~~~
•. l • i .•
-, 1:::::,.,:- .... _/::::1·· .,.. -21'2 -1 o +1 +21'2
Figura 5.8 - Esboço do Planejamento em Estrela Ortogonal.
87
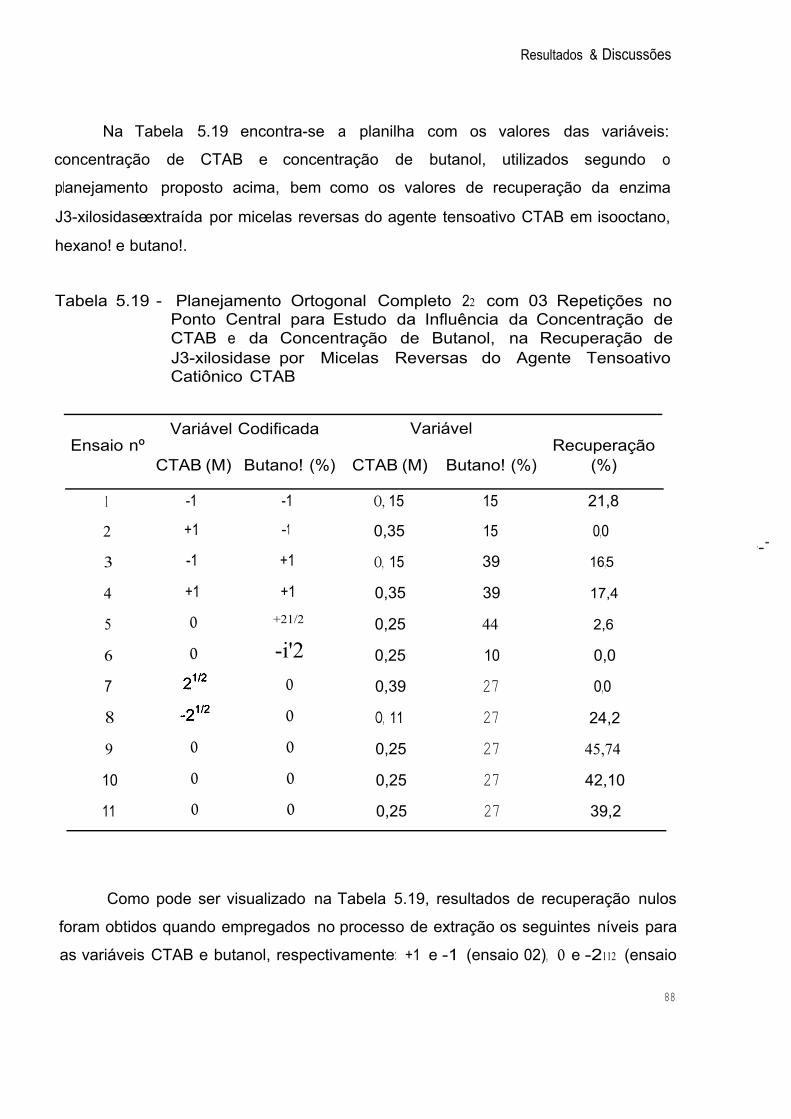
Resultados & Discussões
Na Tabela 5.19 encontra-se a planilha com os valores das variáveis:
concentração de CTAB e concentração de butanol, utilizados segundo o
planejamento proposto acima, bem como os valores de recuperação da enzima
J3-xilosidase extraída por micelas reversas do agente tensoativo CTAB em isooctano,
hexano! e butano!.
Tabela 5.19 - Planejamento Ortogonal Completo 22 com 03 Repetições no Ponto Central para Estudo da Influência da Concentração de CTAB e da Concentração de Butanol, na Recuperação de J3-xilosidase por Micelas Reversas do Agente Tensoativo Catiônico CTAB
Variável Codificada Variável Ensaio nº Recuperação
CTAB (M) Butano! (%) CTAB (M) Butano! (%) (%)
1
2
3
4
5
6
7
8
9
10
11
-1 -1
-1
o o o
o o o o o
O, 15
0,35
O, 15
0,35
0,25
0,25
0,39
O, 11
0,25
0,25
0,25
15
15
39
39
44
10
27 27 27 27 27
21,8
0,0
16,5
17,4
2,6
0,0
0,0
24,2
45,74
42,10
39,2
- ·- +1
-1 +1
+1
o o
+1 +21/2
-i'2
Como pode ser visualizado na Tabela 5.19, resultados de recuperação nulos
foram obtidos quando empregados no processo de extração os seguintes níveis para
as variáveis CTAB e butanol, respectivamente: +1 e -1 (ensaio 02), o e -2112 (ensaio
88

Resultados & Discussões
06), +i12 e O (ensaio 07) Observa-se que tais ensaios têm em comum a utilização de
nível mais elevado para o CTAB (A) em detrimento do butanol (B). Assim conclui-se
que a concentração do tensoativo, nos três casos, é excessiva para concentração de
butanol presente, ocasionando provavelmente desaparecimento da fase micelar, que
existe numa determinada região de equilíbrio para os diferentes componentes que
formam o sistema. A Figura 2.9 (Revisão Bibliográfica) ilustra as mudanças ocorridas
nos sistemas micelares quando da modificação das proporções entre seus
componentes. No entanto, tal fato somente poderia ser confirmado com a utilização
do diagrama de fases dos componentes do sistema: CT AB, isooctano, hexanol e
butanol. Porém este diagrama não encontra-se disponível na literatura.
Por outro lado, nas extrações realizadas nas condições definidas pelo ponto
central (CTAB 0,25 e 27% de Butanol) a recuperação da enzima í3-xilosidase
alcançou valores médios de 40%. Isto sugere a existência de efeitos quadráticos
para ambas as variáveis avaliadas (CTAB e butanol), e que o ponto máximo de
recuperação encontra-se na região estudada.
Quando utiliza-se tensoativos catiônicos, o transporte de enzimas para o
interior das micelas reversas é favorecido quando estas possuem carga elétrica
negativa, o que é alcançado com o uso de valores de pH superiores ao pi da enzima
(pi í3-xilosidase entre 4,9 e 6,0). Portanto espera-se que com o uso de agentes
tensoativos catiônicos a enzima migre para o interior das micelas reversas quando o
pH do meio é superior ao seu pi. Nos ensaios em análise usou-se pH 8,0, tendo sido
portanto, a transferência da enzima para a fase micelar governada por interações
eletrostáticas. Desta forma, o uso de concentrações elevadas de CT AB se faz
necessário de modo a se obter micelas maiores, capazes de englobar a í3-xilosidase
(100kDa).
A análise de variância para os parâmetros estudados encontra-se na Tabela
5.20.
89

Resultados & Discussões
Tabela 5.20 - Parâmetros de Análise de Variância para os Efeitos Significativos ao Processo de Extração de (3-xilosidase por Micelas Reversas de CTAB
Causa de SQ GL MQ F p
Variação
pH (A) 75,50 1 75,50 7,03 O, 117 CTAB (B) 268,24 1 268,24 24,98 0,037*
AB 128,82 1 128,82 12,00 0,074*
AA 1978,32 1 1978,32 184,23 0,005*
BB 1001,47 1 1001,47 93,26 0,011 *
Falta de Ajuste 202,67 3 67,56 6,29 0,140
Erro Puro 21,48 2 10,74
Total 3052,30 10 R2=0,93 *95% de significãncia GL=graus de liberdade, SQ=soma quadrática, MQ=média quadrática
Observa-se, que o modelo não possui falta de ajuste significativo para o
intervalo de confiança considerado e que existe interação quadrática (AA e BB)
significativa entre os parâmetros avaliados. Assim, os parâmetros de um modelo de
segunda ordem (quadrático) usados para estimar a porcentagem da enzima
recuperada em função do CT AB e butanol foram obtidos através da análise de
regressões múltiplas, mostrada na Tabela 5.21.
Tabela 5.21 - Variáveis, Coeficientes, Teste t (Student), Valores de p do Planejamento Completo Ortogonal 22 com 03 Repetições no Ponto Central para Extração de (3-xilosidase por Micelas Reversas de CTAB
Variáveis Coeficientes t p
Média 42,35 10,97 <0,005 CTAB (A) -3,07 -0,65 O, 12
Butano! (B) 5,79 1,22 0,03 AB 5,67 0,84 0,07 AA -18,72 -3,32 0,005 BB -13,32 -2,36 0,01
90

Resultados & Discussões
A significância estatística do modelo (Tabela 5.22) foi avaliada pelo teste F e
revelou que esta regressão é estatisticamente significativa (Fmode1o>Ftabelado=5,05). O
modelo não apresenta falta de ajuste e o coeficiente de determinação (R2 = 0,93)
indica que 93% das variações na recuperação da enzima por este sistema podem
ser explicadas pelo modelo.
Tabela 5.22 - Parâmetros da Análise de Variância (ANOVA) dos Fatores Significativos ao Processo de Extração de J3-xilosidase por Micelas Reversas de CTAB
Fonte de SQ GL MQ F Variação Modelo 3452,35 5 690,47 15,40
Erro 224, 15 5 44,83
Total 3676,50 10
GL=graus de liberdade, SQ=soma quadrática, MQ=média quadrática
Assim, um modelo matemático que represente a recuperação de !3-xilosidase
por micelas reversas de CTAB (em isooctano e hexanol), pode ser expresso pela
Equação 5.4:
Y = 42,35-3,07A + 5,798 + 5,67A8-18,72A2 -13,3282 Equação 5.4
Onde: Y = recuperação da enzima(%);
A = concentração de CTAB;
B = concentração de Butano!.
O máximo valor de recuperação de J3-xilosidase, 43%, corresponde ao ponto
definido pela concentração de CTAB igual a 0,26 M (A = -0,05) e concentração de
butanol de 29,4% (B = 0,2). A Figura 5.9 corresponde à superfície de resposta obtida
usando os valores ótimos previstos para as variáveis.
Ensaios nas condições ótimas previstas, foram feitos com o intuito de se
validar o modelo. O valor médio obtido para a recuperação foi de 44,3%, confirmando
assim o valor previsto pelo modelo que foi de 43%.
91

Resultados & Discussões
Figura 5.9 - Superfície de Resposta Descrita pelo Modelo da Equação 5.4, que Representa a Extração em Condições Otimizadas de f3-xilosidase por Micelas Reversas do Agente Tensoativo Catiônico CTAB em lsooctano, Hexanol e Butanol.
A extração da enzima f3-xilosidase por micelas reversas de CTAB, nas
condições otimizadas previstas pelo modelo obtido, proporcionou um aumento de
pureza de 5,34 vezes para a enzima extraída para FAII (em relação ao precipitado
solubilizado), como pode ser visto na Tabela 5.23.
Tabela 5.23 - Aumento de Pureza para Enzima f3-xilosidase
Atividade Específica U/mg
Ae ( após a extração) Ae ( antes da extração)
21,58 4,04
Ap 5,34 Ae = atividade específica; Ap = aumento de pureza
Os resultados mostram mais uma vez ser possível, com a utilização de um
método simples a extração por micelas reversas, o isolamento de enzimas de
92

Resultados & Discussões
interesse, sem o inconveniente das várias etapas utilizadas nos processos
convencionais.
5.4.2- Determinação do Raio Mice/ar
Foi determinado o raio das micelas reversas no ponto de ótimo do
planejamento, utilizando as seguintes condições experimentais: pH 8,0,
concentração de hexanol 1 O %, condutividade elétrica de 4 mS/cm, temperatura de
26ºC, concentração de CTAB de 0,26M e concentração de butanol de 29,4%.
Na Tabela 5.24 encontram-se os raios micelares obtidos. O raio médio para as
micelas reversas formadas sob as condições experimentais anteriormente citadas. O
valor apresentado é superior a 5nm, valor indicado por KREI & HUSTEDT (1992)
para que uma proteína de 100 kDa possa ser englobada. Como a 13-Xilosidase
possui massa molar nesta ordem de grandeza (100kDa), o raio micelar permitiu o
englobamento da enzima contribuindo para o processo de extração da enzima, o que
reforça a existência de interações eletrostáticas agindo no processo de extração
desta enzima.
Tabela 5.24 - W0 e Rm Determinados na FAI E FAII Obtidas após Extração por Micelas Reversas de CT AB a 0,26M, com 10% de Hexanol e 29,4% de butanol, em pH 8,0, Condutividade Elétrica de 4 mS/cm, Temperatura de 26ºC
Fase da Extração Rm (nm)
FMI
FMII
85,13
11,00
13,95
1,80
Da mesma forma que o raio das micelas reversas presentes na FMI é suficiente para o englobamento da enzima, na FMII o valor de 1,80 nm mostra ser
improvável que a enzima {3-xilosidase possa estar contida no interior das micelas
reversas, indicando que o processo de reextração da enzima para a FAII foi
eficiente.
93

Resultados & Discussões
5.5 - EXTRAÇÃO DA ENZIMA (3-XILOSIDASE POR MICELAS REVERSAS DE BDBAC E CTAB, DIRETAMENTE DO MEIO FERMENTADO
Uma vez obtidas as condições otimizadas de extração da enzima f3-xilosidase
pré purificada por precipitação fracionada com etanol, utilizando micelas reversas de
BDBAC e CTAB, partiu-se para utilização dessas condições na condução da
extração da enzima diretamente do meio fermentado.
Tais experimentos visaram confirmar a possibilidade do uso da extração por
micelas reversas como uma única etapa; uma vez que a cada etapa de DSP que se
adiciona ao processo de recuperação de um bioproduto, agrega-se custo ao produto
final devido às perdas inevitáveis em cada etapa e a conseqüente diminuição do
rendimento final, como pode ser observado na Figura 5.1 o.
% Final 100
20
Rendimento da Etapa 80
60
40
1 2 3 4 5 6 7 8 9 10 Nº de Etapas de DSP
Figura 5.10 - Efeito do Número de Etapas de DSP no Rendimento Final do Bioproduto (Adaptado de PESSOA JR., 1997).
Os resultados de extração da enzima (3-xilosidase diretamente do meio
fermentado constam da Tabela 5.25.
94

Resultados & Discussões
Tabela 5.25 - Comparação entre a Porcentagem de Recuperação da Extração por Micelas Reversas de BDBAC e CT AB de f3-xilosidase sem Precipitação e com Precipitação por Etanol
Recuperação Recuperação Tensoativo Condições de Extração % %
sem precipitação com precipitação
Concentração de Tensoativo = 0,2M
Concentração de Hexano! = 10% 43 BDBAC 8,5
Temperatura= 24ºC
pH 3,3
Concentração de Tensoativo = 0,26M
Concentração de Hexano! = 10% 44,3 CTAB Concentração de Butano! = 29 ,4 % 35
(aproximadamente) Temperatura= 26ºC pH 8,0
Nos ensaios realizados sem prévia precipitação com etanol, não foi possível
incluir a variável condutividade elétrica em seu nível previsto nos modelos
otimizados, ou seja, em 4,0 mS/cm. Como objetivava-se estudar a viabilidade do uso
da técnica de extração líquido-líquido por micelas reversas em uma única etapa de
DSP, não fez-se a diálise da amostra contendo 13-xilosidase. Assim, a condutividade
elétrica utilizada nestes ensaios foi de 14,98 mS/cm.
A elevada condutividade elétrica do meio, apresenta-se como um obstáculo à
extração de f3-xilosidase por micelas reversas de BDBAC. Como anteriormente
proposto, a extração desta enzima no sistema formado por este agente tensoativo
catiônico ocorre via interações hidrofóbicas. Ao que tudo indica, a elevada
concentração de íons em solução promove o enfraquecimento destas forças inibindo
a extração da enzima, pois grande parte da atividade enzimática é encontrada na
FAI, ou seja, não foi transferida para o interior das micelas reversas.
Por outro lado, os resultados de recuperação da enzima 13-xilosidase por
micelas reversas formadas a partir do agente tensoativo CTAB, mostraram ser
possível a utilização da técnica de extração por micelas reversas em uma única
95

Resultados & Discussões
etapa. Os resultados alcançados para o sistema sem pré purificação da enzima
(35%) são semelhantes aos obtidos com a prévia precipitação com etanol (da ordem
de 40%). Para este sistema, a condutividade elétrica do meio não interferiu no
processo de extração. Esta "imunidade" do sistema micelar à elevada condutividade
elétrica pode ser ocasionada pela elevada concentração de solventes na
microemulsão, que auxiliam na formação do centro micelar com maiores diâmetros
(HATION, 1989). Assim extrações governadas por interações eletrostáticas (como no
caso do CTAB), não seriam influenciados pela força iônica da fase aquosa inicial,
uma vez que o diâmetro do núcleo interno da micela obtida com o uso de dois co-
solventes (hexano! e butano!) seria suficiente para o englobamento da enzima.
96

Conclusões
6 • CONCLUSÕES
Os resultados alcançados no presente trabalho, levaram às conclusões que se
seguem:
1. A enzima (3-xilosidase produzida pelo fungo P. janthinellum apresentou, nas
condições averiguadas na presente dissertação, estabilidade frente à estocagem
sob refrigeração a -4ºC e 4°C, na faixa ampla de pH de 3,0 a 7,0, por 96 dias.
2. Para as condições de extração por micelas reversas estudadas no presente
trabalho, os agentes tensoativos catiônicos BDBAC e CT AB, promovem
resultados satisfatórios de recuperação (em atividade) para a enzima í3-xilosidase.
No entanto o agente tensoativo aniônico AOT (em isooctano), não mostrou-se
efetivo ao processo nas condições averiguadas: concentração de 0, 1 O a 0,2 M,
temperatura de 4° a 26ºC, condutividade elétrica de 4,0 a 10 mS/cm e pH de 3,0 a
9,0.
3. Com o uso do agente tensoativo BDBAC, construi-se um modelo matemático
otimizado, que prevê valores de recuperação para enzima í3-xilosidase de 42%,
sendo que nos ensaios para confirmação do modelo foi observado 43% de
recuperação.
4. Com a otimização do processo de extração de í3-xilosidase por micelas reversas
do agente tensoativo CT AB, o modelo obtido prevê valores de recuperação para
97

Conclusões
enzima de 43%, tal valor foi confirmado experimentalmente, onde se obteve
44,3% de recuperação.
5. O agente tensoativo catiônico CTAB, utilizado num dos sistemas de extração por
micelas reversas estudados no presente trabalho, ocasionou um aumento na
atividade específica da enzima de 260% maior que o com o uso do agente
tensoativo BDBAC.
6. Os resultados obtidos na extração de 13-xilosidase por micelas reversas do agente
tensoativo catiônico CT AB diretamente do meio fermentado, sem pré purificação
por etanol, mostrou mais uma vez ser este processo, viável para a extração direta
do meio fermentado, diminuindo portanto etapas dispendiosas de DSP.
7. As extrações feitas com o agente tensoativo CTAB mostraram que o efeito da
força iônica do meio sobre a extração da enzima !3-xilosidase é menor quando se
utiliza dois co-solventes (hexanol e butanol), pois a força iônica não afeta
significativamente o diâmetro interno da micela reversa.
8. Por tudo o que foi exposto acima, conclui-se portanto que a extração por micelas
reversas oferece uma alternativa para purificação de enzimas bastante
satisfatória: consiste de uma técnica de desenvolvimento simples, onde os
reagentes e equipamentos utilizados são comuns aos laboratórios.
98

Referências Bibliográficas
7 - REFERÊNCIAS BIBLIOGRÁFICAS
AIRES-BARROS, M.R., CABRAL, J.M.S. Selective separation and purification of two Lipases from Chromobacterium viscosum using AOT reversed micelles. Biotechnology and Bioengineering, NewYork, v.38, p. 1302-1307, 1991.
ANDREWS, B.A., PYLE, D.L., ASENJO, J.A. The effect of pH and ionic stregh on partitioning of four proteins in reverse micelle systems. Biotechonology and Bioengineering, v. 43, p. 1048-1052, 1994.
ANDREWS, B.A.,HAYWOOD, K. Effect of pH, ion type and ionic stregh on partiong of proteins in reverse micele systems. Journal of Chromatography, v. 688, p.55- 60, 1994.
ASENJO, J.A. Separation Processes in Biotechnology. New York: Marcel! Dekker, 1990. 801p.
BACHMAN, S.L., McCARTHY, A.J. Purification and characterization of a thermostable p-xylosidase from Thermonospora fusca. Journal Gen. Microbial., v.135, p293-299, 1989.
BAILEY, M.J, BIEL Y, P., POUTANEN, K. lnterlaboratory testing of methods for assay of xylanase activity. Journal of Biotechnology, v.23, n.3, p.257-270, 1992.
BIELY, P. Microbial xylanolitic systems. Trends in Biotechnology, v. 3, p. 286-290, 1985.
99

Referências Bibliogrãficas
BOX, G.E.P., HUNTER, G.W., HUNTER, J.S. Statistics for experimenters - An introduction to design. data analysis, and model building. New York: John Wiley & Sons, 1978. 653p.
BRADFORD, M. M. A rapid and sensitiva method for the quantitation of microgram quantities of protein utilizing the principie of protein-dye binding. Analytical. Biochemistry, v. 72, p. 248-54, 1976.
BRANDANl,V., DI GIACOMO, G., SPERA, L. Extraction of a-amilase protein by reverse micelles. Process Biochemistry, v.28, p.9-15, 1991.
SRICE, R.E., MORRISON, I.M. Effect of the addition of D-xylose on xylanase activity and digestibility of fiber in an artificial rumen Applied Biochemistry and Biotecnology, v.30, p.9-15, 1991.
CASTRO, M.J.M, CABRAL, J.M.S. Reversed micelles in biotechnological processes. Biotechnology Advances, New York, v.6, p.151-167, 1988.
CHANG, Q.-L., CHEN, J.-Y. Selective recovery of two enzymes from Bacillus subtílís using Aliquat 336 reversed micelles. Applied Biochemistry and Biotechnology, v.56, p. 197-204, 1996.
' -- ' --
CLARKSON, A.I., BULMER, M., TITCHENER-HOOKER, N.J. Pilot-scale verification of a computer-bsed simulation for fractional protein purification. Bioprocess Engineering, v.14, n.2, p.69-80, 1996.
CORTEZ, E. V. Recuperação de enzimas xilanolíticas por precipitação. Lorena- SP, FAENQUIL 77p, 1998 (Dissertação de Mestrado).
CORTEZ, E.V., PESSOA JUNIOR, A, ASSIS, A.N. Xylanase recovery by ethanol and Na2S04 precipitation. Applied Biochmistry and Biotechnology, v. 70-72, 1998.
CORTEZ, E.V., PESSOA JUNIOR, A. Xylanase and J3-xylosidase separation by fractional precipitation. Process Biochemistry, v.35, p.277-283, 1999.
COSTA, S.A. Extração líquido-líquido de xilanase em sistema de duas fases aquosas. Lorena-SP, FAENQUIL 93p., 1998 (Dissertação de Mestrado).
100

Referências Bibliogrãficas
COSTA, S.A.; PESSOA JUNIOR, A.; ROBERTO, I.C. Xylanase recovery: effect of extraction conditions on the aqueous two-phase systems using experimental design. Applied Biochmistry and Biotechnology. v.70/72, p.629-639, 1998.
COUGHLAN, M.P.; HAZLEWOOD, G.P. 13-1,4-D-xylan - degrading enzyme systems: biochemistry molecular biology and applications, Biotechnology Applied Biochemistry, v. 17, p. 259-289, 1993.
CHRISTOV, L.P.; PRIOR, B.A. Repeated treatments with Aureobasidium pullulans hemicellulases and alkali enhace biobleanching of sulfite pulps Enzime Microbiology and Technology, v. 18, n.4, p. 244-250, 1996.
DEKKER, R.F.H, RICHARDS, G.N. Hemicellulase: their occurence, purification, properties and mode of action. Advances Carbohydrate Chemistry Biochemistry, v.32, p.277-352, 1976.
DEKKER, R.F.H. The enzymatic hydrolisis of biomass. ln: SEMINÁRIO DE HIDRÓLISE ENZIMÁTICA DE BIOMASSA, v.2. Maringá, 1985.
DEKKER, M.; HILHORST, R.; LAANE, C., lsolation enzymes by reversed micelles. Anallytical Biochemistry, 178, 217-26, 1989.
DESHPANDE, V., LACHKE, A., VALADE, J.L. The use kraft pulp bleaching: a rewiew. Tappi Journal, v.77, n.6, p.125-131, 1994.
DOBBERSTEIN, J., EMEIS, C.C. Purification and characterization of 13-xylosidase from Aerobasidium pullulans. Applied Microbiology and Biotechnology, v. 56, p43-50, 1978.
DUNGAN, S.R., BAUSCH, T., HATTON, T.A., PLUCINSKI, P. NITSCH, W. lnterfacial transport processes in the reversed micellar extraction of proteins, Journal of Colloid and Interface Science, v. 145, n.1, p. 33-50, 1991.
ENZITEC, 1999, Folheto de apresentação do IV ENZITEC: Seminário Internacional de Tecnologia Enzimática, UFRJ, Rio de Janeiro, Outubro de 1999.
GARCIA, C.A. Protein Precipitation ln: Recovery Processes from Biological Material. Edited by John F. Kennedy and Joaquim M.S. Cabral, John Wiley & Sons, New York (1993)
101

Referências Bibliogrãficas
GILBERT, M., BREU, C., YAGUCHI, M., SADDLER, J.N. Purification and characterization of a xylanase. Applied Biochemistry and Biotechnology, v.34/35, p.247-59, 1992.
GLATZ, C.E. Precipitation. ln: ASENJO, J.A. Separation Processes in Biotechnology. New York: Marcel Dekker, 1990, p.329-56.
GIOVENCO, S., VERHEGGEN, F. Purification of intracellular enzymes from whole bacterial cells using reversed micelles. Enzyme Microbiology and Technology, v. 9, p.470-473, 1987.
HATTON, T.A. Extraction of proteins and amino acids using reversed micelles. Washington: American Chemical Society [ACS Symposium Series], 1987.
HENTSCH, M., MENOUD, P., STEINER, L., FLASCHEL, E., RENKEN, A. Optmization of the surfactant (AOT) concentration in a reverse micellar extration process. Biotechnlogy Technology, Keue, v.6, n.4, p.359-354, 1992.
HILHORST, R., SEGEEVA, M., HEERING, D., RIETVELD, P., FIJNEMAN, P., WOLBET, R.B.G., DEKKER, M., BIJSTERBOSH, B.H. Protein extraction from aqueous phase into a reversed micellar phase: effect of water content and reversed micellar composition. Biotechnology and Bioengineering, n.46, v.4, p.375-387, 1995.
HUANG, S-Y., LEE, Y-C. Separation and purification of herseradish perosidase from Armoracia rusticana root using reversed micellar extraction. Bioseparation, Dordrecht, v.4, p.1-5, 1994.
ISHIKAWA, Y., HAMASAKI, T., UEDA, Y., FUJIWARA, T. Aurantionone, a potent antioxidant and synegist from Penicillium aurantio-virens, Bioscience Biotechnology and Biochemistry, n.56, v.9, p.1486-1487, 1992.
JANES, R. L. The chemistry of wood and fibers. ln: Mac Donald, R.G.; Franklin, J.N. the Pulping ofWood. NewYork, McGraw-Hill, p. 33, 1969.
JEFFRIES, T.W., Utilization of xylose by bacteria, yeasts, and fungi. Advances Biochemical Engineering, v.27, p.1-32, 1983.
JOSELEAU, J. P. COMTAT, J., RUEL, K. Chemical structure of xylans and their
102

Referências Bibliográficas
interaction in the plant cell walls. ln: VISSER, J., BELDMAN, G., KUSTERS- van SOMEREN, M.A., VORAGEN, A.G.J. (eds.). Xylans and Xylanases. Amsterdam: Elsevier, 1992, p.1-15. (Progress in Biotechnology, 7).
KADAM, K. 1. Reverse micelles as a bioseparation toai. Enzyme Microbiology and Technology, v. 8, p. 266-273, 1986.
KERSTERS-HILDERSON, H., LOONTIENS, F.G., CLAEYSSENS, M., DE BRUYNE, C. K. Partial purification and some properties of na induced f3-D-xylosidase of Bacillus pumilus 12. European Journal Biochemistry, v. 7, p.434-441, 1969.
KREI, G.A., HUSTEDT, H. Extraction of enzymes by reverse micelles. Chemical Engineering Science, New York, v.47, n.1, p.99-111, 1992.
KREI, G.A., HUSTEDT, H. Extraktion von enzymen mittels reversar mizellen. Bioengineering, v.3/4, p.32-41, 1989.
KREI, G., MEYER, U., BÔRNER, B., HUSTEDT, H. Extraction of a-amylase using BDBAC reversed micelles. Bioseparation, v.5, p. 175-183, 1995.
KRIEGER, N. Produção, purificação e caracterizaçao de lipase de Penicillium citrinum. Curitiba-PR, Universidade Federal do Paraná, Bioquímica, 1995. (Tese de Doutorado).
KRIJGSMAN, J. Downstream Processing: lntroduction. Delf-Leiden: lnstitute for Biotechnology Studies, 1991. (Apostila de curso).
KUMAR, S. & RÁMON, D. Purification and regulation of the synthesis of a f3- Xylosidase from Aspergillus nidulans. FEMS Microbiology Letters. 135 (1996) 287-293.
LEHNINGER, A.L. Bioquímica. São Paulo: Edgard Blücher, 1976. p.126-145. L YE, G. J., ASENJO, J. A., PYLE, D. L., Extraction of lysozyme and ribonuclease-a
using reversed micelles: limits to protein solubilization. Biotechnology and Bioenginnering,V. 47, P. 509-519, 1995.
LOWRY, O. H., ROSEBROUGH, N. J., FARR, A . L., RANDALL, R. J., Protein measurement woth the Folin-phenol-reagent, Journal Biotechnology, v. 193, p. 265-275, 1959.
LUISI, P.L., GIOMINI, M., PILENI, M.P., ROBINSON, B.H. Reversed micelles as hosts for proteins and small molecules. Biochimica et Biophysica Acta, Amsterdam, v.947, p.209-246, 1988.
103

Referências Bibliográficas
MARCOZZI, G., CORREA, N., LUISI, P.L., CASELLI, M. Protein extraction by reverse micelles: A Study of the factors affecting the forward and backward transfer of a- chymotrypsin and its activit. Biotechnology and Bioengineering, v. 38, p. 1239- 1246, 1991.
MATSUO, M., YASUI, T. Purication and some properties of j3-xylosidase from Trichoderma viride. Agriculture Biology and Chemical., v. 48, p.1845-1852, 1984.
MEYER, U. Extraktion von Enzymen Mittels lnverser Mizellen. Braunschweig: Technische Universitãt Braunschweig, 1992, 58p. (Dissertação de Graduação).
MILAGRES, A.M.F. Alguns aspectos da regulação de 8-xilanases extracelulares de Penicillium ianthinellum. Viçosa, UFV, 1988 (Dissertação de Mestrado).
MILAGRES, A.M.F., LACIS, L.S. Efficient screening of process variables in Penicillium janthinellum fermentation Biotechnology Letters, v.13, n.2, p.113- 118, 1991.
MILAGRES, A.M.F., DURAN, N. Xylanolytic enzymes from Penicillium janthinellum and its applications in bleaching of pulp. Progress Biotechnology, v.7, p 539- 545, 1992.
MILAGRES, A.M.F. Produção de xilanases por Penicillium ianthinellum e aplicação das enzimas no branqueamento de polpas Kraft. Campinas: UNICAMP, 1994. 145p. (Tese de Doutorado).
MILAGRES, A.M.F., PRADE, R.A. Production of xylanases from Penicillium janthinel/um and its use in the recovery of cellulosic textile fibers. Enzyme and Microbial Technology, v.16, p.627-631, 1994.
MILLER, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar, Analytical Chemistry, v.31, p.426-428, 1959.
MUTSAERS, J.H.G.M. Xylanase in breadmaking. ln: XYLANS ANO XYLANASES INTERNATIONAL SYMPOSIUM Wageningen, The Netherlands, Novo nordisk, December 8-11, p.48, 1991.
PATEL, M.T., NAGARAJAN, R., KILARA, A. lnteractive influence and optimization of
104

Referências Bibliográficas
reaction parameters in the hydrolysis of olive and coconut triacylglycerols by C. cylindracea and R. javanicus lipases in reverse micellar media. Applied Biochemistry and Biotechnology, v. 59, p. 109-124, 1996.
PESSOA JUNIOR, A., Obtenção de lnulinase: Recuperação e Ampliação de Escala. São Paulo, USP, 1995 (Tese de Doutorado).
PESSOA JUNIOR, A., VITOLO, M. Recovery of inulinase using BDBAC reversed micelles. Process Biochemistry, v.33, n.3, p.291-97, 1998.
PESSOA JUNIOR, A., MANCILHA, I.M., SATO, S. Acid hydrolysis of hemicellulose from sugarcane bagasse. Brazilian Journal of Chemical Engeneering, v.14, n.3, p. 291-97, 1997.
PESSOA JUNIOR, A. Apostila de Aula. FAENQUIL, Lorena, 1997.
PIRES, M.J., AIRES-BARROS, M.R., CABRAL, J.M.S. Liquid-Liquid extraction of protein with reversed micelles. Biotechnology Progress, v.12, p.290-01, 1996.
PULS, J., CLAEYSSENS, M., VRANSKA, M., COUGHLAN, M.P., Production of exo-glycosidase enzymes by Talaromyces emersonii CBS814.70 and importance in the overall hydrolysis of cell wall polysaccharides. Abstracts, v.13, n.14, 1995.
PULS, J., POUNTANEN, K. Mechanisms of enzymic hydrolysis hemicelluloses (xylans) and procedures of determination of the enzyme activities involved. ln: COUGHLAN, M. P. (ed.). Enzyme Systems for Lignocellulose Degradation. London: Elsevier, 1989, p. 151-165.
RAMAHAN, R. S., HATTON, T. A. ln :LI, N.N., STRATHMAN, H., Separation Technology, New York, United Eng. Trustees, lnc. N.Y., 1988.
REGALADO, C., ASENJO, J.A., PYLE, D.L. Studus on the purification of peroxidase from horseradish roots using reversed micelles. Enzyme and Microbial Technology, v. 18, p. 332-339, 1996.
RIELL Y, P.J. Xylanases: structure and function. ln: HOLLAENDER, A.E.(ed.). Trends in the Biology of Fermentation for Fuels and Chemicals. New York: Plenum Press, 1981. P.111-129.
RISTROPH, D.L., HUMPHREY, A.E. The j3-xylosidase of Thermonospora. Biotechnology and Bioengineering, 27, p.909-913, 1985.
RODIONOVA, N. A.; TAVOBILOV, J. M.; BEZBORODOV, A. M. j3-xylosidase from A. niger 15: Purification and properties. J. Appl. Biochem.5, 300-312, 1983.
105

Referências Bibliogrãficas
RODRIGUES, E.M.G. Extração Líquido-líquido de Xilanase por Micela Reversa, Lorena, FAENQUIL, 1997 (Dissertação de Mestrado).
RODRIGUES, E.M.G.; PESSOA-JR, A.; MILAGRES, A.M.F., Screening of variables in xylanase recovery using BDBAC reversed Micelles, Applied Biochemistry and Biotechnology, v. 77-79, p. 779-788, 1999.
RUTTERSMITH, L.D., DANIEL, R.M. Termostable J3-glucosidase and J3-xylosidase from Thermotoga sp strain FJSS3-B.1. Biochim, Bioph. Acta, v.1156, p.167- 172, 1993.
SCAWEN, M.D., MELLING, J. Large-scale extraction and purification of enzymes and others proteins. ln: WISEMAN, A Handbook of Enzyme Biotechnology. West Sussex: Ellis Horwood, 1985, p. 15-48.
SCOPES, R.K. Protein Purification: principies and practice. New York: Springer- Verlag, 1994. 329p.
SEOUD, O.A., SANCTIS, D.S., PALMA, E.J. Tensoativos: química e aplicações em cosméticos e detergentes. São Paulo:ABQ, 1999.(Apostila do Curso de T ensoativos)
SHAMEL, R.E., KEOGH, M. Double-digt growth predicted for biotechnology products in next decade. Genetic Engeneering News, 6 dez, 1995.
SILVA, R.R.P., CORTEZ, E.V.; COSTA, S.A.; ROBERTO, I.C.; HASMANN, F.A.; RODRIGUES, R.C.L.B.; ASSIS, A.N.; PESSOA-JR, A., Influência do pH na extração líquido-líquido de J3-xilosidase por micelas reversas de agente tensoativo catiônico. XII SINAFERM - Simpósio Nacional de Fermentações, Uberlândia-MG, 04-07 /08/98.
SULISTYO, J., KAMIYAMA, Y., YASUI, T. Purification and some properties of Aspergillus pulverulentus J3-xylosidase with transxylosylation capacity. Journal of Fermentation and Bioengineering. V. 79, n.1, p. 77-22, 1995.
SUNNA, A., ANTRANIKIAN, G. Xylanolitic enzymes from fungi and bacteria. Criticai Reviews in Biotechnology, v. 17, n.1, p. 39-67, 1997.
TAMAMUSHI, B., WATANABE, N. The formation of molecular aggregation structures in ternary system: aerosol OT/water/iso-octane. Colloid & Polymer Science., v.258, p.174-78, 1980.
UZIIE, M., MATSUO, M., YASUI, TL Possible identify of J3-xylosidase and J3-glucosidase of Chaetomium trilaterale. Agriculture Biology Chemical, v.49, p.1167-1173, 1985.
106

Referências Bibliográficas
VAN DER BROECK, H.C. DE GRAAF, L.L., HILLE, J.D.R., VAN OOYEN, A.J.J., VESSER, J.HARDER, A Cloning and expression of fungai xylanase genes and use of the xylanase in bread making and in preparation of feed and paper products. European Patent Applied, v.90, p.202-220, 1990.
VAN-PEIJ-N-N-M-E, BRINKMANN, J., VRSANSKA, M., VISSER, J., GRAAFF, F. p-xylosidase activity, encoded by xinD, is essential for complete hydrolisis of xilan by Aspergillus niger but not for induction of xylanolytic enzime spectrum. European Journal of Biochemistry, v.245, n. 1, p.164-173, 1997.
VIIKARI, L., SUNDQUIST, J., KETTUNEN, J. Xylanases enzymes promete pul bleaching. Paper timber, v. 73, p.382-389, 1991.
VIIKARI, L. KANTELINEN, A, SUDQUIST, J., KLINKO, M. Xylanases in bleaching: from na idea to the industry. FEMS Microbiology Reviews, v.13, p.335-350, 1994.
VOGEL, H.J. A Convenient growth medium for Neurospora crassa (Medium N) Microbial Genetics Bulletin, v.13, p.42-43, 1956.
WIELAND, G. Water determination by Karl Fisher titration: theory and applications. Git Verlag, 1987.
WEBER, K., PRINGLE, J.R., OSBORN, M., Measurement of molecular weights by eletrophoresis on SDS-acrylamid gel. Methods in Enzymology, v.26, p. 3-27, 1972.
WONG, K.K.Y., TAN, L.U.L., SADDLER, J.N. Multiplicity of P-1,4-xylanase in microorganisms; Functions, Properties and Applications Microbiology Reviews, v.52, n.13, p. 305-317, 1988.
WOODWARD, J. Xylanases: Functions, properties and applications. ln: WISEMAN, A Topics in Enzyme and Fermentation Biotechnology, p. 9-30, 1984.
ZAMARRO, M.T., DOMINGO, M.J., ORTEGA, F., ESTRADA, P. Recovery of cellulolytic enzymes from wheat-straw hydrolysis broth with dioctyl sulphosuccinate/iso-octane reverse micelles. Biotechnology and Applied Biochemistry, v. 23, p. 117-125, 1996.
107


















