ENERGIA METABOLIZÁVEL DE ALIMENTOS E QUALIDADE … · energia metabolizÁvel de alimentos e...
113
ENERGIA METABOLIZÁVEL DE ALIMENTOS E QUALIDADE DE OVOS E CARNE DE CODORNAS JAPONESAS ALIMENTADAS COM RAÇÕES CONTENDO COLORÍFICO DO URUCUM E NIACINA SUPLEMENTAR NEWTON TAVARES ESCOCARD DE OLIVEIRA Tese apresentada ao Centro de Ciências e Tecnologias Agropecuárias da Universidade Estadual do Norte Fluminense, como parte das exigências para obtenção do título de Doutor em Produção Animal Orientador: Prof. José Brandão Fonseca CAMPOS DOS GOYTACAZES - RJ AGOSTO – 2004
Transcript of ENERGIA METABOLIZÁVEL DE ALIMENTOS E QUALIDADE … · energia metabolizÁvel de alimentos e...

ENERGIA METABOLIZÁVEL DE ALIMENTOS E QUALIDADE DE OVOS E CARNE DE CODORNAS JAPONESAS ALIMENTADAS COM
RAÇÕES CONTENDO COLORÍFICO DO URUCUM E NIACINA SUPLEMENTAR
NEWTON TAVARES ESCOCARD DE OLIVEIRA
Tese apresentada ao Centro de Ciências e Tecnologias Agropecuárias da Universidade Estadual do Norte Fluminense, como parte das exigências para obtenção do título de Doutor em Produção Animal
Orientador: Prof. José Brandão Fonseca
CAMPOS DOS GOYTACAZES - RJ AGOSTO – 2004

ENERGIA METABOLIZÁVEL DE ALIMENTOS E QUALIDADE DE OVOS E CARNE DE CODORNAS JAPONESAS ALIMENTADAS COM
RAÇÕES CONTENDO COLORÍFICO DO URUCUM E NIACINA SUPLEMENTAR
NEWTON TAVARES ESCOCARD DE OLIVEIRA
Tese apresentada ao Centro de Ciências e Tecnologias Agropecuárias da Universidade Estadual do Norte Fluminense, como parte das exigências para obtenção do título de Doutor em Produção Animal
Aprovada em 19 de agosto de 2004
Comissão Examinadora:
___________________________________________________________________ Prof. Rita da Trindade Nobre Soares (Doutor, Nutrição de Monogástricos) - UENF
___________________________________________________________________ Prof. Karla Silva Ferreira (Doutor, Tecnologia de Alimentos) - UENF
___________________________________________________________________ Prof. Augusto Vidal da Costa Gomes (Doutor, Nutrição Animal) - UFRRJ
___________________________________________________________________ Prof. José Brandão Fonseca (PhD, Nutrição de Monogástricos) - UENF
(Orientador)

ii
“Ao meu pai, Antônio, e à minha querida irmã, Marta, ambos ao lado do
Criador, dedico esta vitória. Deus os abençoe”.

iii
AGRADECIMENTOS
Ao meu Deus e a meus amigos espirituais, pela presença fiel e constante em
todos os momentos da minha existência.
`A minha esposa, Cláudia, pela afinidade espiritual, dedicação, carinho,
paciência, companheirismo, entre muitas outras qualidades.
Aos meus filhos, Natália e Vítor, por serem os grandes amores da minha
vida.
À minha mãe, Rosita, pelo referencial e bem-querer.
Ao meu pai, Antônio, e minha irmã, Marta, falecidos recentemente, pela
certeza de que estivemos, estamos e estaremos sempre juntos.
Ao meu querido sobrinho, Wágner, pela sua luz interior.
Ao meu orientador, José Brandão Fonseca, pela confiança, respeito e
amizade.
`A professora Rita da Trindade Ribeiro Nobre Soares, pela preciosa
orientação, atenção, simplicidade e apoio.
À professora Karla Silva Ferreira, pela participação constante na minha vida
acadêmica, amizade e apoio.

iv
Aos demais amigos do Centro de Ciências e Tecnologias Agropecuárias e a
todos aqueles que contribuíram direta ou indiretamente para a realização deste
trabalho, pelo prazer da convivência nesta caminhada.
`A Universidade Estadual do Norte Fluminense (UENF), pela concessão de
bolsa de estudo.

v
BIOGRAFIA
NEWTON TAVARES ESCOCARD DE OLIVEIRA, filho de Antônio Escocard
de Oliveira e Rosita Tavares Escocard de Oliveira, nasceu em 26 de janeiro de 1966,
na cidade de Campos dos Goytacazes – RJ.
Cursou Zootecnia na Universidade Federal Rural do Rio de Janeiro no
período de 1993 a 1997.
Em agosto de 1998, foi admitido no Curso de Pós-Graduação em Produção
Animal, Mestrado, Nutrição Animal, da Universidade Estadual do Norte Fluminense
(UENF), em Campos dos Goytacazes – RJ, submetendo-se à defesa de dissertação
para conclusão do curso em agosto de 2000.
Em agosto de 2000, ingressou no Curso de Pós-Graduação em Produção
Animal, Doutorado, Nutrição Animal, da Universidade Estadual do Norte Fluminense
(UENF), em Campos dos Goytacazes – RJ, submetendo-se à defesa de tese para
conclusão do curso em agosto de 2004.

vi
CONTEÚDO
Páginas RESUMO.................................................................................................
ABSTRACT.............................................................................................
1. INTRODUÇÃO.....................................................................................
x
xii
1 2. REVISÃO DE LITERATURA............................................................... 4
2.1. Energia metabolizável.................................................................... 4
2.2. Pigmentação de gemas................................................................. 9
2.3. Triglicerídeos.................................................................................. 12
2.3.1 Digestão e absorção de lipídeos da ração................................
2.3.2 Rota de absorção dos lipídeos da ração...................................
2.3.3 Biossíntese de triglicerídeos endógenos e transporte..............
2.3.4 Deposição de triglicerídeos nos tecidos....................................
2.3.5 Ação biológica da bixina e niacina............................................
2.3.6 Niacina e o metabolismo dos lipídeos.......................................
2.3.7 Ação biológica da niacina......................................................... 3. REFERÊNCIAS BIBLIOGRÁFICAS...................................................
4. TRABALHOS.......................................................................................
DETERMINAÇÃO DA ENERGIA METABOLIZÁVEL DE ALGUNS
12
13
14
14
16
17
20
22
29
31

vii
ALIMENTOS TESTADOS EM CODORNAS JAPONESAS FÊMEAS
RESUMO...........................................................................................
ABSTRACT........................................................................................
INTRODUÇÃO...................................................................................
MATERIAL E MÉTODOS...................................................................
Local do experimento......................................................................
Distribuição das codornas no delineamento experimental e
tratamentos......................................................................................
Alojamento das codornas nas instalações e manejo......................
Análises químicas...........................................................................
Ração referência.............................................................................
Coleta total e preparação da excreta..............................................
Variáveis avaliadas.........................................................................
RESULTADOS e DISCUSSÃO..........................................................
CONCLUSÕES..................................................................................
REFERÊNCIAS BIBLIOGRÁFICAS...................................................
PIGMENTAÇÃO DE GEMAS DE OVOS DE CODORNAS
JAPONESAS ALIMENTADAS COM RAÇÕES CONTENDO
COLORÍFICO
RESUMO...........................................................................................
ABSTRACT........................................................................................
INTRODUÇÃO...................................................................................
MATERIAL E MÉTODOS...................................................................
Local do experimento......................................................................
Distribuição das codornas no delineamento experimental e
tratamentos......................................................................................
Alojamento das codornas nas instalações e manejo......................
Rações experimentais.....................................................................
Variáveis avaliadas.........................................................................
Coleta de dados..............................................................................
Procedimentos estatísticos.............................................................
RESULTADOS e DISCUSSÃO..........................................................
Coloração de gema (CG)................................................................
31
31
32
34
34
34
35
35
37
37
39
39
44
44
46
46
46
47
50
50
50
50
51
53
53
54
56
56

viii
Consumo de ração (CR).................................................................
Peso médio dos ovos (PMO)..........................................................
Percentagem de postura (PP), Conversão alimentar por dúzia de
ovos (CAD) e Conversão alimentar por grama de ovos (CAG).......
CONCLUSÕES..................................................................................
REFERÊNCIAS BIBLIOGRÁFICAS...................................................
COLORÍFICO E NIACINA SUPLEMENTAR EM RAÇÕES DE
CODORNAS JAPONESAS MACHOS: TRIGLICERÍDEOS
SANGÜÍNEOS E COMPOSIÇÃO QUÍMICA DA CARNE E
CARCAÇA.............................................................................................
RESUMO...........................................................................................
ABSTRACT........................................................................................
INTRODUÇÃO...................................................................................
MATERIAL E MÉTODOS...................................................................
Local do experimento......................................................................
Distribuição das codornas no delineamento experimental e
tratamentos......................................................................................
Alojamento das codornas nas instalações e manejo......................
Análises químicas dos ingredientes................................................
Rações experimentais.....................................................................
Variáveis avaliadas.........................................................................
Coleta de dados..............................................................................
Procedimentos estatísticos.............................................................
RESULTADOS e DISCUSSÃO..........................................................
Consumo de ração (CR), ganho de peso (GP), conversão
alimentar (CA) e peso (P)................................................................
Triglicerídeos (TS) e lipoproteínas de densidade muito baixa
(VLDL) no sangue............................................................................
Matéria seca (MS), extrato etéreo (EE) e proteína bruta (PB) na
carne de peito..................................................................................
Matéria seca (MS), extrato etéreo (EE) e proteína bruta (PB) na
carne de coxa e sobrecoxa..............................................................
Matéria seca (MS), extrato etéreo (EE) e proteína bruta (PB) na
59
61
62
63
63
65
65
66
66
69
69
69
69
70
71
73
73
75
76
76
77
78
81
82

ix
carcaça............................................................................................
CONCLUSÕES..................................................................................
REFERÊNCIAS BIBLIOGRÁFICAS................................................... 5. CONCLUSÕES GERAIS.....................................................................
83
84
87 APÊNDICES............................................................................................ 88
APÊNDICE A........................................................................................... 89 APÊNDICE B...........................................................................................
APÊNDICE C...........................................................................................
95
99

x
RESUMO
OLIVEIRA, Newton Tavares Escocard de, D.S., Universidade Estadual do Norte Fluminense; Agosto/2004; Energia metabolizável de alimentos e qualidade de ovos e carne de codornas japonesas alimentadas com rações contendo colorífico do urucum e niacina suplementar; Professor orientador: José Brandão Fonseca; Professor conselheiro: Rita da Trindade Ribeiro Nobre Soares.
Esta pesquisa objetivou determinar a energia metabolizável aparente e a
energia metabolizável aparente corrigida pelo balanço de nitrogênio de sete
alimentos para codornas japonesas (Coturnix japonica); avaliar a pigmentação de
gemas de ovos de codornas japonesas por meio da inclusão de diferentes níveis de
colorífico nas rações e mensurar o teor de triglicerídeos e lipoproteínas de densidade
muito baixa no sangue, e os níveis de gordura na carne de peito e coxa + sobrecoxa
e na carcaça de codornas japonesas por meio da inclusão de colorífico e niacina
suplementar nas rações. No experimento sobre digestibilidade de alimentos,
utilizaram-se 280 codornas fêmeas com idade inicial de vinte e seis dias, em
delineamento experimental inteiramente casualizado, com cinco repetições e sete
codornas por unidade experimental. Os tratamentos consistiram de sete rações
experimentais e uma ração referência. Cada ração experimental foi constituída, na

xi
base da matéria natural, por 70 % da ração referência e 30 % do alimento, à
exceção da ração contendo óleo de soja degomado, que foi testado com 10 % de
inclusão e 90 % da ração referência. Os respectivos valores de energia
metabolizável aparente e energia metabolizável aparente corrigida pelo balanço de
nitrogênio (kcal por kg de matéria natural) do milho moído, colorífico, quirera de
arroz, óleo de soja degomado, farelo de soja, semente de linhaça e casca + polpa de
abacaxi foram iguais a 3.124 e 3.122; 3.152 e 3.102; 2.980 e 2.947; 8.065 e 7.940;
2.633 e 2.651; 2.477 e 2.492 e 1.323 e 1.274. No experimento sobre pigmentação
de gemas, foram utilizadas 240 codornas fêmeas, em delineamento experimental de
blocos inteiramente casualizados, com oito tratamentos, cinco repetições e seis
codornas por unidade experimental. Os tratamentos foram constituídos por oito
rações experimentais oriundas da combinação de quatro níveis de colorífico na
ração (0, 1,5, 3,0 e 4,5 %) x duas fontes energéticas (milho e quirera de arroz). Nos
dias 7, 14, 21 e 28 do experimento, os resultados estimados de máxima coloração
de gemas ocorreram com o uso de 3,15; 2,91; 2,97 e 2,77 % de colorífico nas
rações, respectivamente. Codornas alimentadas com rações à base de milho
apresentaram gemas mais pigmentadas do que as gemas de ovos de codornas que
receberam rações à base de quirera de arroz. No experimento sobre mensuração da
gordura da carne, utilizaram-se 240 codornas machos, em delineamento
experimental inteiramente casualizado, com quatro tratamentos, cinco repetições e
doze codornas por unidade experimental. Os tratamentos consistiram de uma ração
referência à base de milho e farelo de soja, sem inclusão de colorífico (COL) e de
niacina suplementar (NS), e de outras três rações, uma com 4,5 % de COL, outra
com 0,08 % de NS e outra com 4,5 % de COL e 0,08 % de NS. Não houve efeito de
tratamento nos níveis de triglicerídeos e lipoproteínas de densidade muito baixa no
sangue e nos teores de gordura na carne de coxa + sobrecoxa e carcaça das
codornas. No 49° dia de idade, as codornas que receberam rações com 0,08 % de
NS tiveram 1,50 % de gordura na carne de peito, sendo esse nível superior (P≤0,05)
a 0,85 %, apresentado por codornas alimentadas com a ração referência. O uso do
COL e da NS não reduziram os níveis de gordura no sangue, carne e carcaça de
codornas japonesas machos.
Palavras-chave: codorna japonesa, colorífico, gordura, niacina suplementar, energia
metabolizável, pigmentação de gema de ovo

xii
ABSTRACT
OLIVEIRA, Newton Tavares Escocard de, D.S., Universidade Estadual do Norte Fluminense; August/2004; Metabolizable food energy, egg and meat quality of japanese quails fed on spice and supplementary niacin in the rations; Adviser: José Brandão Fonseca; Committee Member: Rita da Trindade Ribeiro Nobre Soares.
This research aimed at determining the apparent metabolizable energy and
apparent metabolizable energy corrected for nitrogen balance in seven foodstuffs for
japanese quails (Coturnix japonica); evaluating the egg yolk colour grade of japanese
quails by means of addition of different levels of spice to the rations and measuring
triglyceride contents, very low-density lipoprotein levels in plasma, and fat levels in
breast, drumstick and thigh meat, as well as on carcass of japanese quails by means
of addition of spice and supplementary niacin to the rations. In the experiment of food
digestibility, two hundred and eighty female quails, twenty-six days of age were used
in a completely randomized design, with five replicates and seven quails per
experimental unit. The treatments have consisted of seven experimental rations and
one reference ration. Each experimental ration was constituted, as fed basis, for 70
% of the reference ration and 30 % of food, save the ration that contained soy oil
without phospholipid, that was tested for 10 % of addition and 90 % of the reference

xiii
ration. The respective values of apparent metabolizable energy and apparent
metabolizable energy corrected for nitrogen (kcal/kg of fed basis) to ground corn,
spice, broken rice grains, soy oil without phospholipid, soybean meal, linseed meal
and pineapple (peel and pulp) were 3.124 and 3.122; 3.152 and 3.102; 2.980 and
2.947; 8.065 and 7.940; 2.633 and 2.651; 2.477 and 2.492 and 1.323 and 1.274. In
the experiment of yolk colour, two hundred and forty female quails were used in a
completely randomized block design, with eight treatments, five replicates and six
quails per experimental unit. The treatments consisted of eight experimental rations
derived from the combination of four spice levels in the rations (0, 1,5, 3,0 e 4,5 %) x
two energetic sources (corn and broken rice grains). On the 7th, 14th, 21st and 28th
experimental days, the estimated results for maximum yolk colour occurred with the
use of 3,15; 2,91; 2,97 e 2,77 % spice rations, respectively. Quails fed on corn based
rations had higher egg yolk colour than egg yolks of quails that received broken rice
grains based rations. In the experiment of measuring fat meat, two hundred and forty
male quails were used in a completely randomized design, with four treatments, five
replicates and twelve quails per experimental unit. The treatments consisted of one
reference ration with corn and soybean meal, without spice and supplementary niacin
(SN), and three other rations, one with 4,5 % of spice, the other with 0,08 % of SN
and an other one with 4,5 % of spice and 0,08 % of SN. The treatments had no effect
upon triglycerides and very low-density lipoprotein levels in plasma and fat contents
in the drumsticks and thigh meat and carcass of male quails. On the 49th day of age,
the quails that received rations with 0,08 % of SN had 1,50 % of fat in the breast
meat, higher (P≤0,05) than 0,85 % presented by japanese quails fed on the reference
ration. The utilization of spice and SN did not reduce the fat levels in plasma, meat
and carcass of japanese male quails. Key words: egg yolk colour, fat, japanese quail, metabolizable energy, spice,
supplementary niacin

1
1. INTRODUÇÃO
O Estado do Rio de Janeiro tem apresentado, nos últimos anos, baixos
índices produtivos no setor avícola, contribuindo com pequena parcela da produção
nacional. Dados estatísticos do IBGE (1997), referentes ao ano de 1995, mostram
um efetivo de 18.896.655 aves, incluindo galinhas, galos, frangos, frangas, pintos e
codornas, e uma produção de ovos de galinha da ordem de 30.557 mil dúzias. Estes
valores representaram, aproximadamente, 2,58 % do rebanho nacional e 1,29 % do
total de ovos produzidos, ocupando posições bem inferiores aos Estados de São
Paulo, Minas Gerais, Rio Grande do Sul, Paraná, Santa Catarina, entre outros. Em
relação ao setor de insumos básicos, a produção de milho em grão no ano de 1996
correspondeu a 42.144 toneladas ou 0,13 % do total e a produção de soja no ano de
1996 foi inexistente (IBGE, 1997).
O re-direcionamento da produção de ovos e carne de aves para regiões com
elevado potencial para produção de grãos tem-se constituído em um dos fatores
mais importantes na redução dos custos de produção e, conseqüentemente, no
aumento da produção avícola, tornando as empresas avícolas mais competitivas. As
regiões Sul, Sudeste, Nordeste + Norte e Centro-Oeste foram responsáveis por 53,8,
28,9, 10,6 e 6,7 % da produção nacional de grãos, respectivamente, no período de
janeiro a outubro de 1999. Contudo, as dez maiores empresas avícolas

2
concentraram 50,1 % da produção avícola nacional no ano de 1999, sendo que
nenhuma delas foi implantada no Estado do Rio de Janeiro (BENÍCIO, 2000).
Em razão da visível dificuldade de competir e conquistar mercado com
Estados possuidores de empresas avícolas altamente produtivas, da carência de
políticas que estimulem uma maior produção de grãos e aumento do efetivo avícola
no Estado do Rio de Janeiro e da demanda crescente de mercados consumidores
por produtos especializados, há relevante necessidade da realização de pesquisas
nutricionais visando a obter produtos avícolas diferenciados, de maior valor
agregado, cuja composição química determine efeitos benéficos à saúde humana e
animal.
Várias pesquisas têm sido desenvolvidas visando ao melhor entendimento
dos diversos fatores que influem na qualidade dos ovos e da carne de aves e,
conseqüentemente, na saúde humana. A influência dos carotenóides, das vitaminas
e de outros nutrientes no controle de diversas doenças humanas tem sido marcante.
Isto tem incentivado pesquisadores, nutricionistas e técnicos da área animal a
trabalharem na produção de produtos avícolas funcionais. O enriquecimento dos
produtos avícolas e sua inclusão na dieta humana visam a ajustar deficiências
decorrentes do consumo inadequado de nutrientes importantes para o metabolismo,
causado principalmente pela agitação da vida moderna. Tais produtos têm sido
procurados cada vez mais, tendo em vista a maior exigência dos consumidores.
Vale destacar que, em passado recente, características como peso corporal
e conversão alimentar foram consideradas de maior importância nos programas de
melhoramento genético. Entretanto, com o mercado cada vez mais especializado e
segmentado, características como rendimento de carcaça eviscerada, rendimento de
carne de peito e de perna, teor de gordura, além das relacionadas com a qualidade
da carne, como cor, textura, entre outras, e com a saúde e bem-estar humano e
animal têm sido incorporadas nos programas (SOUZA, 2000).
Sendo assim, foram realizados três experimentos com a finalidade de
alcançar os seguintes objetivos:
1. Determinar a energia metabolizável aparente e a energia metabolizável aparente
corrigida pelo balanço de nitrogênio do milho, colorífico, quirera de arroz, óleo de
soja degomado, farelo de soja, semente de linhaça e polpa + casca de abacaxi
para codornas japonesas;

3
2. Avaliar a pigmentação de gemas de ovos de codornas por meio da inclusão de
diferentes níveis de colorífico nas rações;
3. Mensurar o teor de triglicerídeos e lipoproteínas de densidade muito baixa do
soro sangüíneo, e os níveis de triglicerídeos do peito, coxa + sobrecoxa e
carcaça de codornas japonesas por meio da inclusão de colorífico e niacina
suplementar nas rações;
O agrupamento de informações sobre digestibilidade de alimentos e do
efeito da inclusão do colorífico (fonte do carotenóide bixina, pigmento natural que
pode atuar como agente antioxidante no organismo humano) e da niacina
suplementar (vitamina hidrossolúvel que possui ação hipolipidêmica) em rações para
codornas japonesas será útil para subsidiar decisões sobre a qualidade dos ovos e
da carne de codornas.

4
2. REVISÃO DE LITERATURA
2.1 Energia metabolizável
Segundo BERTECHINI (1991), a energia contida nos alimentos é dividida
em energia bruta (EB), energia digestível, energia metabolizável e energia líquida
(Figura 1).
Figura 1. Partição biológica da energia. A EB é determinada em bomba calorimétrica através da oxidação total da
matéria orgânica na presença de alta pressão de oxigênio (25 ATMs), onde é
- Energia fecal
- Energia urinária
- Incremento calórico
MantençaMetabolismo basal
AtividadesCalor corporal
Energia produtiva paraCrescimento, Reprodução,
Gestação, Lactação
ENERGIA LÍQUIDA
ENERGIA METABOLIZÁVEL
ENERGIA DIGESTÍVEL
ENERGIA BRUTA INGERIDA

5
medida a produção de calor liberado. Para aves, a medida normalmente utilizada é a
energia metabolizável (EM), que corresponde à diferença entre a energia bruta (EB)
da ração e a soma da EB das fezes e urina. Como os dutos dos tratos digestivo e
urinário convergem para a cloaca, parte final que se abre externamente através do
ânus, a determinação da energia das fezes é realizada junto com a da excreção
urinária (BERTECHINI, 1991).
A EM é denominada aparente porque ocorre interferência de elementos que
não são oriundos do alimento ingerido, como escoriações de células da mucosa
intestinal, bílis, fluidos digestivos e produtos do catabolismo dos tecidos, e que são
computados no cálculo da energia metabolizável aparente (EMA). Quando se retira
do valor energético da excreta a energia das secreções metabólicas fecais e
endógenas urinárias, que não são provenientes do alimento ingerido, tem-se a
energia metabolizável verdadeira. Para reduzir a variação entre estes valores, tem-
se recomendado corrigir os valores de EMA pelo balanço de nitrogênio (retenção e
excreção), pois o nitrogênio participa, juntamente com outros elementos, no
atendimento das exigências de mantença, prioritárias no metabolismo animal
(SIBBALD, 1980).
A correção da EMA pela retenção de nitrogênio tem permitido maior precisão
nas formulações de rações para aves, mas especificamente em relação a estudos e
ensaios envolvendo a codorna japonesa, tem sido freqüente o uso de valores de EM
de alimentos testados com frangos de corte ou galinhas poedeiras em formulações
de rações para codornas japonesas. Isso tem comprometido o desempenho
produtivo das codornas, porque há diferenças anatômicas, histológicas e de
tamanho e comprimento do trato gastrintestinal entre essas espécies, além de haver
diversas interações entre alimento e trato gastrintestinal, causadas pelas diferenças
de composição química e estrutura física dos alimentos, interferindo na eficiência
dos processos digestivo e absortivo.
O comprometimento do desempenho produtivo de codornas japonesas pode
ser evidenciado quando essas aves consomem rações cujos valores de EM estão
discrepantes das suas exigências energéticas. Considerar, durante a formulação de
rações, a influência conjunta de fatores nutricionais, como o valor nutritivo de um
alimento, expresso pela composição química da ração e digestibilidade dos seus
nutrientes, de fatores não nutricionais, como estrutura física da ração, idade, sexo,
entre outros, e, principalmente, da interação entre dois ou mais fatores sobre a

6
quantidade de energia da ração que as codornas conseguem metabolizar pode
minimizar a diferença entre exigência de energia e energia metabolizável (EM) da
ração.
Como exemplo das peculiaridades apresentadas, podem-se citar
observações feitas no trabalho de ANGULO et al. (1993), que verificaram que
codornas japonesas, de ambos os sexos, alimentadas com rações peletizadas
contendo 3.200 kcal de EM por kg de ração foram mais eficientes no uso da energia
(maior ganho de peso, menor consumo de ração, melhor conversão alimentar) do
que codornas alimentadas com rações fareladas contendo 3.000 kcal de EM por kg
de ração, nos períodos de 1 a 14, 14 a 33 e 1 a 33 dias de idade.
Em relação ao efeito de idade, FARRELL et al. (1982) observaram que
codornas japonesas de ambos os sexos consumiram diariamente 57,0, 46,8 e 46,7
kcal de EM por kg de peso corporal (PC) entre o 12° e 15°, 19° e 22° e 26° e 29°
dias de idade, respectivamente. Estes resultados mostraram que as codornas
apresentaram eficiência alimentar constante a partir do 19° ao 22° dias, pois o
consumo de EM e o ganho de peso em gramas por kg de PC diminuíram do 12° ao
15° para o 19° ao 22° dias de idade, mas mantiveram-se sem variações desse
período em diante.
FURLAN et al. (1998a) determinaram os valores de energia metabolizável
aparente corrigida pelo balanço de nitrogênio (EMAn) e os coeficientes de
metabolização da energia bruta (CMEB) na matéria natural, para codornas
japonesas machos, em farinha de carne e ossos, farinha de peixe, farelo de algodão,
sorgo e triticale. Os resultados para a EMAn foram de 1.346, 2.336, 1.120, 3.047 e
2.908 kcal por kg e para o CMEB foram de 39,48, 52,59, 27,05, 82,06 e 77,69 %,
respectivamente.
Em codornas japonesas machos com idade de 65 dias, os valores de
energia metabolizável aparente (EMA) e EMAn do milho e dos farelos de trigo, arroz
integral, soja e semente de colza foram iguais a 3.444 e 3.429, 2.400 e 2.400, 3.001
e 3.014, 2.565 e 2.593 e 2.020 e 1.996 kcal por kg de matéria natural,
respectivamente (FURLAN et al., 1998b).
ALBINO et al. (1986) encontraram, em frangos com quarenta e dois dias de
idade, 3.992 kcal de EMA por kg, 3.909 kcal de EMAn por kg e 82,91 % de matéria
seca metabolizável aparente (MSMA) para o milho moído e 2.906 kcal por kg de
EMA, 2.852 kcal por kg de EMAn e 42,74 % de MSMA para o farelo de soja.

7
Houve uma relação linear entre os valores de energia metabolizável
aparente (EMA) com o conteúdo de matéria seca metabolizável aparente (MSMA) de
dezenove alimentos testados (R2 = 0,99) em rações de frangos, expressos na base
da matéria seca, indicando que estas equações podem ser usadas para estimar os
valores de EMA dos alimentos, principalmente nos locais onde não existe bomba
calorimétrica, pois esta metodologia só necessita determinar a MSMA dos alimentos
(HAN et al., 1976).
Por meio do método de coleta total de excretas, foram determinados e
tabelados os valores de energia metabolizável aparente corrigida por retenção de
nitrogênio (EMAn) e de energia metabolizável verdadeira corrigida por retenção de
nitrogênio (EMVn) de diversos alimentos usados em rações de aves, utilizando-se
aves de diversas idades (galos, galinhas poedeiras e pintos). Os valores de EMAn e
EMVn do milho em grão, farelo de soja (45 % de proteína bruta), quirera de arroz,
óleo de soja e óleo de milho foram, respectivamente, de 3.371 e 3.450, 2.266 e
2.495, 3.273 e 3.507, 8.790 e 9.200 e 8.886 e 9.250 kcal por kg de matéria natural
(ROSTAGNO et al., 2000).
Não houve diferença entre valores de EMAn de dezenove ingredientes
utilizados em rações de frangos, expressos na base da matéria seca, no período de
14 a 28 dias de idade, obtidos com os métodos de coleta total de excretas e do
óxido crômico como indicador fecal (HAN et al., 1976).
Os valores de energia metabolizável (EM) para codornas foram similares aos
de galinhas para o milho, sorgo, farinha de peixe, amido de milho e banha, e
maiores para alimentos fibrosos, como o farelo de trigo e o feno de alfafa
(MURAKAMI, 1998). O maior valor de EM dos alimentos fibrosos para codornas foi
atribuído à digestibilidade da fibra, que está em função do maior tamanho do ceco
em relação ao tamanho do corpo, quando comparado com o de galinhas.
Segundo MURAKAMI (1998), quanto menor o tempo de permanência do
bolo alimentar no trato digestório (maior taxa de passagem), menor será o
aproveitamento energético do alimento. Em razão disso, o tempo de passagem do
milho moído e dos farelos de trigo, arroz e canola através do trato digestório de
codornas japonesas foi mensurado. Os valores encontrados foram de 97, 82, 76 e
77 minutos, respectivamente.
Em frangos de corte, a digestibilidade de substâncias pécticas, celulose e
hemicelulose foi quase zero em função do pouco tempo de permanência do material

8
fibroso no aparelho digestório, que foi de cinco a nove horas, permitindo baixa
fermentação da fibra pela flora microbiana intestinal (ROSTAGNO, 1994).
Entretanto, o tempo de passagem de uma digesta (não revelada) através do trato
digestório de codornas japonesas variou entre 60 e 90 minutos (VOHRA, 1971).
Usando rações com baixo e alto teor de celulose, em codornas de ambos os
sexos com idade de dezesseis semanas, SAVORY e GENTLE (1976a) verificaram
que a máxima excreção de óxido crômico ocorreu duas a três horas após a
introdução do marcador na ração, cessando quatro a cinco horas após sua remoção.
Segundo SAVORY e GENTLE (1976a), codornas consumiram em maior
quantidade a ração com alto teor fibroso do que a ração com pouca fibra, sendo
atribuído ao elevado teor fibroso o decréscimo da digestibilidade e da energia
metabolizável (EM) da ração.
Em razão do conteúdo energético das rações ser inversamente proporcional
ao conteúdo fibroso das mesmas, comportamento similar foi observado por
MURAKAMI et al. (1993), que revelaram ter havido redução do consumo de ração
(CR) de codornas japonesas não sexadas com 42 dias de idade com o aumento de
2.800 para 3.000 kcal de EM por kg de ração.
WILSON et al. (1977) verificaram que codornas Bobwhite alimentadas com
2.850 kcal de EM por kg de ração tiveram maior CR às cinco semanas de idade do
que codornas alimentadas com 3.010 e 3.170 kcal de EM por kg de ração,
mostrando que as codornas que consumiram rações com menor teor calórico
compensaram o insuficiente consumo energético ingerindo maior quantidade de
ração, a fim de manterem estáveis suas reservas energéticas corporais, ratificando a
existência de um mecanismo fisiológico regulador do consumo, em longo prazo,
onde estas reservas funcionam como referencial.
SAVORY e GENTLE (1976b) verificaram menor peso em codornas
alimentadas com rações com alto teor de fibra do que em codornas alimentadas com
rações com baixo teor fibroso. O contraste de peso corporal foi atribuído a diferenças
no uso da energia, pois o elevado conteúdo fibroso promove maior gasto de energia
em função do maior tempo e trabalho gasto no ato da alimentação. Além disto, os
intestinos de aves alimentadas com altos níveis de fibra são maiores, exigindo maior
demanda energética metabólica.

9
2.2 Pigmentação de gemas As sementes do urucum possuem um corante vermelho situado em suas
partes externas. A bixina, um mono metil éster da norbixina, é o corante majoritário
encontrado nessas partes. Em função do extensivo sistema de ligações duplas
conjugadas na molécula (Figura 2), a bixina pode ocorrer na forma cis e trans, sendo
a forma cis a mais encontrada nas sementes. A trans-bixina e a cis-norbixina são
encontradas como constituintes minoritários em sua composição (HENRY, 1992).
Figura 2. Estrutura química da bixina. Do total de sementes de urucum produzidas no Brasil, cerca de 75% são
utilizadas na fabricação do colorífico, totalmente consumido no mercado interno e
definido pela Resolução 12/78 da Comissão Nacional de Normas e Padrões para
Alimentos do Ministério da Saúde (CNNPA-MS) como um produto constituído pela
mistura de fubá ou farinha de mandioca com urucum em pó ou extrato oleoso de
urucum, adicionado ou não de sal e de óleos comestíveis (TOCCHINI e
MERCADANTE, 2001).
Em razão da facilidade de obtenção do produto no mercado e da elevada
proporção do carotenóide bixina, existe um grande potencial de uso do colorífico em
rações de aves a fim de se obter gemas de ovos mais pigmentadas e,
conseqüentemente, enriquecidas nutricionalmente. Entretanto, isto tem sido pouco
freqüente porque tem havido variações nos teores de bixina em diversas amostras
de colorífico, o que dificulta as recomendações de uso do colorífico nas rações.
TOCCHINI e MERCADANTE (2001) observaram uma grande variação nos teores de
bixina em sete marcas diferentes de colorífico presentes no mercado, determinadas
por cromatografia líquida de alta eficiência. Nesse estudo, os valores variaram de
154 a 354 mg de bixina por 100 g de colorífico.
Embora com custos de produção inferiores aos dos pigmentos naturais, em
função da exigência do consumidor por alimentos sem aditivos químicos, os
corantes artificiais têm sido cada vez mais substituídos por pigmentos naturais.

10
Entretanto, na avaliação toxicológica, nem sempre o caráter natural de um pigmento
indica toxicidade menor que um pigmento artificial (CARVALHO, 1992). Pelo
extensivo uso do colorífico, o consumo estimado de bixina por cerca de 44 milhões
de brasileiros foi em torno de 150 % superior ao consumo diário máximo de bixina
para humanos, estabelecido em 0,065 mg por kg de peso corporal (WHO, 2000).
O consumo excessivo de corantes em relação a sua ingestão diária aceitável
tem estimulado a realização de estudos sobre toxidade. PAUMGARTTEN et al.
(2002) não constataram redução no peso corporal (PC) em ratas com 21 dias de
gestação e nem mortalidade, redução no PC, anormalidades viscerais e esqueléticas
nas suas proles, decorrentes da administração oral de extrato seco de urucum (28 %
de bixina) suspenso em óleo de milho, nas dosagens de 0; 31,2; 62,5; 125 e 500 mg
de extrato por kg de PC por dia, durante o 6º e 15º dia de gestação das ratas. A
dose de 500 mg, equivalente a 140 mg de bixina por kg de PC por dia, correspondeu
a cerca de 2153 vezes a dose máxima pré-estabelecida para humanos, que é de
0,065 mg de bixina por kg de PC por dia.
A não-toxicidade da bixina, quando usada em doses elevadas, mostra que
este aspecto não se tem constituído em fator limitante para seu uso. Considerando
isso, pesquisas têm sido realizadas utilizando-se o urucum, rico em bixina, na
pigmentação de gemas de ovos de poedeiras, principalmente quando se utiliza uma
fonte energética isenta de pigmentos, como o sorgo, a quirera de arroz, entre outras,
em substituição ao milho amarelo.
Segundo CAMPOS (1955), as gemas de ovos colocados por poedeiras
alimentadas com rações contendo 1 % de sementes de urucum não trituradas
(farinha de urucum) e 30 % de adlai (isento de pigmentos) apresentaram coloração
variável entre o amarelo e o amarelo alaranjado, sendo essa cor similar às gemas
produzidas por galinhas alimentadas com 30 % de milho e sem farinha de urucum.
Entretanto, as aves que receberam rações contendo 2 % de sementes de urucum
não trituradas e 30 % de adlai produziram gemas mais escuras, cuja tonalidade
variou do laranja ao laranja forte, de maior preferência do consumidor brasileiro.
PEREIRA et al. (2000) verificaram que o uso de níveis crescentes (500,
1000, 1500 e 2000 ppm) de extrato oleoso de urucum aumentou linearmente a
coloração das gemas dos ovos de galinhas poedeiras, produzidos durante dois
ciclos de 28 dias. A adição de 2000 ppm de bixina em ração com 63 % de sorgo
promoveu a mesma cor de gema obtida com ração contendo 61 % de milho.

11
Contudo, a produção e o peso dos ovos, o consumo de ração e a conversão
alimentar não foram afetados pelos tratamentos.
PEREIRA et al. (2001) verificaram que em rações à base de milho contendo
0; 0,05; 0,10; 0,15 e 0,20 % de extrato oleoso de urucum (EU), houve intensificação
da cor das gemas dos ovos de poedeiras à medida que os níveis de extrato foram
aumentados. Entretanto, o extrato não interferiu na produção e qualidade interna dos
ovos, no consumo de ração e na conversão alimentar.
Poedeiras leves e pesadas que receberam rações com 21,4 % de farelo de
soja (FS), 25,4 % de milho, 41,4 % de sorgo e 0,10 % de EU produziram ovos com
cor de gemas similar às aves que receberam a ração referência (22,8 % de FS e
63,7 % de milho). Os valores de pigmentação de gemas, obtidos por meio do leque
colorimétrico da Roche, foram de 5,10 e 5,02 para as poedeiras leves, e 5,83 e 5,35
para as poedeiras pesadas tratadas com ração referência e com ração com sorgo +
0,10 % de EU, respectivamente. A adição de 0,45 % de EU à ração contendo sorgo
produziu pigmentação da gema de ovos de poedeiras leves igual a 8,9 pontos no
leque da Roche, valor dentro da faixa referencial para aves caipiras, que gira em
torno de 8,5 - 9,0 pontos (SILVA et al., 2000).
Outros estudos têm sido realizados a fim de avaliar a inclusão de fontes de
xantofilas diferentes do colorífico e de outros agentes pigmentantes derivados do
urucum, em rações de aves e, dessa forma, avaliar a eficiência de incorporação de
pigmentos nas gemas ao longo de um período. Verificando o valor pigmentante do
feno de capim bermuda ao longo do tempo, CHEN e BAILEY (1988) observaram que
galinhas poedeiras no primeiro ciclo de produção colocaram ovos cujas gemas
alcançaram um platô (pigmentação máxima) entre a terceira e quarta semana após a
inclusão do feno. Em período pré-experimental de duas semanas, as galinhas
receberam ração isenta de xantofilas a fim de depletar seus níveis no organismo.
Entretanto, BORNSTEIN e BARTOV (1965) mencionaram que, quando a
avaliação é feita por meio de métodos que utilizam o escore visual, nos quais se
inclui o leque colorimétrico da Roche, as respostas de coloração de gemas em
função dos incrementos de xantofilas na ração seguem um padrão logaritmico. Os
autores revelaram que o modelo quadrático é inadequado porque parece improvável
que a cor das gemas decresça, devido ao excesso ou constância de suplementação
de pigmento na ração, após ter alcançado um ponto de coloração máxima.

12
2.3 Triglicerídeos Os lipídeos são substâncias orgânicas heterogêneas, solúveis em solventes
apolares com pequena ou nenhuma solubilidade em água (BERTECHINI, 1991) e os
triglicerídeos são ésteres formados pela união do glicerol com três moléculas de
ácidos graxos. Eles são o tipo de lipídeo encontrado em maior proporção nas
gorduras, que são fontes de lipídeos normalmente encontradas em produtos de
origem animal. Na composição centesimal das gorduras, as moléculas de
triglicerídeos são formadas, em sua maioria, por ácidos graxos saturados, sendo, por
isso, sólidas à temperatura ambiente. Os óleos são líquidos à temperatura ambiente
e são obtidos geralmente de fontes de origem vegetal, sendo compostos, em sua
maioria, por moléculas de triglicerídeos formadas por ácidos graxos insaturados. Se
os três ácidos graxos constituintes da gordura ou óleo forem iguais, forma-se um
triglicerídeo simples, caso contrário, tem-se um triglicerídeo misto.
2.3.1 Digestão e absorção de lipídeos da ração
Os triglicerídeos, ácidos graxos livres e fosfolipídeos presentes na ração não
sofrem transformação química na boca, faringe e esôfago. No estômago glandular, a
temperatura, o pH e a atividade da pepsina atuam em conjunto para unir e formar
glóbulos de gordura. O efeito final da digestão no proventrículo e moela é destruir a
integridade estrutural do alimento e formar um "pool" de lipídeos. No ambiente
neutro do duodeno, a presença de produtos da digestão gástrica estimula a
liberação da bile e dos sucos pancreáticos. Ao mesmo tempo, aumenta-se a
motilidade, resultando em eficiente emulsificação. Os ácidos biliares e a lecitina
presente na bile aumentam a área superficial e otimizam a eficiência digestiva. O
lipídeo emulsificado fica com os ácidos biliares e a lecitina na parte externa, em
contato com a água, e os triglicerídeos posicionam-se na parte interna dos glóbulos
lipídicos (MORAN JR., 1994).
A colipase é um cofator protéico que permite que a lipase se fixe na
superfície óleo-água, tendo acesso aos triglicerídeos de camadas menos expostas.
A lipase cliva ácidos graxos das posições 1 e 3, liberando dois ácidos graxos livres e
um 2-monoglicerídeo por molécula de triglicerídeo. A produção e liberação de lipase
e colipase são ajustadas à quantidade de gordura alimentar. A motilidade possibilita
uma remoção contínua de lipídeos da interface óleo-água e a formação de micelas

13
ocorre a partir da combinação dos produtos da digestão de gorduras com os ácidos
biliares e lecitina da bile (MORAN JR., 1994).
A absorção das micelas ocorre contra a camada de água estacionária,
criada pelas microvilosidades e o glicocálix do enterócito em si. A mucina contribui
para a estabilidade dessa água por atuar como um gel e impedir o movimento
macromolecular. As micelas lipídicas se desintegram em seus componentes
moleculares, dentro da camada de mucina, assim que se aproximam da superfície
celular do enterócito. O movimento das micelas é dependente de sua concentração
no lúmen e da atividade convectiva do seu conteúdo (MORAN JR., 1994).
2.3.2 Rota de absorção dos lipídeos da ração
Após a absorção, as moléculas de ácidos graxos livres, 2-monoglicerídeos,
lisolecitina e colesterol (COL) são captados por carreadores e levadas para o retículo
endoplasmático. A montagem das lipoproteínas, macromoléculas formadas por um
centro hidrofóbico de triacilgliceróis e éster de COL, circundado por uma camada
superficial de fosfolipídeos, COL livre e proteínas (BRAGAGNOLO, 1992), ocorre
pela reesterificação dos 2-monoglicerídeos para triglicerídeos no retículo
endoplasmático liso. Apoproteínas são adicionadas para dar suporte às esferas de
lipoproteínas, que são agrupadas no complexo de Golgi, transferidas para a
membrana basolateral e liberadas na lâmina própria (MORAN JR., 1994).
O intestino das aves secreta diretamente na veia porta grandes lipoproteínas
ricas em triglicerídeos (portamícrons) oriundos da gordura da ração. Contudo, a
reesterificação dos ácidos graxos absorvidos na mucosa intestinal não é completa
em aves e acima de 50 % da gordura absorvida pode ser liberada para dentro da
veia porta como ácidos graxos não esterificados (livres), os quais são provavelmente
removidos da circulação durante sua passagem inicial através do fígado. Após
hidrólise, uma proporção de triglicerídeos contidos nos portamícrons é captada pelo
fígado, liberando portamícrons remanescentes na circulação periférica. Esses dois
eventos podem contribuir para os efeitos do conteúdo de gordura da ração sobre a
composição corporal em aves, pois a síntese hepática de ácidos graxos é fortemente
inibida pela gordura da ração (GRIFFIN et al., 1999).
Os portamícrons formados são secretados diretamente no sistema portal,
não são metabolizados no fígado em função do elevado tamanho, e atingem a
grande circulação (GRIFFIN et al., 1999), onde os triglicerídeos presentes podem ser

14
catabolisados pela lipase lipoprotéica para que os ácidos graxos resultantes
penetrem na célula e, no tecido adiposo, sejam reesterificados e armazenados como
triglicerídeos, ou servirem como fonte de energia para o metabolismo celular. 2.3.3 Biossíntese de triglicerídeos endógenos e transporte
Os lipídeos de origem hepática são sintetizados e secretados como
partículas de lipoproteínas, cuja biossíntese, em aves, ocorre principalmente no
fígado. A taxa de lipogênese hepática em aves é muito maior do que no tecido
adiposo, sendo o fígado responsável por cerca de 95 % da síntese de novos ácidos
graxos em pintos. Tem sido unânime que quase toda gordura que se acumula no
tecido adiposo de aves é sintetizada no fígado ou oriunda da ração (GRIFFIN et al.,
1999).
O transporte de lipídeos do fígado para outros tecidos em aves fêmeas
imaturas e em machos é feito pelas lipoproteínas. O plasma das aves contém
lipoproteínas de densidade muito baixa (VLDL), lipoproteínas de densidade baixa
(LDL) e lipoproteínas de densidade alta (HDL) cujo tamanho, composição lipídica e
funções são similares aos de mamíferos, contudo as aves não têm a apolipoproteína
apo-E ou apo-A-2 encontradas nos mamíferos, e sintetizam somente a forma
equivalente apo-100 da apo-B (GRIFFIN et al., 1999).
No sangue das aves, as lipoproteínas são chamadas de portamícrons
quando transportam os lipídeos absorvidos (90 % de triglicerídeos, 5 % de colesterol
e 2 % de proteínas) e de VLDL (60 % de triglicerídeos, 12 % de colesterol e 10 % de
proteínas) quando transportam os lipídeos de origem endógena (HERMIER, 1997). 2.3.4 Deposição de triglicerídeos nos tecidos
Em aves, a gordura extra hepática depende do nível de síntese e secreção
das lipoproteínas pelo fígado, pois o armazenamento de triglicerídeos nos
adipócitos, hepatócitos e ovócitos em crescimento depende da disponibilidade de
lipídeos do plasma, oriundos da ração ou da lipogênese no fígado. Normalmente a
concentração de lipídeos das rações para aves comerciais não tem ultrapassado o
limite de 10 %, o que tem contribuído para que a lipogênese seja dependente, em
sua maior parte, da disponibilidade de carboidratos nas rações, que darão origem a
moléculas de acetil-CoA, pois a síntese de ácidos graxos é muito limitada no tecido
adiposo e não ocorre no ovário (HERMIER, 1997).

15
Medidas “in vivo” da taxa de secreção de lipoproteínas de densidade muito
baixa (VLDL) e do destino do carbono biologicamente marcado e ligado a moléculas
de VLDL ([14C]-VLDL) indicaram que o metabolismo das lipoproteínas garantiu cerca
de 85 % da deposição de ácidos graxos na gordura abdominal de frangos com seis
semanas de idade, ao passo que a lipogênese no tecido adiposo foi suficiente para
garantir os 15 % restantes. A baixa taxa de lipogênese no tecido adiposo em aves foi
atribuída à supressão feita pelos ácidos graxos produzidos no fígado ou vindos do
intestino. Entretanto, somente 25 % do [14C]-VLDL injetado via intravenosa foi
oxidado dentro de oito horas, indicando que apenas uma pequena proporção dos
triglicerídeos contidos nas moléculas de VLDL foi captada pelos músculos para
servir como fonte de energia para o metabolismo celular (GRIFFIN et al., 1999).
As concentrações plasmáticas de VLDL em frangos e perus alimentados à
vontade são bem correlacionadas com conteúdo de gordura corporal, agindo como
medida indireta de adiposidade. O coeficiente de correlação encontrado (r = 0,6 –
0,7) indicou que aproximadamente 50 % da variação genética sobre adiposidade em
populações comerciais normais foram atribuídas a diferenças nas concentrações
plasmáticas de VLDL (GRIFFIN et al., 1999).
A seleção divergente para baixa e alta concentração de VLDL no plasma
tem produzido linhagens “magras” e “gordas” com diferenças marcantes em
conversão alimentar e eficiência protéica. Após dez gerações de seleção, a
diferença nas concentrações plasmáticas de VLDL entre linhagens foi cerca de vinte
vezes, sendo que a diferença na taxa de secreção de VLDL do fígado foi de quatro a
cinco vezes. Uma relativa alta taxa de β-oxidação no fígado combinada com uma
alta taxa de síntese de ácidos graxos podem explicar como pequenas trocas na
lipogênese hepática de aves podem ter um grande efeito na secreção de VLDL, com
reflexo nas concentrações de VLDL no plasma (GRIFFIN et al., 1999).
Outro ponto importante para a formação da gordura extra hepática é o
catabolismo intravascular das lipoproteínas, que controla a entrada e o
armazenamento de lipídeos no tecido adiposo. A transferência de triglicerídeos do
núcleo das VLDL e dos portamícrons para o tecido adiposo envolve seu catabolismo
pela lipase lipoprotéica, que catalisa a hidrólise de triglicerídeos a ácidos graxos e
glicerol. Os ácidos graxos entram nos tecidos vizinhos e, no tecido adiposo, são
reesterificados e armazenados como triglicerídeos. A lipase lipoprotéica é sintetizada
nos adipócitos, músculos e em outros tipos de células, mas só a enzima que foi

16
secretada e encontrada na superfície interna das paredes capilares dos tecidos
periféricos é funcionalmente ativa (HERMIER, 1997).
A seleção divergente de frangos para concentrações de lipoproteínas de
densidade muito baixa (VLDL) no plasma tem criado diferenças significativas na
atividade da lipase lipoprotéica do plasma, tecido adiposo e músculo. Em todos os
casos, a maior atividade foi verificada nas linhagens magras, com seleção para
baixos níveis plasmáticos de VLDL. Contudo, a maior atividade da lipase lipoprotéica
no músculo das linhagens magras não foi acompanhada por diferença significativa
na proporção de triglicerídeos ligados a moléculas de VLDL que podem ser oxidados
a CO2 (GRIFFIN et al., 1999). 2.3.5 Ação biológica da bixina
Amostras de colorífico têm apresentado de 154 a 354 mg do carotenóide
bixina (TOCCHINI e MERCADANTE, 2001). A bixina tem sido utilizada no controle
da hiperlipidemia e da aterosclerose e seu efeito na prevenção e controle da
hiperlipidemia tem sido atribuído à sua ação ativadora sobre a lipase lipoprotéica.
Sugeriu-se, para explicar o mecanismo de ação, a formação de quelatos entre a
enzima e a bixina, que induzem a mudança conformacional da enzima, modificando
seu centro ativo (LIMA et al., 1999).
Todavia, o efeito hipolipidêmico da bixina na prevenção e controle da
aterosclerose de organismos animais tem sido atribuído à sua ação antioxidante.
Após serem captados, a bixina, a norbixina e outros carotenóides podem proteger o
endotélio dos vasos sangüíneos da oxidação da lipoproteína de densidade baixa
(LDL). O endotélio danificado pode gerar lesões ateroscleróticas (LIMA et al., 2001).
A concentração de 5,4 x 10-5 mol/L de bixina e de norbixina promoveu
aumento máximo da atividade da lipase lipoprotéica. Em relação ao tratamento sem
adição de modificadores, os aumentos foram de 145 % para a bixina e 71 % para a
norbixina (LIMA et al., 1999).
Após três meses e em relação ao grupo que recebeu somente ração, ratos
fêmeas alimentadas com ração diária contendo 70, 350 e 700 mg de corante (28 %
de bixina) apresentaram uma redução de 9,30; 15,24 e 10,85 % dos níveis de
triglicerídeos séricos (TS), respectivamente. Machos que receberam 70 mg
apresentaram aumento de 4,77 %, porém os alimentados com 350 e 700 mg de
corante tiveram os níveis de TS reduzidos em 37,36 e 43,82 %, respectivamente.

17
Após seis meses e em relação ao grupo referência, as fêmeas responderam com
redução de 6,40 e 12,58 %, nos tratamentos com 350 e 700 mg, respectivamente, e
com aumento de 1,32 % com o uso de 70 mg. Os níveis de triglicerídeos séricos dos
machos foram reduzidos em 25,91; 9,21 e 13,28 % com o uso de 70, 350 e 700 mg
de corante, respectivamente. Todas as doses de bixina reduziram a lipidemia em
ratos que não tiveram hiperlipidemia induzida (SOUZA et al., 2000).
Com hiperlipidemia induzida por 1 % de colesterol (COL) e 0,1 % de ácido
cólico (AC) na ração (R) diária, durante 28 dias, coelhos da raça Nova Zelândia que
receberam R + COL + AC e a dose diária de 0,01 mol de corante (30 % bixina) por
Kg de peso corporal (via oral e em cápsula) apresentaram 228,79 mg de triglicerídeo
por dl de soro no 28° dia, valor superior (P≤0,05) a 113,78 mg/dl, encontrado nos
coelhos que receberam somente ração comercial, e inferior numericamente a 266,72
mg/dl, obtido por coelhos que receberam R + COL + AC (LIMA et al., 2001).
Em relação a sua toxidade, não foram verificados redução no peso corporal
em ratas com 21 dias de gestação e nas suas proles, com o uso da dose de 500 mg
de extrato seco de urucum (28 % de bixina) por kg de peso corporal por dia,
dosagem que equivale a um consumo médio de 140 mg de bixina por kg de peso
corporal por dia (PAUMGARTTEN et al., 2002).
2.3.6 Niacina e o metabolismo dos lipídeos A niacina é uma vitamina hidrossolúvel que se refere tanto ao ácido nicotínico
(Figura 3) como a nicotinamida.
Figura 3. Estrutura química do ácido nicotínico.
No organismo, a nicotinamida faz parte do grupo ativo das coenzimas
nicotinamida adenina dinucleotídeo e nicotinamida adenina dinucleotídeo fosforilada.
O grupo fosfato ocorre no grupo 2-hidroxila da adenosina monofosfato (AMP) da

18
coenzima. A nicotinamida adenina dinucleotídeo (NAD) e a nicotinamida adenina
dinucleotídeo fosforilada (NADP) podem ser encontradas na forma oxidada (NAD+ e
NADP+) e na forma reduzida (NADH + H+ e NADPH + H+), respectivamente (Figura
4). Nos alimentos, a niacina ocorre principalmente na forma de NAD e NADP e suas
versões reduzidas. A NAD pode ser hidrolisada por enzimas na mucosa intestinal,
resultando na produção de nicotinamida e de nucleotídeo nicotinamida, os quais
podem ser clivados, possivelmente por enzimas intestinais e hepáticas, para formar
ácido nicotínico (BRODY, 1994).
Figura 4. Estrutura química das coenzimas da niacina e da nicotinamida. Por outro lado, o primeiro passo da conversão do ácido nicotínico a NAD
envolve a transferência de um grupo fosfato ribose do pirofosfato para o ácido
nicotínico, formando ácido nicotínico nucleotídeo. O segundo passo envolve a
transferência de um grupo adenosina difosfato (ADP) da adenosina trifosfato (ATP),
formando ácido nicotínico adenina dinucleotídeo e o passo final é uma reação de
amidação, onde a glutamina doa um grupo amida para um grupo caboxila, formando
a NAD (BRODY, 1994).
A função da nicotinamida da forma oxidada NAD+ é atuar como receptora de
dois elétrons e transportadora temporária de um íon hidreto (o próton H+), que é
removido enzimaticamente da molécula do substrato pela ação de desidrogenases

19
(LEHNINGER, 1991), em reações catabólicas envolvendo a degradação de ácidos
graxos usadas na produção de energia (BRODY, 1994).
As reações reversíveis envolvendo as enzimas NAD(P) desidrogenases
seguem o seguinte esquema geral:
Substrato reduzido + NAD+ ↔ Substrato oxidado + NADH + H+
Substrato reduzido + NADP+ ↔ Substrato oxidado + NADPH + H+
Como a reação é reversível, a função da forma fosforilada reduzida (NADPH +
H+) é atuar como força redutora, na forma de íons H+ (alta energia livre). A produção
de NADPH + H+ pode ocorrer normalmente nas células adiposas, sendo importante
nos tecidos com grande atividade de biossíntese de ácidos graxos a partir de
precursores pequenos, como no tecido adiposo. A biossíntese ocorre por meio de
reações citossólicas, que requerem força de redução química na forma de NADPH
para redução das duplas ligações a ligações simples, ou seja, a forma fosforilada
reduzida (NADPH + H+) pode doar dois elétrons e um íon H+ para o substrato
oxidado, catalisada por desidrogenases específicas (LEHNINGER, 1991).
O efeito da niacina no metabolismo dos lipídeos pode ser explicado por meio da
ação de desidrogenases, enzimas que têm a nicotinamida como grupo ativo das
coenzimas de niacina. Alguns passos metabólicos mediados pelas desidrogenases
encontram-se apresentados a seguir: Na degradação de ácidos graxos:
L-3-hidroxiacil-S-CoA graxo + NAD+ ↔ 3-cetoacil-S-CoA + NADH + H+ (3-
hidroxiacil-CoA desidrogenase).
No ciclo de Krebs (oxidação de unidades de dois carbonos):
Isocitrato + NADP+ (NAD+) ↔ ∝-cetoglutarato + CO2 + NADPH (NADH) + H+
(isocitrato desidrogenase).
∝-cetoglutarato + NAD+ + CoA ↔ Succinil-CoA + NADH + H+ + CO2 (∝-
cetoglutarato desidrogenase).
L-malato + NAD+ ↔ oxaloacetato + NADH + H+ (malato desidrogenase).
Na síntese de ácidos graxos: Oxaloacetato + NADH + H+ ↔ L-malato + NAD+ (malato desidrogenase
citossólica).

20
Acetoacetil + NADPH + H+ ↔ D-3-hidroxibutiril + NADP+ (3-cetoacil-ACP-
redutase).
Trans-butenoil-S-ACP + NADPH + H+ ↔ Butiril-S-ACP + NADP+ (enoil-ACP-
redutase).
Na síntese de glicerol 3-fosfato (substrato para síntese de triglicerídeos): Diidroxiacetona fosfato (da glicólise) + NADH + H+ ↔ L-glicerol 3-fosfato + NAD+
(glicerol fosfato desidrogenase). Na síntese de mevalonato (substrato para síntese de carotenóides): 3-hidroxi-3-metilglutaril-CoA + 2NADPH + 2H+ ↔ mevalonato + CoA + 2NADP+
(hidroximetil glutaril-CoA redutase). Na dessaturação de ácidos graxos:
Palmitoil-CoA + NADPH + H+ + O2 ↔ palmitoleil-CoA + NADP+ + 2OH (acil-CoA
oxigenase).
Estearoil-CoA + NADPH + H+ + O2 ↔ oleil-CoA + NADP+ + 2H2O (acil-CoA
oxigenase).
No metabolismo dos corpos cetônicos:
Acetoacetato + NADH + H+ ↔ D-β-hidroxibutirato + NAD+ (Acetoacetato
desidrogenase).
D-β-hidroxibutirato + NAD+ ↔ Acetoacetato + NADH + H+ (D-β-hidroxibutirato
desidrogenase). 2.3.7 Ação biológica da niacina
O ácido nicotínico tem sido usado com freqüência como medicamento para
controlar os níveis de lipídeos no organismo. Sua atividade hipolipidêmica decorre
da capacidade de inibir a lipólise do tecido adiposo, de reduzir a esterificação dos
triglicerídeos no fígado e de aumentar a atividade da lipase lipoprotéica
(GOODMANS et al., 1996).
Segundo GOTTO JR. (1998), o controle da hipertrigliceridemia deve ser
iniciado por meio de terapia não farmacológica, como freqüência de atividade física,
controle de peso, redução do uso de bebidas alcoólicas e, em indivíduos diabéticos,

21
um rígido controle de glicemia. Entretanto, se essas medidas forem ineficientes,
opções farmacológicas como o uso de niacina, que reduz os triglicerídeos, as
lipoproteínas de densidade baixa (LDL) e o fibrinogênio, além de aumentar a
lipoproteína de densidade alta (HDL) no plasma, podem ser apropriadas.
Imediatamente após a indução de hiperlipidemia com Triton e após 20
horas, o ácido nicotínico (AN) foi administrado na dose de 5 mg por Kg de peso
corporal, por via intraperitoneal, em ratos que receberam ração comercial (R) e água
à vontade. Após 43 horas da aplicação do Triton, os ratos tratados com R + T + AN
apresentaram 104,56 mg de triglicerídeo por dl de soro, valor inferior (P≤0,05) ao
apresentado por ratos tratados com R + T, que foi de 311,36 mg/dl. O tratamento
com ração proporcionou o nível de 162,30 mg/dl, sendo numericamente superior ao
tratamento com R + T + AN (SANTOS et al., 1999).

22
3. REFERÊNCIAS BIBLIOGRÁFICAS ALBINO, L.F.T., FIALHO, E.T., BLUME, E. (1986) Energia metabolizável e
composição química de alguns alimentos para frangos de corte. Revista da
Sociedade Brasileira de Zootecnia, 15 (3): 184-192.
ANGULO, E., BRUFAU, J., MIQUEL, A., ESTEVE-GARCIA, E. (1993) Effect of diet
density and pelleting on productive parameters of Japanese quail. Poultry
Science, 72:607-610.
BENÍCIO, L.S. (2000) Avicultura: Passado, Presente e Futuro. In: Anais do Simpósio
da XXXVII Reunião Anual da Sociedade Brasileira de Zootecnia, Viçosa - MG:
SBZ, p.61-100.
BERTECHINI, A.G. (1991) Nutrição de Monogástricos. Lavras - MG: Esal/Faepe,
142p.
BORNSTEIN, S., BARTOV, I. (1965) Studies on egg yolk pigmentation: A
comparison between visual scoring of yolk color and colorimetric assay of yolk
carotenoids. Poultry Science, 45:287-296.

23
BRAGAGNOLO, N. (1992) Determinação dos teores de colesterol em carnes, ovos e
massas com ovos. Dissertação (Mestrado em Ciência de Alimentos) - Campinas –
SP, Universidade Estadual de Campinas – UNICAMP - Faculdade de Engenharia
de Alimentos, 105p.
BRODY, T. (1994) Nutritional Biochemistry. California: Academic Press, 658p.
CAMPOS, J. (1955) Efeito do urucum na cor da gema do ovo. Revista Ceres, 9
(53):349-353.
CARVALHO, P.R.N. (1992) Potencialidade dos corantes naturais. Revista Brasileira
de Corantes Naturais, 1 (1):244-245.
CHEN, B.H., BAILEY, C.A. (1988) Effect on turf bermudagrass meal on egg
production, feed utilization, yolk color, and egg weight. Poultry Science, 67:1154-
1156.
FARRELL, D. J., ATMAMIHARDJA, S.I., PYM, R.A.E. (1982) Calorimetric
measurements of the energy and nitrogen metabolism of Japanese quail. British
Poultry Science, 23 (5):375-382.
FURLAN, A. C., OLIVEIRA, A. M., MURAKAMI, A. E., SCAPINELLO, C., MOREIRA,
I., ANDREOTI, M.O. (1998a) Valor energético de alguns alimentos determinados
com codornas Japonesas (Coturnix coturnix japonica). In: Anais da XXXV
Reunião Anual da Sociedade Brasileira de Zootecnia, 35, BOTUCATU-SP:
FMVZ-UNESP, Vol. IV, p. 183-185.
FURLAN, A. C., OLIVEIRA, A. M., MURAKAMI, A. E., SCAPINELLO, C., MOREIRA,
I., CAVALIERI, F.L.B., ANDREOTI, M.O. (1998b) Valores energéticos de alguns
alimentos determinados com codornas Japonesas (Coturnix coturnix japonica).
Revista da Sociedade Brasileira de Zootecnia, 27 (6):1147-1150.
GOODMANS, J.G.; GILMAN, A.G.; LIMBIRD, L.E. (1996) The pharmacological basis
of therapeutics. 9a Ed. The McGraw-Hill Company, 1843p.

24
GOTTO JR., A.M. (1998) Triglyceride as a risk factor for coronary artery disease.
American Journal of Cardiology, 82:22Q-25Q.
GRIFFIN, H.D., GODDARD, C., BUTTERWITH, S.C. (1999) Control of lean and fat
deposition in birds. In: Boorman, K.N., Buttery, P.J., Lindsay, D.B. (eds.) The
control of fat and lean deposition. Oxford: Butterworth-Heinemann, p. 231-248.
HAMMAD, S.M., SIEGEL, H.S., MARKS, H.L. (1998) Total cholesterol, total
triglycerides, and cholesterol distribution among lipoproteins as predictors of
atherosclerosis in selected lines of Japanese quail. Comparative Biochemistry and
Physiology, 119A (2):485-492.
HAN, I.K., HOCHSTETLER, H.W., SCOTT, M.L. (1976) Metabolizable energy values
of some poultry feeds determined by various methods and their estimation using
metabolizability of the dry matter. Poultry Science, 55:1335-1342.
HENRY, B.S. (1992) Natural food colours. In: Hendry, G.A.F., Houghton, J.D. (eds.)
Natural Food Colorants. New York: AVI, p. 39-78.
HERMIER, D. (1997) Lipoprotein metabolism and fattening in poultry. Journal of
Nutrition, 127:805S-808S.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA – IBGE (1997) Anuário
estatístico do Brasil, p. 3-52, 3-53, 3-75, 3-85.
JÚNIOR, H.M.E. (1981) Carcass composition studies: Influences of age, sex and
calorie-protein content of the diet on carcass composition of japanese quail.
Poultry Science, 60 (11):2506-2512.
KESSLER, A.M., JÚNIOR, P.N.S. (2001) Considerações sobre a quantidade de
gordura na carcaça do frango. A produção animal na visão dos brasileiros.
Piracicaba - SP: Fealq, p. 111-133.

25
LEHNINGER, A.L. (1991) Princípios de Bioquímica. Ribeirão Preto - SP: USP,
Departamento de Bioquímica, 725p.
LEVY, L.W., REGALADO, E., NAVARRETE, S., WATKINS, R.H. (1997) Bixin and
norbixin in human plasma: determination and study of the absorption of a single
dose of annatto food color. Analyst, 122:977-980.
LIMA, L.R.P., OLIVEIRA, T.T., NAGEM, T.J., OLIVEIRA, M.G.A., PINTO, A.S.,
TINOCO, A.L.A., STRINGHETA, P.C., OLIVEIRA, M.L. (1999) Efeito de
flavonóides (luteolina e apigenina) e corantes naturais (norbixina e bixina) do
urucum sobre a atividade da lipase. Revista da Escola de Farmácia e Odontologia
de Alfenas, 1 (21):29-36.
LIMA, L.R.P., OLIVEIRA, T.T., NAGEM, T.J., PINTO, A.S., STRINGHETA, P.C.,
TINOCO, A.L.A., SILVA, J.F. (2001) Bixina, Norbixina e Quercetina e seus efeitos
no metabolismo lipídico de coelhos. Brazilian Journal of Veterinary Research and
Animal Science, 38 (4):196-200.
McCREADY, R.M., McCOMB, E.A. (1952) Extraction and determination of total
pectic materials in fruits. Analytical Chemistry, 24 (12):1986-1988.
MATTERSON, L.B., POTTER, L.M., STUTZ, M.W., SINGSEN, E.P. (1965) The
metabolizable energy of feed ingredients for chickens. Research Report, 7:3-11.
MORAN JR., E.T. (1994) Digestão e absorção de gorduras. In: Mendes, A.A. (ed.)
Fisiologia da Digestão e Absorção das Aves. Campinas - SP: FACTA, p. 71-82.
MURAKAMI, A.E. (1998) Nutrição e alimentação de codornas em postura. In: Anais
do Simpósio sobre Nutrição Animal e Tecnologia da Produção de Rações,
Campinas - SP: CBNA, p. 19-38.
MURAKAMI, A.E., MORAES, V.M.B., ARIKI, J., JUNQUEIRA, O.M., KRONKA, S.N.
(1993) Níveis de proteína e energia em rações para codornas Japonesas

26
(Coturnix coturnix japonica) em crescimento. Revista da Sociedade Brasileira de
Zootecnia, 22 (4):534-540.
NATIONAL RESEARCH COUNCIL – NRC (1994) Nutrient Requirements of Poultry.
9.ed. Washington: D.C. National Academy Press, 155p.
OLIVEIRA, N.T.E. (2000) Exigências de proteína bruta e energia metabolizável para
codornas japonesas criadas para a produção de carne. Dissertação (Mestrado
em Produção Animal) – Campos dos Goytacazes – RJ, Universidade Estadual do
Norte Fluminense – UENF, 98p.
PAUMGARTTEN, F.J.R., CARVALHO, R.R., ARAÚJO, I.B., PINTO, F.M., BORGES,
O.O., SOUZA, C.A.M., KURIYAMA, S.N. (2002) Evaluation of the developmental
toxicity of annatto in the rat. Food and Chemical Toxicology, 40:1595-1601.
PEREIRA, A.V., KISHIBE, R., ARIKI, J., BORGES, S.A., LODDI, M.M. (2000) Bixina
e norbixina como agentes pigmentantes da gema de ovos de poedeiras
comerciais. In: Anais dos trabalhos da XXXVII Reunião Anual da Sociedade
Brasileira de Zootecnia. Viçosa - MG: SBZ, p.259.
PEREIRA, A.V., ARIKI, J., LODDI, M.M., KISHIBE, R. (2001) Bixina como agente
pigmentante das gemas de ovos de poedeiras comerciais. In: Anais dos trabalhos
da XXXVIII Reunião Anual da Sociedade Brasileira de Zootecnia. Piracicaba - SP:
SBZ, p.839-840.
ROSTAGNO, H.S. (1994) Carboidratos. In: Mendes, A.A. (ed.) Fisiologia da Digestão
e Absorção das Aves. Campinas - SP: FACTA, p. 43-58.
ROSTAGNO, H.S., ALBINO, L.F.T., DONZELE, J.L., GOMES, P.C., FERREIRA,
A.S., OLIVEIRA, R.F., LOPES, D.C. (2000) Tabelas Brasileiras para Aves e
Suínos: Composição de Alimentos e Exigências Nutricionais. 1. ed. Viçosa - MG:
UFV, Departamento de Zootecnia, 141p.

27
SANTOS, K.F.R., OLIVEIRA, T.T., NAGEM, T.J., PINTO, A.S., OLIVEIRA, M.G.A.,
SOARES, J.F. (1999) Efeitos das associações de morina-ácido nicotínico e
quercetina-ácido nicotínico no controle de lipídeos. Revista Brasileira de Análises
Clínicas, 31 (1):5-7.
SAVORY, C.J., GENTLE, J. (1976ª) Effects of dietary dilution with fibre on the food
intake and gut dimensions of Japanese quail. British Poultry Science, 17:561-570.
SAVORY, C.J., GENTLE, J. (1976b) Changes in food intake and gut size in
Japanese quail in response to manipulation of dietary fibre content. British Poultry
Science, 17:561-570.
SIBBALD, I.R. (1980) The effects of dietary cellulose and saud on the combined
metabolic plus endogenous energy and aminoacid outputs of adult cockerels.
Poultry Science, 59:836-844.
SILVA, D.J. (1990) Análise de alimentos: Métodos químicos e biológicos. 2 ed.
Viçosa - MG: UFV, Imprensa Universitária, 165 p.
SILVA, J.H.V., ALBINO, L.F.T. GODOI, M.J.S. (2000) Efeito do extrato de urucum na
pigmentação da gema dos ovos. Revista Brasileira de Zootecnia, 29 (5):1435-
1439.
SOUZA, E.C.G., STRINGHETA, P.C., OLIVEIRA, T.T., NAGEM, T.J., PINTO, A.S.,
SILVA, J.F., LEÃO, M.A. (2000) Efeitos de bixina sobre o metabolismo lipídico. In:
Anais da 23a Reunião Anual da Sociedade Brasileira de Química, Poços de
Caldas - MG: SBQ, QB-017.
SOUZA, E.M. (2000) Melhoramento do frango de corte. In: Anais do Simpósio da
XXXVII Reunião Anual da Sociedade Brasileira de Zootecnia. Viçosa - MG: SBZ,
p.1-8.

28
TOCCHINI, L., MERCADANTE, A.Z. (2001) Extração e determinação, por CLAE, de
bixina e norbixina em coloríficos. Ciência e Tecnologia de Alimentos, 21 (3):310-
313.
UNIVERSIDADE FEDERAL DE VIÇOSA (1997) SAEG - Sistema de análises
estatísticas e genéticas: Versão 7.1. Viçosa - MG: UFV, 150p.
VOHRA, P. (1971) A review of the nutrition of Japanese quail. World's Poultry
Science Journal, 27 (1)26-34.
YALÇIN, S., OGUZ, I., OTLES, S. (1995) Carcase characteristics of quail (Coturnix
coturnix japonica) slaughtered at different ages. British Poultry Science, 36
(3):393-399.
WILSON, H.R., DOUGLAS, C.R., NESBETH, W.G. (1977) Feed consumption and
protein efficiency by bobwhite quail in response to dietary energy levels. Poultry
Science, 56:1127-1129.
WHO (2000) Safety evaluation of certain food additives and contaminants: Evaluation
of national assessments of intake of annatto extracts (bixin). WHO Food Additive
Series, 44:485-492.

29
4. TRABALHOS
Os trabalhos a seguir foram elaborados segundo as normas da Revista
Arquivo Brasileiro de Medicina Veterinária e Zootecnia:
DETERMINAÇÃO DA ENERGIA METABOLIZÁVEL DE ALGUNS ALIMENTOS TESTADOS EM CODORNAS JAPONESAS FÊMEAS1
NEWTON TAVARES ESCOCARD DE OLIVEIRA2, JOSÉ BRANDÃO FONSECA3, RITA DA
TRINDADE RIBEIRO NOBRE SOARES3, CLÁUDIO TEIXEIRA LOMBARDI4, MARIA BEATRIZ MERCADANTE4
1 Parte da tese de doutorado que será apresentada à Universidade Estadual do Norte Fluminense pelo primeiro autor. 2 Estudante de doutorado do Centro de Ciências e Tecnologias Agropecuárias - Universidade Estadual do Norte Fluminense. Av. Alberto Lamego, 2000 - Campos dos Goytacazes - RJ CEP: 28015 - 620. 3 Docentes do Centro de Ciências e Tecnologias Agropecuárias - Universidade Estadual do Norte Fluminense. Av. Alberto Lamego, 2000 - Campos dos Goytacazes - RJ CEP: 28015 - 620. 4 Técnicos de Nível Superior do Laboratório de Zootecnia e Nutrição Animal do Centro de Ciências e Tecnologias Agropecuárias - Universidade Estadual do Norte Fluminense. Av. Alberto Lamego, 2000 - Campos dos Goytacazes - RJ CEP: 28015 - 620.
PIGMENTAÇÃO DE GEMAS DE OVOS DE CODORNAS JAPONESAS
ALIMENTADAS COM RAÇÕES CONTENDO COLORÍFICO1
NEWTON TAVARES ESCOCARD DE OLIVEIRA2, JOSÉ BRANDÃO FONSECA3, RITA DA TRINDADE RIBEIRO NOBRE SOARES3, KARLA SILVA FERREIRA3
1 Parte da tese de doutorado que será apresentada à Universidade Estadual do Norte Fluminense pelo primeiro autor. 2 Estudante de doutorado do Centro de Ciências e Tecnologias Agropecuárias - Universidade Estadual do Norte Fluminense. Av. Alberto Lamego, 2000 - Campos dos Goytacazes - RJ CEP: 28015 - 620. 3 Docentes do Centro de Ciências e Tecnologias Agropecuárias - Universidade Estadual do Norte Fluminense. Av. Alberto Lamego, 2000 - Campos dos Goytacazes - RJ CEP: 28015 - 620.

30
COLORÍFICO E NIACINA SUPLEMENTAR EM RAÇÕES DE CODORNAS JAPONESAS MACHOS: TRIGLICERÍDEOS SANGÜÍNEOS E COMPOSIÇÃO
QUÍMICA DA CARNE E CARCAÇA1
NEWTON TAVARES ESCOCARD DE OLIVEIRA2, JOSÉ BRANDÃO FONSECA3, RITA DA TRINDADE RIBEIRO NOBRE SOARES3, KARLA SILVA FERREIRA3
1 Parte da tese de doutorado que será apresentada à Universidade Estadual do Norte Fluminense pelo primeiro autor. 2 Estudante de doutorado do Centro de Ciências e Tecnologias Agropecuárias - Universidade Estadual do Norte Fluminense. Av. Alberto Lamego, 2000 - Campos dos Goytacazes - RJ CEP: 28015 - 620. 3 Docentes do Centro de Ciências e Tecnologias Agropecuárias - Universidade Estadual do Norte Fluminense. Av. Alberto Lamego, 2000 - Campos dos Goytacazes - RJ CEP: 28015 - 620.

31
DETERMINAÇÃO DA ENERGIA METABOLIZÁVEL DE ALGUNS ALIMENTOS TESTADOS EM CODORNAS JAPONESAS FÊMEAS
RESUMO
Foram determinados os valores de energia metabolizável aparente (EMA),
energia metabolizável aparente corrigida por retenção de nitrogênio (EMAn) e
matéria seca metabolizável aparente (MSMA) de sete alimentos testados em rações
para codornas japonesas. Utilizaram-se 280 codornas japonesas (Coturnix japonica)
fêmeas com idade inicial de vinte e seis dias, em delineamento experimental
inteiramente casualizado, com cinco repetições e sete codornas por unidade
experimental. Os tratamentos consistiram de sete rações experimentais e uma ração
referência. Cada ração experimental foi constituída, na base da matéria natural, por
70 % da ração referência e 30 % do alimento a ser testado, à exceção da ração
contendo óleo de soja degomado, que foi testado, na base da matéria natural, com
10 % de inclusão e 90 % da ração referência. O ensaio biológico foi conduzido em
gaiolas de arame galvanizado distribuídas em baterias de madeira. Os respectivos
valores de EMA e EMAn (kcal por kg de matéria natural) e MSMA (%) do milho
moído, colorífico, quirera de arroz, óleo de soja degomado, farelo de soja, semente
de linhaça e casca + polpa de abacaxi foram iguais a 3.124, 3.122 e 82,95; 3.152,
3.102 e 82,30; 2.980, 2.947 e 89,12; 8.065, 7.940 e 72,47; 2.633, 2.651 e 46,29;
2.477, 2.492 e 32,11 e 1.323, 1.274 e 42,11. Os valores de EMAn do milho e do
farelo de soja foram cerca de 7,4 % inferior e 14,6 % superior, respectivamente, aos
encontrados em tabela brasileira de composição de alimentos. Há necessidade de
se realizar outras pesquisas objetivando determinar a energia metabolizável de
alimentos para codornas japonesas.
Palavras-chave: alimento, codorna japonesa, digestibilidade, energia metabolizável,
matéria seca metabolizável aparente
ABSTRACT
The values of the apparent metabolizable energy (AME), apparent
metabolizable energy corrected for nitrogen retention (AMEn) and dry matter

32
apparently metabolizable (DMAM) were determined in seven foodstuffs tested in
japanese quail feedings. Two hundred and eighty japanese female quails (Coturnix
japonica), twenty-six days of age were used in a completely randomized design, with
five replicates and seven quails per experimental unit. The treatments consisted of
seven experimental rations and one reference ration. Each experimental ration was
constituted, as fed basis, for 70 % of the reference ration and 30 % of food that would
be tested, save the ration that contained soy oil without phospholipid, that was
tested, as fed basis, for 10 % of addition and 90 % of the reference ration. The
biological assay was carried out in wired floor cages distributed in wood batteries.
The respective values of AME and AMEn (kcal/kg of fed basis) and DMAM (%) to
ground corn, spice, broken rice grains, soy oil without phospholipid, soybean meal,
linseed meal and pineapple (peel and pulp) were 3.124, 3.122 and 82,95; 3.152,
3.102 and 82,30; 2.980, 2.947 and 89,12; 8.065, 7.940 and 72,47; 2.633, 2.651 and
46,29; 2.477, 2.492 and 32,11 and 1.323, 1.274 and 42,11. AMEn values to corn
ground and soybean meal were about 7,4 % lower and 14,6 % higher, respectively,
to those found in the Brazilian table foodstuff composition. It’s necessary to carry out
other researches to establish the metabolizable energy of foodstuffs for japanese
quails. Key words: digestibility, dry matter apparently metabolizable, food, japanese quail,
metabolizable energy
INTRODUÇÃO
Como conseqüência do potencial de produção da codorna japonesa
(Coturnix japonica), tem havido um recente e crescente interesse em pesquisar
aspectos nutricionais desta espécie. Dentre esses aspectos, os estudos sobre
exigências de energia metabolizável (EM) e de determinação da EM de alimentos
têm se destacado. Uma das explicações para tal importância reside no fato de que
as exigências de proteínas, aminoácidos e outros nutrientes são freqüentemente
expressas em função dos níveis de EM da ração, os quais influem ativamente sobre
o consumo de ração e, conseqüentemente, sobre o desempenho das codornas.
Um problema que tem ocorrido com freqüência nos estudos e ensaios com a
codorna japonesa é o uso de valores de EM de alimentos testados com frangos de

33
corte ou galinhas poedeiras em formulações de rações para codornas japonesas.
Essa situação não se justifica em razão da existência de diferenças anatômicas,
histológicas e de tamanho e comprimento do trato gastrintestinal entre essas
espécies, além de haver interações entre alimento e trato gastrintestinal, causadas
pelas diferenças de composição química e estrutura física dos alimentos, interferindo
na eficiência dos processos digestivo e absortivo.
O comprometimento do desempenho produtivo de codornas japonesas pode
ser evidenciado quando essas aves consomem rações cujos valores de energia
metabolizável (EM) estão discrepantes das suas exigências energéticas.
A recomendação de se utilizar 2.900 kcal de EM por kg de ração (NRC,
1994), para atendimento das exigências de manutenção e de produção de codornas
japonesas em fase inicial, de crescimento e reprodutiva, tem sido freqüentemente
utilizada como nível energético padrão em pesquisas envolvendo nutrição de
codornas (OLIVEIRA, 2000 e MURAKAMI et al., 1993). Contudo, o uso de valores de
EM de alimentos testados com outras espécies e extrapolados para a formulação de
rações para codornas pode superestimar ou subestimar o conteúdo real de EM das
rações. Assim, as diversas recomendações, feitas por diferentes autores, podem ter
sido baseadas em diferentes conteúdos reais de EM nas rações, causando
desuniformidade nas respostas.
Testando o milho e o farelo de soja em machos de codornas japonesas com
idade de 65 dias, os valores de energia metabolizável aparente (EMA) foram de
3.444 e 2.565 e os de energia metabolizável aparente corrigida por retenção de
nitrogênio (EMAn) foram iguais a 3.429 e 2.593 kcal por kg de matéria natural,
respectivamente (FURLAN et al., 1998b). Por sua vez, ALBINO et al. (1986)
encontraram valores energéticos bem superiores aos reportados por FURLAN et al.
(1998b). Testados em frangos com quarenta e dois dias de idade, o milho moído
proporcionou 3.992 kcal de EMA por kg, 3.909 kcal de EMAn por kg e 82,91 % de
matéria seca metabolizável aparente (MSMA) enquanto o farelo de soja forneceu
2.906 kcal de EMA por kg, 2.852 kcal de EMAn por kg e 42,74 % de MSMA.
De acordo com as tabelas brasileiras, os valores de EMAn e de energia
metabolizável verdadeira corrigida por retenção de nitrogênio (EMVn) do milho em
grão, farelo de soja (45 % de proteína bruta), quirera de arroz, óleo de soja e óleo de
milho são, respectivamente, de 3.371 e 3.450, 2.266 e 2.495, 3.273 e 3.507, 8.790 e
9.200 e 8.886 e 9.250 kcal por kg de matéria natural. Esses valores têm sido

34
freqüentemente usados em formulações de rações para codornas, mas foram
obtidos utilizando-se galos, galinhas poedeiras e pintos (ROSTAGNO et al., 2000).
Segundo MURAKAMI (1998), os valores de energia metabolizável (EM) de
alimentos fibrosos, como farelo de trigo e feno de alfafa, testados com codornas
foram superiores aos valores de EM desses alimentos, testados com galinhas. Em
relação ao milho e à banha, os valores de EM obtidos com codornas foram similares
aos de galinhas.
Nesse contexto, torna-se relevante determinar, por meio de ensaio de
digestibilidade, os valores de energia metabolizável aparente e energia
metabolizável aparente corrigida por retenção de nitrogênio de alguns alimentos
utilizados em rações para codornas japonesas em fase de crescimento. Estes
resultados podem proporcionar maior disponibilidade de dados sobre digestibilidade
de alimentos para essa espécie, servindo de fonte de pesquisa para trabalhos
futuros, haja vista que alguns desses alimentos são pouco freqüentes nas tabelas
nutricionais brasileiras.
MATERIAL E MÉTODOS
Local do experimento O experimento foi realizado no Núcleo de Pesquisa do Laboratório de
Zootecnia e Nutrição Animal (LZNA), situado na Escola Técnica Estadual Agrícola
“Antônio Sarlo”, Campus Experimental da Universidade Estadual do Norte
Fluminense (UENF), localizada no município de Campos dos Goytacazes. Distribuição das codornas no delineamento experimental e tratamentos
Foram utilizadas 280 codornas japonesas (Coturnix japonica) fêmeas, em
delineamento experimental inteiramente casualizado, com oito tratamentos, cinco
repetições e sete codornas por unidade experimental. Os tratamentos foram constituídos por oito rações, sendo sete experimentais
e uma ração referência. Cada ração experimental foi constituída, na base da matéria
natural, por 70 % da ração referência e 30 % do alimento a ser testado, à exceção
da ração contendo óleo de soja degomado, que foi testado, na base da matéria
natural, com 10 % de inclusão e 90 % da ração referência. Os alimentos testados

35
foram: milho, colorífico, quirera de arroz, óleo de soja degomado, farelo de soja,
semente de linhaça e casca + polpa de abacaxi, oriunda da Fábrica de Sucos Bela
Joana, situada no município de Campos dos Goytacazes - RJ. Alojamento das codornas nas instalações e manejo
As codornas com idade inicial de vinte e seis dias foram alojadas em galpão
contendo duas baterias de madeira, as quais agruparam dez gaiolas metálicas de
quatro divisões internas cada. Cada gaiola mediu 1,00 m de comprimento x 0,34 m
de largura x 0,24 m de altura e abrigou 28 codornas.
As codornas receberam água e ração à vontade, em bebedouros e
comedouros de chapa galvanizada, respectivamente, e ficaram expostas a 24 horas
de luz diária, natural ou artificial, durante todo o período experimental, a fim de
incentivar o consumo das rações.
Análises químicas
Em todas as rações e excretas, realizaram-se análises químicas referentes
aos teores de matéria seca (MS) e nitrogênio (SILVA, 1990), no Laboratório de
Zootecnia e Nutrição Animal (LZNA), do Centro de Ciências e Tecnologias
Agropecuárias (CCTA), da Universidade Estadual do Norte Fluminense (UENF).
Os valores de energia bruta (EB) das rações e das excretas foram
quantificados por meio de bomba calorimétrica PARR. Tais análises foram feitas
utilizando-se as instalações e equipamentos do Laboratório de Nutrição Animal e
Pastagens, do Instituto de Zootecnia, da Universidade Federal Rural do Rio de
Janeiro (UFRRJ), situada no Km 47, município de Seropédica - RJ.
Os valores de MS, nitrogênio e EB das rações encontram-se no Quadro 1.
Quadro 1. Valores médios de matéria seca (MS), nitrogênio (N) e energia bruta (EB)
da ração referência (RR) e das rações experimentais (RE)
Tratamentos MS (%) N na MS (%) EB (kcal/kg) RR 87,5 3,75 3.697 RE + Milho 87,0 2,94 3.723 RE + Colorífico 87,7 2,80 3.785 RE + Quirera de Arroz 87,0 2,64 3.637 RE + Óleo de Soja Degomado 88,7 3,32 4.269 RE + Farelo de Soja 87,5 4,91 3.928 RE + Semente de Linhaça 88,7 3,66 4.405 RE + Casca + Polpa de Abacaxi 87,4 2,74 3.670
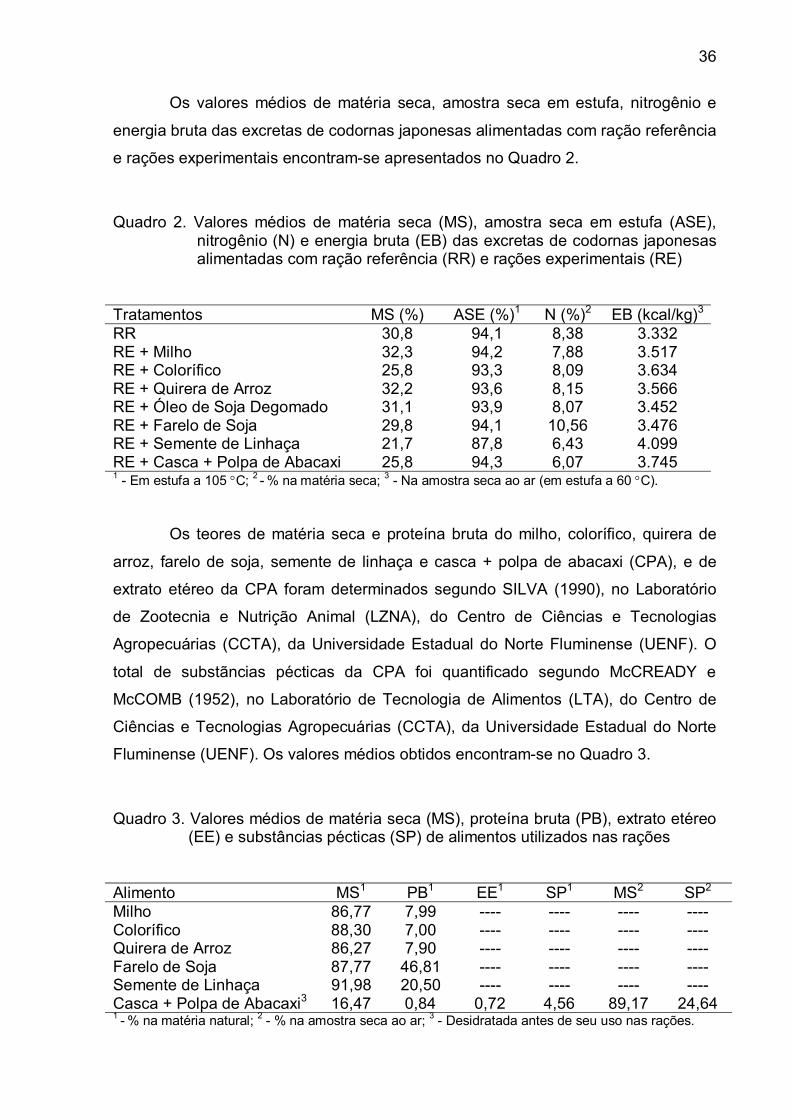
36
Os valores médios de matéria seca, amostra seca em estufa, nitrogênio e
energia bruta das excretas de codornas japonesas alimentadas com ração referência
e rações experimentais encontram-se apresentados no Quadro 2.
Quadro 2. Valores médios de matéria seca (MS), amostra seca em estufa (ASE),
nitrogênio (N) e energia bruta (EB) das excretas de codornas japonesas alimentadas com ração referência (RR) e rações experimentais (RE)
Tratamentos MS (%) ASE (%)1 N (%)2 EB (kcal/kg)3 RR 30,8 94,1 8,38 3.332 RE + Milho 32,3 94,2 7,88 3.517 RE + Colorífico 25,8 93,3 8,09 3.634 RE + Quirera de Arroz 32,2 93,6 8,15 3.566 RE + Óleo de Soja Degomado 31,1 93,9 8,07 3.452 RE + Farelo de Soja 29,8 94,1 10,56 3.476 RE + Semente de Linhaça 21,7 87,8 6,43 4.099 RE + Casca + Polpa de Abacaxi 25,8 94,3 6,07 3.745 1 - Em estufa a 105 °C; 2 - % na matéria seca; 3 - Na amostra seca ao ar (em estufa a 60 °C).
Os teores de matéria seca e proteína bruta do milho, colorífico, quirera de
arroz, farelo de soja, semente de linhaça e casca + polpa de abacaxi (CPA), e de
extrato etéreo da CPA foram determinados segundo SILVA (1990), no Laboratório
de Zootecnia e Nutrição Animal (LZNA), do Centro de Ciências e Tecnologias
Agropecuárias (CCTA), da Universidade Estadual do Norte Fluminense (UENF). O
total de substãncias pécticas da CPA foi quantificado segundo McCREADY e
McCOMB (1952), no Laboratório de Tecnologia de Alimentos (LTA), do Centro de
Ciências e Tecnologias Agropecuárias (CCTA), da Universidade Estadual do Norte
Fluminense (UENF). Os valores médios obtidos encontram-se no Quadro 3.
Quadro 3. Valores médios de matéria seca (MS), proteína bruta (PB), extrato etéreo
(EE) e substâncias pécticas (SP) de alimentos utilizados nas rações
Alimento MS1 PB1 EE1 SP1 MS2 SP2 Milho 86,77 7,99 ---- ---- ---- ---- Colorífico 88,30 7,00 ---- ---- ---- ---- Quirera de Arroz 86,27 7,90 ---- ---- ---- ---- Farelo de Soja 87,77 46,81 ---- ---- ---- ---- Semente de Linhaça 91,98 20,50 ---- ---- ---- ---- Casca + Polpa de Abacaxi3 16,47 0,84 0,72 4,56 89,17 24,64 1 - % na matéria natural; 2 - % na amostra seca ao ar; 3 - Desidratada antes de seu uso nas rações.
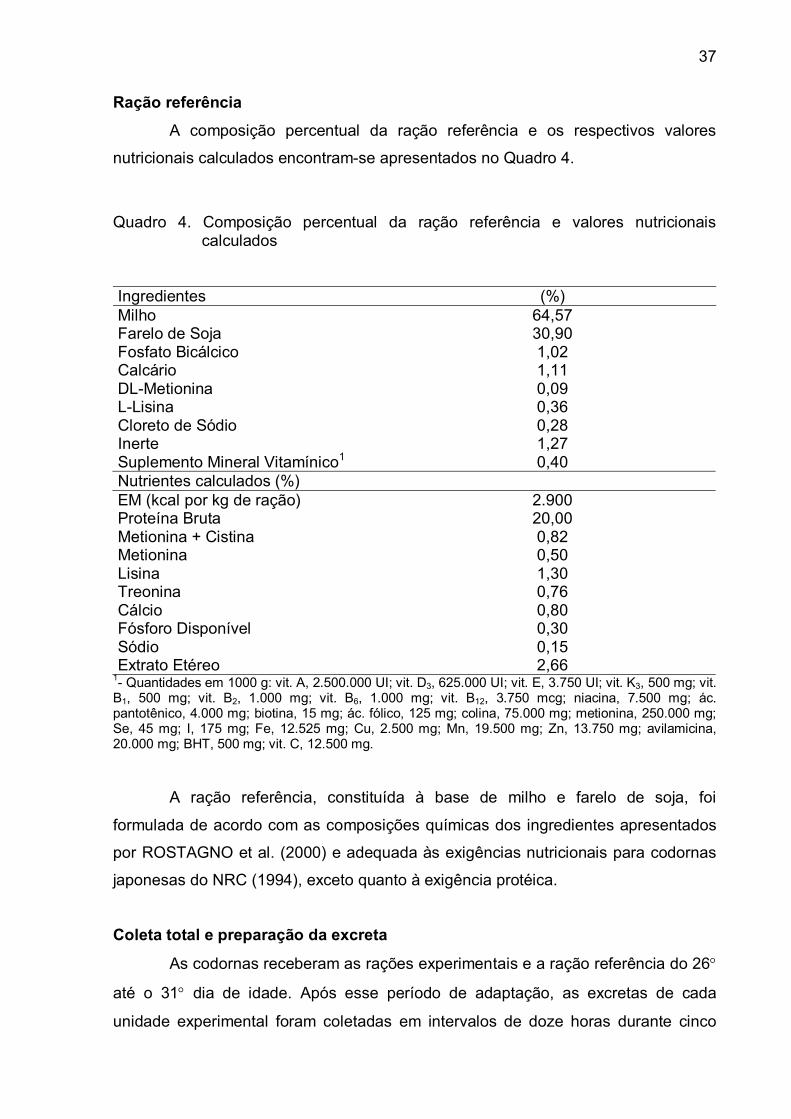
37
Ração referência
A composição percentual da ração referência e os respectivos valores
nutricionais calculados encontram-se apresentados no Quadro 4.
Quadro 4. Composição percentual da ração referência e valores nutricionais calculados
Ingredientes (%) Milho 64,57 Farelo de Soja 30,90 Fosfato Bicálcico 1,02 Calcário 1,11 DL-Metionina 0,09 L-Lisina 0,36 Cloreto de Sódio 0,28 Inerte 1,27 Suplemento Mineral Vitamínico1 0,40 Nutrientes calculados (%) EM (kcal por kg de ração) 2.900 Proteína Bruta 20,00 Metionina + Cistina 0,82 Metionina 0,50 Lisina 1,30 Treonina 0,76 Cálcio 0,80 Fósforo Disponível 0,30 Sódio 0,15 Extrato Etéreo 2,66
1- Quantidades em 1000 g: vit. A, 2.500.000 UI; vit. D3, 625.000 UI; vit. E, 3.750 UI; vit. K3, 500 mg; vit. B1, 500 mg; vit. B2, 1.000 mg; vit. B6, 1.000 mg; vit. B12, 3.750 mcg; niacina, 7.500 mg; ác. pantotênico, 4.000 mg; biotina, 15 mg; ác. fólico, 125 mg; colina, 75.000 mg; metionina, 250.000 mg; Se, 45 mg; I, 175 mg; Fe, 12.525 mg; Cu, 2.500 mg; Mn, 19.500 mg; Zn, 13.750 mg; avilamicina, 20.000 mg; BHT, 500 mg; vit. C, 12.500 mg.
A ração referência, constituída à base de milho e farelo de soja, foi
formulada de acordo com as composições químicas dos ingredientes apresentados
por ROSTAGNO et al. (2000) e adequada às exigências nutricionais para codornas
japonesas do NRC (1994), exceto quanto à exigência protéica. Coleta total e preparação da excreta
As codornas receberam as rações experimentais e a ração referência do 26°
até o 31° dia de idade. Após esse período de adaptação, as excretas de cada
unidade experimental foram coletadas em intervalos de doze horas durante cinco

38
dias consecutivos (31° ao 36° dia de idade) nas bandejas das gaiolas, as quais
foram removidas a cada coleta. Antecedendo ao início do período de coleta das
excretas, as codornas foram submetidas a um jejum alimentar durante
aproximadamente três horas, visando a remover qualquer resíduo alimentar do trato
gastrintestinal, referente à ração fornecida no período de adaptação. O final da
coleta ocorreu após novo jejum alimentar, que teve a finalidade de retirar e coletar
totalmente o conteúdo alimentar relativo ao período experimental propriamente dito
(31° ao 36° dia de idade).
Foram mensurados o consumo de ração e a quantidade de excretas das
codornas alimentadas com ração referência e com rações experimentais, cujos
valores médios encontram-se apresentados no Quadro 5. Atenção especial foi dada
à redução da contaminação das excretas com penas, resíduos de ração e outros.
Quadro 5. Valores médios1 de consumo de ração e quantidade de excretas das
codornas no período de 31 a 36 dias de idade
Tratamentos Consumo de ração (kg) Total de excretas (kg) Ração Referência 0,629 0,565 RE2 + Milho 0,600 0,437 RE + Colorífico 0,547 0,541 RE + Quirera de Arroz 0,616 0,422 RE + Óleo de Soja Degomado 0,558 0,497 RE + Farelo de Soja 0,642 0,728 RE + Semente de Linhaça 0,591 1,026 RE + Casca + Polpa de Abacaxi 0,655 0,878
1 - Obtidos de cinco observações; 2 – Ração Experimental. A cada coleta, as excretas de cada repetição foram acondicionadas em
sacos plásticos, identificadas, pesadas e armazenadas em congelador até o final do
período de coleta, quando foram descongeladas, reunidas por repetição,
homogeneizadas, pesadas e colocadas em estufa de ventilação forçada por 72
horas a 60 °C, onde foi realizada a pré-secagem. A seguir, foram moídas e
armazenadas para análises posteriores, juntamente com amostras das rações
contendo os alimentos a serem testados e da ração referência.

39
Variáveis avaliadas Para cada alimento testado, foram avaliadas a energia metabolizável
aparente (EMA) e a energia metabolizável aparente corrigida pelo balanço de
nitrogênio (EMAn) em kcal por kg de matéria natural e a matéria seca metabolizável
aparente (MSMA) em percentagem.
Os valores de EMA e EMAn (MATTERSON et al., 1965) e de MSMA dos
alimentos (HAN et al., 1976) foram obtidos por meio das seguintes equações:
EMA Ração (kcal/kg MS) = [(MS ingerida (g) x EB Ração) - (MS excretada (g) x EB excretada)] ÷
MS ingerida (g)
EMA Alimento (kcal/kg MS) = EM RR + [(EM RE – EM RR) ÷ PSA RR]
EMAn Ração (kcal/kg MS) = [(MS ingerida (g) x EB Ração) - (MS excretada (g) x EB excretada)]
± 8,22 BN ÷ MS ingerida (g)
BN = (MS ingerida (g) x % N Ração ÷ 100) – (MS excretada (g) x % N Excreta ÷ 100)
EMAn Alimento (kcal/kg MS) = EMAn RR + [(EMAn RE – EMAn RR) ÷ PSA RR]
MSMA Ração (%) = (MS ingerida (g) – MS excretada (g)) ÷ MS ingerida (g) x 100
MSMA Alimento (%) = MSMA RR (%) – [(MSMA RR - MSMA RE) ÷ PSA RR]
Legenda:
MS – matéria seca; RR – ração referência; RE – ração experimental; PSA RR –
proporção de substituição do alimento na ração referência; EB – energia bruta; BN –
balanço de nitrogênio; N – nitrogênio.
8,22 = fator que corresponde a 8,22 kcal de energia por grama de nitrogênio retido.
RESULTADOS E DISCUSSÃO
Os valores médios e os desvios-padrão de EMA e EMAn, expressos em kcal
por kg de matéria natural, e de MSMA (%) encontram-se apresentados no Quadro 6.
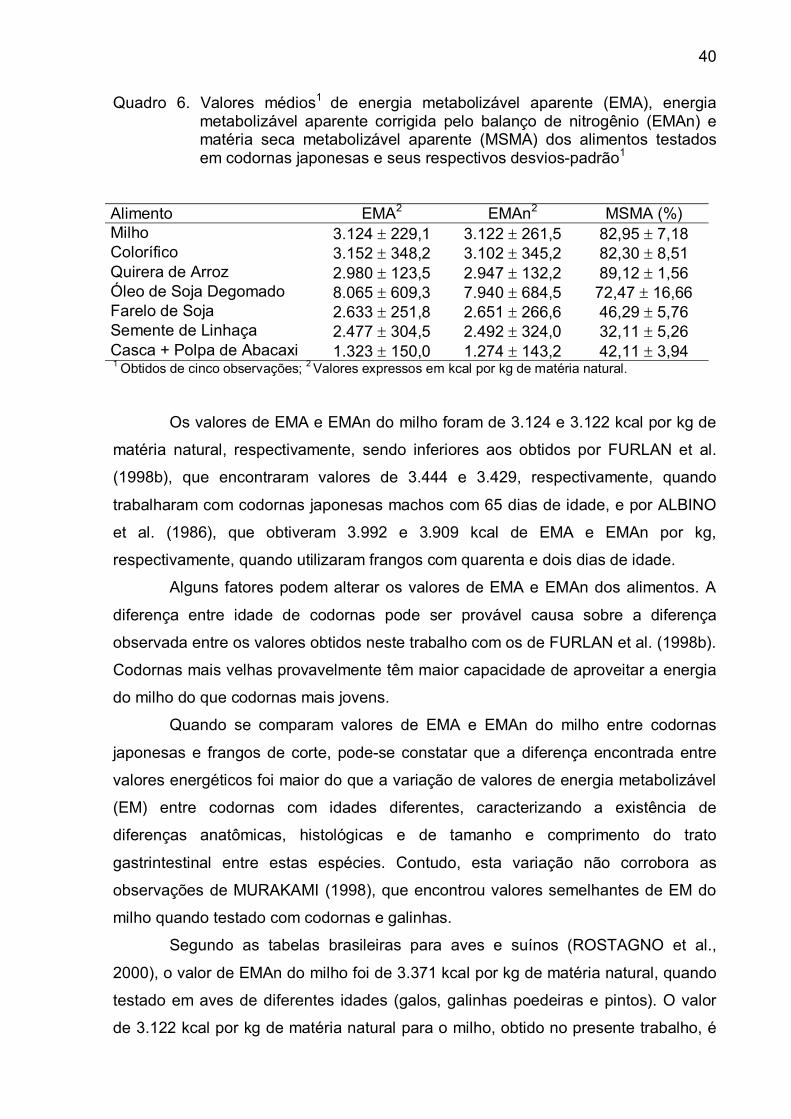
40
Quadro 6. Valores médios1 de energia metabolizável aparente (EMA), energia metabolizável aparente corrigida pelo balanço de nitrogênio (EMAn) e matéria seca metabolizável aparente (MSMA) dos alimentos testados em codornas japonesas e seus respectivos desvios-padrão1
Alimento EMA2 EMAn2 MSMA (%) Milho 3.124 ± 229,1 3.122 ± 261,5 82,95 ± 7,18 Colorífico 3.152 ± 348,2 3.102 ± 345,2 82,30 ± 8,51 Quirera de Arroz 2.980 ± 123,5 2.947 ± 132,2 89,12 ± 1,56 Óleo de Soja Degomado 8.065 ± 609,3 7.940 ± 684,5 72,47 ± 16,66 Farelo de Soja 2.633 ± 251,8 2.651 ± 266,6 46,29 ± 5,76 Semente de Linhaça 2.477 ± 304,5 2.492 ± 324,0 32,11 ± 5,26 Casca + Polpa de Abacaxi 1.323 ± 150,0 1.274 ± 143,2 42,11 ± 3,94 1 Obtidos de cinco observações; 2 Valores expressos em kcal por kg de matéria natural.
Os valores de EMA e EMAn do milho foram de 3.124 e 3.122 kcal por kg de
matéria natural, respectivamente, sendo inferiores aos obtidos por FURLAN et al.
(1998b), que encontraram valores de 3.444 e 3.429, respectivamente, quando
trabalharam com codornas japonesas machos com 65 dias de idade, e por ALBINO
et al. (1986), que obtiveram 3.992 e 3.909 kcal de EMA e EMAn por kg,
respectivamente, quando utilizaram frangos com quarenta e dois dias de idade.
Alguns fatores podem alterar os valores de EMA e EMAn dos alimentos. A
diferença entre idade de codornas pode ser provável causa sobre a diferença
observada entre os valores obtidos neste trabalho com os de FURLAN et al. (1998b).
Codornas mais velhas provavelmente têm maior capacidade de aproveitar a energia
do milho do que codornas mais jovens.
Quando se comparam valores de EMA e EMAn do milho entre codornas
japonesas e frangos de corte, pode-se constatar que a diferença encontrada entre
valores energéticos foi maior do que a variação de valores de energia metabolizável
(EM) entre codornas com idades diferentes, caracterizando a existência de
diferenças anatômicas, histológicas e de tamanho e comprimento do trato
gastrintestinal entre estas espécies. Contudo, esta variação não corrobora as
observações de MURAKAMI (1998), que encontrou valores semelhantes de EM do
milho quando testado com codornas e galinhas.
Segundo as tabelas brasileiras para aves e suínos (ROSTAGNO et al.,
2000), o valor de EMAn do milho foi de 3.371 kcal por kg de matéria natural, quando
testado em aves de diferentes idades (galos, galinhas poedeiras e pintos). O valor
de 3.122 kcal por kg de matéria natural para o milho, obtido no presente trabalho, é

41
7,4 % inferior ao valor relacionado nas tabelas brasileiras, caracterizando as
diferenças encontradas na literatura sobre recomendações de exigências de energia
metabolizável (EM), haja vista que o valor apresentado nas tabelas brasileiras tem
sido utilizado como referencial nas formulações de rações para codornas.
O valor de matéria seca metabolizável aparente (MSMA) do milho
encontrado no presente trabalho (82,95 %) foi similar ao encontrado por ALBINO et
al. (1986), que obtiveram 82,91 % trabalhando com frangos com quarenta e dois
dias de idade. Mesmo considerando a diferença entre espécies de aves, essa
pequena ou quase inexistente variação entre valores pode consolidar as
observações feitas por HAN et al. (1976), que mencionaram que a MSMA pode se
constituir em um excelente parâmetro para estimar a EM de alimentos ricos em
carboidratos e com baixo conteúdo em fibra bruta.
Os valores obtidos neste trabalho referentes à energia metabolizável
aparente (EMA) e energia metabolizável aparente corrigida pelo balanço de
nitrogênio (EMAn) do colorífico foram de 3.152 e 3.102 kcal por kg de matéria
natural, respectivamente. O colorífico é um produto constituído pela mistura de fubá
com urucum em pó ou extrato oleoso de urucum, adicionado ou não de sal e de
óleos comestíveis. Não foram encontrados valores de EM do colorífico na literatura,
haja vista que se trata de um alimento pouco freqüente na formulação de rações,
entretanto, pela sua constituição, pode-se deduzir que seus valores de EMA e EMAn
devem ficar próximos aos do milho. Essa dedução pôde ser confirmada, pois 3.152 e
3.102 kcal por kg de matéria natural foram bem próximos a 3.124 e 3.122 kcal por
kg, valores respectivos de EMA e EMAn encontrados para o milho.
O valor de MSMA para o colorífico foi de 82,30 %, bem próximo do
encontrado para o milho (82,95 %). A MSMA contida no colorífico foi facilmente
convertida em energia pelas codornas, fato que pode ser comprovado pelos valores
de EMA (3.152) e EMAn (3.102) do colorífico, em kcal por kg de matéria natural.
Os valores obtidos neste trabalho referentes à EMA e EMAn foram de 2.980
e 2.947 para a quirera de arroz e 8.065 e 7.940 kcal por kg de matéria natural para o
óleo de soja degomado, respectivamente. Comparando os mesmos alimentos, esses
valores são inferiores aos valores de EMAn da quirera de arroz, óleo de milho e óleo
de soja obtidos nas tabelas brasileiras de composição de alimentos (ROSTAGNO et
al., 2000), que foram de 3.273, 8.886 e 8.790 kcal por kg de matéria natural,
respectivamente, utilizando-se galinhas poedeiras. Em contrapartida, MURAKAMI

42
(1998) revelou que codornas e galinhas obtiveram valores de energia metabolizável
(EM) similares quando utilizaram alimentos ricos em carboidratos, como o milho e a
gordura (banha).
Foram encontrados os valores de 89,12 e 72,47 (%) para a matéria seca
metabolizável aparente (MSMA) da quirera de arroz e do óleo de soja degomado,
respectivamente. Esses resultados mostram que as codornas metabolizaram a maior
parte da matéria seca destes alimentos, convertendo-a em energia para seu
metabolismo, o que pode ser observado pelos valores de EM destes alimentos.
Os valores de energia metabolizável aparente (EMA) e energia
metabolizável aparente corrigida pelo balanço de nitrogênio (EMAn) do farelo de
soja foram de 2.633 e 2.651 kcal por kg de matéria natural. Estes valores são
ligeiramente superiores aos encontrados por FURLAN et al. (1998b), que obtiveram
2.565 e 2.593 kcal por kg de matéria natural, respectivamente, usando codornas
japonesas machos com idade de 65 dias. Esta concordância entre tais valores indica
que devem ser usados valores de EM específicos para codornas, em vez do uso de
valores de EM de alimentos testados com outras espécies de aves.
Nas tabelas brasileiras de composição de alimentos (ROSTAGNO et al.,
2000), é encontrado o valor de 2.266 kcal de EMAn por kg de matéria natural para o
farelo de soja com 45 % de proteína bruta, obtido com galinhas poedeiras. Esse
valor corresponde a, aproximadamente, 14,6 % a menos do que o valor obtido neste
trabalho (2.651 kcal por kg de matéria natural). Todavia, ALBINO et al. (1986)
encontraram 2.906 e 2.852 kcal de EMA e EMAn por kg, respectivamente, para o
farelo de soja testado em frangos com quarenta e dois dias de idade.
Do mesmo modo que ocorreu com os valores de EMAn do milho, houve
variação entre valores de EMAn do farelo de soja quando testado em diferentes
espécies. Outros experimentos sobre digestibilidade de alimentos devem ser
realizados a fim de se obter maior disponibilidade de dados para codornas
japonesas e ratificar as observações sobre o uso indevido de valores de EM de
alimentos testados com outras espécies de aves em rações de codornas.
Foi obtido o valor de 46,29 % para a MSMA do farelo de soja para codornas.
Este valor está próximo e pouco superior ao encontrado por ALBINO et al. (1986),
que mensuraram 42,74 % de MSMA para frangos com quarenta e dois dias de
idade.

43
Os valores de energia metabolizável aparente (EMA) e energia
metabolizável aparente corrigida pelo balanço de nitrogênio (EMAn) da semente de
linhaça foram de 2.477 e 2.492 kcal por kg de matéria natural (MN), sendo um pouco
inferiores aos de EMA (2.633) e EMAn (2.651) do farelo de soja. A moagem das
sementes antes do fornecimento às codornas possivelmente possibilitou um melhor
aproveitamento da energia dos nutrientes, principalmente da parte oleosa, o que
pode ter contribuído para a obtenção de valores satisfatórios de energia
metabolizável (EM), em relação ao baixo valor de matéria seca metabolizável
aparente (MSMA), que foi de 32,11 %.
Esse baixo valor provavelmente foi causado pelo seu conteúdo fibroso, que
promoveu aumento da taxa de passagem e, conseqüentemente, redução do tempo
de permanência da ração no trato gastrintestinal das codornas e aumento das
perdas. Essas perdas foram observadas com a mensuração do total de excretas das
codornas que receberam ração com semente de linhaça, que foi de 1,026 kg e
numericamente bem superior aos demais tratamentos (Quadro 5). Além disso, pode-
se constatar que o percentual de matéria seca das fezes das codornas que
receberam o tratamento com semente de linhaça (21,7 %) foi inferior numericamente
aos demais tratamentos (Quadro 2). A grande quantidade de excretas e o elevado
teor de umidade nas fezes (efeito higroscópico da fibra) podem, possivelmente,
caracterizar o baixo valor de MSMA.
Os valores de EMA e EMAn da casca + polpa de abacaxi (CPA) foram de
1.323 e 1.274 kcal por kg de MN. A literatura não disponibiliza valores de EMA e
EMAn da CPA, um subproduto da indústria do abacaxi cuja composição em pectina,
uma fibra solúvel em água, foi de 24,64 % na amostra seca ao ar, forma pela qual a
CPA foi utilizada nas rações, e 4,56 % na MN (Quadro 3). O valor de EM obtido
indica que as codornas não metabolizam com eficiência a energia contida neste
alimento, possivelmente porque elas não possuem a enzima pectinase e pelo efeito
higroscópico da pectina no trato gastrintestinal, que ocasionou a excreção de 0,878
kg de fezes (Quadro 5) com 25,8 % de matéria seca (Quadro 2). Esta reduzida
metabolização da energia pode ser confirmada pelo baixo valor de MSMA (42,11 %).

44
CONCLUSÕES
1. Os valores de energia metabolizável aparente corrigida pelo balanço de
nitrogênio dos alimentos energéticos milho, quirera de arroz e óleo de soja
degomado foram de 3.122, 2.947 e 7.940 kcal por kg de matéria natural,
respectivamente, sendo bastante inferiores aos valores de energia metabolizável
aparente corrigida pelo balanço de nitrogênio desses alimentos expressos na
literatura.
2. O valor de energia metabolizável aparente corrigida pelo balanço de nitrogênio
do farelo de soja foi de 2.651 kcal por kg de matéria natural, sendo similar ao
reportado na literatura, quando o farelo de soja foi testado com codornas
japonesas, mas bastante superior ao valor de energia metabolizável aparente
corrigida pelo balanço de nitrogênio encontrado nas tabelas brasileiras de
composição de alimentos, quando o farelo de soja foi testado com outras
espécies de aves.
REFERÊNCIAS BIBLIOGRÁFICAS
ALBINO, L.F.T., FIALHO, E.T., BLUME, E. (1986) Energia metabolizável e
composição química de alguns alimentos para frangos de corte. Revista da
Sociedade Brasileira de Zootecnia, 15 (3): 184-192.
FURLAN, A. C., OLIVEIRA, A. M., MURAKAMI, A. E., SCAPINELLO, C., MOREIRA,
I., CAVALIERI, F.L.B., ANDREOTI, M.O. (1998b) Valores energéticos de alguns
alimentos determinados com codornas Japonesas (Coturnix coturnix japonica).
Revista da Sociedade Brasileira de Zootecnia, 27 (6):1147-1150.
HAN, I.K., HOCHSTETLER, H.W., SCOTT, M.L. (1976) Metabolizable energy values
of some poultry feeds determined by various methods and their estimation using
metabolizability of the dry matter. Poultry Science, 55:1335-1342.
MATTERSON, L.B., POTTER, L.M., STUTZ, M.W., SINGSEN, E.P. (1965) The
metabolizable energy of feed ingredients for chickens. Research Report, 7:3-11.

45
McCREADY, R.M., McCOMB, E.A. (1952) Extraction and determination of total
pectic materials in fruits. Analytical Chemistry, 24 (12):1986-1988.
MURAKAMI, A.E. (1998) Nutrição e alimentação de codornas em postura. In: Anais
do Simpósio sobre Nutrição Animal e Tecnologia da Produção de Rações,
Campinas - SP: CBNA, p. 19-38.
MURAKAMI, A.E., MORAES, V.M.B., ARIKI, J., JUNQUEIRA, O.M., KRONKA, S.N.
(1993) Níveis de proteína e energia em rações para codornas Japonesas
(Coturnix coturnix japonica) em crescimento. Revista da Sociedade Brasileira de
Zootecnia, 22 (4):534-540.
NATIONAL RESEARCH COUNCIL – NRC (1994) Nutrient Requirements of Poultry.
9.ed. Washington: D.C. National Academy Press, 155p.
OLIVEIRA, N.T.E. (2000) Exigências de proteína bruta e energia metabolizável para
codornas japonesas criadas para a produção de carne. Dissertação (Mestrado
em Produção Animal) – Campos dos Goytacazes – RJ, Universidade Estadual do
Norte Fluminense – UENF, 98p.
ROSTAGNO, H.S., ALBINO, L.F.T., DONZELE, J.L., GOMES, P.C., FERREIRA,
A.S., OLIVEIRA, R.F., LOPES, D.C. (2000) Tabelas Brasileiras para aves e
suínos - Composição de alimentos e exigências nutricionais. Viçosa: UFV,
Departamento de Zootecnia, 141p.
SILVA, D.J. (1990) Análise de alimentos (Métodos químicos e biológicos). 2 ed.
Viçosa - MG: UFV, Imprensa Universitária, 165 p.

46
PIGMENTAÇÃO DE GEMAS DE OVOS DE CODORNAS JAPONESAS
ALIMENTADAS COM RAÇÕES CONTENDO COLORÍFICO
RESUMO
O objetivo do trabalho foi avaliar o grau de pigmentação de gemas de ovos
de codornas japonesas alimentadas com rações contendo níveis diferentes de
colorífico. Foram utilizadas 240 codornas japonesas (Coturnix japonica) fêmeas, em
delineamento experimental de blocos inteiramente casualizados, com oito
tratamentos, cinco repetições e seis codornas por unidade experimental. Os
tratamentos foram constituídos por oito rações experimentais oriundas da
combinação de quatro níveis de colorífico na ração (0, 1,5, 3,0 e 4,5 %) x duas
fontes energéticas (milho e quirera de arroz). Nos dias 7, 14, 21 e 28 do
experimento, os resultados estimados de máxima coloração de gemas ocorreram
com o uso de 3,15; 2,91; 2,97 e 2,77 % de colorífico nas rações, respectivamente,
que equivalem a estimativas de escores máximos de CG iguais a 9,63; 10,19; 10,34
e 11,08 pontos no leque colorimétrico da Roche, respectivamente. Codornas
alimentadas com rações à base de milho apresentaram gemas de ovos mais
pigmentadas do que as gemas de ovos de codornas que receberam rações à base
de quirera de arroz, em todos os níveis de colorífico utilizados nas rações,
independente do dia experimental. À medida que a idade aumentou, houve
incorporação linear de pigmentos nas gemas, em todos os níveis de colorífico
utilizados nas rações, independente da fonte energética.
Palavras-chave: bixina, codorna japonesa, colorífico, pigmentação de gema de ovo,
quirera de arroz
ABSTRACT
The objective of the research was to evaluate the egg yolk colour grade of
japanese quails fed on rations with different levels of spice. Two hundred and forty
japanese female quails (Coturnix japonica) were used in a completely randomized
block design, with eight treatments, five replicates and six quails per experimental
unit. The treatments consisted of eight experimental rations derived from the

47
combination of four spice levels in the rations (0, 1,5, 3,0 e 4,5 %) x two energetic
sources (corn and broken rice grains). On the 7th, 14th, 21st and 28th experimental
days, the estimated results for maximum yolk colour occurred with the use of 3,15;
2,91; 2,97 e 2,77 % of spice rations, respectively, that are equal to maximum egg
yolk colour scores of 9,63; 10,19; 10,34 and 11,08 points by Roche colorimetric fan,
respectively. Quails fed on corn based rations had greater egg yolk colour than egg
yolk colour of quails that received broken rice grains based rations, in all spice levels
used in the rations, independent of the experimental day. As long as age increased,
linear incorporation of egg yolk pigments occurred, in all spice levels used in the
rations, independent of the energetic source. Key words: bixin, broken rice grains, egg yolk colour, japanese quail, spice
INTRODUÇÃO
Dentre os atributos sensoriais, a cor tem sido correlacionada como indicador
de qualidade da maioria dos alimentos, exercendo papel importante no que se refere
à aceitação dos alimentos pelos consumidores.
Em escala comercial e de forma idêntica à produção de ovos de galinhas
poedeiras, os ovos de codornas são produzidos por meio do fornecimento, às
codornas, de rações de postura à base de milho amarelo e farelo de soja, o que tem
garantido uma pigmentação de gema que varia do amarelo ao laranja. Esta cor é
oriunda da incorporação de xantofilas presentes no milho, principalmente luteína e
zeaxantina.
Embora não ocorra rejeição, é comum a visualização, no momento de seu
consumo, de gemas de ovos de codornas produzidos com pouca coloração. A
intensidade de cor das gemas dos ovos de codornas encontrados no mercado tem
variado em função dos diferentes níveis de inclusão do milho amarelo nas rações de
poedeiras comerciais. Vale realçar que o incremento da pigmentação das gemas “in
natura” resulta em maior pigmentação das gemas cozidas, a forma mais comum de
consumo de ovos de codornas.
Do ponto de vista do criador de aves, a obtenção de gemas com coloração
mais intensa ainda não permite que os ovos alcancem um maior valor comercial no
mercado. Todavia, do ponto de vista nutricional e conseqüentemente de saúde

48
pública, a intensidade da pigmentação de gemas deve ter outro significado, haja
vista os importantes efeitos fisiológicos no organismo humano atribuído aos
carotenóides, substâncias que têm sido relacionadas como agentes preventivos de
diversas patologias. A bixina, carotenóide predominante nos coloríficos, têm sido
mencionada como uma das substâncias naturais mais efetivas na eliminação de
radicais livres, os quais podem causar patologias como câncer, arteriosclerose, entre
outras (LEVY et al., 1997).
Muitas pesquisas têm mostrado ser possível e viável modular a
concentração de alguns nutrientes, em produtos de origem animal, por meio da
manipulação nutricional das rações. A produção de gemas de maior pigmentação e,
conseqüentemente, com maior deposição de carotenóides, pode ser obtida pela
inclusão de corantes lipossolúveis às rações.
A maior exigência dos consumidores por produtos saudáveis e as proibições
legais de uso de corantes sintéticos às rações animais e a alimentos de consumo
humano têm promovido crescente aceitação na incorporação de corantes naturais a
tais alimentos. O urucum (Bixa orellana L) tem sido o mais usado pela indústria
brasileira, representando cerca de 90% dos corantes naturais usados no Brasil e
70% no mundo. Suas preparações têm sido usadas para colorir queijos, pães, óleos,
cereais, entre outros. Do total de sementes de urucum produzidas no Brasil, cerca
de 75% são usadas na fabricação do colorífico, totalmente consumido no mercado
interno e definido pela Resolução 12/78 da Comissão Nacional de Normas e
Padrões para Alimentos do Ministério da Saúde (CNNPA-MS) como um produto
constituído pela mistura de fubá ou farinha de mandioca com urucum em pó ou
extrato oleoso de urucum, adicionado ou não de sal e de óleos comestíveis
(TOCCHINI e MERCADANTE, 2001).
Embora o uso do colorífico seja quase que exclusivamente de uso
doméstico, pois tem emprestado cor avermelhada a alimentos como o arroz, bifes,
frangos, farofas, molhos, entre outros, seu uso em rações de poedeiras pode
constituir-se em alternativa viável para a obtenção de gemas de ovos de codornas
mais pigmentadas, haja vista a elevada proporção do carotenóide lipossolúvel bixina
e a facilidade de obtenção do produto no mercado. Por outro lado, a grande variação
nos teores de bixina encontrados em diversas amostras de colorífico se constitui em
um fator negativo quanto a sua inclusão na alimentação de poedeiras.

49
Pesquisas têm sido realizadas aproveitando-se o potencial de pigmentação
do urucum na pigmentação de gemas de ovos de poedeiras, principalmente quando
se utiliza uma fonte energética isenta de pigmentos, como o sorgo, a quirera de
arroz, entre outras, em substituição ao milho amarelo.
O uso de 500, 1000, 1500 e 2000 ppm de extrato oleoso de urucum
aumentou linearmente a coloração das gemas dos ovos de galinhas poedeiras,
produzidos durante dois ciclos de 28 dias. A mesma cor de gema foi obtida quando
se comparou o resultado obtido com o uso de ração com 63 % de sorgo, adicionada
de 2000 ppm de bixina, com a ração contendo 61 % de milho e sem bixina. A
produção e o peso dos ovos, o consumo de ração e a conversão alimentar não
foram afetados pelos tratamentos (PEREIRA et al., 2000).
Em rações à base de milho contendo 0; 0,05; 0,10; 0,15 e 0,20 % de extrato
oleoso de urucum, houve intensificação da cor das gemas dos ovos de poedeiras à
medida que os níveis de extrato foram aumentados. Entretanto, o extrato oleoso de
bixina não interferiu na produção e qualidade interna dos ovos, no consumo de
ração e na conversão alimentar (PEREIRA et al., 2001).
Poedeiras leves e pesadas que receberam rações contendo 21,45 % de
farelo de soja, 25,45 % de milho, 41,4 % de sorgo e 0,10 % de extrato de urucum
produziram ovos com coloração de gemas similar às aves que receberam a ração
referência (22,85 % de farelo de soja e 63,66 % de milho). Os valores de
pigmentação de gemas, obtidos por meio do leque colorimétrico da Roche, foram de
5,10 e 5,02, para as poedeiras leves, e 5,83 e 5,35 para as poedeiras pesadas
tratadas com ração referência e com ração com sorgo + 0,10 % de extrato de
urucum, respectivamente. A adição de 0,45 % de extrato de urucum à ração
contendo sorgo produziu pigmentação da gema de ovos de poedeiras leves igual a
8,87 pontos no leque da Roche, valor dentro da faixa referencial para aves caipiras,
que gira em torno de 8,5 - 9,0 pontos (SILVA et al., 2000).
De outro modo, outros estudos têm sido realizados a fim de avaliar a
inclusão de fontes de xantofilas diferentes do colorífico, em rações de aves, e dessa
forma avaliar a eficiência de incorporação de pigmentos nas gemas ao longo de um
período. CHEN e BAILEY (1988) observaram que galinhas poedeiras colocaram
ovos cujas gemas alcançaram um platô (pigmentação máxima) entre a terceira e
quarta semana após a inclusão do feno de capim bermuda.

50
Sendo assim, esta pesquisa teve o objetivo de verificar o efeito da inclusão
de diferentes níveis de colorífico, em rações de codornas japonesas em postura,
sobre a pigmentação das gemas dos ovos “in natura”.
MATERIAL E MÉTODOS
Local do experimento O experimento foi realizado no Núcleo de Pesquisa do Laboratório de
Zootecnia e Nutrição Animal (LZNA), situado na Escola Técnica Estadual Agrícola
“Antônio Sarlo”, Campus Experimental da Universidade Estadual do Norte
Fluminense (UENF), localizada no município de Campos dos Goytacazes. Distribuição das codornas no delineamento experimental e tratamentos
Foram utilizadas 240 codornas japonesas (Coturnix japonica) fêmeas, em
delineamento experimental de blocos inteiramente casualizados, com cinco
repetições de seis codornas por unidade experimental. Os tratamentos, constituídos
por oito rações experimentais oriundas da combinação de quatro níveis de colorífico
na ração (0; 1,5; 3,0 e 4,5 %) x duas fontes energéticas (milho e quirera de arroz),
foram casualizados em cada bloco e distribuídos nas parcelas, enquanto os quatro
períodos experimentais constituíram as subparcelas. Cada bloco foi composto por
duas gaiolas, que correspondeu a um nível de altura de gaiolas inseridas em
baterias de madeira, e teve a finalidade de eliminar uma possível influência da
luminosidade nos cinco diferentes andares de gaiolas. Alojamento das codornas nas instalações e manejo
As 240 codornas (80 dias de idade) foram alojadas em galpão de alvenaria
com piso cimentado, telhas de barro, telas de arame da mureta lateral ao teto e sem
cortinas laterais, contendo duas baterias de madeira, as quais agruparam dez
gaiolas metálicas de quatro divisões internas cada. Cada gaiola mediu 1,00 m de
comprimento x 0,34 m de largura x 0,24 m de altura e abrigou 24 codornas, que
receberam água e ração à vontade, em bebedouros e comedouros de chapa
galvanizada, respectivamente, e 24 horas de luz diária, seja natural ou artificial, durante todo o período experimental (80° ao 108° dia de idade).

51
Durante todo o período experimental, a amplitude de temperatura no interior
do galpão, registrada por meio de um termômetro de máxima e mínima, variou de 19,5 a 33 °C. A temperatura matinal, registrada entre 8:00 e 10:30 horas, variou de
21 a 26,5°C.
Antes do início do experimento, o ambiente experimental foi limpo e
desinfetado. Os bebedouros foram limpos diariamente e a limpeza das bandejas
coletoras de fezes foi feita de dois em dois dias.
Rações experimentais
Antes do experimento, por um período de 43 dias, as codornas receberam
ração formulada para conter níveis mínimos ou nulos de pigmentos, visando a
esgotar o estoque de pigmentos contido no organismo das mesmas. Essa ração
apresentou 46,00 % de quirera de arroz; 36,83 % de farelo de soja; 1,23 % de
fosfato bicálcico; 5,34 % de calcário; 0,11 % de DL-metionina; 0,27 % de cloreto de
sódio; 6,99 % de óleo de soja refinado; 2,83 % de inerte e 0,40 % de suplemento
mineral e vitamínico. Os valores nutricionais calculados foram de 2.900 kcal de
energia metabolizável por kg de ração; 21 % de proteína bruta; 0,83 % de metionina
+ cistina; 0,54 % de metionina; 1,15 % de lisina; 0,78 % de treonina; 2,50 % de
cálcio; 0,35 % de fósforo disponível; 0,15 % de sódio e 7,88 % de extrato etéreo.
A partir do 80° dia de idade, as codornas receberam rações experimentais,
que foram formuladas para conterem valores iguais de energia metabolizável,
proteína bruta, cálcio, fósforo disponível, metionina + cistina, metionina e lisina. A
composição percentual das rações experimentais e os respectivos valores
nutricionais calculados encontram-se apresentados no Quadro 1.
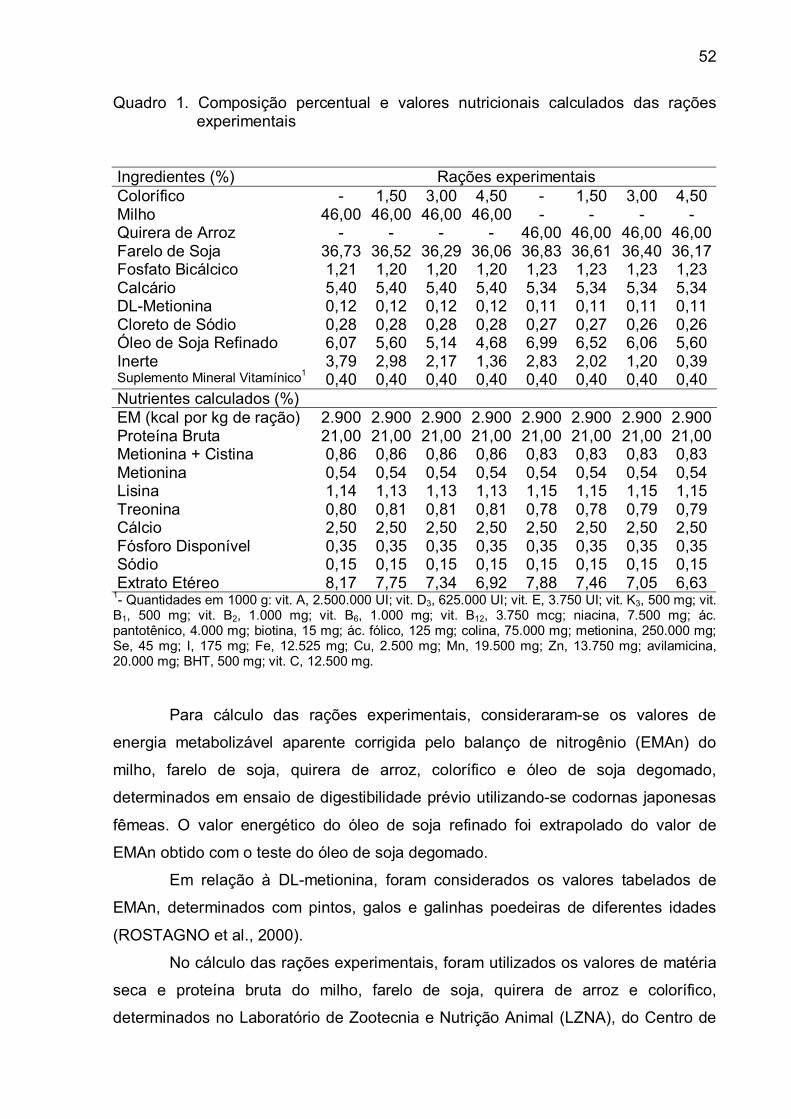
52
Quadro 1. Composição percentual e valores nutricionais calculados das rações experimentais
Ingredientes (%) Rações experimentais Colorífico - 1,50 3,00 4,50 - 1,50 3,00 4,50 Milho 46,00 46,00 46,00 46,00 - - - - Quirera de Arroz - - - - 46,00 46,00 46,00 46,00 Farelo de Soja 36,73 36,52 36,29 36,06 36,83 36,61 36,40 36,17 Fosfato Bicálcico 1,21 1,20 1,20 1,20 1,23 1,23 1,23 1,23 Calcário 5,40 5,40 5,40 5,40 5,34 5,34 5,34 5,34 DL-Metionina 0,12 0,12 0,12 0,12 0,11 0,11 0,11 0,11 Cloreto de Sódio 0,28 0,28 0,28 0,28 0,27 0,27 0,26 0,26 Óleo de Soja Refinado 6,07 5,60 5,14 4,68 6,99 6,52 6,06 5,60 Inerte 3,79 2,98 2,17 1,36 2,83 2,02 1,20 0,39 Suplemento Mineral Vitamínico1 0,40 0,40 0,40 0,40 0,40 0,40 0,40 0,40 Nutrientes calculados (%) EM (kcal por kg de ração) 2.900 2.900 2.900 2.900 2.900 2.900 2.900 2.900 Proteína Bruta 21,00 21,00 21,00 21,00 21,00 21,00 21,00 21,00 Metionina + Cistina 0,86 0,86 0,86 0,86 0,83 0,83 0,83 0,83 Metionina 0,54 0,54 0,54 0,54 0,54 0,54 0,54 0,54 Lisina 1,14 1,13 1,13 1,13 1,15 1,15 1,15 1,15 Treonina 0,80 0,81 0,81 0,81 0,78 0,78 0,79 0,79 Cálcio 2,50 2,50 2,50 2,50 2,50 2,50 2,50 2,50 Fósforo Disponível 0,35 0,35 0,35 0,35 0,35 0,35 0,35 0,35 Sódio 0,15 0,15 0,15 0,15 0,15 0,15 0,15 0,15 Extrato Etéreo 8,17 7,75 7,34 6,92 7,88 7,46 7,05 6,63
1- Quantidades em 1000 g: vit. A, 2.500.000 UI; vit. D3, 625.000 UI; vit. E, 3.750 UI; vit. K3, 500 mg; vit. B1, 500 mg; vit. B2, 1.000 mg; vit. B6, 1.000 mg; vit. B12, 3.750 mcg; niacina, 7.500 mg; ác. pantotênico, 4.000 mg; biotina, 15 mg; ác. fólico, 125 mg; colina, 75.000 mg; metionina, 250.000 mg; Se, 45 mg; I, 175 mg; Fe, 12.525 mg; Cu, 2.500 mg; Mn, 19.500 mg; Zn, 13.750 mg; avilamicina, 20.000 mg; BHT, 500 mg; vit. C, 12.500 mg.
Para cálculo das rações experimentais, consideraram-se os valores de
energia metabolizável aparente corrigida pelo balanço de nitrogênio (EMAn) do
milho, farelo de soja, quirera de arroz, colorífico e óleo de soja degomado,
determinados em ensaio de digestibilidade prévio utilizando-se codornas japonesas
fêmeas. O valor energético do óleo de soja refinado foi extrapolado do valor de
EMAn obtido com o teste do óleo de soja degomado.
Em relação à DL-metionina, foram considerados os valores tabelados de
EMAn, determinados com pintos, galos e galinhas poedeiras de diferentes idades
(ROSTAGNO et al., 2000).
No cálculo das rações experimentais, foram utilizados os valores de matéria
seca e proteína bruta do milho, farelo de soja, quirera de arroz e colorífico,
determinados no Laboratório de Zootecnia e Nutrição Animal (LZNA), do Centro de

53
Ciências e Tecnologias Agropecuárias (CCTA), da Universidade Estadual do Norte
Fluminense (UENF), segundo metodologia proposta por SILVA (1990). Para
cômputo nas rações experimentais, os valores de cálcio e fósforo total dos alimentos
mencionados foram analisados no Laboratório de Solos da Universidade Federal
Rural do Rio de Janeiro, campus Dr. Leonel Miranda, situado na cidade de Campos
dos Goytacazes (Quadro 2).
Quadro 2. Composição bromatológica (%) de alguns ingredientes utilizados nas rações experimentais, expressa na base da matéria natural
Ingredientes Matéria Seca Proteína Bruta Cálcio Fósforo Total Milho 86,77 7,99 0,04 0,32 Farelo de soja 87,77 46,81 0,29 0,63 Quirera de arroz 86,27 7,89 0,07 0,29 Colorífico 88,30 7,00 0,05 0,15
Exceto os nutrientes cujas análises foram mencionadas, todas as rações
foram formuladas de acordo com as composições químicas dos ingredientes
segundo ROSTAGNO et al. (2000) e adequadas às exigências nutricionais para
codornas japonesas segundo o NRC (1994), com exceção dos níveis de proteína
bruta, metionina e lisina. Variáveis avaliadas
Foi avaliada a cor das gemas no 87°; 94°; 101° e 108° dias de idade. O
consumo de ração (g), a conversão alimentar em quantidade de ração consumida
por dúzia e grama de ovos, o percentual de postura e o peso médio dos ovos (g)
foram avaliados no período total (80 a 108 dias de idade). O peso (g) das codornas foi avaliado no 80° e 108° dias de idade.
Coleta de dados
As rações de cada unidade experimental foram pesadas em sacos de
plástico transparente, identificadas e acondicionadas em baldes plásticos com tampa
no 80º dia de idade. O consumo médio de ração por codorna, em cada tratamento e
repetição, foi medido no 108º dia de idade, por diferença entre o total de ração dos
sacos antes e após o consumo pelas codornas, computando-se as sobras de ração
nos comedouros.

54
A avaliação do ganho de peso e do peso das codornas foi obtida pesando-se
as aves dentro de caixa de papelão limpa e de peso conhecido, usando balança
digital, no 80º e 108º dia de idade. A conversão alimentar, expressa em quantidade
de ração consumida por dúzia de ovo produzida, foi calculada pela divisão entre o
consumo de ração e a quantidade de dúzias de ovos produzida em cada tratamento
(média de cinco repetições). Para calcular a conversão alimentar em quantidade de
ração consumida por grama de ovo produzido, dividiram-se os valores médios de
consumo de ração pelos de peso dos ovos, registrados em cada repetição de cada
tratamento.
O percentual de postura foi registrado diariamente, considerando-se o
número de codornas por unidade experimental, porém seu valor médio foi calculado
no período total.
Para cálculo do peso médio dos ovos, dois ovos de cada unidade
experimental, colocados no primeiro, quarto e sétimo dia de cada semana (três
vezes por semana), foram pesados imediatamente após a coleta, que foi feita na
parte da manhã do dia consecutivo à postura. Os valores médios de cada tratamento
foram calculados pela média dos pesos dos ovos das respectivas repetições.
Na manhã dos dias 8, 15, 22 e 29 do experimento, 80 ovos (dez por
tratamento e dois por repetição) produzidos nos dias 7, 14, 21 e 28 foram coletados
aleatoriamente, identificados com caneta para retro-projetor e acondicionados em
embalagens apropriadas para ovos de codorna. Na parte da tarde, após cada coleta,
um ovo por vez foi aberto por meio de quebra da casca na região da câmara de ar e
sua gema foi separada do albúmem e dos resíduos de chalazas e membranas por
retirada manual, sendo colocada em tampa plástica disposta em superfície lisa, de
cor branca e de pequena profundidade. A cor de cada gema de ovo “in natura” foi
visualmente comparada e classificada utilizando-se um leque colorimétrico da Roche
(escore de um a quinze). A tampa plástica foi limpa e seca após cada classificação.
Procedimentos estatísticos
Para análise dos dados de coloração de gemas, utilizou-se o seguinte
modelo estatístico:
Yijkl = m + Bk + Ci + Fj + CFij + eijk + Dl + CDil + FDjl + CFDijl + eijkl.
Em que,

55
Yijkl = Observação individual média no nível i de colorífico da ração, na classe j de
fonte energética da ração, no bloco k e no dia experimental l.
m = Média geral.
Bk = Efeito dos blocos; k = (1, 2, 3, 4 e 5).
Ci = Efeito dos níveis de colorífico da ração; i = (1, 2, 3 e 4).
Fj = Efeito da fonte energética da ração; j = (1 e 2).
CFij = Efeito de interação dos níveis de colorífico com a fonte energética da ração.
eijk = Erro aleatório da parcela associado a cada observação ijk.
Dl = Efeito do dia experimental; l = (1, 2, 3 e 4).
CDil = Efeito de interação dos níveis de colorífico da ração com o dia experimental.
FDjl = Efeito de interação da fonte energética da ração com o dia experimental.
CFDijl = Efeito de interação tripla níveis de colorífico vs fonte energética vs dia
experimental.
eijkl = Erro aleatório da subparcela associado a cada observação ijkl.
Para a análise do percentual de postura, peso médio dos ovos, consumo de
ração, ganho de peso, peso e conversão alimentar, foi utilizado o seguinte modelo
estatístico:
Yijk = m + Bk + Ci + Fj + CFij + eijk.
Em que,
Yijk = Observação individual média no nível i de colorífico da ração, na classe j de
fonte energética da ração e no bloco k.
m = Média geral.
Bk = Efeito dos blocos; k = (1, 2, 3, 4 e 5).
Ci = Efeito dos níveis de colorífico da ração; i = (1, 2, 3 e 4).
Fj = Efeito da fonte energética da ração; j = (1 e 2).
CFij = Efeito de interação dos níveis de colorífico com a fonte energética da ração.
eijk = Erro aleatório associado a cada observação ijk.
Os efeitos dos níveis de colorífico da ração e dos dias experimentais sobre
as diversas variáveis foram verificados pela análise de variância e estimados por
meio de equações polinomiais, respeitando-se as interações significativas,
porventura existentes, entre os fatores incluídos no modelo.
Os efeitos de fonte energética da ração foram comparados por meio do teste
F, ao nível de 5 % de probabilidade.

56
Todas as análises estatísticas foram efetuadas usando-se o programa SAEG
(Sistema de análises estatísticas e genéticas), desenvolvido na Universidade
Federal de Viçosa - UFV (1997).
RESULTADOS E DISCUSSÃO
Coloração de gema (CG) As interações nível de colorífico vs dia experimental e nível de colorífico vs
fonte energética e o efeito isolado de nível de colorífico, fonte energética e dia
experimental foram significativos (P≤0,05).
Observou-se efeito quadrático do nível de colorífico sobre a CG de ovos de
codornas nos dias experimentais 7, 14, 21 e 28, independente da fonte energética
da ração. Nos dias 7, 14, 21 e 28 do experimento, as estimativas de máxima CG
ocorreram com o uso de 3,15; 2,91; 2,97 e 2,77 % de colorífico nas rações,
respectivamente, que equivalem a estimativas de escores máximos de CG iguais a
9,63; 10,19; 10,34 e 11,08 pontos no leque da Roche, respectivamente (Quadro 3).
Estes resultados mostram que o uso de níveis superiores a 3,15 % de
colorífico, em rações contendo 46,00 % de milho ou quirera de arroz, causa redução
na eficiência de incorporação de pigmentos pela saturação de sua capacidade de
armazenamento nas gemas. Essa baixa eficiência pôde ser visualmente observada
e comprovada no decorrer do experimento, visto que as fezes das codornas que
receberam rações com 3,0 e 4,5 % de colorífico se apresentavam com intensa
coloração vermelha, o que significa elevada perda de pigmentos ou baixa taxa de
absorção de bixina.
Quadro 3. Equações de regressão de coloração de gema em função do nível de
colorífico (C), avaliadas em cada dia experimental (D)
Fonte de variação Equações CG máxima R2 C/D7 Ŷ = 3,59 + 3,84 X - 0,61 X2 9,63 1,00 C/D14 Ŷ = 3,73 + 4,43 X - 0,76 X2 10,19 1,00 C/D21 Ŷ = 4,07 + 4,22 X - 0,71 X2 10,34 0,99 C/D28 Ŷ = 4,89 + 4,48 X - 0,81 X2 11,08 1,00
D7 - 7º dia do experimento; D14 - 14º dia do experimento; D21 - 21º dia do experimento; D28 - 28º dia do experimento.

57
Observou-se efeito quadrático do nível de colorífico sobre a CG de ovos de
codornas alimentadas com rações contendo milho e quirera de arroz, independente
do dia experimental (Quadro 4). Os níveis de colorífico que reproduziram estimativas
de máxima coloração de gema foram de 3,15 e 2,80 %, em rações à base de milho e
quirera de arroz, respectivamente. As estimativas de escores máximos de CG foram
de 10,67 pontos, para o uso de colorífico em rações com milho, e 9,91 pontos no
leque da Roche, para o uso de colorífico em rações contendo quirera de arroz.
Estes resultados indicam que não são recomendados os usos de níveis
superiores a 3,15 % de colorífico, em rações à base de milho, e 2,80 % de colorífico,
em rações à base de quirera de arroz, porque ocorre saturação da capacidade de
incorporação e armazenamento de pigmentos nas gemas.
Os usos de 3,0 % de colorífico, em rações à base de quirera de arroz, e de
1,5 % de colorífico, em rações à base de milho, produziram CG estimada igual a
9,87 e 9,29 pontos no leque da Roche (Quadro 4), respectivamente. Estas
estimativas são similares à encontrada por SILVA et al. (2000), que obtiveram o
valor médio de 8,87 pontos em gemas de ovos de galinhas poedeiras leves
alimentadas com ração contendo 25,45 % de milho, 41,40 % de sorgo, 21,45 % de
farelo de soja e com a adição de 0,45 % de extrato de urucum. Esta semelhança
entre valores de CG pode estar associada ao teor de bixina do extrato de urucum e
do colorífico e ao percentual de uso nas rações.
Quadro 4. Equações de regressão de coloração de gema em função do nível de colorífico (C), avaliadas em cada fonte energética (FE)
Fonte de variação Equações CG máxima R2 C/FE1 Ŷ = 5,62 + 3,21 X - 0,51 X2 10,67 1,00 C/FE2 Ŷ = 2,52 + 5,27 X - 0,94 X2 9,91 1,00
FE1 - milho; FE2 – quirera de arroz. Gemas de ovos de codornas alimentadas com rações à base de milho foram
mais pigmentadas (P≤0,05) do que gemas de ovos de codornas alimentadas com
rações à base de quirera de arroz, em todos os níveis de colorífico utilizados nas
rações, independente do dia experimental (Quadro 5 e Figura 1). Isto evidencia a
contribuição dos carotenóides presentes no milho na coloração das gemas.

58
As codornas alimentadas com ração contendo quirera de arroz e colorífico
apresentaram maiores valores médios de coloração de gema do que codornas
alimentadas com rações contendo milho e que receberam níveis de colorífico
imediatamente inferiores aos utilizados em rações com quirera de arroz (Quadro 5).
Esses resultados indicam que este colorífico é um agente pigmentante eficaz em
rações de aves, pois o uso de níveis mais elevados de colorífico, em rações à base
de quirera de arroz, suplantou a contribuição dos carotenóides do milho.
Quadro 5. Médias de coloração de gema de ovos de codornas japonesas
alimentadas com rações contendo milho ou quirera de arroz, referentes a cada nível de colorífico (C)
Fonte de energia Nível de colorífico (%) 0 1,5 3,0 4,5 Milho 5,07* 8,50* 10,55* 11,92* Quirera de arroz 1,50 6,95 9,65 11,35 * Significativo pelo teste F ao nível de 5 % de probabilidade.
Figura 1. Diferença visual entre cor de gemas de ovos de codornas alimentadas com rações à base de milho (M) e quirera de arroz (Q), nos diversos níveis de colorífico (0; 1,5; 3,0 e 4,5 %).

59
Houve efeito linear positivo de dia experimental sobre a coloração de gema
(CG) em todos os níveis de colorífico das rações, independente da fonte energética
utilizada nas rações (Quadro 6). Entre o sétimo e o vigésimo oitavo dia do
experimento, as estimativas médias de coloração de gema variaram de 2,76 a 3,79;
6,93 a 8,53; 9,53 a 10,66 e 11,11 a 12,14 pontos no leque da Roche, para codornas
alimentadas com 0; 1,5; 3,0 e 4,5 % de colorífico na ração, respectivamente.
Estes resultados demonstraram que, à medida que a idade aumentou,
ocorreu baixa incorporação de pigmentos nas gemas. Isto pode ser evidenciado pela
pequena magnitude dos coeficientes angulares das equações de regressão. CHEN
e BAILEY (1988) não observaram este comportamento quando avaliaram, ao longo
de quatro semanas, o efeito da inclusão de 0; 3; 6 e 9 % de feno de capim bermuda
como única fonte de xantofilas em rações para galinhas poedeiras submetidas a um
período prévio de quatorze dias de depleção de xantofilas. A pigmentação das
gemas alcançou um platô, na escala de pontos do leque da Roche, após três
semanas de alimentação, em cada nível de inclusão do feno de capim bermuda.
Quadro 6. Equações de regressão de coloração de gema em função do dia experimental (D), avaliadas em cada nível de colorífico (C)
Fonte de variação Equações R2 D/C0 Ŷ = 2,42 + 0,049 X 0,82 D/C1,5 Ŷ = 6,40 + 0,076 X 0,88 D/C3 Ŷ = 9,15 + 0,054 X 0,60 D/C4,5 Ŷ = 10,77 + 0,049 X 0,99
C0 – 0 % de colorífico; C1,5 – 1,5 % de colorífico; C3 – 3 % de colorífico; C4,5 – 4,5 % de colorífico.
Consumo de ração (CR)
Houve efeito significativo (P≤0,05) para a interação fonte energética vs
níveis de colorífico e para o efeito de fonte energética. O efeito isolado de nível de
colorífico foi não-significativo (P>0,05).
À medida que os níveis de colorífico foram aumentados, observou-se uma
redução do CR de codornas alimentadas com rações à base de milho e um aumento
linear do CR de codornas que receberam quirera de arroz nas rações (Quadro 7). Os
resultados indicam que, em rações à base de milho, as quais já contêm pigmentos, o
uso crescente de colorífico prejudicou o CR das codornas. Contudo, em rações à
base de quirera de arroz, observa-se que a adição de colorífico favoreceu o CR das

60
codornas. Esses resultados são discordantes dos apresentados por PEREIRA et al.
(2000), SILVA et al. (2000) e PEREIRA et al. (2001), que não encontraram efeito da
inclusão de diferentes níveis de extrato oleoso de urucum (EU) sobre o consumo de
ração em poedeiras. As diferenças entre os resultados encontrados neste trabalho e
os observados na literatura talvez possam ser explicadas pela maior concentração
de bixina presente nas rações com colorífico, em relação às rações contendo EU.
Quadro 7. Equações de regressão do consumo de ração em função dos níveis de
colorífico (C), avaliadas em cada fonte energética (FE)
Fonte de variação Equações R2 C/FE1 Ŷ = 655,40 - 9,31 X 0,75 C/FE2 Ŷ = 507,25 + 12,12 X 0,76
FE1 - milho; FE2 – quirera de arroz.
O consumo de ração (CR) de codornas japonesas alimentadas com rações à
base de milho foi maior (P≤0,05) do que o CR de codornas que receberam quirera
de arroz, em todos os níveis de colorífico utilizados nas rações (Quadro 8).
Quadro 8. Médias do consumo de ração (g/ave) de codornas japonesas, referentes a
cada combinação de fonte energética (FE) vs níveis de colorífico (C)
Níveis de colorífico (%) FE 0,0 1,5 3,0 4,5
Milho 654,83* 649,61* 612,85* 620,54* Quirera 518,99 513,21 532,84 573,06
* Significativo pelo teste F ao nível de 5 % de probabilidade. É muito provável que os menores CR de codornas que receberam quirera de
arroz sejam atribuídos a algum fator presente na quirera de arroz que prejudicou o
CR, haja vista que foi observado um percentual maior (não registrado) de
mortalidade, durante os períodos de adaptação e experimental, em codornas
alimentadas com quirera de arroz. Esses resultados são divergentes dos
encontrados por SILVA et al. (2000), que não verificaram diferenças entre os valores
médios de CR de poedeiras alimentadas com ração referência à base de milho e
com rações à base de sorgo, suplementadas ou não com níveis crescentes de
extrato oleoso de urucum.

61
Embora explicado pelas equações de regressão (Quadro 7), vale mencionar
que a diferença entre valores de consumo de ração (CR) das codornas alimentadas
com milho e quirera de arroz foi reduzida com o uso de 3,0 e, principalmente, 4,5 %
de colorífico em rações à base de quirera de arroz (Quadro 8). Com base na
melhora do CR, pode-se afirmar que os pigmentos oriundos do colorífico interferiram
no metabolismo e, conseqüentemente, no desempenho das codornas que
receberam quirera de arroz.
Peso médio dos ovos (PMO)
Houve efeito significativo (P≤0,05) para a interação fonte energética vs nível
de colorífico. Os efeitos isolados de fonte energética e nível de colorífico foram não-
significativos (P>0,05).
Observou-se efeito linear negativo dos níveis de colorífico sobre o PMO de
codornas japonesas alimentadas com rações à base de milho. Entretanto, codornas
alimentadas com rações à base de quirera de arroz apresentaram PMO mais
elevados à medida que os níveis de inclusão do coloríficos foram aumentados nas
rações, embora o coeficiente de determinação encontrado (0,45) mostre que houve
grande dispersão dos dados, o que dificulta sua avaliação (Quadro 9).
Esses resultados são discordantes dos apresentados por SILVA et al.
(2000), que não encontraram efeito na adição de 0; 0,1; 0,2; 0,3; 0,4; 0,5 e 0,6 % de
extrato oleoso de urucum (EU) sobre o PMO de galinhas poedeiras no segundo ciclo
de produção, e por PEREIRA et al. (2000), que não observaram efeito na adição de
500; 1000; 1500 e 2000 ppm de EU sobre o PMO de poedeiras durante dois ciclo de
28 dias. Essas divergências possivelmente podem ser atribuídas à maior
concentração de bixina nas rações com colorífico, em relação às rações com EU.
Quadro 9. Equações de regressão de peso médio dos ovos em função do nível de
colorífico (C), avaliadas em cada fonte energética (FE)
Fonte de variação Equações R2 C/FE1 Ŷ = 10,67 – 0,095 X 0,70 C/FE2 Ŷ = 10,30 + 0,086 X 0,45
FE1 - milho; FE2 – quirera de arroz.

62
O peso médio dos ovos (PMO) de codornas alimentadas com rações à base
de milho foi maior (P≤0,05) do que o PMO de codornas que receberam quirera de
arroz, quando ambas as rações apresentaram 0 % de colorífico. Todavia, com o uso
de 3,0 % de colorífico nas rações, as codornas que receberam ração com quirera de
arroz apresentaram maior PMO (P≤0,05) do que aquelas que receberam rações à
base de milho. Quando se utilizaram 1,5 e 4,5 % de colorífico nas rações, os valores
médios de PMO de codornas alimentadas com rações à base de milho e quirera de
arroz não diferiram estatisticamente (P>0,05) (Quadro 10).
Quadro 10. Médias do peso médio (g) de ovos de codornas japonesas, referentes a
cada combinação de fonte energética (FE) vs nível de colorífico (C)
Níveis de colorífico (%) Fonte de energia 0,0 1,5 3,0 4,5 Milho 10,78* 10,37 NS 10,35* 10,31 NS
Quirera 10,37 10,22 10,78 10,61 * Significativo pelo teste F ao nível de 5 % de probabilidade; NS - Não significativo (P>0,05). Percentagem de postura (PP), Conversão alimentar por dúzia de ovos (CAD) e Conversão alimentar por grama de ovos (CAG)
Em relação ao PP, à CAD e à CAG, somente houve efeito significativo
(P≤0,05) para o fator fonte energética. A interação fonte energética vs nível de
colorífico e o efeito isolado de nível de colorífico foram não-significativos (P>0,05).
Observou-se que codornas japonesas alimentadas com rações à base de
milho apresentaram maior PP e melhores CAD e CAG do que codornas que
receberam rações à base de quirera de arroz, independente do nível de colorífico
utilizado nas rações (Quadro 11).
Esses resultados são discordantes dos apresentados por SILVA et al.
(2000), que observaram que não houve diferença entre a PP e a conversão
alimentar de poedeiras alimentadas com rações à base de milho e com rações à
base de sorgo, suplementada ou não por níveis crescentes de extrato oleoso de
urucum.

63
Quadro 11. Médias do percentual de postura (PP) e da conversão alimentar por dúzia (CAD) e por grama (CAG) de ovos de codornas japonesas, referentes a cada fonte energética (FE)
FE PP CAD (g de ração por dúzia) CAG (g de ração por g de ovos) Milho 77,69* 352,17* 2,81*
Quirera 59,26 392,08 3,11 * Significativo pelo teste F ao nível de 5 % de probabilidade.
CONCLUSÕES
1. A inclusão de níveis crescentes de colorífico nas rações à base de milho e
quirera de arroz aumentou de forma quadrática a pigmentação das gemas de
ovos “in natura” de codornas japonesas.
2. O uso de 1,5 % de colorífico nas rações com 46 % de milho ou quirera de arroz
promoveu intensa pigmentação das gemas de ovos de codornas japonesas,
tornando-os mais atrativos para o consumidor.
REFERÊNCIAS BIBLIOGRÁFICAS CHEN, B.H., BAILEY, C.A. (1988) Effect on turf bermudagrass meal on egg
production, feed utilization, yolk color, and egg weight. Poultry Science, 67:1154-
1156.
LEVY, L.W., REGALADO, E., NAVARRETE, S., WATKINS, R.H. (1997) Bixin and
norbixin in human plasma: determination and study of the absorption of a single
dose of annatto food color. Analyst, 122:977-980.
NATIONAL RESEARCH COUNCIL – NRC (1994) Nutrient Requirements of Poultry.
9.ed. Washington: D.C. National Academy Press, 155p.
PEREIRA, A.V., KISHIBE, R., ARIKI, J., BORGES, S.A., LODDI, M.M. (2000) Bixina
e norbixina como agentes pigmentantes da gema de ovos de poedeiras
comerciais. In: Anais dos trabalhos da XXXVII Reunião Anual da Sociedade
Brasileira de Zootecnia. Viçosa - MG: SBZ, p.259.

64
PEREIRA, A.V., ARIKI, J., LODDI, M.M., KISHIBE, R. (2001) Bixina como agente
pigmentante das gemas de ovos de poedeiras comerciais. In: Anais dos trabalhos
da XXXVIII Reunião Anual da Sociedade Brasileira de Zootecnia. Piracicaba - SP:
SBZ, p.839-840.
ROSTAGNO, H.S., ALBINO, L.F.T., DONZELE, J.L., GOMES, P.C., FERREIRA,
A.S., OLIVEIRA, R.F., LOPES, D.C. (2000) Tabelas Brasileiras para aves e
suínos - Composição de alimentos e exigências nutricionais. Viçosa - MG: UFV,
Departamento de Zootecnia, 141p.
SILVA, D.J. (1990) Análise de alimentos (Métodos químicos e biológicos). 2 ed.
Viçosa - MG: UFV, Imprensa Universitária, 165 p.
SILVA, J.H.V., ALBINO, L.F.T. GODOI, M.J.S. (2000) Efeito do extrato de urucum na
pigmentação da gema dos ovos. Revista Brasileira de Zootecnia, 29 (5):1435-
1439.
TOCCHINI, L., MERCADANTE, A.Z. (2001) Extração e determinação, por CLAE, de
bixina e norbixina em coloríficos. Ciência e Tecnologia de Alimentos, 21 (3):310-
313.
UNIVERSIDADE FEDERAL DE VIÇOSA (1997) SAEG - Sistema de análises
estatísticas e genéticas: Versão 7.1. Viçosa - MG: UFV, 150p.

65
COLORÍFICO E NIACINA SUPLEMENTAR EM RAÇÕES DE CODORNAS JAPONESAS MACHOS: TRIGLICERÍDEOS SANGÜÍNEOS E COMPOSIÇÃO
QUÍMICA DA CARNE E CARCAÇA
RESUMO
O objetivo deste trabalho foi avaliar o efeito da inclusão de colorífico e de
niacina suplementar (NS) em rações nos níveis plasmáticos de triglicerídeos (T) e
lipoproteínas de densidade muito baixa (VLDL) e na gordura da carne de peito, coxa
e sobrecoxa e da carcaça de codornas japonesas, sob condições normais de
alimentação. Foram utilizadas 240 codornas japonesas (Coturnix japonica) machos,
em delineamento experimental inteiramente casualizado, com quatro tratamentos,
cinco repetições e doze codornas por unidade experimental (boxe). Os tratamentos
consistiram de uma ração referência à base de milho e farelo de soja, sem inclusão
de colorífico e de NS, e de outras três rações, uma com 4,5 % de colorífico, outra
com 0,08 % de NS e outra com 4,5 % de colorífico e 0,08 % de NS. Não houve efeito
de tratamento no desempenho, nos níveis de T e VLDL no sangue e nos teores de
matéria seca, extrato etéreo e proteína bruta (PB) na carcaça com osso e sem
cabeça, patas, pele, penas, vísceras, peito, coxas e sobrecoxas de codornas
japonesas machos. No 49° dia de idade, as codornas que receberam rações com
0,08 % de NS tiveram 1,50 % de gordura na carne de peito, sendo esse nível
superior (P≤0,05) a 0,85 %, apresentado por codornas alimentadas com a ração
referência. No 35° dia de idade, as codornas alimentadas com ração + colorífico +
NS apresentaram 17,8 % de PB na carne de coxa e sobrecoxa, sendo esse valor
maior (P≤0,05) que 17,2 %, apresentado por codornas alimentadas com a ração
referência. Os resultados mostraram que o colorífico e a NS não reduziram os níveis
de gordura no sangue, carne e carcaça de codornas japonesas machos.
Palavras-chave: colorífico, codorna japonesa, gordura, niacina suplementar,
triglicerídeos plasmáticos

66
ABSTRACT
The objective of this research was to evaluate the effect of addition of spice
and supplementary niacin (SN) to the rations upon triglyceride (T) and very low-
density lipoprotein (VLDL) levels in plasma and fat in breast, drumstick and thigh
meat and carcass of japanese quails, under normal condition of feeding. Two
hundred and forty japanese male quails (Coturnix japonica) were used in a
completely randomized design, with four treatments, five replicates and twelve quails
per experimental unit (box). The treatments consisted of one reference ration with
corn and soybean meal, without spice and SN, and three other rations, one with 4,5
% of spice, an other with 0,08 % of SN and an other one with 4,5 % of spice and 0,08
% of SN. The treatments had no effect upon performance, T and VLDL levels in
plasma, as well as on dry matter, ether extract and crude protein (CP) contents in the
carcass with bone and without head, paws, skin, feathers, entrails, breast, drumsticks
and thighs of the japanese male quails. On the 49th day of age, the quails that
received rations with 0,08 % of SN had 1,50 % of fat in the breast meat, higher
(P≤0,05) than 0,85 % presented by japanese quails fed with reference ration. On the
35th day of age, the quails fed on rations with spice and SN had 17,8 % of CP in the
drumstick and thigh meat, higher (P≤0,05) than 17,2 % presented by quails fed with
the reference ration. The results indicated that spice and SN did not reduce the fat
levels in plasma, meat and carcass of japanese male quails. Key words: fat, japanese quail, plasma triglycerides, spice, supplementary niacin
INTRODUÇÃO
Um grama de gordura contribui com cerca de 9,4 kcal de energia bruta (EB),
representando cerca de 2,25 vezes mais calorias do que um grama de carboidrato,
que produz em torno de 4,2 kcal de EB (BERTECHINI, 1991). Isso significa que a
produção de aves com carcaças “magras” ou com pouco acúmulo de gordura pode
contribuir para reduzir a ingestão de calorias nas dietas humanas e obter maior
controle dos níveis de triglicerídeos sangüíneos de humanos, refletindo
possivelmente em redução da obesidade e melhor qualidade de vida dos indivíduos.
Como exemplo, em cortes sem pele e osso, a carne de peito de frangos de
corte tem apresentado de 1,6 a 2,4 % de gordura, sendo os valores para frangos

67
machos e fêmeas bem semelhantes (KESSLER e JÚNIOR, 2001). De outro modo,
codornas japonesas machos apresentaram 4,0 % de extrato etéreo (EE) na carne de
peito no 35° dia e 7,1 % no 49° dia de idade (YALÇIN et al., 1995), e 5,2 % de EE na
carcaça (matéria natural) no 35° dia e 9,9 % no 49° dia de idade (JÚNIOR, 1981).
De forma geral, as concentrações elevadas de triglicerídeos sangüíneos
(TS) têm sido consideradas como um fator de risco sinérgico para as doenças
coronarianas arteriais (DCA). A avaliação da hipertrigliceridemia no risco de DCA é
feita por meio de mensurações dos níveis de TS em jejum, em conjunto com os
níveis de lipoproteínas de baixa densidade (LDL) e lipoproteínas de alta densidade
(HDL), cujas correlações são elevadas e positivas com DCA (↑TS, ↑LDL e ↓ HDL →
DCA) (GOTTO JR., 1998).
O sangue é o veículo pelo qual os triglicerídeos são transportados e
posteriormente depositados nos produtos. Baixas concentrações plasmáticas de
lipoproteínas de densidade muito baixa (VLDL) em frangos de corte e perus
alimentados à vontade têm sido bem correlacionadas (r = 0,6 – 0,7) com baixo
conteúdo de gordura corporal (GRIFFIN et al., 1999).
Segundo KESSLER e JÚNIOR (2001), aves mais magras têm apresentado
uma menor concentração plasmática de VLDL e uma menor atividade da lipase
lipoprotéica, enzima que catalisa a hidrólise de triglicerídeos do núcleo das VLDL e
dos portamícrons a ácidos graxos e glicerol. No tecido adiposo, os ácidos graxos são
reesterificados e armazenados como triglicerídeos.
O acúmulo de gordura em aves depende do nível de síntese e secreção de
lipoproteínas pelo fígado, já que o armazenamento de triglicerídeos nos adipócitos
depende da disponibilidade de lipídeos do plasma, oriundos da ração ou da
lipogênese no fígado. Como o nível de lipídeos nas rações de aves comerciais não
tem ultrapassado o limite de 10 %, a lipogênese tem sido dependente, em sua maior
parte, da disponibilidade de carboidratos nas rações, que darão origem a moléculas
de acetil-CoA, pois a síntese de ácidos graxos é muito limitada no tecido adiposo
(HERMIER, 1997).
Por outro lado, em estudo sobre toxicidade, PAUMGARTTEN et al. (2002)
não constataram redução no peso corporal em ratas com 21 dias de gestação e nas
suas proles, com o uso da dose de 500 mg de extrato seco de urucum (28 % de
bixina) por kg de peso corporal (PC) por dia, dosagem que equivale a um consumo
médio de 140 mg de bixina por kg de PC por dia.

68
Considerando a baixa toxicidade, faz-se necessário investigar a contribuição
da atividade hipolipidêmica da bixina e niacina suplementar (NS) sobre o
metabolismo lipídico de animais normolipidêmicos. A bixina e a NS têm sido mais
ativas para reduzir a lipidemia quando é provocada uma hiperlipidemia, normalmente
com a inclusão, nas rações, de colesterol, ácido cólico ou gordura animal, fato que
foge da realidade sobre a alimentação das aves comerciais.
O efeito hipolipidêmico da bixina, presente no colorífico, na prevenção e
controle da hiperlipidemia tem sido atribuído à sua ação ativadora sobre a lipase
lipoprotéica. A formação de quelatos entre a enzima e a bixina induz a mudança
conformacional da enzima, modificando seu centro ativo (LIMA et al., 1999).
A atividade hipolipidêmica do ácido nicotínico decorre da capacidade de
inibir a lipólise do tecido adiposo, de reduzir a esterificação dos triglicerídeos no
fígado e de aumentar a atividade da lipase lipoprotéica (GOODMANS et al., 1996).
Com hiperlipidemia induzida por 1 % de colesterol (COL) e 0,1 % de ácido
cólico (AC) na ração (R) diária, durante 28 dias, coelhos que receberam R + COL +
AC e a dose oral (em cápsula) diária de 0,01 mol de bixina por Kg de peso corporal
apresentaram 228,79 mg de triglicerídeo por dl de soro, valor superior (P≤0,05) a
113,78 mg/dl, obtido por coelhos que receberam somente ração (LIMA et al., 2001).
Por sua vez, SOUZA et al. (2000) verificaram que após três meses e em
relação ao grupo que recebeu somente ração, fêmeas de ratos alimentadas com
ração diária contendo 70, 350 e 700 mg de corante (28 % de bixina) tiveram uma
redução de 9,30; 15,24 e 10,85 % dos níveis de triglicerídeos séricos (TS),
respectivamente. Machos alimentados com 350 e 700 mg de corante tiveram os
níveis de TS reduzidos em 37,36 e 43,82 %, respectivamente. Todas as doses de
bixina causaram efeito hipolipidêmico em ratos sem hiperlipidemia induzida.
Imediatamente após a indução da hiperlipidemia com Triton (T) e após 20
horas, o ácido nicotínico (AN) foi administrado na dose de 5 mg por Kg de peso
corporal, por via intraperitoneal, em ratos que receberam ração comercial (R) à
vontade. Após 43 horas da aplicação do Triton, os ratos tratados com R + T + AN
apresentaram 104,56 mg de triglicerídeo por dl de soro, valor inferior (P≤0,05) ao
apresentado por ratos tratados com R + T, que foi de 311,36 mg/dl. O tratamento
somente com ração (R) proporcionou o nível de 162,30 mg/dl, sendo numericamente
superior ao tratamento com R + triton + ácido nicotínico (SANTOS et al., 1999).

69
Nesse contexto, torna-se relevante verificar se a inclusão de colorífico e de
niacina suplementar em rações reduz os níveis de triglicerídeos e lipoproteínas de
densidade muito baixa no sangue e a gordura na carne de codornas japonesas, sob
condições normais de alimentação.
MATERIAL E MÉTODOS
Local do experimento O experimento foi realizado no Núcleo de Pesquisa do Laboratório de
Zootecnia e Nutrição Animal (LZNA), situado no Colégio Agrícola Antônio Sarlo,
Campus Experimental da Universidade Estadual do Norte Fluminense (UENF),
localizada no município de Campos dos Goytacazes. Distribuição das codornas no delineamento experimental e tratamentos
Foram utilizadas 240 codornas japonesas (Coturnix japonica) machos, em
delineamento experimental inteiramente casualizado, com quatro tratamentos, cinco
repetições e doze codornas por unidade experimental (boxe). Os tratamentos
consistiram de uma ração referência, que foi constituída à base de milho e farelo de
soja, sem inclusão de colorífico e de niacina suplementar (NS), e de outras três
rações, uma com 4,5 % de colorífico, outra com 0,08 % de NS e outra com 4,5 % de
colorífico e 0,08 % de NS.
Alojamento das codornas nas instalações e manejo
As codornas com idade inicial de vinte e um dias foram alojadas em galpão
de alvenaria contendo vinte boxes feitos de madeira e tela. Cada boxe mediu 2,05 m
de comprimento x 1,75 m de largura x 1,77 m de altura e abrigou doze codornas.
Antes de serem alojadas em cada unidade experimental, procedeu-se ao
corte, por meio de tesoura, das penas primárias da asa direita de cada codorna,
visando a evitar vôos que pudessem resultar em fuga das aves. Após isso, as
codornas foram abrigadas com pesos iniciais semelhantes e receberam água e
ração à vontade, em bebedouros tipo copo de pressão e comedouros do tipo
bandeja contendo folhas de jornal embaixo, respectivamente, além de receberem
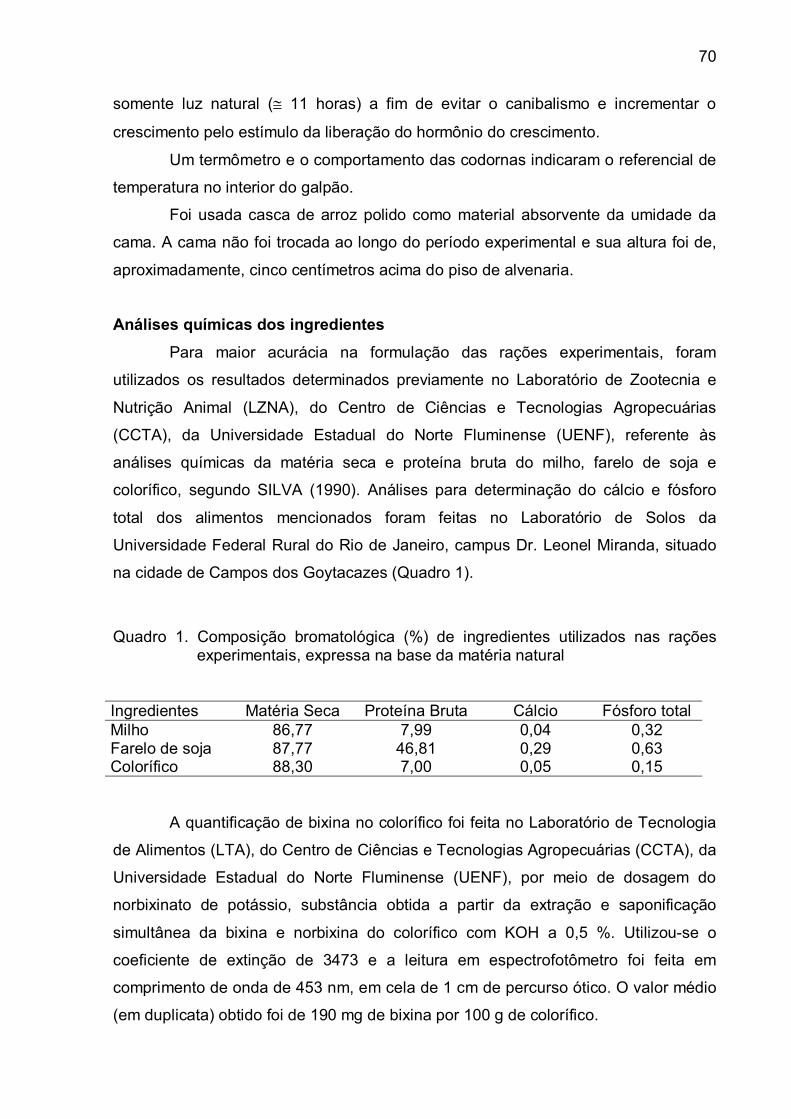
70
somente luz natural (≅ 11 horas) a fim de evitar o canibalismo e incrementar o
crescimento pelo estímulo da liberação do hormônio do crescimento.
Um termômetro e o comportamento das codornas indicaram o referencial de
temperatura no interior do galpão.
Foi usada casca de arroz polido como material absorvente da umidade da
cama. A cama não foi trocada ao longo do período experimental e sua altura foi de,
aproximadamente, cinco centímetros acima do piso de alvenaria.
Análises químicas dos ingredientes Para maior acurácia na formulação das rações experimentais, foram
utilizados os resultados determinados previamente no Laboratório de Zootecnia e
Nutrição Animal (LZNA), do Centro de Ciências e Tecnologias Agropecuárias
(CCTA), da Universidade Estadual do Norte Fluminense (UENF), referente às
análises químicas da matéria seca e proteína bruta do milho, farelo de soja e
colorífico, segundo SILVA (1990). Análises para determinação do cálcio e fósforo
total dos alimentos mencionados foram feitas no Laboratório de Solos da
Universidade Federal Rural do Rio de Janeiro, campus Dr. Leonel Miranda, situado
na cidade de Campos dos Goytacazes (Quadro 1).
Quadro 1. Composição bromatológica (%) de ingredientes utilizados nas rações
experimentais, expressa na base da matéria natural
Ingredientes Matéria Seca Proteína Bruta Cálcio Fósforo total Milho 86,77 7,99 0,04 0,32 Farelo de soja 87,77 46,81 0,29 0,63 Colorífico 88,30 7,00 0,05 0,15
A quantificação de bixina no colorífico foi feita no Laboratório de Tecnologia
de Alimentos (LTA), do Centro de Ciências e Tecnologias Agropecuárias (CCTA), da
Universidade Estadual do Norte Fluminense (UENF), por meio de dosagem do
norbixinato de potássio, substância obtida a partir da extração e saponificação
simultânea da bixina e norbixina do colorífico com KOH a 0,5 %. Utilizou-se o
coeficiente de extinção de 3473 e a leitura em espectrofotômetro foi feita em
comprimento de onda de 453 nm, em cela de 1 cm de percurso ótico. O valor médio
(em duplicata) obtido foi de 190 mg de bixina por 100 g de colorífico.

71
Rações experimentais Na fase inicial de vida (1 a 21 dias), as codornas foram alimentadas com
ração comercial contendo um nível mínimo de 28,00 % de proteína bruta e 0,70 %
de fósforo, um nível máximo de 1,50 % de cálcio, e suplementada com 48 mg de
niacina por kg de ração. A partir do 21° dia de idade, as codornas receberam as
rações experimentais, cuja composição percentual e os valores nutricionais
calculados encontram-se apresentados no Quadro 2.
Quadro 2. Composição percentual e valores nutricionais calculados das rações
experimentais
Ingredientes (%) Rações Experimentais Milho 62,63 57,89 62,47 57,73 Farelo de Soja 33,96 34,10 33,99 34,13 DL-Metionina 0,01 0,01 0,01 0,01 Fosfato Bicálcico 0,87 0,88 0,87 0,88 Calcário 1,19 1,18 1,19 1,18 Cloreto de Sódio 0,27 0,27 0,27 0,27 Inerte 0,16 0,16 0,16 0,16 Óleo de Soja Refinado 0,51 0,61 0,56 0,66 Suplemento Mineral Vitamínico1 0,40 0,40 0,40 0,40 Colorífico ---- 4,50 ---- 4,50 Nicotinamida em Pó2 ---- ---- 0,08 0,08 Nutrientes Calculados (%) Proteína Bruta 21,00 21,00 21,00 21,00 EMAn3 2.900 2.900 2.900 2.900 Extrato Etéreo 3,14 3,23 3,19 3,28 Metionina + Cistina 0,84 0,84 0,84 0,84 Metionina 0,50 0,50 0,50 0,50 Lisina 1,10 1,10 1,10 1,10 Treonina 0,81 0,81 0,81 0,81 Cálcio 0,80 0,80 0,80 0,80 Fósforo Disponível 0,30 0,30 0,30 0,30 Sódio 0,15 0,15 0,15 0,15 1- Vaccinar postura/reprodução inicial completo - quantidades em 1000 g: vit. A, 3.750.000 UI; vit. D3, 750.000 UI; vit. E, 7.500 mg; vit. K3, 1.000 mg; vit. B1, 750 mg; vit. B2, 1.500 mg; vit. B6, 1.500 mg; vit. B12, 7.500 mcg; niacina, 10.000 mg; ác. pantotênico, 3.750 mg; biotina, 30 mg; ác. fólico, 375 mg; colina, 10.000 mg; metionina, 400.000 mg; Se, 45 mg; I, 175 mg; Fe, 12.525 mg; Cu, 2.500 mg; Mn, 19.500 mg; Zn, 13.750 mg; avilamicina, 15.000 mg; nicarbazina, 12.500 mg; narasin, 10.000 mg; BHT, 500 mg; vit. C, 12.500 mg. 2- Nicotinamida com teor de 100 % de pureza - Pharma Nostra Comercial Ltda. 3- Energia metabolizável aparente corrigida por retenção de nitrogênio.
Para cálculo das rações experimentais, consideraram-se os valores médios
de energia metabolizável aparente corrigida pelo balanço de nitrogênio (EMAn) do

72
milho, farelo de soja, colorífico e óleo de soja degomado, determinados em ensaio
de digestibilidade com codornas japonesas fêmeas. O valor energético do óleo de
soja refinado foi extrapolado do valor de EMAn obtido com o teste do óleo de soja
degomado.
Considerando-se um consumo diário de ração de 25 gramas por codorna por
dia, um peso médio por codorna de 100 g e o teor médio de bixina do colorífico,
determinado em laboratório, igual a 190 mg por 100 g de colorífico, o uso de 4,5 %
de colorífico nas rações (45 g por kg de ração) proporcionou um consumo médio
diário de, aproximadamente, 2,14 mg de bixina por codorna por dia ou 21,4 mg de
bixina por kg de peso corporal. Este nível é inferior à dosagem que tem sido utilizada
em coelhos, que é de 0,01 mol ou 3,94 g de bixina por kg de peso corporal, mas é
suficiente para promover intensa coloração vermelha nas fezes, indicativo de que a
quantidade de bixina presente na ração supre a quantidade de bixina absorvida
pelas codornas.
A inclusão de 0,08 % de nicotinamida em pó nas rações (800 mg por kg de
ração) corresponde a uma tolerância vinte vezes superior à exigência nutricional de
niacina para codornas japonesas recomendada pelo NRC (1994), que é de 40 mg
por kg de ração. Considerando-se um consumo médio de ração de 25 gramas por
codorna por dia e um peso médio por codorna de 100 g, o uso dessa dosagem
possibilitou um consumo médio de 20 mg de niacina por codorna por dia, ou 200 mg
de niacina por kg de peso corporal, nível 40 vezes superior à dosagem utilizada em
ensaios com ratos (5 mg por kg de peso corporal).
Em relação à DL-metionina, foram considerados os valores tabelados de
EMAn, determinados com pintos, galos e galinhas poedeiras de diferentes idades
(ROSTAGNO et al., 2000).
Exceto os nutrientes que foram analisados em laboratório, todas as rações
experimentais (isoenergéticas, isoprotéicas, isocálcicas e isofosfóricas) foram
formuladas de acordo com as composições químicas dos ingredientes apresentados
por ROSTAGNO et al. (2000) e adequadas às exigências nutricionais para codornas
japonesas do NRC (1994), à exceção dos níveis de proteína bruta.
Variáveis avaliadas
Nos períodos de vinte e um a trinta e cinco, e trinta e cinco a quarenta e
nove dias de idade, foram avaliados ganho de peso, consumo de ração e conversão

73
alimentar. No 35° e 49° dias de idade, foram avaliados peso, triglicerídeos e
lipoproteínas de densidade muito baixa no plasma sangüíneo, e matéria seca,
extrato etéreo e proteína bruta na carne de peito, na carne de coxa + sobrecoxa e na
carcaça. Coleta de dados
As rações de cada unidade experimental foram pesadas, identificadas e
acondicionadas em baldes plásticos com tampa no 21º dia de idade. O consumo
médio por codorna, em cada tratamento e repetição, foi medido no 35° e 49º dia de
idade, por diferença entre o total de ração dos baldes antes e após o consumo pelas
codornas, computando-se as sobras de ração nos comedouros.
Para avaliação do ganho de peso, conversão alimentar (grama de ração
consumida por grama de ganho de peso) e peso, as codornas de cada unidade
experimental foram colocadas em caixa de papelão limpa e pesadas, em balança
digital, no 21º, 35° e 49º dias de idade.
Na noite que antecedeu o 35° e 49º dia de idade, foi iniciado um jejum
alimentar, com duração de, aproximadamente, doze horas, visando a preparar as
codornas para a coleta de amostras de sangue e, em conseqüência, evitar a
contaminação da carne pelo conteúdo intestinal. Para isso, duas codornas de cada
repetição, num total de quarenta, foram retiradas aleatoriamente dos boxes e
transferidas para cinco gaiolas metálicas de quatro divisões internas cada, com
dimensão de 1,0 m de comprimento, 0,34 m de largura e 0,24 m de altura,
localizadas em outro galpão e próximas do local do abate. Cada compartimento de
gaiola foi identificado e representou um boxe distinto. Na manhã do 35° e 49º dias de
idade (término do experimento), foram coletados de dois a três mililitros de sangue
por codorna, por meio de degola completa com faca de lâmina reta, entre os ossos
occipital e atlas, das quarenta codornas, sendo duas por repetição.
As amostras de sangue foram drenadas individualmente para tubos
esterilizados de vidro, os quais foram colocados verticalmente em suportes de ferro,
acondicionados dentro de isopor e levados imediatamente para o Laboratório Plínio
Bacelar, localizado em Campos dos Goytacazes – RJ. A seguir, os tubos foram
centrifugados em centrífuga Fanem a 3.000 rotações por minuto, por cinco minutos,
os plasmas foram retirados com pipeta, reunidos por repetição e utilizados para
determinar os níveis de triglicerídeos e lipoproteínas de densidade muito baixa

74
(VLDL). Os triglicerídeos foram quantificados em aparelho Vitros, utilizando-se
metodologia denominada química seca. A VLDL foi estimada utilizando-se a
equação de Friedewald et al. (1972), citada por HAMMAD et al. (1998). Cada
amostra foi constituída pela reunião de dois plasmas, provenientes de duas
codornas por repetição, totalizando vinte amostras analisadas.
Após a coleta do sangue, foi procedida a sangria individual, por
aproximadamente dois minutos. A seguir, utilizando-se uma faca de lâmina reta, as
patas (canela e dedos) foram descartadas por meio de corte na articulação tíbio-
társica. A pele e, conseqüentemente, as penas foram retiradas completamente por
meio de corte superficial realizado no dorso das codornas e de retirada do braço e
dedos (tulipas), realizada através de corte na articulação úmero-cubital. A
evisceração manual foi feita por meio de corte transversal da cloaca e corte ventral
entre o tórax (ponta do esterno) e o abdômen, tornando possível proceder ao toalete
completo das carcaças, as quais foram, ao mesmo tempo, lavadas em água
corrente.
Terminado esse processo, as carcaças foram reunidas por repetição,
colocadas em sacos plásticos, identificadas, mergulhadas em isopor contendo gelo
picado por vinte a trinta minutos, retiradas dos sacos plásticos e deixadas gotejar por
dez minutos, à temperatura ambiente, após terem sido colocadas em redes plásticas
e penduradas em ganchos metálicos. Após o gotejamento, foram reembaladas em
novos sacos plásticos, reidentificadas, colocadas em saco plástico de maior
tamanho, embrulhadas em papel alumínio para evitar a oxidação e, finalmente,
congeladas em freezer para análises futuras.
Com antecedência de aproximadamente 48 horas à manipulação e
processamento da carne, as carcaças foram retiradas do freezer e deixadas degelar
lentamente em geladeira a aproximadamente 4°C. Após o degelo completo, as duas
carcaças completas de cada repetição foram pesadas e as coxas + sobrecoxas e o
peito de cada codorna foram separados nas articulações, com o auxílio de uma faca
de lâmina reta, reunidos por parte e pesados separadamente. O peso das carcaças
incompletas foi obtido por diferença.
A seguir, procedeu-se à retirada da carne de peito e de coxa e sobrecoxa, as
quais foram cortadas em pedaços pequenos, reunidas por repetição, transferidas
para bandejas de alumínio devidamente identificadas e pesadas. As carcaças
incompletas (com ossos) foram submetidas ao mesmo procedimento adotado para a

75
carne de peito e de coxa e sobrecoxa. As tíbias, os fêmures e os esternos retirados
foram descartados e toda gordura visível foi colocada junto da parte correspondente
da carcaça.
Após a pesagem, cada amostra composta de carne de peito, de carne de
coxa + sobrecoxa e de carcaça incompleta foi colocada em estufa a 105 °C por
aproximadamente 24 horas, retirada da estufa, deixada esfriar em temperatura
ambiente por aproximadamente uma hora até que a umidade das amostras entrasse
em equilíbrio com a umidade do ar, pesada em balança de precisão (0,01 g), moída
em moinho de bola até adquirir granulometria bem fina e armazenada em pote
plástico devidamente identificado para posteriores análises químicas de matéria
seca definitiva, extrato etéreo e proteína bruta de acordo com SILVA (1990).
Foram duas unidades por amostra, cinco amostras compostas por
tratamento, vinte amostras compostas por tipo de tecido e sessenta amostras
compostas analisadas vezes dois (duplicata), totalizando cento e vinte análises de
matéria seca, proteína bruta e extrato etéreo.
Procedimentos estatísticos
O seguinte modelo estatístico foi utilizado para as análises dos dados:
Yij = m + Ti + eij.
Em que,
Yij = Observação individual média no tratamento i e na repetição j.
m = Média geral.
Ti = Efeito de tratamento; i = (1, 2, 3 e 4).
eij = Erro aleatório associado a cada observação ij.
Os efeitos da inclusão do colorífico e da niacina suplementar sobre as
diversas variáveis foram verificados pela análise de variância e suas médias foram
comparadas por meio do teste de Tukey, ao nível de 5 % de probabilidade.
Foi realizada análise de correlação entre as variáveis dependentes para
verificar o grau de associação entre as mesmas.
Todas as análises estatísticas foram efetuadas usando-se o programa SAEG
(Sistema de análises estatísticas e genéticas), desenvolvido na Universidade
Federal de Viçosa - UFV (1997).

76
RESULTADOS E DISCUSSÃO
Consumo de ração (CR), ganho de peso (GP), conversão alimentar (CA) e peso (P)
Não foi observado efeito significativo (P>0,05) de tratamentos sobre as
variáveis de desempenho, tanto no período de 21 a 35 quanto no período de 35 a 49
dias de idade das codornas japonesas machos (Quadros 3 e 4).
Quadro 3. Médias do consumo de ração (CR), ganho de peso (GP) e conversão
alimentar (CA) de codornas japonesas machos no período de 21 a 35 dias de idade e peso (P) aos 35 dias de idade
Tratamentos CR (g/ave) GP (g/ave) CA P (g/ave) Ração 183,5 37,87 4,85 102,4
Ração + Colorífico1 182,1 38,47 4,74 102,6 Ração + NS2 187,3 40,11 4,68 104,0
Ração + Colorífico1 + NS2 178,6 38,03 4,73 103,3 1- 4,5 % da ração; 2- niacina suplementar (0,08 % da ração). Quadro 4. Médias do consumo de ração (CR), ganho de peso (GP) e conversão
alimentar (CA) de codornas japonesas no período de 35 a 49 dias de idade e peso (P) aos 49 dias de idade
Tratamentos CR (g/ave) GP (g/ave) CA P (g/ave) Ração 204,9 18,16 11,55 121,7
Ração + Colorífico1 196,3 16,77 11,97 120,5 Ração + NS2 204,9 16,47 12,64 122,9
Ração + Colorífico1 + NS2 200,4 17,10 11,90 120,3 1- 4,5 % da ração; 2- niacina suplementar (0,08 % da ração).
Estes resultados indicam que não houve efeito deletério no uso de doses
elevadas de bixina e niacina sobre o CR, GP, CA e P das codornas.
PAUMGARTTEN et al. (2002) também não verificaram redução no peso corporal
(toxicidade) em ratas com 21 dias de gestação e nas suas proles, com o uso de 500
mg de extrato seco de urucum (28 % de bixina) por kg de peso corporal por dia.
Neste trabalho, o peso corporal das codornas aos 35 e 49 dias de idade
ficou em torno de 103 e 121 g, respectivamente. Entretanto, quando alimentadas
com ração contendo 20 % de proteína bruta e 2.870 kcal de energia metabolizável
por kg de ração, codornas japonesas machos com 35 e 49 dias de idade atingiram o

77
peso corporal de 144 e 166 g, respectivamente (YALÇIN et al., 1995). Embora
tenham sido obtidos em trabalhos diferentes, a diferença no peso para codornas da
mesma espécie pode ser atribuída ao efeito de linhagem, o que pode gerar
diferenças na composição corporal, principalmente quanto à capacidade de
acumular gordura na carcaça.
Triglicerídeos (T) e lipoproteínas de densidade muito baixa (VLDL) no sangue
Não foi observado efeito significativo (P>0,05) de tratamentos sobre os
níveis de T e VLDL no sangue de codornas japonesas machos, tanto no 35° quanto
no 49° dia de idade (Quadro 5).
Quadro 5. Médias dos valores (mg/dl) de triglicerídeos (T) e de lipoproteínas de
densidade muito baixa (VLDL) no sangue de codornas japonesas machos aos 35 e 49 dias de idade
Tratamentos T 35 VLDL 35 T 49 VLDL 49 Ração 40,6 8,12 75,6 15,12
Ração + Colorífico1 39,0 7,80 69,8 13,96 Ração + NS2 34,6 6,92 71,2 14,24
Ração + Colorífico1 + NS2 38,0 7,60 66,8 13,36 1- 4,5 % da ração; 2- niacina suplementar (0,08 % da ração).
Com base nesses valores, nota-se que a bixina e a niacina suplementar não
exerceram atividade hipolipidêmica sobre o metabolismo lipídico de codornas que
receberam alimentação normal ou sem indução de hiperlipidemia. Estes resultados
não corroboram com os resultados dos trabalhos apresentados por SOUZA et al.
(2000), que encontraram reduções significativas na incorporação diária na ração de
70, 350 e 700 mg de um corante industrial contendo 28 % de bixina sobre os níveis
de T no sangue de fêmeas de ratos que não tiveram hiperlipidemia induzida. Em
adição, SANTOS et al. (1999) verificaram um decréscimo numérico no valor de T no
sangue de ratos que receberam ração + triton + 5 mg de ácido nicotínico por Kg de
peso corporal (104,56 mg/dl), em relação a ratos que receberam somente ração
(162,30 mg/dl).
Embora não tenham sido comparados estatisticamente, pode-se observar
que os valores médios de triglicerídeos (T 49) e lipoproteínas de densidade muito
baixa (VLDL 49) no 49° dia foram bem superiores aos valores médios de

78
triglicerídeos (T 35) e lipoproteínas de densidade muito baixa (VLDL 35) no 35° dia
(Quadro 5). Os aumentos médios dos valores de T e VLDL foram de 86,2; 79,0;
105,8 e 75,8 %, para os tratamentos com ração, ração + colorífico, ração + niacina
suplementar e ração + colorífico + niacina suplementar, respectivamente.
A disponibilidade de lipídeos do plasma depende, fundamentalmente, da
lipogênese no fígado, que, por sua vez, depende, em sua maior parte, da
disponibilidade de carboidratos nas rações, que darão origem a moléculas de acetil-
CoA (HERMIER, 1997). Assim, o aumento dos níveis de VLDL e triglicerídeos no
sangue das codornas com 49 dias de idade talvez possa ser explicado pela maior
atividade do fígado, ou seja, pode ter havido, a partir do 35° dia de idade, uma
transformação metabólica crescente de carboidratos oriundos da ração em lipídeos
sangüíneos, indicando que o aporte energético fornecido pela ração poderia estar
acima da exigência das codornas. Os carboidratos podem ter contribuído mais que
os lipídeos porque os níveis de lipídeos contidos nas rações foram baixos (≅ 3,2 %).
Matéria seca (MS), extrato etéreo (EE) e proteína bruta (PB) na carne de peito
Não foi observado efeito significativo (P>0,05) de tratamentos sobre os
teores de MS e PB na carne de peito de codornas japonesas machos com 35 e 49
dias de idade. O mesmo resultado ocorreu em relação aos níveis de EE no 35° dia
de idade. Houve efeito significativo (P≤0,05) de tratamento sobre os níveis de EE no
49° dia de idade das codornas. Codornas alimentadas com 0,08 % de niacina
suplementar (NS) e com 4,5 % de colorífico + 0,08 % de NS nas rações
apresentaram níveis mais elevados de gordura na carne de peito do que codornas
alimentadas com ração e com ração + colorífico (Quadro 6).
Quadro 6. Médias dos valores (% na matéria natural) de matéria seca (MS), extrato
etéreo (EE) e proteína bruta (PB) da carne de peito de codornas japonesas machos aos 35 e 49 dias de idade
Tratamentos MS 35 EE 35 PB 35 MS 49 EE 49 PB 49 Ração 23,7 0,56 20,2 24,0 0,92 b 20,6
Ração + Colorífico1 23,9 0,53 20,0 23,8 0,92 b 20,4 Ração + NS2 23,6 0,46 19,9 24,1 1,42 a 20,6
Ração + Colorífico1 + NS2 23,8 0,52 19,8 24,2 1,33 a 20,5 1- 4,5 % da ração; 2- niacina suplementar (0,08 % da ração); médias seguidas por letras diferentes diferem entre si, pelo teste de Tukey ao nível de 5 % de probabilidade.

79
A literatura não disponibiliza dados sobre os efeitos da inclusão de niacina
suplementar, em rações para aves, na composição química da carne de peito, mas
GOODMANS et al. (1996) revelaram que o efeito do ácido nicotínico sobre o
metabolismo lipídico decorre da capacidade de inibir a lipólise do tecido adiposo, de
reduzir a esterificação dos triglicerídeos no fígado e de aumentar a atividade da
lipase lipoprotéica. Com base nos resultados, pode-se inferir que o mecanismo de
ação do ácido nicotínico explica a elevação do conteúdo de gordura na carne de
peito das codornas, no 49° dia de idade. Todavia, ao contrário do esperado, esse
aumento não foi decorrente da redução dos níveis de triglicerídeos sangüíneos, pois
isto não ocorreu neste trabalho.
Este fato não pôde ser evidenciado e explicado por meio da análise de
correlação, na qual foi obtido um coeficiente baixo e negativo (- 0,18) para teor de
triglicerídeos (T) ou de lipoproteínas de densidade muito baixa (VLDL) no sangue e
teor de gordura na carne de peito, ambas aos 49 dias de idade das codornas
(Quadro 10B). Esses resultados mostram que o aumento no teor de gordura na
carne de peito das codornas aos 49 dias de idade não refletiu em decréscimo nos
níveis de T e VLDL no sangue aos 49 dias de idade das codornas decorrente do
efeito da niacina suplementar (NS), embora tenha havido uma pequena redução
numérica dos T no 49° dia de idade com o tratamento ração + NS (71,2 mg/dl) em
relação ao tratamento com ração (75,6 mg/dl) (Quadro 5).
Mesmo com valor médio inferior ao tratamento ração + NS (1,42 %), o
consumo médio diário de bixina pelas codornas, oriunda da inclusão do colorífico na
ração (0,92 %), não induziu um efeito hipolipidêmico na carne de peito no 49° dia de
idade, em relação ao tratamento com ração (0,92 %) (Quadro 6). LIMA et al. (1999)
sugeriram que o efeito da bixina no metabolismo lipídico ocorre pela ativação da
lipase lipoprotéica, enzima que hidrolisa os triglicerídeos das lipoproteínas de
densidade muito baixa e libera ácidos graxos e glicerol. Os resultados mostram que
esse mecanismo não ocorreu, embora tenha havido uma redução numérica dos
níveis de triglicerídeos no sangue no 49° dia de idade (69,8 mg/dl) com o uso de
ração + colorífico, comparado ao tratamento com ração (75,6 mg/dl) (Quadro 5).
Os teores de matéria seca (MS), proteína bruta (PB) e extrato etéreo (EE) na
carne de peito de codornas japonesas machos com 35 dias de idade foram de 27,1;
18,2 e 4,0 %, respectivamente. No 49° dia, obtiveram-se 29,5 % de MS, 19,4 % de
PB e 7,1 % de EE (YALÇIN et al., 1995). Considerando o 35° dia de idade, os

80
valores de MS e EE obtidos no trabalho de YALÇIN et al. (1995) foram,
respectivamente, 14,0 e 670 % superiores aos valores de MS (23,8 %) e EE (0,52
%) do tratamento ração + colorífico + NS obtidos neste trabalho (Quadro 6). No 49°
dia de idade, a superioridade dos valores de MS na carne de peito obtida no trabalho
de YALÇIN et al. (1995) foi ainda maior: 21,9 %. O aumento do teor de gordura foi
de 434 %.
Em cortes sem pele e osso, a carne de peito de frangos de corte tem
apresentado de 1,6 a 2,4 % de gordura, sendo os valores para frangos machos e
fêmeas bem semelhantes (KESSLER e JÚNIOR, 2001). Verifica-se que o valor de
1,42 % de extrato etéreo (EE), obtido no 49° dia de idade com o uso de niacina
suplementar nas rações, é similar ao relatado por KESSLER e JÚNIOR (2001) e
inferior ao encontrado por YALÇIN et al. (1995). Apesar do objetivo do trabalho ser
redução de gordura corporal, esse aumento do percentual de gordura na carne de
peito das codornas possivelmente não deprecia sua qualidade.
YALÇIN et al. (1995) mencionaram que o conteúdo de proteína bruta (PB) e
gordura da carne de peito foi maior e menor, respectivamente, que o teor de PB e
gordura da carne de coxa + sobrecoxa. Esses dados são coincidentes com os
obtidos neste trabalho, no qual o nível de PB (19,8 %) e EE (0,52 %) na carne de
peito das codornas alimentadas com o tratamento ração + colorífico + niacina
suplementar, com 35 dias de idade (Quadro 6), foi superior e inferior,
respectivamente, ao nível de PB (17,8 %) e EE (0,64 %) na carne de coxa +
sobrecoxa, considerando-se o mesmo tratamento e idade das codornas (Quadro 7).
Matéria seca (MS), extrato etéreo (EE) e proteína bruta (PB) na carne de coxa e sobrecoxa
Não foi observado efeito significativo (P>0,05) de tratamento sobre os teores
de MS e EE na carne de coxa e sobrecoxa de codornas japonesas machos com 35
e 49 dias de idade. O mesmo resultado ocorreu em relação aos níveis de PB no 49°
dia de idade. Houve efeito significativo (P≤0,05) de tratamento sobre os níveis de PB
no 35° dia de idade das codornas. Codornas alimentadas com ração + colorífico +
niacina suplementar apresentaram maior teor de PB na carne de coxa e sobrecoxa
do que codornas alimentadas com ração e com ração + colorífico (Quadro 7).

81
Quadro 7. Médias dos valores (% na matéria natural) de matéria seca (MS), extrato etéreo (EE) e proteína bruta (PB) da carne de coxa e sobrecoxa de codornas japonesas machos aos 35 e 49 dias de idade
Tratamentos MS 35 EE 35 PB 35 MS 49 EE 49 PB 49 Ração 20,7 0,85 17,2 a 20,6 0,72 17,8
Ração + Colorífico1 20,6 0,72 17,6 a 20,3 0,90 17,4 Ração + NS2 20,8 0,79 17,7ab 20,6 1,06 17,5
Ração + Colorífico1 + NS2 21,0 0,64 17,8 b 20,6 0,97 17,6 1- 4,5 % da ração; 2- niacina suplementar (0,08 % da ração); médias seguidas por letras diferentes diferem entre si, pelo teste de Tukey ao nível de 5 % de probabilidade.
Dados sobre os efeitos da inclusão de niacina suplementar (NS) e bixina, em
rações para aves, na composição química da carne de coxa + sobrecoxa são raros
na literatura, contudo, pode-se observar que, no 35° dia de idade, fase em que as
codornas estão em pleno crescimento (Quadro 3), a maior deposição de PB no
tecido muscular (17,8 %) ocorreu às custas da redução do teor de EE (0,64 %) e
aumento do percentual de MS (21,0 %), quando se utilizou o tratamento com ração +
colorífico + NS e comparado ao tratamento com ração (Quadro 7).
O efeito de colorífico e NS na elevação do teor de PB no tecido muscular da
coxa + sobrecoxa pode estar relacionado com a atividade da enzima lipase
lipoprotéica. Niacina e bixina têm sido mencionadas como ativadoras da lipase e isto
pode ser ratificado pelos seus efeitos potencializadores, quando usadas em conjunto
na ração (Quadro 7). A lipase ativada hidrolisa os triglicerídeos presentes nas
lipoproteínas de densidade muito baixa e libera ácidos graxos no sangue. Como há
uma maior demanda de energia para mantença e crescimento, os ácidos graxos
podem ter sido usados preferencialmente para servirem como fonte de energia para
as codornas, em vez de serem incorporados no tecido adiposo, o que pode ser
observado pelo percentual de extrato etéreo (0,64) no 35° dia (Quadro 7). Este efeito
indireto de aumento da proteína bruta (PB) pela redução do extrato etéreo (EE) deve
ser confirmado em outros trabalhos.
Codornas japonesas machos com 35 dias de idade apresentaram 25,0 % de
matéria seca (MS), 17,9 % de PB e 4,6 % de EE na carne de coxa e sobrecoxa. No
49° dia de idade, os valores foram de 27,1 % de MS, 19,5 de PB e 8,3 % de EE
(YALÇIN et al., 1995). Os resultados referentes a MS e EE são destoantes dos
encontrados neste trabalho. Os valores apresentados por YALÇIN et al. (1995)
mostram um aumento de cerca de 19,2 % no valor de MS e de 620 % no valor de

82
EE, quando comparados com o respectivo valor médio de MS (21,0 %) e EE (0,64
%) do tratamento constituído por ração + colorífico + NS, no 35° dia de idade. No 49°
dia de idade, as diferenças foram ainda maiores: 31,6 % para MS e 756 % para EE.
O genótipo das codornas, entre outros fatores, pode ter sido responsável por esta
discrepância.
Matéria seca (MS), extrato etéreo (EE) e proteína bruta (PB) na carcaça Não foi observado efeito significativo (P>0,05) de tratamentos sobre os
teores de MS, EE e PB na carcaça (com osso e sem cabeça, patas, pele, penas,
vísceras, peito, coxa e sobrecoxa) de codornas japonesas machos com 35 e 49 dias
de idade (Quadro 8).
Quadro 8. Médias dos valores (% na matéria natural) de matéria seca (MS), extrato
etéreo (EE) e proteína bruta (PB) da carcaça3 de codornas japonesas machos aos 35 e 49 dias de idade
Tratamentos MS 35 EE 35 PB 35 MS 49 EE 49 PB 49 Ração 24,0 0,88 15,8 24,2 2,08 15,9
Ração + Colorífico1 24,0 0,90 15,9 24,0 2,48 15,6 Ração + NS2 23,7 0,96 15,6 24,6 2,59 15,6
Ração + Colorífico1 + NS2 24,1 0,88 16,0 24,5 2,81 15,8 1- 4,5 % da ração; 2- niacina suplementar (0,08 % da ração); 3- Carcaça com osso e sem cabeça, patas, pele, penas, vísceras, peito, coxa e sobrecoxa.
No entanto, observa-se que houve uma tendência a maior acúmulo de
gordura na carcaça em codornas alimentadas com ração + niacina suplementar (NS)
e com ração + colorífico + NS, comparadas ao tratamento com ração, no 49° dia de
idade. Embora a literatura não disponibilize trabalhos científicos que tratem dos
efeitos da inclusão de bixina e niacina suplementar, em rações de aves, sobre a
composição química da carcaça, os valores obtidos de extrato etéreo (EE) na
carcaça (Quadro 8) e nas partes da carcaça (Quadros 6 e 7) no 49° dia de idade
mostram um comportamento similar no que se refere ao efeito da niacina
suplementar e colorífico em elevar o teor de gordura.
JÚNIOR (1981) encontrou valores bem diferentes de matéria seca (MS),
proteína bruta (PB) e EE na carcaça (matéria natural) de codornas japonesas
machos aos 35 e 49 dias de idade, em relação aos valores obtidos neste trabalho.
Utilizando-se rações com 20 % de PB e 3.200 kcal de energia metabolizável por kg,

83
as codornas apresentaram 30,4; 20,8 e 5,2 % no 35° dia e 34,7; 20,5 e 9,9 % no 49°
dia de idade para MS, PB e EE, respectivamente.
Segundo YALÇIN et al. (1995), muitos fatores podem afetar as
características de carcaça, e estes incluem idade, sexo, genótipo e temperatura de
criação. As diferenças encontradas nos valores de composição química da carcaça e
observadas em trabalhos diferentes podem ser explicadas por esses fatores, bem
como pelo uso de metodologias distintas. No presente trabalho, no cômputo dos
valores de EE na carcaça, não foi incluída a pele, rica em gordura, que foi retirada
no momento do abate das codornas.
Codornas japonesas machos com 49 dias de idade tiveram menores
conteúdos de MS na carcaça do que codornas com 35 dias de idade (JÚNIOR,
1981). Este resultado difere do encontrado no presente trabalho, no qual se observa
uma tendência a maior teor de MS no 49° dia de idade, à exceção do tratamento
com ração + colorífico, embora as diferenças entre valores numéricos tenham sido
de pequena magnitude (Quadro 8).
CONCLUSÕES 1. A bixina do colorífico e a niacina suplementar não exerceram atividade
hipolipidêmica sobre os níveis de triglicerídeos e de lipoproteínas de densidade
muito baixa no sangue e sobre a gordura na carne de peito, de coxa + sobrecoxa
e na carcaça de machos de codornas japonesas alimentadas com rações sem
indução de hiperlipidemia.
2. O uso de 0,08 % de niacina suplementar na ração promoveu aumento dos níveis
de gordura na carne de peito de codornas japonesas machos com 49 dias de
idade.
REFERÊNCIAS BIBLIOGRÁFICAS BERTECHINI, A.G. (1991) Nutrição de Monogástricos. Lavras - MG: Esal/Faepe,
142p.

84
GOODMANS, J.G.; GILMAN, A.G.; LIMBIRD, L.E. (1996) The pharmacological basis
of therapeutics. 9a Ed. The McGraw-Hill Company, 1843p.
GOTTO JR., A.M. (1998) Triglyceride as a risk factor for coronary artery disease.
American Journal of Cardiology, 82:22q-25q.
GRIFFIN, H.D., GODDARD, C., BUTTERWITH, S.C. (1999) Control of lean and fat
deposition in birds. In: Boorman, K.N., Buttery, P.J., Lindsay, D.B. (eds.) The
control of fat and lean deposition. Oxford: Butterworth-Heinemann, p. 231-248.
HAMMAD, S.M., SIEGEL, H.S., MARKS, H.L. (1998) Total cholesterol, total
triglycerides, and cholesterol distribution among lipoproteins as predictors of
atherosclerosis in selected lines of Japanese quail. Comparative Biochemistry and
Physiology, 119A (2):485-492.
HERMIER, D. (1997) Lipoprotein metabolism and fattening in poultry. Journal of
Nutrition, 127:805S-808S.
JÚNIOR, H.M.E. (1981) Carcass composition studies: Influences of age, sex and
calorie-protein content of the diet on carcass composition of japanese quail.
Poultry Science, 60 (11):2506-2512.
KESSLER, A.M., JÚNIOR, P.N.S. (2001) Considerações sobre a quantidade de
gordura na carcaça do frango. A produção animal na visão dos brasileiros.
Piracicaba - SP: Fealq, p. 111-133.
LIMA, L.R.P., OLIVEIRA, T.T., NAGEM, T.J., OLIVEIRA, M.G.A., PINTO, A.S.,
TINOCO, A.L.A., STRINGHETA, P.C., OLIVEIRA, M.L. (1999) Efeito de
flavonóides (luteolina e apigenina) e corantes naturais (norbixina e bixina) do
urucum sobre a atividade da lipase. Revista da Escola de Farmácia e Odontologia
de Alfenas, 1 (21):29-36.
LIMA, L.R.P., OLIVEIRA, T.T., NAGEM, T.J., PINTO, A.S., STRINGHETA, P.C.,
TINOCO, A.L.A., SILVA, J.F. (2001) Bixina, Norbixina e Quercetina e seus efeitos

85
no metabolismo lipídico de coelhos. Brazilian Journal of Veterinary Research and
Animal Science, 38 (4):196-200.
NATIONAL RESEARCH COUNCIL – NRC (1994) Nutrient Requirements of Poultry.
9.ed. Washington: D.C. National Academy Press, 155p.
PAUMGARTTEN, F.J.R., CARVALHO, R.R., ARAÚJO, I.B., PINTO, F.M., BORGES,
O.O., SOUZA, C.A.M., KURIYAMA, S.N. (2002) Evaluation of the developmental
toxicity of annatto in the rat. Food and Chemical Toxicology, 40:1595-1601.
ROSTAGNO, H.S., ALBINO, L.F.T., DONZELE, J.L., GOMES, P.C., FERREIRA,
A.S., OLIVEIRA, R.F., LOPES, D.C. (2000) Tabelas Brasileiras para aves e
suínos - Composição de alimentos e exigências nutricionais. Viçosa - MG: UFV,
Departamento de Zootecnia, 141p.
SANTOS, K.F.R., OLIVEIRA, T.T., NAGEM, T.J., PINTO, A.S., OLIVEIRA, M.G.A.,
SOARES, J.F. (1999) Efeitos das associações de morina-ácido nicotínico e
quercetina-ácido nicotínico no controle de lipídeos. Revista Brasileira de Análises
Clínicas, 31 (1):5-7.
SILVA, D.J. (1990) Análise de alimentos (Métodos químicos e biológicos). 2 ed.
Viçosa - MG: UFV, Imprensa Universitária, 165 p.
SOUZA, E.C.G., STRINGHETA, P.C., OLIVEIRA, T.T., NAGEM, T.J., PINTO, A.S.,
SILVA, J.F., LEÃO, M.A. (2000) Efeitos de bixina sobre o metabolismo lipídico. In:
Anais da 23a Reunião Anual da Sociedade Brasileira de Química, Poços de
Caldas - MG: SBQ, QB-017.
UNIVERSIDADE FEDERAL DE VIÇOSA (1997) SAEG - Sistema de análises
estatísticas e genéticas: Versão 7.1. Viçosa - MG: UFV, 150p.
YALÇIN, S., OGUZ, I., OTLES, S. (1995) Carcase characteristics of quail (Coturnix
coturnix japonica) slaughtered at different ages. British Poultry Science, 36
(3):393-399.

86
WHO (2000) Safety evaluation of certain food additives and contaminants: Evaluation
of national assessments of intake of annatto extracts (bixin). WHO Food Additive
Series, 44:485-492.

87
5. CONCLUSÕES GERAIS
1. Os valores de energia metabolizável do milho, quirera de arroz, óleo de soja
degomado e farelo de soja testados em codornas foram bastante variáveis em
relação aos encontrados na literatura. Torna-se de fundamental importância a
realização de novos ensaios de digestibilidade visando a determinar valores de
energia metabolizável aparente e energia metabolizável aparente corrigida por
retenção de nitrogênio de alimentos a serem testados em codornas japonesas, a
fim de uso específico em formulações de rações para a espécie Coturnix
japonica.
2. Foi eficaz a inclusão do colorífico, em rações de codornas poedeiras, como
agente pigmentante de gemas de ovos. Estudos sobre determinação analítica
dos níveis de bixina em gemas de ovos e viabilidade econômica do uso do
colorífico em rações devem ser realizados para complementar estes resultados.
3. Foi ineficaz a utilização do colorífico e da niacina suplementar como agentes
hipolipidêmicos em codornas japonesas, principalmente em relação ao teor de
gordura na carne de peito e coxa + sobrecoxa. Outras pesquisas devem ser
conduzidas visando à confirmação sobre o uso de colorífico e niacina
suplementar no controle dos níveis de triglicerídeos e lipoproteínas de densidade
muito baixa no sangue.

88
APÊNDICES

89
APÊNDICE A Quadro 1A. Resumo da análise de variância de coloração de gema (CG)
CG Fonte de variação GL QM Bloco 4 0,2422 NS Fonte energética (FE) 1 108,9000* Níveis de colorífico (C) 3 530,4542* Interação FE x C 3 18,1125* Erro (A) 28 0,4529 Dias experimentais (D) 3 11,1333* Interação D x FE 3 0,0417 NS Interação D x C 9 0,8681* Interação D x FE x C 9 0,3819 NS Resíduo 96 0,3734 Coeficiente de Variação (%) 7,4638
* - Significativo pelo teste F ao nível de 5 % de probabilidade; NS - Não Significativo (P>0,05).

90
Qua
dro
2A. R
esum
o da
s an
ális
es d
e va
riânc
ia d
o co
nsum
o de
raçã
o (C
R),
perc
entu
al d
e po
stur
a (P
P), p
eso
méd
io d
os o
vos
(PM
O),
conv
ersã
o al
imen
tar p
or d
úzia
de
ovos
(CAD
) e c
onve
rsão
alim
enta
r por
gra
ma
de o
vos
(CAG
)
CR
PP
PM
O
CAD
C
AG
Font
e de
var
iaçã
o G
L Q
M
QM
Q
M
QM
Q
M
Bloc
o 4
1446
,988
0 N
S 11
,666
8 N
S 0,
0796
NS
515,
7467
NS
0,06
65 N
S Fo
nte
ener
gétic
a (F
E)
1 99
861,
9600
* 33
95,7
290*
0,
0181
NS
1592
6,49
00*
0,93
64*
Nív
el d
e co
loríf
ico
(C)
3 10
08,1
370
NS
92,4
569
NS
0,16
70 N
S 25
80,9
850
NS
0,12
48 N
S In
tera
ção
FE x
C
3 48
05,6
880*
12
5,65
99 N
S 0,
3789
* 14
96,4
760
NS
0,16
20 N
S R
esíd
uo
28
1140
,270
0 58
,258
3 0,
0953
11
36,0
080
0,06
17
Coe
ficie
nte
de V
aria
ção
(%)
5,
78
11,1
5 2,
95
9,06
8,
39
* - S
igni
ficat
ivo
pelo
test
e F
ao n
ível
de
5 %
de
prob
abili
dade
; NS
- Não
Sig
nific
ativ
o (P
>0,0
5).

91
Quadro 3A. Resumo da análise de variância de coloração de gema referente ao efeito dos níveis de colorífico (C), avaliada em cada dia experimental (D)
Fonte de variação GL QM C/D7
Linear 1 363,1512* Quadrático 1 15,0064* Desvio da regressão 1 1,3300
C/D14 Linear 1 401,8613* Quadrático 1 23,2561* Desvio da regressão 1 1,2353
C/D21 Linear 1 385,0312* Quadrático 1 20,3063* Desvio da regressão 1 5,2126
C/D28 Linear 1 355,1112* Quadrático 1 26,4060* Desvio da regressão 1 1,2270
Resíduo 121 0,3933 * - Significativo pelo teste F ao nível de 5 % de probabilidade; D7 - 7º dia do experimento; D14 - 14º dia do experimento; D21- 21º dia do experimento; D28- 28º dia do experimento. Quadro 4A. Resumo da análise de variância de coloração de gema referente ao
efeito dos níveis de colorífico (C), avaliada em cada fonte energética (FE)
Fonte de variação GL QM C/FE1
Linear 1 510,7600* Quadrático 1 21,0128* Desvio da regressão 1 0,4567
C/FE2 Linear 1 1040,0630* Quadrático 1 70,3120* Desvio da regressão 1 2,9857
Resíduo 28 0,4529 * - Significativo pelo teste F ao nível de 5 % de probabilidade; FE1 - milho; FE2 – quirera de arroz.
92
Quadro 5A. Resumo da análise de variância de coloração de gema, referente ao efeito de fonte energética (milho ou quirera de arroz), avaliada em cada nível de colorífico (C)
Fonte de variação GL QM FE/C0 1 127,8063* FE/C1,5 1 24,0250* FE/C3 1 8,1000* FE/C4,5 1 3,3062*
Resíduo 28 0,4529 * - Significativo pelo teste F ao nível de 5 % de probabilidade; FE – fonte energética; C0 – 0 % de colorífico; C1,5 – 1,5 % de colorífico; C3 – 3 % de colorífico; C4,5 – 4,5 % de colorífico. Quadro 6A. Resumo da análise de variância de coloração de gema referente ao
efeito de dia experimental (D), avaliada em cada nível de colorífico (C)
Fonte de variação GL QM D/C0
Linear 1 5,9512* Quadrático 1 1,0562 NS Desvio da regressão 1 0,2105
D/C1,5 Linear 1 14,0450* Quadrático 1 0,9000 NS Desvio da regressão 1 0,9782
D/C3 Linear 1 7,2200* Quadrático 1 0,0250 NS Desvio da regressão 1 4,7981
D/C4,5 Linear 1 5,9513* Quadrático 1 0,0062 NS Desvio da regressão 1 0,0606
Resíduo 96 0,3734 * - Significativo pelo teste F ao nível de 5 % de probabilidade; NS - Não significativo (P>0,05); C0 – 0 % de colorífico; C1,5 – 1,5 % de colorífico; C3 – 3 % de colorífico; C4,5 – 4,5 % de colorífico.

93
Quadro 7A. Resumo da análise de variância do peso médio de ovos referente ao efeito de nível de colorífico (C), avaliada em cada fonte energética (FE)
Fonte de variação GL QM C/FE1
Linear 1 0,5041* Quadrático 1 0,1730 NS Desvio da regressão 1 0,0441
C/FE2 Linear 1 0,4134* Quadrático 1 0,0006 NS Desvio da regressão 1 0,5027
Resíduo 28 0,0953 * - Significativo pelo teste F ao nível de 5 % de probabilidade; NS - Não significativo (P>0,05); FE1 - milho; FE2 – quirera de arroz. Quadro 8A. Resumo da análise de variância de peso médio de ovos, referente ao
efeito de fonte energética (milho ou quirera de arroz), avaliada em cada nível de colorífico (C)
Fonte de variação GL QM FE/C0 1 0,4244* FE/C1,5 1 0,0518 NS FE/C3 1 0,4537* FE/C4,5 1 0,2250 NS
Resíduo 28 0,0953 * - Significativo pelo teste F ao nível de 5 % de probabilidade; NS - Não significativo (P>0,05); FE – fonte energética; C0 – 0 % de colorífico; C1,5 – 1,5 % de colorífico; C3 – 3 % de colorífico; C4,5 – 4,5 % de colorífico. Quadro 9A. Resumo da análise de variância do consumo de ração, referente ao
efeito de nível de colorífico (C), avaliada em cada fonte energética (FE)
Fonte de variação GL QM C/FE1
Linear 1 4874,1320* Quadrático 1 208,3325 NS Desvio da regressão 1 1443,7030
C/FE2 Linear 1 8264,8070* Quadrático 1 2644,7730 NS Desvio da regressão 1 6,0459
Resíduo 28 1140,2700 * - Significativo pelo teste F ao nível de 5 % de probabilidade; NS - Não significativo (P>0,05); FE1 - milho; FE2 – quirera de arroz.

94
Quadro 10A. Resumo da análise de variância do consumo de ração, referente ao efeito do fator fonte energética (milho ou quirera de arroz), avaliada em cada nível de colorífico (C)
Fonte de variação GL QM FE/C0 1 46128,5500* FE/C1,5 1 46511,0400* FE/C3 1 16003,2000* FE/C4,5 1 5636,3560*
Resíduo 28 1140,2700 * Significativo pelo teste F ao nível de 5 % de probabilidade; FE – fonte energética; C0 – 0 % de colorífico; C1,5 – 1,5 % de colorífico; C3 – 3 % de colorífico; C4,5 – 4,5 % de colorífico.

95
APÊNDICE B Quadro 1B. Resumo das análises de variância do consumo de ração (CR), ganho de
peso (GP), conversão alimentar (CA) e peso (P) no período de 21 a 35 dias de idade
CR GP CA P Fonte de variação GL QM QM QM QM Tratamento 3 64,7388 NS 5,2712 NS 0,0267 NS 2,6620 NS Resíduo 16 29,1950 6,3109 0,0762 5,8570 C.V. (%) 3,23 6,42 5,51 2,24 NS – Não significativo pelo teste F ao nível de 5 % de probabilidade. Quadro 2B. Resumo das análises de variância do consumo de ração (CR), ganho de
peso (GP), conversão alimentar (CA) e peso (P) no período de 35 a 49 dias de idade
CR GP CA P Fonte de variação GL QM QM QM QM Tratamento 3 85,8008 NS 2,6957 NS 1,0501 NS 7,0893 NS Resíduo 16 60,1135 8,6177 3,0989 8,3876 C.V. (%) 3,97 16,19 13,86 2,36 NS – Não significativo pelo teste F ao nível de 5 % de probabilidade. Quadro 3B. Resumo das análises de variância da matéria seca (MS), extrato etéreo
(EE) e proteína bruta (PB) na carne de peito de codornas com 35 dias de idade
MS EE PB Fonte de variação GL QM QM QM Tratamento 3 0,0534 NS 0,0080 NS 0,1193 NS Resíduo 16 0,1946 0,0213 0,1061 C.V. (%) 1,75 26,76 1,65 NS - Não significativo pelo teste F ao nível de 5 % de probabilidade.

96
Quadro 4B. Resumo das análises de variância da matéria seca (MS), extrato etéreo (EE) e proteína bruta (PB) na carne de coxa e sobrecoxa de codornas com 35 dias de idade
MS EE PB Fonte de variação GL QM QM QM Tratamento 3 0,1465 NS 0,0416 NS 0,4011* Resíduo 16 0,2063 0,0445 0,0835 C.V. (%) 2,14 27,95 2,08 * - Significativo pelo teste F ao nível de 5 % de probabilidade; NS - Não significativo (P>0,05). Quadro 5B. Resumo das análises de variância da matéria seca (MS), extrato etéreo
(EE) e proteína bruta (PB) da carcaça de codornas com 35 dias de idade
MS EE PB Fonte de variação GL QM QM QM Tratamento 3 0,1028 NS 0,0071 NS 0,1360 NS Resíduo 16 0,4908 0,0226 0,1112 C.V. (%) 2,74 15,70 2,14 NS - Não significativo pelo teste F ao nível de 5 % de probabilidade. Quadro 6B. Resumo das análises de variância dos triglicerídeos (T) e lipoproteínas
de densidade muito baixa (VLDL) no sangue de codornas com 35 e 49 dias de idade
T 35 VLDL 35 T 49 VLDL 49 Fonte de variação GL QM QM QM QM Tratamento 3 32,1833 NS 1,2873 NS 66,9833 NS 2,6793 NS Resíduo 16 58,0250 2,3210 297,4750 11,8990 C.V. (%) 19,30 19,30 22,81 22,81 NS – Não significativo pelo teste F ao nível de 5 % de probabilidade.
97
Quadro 7B. Resumo das análises de variância da matéria seca (MS), extrato etéreo (EE) e proteína bruta (PB) na carne de peito de codornas com 49 dias de idade
MS EE PB Fonte de variação GL QM QM QM Tratamento 3 0,1351 NS 0,3506* 0,0698 NS Resíduo 16 0,1530 0,0207 0,1633 C.V. (%) 1,61 23,58 1,88 * - Significativo pelo teste F ao nível de 5 % de probabilidade; NS - Não significativo (P>0,05). Quadro 8B. Resumo das análises de variância da matéria seca (MS), extrato etéreo
(EE) e proteína bruta (PB) na carne de coxa e sobrecoxa de codornas com 49 dias de idade
MS EE PB Fonte de variação GL QM QM QM Tratamento 3 0,0935 NS 0,0996 NS 0,1685 NS Resíduo 16 0,1269 0,0520 0,0813 C.V. (%) 1,70 26,70 1,75 NS – Não significativo pelo teste F ao nível de 5 % de probabilidade. Quadro 9B. Resumo das análises de variância da matéria seca (MS), extrato etéreo
(EE) e proteína bruta (PB) na carcaça de codornas com 49 dias de idade
MS EE PB Fonte de variação GL QM QM QM Tratamento 3 0,3905 NS 0,4634 NS 0,1335 NS Resíduo 16 0,2504 0,1947 0,0832 C.V. (%) 2,15 19,56 1,92 NS – Não significativo pelo teste F ao nível de 5 % de probabilidade.

98
Qua
dro
10B.
Cor
rela
ções
de
Pear
son
entre
trig
licer
ídeo
s no
san
gue
(TS)
, lip
opro
teín
as d
e m
uito
bai
xa d
ensi
dade
(VL
DL)
, ex
trato
et
éreo
no
peito
(EEP
), ex
trato
eté
reo
na c
oxa
e so
brec
oxa
(EEC
SC) e
ext
rato
eté
reo
na c
arca
ça (E
ECAR
)
TS 3
5a VL
DL
35
EEP
35
EEC
SC 3
5 EE
CAR
35
TS 4
9b VL
DL
49
EEP
49
EEC
SC 4
9 EE
CAR
49
TS 3
5 a
----
1,0
(0,0
001)
1 0,
72 (0
,000
2)
0,56
(0,0
050)
0,
44 (0
,024
8)
0,10
(0,3
349)
0,
10 (0
,334
9)
- 0,0
9(0,
358)
0,
17 (0
,236
4)
0,23
(0,1
632)
VL
DL
35
1,00
(0,0
001)
---
- 0,
72 (0
,000
2)
0,56
(0,0
050)
0,
44 (0
,024
8)
0,10
(0,3
349)
0,
10 (0
,334
9)
- 0,0
9(0,
358)
0,
17 (0
,236
4)
0,23
(0,1
632)
EE
P 35
0,
72 (0
,000
2)
0,72
(0,0
002)
---
- 0,
73 (0
,000
1)
0,66
(0,0
009)
0,
005
(0,4
91)
0,00
5 (0
,491
) - 0
,08(
0,37
4)
- 0,2
0(0,
199)
0,
04 (0
,425
6)
EEC
SC 3
5 0,
56 (0
,005
0)
0,56
(0,0
050)
0,
73 (0
,000
1)
----
0,52
(0,0
093)
- 0
,13(
0,29
0)
- 0,1
3(0,
290)
0,
007(
0,48
9)
0,02
(0,4
718)
0,
15 (0
,267
8)
EEC
AR 3
5 0,
44 (0
,024
8)
0,44
(0,0
248)
0,
66 (0
,000
9)
0,52
(0,0
093)
---
- - 0
,11(
0,33
0)
- 0,1
1(0,
330)
0,
13 (0
,291
4)
0,14
(0,2
776)
0,
15 (0
,261
0)
TS 4
9 b
0,10
(0,3
349)
0,
10 (0
,334
9)
0,00
5 (0
,491
) - 0
,13(
0,29
0)
- 0,1
1(0,
330)
---
- 1,
00 (0
,000
1)
- 0,1
8(0,
229)
- 0
,18(
0,22
6)
- 0,3
8(0,
049)
VL
DL
49
0,10
(0,3
349)
0,
10 (0
,334
9)
0,00
5 (0
,491
) - 0
,13(
0,29
0)
- 0,1
1(0,
330)
1,
00 (0
,000
1)
----
- 0,1
8(0,
229)
- 0
,18(
0,22
6)
- 0,3
8(0,
049)
EE
P 49
- 0
,09(
0,35
8)
- 0,0
9(0,
358)
- 0
,08(
0,37
4)
0,00
7(0,
489)
0,
13 (0
,291
4)
- 0,1
8(0,
229)
- 0
,18(
0,22
9)
----
0,65
(0,0
009)
0,
64 (0
,001
3)
EEC
SC 4
9 0,
17 (0
,236
4)
0,17
(0,2
364)
- 0
,20(
0,19
9)
0,02
(0,4
718)
0,
14 (0
,277
6)
- 0,1
8(0,
226)
- 0
,18(
0,22
6)
0,65
(0,0
009)
---
- 0,
78 (0
,000
1)
EEC
AR 4
9 0,
23 (0
,163
2)
0,23
(0,1
632)
0,
04 (0
,425
6)
0,15
(0,2
678)
0,
15 (0
,261
0)
- 0,3
8(0,
049)
- 0
,38(
0,04
9)
0,64
(0,0
013)
0,
78 (0
,000
1)
----
1 - O
nív
el d
e si
gnifi
cânc
ia d
o co
efic
ient
e de
cor
rela
ção
está
apr
esen
tado
ent
re p
arên
tese
s; a -
35°
dia
de id
ade;
b - 49°
dia
de id
ade.
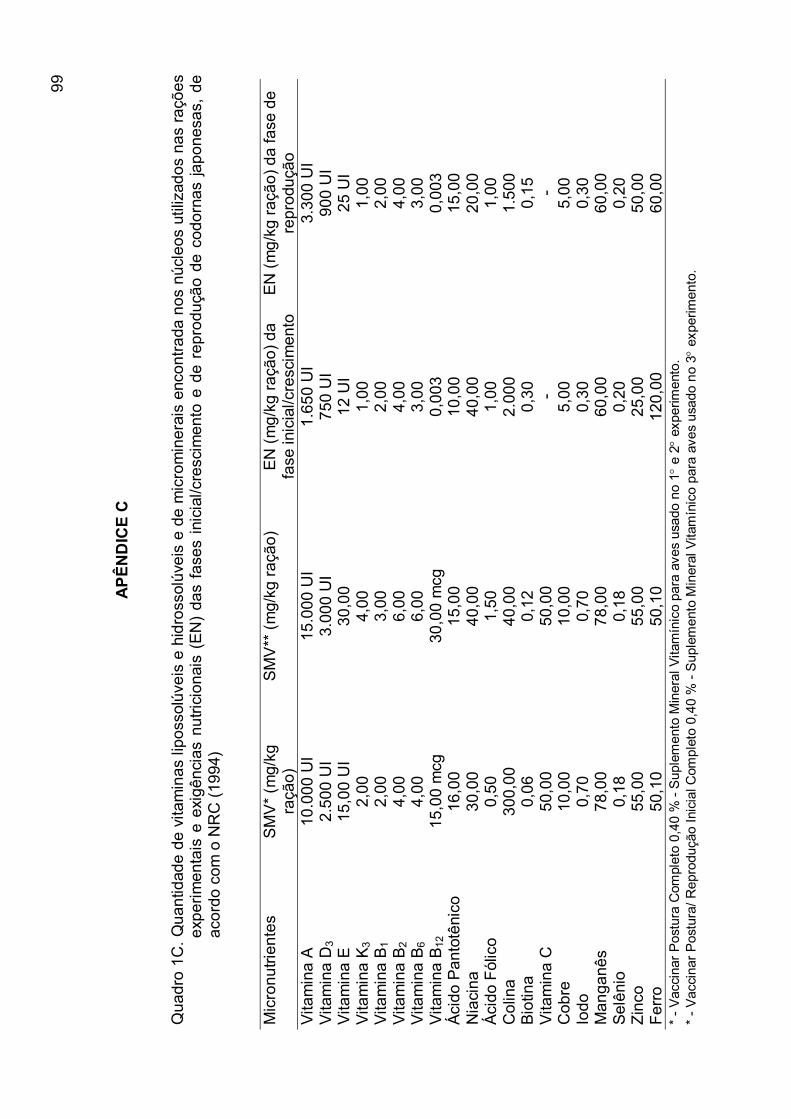
99
APÊN
DIC
E C
Q
uadr
o 1C
. Qua
ntid
ade
de v
itam
inas
lipo
ssol
úvei
s e
hidr
osso
lúve
is e
de
mic
rom
iner
ais
enco
ntra
da n
os n
úcle
os u
tiliz
ados
nas
raçõ
es
expe
rimen
tais
e e
xigê
ncia
s nu
trici
onai
s (E
N)
das
fase
s in
icia
l/cre
scim
ento
e d
e re
prod
ução
de
codo
rnas
japo
nesa
s, d
e ac
ordo
com
o N
RC
(199
4)
Mic
ronu
trien
tes
SMV*
(mg/
kg
raçã
o)
SMV*
* (m
g/kg
raçã
o)
EN (m
g/kg
raçã
o) d
a fa
se in
icia
l/cre
scim
ento
EN
(mg/
kg ra
ção)
da
fase
de
repr
oduç
ão
Vita
min
a A
10.0
00 U
I 15
.000
UI
1.65
0 U
I 3.
300
UI
Vita
min
a D
3 2.
500
UI
3.00
0 U
I 75
0 U
I 90
0 U
I Vi
tam
ina
E 15
,00
UI
30,0
0 12
UI
25 U
I Vi
tam
ina
K 3
2,00
4,
00
1,00
1,
00
Vita
min
a B 1
2,
00
3,00
2,
00
2,00
Vi
tam
ina
B 2
4,00
6,
00
4,00
4,
00
Vita
min
a B 6
4,
00
6,00
3,
00
3,00
Vi
tam
ina
B 12
15,0
0 m
cg
30,0
0 m
cg
0,00
3 0,
003
Ácid
o Pa
ntot
ênic
o 16
,00
15,0
0 10
,00
15,0
0 N
iaci
na
30,0
0 40
,00
40,0
0 20
,00
Ácid
o Fó
lico
0,50
1,
50
1,00
1,
00
Col
ina
300,
00
40,0
0 2.
000
1.50
0 Bi
otin
a 0,
06
0,12
0,
30
0,15
Vi
tam
ina
C
50,0
0 50
,00
- -
Cob
re
10,0
0 10
,00
5,00
5,
00
Iodo
0,
70
0,70
0,
30
0,30
M
anga
nês
78,0
0 78
,00
60,0
0 60
,00
Selê
nio
0,18
0,
18
0,20
0,
20
Zinc
o 55
,00
55,0
0 25
,00
50,0
0 Fe
rro
50,1
0 50
,10
120,
00
60,0
0 * -
Vac
cina
r Pos
tura
Com
plet
o 0,
40 %
- Su
plem
ento
Min
eral
Vita
mín
ico
para
ave
s us
ado
no 1°
e 2°
exp
erim
ento
. * -
Vac
cina
r Pos
tura
/ Rep
rodu
ção
Inic
ial C
ompl
eto
0,40
% -
Supl
emen
to M
iner
al V
itam
ínic
o pa
ra a
ves
usad
o no
3°
expe
rimen
to.