
EFEITOS DA PRIVAÇÃO DE SONO SOBRE A PLASTICIDADE ... · Por todos os desafios e sugestões que...
133
i FRANCISCO PAULINO DUBIELA EFEITOS DA PRIVAÇÃO DE SONO SOBRE A PLASTICIDADE HIPOCAMPAL: a importância dos receptores AMPA Tese apresentada à Universidade Federal de São Paulo – Escola Paulista de Medicina como parte dos requisitos para obtenção do Título de Doutor em Ciências. UNIVERSIDADE FEDERAL DE SÃO PAULO SÃO PAULO 2012
Transcript of EFEITOS DA PRIVAÇÃO DE SONO SOBRE A PLASTICIDADE ... · Por todos os desafios e sugestões que...

i
FRANCISCO PAULINO DUBIELA
EFEITOS DA PRIVAÇÃO DE SONO SOBRE A
PLASTICIDADE HIPOCAMPAL:
a importância dos receptores AMPA
Tese apresentada à Universidade
Federal de São Paulo – Escola
Paulista de Medicina como parte dos
requisitos para obtenção do Título de
Doutor em Ciências.
UNIVERSIDADE FEDERAL DE SÃO PAULO
SÃO PAULO
2012

ii
Dubiela, Francisco Paulino Efeitos da Privação de Sono sobre a Plasticidade Hipocampal: a importância dos receptores AMPA/ Francisco Paulino Dubiela – São Paulo, 2012. xxi, 110 f. Tese (Doutorado) - Universidade Federal de São Paulo. Escola Paulista de Medicina. Programa de Pós-Graduação em Psicobiologia. Título em inglês: Effects of sleep deprivation on hippocampal plasticity: the relevance of AMPA receptors 1. privação de sono 2. receptores AMPA 3. memória 4. hipocampo.

i
FRANCISCO PAULINO DUBIELA
EFEITOS DA PRIVAÇÃO DE SONO SOBRE A
PLASTICIDADE HIPOCAMPAL:
a importância dos receptores AMPA
Tese apresentada à Universidade
Federal de São Paulo – Escola
Paulista de Medicina como parte dos
requisitos para obtenção do Título de
Doutor em Ciências.
Orientadora: Profa. Dra. Débora Cristina Hipólide
Co- Orientador: Prof. Dr. Claudio Marcos Teixeira de Queiroz
UNIVERSIDADE FEDERAL DE SÃO PAULO
SÃO PAULO
2012

ii
UNIVERSIDADE FEDERAL DE SÃO PAULO
ESCOLA PAULISTA DE MEDICINA
DEPARTAMENTO DE PSICOBIOLOGIA
Chefe do departamento: Profa. Dra. Maria Lucia Oliveira de Souza Formigoni
Coordenador do curso de pós-graduação: Profa. Dra. Vânia D’Almeida

iii
Esta tese de doutoramento foi realizada no Departamento de
Psicobiologia da Universidade Federal de São Paulo – Escola Paulista de
Medicina, com o apoio financeiro das fundações:
FAPESP – Fundação de Amparo à Pesquisa do Estado de São Paulo,
bolsa de doutoramento, processo n°: 2007/53176-8.
CNPq – Conselho Nacional de Desenvolvimento Científico e Tecnológico,
bolsa de doutoramento, processo n° 142594/2007-5.
CAPES – Coordenação de Aperfeiçoamento de Pessoal de Nível
Superior, bolsa de doutorado sanduiche no exterior, processo n° 1324-09-8.
AFIP – Associação Fundo de Incentivo a Pesquisa.

iv
À minha família, que cresce a cada dia!

v
AGRADECIMENTOS
À Profa. Dra. Débora Cristina Hipólide, por sua orientação dedicada e amizade
constante. Por dar o impulso inicial de minha carreira científica e auxiliar na
conclusão dessa etapa tão importante.
Ao Prof. Dr. Claudio Marcos Teixeira de Queiroz, pela co-orientação e
motivação. Por todos os desafios e sugestões que tornaram esse trabalho
possível.
Ao Prof. Dr. Sergio Tufik, por acreditar e apoiar incondicionalmente meu
potencial.
Ao Prof. Dr. Pierre Hervé Luppi, por seu acolhimento e atenção em seu
laboratório. Por seu espírito sagaz e carisma contagiante.
Ao Prof. Dr. Damien Gervasoni, pelo suporte, confiança e perseverança. Por
sua amizade e profissionalismo.
Aos Prof. Dr. Jackson Bittencourt e a Profa. Dra. Luciane Sita, por sua
recepção e atenção em seu laboratório.
À Profa. Dra. Maria Gabriela Menezes de Oliveira, pelos conselhos valiosos
nos momentos cruciais desse trabalho.

vi
À Dra. Juliana Carlota, por sua valiosa colaboração nesse estudo e em outros
tantos experimentos.
Aos membros participantes da banca de qualificação e defesa.
Aos amigos e colegas do Departamento de Psicobiologia, que compartilharam
os sonhos, angústias e desafios das neurociências.
Aos técnicos José Bernardo da Costa e Danilo Carlos Machado pelo auxílio
dos experimentos farmacológicos deste estudo.
Às técnicas Joelcimar Martins da Silva e Diva Lima, pela valiosa ajuda na
realização dos experimentos neuroquímicos e moleculares.
Às secretárias do Departamento de Psicobiologia, Nereide Lourdes Garcia,
Mara Vianna e Jacqueline Wetzl, por se dedicarem à resolução dos assuntos
burocráticos da minha pós graduação.
À Leticia Lobo Luppi, por sua imensa e carinhosa asa em terras
desconhecidas.
À minha esposa, Larissa Zeggio, meu primeiro e último amor, que trouxe
bonança nos mares revoltos desse trabalho.

vii
Aos meus pais, Valdir Luiz Dubiela e Ana Yara Paulino, pelo amor incondicional
e amparo constante.
Aos meus irmãos, Joana, Pablo, Rafael, Suiá e Felipe, pelo apoio moral.
À minha sogra, Maria Teresa Zeggio, pela convivência agradável e por apoiar a
realização de novos projetos de vida.
Aos ratos, que involuntariamente permitiram que esse trabalho fosse realizado.
Ao CNPq, à FAPESP, à CAPES e à AFIP pelo apoio financeiro.
A você, que lê esta tese.

viii
Do you see the way that tree bends?
Does it inspire?
Leaning out to catch the sun’s rays
A lesson to be applied
I remember when I swore I knew everything
Let's say knowledge is a tree and it's growing up just like me
I'm so light the wind it shakes
I'm so high the sky I scrape
I'm so light I hold just one breath and go back to my nest
Sleep with innocence...
Present Tense/ Up my Tree
Eddie Vedder

ix
Dubiela, FP. Efeitos da Privação de Sono sobre a Plasticidade Hipocampal: a
importância dos receptores AMPA (tese de doutoramento). Universidade
Federal de São Paulo, 2012.
1. RESUMO
Diversos estudos mostram que a falta de sono interfere na formação de novas
memórias, especialmente as dependentes do hipocampo. Em consonância, há
trabalhos que mostram que a privação de sono (PS) interfere sobre o ritmo teta
hipocampal (4 a 12 Hz), mas não há dados consistentes sobre seus efeitos
durante a aprendizagem de tarefas de memóra. Há evidências indiretas que
sugerem o envolvimento de receptores glutamatérgicos ionotrópicos do tipo
AMPA, que constituem um fator essencial tanto para a aprendizagem de
tarefas como para a plasticidade hipocampal. Tratamentos com fármacos que
atuam sobre os receptores AMPA têm se mostrado eficazes na prevenção de
prejuízos de memória induzidos por patologias neurais, bem como na
modulação do ritmo teta hipocampal. Portanto, o objetivo do presente estudo
foi investigar o envolvimento dos receptores AMPA hipocampais sobre os
prejuízos de memória de ratos, induzidos pela privação de sono. Na primeira
etapa, foram realizadas análises da marcação autorradiográfica de receptores
AMPA e da transcrição da subunidade GluR1 após a privação de sono, bem
como experimentos farmacológicos com potenciador (aniracetam) e
antagonista (GYKI 52466) de receptores AMPA, com o propósito de prevenir os

x
prejuízos apresentados por ratos privados de sono na tarefa de esquiva
inibitória. Na segunda etapa, foi realizada a caracterização do ritmo teta
hipocampal e seus subtitpos (teta tipo 1 e tipo 2) durante uma tarefa de
reconhecimento de objetos em após PS. Na primeira etapa, foi observado uma
diminuição da marcação autorradiográfica de receptores AMPA
especificamente na formação hipocampal de ratos privados de sono por 96 h,
que se normaliza com uma recuperação de sono por 24 h. Por outro lado, não
foram encontradas alterações na hibridização in situ da subunidade GluR1
hipocampal. Dados de experimentos farmacológicos mostraram que a
administração aguda de aniracetam (100 mg/kg) foi eficaz na prevenção de
prejuízo de desempenho de esquiva inibitória de ratos privados de sono, ao
passo que tratamentos agudo e crônico com GYKI 52466 não surtiram efeito.
Na segunda etapa, ratos com implantes de eletrodos na formação hipocampal
desempenharam uma tarefa de reconhecimento de objetos após PS de 72 h, e
apresentaram prejuízo nessa condição. A análise da potência do ritmo teta
hipocampal mostrou que o ritmo teta tipo 2 está reduzido durante o treino da
tarefa, e que se normaliza na ocorrência do teste após a recuperação de sono.
A partir dos dados obtidos nas duas etapas, conclui-se que: 1) a privação de
sono reduz a marcação de receptores AMPA hipocampais, por meio de
modificações pós-transcricionais de suas subunidades, e que a recuperação de
sono normaliza esse efeito; 2) o potenciador aniracetam previne o prejuízo de
memória de ratos privados de sono na tarefa de esquiva inibitória; 3) a privação
de sono modifica o ritmo teta hipocampal durante a aprendizagem de uma
tarefa de reconhecimento de objetos, e sugere a participação dos receptores
AMPA hipocampais nessa condição.

xi
Palavras-chave: privação de sono, receptores AMPA, memória, hipocampo.

xii
Dubiela, FP. Effects of sleep deprivation on hippocampal plasticity: the
relevance of AMPA receptors (P.h.D. thesis). Universidade Federal de São
Paulo, 2012.
ABSTRACT
Several studies have shown that lack of sleep influences formation of new
memories, specially hippocampus dependent ones. In addition, reports have
shown that sleep deprivation (SD) disturbs hippocampal theta rhythm (4-12 Hz),
but there is no consistent data regarding its effects on task learning. There is a
body of indirect evidence suggesting the involvement of glutamatergic
ionotropic AMPA receptors, which represent an essential factor both for task
learning as for hippocampal plasticity. Pharmacological treatment with drugs
acting on AMPA receptors has been regarded as an useful therapy to prevent
memory deficits induced by neural pathologies and to modulate hippocampal
theta rhythm. Therefore, the objective of the present study was to investigate
AMPA receptors involvement on memory deficits of rats induced by SD. The
first part relayed on analyses of AMPA receptor binding and GluR1 subunit
transcription after SD, and pharmacological experiments with a potentiator
(aniracetam) and an antagonist (GYKI 52466) of AMPA receptors in order to
prevent SD induced deficits in an inhibitory avoidance task. In the second part,
the hippocampal theta rhythm and its components (type 1 and type 2 theta)

xiii
were studied during an object recognition task after SD. Results from the first
part showed AMPA receptor binding reduction specifically on hippocampal
areas after 96 h SD, which are normalized after 24 h sleep recovery. No
changes were found in GluR1 subunit transcription within the hippocampus of
sleep deprived rats. Pharmacological results displayed an efficacious effect of
acute aniracetam treatment (100 mg/kg) in order to prevent SD induced
performance deficit in the inhibitory avoidance task, while both acute and
chronic treatments with GYKI 52466 were devoid of any mnesic effect. In the
second part, rats with electrodes implants in the hippocampal formation
performed an object recognition task after 72 h SD, and displayed a
performance deficit in this condition. Power spectrum analyses of the
hippocampal theta rhythm showed a reduction of type 2 theta during learning,
which is normalized after sleep recovery in between sessions of the task.
Altogether data from the two parts one conclude that 1) sleep deprivation
reduces hippocampal AMPA receptor binding, through post transcriptional
mechanisms, which is normalized after sleep recovery; 2) AMPA potentiator
aniracetam prevents memory deficit of sleep deprived rats in the inhibitory
avoidance task; 3) sleep deprivation changes hippocampal theta rhythm during
task learning, with potential involvement of AMPA receptors of the hippocampal
formation.
Keywords: sleep deprivation, AMPA receptors, memory, hippocampus.

xiv
ÍNDICE
1. RESUMO ............................................................................................................ IIX
1.1. Lista de Abreviaturas..................................................................................XVII
1.2. Lista de Figuras.........................................................................................XVIII
1.3. Lista de Tabelas..........................................................................................XXI
2. INTRODUÇÃO ...................................................................................................... 1
2.1. Perda de Sono e de Memória ........................................................................ 1
2.2. A Privação de Sono e os Receptores AMPA ................................................. 6
2.3. Modulação Farmacológica de Receptores AMPA ........................................ 11
2.4. Receptores AMPA e o Ritmo Teta Hipocampal ........................................... 14
2.5. Justificativa .................................................................................................. 18
3. OBJETIVOS ....................................................................................................... 20
4. PRIMEIRA ETAPA: AVALIAÇÃO NEUROQUÍMICA E FARMACOLÓGICA DOS
RECEPTORES AMPA NA FORMAÇÃO DE MEMÓRIAS EM RATOS PRIVADOS DE
SONO ......................................................................................................................... 22
4.1. Experimento 1: Efeitos da Privação e Recuperação de Sono na marcação
de [3H]AMPA e hibridização in situ de GluR1 ...................................................... 22
4.1.1. Experimento 1-1: Autorradiografia de Receptores AMPA após Privação e
Recuperação de Sono ........................................................................................ 22
4.1.2. Experimento 1-2: Hibridização in Situ da Subunidade GluR1 após Privação
e Recuperação de Sono...................................................................................... 27

xv
4.2. Experimento 2 – Efeitos da Administração de Potenciador e Antagonista
de receptores AMPA no desempenho de Esquiva Inibitória de Ratos Privados de
Sono 30
4.2.1. Experimento 2-1: Efeitos da Administração Aguda de Aniracetam em
Ratos Privados de Sono...................................................................................... 33
4.2.3. Experimento 2-2: Efeitos da Administração Aguda de GYKI 52466 em
Ratos Privados de Sono...................................................................................... 34
4.2.4. Experimento 2-3: Efeitos da Administração Crônica de GYKI 52466 em
Ratos Privados de Sono...................................................................................... 35
4.3. RESULTADOS ........................................................................................ 37
4.3.1. Experimento 1-1: Autorradiografia de Receptores AMPA após Privação e
Recuperação de Sono ........................................................................................ 37
4.3.2. Experimento 1-2: Hibridização in Situ da Subunidade GluR1 após Privação
e Recuperação de Sono...................................................................................... 39
4.3.3. Experimento 2-1: Efeitos da Administração Aguda de Aniracetam em
Ratos Privados de Sono...................................................................................... 40
4.3.4. Experimento 2-2: Efeitos da Administração Aguda de GYKI 52466 em
Ratos Privados de Sono...................................................................................... 42
4.3.5. Experimento 2-3: Efeitos da Administração Crônica de GYKI 52466 em
Ratos Privados de Sono...................................................................................... 43
4.4. DISCUSSÃO PARCIAL ........................................................................... 44
4.5. Experimento Complementar: Efeitos da Administração Intra-Hipocampal
de Aniracetam em Ratos Privados de Sono no Condicionamento de Medo ........ 50
4.5.1. Resultados .............................................................................................. 57
4.5.2. Discussão do Experimento Complementar .............................................. 59

xvi
5. SEGUNDA ETAPA: AVALIAÇÃO DO RITMO TETA HIPOCAMPAL DE RATOS
PRIVADOS DE SONO DURANTE A TAREFA DE RECONHECIMENTO DE
OBJETOS .................................................................................................................. 61
5.1. Metodologia Geral ................................................................................... 61
5.1.1. Experimento 1 – Efeitos da Privação de Sono na Tarefa de
Reconhecimento de Objetos ............................................................................... 66
5.1.2. Experimento 2 – Análise da Potência de Ritmo Teta Hipocampal durante
uma Tarefa de Reconhecimento de Objetos em Ratos Privados de Sono .......... 70
5.2. RESULTADOS ........................................................................................ 75
5.2.1. Experimento 1 – Efeitos da Privação de Sono na Tarefa de
Reconhecimento de Objetos ............................................................................... 75
5.2.2. Experimento 2 – Análise da Potência de Ritmo Teta Hipocampal durante
uma Tarefa de Reconhecimento de Objetos em Ratos Privados de Sono .......... 81
5.3. DISCUSSÃO PARCIAL ........................................................................... 85
6. DISCUSSÃO GERAL ......................................................................................... 89
7. CONCLUSÕES ................................................................................................... 95
8. REFERÊNCIAS BIBLIOGRÁFICAS ................................................................... 96
9. ANEXO ............................................................................................................. 108

xvii
1.1. LISTA DE ABREVIATURAS
Aniracetam – (1-(4-metoxibenzoil)-2-pirrolidinona
AMPA – ácido α-amino-3-hidroxi-5-metil-4-isoxazol propiônico
ANOVA – Análise de Variância
EEG – eletroencefalografia
GYKI 52466 – (1-(4-aminofenil)-4-metil-7,8-metilenedioxi-5H-2,3-
benzodiazepina
i.p. – intraperitoneal
LTP – Long term potentiation (potenciação de longa duração)
NMDA – N-metil-D-aspartato
PS – Privação de sono
REM – rapid eye movements (movimentos rápidos dos olhos)
SOL – Sono de ondas lentas
SP – Sono paradoxal

xviii
1.2. LISTA DE FIGURAS
Figura 1: Atenuação do prejuízo de esquiva inibitória de ratos privados de sono porde
sono por 96 h após recuperação por 24 h.......................................................................6
Figura 2: Privação de sono por meio do método da plataforma múltipla
modificada.....................................................................................................................25
Figura 3: Desenho esquemático do experimento 1.......................................................26
Figura 4: Aparato de esquiva inibitória..........................................................................32
Figura 5: Desenho esquemático do experimento 2-1....................................................33
Figura 6: Desenho esquemático do experimento 2-2....................................................35
Figura 7: Desenho esquemático do experimento 2-3....................................................36
Figura 8: Imagens computadorizadas da marcação de [3H]AMPA em animais controle,
privado de sono e recuperado de sono.........................................................................38
Figura 9: Imagem computadorizada da hibridização in situ da subunidade GluR1 na
formação hipocampal de um animal controle................................................................40
Figura 10: Latência de passagem de ratos privados de sono tratados com aniracetam
na tarefa de esquiva inibitória........................................................................................41
Figura 11: Latência de passagem de ratos privados tratados agudamente com GYKI
52466 na tarefa de esquiva inibitória.............................................................................42
Figura 12: Latência de passagem de ratos privados tratados cronicamente com GYKI
52466 na tarefa de esquiva inibitória.............................................................................43
Figura 13: Desenho experimental do experimento complementar................................53
Figura 14: Desenho experimental da tarefa de condicionamento clássico de medo....55
Figura 15: Localização histológica de implante de cânula no hipocampo dorsal..........56
Figura 16: Tempo médio por minuto de congelamento de animais privados de sono
tratados com injeção intrahipocampal de aniracetam durante o treino de
condicionamento de medo............................................................................................57

xix
Fig 17: Tempo médio por minuto de congelamento de animais privados de sono
tratados com injeção intrahipocampal de aniracetam durante o teste de
condicionamento de medo ao contexto.........................................................................58
Fig 18: Tempo médio por minuto de congelamento de animais privados de sono
tratados com injeção intrahipocampal de aniracetam durante o teste de
condicionamento de medo ao som................................................................................59
Figura 19: A e C: exemplos de arranjos de eletrodos implantados no córtex S1 (matriz
de 8x2 eletrodos) e no hipocampo (matriz de 4x4 eletrodos); B: amostra da localização
do posicionamento dos eletrodos por reconstrução histológica na formação
hipocampal; D: diagrama contendo agrupamentos de estados do ciclo sono-vigília
(QW: vigília quieta, AE: vigília ativa, SWS: sono de ondas lentas, REM: sono
paradoxal, IS: sono intermediário, WT: movimento de vibrissas), baseados nas
análises de componente das razão 1 (r1, 0.5-20/0.5-55 Hz) e razão 2 (r2, 0.5-4.5/0.5-9
Hz) dos valores obtidos do espectro de frequência de potenciais de campo locais por
segundo.........................................................................................................................65
Figura 20: Acima, desenho experimental da tarefa de reconhecimento de objetos;
abaixo, exemplos de objetos utilizados na tarefa com diferentes formas e texturas....67
Figura 21: A e B: amostras de registros de disparos neuronais e potenciais de campo
locais com visualização simultânea do comportamento do animal; C: desenho
experimental do estudo. Animais foram habituados á arena de registro 2 h antes da
primeira sessão da tarefa de reconhecimento de
objetos...........................................................................................................................68
Figura 22: Registro de potenciais de campo locais hipocampais durante uma sessão
da tarefa de reconhecimento de objetos. A: trecho de registro de um canal localizado
na formação hipocampal. B: espectrograma do canal ao longo da sessão (note a
predominância de ritmo teta hipocampal na faixa de 4 a 12 Hz). C: animal exposto aos
objetos da tarefa de reconhecimento............................................................................71

xx
Figura 23: Razão entre pico e média de faixa de frequências do ritmo teta hipocampal
(4-12 Hz). Os canais hipocampais (numerados de 16 a 32) de cada animal foram
analisados para seleção daquele com maior valor de pico (em vermelho) e menor valor
de média (em azul)........................................................................................................72
Figura 24: Localização histológica do posicionamento dos canais selecionados para
cada animal...................................................................................................................73
Figura 25: Tempo relativo de exploração de objetos idênticos durante a sessão 1 da
tarefa de reconhecimento de objetos, em condição controle (A) e após privação de
sono por 72 h (B)...........................................................................................................76
Figura 26: Tempo total de exploração dos objetos durante a sessão 1 da tarefa em
condição controle (CT) e após privação de sono por 72 h (PS)....................................76
Figura 27: Tempo relativo de exploração de objetos durante a sessão 2 da tarefa de
reconhecimento de objetos, em condição controle (A) e após privação de sono por 72
h (B)..............................................................................................................................77
Figura 28: Tempo total de exploração dos objetos durante a sessão 2 da tarefa em
condição controle (CT) e após privação de sono por 72 horas (PS).............................77
Figura 29: Efeitos da privação de sono no ciclo sono-vigília durante intervalo de 4
horas entre sessões da tarefa de reconhecimento de objetos......................................79
Figura 30: Efeitos da privação de sono em parâmetros específicos dos estados de
sono. A: número de episódios de sono paradoxal (SP); B: duração média de episódio
de SP; C: número de episódios de sono de ondas lentas (SOL); D: duração média de
episódio de SOL............................................................................................................80
Figura 31: Espectro de potência normalizado do registro hipocampal durante as duas
sessões da tarefa de reconhecimento de objetos.........................................................82
Figura 32: Efeitos da privação de sono sobre potência normalizada de teta total (4-12
Hz, A), teta 1 (8-12 Hz, B), e teta 2 (4-8 Hz, C).............................................................83
Figura 33: Pico de frequência de ritmo teta hipocampal nas condições controle (CT) e
privação de sono por 72 horas (PS) na tarefa de reconhecimento de objetos.............84

xxi
1.3. LISTA DE TABELAS
Tabela 1: Marcação de [3H]AMPA após 96 horas de privação de sono e recuperação
de 24 horas....................................................................................................................59
Tabela 2: Hibridização in situ da subunidade GluR1 após 96 horas de privação de
sono e recuperação de 24 horas...................................................................................60

1
2. INTRODUÇÃO
2.1. Perda de Sono e de Memória
O sono é um estado recorrente e dinâmico que compõe, em conjunto
com a vigília, a base do ritmo circadiano dos animais, representado pelo ciclo
sono-vigília (Marques e Menna Barreto, 2003). Com a descoberta das
oscilações elétricas cerebrais, descritas pela primeira vez por Hans Berger em
humanos (1929), o ciclo sono-vigília em mamíferos se divide três fases
principais, definidas por características eletrofisiológicos únicas (Siegel, 2008):
a vigília, com predomínio de dessincronia de ondas corticais de baixa amplitude
e alta frequência; o sono de ondas lentas, com predomínio de ondas de baixa
freqüência e alta amplitude, e uma fase de sono em que há ondas
dessincronizadas similares às da vigília, mas com atonia muscular e baixa
responsividade sensorial: o sono REM (rapid eye movement), descrito
primeiramente em humanos por Aserinsky e Kleitman (1953), também
chamado de sono dessincronizado (Timo-Iaria et al., 1970) e sono paradoxal
(Jouvet, 1969). Essa última denominação será usada no presente trabalho.
Vários autores sugerem que o sono representa um estado fisiológico
importante para o processamento eficiente das informações adquiridas durante
a vigília. As evidências experimentais e clínicas incluem a observação de
melhora de desempenho cognitivo após períodos de sono em humanos
(Mednick et al. 2002), aumento de sono paradoxal após tarefas de
aprendizagem em roedores (Smith e Lapp 1991; Smith e Rose 1997),

2
reverberação de atividade neuronal da vigília e aumento de oscilações lentas
durante o sono de ondas lentas (Ribeiro e Nicolelis 2004), e aumento de
expressão de genes de fatores neurotróficos durante o sono paradoxal (Ribeiro
et al. 1999; Ribeiro et al. 2002).
A perda de sono é um problema cada vez mais presente na sociedade
atual. Segundo levantamento realizado pela National Sleep Foundation (2002),
27% dos adultos americanos tem dificuldades para dormir e 16% apresentam
sonolência com consequências significativas em suas atividades diárias. Um
estudo recente de nosso laboratório levantou que, dentro de um conjunto de
mais de dois mil brasileiros distribuídos em 150 cidades pelo país, 63% destes
sofrem de algum distúrbio de sono (Bittencourt et al., 2009). Especificamente
na cidade de São Paulo, um segundo estudo de nosso laboratório, ao
comparar três levantamentos de queixas de sono obtidos nas últimas décadas,
demonstrou que a prevalência de sonolência diurna aumentou
consideravelmente (Santos-Silva et al., 2010). Sintomas clínicos comuns
associados à perda de sono incluem humor degradado, sonolência e prejuízos
cognitivos (Bonnet e Arand 2003). Esta última relação – perda de sono e
prejuízos cognitivos, especificamente, piora de memória – é o tema central da
presente tese.
A memória não é uma entidade única, podendo, em humanos, ser
classificada em dois tipos principais – memória declarativa e não declarativa –
conforme as áreas cerebrais envolvidas no processo mnemônico (Squire,
1992). A memória declarativa é descrita como a habilidade em reconhecer e
recordar fatos de forma consciente e está relacionada a estruturas do lobo
temporal medial, como a formação hipocampal (Squire & Zola-Morgan, 1991;

3
Eichenbaum, 2000). Por sua vez, a memória não declarativa refere-se à
capacidade de desenvolver habilidades percepto-motoras e cognitivas após
exposições repetidas e está associada a estruturas cortico-estriatais e
cerebelares (Squire & Zola-Morgan, 1991).
De forma similar à divisão de tipos de memória em humanos, é possível
obter modelos animais putativos de memórias declarativas e não declarativas,
com base no aprendizado de tarefas que requerem o envolvimento de
diferentes regiões do sistema nervoso central (Izquierdo e Medina, 1997). Por
exemplo, o hipocampo, uma estrutura essencial para a memória declarativa em
humanos, apresenta uma importante função no processamento de informações
espaciais e contextuais de animais, mas não está relacionado a tarefas que
não dependem deste tipo de informação (O’Keefe, 1990; Kim & Fanselow,
1992; Phillips & LeDoux e col., 1992).
Estudos sobre a influência da falta de sono na memória têm gerado
enfoques diversos, aos quais estão relacionados o momento do impedimento
do sono em relação à aprendizagem, o tipo e a complexidade da tarefa de
memória (Walker e Stickgold, 2006). Muitas evidências sugerem que a privação
de sono (PS) prévia à aprendizagem de novas informações prejudica a
memória do tipo declarativa. Em humanos, um dos primeiros trabalhos sobre o
tema foi realizado por Morris e colaboradores (1960), demonstrando que a
memória declarativa temporal (memória de eventos localizados no tempo)
estava prejudicada após uma noite de privação total de sono antes da
aprendizagem. Essas observações foram reavaliadas em um trabalho posterior
realizado por Harrison e Horne (2000), que observaram um prejuízo similar na
discriminação temporal de fotografias em voluntários privados de sono. Um

4
estudo de neuroimagem funcional avaliou os efeitos de 35 horas de PS na
atividade metabólica de várias áreas cerebrais enquanto os voluntários
aprendiam uma tarefa de memória declarativa verbal, no que se verificou
prejuízo significativo na retenção da tarefa associado a redução de atividade do
lobo temporal medial (Drummond et al. 2000). Em consonância a esse estudo,
Walker e Stickgold (2006) demonstraram que a PS antes da aprendizagem de
palavras neutras e emocionais prejudica o reconhecimento posterior, com
redução expressiva na retenção de palavras neutras e emocionalmente
positivas, ao passo que não há alteração na retenção de palavras negativas.
Em animais não humanos, há um volume considerável de trabalhos
relacionando os efeitos da perda de sono antes da aprendizagem (para
revisão: Pearlman 1979, Smith 1995, Walker e Stickgold 2006). Animais
privados de sono antes de aprenderem uma tarefa apresentam prejuízos de
memórias dependentes do hipocampo. Dessa forma, a privação de sono em
animais induz piora de desempenho em diversas tarefas que avaliam o
componente espacial ou contextual do processo de aprendizagem, como a
versão espacial do labirinto aquático de Morris (Youngblood et al. 1999,
Youngblood et al. 1997), a esquiva inibitória (Moreira et al., 2003, 2010; Dubiela
et al, 2005, 2010) e o condicionamento de medo ao contexto (Bueno et al.,
1994, McDermott et al., 2003; Ruskin et al. 2004). Tarefas que não dependem
do hipocampo, e classicamente não dependem de elementos espaciais ou
contextuais para sua execução, como o condicionamento de medo ao som
(O’Keefe, 1990), geralmente não são afetadas pela privação de sono (Bueno et
al. 1994, McDermott et al. 2003, Ruskin et al. 2004), embora Dametto et al.
(2002) tenham encontrado prejuízo no condicionamento de medo ao som em

5
ratos privados de sono. Nesses estudos, a privação de sono é realizada de 72
a 96 horas (3 a 4 dias), com o impedimento total da fase de sono paradoxal,
embora esta seja acompanhada por uma redução significativa do sono de
ondas lentas (Machado et al. 2004). Ainda, foi verificado que a privação de
sono por períodos mais curtos (por exemplo, 24 horas) não interfere
sobremaneira no desempenho da tarefa de esquiva inibitória (Bueno et al.,
1994), o que sugere uma relação análoga com os resultados obtidos em
humanos pelo estudo de Walker e Stickgold (2006) quanto a resistência de
traços de memórias aversivas aos efeitos da PS.
Especificamente em nosso laboratório, uma série de estudos
demonstrou que ratos privados de sono por 96 horas antes da realização da
tarefa de esquiva inibitória apresentam prejuízos de desempenho, verificados
tanto em intervalos de retenção curtos (30 minutos; Dubiela et al. 2005, 2010),
como em intervalos longos (24 horas; Bueno et al. 1994, Dametto et al. 2002,
Moreira et al. 2003, Dubiela et al., 2010) após o treino da tarefa. Ainda, ratos
submetidos a PS por 4 dias apresentam um aumento expressivo de sono
paradoxal nas primeiras horas de repouso (efeito rebote), que se normaliza
após 24 horas (Machado et al. 2004), e observamos que este mesmo período
de recuperação promove atenuação do prejuízo da PS sobre a tarefa de
esquiva inibitória (Dubiela et al. 2010, Fig 1). Dessa forma, é plausível supor
que os efeitos deletérios da privação de sono sobre os mecanismos neurais
relacionados com a formação de memórias estão vinculados à pressão
homeostática de sono, caracterizada pelo efeito rebote (Machado et al. 2004).
Por conseguinte, as modificações induzidas pela PS em tais mecanismos
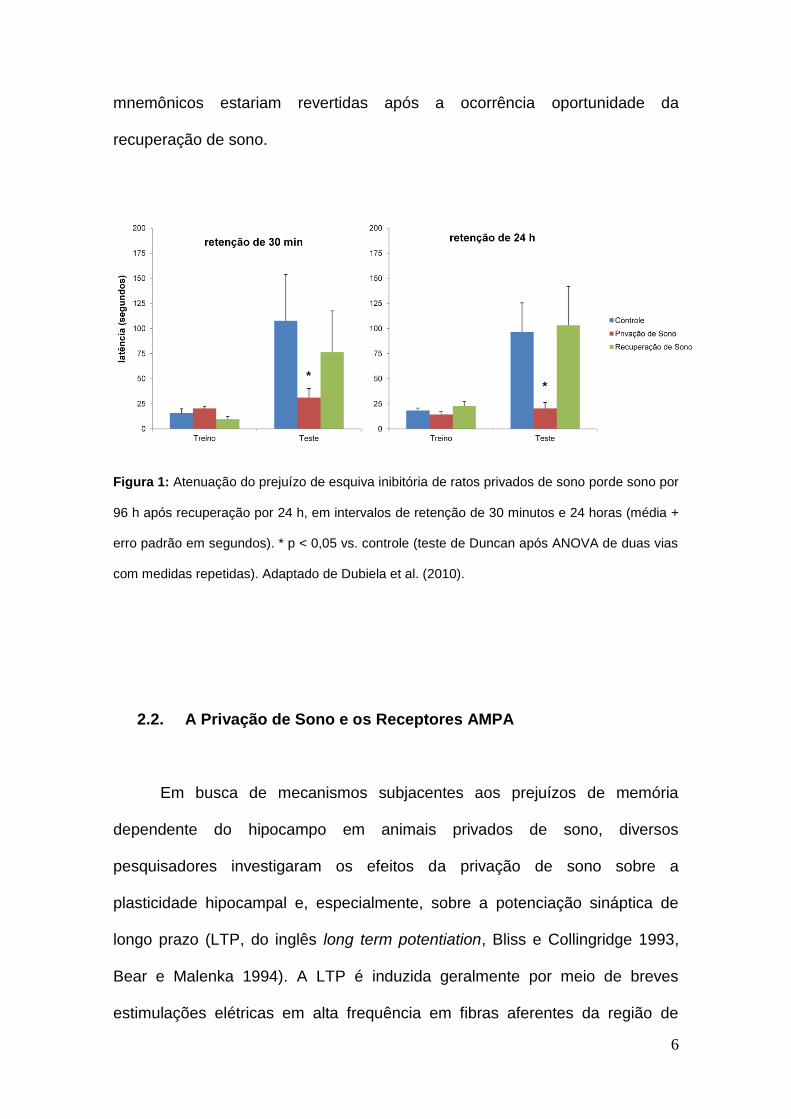
6
mnemônicos estariam revertidas após a ocorrência oportunidade da
recuperação de sono.
Figura 1: Atenuação do prejuízo de esquiva inibitória de ratos privados de sono porde sono por
96 h após recuperação por 24 h, em intervalos de retenção de 30 minutos e 24 horas (média +
erro padrão em segundos). * p < 0,05 vs. controle (teste de Duncan após ANOVA de duas vias
com medidas repetidas). Adaptado de Dubiela et al. (2010).
2.2. A Privação de Sono e os Receptores AMPA
Em busca de mecanismos subjacentes aos prejuízos de memória
dependente do hipocampo em animais privados de sono, diversos
pesquisadores investigaram os efeitos da privação de sono sobre a
plasticidade hipocampal e, especialmente, sobre a potenciação sináptica de
longo prazo (LTP, do inglês long term potentiation, Bliss e Collingridge 1993,
Bear e Malenka 1994). A LTP é induzida geralmente por meio de breves
estimulações elétricas em alta frequência em fibras aferentes da região de

7
interesse, produzindo aumento da responsividade do neurônio pós sináptico
(Bliss e Collingridge 1993, Bear e Malenka 1994). Desde sua descoberta, a
LTP hipocampal é considerada um modelo eletrofisiológico putativo de
formação de memórias, embora grande parte das evidências que a suporte
sejam indiretas, tais como intervenções farmacológicas que prejudicam tanto a
aprendizagem de tarefas hipocampo dependentes como a indução de LTP
(Albensi et al. 2007). Por outro lado, uma evidência direta foi demonstrada no
estudo de Whitlock et al. (2006), no qual foi observado que a aprendizagem da
tarefa de esquiva inibitória induz mudanças sinápticas idênticas às geradas
pela LTP na região CA1, como aumento de potencial de resposta evocada e
ativação de receptores AMPA
Um dos primeiros trabalhos a avaliar os efeitos da PS sobre a
plasticidade hipocampal foi realizado por Campbell e colaboradores (2002), que
observou redução da LTP na região do CA1 de fatias hipocampais de ratos
privados de sono por 12 horas. McDermott et al. (2003) expandiram esses
dados, ao mostrar que, em preparações in vitro, tanto a LTP da região CA1
quanto do giro denteado estavam inibidas após PS de 72 horas de duração. À
semelhança de nossos dados na tarefa de esquiva inibitória (Fig 1), McDermott
et al. (2003) também demonstraram que a recuperação de sono por 24 horas
após PS reverteu a redução da LTP hipocampal.
Um fator comum entre a formação de memórias hipocampo
dependentes e a LTP hipocampal é a participação do sistema glutamatérgico,
responsável pela maior parte da transmissão excitatória do sistema nervoso
central e, por conseguinte, envolvido em diversos processos de plasticidade
cerebral (Izquierdo et al., 1993; Kandel et al., 2000). As evidências

8
neurofarmacológicas da participação desse sistema em processos
mnemônicos e eletrofisiológicos são obtidas por meio do estudo de seus dois
principais receptores ionotrópicos: os receptores de tipo NMDA e do tipo AMPA.
A privação de sono aumenta os níveis de glutamato extracelular no
córtex (Bettendorff et al., 1996; Mohammed et al., 2011) e no hipocampo
(Mohammed et al., 2011) de animais de laboratório, o que pode estar
associado a alterações em receptores do sistema glutamatérgico. Nesse
sentido, estudos dos efeitos da PS sobre os receptores do tipo NMDA, por
exemplo, demonstram redução na expressão de suas subunidades NR1
(McDermott et al., 2006, Ravassard et al., 2009) e NR2A (McDermott et al.,
2006) no hipocampo de ratos após 72 horas de privação de sono.
Complementarmente, dados de nosso laboratório mostram que ratos privados
de sono por 96 horas apresentam uma resposta farmacológica exacerbada aos
efeitos locomotores de MK-801, um antagonista não-competitivo de receptores
NMDA (Dubiela et al., 2011). Esses dados tomados em conjunto, reforçam a
concepção de que há uma redução da função desses receptores induzida pela
privação de sono.
Por outro lado, evidências sobre a influência da perda de sono sobre a
expressão e funcionalidade dos receptores do tipo AMPA são contraditórias. De
acordo com o estudo apresentado por Vyazovskiy et al. (2008), há aumento da
expressão da subunidade GluR1 no córtex e no hipocampo de ratos privados
totalmente de sono por 4 horas, enquanto que Hagewoud et al. (2009) não
encontraram modificações nos níveis de expressão da mesma subunidade em
camundongos submetidos a privação total de sono por 6 ou 12 horas. Por sua
vez, McDermott et al. (2006) indicam não haver alterações nas correntes

9
iônicas que fluem através do canal de receptores AMPA no hipocampo de ratos
privados de sono por 72 horas, ao passo que Ravassard et al. (2009)
revelaram níveis reduzidos da subunidade GluR1 no hipocampo de ratos
privados pelo mesmo período. Dentro deste contexto, é plausível que uma
análise abrangente em regiões cerebrais envolvidas com a formação de
memórias, por meio da autorradiografia de receptores, possa contribuir para
melhor elucidar essa questão.
A abordagem neuroquímica dos efeitos da privação de sono sobre os
receptores AMPA fornece uma interessante base de investigação, uma vez que
a marcação desses receptores encontra-se aumentada no hipocampo após a
indução de LTP (Maren et al. 1993). De forma similar, há aumento de
marcação de receptores AMPA hipocampais após aprendizagem de tarefas
hipocampo dependentes, como o condicionamento de medo ao contexto
(Tocco et al. 1992) e a esquiva inibitória (Cammarota et al. 1995, 1996; Sadad
et al., 2006). Ademais, a aplicação intra-hipocampal de CNQX, um antagonista
de receptores AMPA, prejudica a formação da memória na tarefa de esquiva
inibitória (Bonini et al. 2003), o que sugere um possível mecanismo que associe
a diminuição da função dos receptores AMPA e os prejuízos de memória
observados após a privação de sono.
Os receptores AMPA são estruturas multiméricas formadas pela
combinação de 4 a 5 subunidades proteicas, classificadas em quatro tipos:
GluR1, GluR2, GluR3 e GluR4 (Monaghan e Wenthold 1997). A identificação
dessa diversidade é crucial devido às características funcionais que cada
subunidade confere ao receptor: homômeros de GluR1 são recrutados do
citosol para o terminal pós sináptico quando há aumento de atividade neuronal

10
no hipocampo (Hayashi et al., 2000), e a presença da subunidade GluR2 inibe
a condutância de cálcio pelo canal iônico (Monaghan e Wenthold, 1997).
No hipocampo, a maior parte dos receptores AMPA são heterômeros
compostos pelas subunidades GluR1-GluR2 e GluR2-GluR3 (Shi et al., 2001).
A redução de receptores AMPA hipocampais é uma característica presente em
algumas patologias neurais associadas a prejuízos de memória, como o mal de
Alzheimer e a hipóxia isquêmica (Westerberg et al. 1987, Dewar et al. 1991,
Sommer et al. 2002, Montori et al., 2010). A diminuição do número de
receptores no mal de Alzheimer parece estar associada a uma redução da
transcrição da subunidade GluR1 (Pellegrini-Giampietro et al. 1994, Dickey et
al. 2003), enquanto que na isquemia cerebral observa-se uma diminuição nos
níveis de RNAm e de proteína das subunidades GluR1 e GluR2 na formação
hipocampal (Montori et al., 2010). Entretanto, um estudo recente demonstrou
que não há modificação da transcrição da subunidade GluR1 na formação
hipocampal de ratos privados de sono por 72 h (Ravassard et al., 2009). Dessa
forma, a caracterização da transcrição da subunidade GluR1 dos receptores
AMPA pode auxiliar a esclarecer mecanismos fisiológicos envolvidos na
modulação da plasticidade hipocampal após a privação e recuperação de sono.

11
2.3. Modulação Farmacológica De Receptores AMPA
Nas últimas décadas um grande esforço foi feito na busca por fármacos
que tenham a propriedade de melhorar a memória, tanto em situações
patológicas como normais. Muitos desses fármacos tem como mecanismo de
ação a facilitação da neurotransmissão glutamatérgica (O'Neill et al. 2004),
porém sem produzir efeitos indesejáveis decorrentes de estimulação intensa
e/ou sustentada, tais como convulsão e atividade paroxística (Yamashita et al.,
2004; Fan et al., 2011). Esta abordagem resultou no desenvolvimento de
potenciadores dos receptores AMPA, moduladores alostéricos positivos cujo
mecanismo de ação envolve o retardo da dessensibilização desses receptores
após sua ativação (Jin et al. 2005). Alguns exemplos incluem as pirrolidinonas
(piracetam, aniracetam) e, mais recentemente, as ampaquinas (CX614, CX717)
(O'Neill et al. 2004). A eficácia dessas drogas foi demonstrada com o uso de
modelos animais, onde os prejuízos de memória decorrentes de lesão cerebral,
envelhecimento, administração de fármacos amnésicos foram atenuados ou
mesmo prevenidos, seja com administração pré ou imediatamente após a
aprendizagem (Cumin et al., 1982; Gouliaev e Senning, 1994; Bartolini et al.,
1996).
Estudos recentes mostram que os potenciadores de receptores AMPA
podem contribuir para a prevenção de prejuízos cognitivos decorrentes da
perda de sono. Porrino et al. (2005) demonstraram que a ampaquina CX717
produz melhora de desempenho em primatas privados de sono e treinados
numa tarefa de memória dependente de hipocampo. Nesse trabalho, macacos-

12
rhesus foram treinados diariamente numa tarefa de reconhecimento espacial
com atraso (delayed match to sample task), na qual precisavam identificar,
numa tela de computador, uma imagem específica dentro de um conjunto, que
havia sido apresentada previamente à tentativa de identificação (1 a 30
segundos de intervalo). Em algumas sessões, os animais foram privados
totalmente de sono por 36 horas antes de começar a tarefa. Nesses casos, os
animais apresentaram prejuízos significativos de desempenho na tarefa, tais
como aumento da quantidade de erros e aumento na latência para resposta
(Porrino et al., 2005). Tais prejuízos foram atenuados quando os animais
receberam uma dose de CX717 (0,8 mg/kg) minutos antes do início da sessão
de teste. Um estudo similar, realizado em humanos privados de sono por 27
horas, mostrou que uma alta dose de CX717 (1000 mg) foi capaz de prevenir
os prejuízos de atenção induzidos pela privação de sono total por 24 horas
(Boyle et al., 2011).
Paralelamente, outra linha de evidências propõe que o bloqueio dos
receptores AMPA, em situações patológicas caracterizadas pelo aumento
excessivo de glutamato liberado, pode atenuar prejuízos de memória (Ruel et
al. 2002). Especificamente, antagonistas não competitivos seletivos de
receptores AMPA, como os 2,3-benzodiazepínicos (GYKI 52466, GYKI 53405) ,
foram avaliados em estudos de isquemia cerebral, nos quais se verificou
função neuroprotetora desses fármacos (Elger et al. 2005Block e Schwarz
1997, Gyertyan et al. 1999). Nesses dois últimos estudos, foi constatado que o
tratamento agudo com GYKI 52466, durante a intervenção isquêmica em
roedores, preveniu prejuízos de memória observados posteriormente, avaliados

13
na versão espacial do labirinto aquático de Morris (Block e Schwarz 1997) e na
tarefa de alternação espontânea (Gyertyan et al. 1999).
Consistente com mecanismos farmacológicos compensatórios entre
disponibilidade de ligante e receptor, a marcação cerebral de receptores AMPA
pode ser reduzida (downregulation) por meio de aumento excessivo de níveis
de glutamato (Montori et al., 2010) ou estimulação contínua desses receptores
por AMPA (Sinha et al., 2004). Por outro lado, o bloqueio crônico por meio de
antagonistas promove aumento da marcação dos receptores AMPA
(upregulation): Sinha et al. (1999) demonstraram que o tratamento crônico (6
dias) com o antagonista GYKI 52466 promove aumento no número de
receptores AMPA, tanto no córtex como no hipocampo de ratos. Tais
resultados sugerem um uso promissor como terapia em patologias associadas
a redução da atividade dos receptores AMPA.
Em suma, a pesquisa de fármacos que atuam sobre os receptores
AMPA sugere que tanto a administração de potenciadores quanto antagonistas
desses receptores possam atenuar os efeitos deletérios da PS em tarefas de
memória dependentes de hipocampo, seja por meio da modulação intrínseca
da função dos receptores, seja pela interferência sobre os níveis de receptores
nas estruturas cerebrais.

14
2.4. Receptores AMPA e o Ritmo Teta Hipocampal
As oscilações elétricas cerebrais foram descritas pela primeira vez por
Hans Berger (1929), ao descobrir sinais periódicos em registros de
eletroencefalograma (EEG) de humanos. Estas oscilações são produzidas
primariamente pela atividade síncrona de populações de neurônios, por meio
da soma resultante de potenciais pós-sinápticos inibitórios e excitatórios. Estes
últimos são gerados, em grande parte, por correntes iônicas dos receptores
AMPA, responsáveis pela despolarização rápida de terminais pós sinápticos
(Platt, 2007).
Os registros eletroencefalográficos podem ser convertidos do domínio do
tempo em que ocorrem ao domínio da frequência e, por meio de análise de
espectro de potência, é possível verificar a contribuição de cada componente
de frequência para o sinal obtido. Essa ferramenta é especialmente útil para
análise de sono, haja vista a ocorrência de faixas de frequências características
em diferentes estados do ciclo sono-vigília em roedores (Timo Iaria, 1970), tais
como o ritmo delta no sono de ondas lentas e o ritmo teta no sono paradoxal e
vigília.
O ritmo teta hipocampal é uma oscilação de grande amplitude, na faixa
de frequência de 4 a 12 Hz, observada na formação hipocampal de roedores
(Buzsaki, 2002). Esse ritmo pode ser verificado em outras espécies animais,
inclusive humanos (Bodizs et al., 2001), embora a compreensão de suas
características não esteja tão consolidada como em ratos (Lega et al., 2011). A
atividade de diferentes células hipocampais muda de acordo com a presença

15
do ritmo teta: a taxa de disparos de neurônios glutamatérgicos diminui, ao
passo que a maioria dos interneurônios GABAérgicos aumenta ou não altera
sua taxa de disparos (Buzsaki, 1983). Além disso, foi observado que a
atividade de neurônios glutamatérgicos e interneurônios pode estar em fase,
resultando em um padrão em que diferentes células disparam em sincronia
com outras durante fases específicas do ciclo (Csicsvari et al., 1999).
Classicamente, considera-se que o sítio da geração do marca passo do ritmo
teta é o núcleo septal medial da banda de Broca, ao passo que as camadas
stratum oriens de CA1 e a camada molecular do giro denteado compõem os
sítios geradores da corrente, embora não haja consenso conclusivo a respeito
(Buzsaki, 2002).
Vários autores buscam estabelecer uma relação entre comportamentos
e oscilações cerebrais. Devido a sua consistência, reproducibilidade e forte
relação com atividade exploratória, o ritmo teta hipocampal vem sendo utilizado
como modelo experimental de aprendizagem. Em ratos, verificou-se que a
potência do ritmo teta aumenta durante comportamentos preparatórios, como
movimentos voluntários, orientação espacial, exploração do ambiente, e
durante o sono paradoxal (Vanderwolf, 1969; Timo-Iaria, 1970). Por outro lado,
o ritmo teta está ausente quando o animal está imóvel, se alimentando ou
executando auto-limpeza (grooming), considerados como comportamentos
consumatórios (Vanderwolf, 1969).
O ritmo teta pode apresentar características diferentes de acordo com as
condições em que é registrado. Assim, dois tipos de ritmo teta foram descritos
com base na sensibilidade farmacológica ao antagonista colinérgico
muscarínico atropina (Kramis et al., 1975): o ritmo teta tipo 1, composto por

16
oscilações na faixa de 8 a 12 Hz, geralmente é observado durante atividade
motora em ratos conscientes, não é afetado pela atropina, mas pode ser
bloqueado pela ação de antagonistas de receptores NMDA (Buzsaki et al.,
2002); por sua vez, oscilações na faixa de 4 a 8 Hz, consideradas como
pertencentes ao ritmo teta tipo 2, são bloqueadas por antagonistas
muscarínicos, e podem ser melhor visualizadas em ratos anestesiados ou em
fatias hipocampais tratadas com carbacol. O ritmo teta tipo 2 tem sido
associado com a aprendizagem de tarefas de memória, ao passo que o ritmo
teta tipo 1 está mais relacionado a locomoção (Macrides et al., 1982; Berry e
Swain, 1989; Seidenbecher et al., 2003).
Além dos receptores muscarínicos e NMDA, os receptores AMPA
também representam um importante elemento na modulação do ritmo teta
hipocampal. A administração de antagonistas de receptores AMPA no núcleo
septal medial, proposto como o gerador de frequência do ritmo teta (Buzsaki,
2002), induz redução da amplitude e da frequência do ritmo teta tipo 2 em ratos
anestesiados (Puma e Bizot, 1999). Além disso, a aplicação de antagonistas
destes receptores diretamente na região hipocampal diminui a potência do
ritmo teta tipo 2 em ratos não anestesiados (Leung e Shen, 2004). Por outro
lado, a aplicação local de aniracetam no hipocampo de ratos preveniu a
redução do ritmo teta hipocampal induzido por um antagonista histaminérgico
(Masuoka et al., 2007). Nesse sentido, é plausível supor que alterações na
função dos receptores AMPA, sobretudo na região hipocampal, estejam
acompanhadas por modificações das oscilações teta.
O ritmo teta, e em especial o teta tipo 2, tem gerado grande interesse em
pesquisadores de memória, em função de muitos comportamentos associados

17
a este ritmo estarem envolvidos com processamento cognitivo. Por exemplo, foi
demonstrado que há aumento da intensidade de ritmo teta tipo 2 durante a
aprendizagem de tarefas de memória espacial (Yang et al., 2011) e
condicionamento de medo ao contexto e ao som (Pape et al., 2005). Ademais,
manipulações farmacológicas que bloqueiam ou facilitam o ritmo teta tipo 2
produzem piora ou melhora de desempenho em tarefas de memória hipocampo
dependentes, como a alternação espacial forçada (Givens e Olton, 1995) e a
versão espacial do labirinto aquático de Morris (Pan e McNaughton, 1997).
Consistente com a hipótese do envolvimento do ritmo teta tipo 2 na
formação de memórias, verificou-se que esta faixa de frequência está
associada com a plasticidade sináptica no hipocampo: a estímulação das vias
colaterais de Schaffer por meio de rajadas em pulsos de frequência de 5 Hz,
chamada estimulação de rajadas em teta (theta burst stimulation), promove
LTP hipocampal (Greenstein et al., 1988; Larson et al., 1986), resultando em
aumento da responsividade do neurônio pós sinápticos da região CA1. Por
outro lado, pulsos de rajadas realizadas em frequências mais altas, dentro da
faixa do teta tipo 1 (10 Hz), ou mais baixas, dentro da faixa do ritmo delta (2
Hz), não produzem o mesmo efeito de potenciação (Greenstein et al., 1988;
Larson et al., 1986).
Estudos de privação de sono em animais têm demonstrado efeitos
variados sobre o ritmo teta hipocampal observado durante a vigília. Vyazovskiy
e Tobler (2005) demonstraram que a potência do ritmo teta tipo 2 aumenta em
ratos privados totalmente de sono por 6 horas, ao passo que Mohammed et al.
(2011) encontraram uma diminuição dos ritmos teta tipo 1 e 2 após uma
privação de sono por 24 h, mas não por 72h. Utilizando o mesmo paradigma,

18
Corsi-Cabrera et al. (1994) encontraram um aumento da potência do ritmo teta
tipo 1, mas não do tipo 2, em ratos submetidos à privação de sono por 72
horas, ao qual atribuíram um aumento da excitabilidade locomotora do animal
privado de sono (Van Hulzen et al., 1984). Com relação à aprendizagem de
tarefas de memória, há apenas um estudo que investigou a combinação dos
efeitos da privação de sono sobre uma tarefa de memória e o ritmo teta
hipocampal (Yang et al, 2011). Neste trabalho, foi verificado que ratos privados
de sono paradoxal por sete dias e treinados, concomitantemente, numa tarefa
versão espacial do labirinto aquático de Morris, apresentam prejuízos de
desempenho a partir do terceiro dia (72 horas) de privação, juntamente com
prevenção de aumento da potência de ritmo teta tipo 2 induzida pela
aprendizagem da tarefa (Yang et al, 2011). Este conjunto de evidências sugere
a importância de avaliar os dois tipos de ritmo teta após a privação de sono,
sobretudo durante uma tarefa de memória, no intuito de verificar a relação
entre a atividade da formação hipocampal e o desempenho cognitivo de
animais privados de sono.
2.5. Justificativa
Atualmente, não é possível definir com clareza quais são os
mecanismos neurais relacionados aos prejuízos de memória induzidos pela
perda de sono, e quais representam uma reação compensatória a essa
exposição. Um possível candidato para estudo são os receptores
glutamatérgicos ionotrópicos do tipo AMPA, envolvidos com a formação de

19
memórias e plasticidade hipocampal. Dados da literatura demonstram
variabilidade de efeitos induzidos pela privação de sono sobre estes receptores
tanto em regiões corticais como hipocampais (Vyazovskiy et al., 2008,
Ravassard et al., 2009), ao passo que outro corpo de evidências indica
aumento de transmissão glutamatérgica generalizada em ratos privados de
sono (Bettendorff et al, 1996; Mohammed et al., 2011). Para melhor elucidar
esta questão, a avaliação autorradiográfica pode oferecer uma visão ampla da
distribuição, localização e quantificação de sítios do receptor AMPA em regiões
cerebrais envolvidas com processos mnemônicos em animais privados de
sono, ao passo que a análise de expressão gênica da subunidade GluR1 pode
incrementar a compreensão de mecanismos relacionados à alteração desses
receptores.
Em paralelo, estudos farmacológicos mostram que potenciadores de
receptores AMPA são capazes de atenuar prejuízos cognitivos em diversas
condições (Gouliaev & Senning, 1994), inclusive privação de sono total em
primatas humanos (Boyle et al., 2011) e não-humanos (Porrino et al. 2005).
Outras evidências demonstram que o tratamento crônico com o antagonista
seletivo GYKI 52466 induz aumento de receptores AMPA no hipocampo (Sinha
et al. 1999), e previne prejuízos de memória espacial, quando administrado de
forma aguda, num modelo animal de isquemia hipocampal (Block e Schwarz,
1997; Gyertyan et al. 1999), uma condição que está associada tanto a níveis
aumentados de glutamato como a redução de receptores AMPA hipocampais
(Montori et al., 2010). Portanto, a modulação farmacológica desses receptores
por potenciadores e/ou antagonistas poderia constituir uma estratégia eficaz

20
para a redução do prejuízo de desempenho de esquiva inibitória em ratos
privados de sono em nosso laboratório.
A alteração da função de receptores AMPA após a perda de sono pode
exercer influência sobre a atividade do hipocampo, e especialmente sobre o
ritmo teta hipocampal. O bloqueio de receptores AMPA por antagonistas reduz
a potência do ritmo teta tipo 2 (Leung e Shen, 2004), que está associado com a
aprendizagem de tarefas de memória (Givens e Olton, 1995). Por outro lado,
estudos de privação de sono apresentam resultados variados sobre o ritmo teta
hipocampal (Corsi-Cabrera et al., 1994; Vyazovskiy e Tobler, 2005;
Mohammed et al., 2011), e há poucos dados relativos à correlação dos efeitos
da perda de sono sobre uma tarefa de memória e o ritmo teta tipo 2 (Yang et al,
2011). Nesse sentido, ao considerar o envolvimento dos receptores AMPA
nesse fenômeno (Puma e Bizot, 1999; Leung e Shen, 2004), apresentamos a
hipótese de que o ritmo teta hipocampal tipo 2 está reduzido em ratos privados
de sono durante a aprendizagem de uma tarefa hipocampo dependente.
3. OBJETIVOS
O principal objetivo da presente tese é avaliar o papel dos receptores AMPA
na modulação dos prejuízos de memória dependente de hipocampo após a
privação de sono, bem como investigar possíveis efeitos decorrentes da
alteração da atividade dos receptores AMPA na formação hipocampal. Para
tanto, esta tese está dividida em duas etapas, com os seguintes objetivos
específicos:

21
PRIMEIRA ETAPA
Analisar a marcação de receptores AMPA e a expressão da subunidade
GluR1 no cérebro de ratos privados e recuperados de sono;
Verificar os efeitos da administração do potenciador aniracetam e do
antagonista GYKI 52466 sobre os prejuízos de memória decorrentes da
privação de sono.
SEGUNDA ETAPA
Caracterizar a atividade do ritmo teta hipocampal em ratos privados de
sono durante o desempenho de uma tarefa hipocampo dependente.

22
4. PRIMEIRA ETAPA: AVALIAÇÃO NEUROQUÍMICA E
FARMACOLÓGICA DOS RECEPTORES AMPA NA
FORMAÇÃO DE MEMÓRIAS EM RATOS
PRIVADOS DE SONO
4.1. Experimento 1: Efeitos da Privação e Recuperação de Sono na
Marcação De [3H]AMPA e Hibridização in situ de Glur1
O experimento teve como objetivo avaliar a distribuição, localização e
quantificação de sítios do receptor AMPA em regiões cerebrais envolvidas com
processos mnemônicos em animais privados de sono, bem como realizar a
análise de expressão gênica da subunidade GluR1 em regiões cuja marcação
de receptores AMPA está modificada
4.1.1. Experimento 1-1: Autorradiografia de Receptores AMPA após
Privação e Recuperação de Sono
Considerando os efeitos da privação de sono sobre a transmissão
glutamatérgica (Bettendorff et al., 1996; Mohammed et al., 2011), bem como a

23
modulação atividade-dependente de receptores AMPA (Sinha et al., 1999,
2004), é plausível supor que a perda de sono modifique a distribuição de
receptores AMPA. Dessa forma, o objetivo específico desse experimento foi
avaliar se há alterações sobre a marcação de [3H]AMPA em ratos submetidos à
privação e recuperação de sono, através da técnica de autorradiografia de
receptores, com ênfase em áreas cerebrais envolvidas na modulação da
memória.
Sujeitos
Foram utilizados ratos Wistar, machos, com três meses de idade,
provenientes do biotério do Departamento de Psicobiologia da Universidade
Federal de São Paulo. Os animais foram mantidos em condições controladas
de temperatura (23 2 ºC) e ciclo claro/escuro de 12:12 h, com início do
período claro às 7:00h. Água e comida foram fornecidas ad libitum. Cada
experimento foi realizado com diferentes conjuntos de animais.
Procedimento de Privação de Sono
Uma das primeiras técnicas amplamente utilizadas para indução de PS é
a técnica da plataforma invertida, proposta por Jouvet et al. (1964), utilizando
gatos, e adaptada para ratos por Cohen e Dement (1965). A técnica é
estruturada a partir de uma plataforma estreita circundada por água. Devido à
perda gradual da tonicidade muscular ao longo do sono até sua fase paradoxal,
em que ocorre a atonia muscular, o animal perde o equilíbrio postural e cai da
plataforma, acordando na água (Jouvet et al., 1964). Com a utilização desse
procedimento, além da perda completa do sono paradoxal, há uma perda

24
parcial do sono de ondas lentas (Grahnstedt e Ursin, 1985, Machado et al.,
2004).
A técnica de plataforma invertida tem sofrido críticas devido à presença
de variáveis intervenientes, como a imobilização do animal, embebição do pêlo
na água, isolamento social e outros fatores que podem ocultar os efeitos
específicos da perda de sono (Pearlman, 1979). Van Hulzen e Coenen (1981),
propuseram a técnica da plataforma múltipla como modelo de PS, na qual se
evita o efeito gerado pela imobilização. Nesse método utiliza-se um tanque de
água contendo várias plataformas para um mesmo animal experimental. Nunes
e Tufik (1994) propuseram um modelo similar ao da plataforma múltipla, mas
com um grupo de ratos ao invés de um único – o método da plataforma múltipla
modificada (MPMM, Figura 2). Com essa alteração, pretendia-se evitar o
estresse gerado pelo isolamento social e imobilização.
No entanto, Suchecki et al. (1998) verificaram que o MPMM induz
estresse maior do que o observado na plataforma invertida, através da análise
da ativação do eixo hipotalâmico-hipófise-adrenal (HPA). Em seqüência aos
seus estudos, Suchecki e Tufik (2000) avaliaram que um fator associado ao
estresse no MPMM se deve à instabilidade social dos ratos privados,
provenientes de várias gaiolas, e que essa instabilidade pode ser atenuada se
utilizado um grupo de ratos provenientes da mesma gaiola de moradia. Dados
posteriores demonstraram que o MPMM priva parcialmente (35%) a fase de
sono de ondas lentas, além de abolir por completo o sono paradoxal (Machado
et al., 2004).
Vinte e quatro animais foram separados em três grupos: privação de
sono (PS; n=8), recuperação de sono (RS; n=8) e controle-gaiola (CG; n=8). Os

25
animais dos grupos PS e RS foram habituados ao MPMM, por um período de
uma hora por dia, durante três dias. Após a habituação, foi iniciada a privação
de sono por 96 horas. Os animais tiveram acesso à água e ração ad libitum
durante todo período de privação de sono. Ao fim do período, os animais do
grupo RS foram devolvidos às suas gaiolas por um período de 24 horas, a fim
de recuperar-se da privação. Os animais do grupo CG permaneceram em suas
gaiolas, sob as mesmas condições ambientais dos outros grupos.
Figura 2: Privação de sono por meio do método da plataforma múltipla modificada.
Processamento Histológico
Ao fim de 96 horas de privação de sono para o grupo PS e 24 horas de
recuperação para o grupo RS, os animais dos três grupos foram sacrificados
por decapitação (Figura 3). Os cérebros foram rapidamente removidos,
congelados em gelo seco e mantidos a -80ºC. Para o processamento
histológico, os cérebros foram seccionados em criostato (-18ºC) da região do

26
bulbo olfatório até o bulbo caudal, em secções de 20 m de espessura, com
intervalos de 400 m. As secções cerebrais foram coletadas em lâminas
revestidas com gelatina, e guardadas em freezer (-80ºC) até o dia da marcação
autorradiográfica.
Figura 3: Desenho esquemático do experimento 1.
Marcação de [3H]AMPA
As lâminas contendo as secções cerebrais foram pré incubadas por 30
minutos em solução tampão Tris-acetato 50 mM contendo KSCN 100 mM (pH
7,2; 4ºC). Em seguida, as lâminas foram incubadas durante 45 minutos em
solução tampão contendo [3H]AMPA 10 nM (55,5 Ci/mmol, Perkin-Elmer, EUA).
A marcação de sítios inespecíficos foi obtida adicionando-se 10 uM de
quisqualato às lâminas preparadas para esse fim. Após o período de
incubação, as lâminas foram lavadas em tampão à 0-4ºC, seguido por um
rápido banho em solução de 100 ml de acetona contendo 2,5 ml de
glutaraldeído. Em seguida, as secções foram secas e expostas a filme sensível
ao trício (Kodak BioMax MR-1, Amersham Pharmacia Biotech) por 6 semanas
na presença de padrão calibrado. Para cada região cerebral, foi calculada a
média dos valores de densidade óptica delimitada dentro de cada seção. Em

27
seguida, a média dos valores dessa mesma região foi quantificada para todas
as secções dentro de cada encéfalo, e convertida em Ci/g de tecido por meio
do padrão calibrado. O valor final é a média da região cerebral de todos os
animais dentro de cada grupo. As regiões anatômicas foram definidas de
acordo com o atlas de Paxinos & Watson (1998).
Análise Estatística
Os dados para a marcação dos sítios dos receptores AMPA foram
analisados através de análise de variância de uma via (ANOVA), utilizando
grupo como fator para cada estrutura cerebral, com nível de significância de p <
0,05. Análises post hoc foram feitas através do Teste de Duncan, com nível de
significância de p < 0,05. Todas as análises estatísticas foram realizadas com o
programa STATISTICA (StatSoft Inc., EUA).
4.1.2. Experimento 1-2: Hibridização in situ da Subunidade GluR1
após Privação e Recuperação de Sono
Estudos mostram que neuropatologias associadas com prejuízos de
memória e redução de receptores AMPA, como mal de Alzheimer (Pellegrini-
Giampietro et al. 1994) e isquemia cerebral (Montori et al., 2010), estão
acompanhadas por uma redução da transcrição da subunidade GluR1 na
formação hipocampal. Por outro lado, dados de Ravassard et al. (2009)
mostram que não há modificação de níveis de RNA mensageiro dessa
subunidade em ratos privados de sono por 72 h. O experimento teve como

28
objetivo especifico verificar os efeitos da privação e recuperação de sono sobre
a expressão da subunidade GluR1 em subáreas hipocampais onde houve
alteração de marcação de [3H]AMPA, de acordo com os dados obtidos no
Experimento 1-1.
Privação de Sono e Processamento Histológico
Um novo conjunto de ratos, subdivididos em três grupos (CG, PS, RS,
todos n=8), foi submetido ao procedimento de privação de sono conforme
descrito para o experimento 1-1. Em seguida, foram sacrificados por
decapitação e os cérebros, rapidamente removidos com instrumentos
cirúrgicos livres de RNases, congelados em gelo seco e mantidos a -80ºC.
Para o processamento histológico, os cérebros foram seccionados em criostato
(-18ºC), sendo selecionado o tecido contendo a formação hipocampal, em
seções de 20 m de espessura, com intervalos de 50 m. As secções
cerebrais foram coletadas em lâminas estéreis com aderência eletrostática
(Microscopic Plus, Fisher), e guardadas em freezer (-80ºC) até o momento do
início de ensaios de hibridização in situ.
Hibridização in situ
As lâminas foram gradualmente aquecidas à temperatura ambiente,
fixadas em formaldeído 4% (pH 7,4) e lavadas em solução PBS 10 mM (pH
7,4). Em seguida, foram tratadas por 30 minutos com proteinase K (0,5 ug/ml;
Life Technologies, EUA), EDTA (0.05 M, pH 8,0) e Tris–HCl (0.1 M, pH 8,0).
Depois, as lâminas foram acetiladas com trietanolamina-HCl (Avocado
Research, Reino Unido), lavadas em 2xSSC e desidratadas em uma série de

29
soluções de etanol. A hibridização foi realizada utilizando uma ribossonda
marcada com 35S-UTP complementar à sequência de interesse, produzida por
meio de transcrição in vitro (Promega, EUA). Durante a transcrição, uma
amostra de DNA complementar (DNAc) foi amplificada por meio de cadeia de
reação de polimerase (PCR) utilizando primers complementares à sequência
da subunidade GluR1 (Genbank # NM_031608, bases 108–405). Os primers
estão incorporados a sequências promotoras tanto da RNA polimerase SP6
(atttaggtgaacactatagaa) na extremidade 5’ do primer sense, como da RNA
polimerase T7 (taatacgactcactataggg) na extremidade 5’ do primer antisense.
Esta incorporação possibilita a produção de ribossondas alvo e controle de
RNA em fitas simples a partir do produto amplificado de DNAc. As ribossondas
foram diluídas (106 cpm/ml) em solução de hibridização, contendo formamida
50%, thiosulfato de sódio 1% e diothiothreitol (DTT) 5 M 2%. A solução de
hibridização foi aplicada nas secções de tecido, e as lâminas foram incubadas
a 57ºC durante 16 horas. Para o procedimento de pós hibridização, as secções
foram imersas em solução contendo RNAse A (20 μg/ml; USB, EUA) por 30
minutos e em seguida lavadas em tampão de RNase (NaCl 5 M 10%, Tris 1 M
4%, EDTA 0,2%, pH 8) por 30 minutos em temperatura ambiente. As lâminas
foram então lavadas em soluções contendo concentrações decrescentes de
SSC, desidratadas e expostas a filme sensível a emissão radioativa (Kodak
BMR-2, EUA) por quatro dias. A quantificação da hibridização foi obtida
mensurando a densidade ótica relativa das áreas cerebrais analisadas,
subtraindo-se o valor verificado no fórnix (região adjacente com menor
expressão de transcrição), usando um sistema de análise imagem (M2 MCID
system, Imaging Research, Canadá).

30
Análise Estatística
De forma similar ao Experimento 1-1, os dados para a hibridização in
situ da subunidade GluR1 foram analisados através de análise de variância de
uma via (ANOVA), utilizando grupo como fator para cada estrutura da formação
hipocampal, com nível de significância de p < 0,05. As análises post hoc foram
feitas através do Teste de Duncan, com nível de significância de p < 0,05.
Todas as análises estatísticas foram realizadas com o programa STATISTICA
(StatSoft Inc., EUA).
4.2. Experimento 2 – Efeitos da Administração de Potenciador e
Antagonista de Receptores AMPA no Desempenho de Esquiva
Inibitória de Ratos Privados de Sono
O objetivo deste experimento foi verificar os efeitos da administração do
potenciador aniracetam e do antagonista GYKI 52466 sobre os prejuízos de
memória decorrentes da privação de sono. Cada fármaco e forma de
tratamento (agudo, crônico) foi avaliado em experimentos separados, que
compartilham o mesmo procedimento metodológico descrito a seguir.
Sujeitos
Foram utilizados ratos Wistar machos, com três meses de idade,
provenientes do biotério do departamento de Psicobiologia (UNIFESP). Os
animais foram mantidos em condições controladas de temperatura (22-24 °C) e

31
ciclo claro escuro de 12/12 hr (período claro iniciado às 7:00h), com livre
acesso ao alimento e água.
Procedimento de Privação de Sono
Os animais foram submetidos ao procedimento de privação de sono por
96 h em plataformas múltiplas modificada. Os animais do grupo PS foram
colocados em grupo em um tanque de água contendo nove plataformas
estreitas (6,2 cm de diâmetro) espaçadas 7-8 cm entre si. Água e ração foram
disponibilizadas ad libitum, e o tanque foi limpo diariamente. Os animais
controles (CG) foram mantidos na mesma sala onde ocorreu a privação de
sono.
Tarefa de Esquiva Inibitória
A tarefa de esquiva inibitória foi dividida em três sessões consecutivas:
treino, teste 1 e teste 2. Durante a sessão de treino, cada animal foi colocado
no compartimento claro de um aparato de esquiva inibitória (AVS Projetos
Especiais, Figura 4) e após 10 segundos, uma porta deslizante foi aberta.
Assim que o animal entrou no compartimento escuro, a porta deslizante foi
fechada e o animal recebeu um choque nas patas (0,8 mA/1 s). O animal foi
mantido no compartimento escuro por mais 15 segundos, e então retirado do
aparato e colocado em sua gaiola-moradia. Duas horas após a sessão de
treino, a primeira sessão de teste (teste 1) foi realizada, sob as mesmas
condições da sessão de treino, com exceção do choque, que foi desligado.
Uma segunda sessão de teste (teste 2) foi realizada 24 horas depois da sessão

32
de treino, seguindo o mesmo procedimento do teste 1. A latência máxima para
que o animal entrasse no compartimento escuro foi de 300 segundos.
Figura 4: Aparato de esquiva inibitória.
Fármacos
Foram utilizados dois ligantes de receptores AMPA:
1. o modulador alostérico positivo aniracetam (1-(4-metoxibenzoil)-2-
pirrolidinona, Sigma). O potenciador aniracetam foi suspenso em
uma solução de carboxi-metil-celulose 0,25%, contendo algumas
gotas de Tween 80.
2. o antagonista não competitivo GYKI 52466 (1-(4-aminofenil)-4-metil-
7,8-metilenedioxi-5H-2,3-benzodiazepina, Sigma). O fármaco foi
dissolvido em gotas de 0,1 M HCl e diluído em água destilada.

33
4.2.1. Experimento 2-1: Efeitos da Administração Aguda de
Aniracetam em Ratos Privados de Sono
O experimento foi realizado no intuito de avaliar se o tratamento agudo com
o potenciador aniracetam poderia prevenir prejuízos de desempenho na tarefa
de esquiva inibitória em animais privados de sono.
Tratamento farmacológico
Ao final do procedimento de privação de sono, o potenciador aniracetam
foi aplicado por via oral (volume de 2 ml/kg) nas doses de 25, 50, e 100 mg/kg.
Animais controles para tratamento receberam veículo (grupos n = 8-10). Dessa
forma, foram obtidos oito grupos: CG-vc, CG-ani25, CG-ani50, CG-ani100, PS-
vc, PS-ani25, PS-ani50, e PS-ani100. A administração foi realizada ao final do
período de privação de sono, cerca de 1 hora antes do início da tarefa de
esquiva inibitória (Figura 5).
Figura 5: Desenho esquemático do experimento 2-1. Ao final do procedimento de privação de
sono, animais recebem administração aguda de aniracetam ou veículo (via oral) e são
treinados na tarefa de esquiva inibitória (EI).

34
4.2.2. Experimento 2-2: Efeitos da Administração Aguda de GYKI
52466 em Ratos Privados de Sono
O objetivo do presente experimento foi verificar se o tratamento agudo de
GYKI 52466 em ratos submetidos à privação de sono é capaz de prevenir
prejuízos de desempenho na tarefa de esquiva inibitória, com base numa
possível modulação de receptores AMPA resultando na prevenção de prejuízos
de memória associados a aumento excessivo de glutamato, tal como verificado
em estudos de isquemia cerebral (Block e Schwarz, 1997; Gyertyan et al.
1999).
Tratamento Farmacológico
O antagonista GYKI 62466 foi aplicado por via intraperitoneal (i.p.)
(volume de 2 ml/kg) nas doses de 5 e 10 mg/kg ao final do procedimento de
PS. Animais controles para tratamento receberam veículo. Dessa forma, foram
obtidos seis grupos: CG-vc, CG-gyki5, CG-gyki10, PS-vc, PS-gyki5, e PS-
gyk10. A administração foi realizada ao final do período de PS, cerca de 1 hora
antes do início da tarefa de esquiva inibitória. (Figura 6).

35
Figura 6: Desenho esquemático do experimento 2-2. Ao final do procedimento de privação de
sono, animais receberam administração aguda de GYKI 52466 ou veículo (i.p.) e foram
treinados na tarefa de esquiva inibitória (EI).
4.2.3. Experimento 2-3: Efeitos da Administração Crônica de GYKI
52466 em Ratos Privados de Sono
O objetivo do presente experimento foi avaliar se o tratamento crônico com
GYKI 52466 em ratos privados de sono seria capaz de prevenir a
subsensibilidade de receptores AMPA hipocampais pela PS e,
consequentemente, impedir o prejuízo de desempenho na tarefa de esquiva
inibitória, de acordo com o estudo de Sinha et al. (1999).
Tratamento Farmacológico
O antagonista GYKI 62466 foi aplicado por via i.p. (volume de 2 ml/kg)
nas doses de 5 e 10 mg/kg. Animais controles para tratamento receberam
veículo (n = 8-14/grupo). As administrações ocorreram durante o período de
privação de sono, duas vezes ao dia (as 11:00 h e as 17:00 h,
respectivamente) ao longo de quatro dias, sendo descontinuadas cerca de 17 h

36
antes do início da tarefa de esquiva inibitória, a fim de evitar efeitos agudos do
fármaco (Figura 7). Dessa forma, foram obtidos seis grupos: CG-vc, CG-gyki5,
CG-gyki10, PS-vc, PS-gyki5, e PS-gyk10.
Figura 7: Desenho esquemático do experimento 2-3. Animais controles e privados de sono
foram tratados sistemicamente com GYKI 52466 ou veículo (duas injeções i.p. por dia) durante
o período de privação de sono, antes de serem treinados na tarefa de esquiva inibitória (EI).
Análise Estatística
Para cada experimento, foi realizada uma análise de variância (ANOVA) de
duas via com medidas repetidas, utilizando grupos (CG e PS sob diferentes
tratamentos) e latência de passagem nas sessões (treino, teste 1, teste 2)
como fatores. Análises post hoc foram feitas através do Teste de Duncan, com
nível de significância de p < 0,05.

37
4.3. RESULTADOS
4.3.1. Experimento 1-1: Autorradiografia de Receptores AMPA após
Privação e Recuperação de Sono
As características da marcação de [3H]AMPA dos grupos controle gaiola
(CG), Privação de Sono (PS) e Recuperação de Sono (RS) estão contidas na
tabela 1. As análises de variância (ANOVA) de uma via das estruturas
estudadas indicam não haver diferença significativa da marcação entre os
grupos, com exceção das regiões CA1, CA3 e giro denteado da formação
hipocampal. As análises posteriores de pelo teste de Duncan revelaram uma
marcação reduzida nos animais privados de sono em comparação aos
controles (p < 0,05).
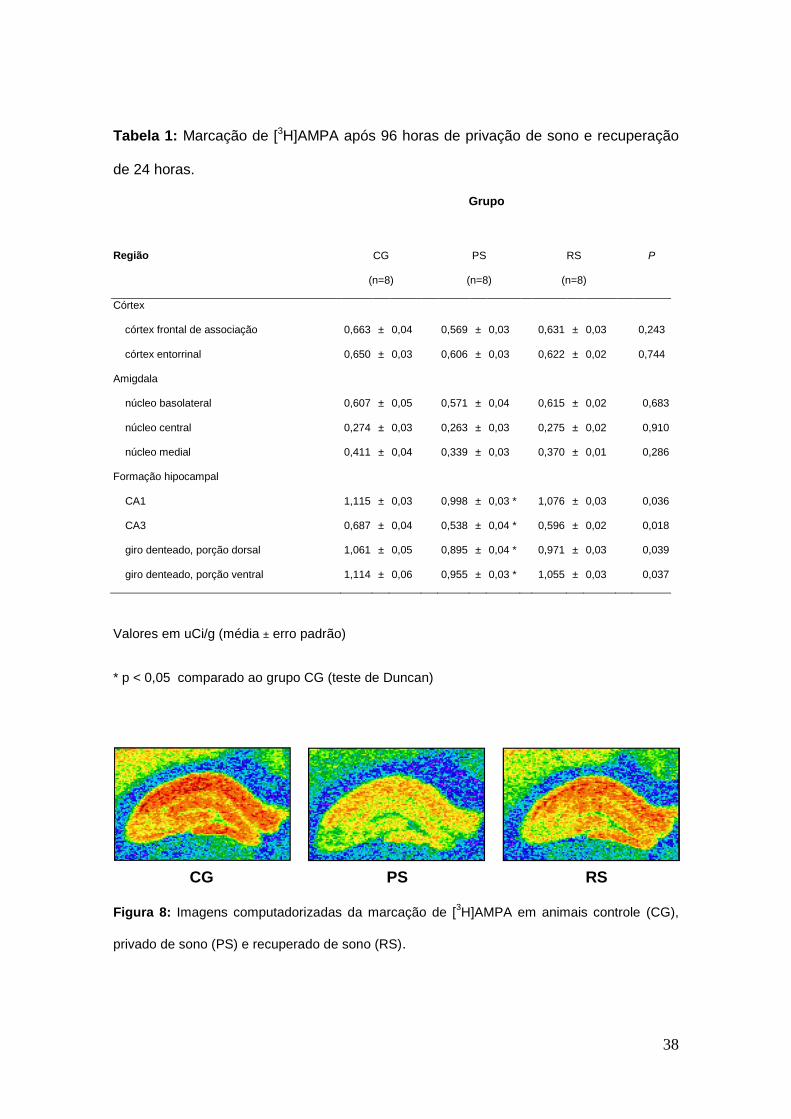
38
Tabela 1: Marcação de [3H]AMPA após 96 horas de privação de sono e recuperação
de 24 horas.
Grupo
Região CG PS RS P
(n=8) (n=8) (n=8)
Córtex
córtex frontal de associação 0,663 ± 0,04 0,569 ± 0,03 0,631 ± 0,03 0,243
córtex entorrinal 0,650 ± 0,03 0,606 ± 0,03 0,622 ± 0,02 0,744
Amigdala
núcleo basolateral 0,607 ± 0,05 0,571 ± 0,04 0,615 ± 0,02 0,683
núcleo central 0,274 ± 0,03 0,263 ± 0,03 0,275 ± 0,02 0,910
núcleo medial 0,411 ± 0,04 0,339 ± 0,03 0,370 ± 0,01 0,286
Formação hipocampal
CA1 1,115 ± 0,03 0,998 ± 0,03 * 1,076 ± 0,03 0,036
CA3 0,687 ± 0,04 0,538 ± 0,04 * 0,596 ± 0,02 0,018
giro denteado, porção dorsal 1,061 ± 0,05 0,895 ± 0,04 * 0,971 ± 0,03 0,039
giro denteado, porção ventral 1,114 ± 0,06 0,955 ± 0,03 * 1,055 ± 0,03 0,037
Valores em uCi/g (média ± erro padrão)
* p < 0,05 comparado ao grupo CG (teste de Duncan)
Figura 8: Imagens computadorizadas da marcação de [3H]AMPA em animais controle (CG),
privado de sono (PS) e recuperado de sono (RS).
CG PS RS

39
4.3.2. Experimento 1-2: Hibridização in Situ da Subunidade GluR1
após Privação e Recuperação de Sono
As características da hibridização in situ da subunidade GluR1 dos
grupos Controle (CG), Privação de Sono (PS) e Recuperação de Sono (RS)
estão contidas na tabela 2. As análises de variância (ANOVA) de uma via das
estruturas hipocampais estudadas indicam não haver diferença significativa da
transcrição da subunidade de GluR1 entre os grupos do experimento.
Tabela 2: Hibridização in Situ da Subunidade GluR1 após 96 horas de privação de
sono e recuperação de 24 horas.
Grupo
Região CG PS RS P
(n=8) (n=8) (n=8)
CA1 1,061 ± 0,25 0,929 ± 0,15 0,947 ± 0,27 0,937
CA3 0,885 ± 0,22 0,678 ± 0,16 0,719 ± 0,23 0,889
Giro denteado, porção dorsal 1,348 ± 0,27 1,355 ± 0,14 1,128 ± 0,25 0,677
Giro denteado, porção ventral 1,566 ± 0,22 1,288 ± 0,15 1,203 ± 0,24 0,641
Valores em densidade óptica relativa (média ± erro padrão)

40
Figura 9: Imagem computadorizada da hibridização in situ da subunidade GluR1 na formação
hipocampal de um animal controle.
4.3.3. Experimento 2-1: Efeitos da Administração Aguda de
Aniracetam em Ratos Privados de Sono
Os dados estão apresentados na Figura 10. Foram encontrada efeitos dos
fatores grupo (F(7, 70)=10.161, p < 0,05), sessão (F(2, 140)=187.19, p < 0,05),
e interação entre os fatores (F(14, 140)=6.1530, p < 0,05). A análise a
posteriori da interação dos fatores revelou que os grupos se comportaram de
forma similar na sessão de treino (p > 0,05), ao passo que nas sessões de
teste 1 e 2 os animais controles apresentaram latências de passagem maiores
do que os animais privados de sono (p < 0,05), com exceção daqueles
privados de sono tratados com 100 mg/kg de aniracetam (p > 0,24), que por
sua vez apresentaram latência de passagem maior do que dos animais
privados de sono tratados com veículo (p < 0,05).

41
Figura 10: Latência de passagem de ratos privados de sono tratados com aniracetam na tarefa
de esquiva inibitória (média ± erro padrão em segundos). CG = controle gaiola, PS = privação
de sono, vc = veículo, ani25 = 25 mg/kg aniracetam, ani50 = 50 mg/kg aniracetam, ani50 = 50
mg/kg, ani100 = 100 mg/kg aniracetam, * p < 0,05 vs. animais controles sob o mesmo
tratamento, na mesma sessão (teste de Duncan), # p < 0,05 vs. animais privados de sono
tratados com veículo, na mesma sessão (teste de Duncan).
0
50
100
150
200
250
300
350
Treino Teste 1 Teste 2
Latê
nci
a (s
egu
nd
os)
Sessão
CG vc (10)
CG ani25 (10)
CG ani50 (10)
CG ani100 (8)
PS vc (10)
PS ani25 (10)
PS ani50 (10)
PS ani100 (10)
* *
# #

42
4.3.4. Experimento 2-2: Efeitos da Administração Aguda de GYKI
52466 em Ratos Privados de Sono
Os dados estão apresentados na Figura 11. Foram encontrados efeitos
dos fatores grupo (F(5, 46)=17.046, p < 0,05), sessão (F(2, 92)=160.63, p <
0,05), e interação entre os fatores (F(10, 92)=8.3627, p < 0,05). A análise a
posteriori da interação dos fatores revelou que a latência de passagem dos
animais privados de sono foi menor do que a latência dos animais controles
(teste de Duncan, p < 0,05), independente do tratamento aplicado.
Figura 11: Latência de passagem de ratos privados tratados agudamente com GYKI 52466 na
tarefa de esquiva inibitória (média ± erro padrão em segundos). CG = controle gaiola, PS =
privação de sono, vc = veículo, gk5 = 5 mg/kg GYKI 52466, gk10 = 10 mg/kg GYKI 52466. * p <
0,05 vs. animais controles sob o mesmo tratamento, na mesma sessão (teste de Duncan).
0
50
100
150
200
250
300
350
Treino Teste 1 Teste 2
Latê
nci
a (s
egu
nd
os)
Sessão
CG vc (8)
CG gyki 5 (8)
CG gyki10 (8)
PS vc (10)
PS gyki5 (8)
PS gyki10 (10)
* *

43
4.3.5. Experimento 2-3: Efeitos da Administração Crônica de GYKI
52466 em Ratos Privados de Sono
Os dados estão apresentados na Figura 12. Foram encontrados efeitos
dos fatores grupo (F(5, 54)=11,676, p < 0,01) e sessão (F(2, 108)=114,36, p <
0,01), bem como interação entre os fatores (F(10, 108)=6,4066, p < 0,01). A
análise a posteriori da interação dos fatores revelou que a latência de
passagem dos animais privados foi menor do que a latência dos animais
controles (teste de Duncan, p < 0,05), independente do tratamento aplicado.
Figura 12: Latência de passagem de ratos privados tratados cronicamente com GYKI 52466 na
tarefa de esquiva inibitória (média ± erro padrão em segundos). CG = controle gaiola, PS =
0
50
100
150
200
250
300
350
Treino Teste 1 Teste 2
Latê
nci
a (s
egu
nd
os)
Sessão
CG vc (14)
CG gk5 (8)
CG gk10 (8)
PS vc (14)
PS gk5 (8)
PS gk10 (8)
* *

44
privação de sono, vc = veículo, gk5 = 5 mg/kg GYKI 52466, gk10 = 10 mg/kg GYKI 52466. * p <
0,05 vs. animais controles sob o mesmo tratamento, na mesma sessão (teste de Duncan).
4.4. DISCUSSÃO PARCIAL
Os dados de marcação autorradiográfica dos receptores AMPA
(Experimento 1-1) revelaram que houve uma redução seletiva em regiões da
formação hipocampal (giro denteado, CA1 e CA3, Figura 8) de ratos privados
de sono por 96 horas. As demais áreas cerebrais analisadas não mostraram
qualquer alteração significativa de marcação, apesar de estarem fortemente
associadas com a formação de memórias emocionais aversivas, tais como o
córtex entorrinal e a amígdala basolateral (McGaugh et al., 1990; Ueki et al.,
1994). Não foram encontradas diferenças na marcação de receptores AMPA
hipocampais entre os animais controles e os animais do grupo RS,
demonstrando uma normalização dos efeitos da privação de sono sobre esses
receptores mediante a oportunidade de dormir 24 horas após o procedimento
comportamental.
Com relação à hibridização in situ da subunidade GluR1 (Experimento 1-
2), não foram encontradas diferenças significativas entre os grupos
experimentais em nenhuma região hipocampal. Esses achados encontram
consonância com o trabalho de Ravassard et al. (2009), no qual a expressão
da proteína GluR1 é reduzida no hipocampo de ratos privados de sono por 72
horas, ao passo que a expressão dos transcritos da subunidade GluR1 não

45
encontra-se alterada nessa condição. É plausível que, ao invés de
modificações na transcrição das subunidades que os compõem, a diminuição
da marcação de receptores AMPA seja decorrente de mecanismos pós
translacionais, tal como a internalização de receptores na membrana dos
terminais pós sinápticos, como verificado para as subunidades que compõe o
receptor NMDA em ratos privados de sono (McDermott et al., 2006). Nesse
sentido, apesar de empregar protocolos de privação de sono distintos (privação
de sono total por 12 horas e privação de sono paradoxal por 72 horas), os
trabalhos de Hagewoud et al. (2009) e Ravassard et al. (2009) coincidem
quanto aos efeitos da perda de sono induzindo redução da fosforilação do sítio
serina 845 da subunidade GluR1, que está associada à incorporação de
receptores AMPA na membrana (Esteban et al., 2003).
A redução e a normalização da marcação de receptores AMPA
hipocampais após a privação e recuperação de sono, respectivamente, abrem
interessantes vias de investigação quanto aos mecanismos subjacentes aos
efeitos deletérios da privação de sono no desempenho de tarefas de memória.
Como verificado em estudos anteriores, a recuperação de sono atenua os
prejuízos decorrentes da privação de sono em tarefas dependentes da
formação hipocampal. Dessa forma, McDermott et al. (2003) demonstraram
que ratos que puderam se recuperar depois de serem privados de sono por 72
horas não apresentam prejuízos de desempenho no condicionamento de medo
ao contexto. Resultados similares foram obtidos em nosso laboratório: ratos
privados de sono por 96 horas que puderam recuperar-se em suas gaiolas
durante 24 horas apresentam uma melhora dos efeitos deletérios da privação
de sono sobre a tarefa de esquiva inibitória (Dubiela et al., 2005, 2010).

46
Tomados em conjunto, esses dados sugerem que os receptores AMPA
hipocampais, participantes relevantes na formação de memórias aversivas
contextuais, parecem estar envolvidos tanto nos prejuízos mnésicos da
privação de sono como na reversão desses efeitos com sua recuperação.
Nossos dados farmacológicos demonstraram que os prejuízos de
desempenho na tarefa de esquiva inibitória, induzidos pela privação de sono,
são consistentes tanto na memória de curto prazo (avaliada 2 horas depois do
treino), como na memória de longo prazo (avaliada 24 horas depois do treino),
corroborando estudos anteriores (Dubiela et al. 2005, 2010). Além disso,
verificamos que o tratamento agudo com o potenciador aniracetam, na dose de
100 mg/kg, é capaz de prevenir tais prejuízos, em ambos os testes
consecutivos da tarefa (Experimento 2-1). Esses resultados encontram
consonância com aqueles de Porrino et al. (2005) e Boyle et al. (2011), que
observaram melhora dos prejuízos cognitivos induzidos pela privação de sono
em humanos e macacos-rhesus por meio de tratamento com o potenciador
CX717. De forma similar aos dados presentes em nosso estudo, o estudo de
Boyle e colaboradores (2011) avaliaram que a prevenção só foi obtida com a
aplicação de uma dose alta do fármaco.
A evidência de redução da função de receptores AMPA após a privação
de sono, obtida em nosso laboratório e em outros (Experimento 1-1, Ravassard
et al., 2009, Hagewoud et al., 2009), sugere uma forte relação com a ação do
aniracetam em ratos privados de sono. Apesar de doses baixas e
intermediárias do fármaco se mostrarem suficientes para promover melhora em
diversos modelos de prejuízo de memória (Cumin et al., 1982; Gouliaev e
Senning, 1994), as doses de 25 e 50 mg/kg não foram eficazes em prevenir os

47
prejuízos induzidos pela privação de sono em nosso experimento. Essa
ausência de melhora pode ser decorrente da baixa disponibilidade de
receptores AMPA após 96 horas de privação de sono, de forma similar à ação
de antagonistas bloqueando os efeitos antiamnésicos de potenciadores desses
receptores (Lu & Wehner, 1997). Por sua vez, a dose mais alta de aniracetam
(100 mg/kg) parece atuar no sentido de promover ativação suficiente sobre os
receptores remanescentes, sendo assim, capaz de anular os prejuízos de
memória induzidos pela privação de sono.
Com base em estudos que mostraram efeitos protetores e
antiamnésicos utilizando antagonistas de receptores AMPA num modelo de
isquemia cerebral (Block e Schwarz, 1997; Gyertyan et al., 1999), buscamos
verificar se um tratamento similar seria capaz de prevenir os prejuízos de
desempenho de esquiva inibitória induzida por PS. A nossa hipótese não foi
confirmada, haja vista que tanto a administração aguda como crônica de GYKI
52466 não produziram efeito sobre o desempenho dos ratos privados de sono.
Apesar das semelhanças encontradas entre os efeitos induzidos pela privação
de sono e a isquemia cerebral sobre o sistema glutamatérgico, tais como
aumento de níveis de glutamato e redução de receptores AMPA (Ravassard et
al., 2009; Montori et al., 2010; Mohammed et al., 2011; Kostandy, 2011), é
necessário reconhecer que a dinâmica dessas alterações ocorre de forma
distinta entre estes dois modelos. Por exemplo, enquanto que o aumento de
glutamato no modelo de isquemia cerebral ocorre de forma aguda e intensa
(Kostandy, 2011), o mesmo não ocorre durante a privação de sono, sendo
mantido em níveis elevados a partir de períodos curtos (12 h, Bettendorff et al.,
1996) a longos de PS (72 h, Mohammed et al., 2011).

48
Há algumas diferenças relevantes entre os trabalhos prévios que
empregaram o modelo de isquemia cerebral (Block e Schwarz, 1997; Gyertyan
et al., 1999) e o presente estudo em relação ao tratamento agudo com GYKI
52466, que podem ter contribuído para a ausência de efeitos do antagonista na
tarefa de esquiva inibitória. Enquanto Gyertyan et al. (1999) utilizaram gerbilos,
Block e Schwarz (1997) aplicaram o antagonista em ratos no momento da
intervenção isquêmica, de forma aguda, uma semana antes de avaliá-los na
versão espacial do labirinto aquático de Morris. Além disso, a alta dose utilizada
nesse estudo (30 mg/kg, Block e Schwarz, 1997) induz ataxia (Danysz et al.,
1994), o que dificultaria sua aplicação antes do treino no paradigma da tarefa
de esquiva inibitória em ratos privados de sono. Dessa forma, optamos por
administrar doses mais baixas do fármaco (5 e 10 mg/kg, Experimento 2-2), na
mesma faixa de doses aplicada no experimento crônico (Experimento 2-3).
É sabido que administração crônica de GYKI 52466 aumenta a
marcação de receptores AMPA (Sinha et al., 1999), e que a facilitação da
atividade desses receptores induz melhora de desempenho em tarefas de
memórias hipocampo dependentes (Gouliaev e Senning, 1994). Entretanto,
não verificamos melhora de desempenho de animais privados de sono ou
controles após 4 dias de tratamento com GYKI 52466 (Experimento 2-3). É
plausível supor que o tratamento crônico possa ter atenuado a redução de
receptores AMPA hipocampais após a privação de sono (Experimento 1-1), e
que não tenha sido suficiente para produzir um efeito sobre o prejuízo de
desempenho de ratos privados de sono. Uma possibilidade para testar esta
hipótese reside na combinação entre o tratamento crônico do antagonista GYKI
52466 com o tratamento agudo de doses subefetivas de aniracetam (por

49
exemplo, 25 e 50 mg/kg, conforme Experimento 2-1). Essa combinação poderia
produzir uma sinergia entre os dois fármacos e, assim, aumentar a eficácia da
reversão dos prejuízos de memória induzidos pela privação de sono com doses
sublimiares do potenciador de receptores AMPA.
A tarefa de esquiva inibitória, utilizada nos experimentos farmacológicos
descritos, é um paradigma que permite avaliar a memória de curto e longo
prazos de forma consecutiva (Izquierdo et al., 1998). Em sua constituição,
utiliza tanto mecanismos de aprendizagem instrumental quanto componentes
do condicionamento clássico de medo (Xavier, 1982). Dentro desse quadro,
para acessar, simultaneamente, efeitos específicos da privação de sono e da
administração do aniracetam sobre memórias hipocampo-dependentes, seria
necessário a utilização de um paradigma que permitisse dissociar memórias
dependentes e independentes do hipocampo, associado a um tratamento local
do fármaco na região de interesse.
O condicionamento clássico de medo é uma tarefa que pareia um
estímulo neutro com um estímulo aversivo e, após algum tempo, o estímulo
neutro assume as propriedades do estímulo aversivo, desencadeando as
mesmas respostas e sendo, então, denominado de estímulo condicionado
(Philips & LeDoux, 1992; Maren & Aharonov, 1996; Goosens & Maren, 2001).
Quando o pareamento é feito entre um estímulo complexo e espacial (como o
ambiente) e o estímulo aversivo (como um choque) a tarefa é denominada de
condicionamento de medo ao contexto e é dependente do hipocampo (Philips
& LeDoux, 1992). Por outro lado, quando o pareamento é feito entre um
estímulo discreto (como um som ou uma luz) e o mesmo tipo de estímulo
aversivo (choque), a tarefa é nomeada como condicionamento de medo ao

50
som. Para esse tipo de aprendizagem, a lesão do hipocampo não provoca
prejuízo no desempenho dos animais (Philips & LeDoux, 1992); por isso, diz-se
que a tarefa de condicionamento ao som é hipocampo-independente.
Baseados nesse panorama planejamos um novo experimento, com o
intuito de acessar os efeitos do potenciador aniracetam administrado
diretamente no hipocampo de ratos privados de sono, utilizando um paradigma
que pudesse dissociar, simultaneamente, memórias dependentes e
independentes do hipocampo: o condicionamento clássico de medo.
4.5. EXPERIMENTO COMPLEMENTAR: EFEITOS DA
ADMINISTRAÇÃO INTRA-HIPOCAMPAL DE ANIRACETAM EM
RATOS PRIVADOS DE SONO NO CONDICIONAMENTO DE
MEDO
Considerando os efeitos seletivos da privação de sono sobre a
marcação de receptores AMPA na região hipocampal (Experimento 1-1) e os
dados farmacológicos obtidos sob o paradigma da esquiva inibitória
(Experimento 2-1), planejamos um novo estudo com o intuito de acessar os
efeitos do potenciador aniracetam administrado diretamente no hipocampo de
ratos privados de sono. Para tanto, um conjunto de ratos foi operado para
implante de cânulas na região hipocampal. Estes animais foram submetidos
posteriormente ao procedimento de privação de sono, administração local de
soluções, e avaliação de desempenho na tarefa de condicionamento clássico
de medo, nessa ordem. Optamos por alterar a tarefa de memória para acessar,

51
simultaneamente, efeitos específicos da privação de sono e da administração
local do aniracetam sobre memórias dependentes do hipocampo (por meio do
condicionamento de medo ao contexto) em relação a memórias independentes
(por meio do condicionamento de medo ao som).
Sujeitos
Foram utilizados ratos Wistar machos, com três meses de idade,
provenientes do biotério do CEDEME (UNIFESP). Os animais foram mantidos
em condições controladas de temperatura (22-24 ºC) e ciclo claro escuro de
12/12 hr (período claro iniciado às 7:00 h), com livre acesso ao alimento e
água.
Cirurgia Estereotáxica
Cada animal foi anestesiado por via i.p. com uma solução associada de
quetamina e xilasina (Agener), na dose de 1 ml/kg para cada um dos fármacos.
Após atingir o plano anestésico, o pêlo da região craniana foi aparado e a
cabeça foi afixada num aparelho de estereotaxia (David Kopf Instruments). A
pele, tecido muscular subcutâneo e periósteo foram divulsionados com auxílio
de um bisturi lâmina 23 até exposição da tábua óssea. Após a ocorrência da
hemostasia, dois pequenos orifícios (cerca de 1 mm) foram realizados para
implantação de cânulas-guia bilaterais na região CA1 da formação hipocampal
(3,8 mm posterior e 2,4 mm lateral ao bregma, 1,6 mm de profundidade),
segundo o atlas de Paxinos & Watson (1998). Mais três orifícios foram
perfurados para implantação de parafusos para manutenção do capacete de
cimento de acrílico dental. Em cada cânula foi colocado um mandril para evitar

52
sua obstrução. Após o procedimento cirúrgico foi administrado pentabiótico (via
intramuscular) e diclofenaco de sódio (via i.p.), e os animais foram colocados
em gaiolas individuais aquecidas por 24 h, antes de retornar às suas gaiolas-
moradia. Os procedimentos comportamentais (privação de sono seguida por
avaliação de desempenho no condicionamento clássico de medo) foram
realizados após o período de recuperação pós-operatória (5-7 dias).
Procedimento de Privação de Sono
Após a recuperação de cirurgia, os animais foram submetidos ao
procedimento de privação de sono por 96 h em plataformas múltiplas. Os
animais do grupo PS foram colocados em duplas (a fim de prevenir isolamento
social) em um tanque de água contendo cinco plataformas estreitas (6,2 cm de
diâmetro) espaçadas 7-8 cm entre si. Água e ração foram disponibilzadas ad
libitum, e o tanque foi limpo diariamente. Os animais controles foram mantidos
em gaiolas individuais (mas com a oportunidade de interagir entre si entre as
grades), na mesma sala onde ocorreu a privação de sono.
Procedimento de Injeção Intra-Hipocampal
Após o procedimento de privação de sono, as soluções de infusão
(veículo ou aniracetam – doses de 0,1 μg e 1 μg, dissolvido em solução de
0,25% DMSO) foram injetadas bilateralmente no hipocampo através de agulhas
de microinjeção (Figura 13). Cada agulha de microinjeção é 1 mm mais longa
do que a cânula guia. Após cuidadosa imobilização do animal, o mandril foi
retirado e a agulha de microinjeção foi colocada dentro da cânula-guia. Cada
agulha foi fixada à extremidade de uma seringa Hamilton de 5 ml por meio de

53
um tubo de polietileno (PE-10). As infusões, em volume de 1 μl, foram
controladas por uma bomba de infusão (Insight), na qual foi acoplada a seringa
Hamilton. A velocidade de infusão foi de 0,5 μl/minuto. Terminada a injeção, a
agulha injetora permaneceu na cânula-guia por um tempo equivalente ao da
microinjeção, a fim de evitar refluxo da solução, e só então a agulha foi retirada
e o mandril recolocado na cânula-guia. O animal foi devolvido a sua gaiola
moradia e, 15 minutos depois, foi submetido à sessão de treino de
condicionamento clássico de medo. As doses escolhidas, bem como o intervalo
entre a administração e a tarefa, foram baseadas nos trabalhos de Lu e
Wehner (1997) e Masuoka et al. (2007).
Figura 13: Desenho experimental do experimento complementar. Ao final de 96 h de privação
de sono, os animais recebem administrações intrahipocampais de aniracetam e são treinados
na tarefa de condicionamento de medo (treino CM). No dia seguinte, os animais são testados
para o condicionamento de medo ao contexto e ao som (testes CMC e CMS).
Aparato de Condicionamento Clássico de Medo
Para a tarefa de condicionamento clássico de medo, utilizamos uma
caixa de condicionamento (21 x 15 cm de base, altura de 27,5 cm) que possui
paredes pretas com cinco quadrados brancos de 6 cm2. A tampa do aparelho
consiste em uma placa de acrílico transparente. A base é formada por grades

54
metálicas conectadas a uma fonte geradora de corrente alternada capaz de
administrar choques elétricos e sinais de som programáveis (AVS Projetos
Especiais). Cada grade da base do aparelho mede 0,4 cm de diâmetro. A
distância entre grades adjacentes é de 1,2 cm. Um recipiente cilíndrico (35 cm
de base e 60 cm de altura) de paredes brancas e piso marrom foi utilizado para
o teste do condicionamento de medo ao som.
Procedimento do Condicionamento Clássico de Medo
A tarefa de condicionamento clássico de medo foi realizada em dois dias
consecutivos. Durante a sessão de treino de condicionamento de medo (CM),
os animais foram colocados no aparato de condicionamento e, 2 minutos após,
receberam cinco pareamentos som-choque (som: 60 dB/5 s; choque: 0,6 mA/1
s), com 30 segundos de intervalo entre os pareamentos. Após o último
pareamento, os animais foram mantidos no aparato por um minuto adicional.
Os animais foram então retirados do aparato e colocados em suas gaiolas
moradias.
Duas sessões de teste foram realizadas após a sessão de treino, a fim
de acessar os efeitos da privação de sono e da administração local do
aniracetam em memórias dependentes e independentes do hipocampo (Phillips
& Ledoux, 1994, Figura 14). O primeiro teste – teste de condicionamento de
medo ao contexto (CMC) – foi realizado 24 horas após a sessão de treino. Os
animais foram colocados por 5 minutos no mesmo aparato de condicionamento
da sessão de treino, sem a presença de som ou choque, e o tempo de
congelamento médio por minuto foi registrado. Em seguida, os animais foram
retirados do aparato e colocados em suas gaiolas moradias. Duas horas

55
depois, foi realizado o segundo teste – teste de condicionamento de medo ao
som (CMS) – em uma nova câmara de teste (novo contexto), numa sala
distinta de onde foram realizadas as sessões anteriores. O animal permaneceu
na câmara por seis minutos. Após três minutos iniciais, os mesmos sons
apresentados na sessão de treino (60 dB/5 s) foram emitidos com intervalos
regulares de 30 segundos por cinco vezes, sem a aplicação de choques nas
patas. O tempo de congelamento médio por minuto foi registrado durante toda
a sessão de CMS, e subdividido em intervalos pré e pós apresentação de som.
Figura 14: Desenho experimental da tarefa de condicionamento clássico de medo. Animais são
treinados na tarefa de condicionamento de medo (treino CM) a contexto e sons específicos, e
consecutivamente testados para cada um desses fatores em separado (testes CMC e CMS).
Análise Estatística
Para o treino de CM foi realizada uma análise de variância (ANOVA) de duas
via com medidas repetidas, utilizando grupos (CG e PS sob diferentes
tratamentos) e intervalos (pré e pós pareamento som-choque) como fatores.
Para o teste de CMC foi realizada uma análise de variância (ANOVA) de uma
via, utilizando grupo como fator. Para o teste de CMS foi realizada uma análise
de variância (ANOVA) de duas via com medidas repetidas, utilizando grupos
X
X
X

56
(CG e PS sob diferentes tratamentos) e intervalos (pré e pós apresentação de
som) como fatores. Análises post hoc foram feitas através do Teste de Duncan,
com nível de significância de p < 0,05.
Histologia
Após os testes comportamentais, a fim de verificar se as cânulas foram
posicionadas corretamente, os animais receberam injeções bilaterais de
corante (azul de metileno 1%) com o mesmo procedimento realizado para a
infusão das soluções, descrito anteriormente. Em seqüência, os animais foram
sacrificados por meio de dose letal do anestésico hidrato de cloral. O encéfalo
foi retirado, armazenado em freezer (-80oC) e, posteriormente, a formação
hipocampal foi seccionada em fatias de 20 μm para confimar as coordenadas
descritas, de acordo com o atlas de Paxinos & Watson (1998, Figura 15).
Figura 15: Localização histológica de implante de cânula no hipocampo dorsal.

57
4.5.1. Resultados
Treino CM: os dados estão apresentados na Figura 16. ANOVA de duas vias
com medidas repetidas revelou efeitos dos fatores grupo (F(5,45)=3,7395, p <
0,05) e intervalo (pré e pós pareamento) (F(1,45)=458,36, p < 0,05), e uma
interação significativa entre eles (F(5,45)=3,0276, p < 0,05). A análise a
posteriori demonstrou que todos os grupos apresentaram mais resposta de
congelamento após os pareamentos som-choque (p < 0.05). Por outro lado, os
animais privados de sono apresentaram menos resposta de congelamento do
que os animais controles, independentemente do tratamento recebido, durante
o intervalo pós pareamento som-choque (p < 0.05).
Figura 16: Tempo médio por minuto de congelamento de animais privados de sono tratados
com injeção intrahipocampal de aniracetam durante o treino de condicionamento de medo
(média ± erro padrão em segundos). CG = controle gaiola, PS = privação de sono, vh = veículo,
ani 0,1 = 0,1 ug/ul aniracetam, ani 1,0 = 1,0 ug/ul aniracetam. * p < 0,05 vs. intervalo pré
pareamento som-choque. # p < 0,05 vs. grupo CG sob mesmo tratamento (teste de Duncan).

58
Teste CMC: os dados estão apresentados na Figura 17. Foi encontrado efeito
do fator grupo (F(5, 45)=11,206, p < 0,01). A análise a posteriori mostrou que o
tempo médio de congelamento dos animais privados de sono foi menor do que
o tempo de congelamento dos animais controles (teste de Duncan, p < 0,01),
independente do tratamento.
Figura 17: Tempo médio por minuto de congelamento de animais privados de sono tratados
com injeção intrahipocampal de aniracetam durante o teste de condicionamento de medo ao
contexto (média ± erro padrão em segundos). CG = controle gaiola, PS = privação de sono, vh
= veículo, ani 0,1 = 0,1 ug/ul aniracetam, ani 1,0 = 1,0 ug/ul aniracetam. * p < 0,05 vs. animais
controles sob o mesmo tratamento, na mesma sessão (teste de Duncan).
Teste CMS: os dados estão apresentados na Figura 18. Não foi encontrado
efeito do fator grupo (F(5, 45)=1,9491, p=0,1049), foi encontrada efeito do fator
intervalo (antes e depois da apresentação do som) (F(1, 45)=177,63, p < 0,01)
e não foi encontrado efeito de interação dos fatores (F(5, 45)=0,4167,
p=0,8346). A análise a posteriori do fator período mostrou que o tempo médio
*
(10) (8) (8) (8) (7) (10)

59
de congelamento durante o intervalo pré som é menor do que o tempo de
congelamento durante o intervalo pós som (teste de Duncan, p < 0,01).
Figura 18: Tempo médio por minuto de congelamento de animais privados de sono tratados
com injeção intrahipocampal de aniracetam durante o teste de condicionamento de medo ao
som (média ± erro padrão em segundos). CG = controle gaiola, PS = privação de sono, vh =
veículo, ani 0,1 = 0,1 ug/ul aniracetam, ani 1,0 = 1,0 ug/ul aniracetam. * p < 0,05 vs. intervalo
pré som (teste de Duncan).
4.5.2. Discussão do Experimento Complementar
O experimento demonstrou que, mesmo administrando aniracetam na
região hipocampal, o fármaco não afetou o desempenho dos animais na tarefa
de condicionamento clássico de medo. Paralelamente, nossos dados mostram
uma redução do tempo de congelamento dos animais privados de sono tanto
após o treino da tarefa como no teste de condicionamento de medo ao
contexto, ao passo que não foi encontrada diferença de desempenho entre os
grupos durante o teste de condicionamento de medo ao som.
* * *
* * *
(10) (10) (8) (8) (8) (7)

60
Estudos prévios mostram que ratos privados de sono podem exibir
respostas comportamentais distintas de animais controles a condições
aversivas, como choque e pressão nas patas (Onen et al., 2000; Dametto et al.,
2002), o que pode influenciar sobremaneira a evocação da tarefa de
condicionamento clássico de medo. Por outro lado, o desempenho dos animais
privados foram diferentes nas sessões de condicionamento de medo ao
contexto e ao som em comparação aos animais controles. Dessa forma,
confirmamos que os prejuízos de memória induzidos pela privação de sono são
específicos para memórias hipocampo dependentes, de acordo com estudos
prévios que utilizaram a tarefa de condicionamento clássico de medo
(McDermott et al., 2003; Ruskin et al. 2004).
Somados aos dados obtidos no Experimento 1-1, os resultados da
administração local de aniracetam indicam que a potenciação alostérica
induzida pelo fármaco, nas doses utilizadas (0,1 e 1 ug/ul), não é suficiente
para prevenir os prejuízos de memória hipocampo-dependentes em ratos
privados de sono. É possível que a redução da marcação de receptores AMPA
no hipocampo previna a ação antiamnéstica da infusão de aniracetam nessa
região, sendo necessária uma ativação mais consistente desses receptores
com doses mais elevadas do potenciador. Por outro lado, conforme observado
no Experimento 1-1, uma possibilidade alternativa reside na necessidade de
ativação de outras subáreas da formação hipocampal, além da região CA1,
para que seja produzida melhora de memórias emocionais aversivas
contextuais. Mais estudos serão necessários para investigar essa hipótese.

61
5. SEGUNDA ETAPA: AVALIAÇÃO DO RITMO TETA
HIPOCAMPAL DE RATOS PRIVADOS DE SONO
DURANTE A TAREFA DE RECONHECIMENTO DE
OBJETOS
O principal objetivo dessa etapa foi investigar o ritmo teta hipocampal de
ratos privados de sono durante a aprendizagem de uma tarefa. De maneira
secundária, também foram avaliados a influência da privação de sono na tarefa
de reconhecimento de objetos, bem como o efeito rebote entre as sessões da
tarefa. Todos os experimentos desta segunda etapa foram realizados durante
estágio de doutorado sanduíche, no laboratório CNRS UMR 5167 Physio-
pathologie des réseaux neuronaux responsables du cycle veille-sommeil (Lyon,
França), dirigido pelo Dr Pierre-Hervé Luppi e com supervisão do Dr Damien
Gervasoni. Dessa forma, algumas alterações metodológicas serão evidentes
ao longo da leitura dessa etapa em relação à anterior (e.g. procedimento de
privação de sono).
5.1. METODOLOGIA GERAL
Sujeitos
Foram utilizados seis ratos Long-Evans machos, com 2,5 meses de
idade, (Harlan Laboratories). Os animais foram mantidos em condições

62
controladas de temperatura (22-24 ºC) e ciclo claro escuro de 12/12 h (período
claro iniciado às 7:00 h), com livre acesso ao alimento e água.
Técnica para registros simultâneos com arranjos de multi-eletrodos
Para realizar esses registros, utilizamos arranjos de feixes de
microeletrodos (microfios de tugstênio, revestidos de teflon, manufaturados em
nosso laboratório), que permitiram amostragem contínua de atividade neuronal
em uma área de 2 mm2 de tecido cerebral por estrutura (Nicolelis et al. 1997;
Kralik et al., 2001). Usando essa abordagem, nós obtemos registros
simultâneos de populações de neurônios individuais localizados no córtex
somatossensorial S1 e nas regiões hipocampais de CA1 e giro denteado.
Os arranjos de microeletrodos implantáveis foram manufaturados em
laboratório. Os microfios foram implantados em arranjos de 16 microfios por
peça, espaçados em intervalos de 250 μm. Esses microfios são acoplados
juntamente com fios terra a conectores Omnetics (34 contatos cada). Os
conectores são cimentados em conjunto com os microfios formando o implante.
O implante foi subdividido para ser inserido em áreas corticais e subcorticais
(Figura 19), incluindo o córtex S1 (matriz de 8x2 eletrodos) e a formação
hipocampal (matriz de 4x4 eletrodos).
Implante cirúrgico dos Arranjos de Eletrodos
A acurácia da implantação dos eletrodos durante a cirurgia foi
assegurada pelo posicionamento estereotáxico de microfios e pelo registro
contínuo de atividade neural posterior à cirurgia. Especificamente para o
hipocampo, o posicionamento dos eletrodos foi guiado pelo perfil de

63
profundidade de atividade neuronal corrente, e pela presença de ritmo teta em
potenciais locais de campo registrados durante o experimento. Esses
procedimentos geralmente fornecem uma estimativa acurada do
posicionamento dos microfios em seus alvos. A reconstrução histológica para
determinar sítios específicos dos eletrodos foi realizada após todos os
experimentos de registro serem concluídos, por meio da inspeção de seções
coronais cerebrais coradas em vermelho neutro, com referência a planos
anatômicos definidos.
Identificação em tempo real de estados de sono e vigília
Um algoritmo de detecção de estado de sono-vigília foi utilizado,
baseado num algoritmo previamente desenvolvido (Gervasoni et al. 2004). Em
resumo, sinais de potenciais de campo locais foram pré amplificados (500X),
filtrados (0.3–400 Hz) e digitalizados a 500 Hz (Multi-Neuron Acquisition
Processor, Plexon, EUA). Em seguida, estes sinais foram recaptados do
computador de registro por meio do programa PlexNet (Plexon Inc, Dallas, TX).
Um computador cliente acessou os sinais de potenciais de campo por meio do
programa Matlab (MathWorks, Natick, MA). Uma variável tampão é gerada
para processar os potenciais de campo mais recentes registrados, enquanto os
dados recém-processados são armazenados. Após um segundo completo de
registro ser obtido, uma transformação de Fourier é aplicada para obter a
densidade espectral e os valores das faixas de amplitude espectral. Em
seguida, duas razões de amplitude de espectro foram calculadas integrando o
valor absoluto da amplitude espectral para cada faixa de freqüência
selecionada: 0.5-20/0.5-55 Hz para a razão 1, e 0.5-4.5/0.5-9 Hz para a razão

64
2. As medidas dessas razões são projetadas para produzir valores
normalizados entre 0 e 1, ou seja, a faixa de frequência do numerador era
incluída no denominador para gerar distribuições simétricas. Análises de
componentes principais foram aplicados em cada razão obtida para todos os
canais, e o primeiro componente principal foi utilizado como a medida da razão
em questão. As duas razões obtidas para cada segundo foram diagramadas
em um plano, de forma a produzir três agrupamentos principais de pontos,
refletindo os estados de vigília, sono de ondas lentas e sono paradoxal. O
agrupamento de sono de ondas lentas ocupa o quadrante superior direito
devido às oscilações lentas (0-20 Hz), que resultam num valor alto no eixo das
ordenadas, e às oscilações delta (1-4 Hz), que corresponde a um valor alto no
eixo das abscissas. Os agrupamentos da vigília e do sono paradoxal têm
valores menores no eixo das ordenadas comparados com o agrupamento do
sono de ondas lentas. O agrupamento da vigília é localizado à direita do sono
paradoxal em função desse último apresentar oscilações teta mais salientes (5-
9 Hz), resultando num valor baixo no eixo das abscissas (Gervasoni et al. 2004,
Figura 19D).

65
Figura 19: A e C: exemplos de arranjos de eletrodos implantados no córtex S1 (matriz de 8x2
eletrodos) e no hipocampo (matriz de 4x4 eletrodos); B: amostra da localização do
posicionamento dos eletrodos por reconstrução histológica na formação hipocampal; D:
diagrama contendo agrupamentos de estados do ciclo sono-vigília (QW: vigília quieta, AE:
vigília ativa, SWS: sono de ondas lentas, REM: sono paradoxal, IS: sono intermediário, WT:
movimento de vibrissas), baseados nas análises de componente das razão 1 (r1, 0.5-20/0.5-55
Hz) e razão 2 (r2, 0.5-4.5/0.5-9 Hz) dos valores obtidos do espectro de frequência de potenciais
de campo locais por segundo.
Habituação à Arena de Registro
Sete dias após a cirurgia de implante de arranjos de multi-eletrodos, os
animais foram habituados à arena de registro (50 cm x 50 cm x 35 cm), durante
quatro dias (4-6 h por sessão diária), e em seguida foram coletados registros

66
basais de disparos neuronais e potenciais de campo locais durante oito horas,
a partir das 10:00 h.
5.1.1. Experimento 1 – Efeitos da Privação de Sono na Tarefa de
Reconhecimento de Objetos
O objetivo desse experimento foi avaliar o desempenho de ratos privados
de sono na tarefa de reconhecimento de objetos (TRO), em comparação a uma
condição prévia sem privação de sono dos mesmos animais. A escolha dessa
tarefa foi baseada na facilidade de executá-la na mesma arena de registro, e
por ser considerada dependente da formação hipocampal (Dere et al., 2007).
Além disso, verificamos os efeitos da privação de sono sobre os estados do
ciclo sono-vigília durante o intervalo entre sessões da tarefa.
Tarefa de Reconhecimento de Objetos
Duas horas após o início de uma sessão de registro basal, os animais
foram expostos a um par de objetos por duas sessões consecutivas de 10 min,
separadas em intervalos de 4 h (Figura 20). Na sessão 1, dois objetos idênticos
entre si foram apresentados, ao passo que na sessão 2 foram introduzidos dois
objetos diferentes, sendo um deles idêntico ao par de objetos da sessão 1
(Figura 20). Nesse paradigma, espera-se que os animais diferenciem o objeto
familiar do novo, por meio do aumento de exploração a este último durante a
sessão 2 (Dere et al 2007), definida pela aproximação do focinho do animal a
menos de 2 cm do objeto. Os objetos são constituídos por peças de plástico

67
similares a Lego (Fig 20), divididos em diferentes conjuntos de acordo com a
forma e textura das peças (lisa, perfurada, rugosa), afixados em plataformas
posicionadas em cantos diagonalmente opostos na arena de registro.
Figura 20: Acima, desenho experimental da tarefa de reconhecimento de objetos; abaixo,
exemplos de objetos utilizados na tarefa com diferentes formas e texturas.
Procedimento de Privação de Sono
Algumas semanas após a primeira exposição à tarefa de
reconhecimento de objetos, os animais foram individualmente submetidos a um
protocolo de privação de sono por 72 h em plataformas múltiplas, de acordo
com protocolo de Ravassard et al, (2009). Cada animal foi colocado em um
tanque de água contendo três plataformas estreitas espaçadas 7-8 cm entre si.
Água e ração foram disponibilizadas ad libitum. Ao final do procedimento cada
animal foi novamente exposto à TRO, sendo pré-habituado à arena por 30 min
antes do início da Sessão 1 (Figura 21). A fim de evitar a habituação dos
animais às características dos objetos, foi utilizada uma nova série de objetos,
2

68
com formatos e texturas diferentes da primeira apresentação à tarefa. Por meio
do acompanhamento dos registros de potenciais de campo locais e da
observação do comportamento do animal, foi possível acompanhar o rebote de
sono do animal especificamente durante o intervalo entre sessões.
Figura 21: A e B: amostras de registros de disparos neuronais e potenciais de campo locais
com visualização simultânea do comportamento do animal; C: desenho experimental do
estudo. Animais foram habituados á arena de registro 2 h antes da primeira sessão da tarefa de
reconhecimento de objetos.
Alterações de Estados do Ciclo Sono-Vigília após Privação de Sono
Os dados obtidos por meio da identificação em tempo de estados de
sono e vigília, especificamente durante o intervalo de 4 horas entre sessões da
tarefa de reconhecimento de objetos, foram analisados por meio do programa
NeuroExplorer e Excel. Os sinais foram classificados de acordo com Gervasoni
A B
C

69
et al. (2004), sendo determinados os estados de vigília, sono de ondas lentas e
sono paradoxal, bem como sua duração e número de episódios.
Análise Estatística
Os dados de reconhecimento de objetos para cada uma das duas
condições experimentais (controle e privação de sono) foram analisados
através de teste t de Student para uma amostra de 50%. Os dados de
exploração de objetos para cada uma das sessões da TRO foram comparados
entre as condições experimentais e analisados através de teste t de Student
pareado. Os estados de ciclo sono vigília foram analisados separadamente
para cada condição através de teste t de Student pareado. Todas as análises
foram realizadas com nível de significância de p < 0,05.

70
5.1.2. Experimento 2 – Análise da Potência de Ritmo Teta
Hipocampal durante uma Tarefa de Reconhecimento de
Objetos em Ratos Privados de Sono
Com a intenção de avaliar com maior detalhe a influência da privação de
sono sobre a aprendizagem da tarefa de reconhecimento de objetos,
procedemos à análise do rimo teta hipocampal durante o comportamento
exploratório nas duas sessões da tarefa.
Seleção de canais de registro
Foi escolhido um canal de registro hipocampal entre os 16 disponíveis,
para posterior realização da análise de potência espectral. Para tanto, para
cada um dos canais foi coletado um intervalo de registro de potenciais de
campo locais durante todo o período da sessão 1 da TRO na condição controle
(Figura 22). Dessa forma, foi obtido um espectrograma do período com a
presença de ritmo teta hipocampal proeminente. Em sequência, esses dados
foram normalizados em relação às demais frequências contidas para cada
amostra de cada animal, e posteriormente comparados por meio da razão do
pico pela média da faixa da frequência do ritmo teta hipocampal (4-12 Hz,
Figura 23). Os maiores valores obtidos determinam o canal de registro
hipocampal a ser utilizado para análises ulteriores. A localização histológica
dos canais selecionados foi confirmada ao final dos experimentos, sendo
verificado que a maioria está situada entre o giro denteado e a região CA1 (Fig
24). Esta observação corrobora achados que demonstram um aumento local do

71
ritmo teta nessas regiões, em comparação à região CA3 (Montgomery et al.,
2009).
Figura 22: Registro de potenciais de campo locais hipocampais durante uma sessão da tarefa
de reconhecimento de objetos. A: trecho de registro de um canal localizado na formação
hipocampal. B: espectrograma do canal ao longo da sessão (note a predominância de ritmo
teta hipocampal na faixa de 4 a 12 Hz). C: animal exposto aos objetos da tarefa de
reconhecimento.
B C
A

72
Figura 23: Razão entre pico e média de faixa de frequências do ritmo teta hipocampal (4-12
Hz). Os canais hipocampais (numerados de 16 a 32) de cada animal foram analisados para
seleção daquele com maior valor de pico (em vermelho) e menor valor de média (em azul).

73
Figura 24: Localização histológica do posicionamento dos canais selecionados para cada
animal. A seta indica a extremidade do eletrodo de registro.

74
Análise Espectral do Ritmo Teta Hipocampal
Para analisar os potenciais de campo locais, uma transformação rápida
de Fourier foi realizada de 0,1 a 50 Hz, com uma resolução de 0,12255 Hz para
cada segundo de atividade exploratória durante as duas sessões da TRO. A
potência da faixa de frequência de interesse foi dividida pela potência da faixa
de 0,1 a 50 Hz, de forma a obtermos a potência normalizada em porcentagem.
Analisamos especificamente a potência relativa do ritmo teta hipocampal na
faixa de 4 a 12 Hz, bem como o valor máximo (pico) dessa faixa. Além disso,
foram também analisadas as potências relativas do teta 1 (8-12 Hz) e do teta 2
(4-8 Hz).
Análise Estatística
Os dados de potência de ritmo teta (total, teta 1 e teta 2) e pico de
frequência obtidos para as duas condições experimentais (controle e privação
de sono), foram analisados através de teste t de Student pareado, com nível de
significância de p < 0,05.

75
5.2. RESULTADOS
5.2.1. Experimento 1 – Efeitos da Privação de Sono na Tarefa de
Reconhecimento de Objetos
Efeitos da Privação de Sono na Tarefa de Reconhecimento de Objetos
Sessão 1: os dados estão apresentados nas Figuras 25 e 26. Não houve
preferência de exploração dos objetos familiares tanto na condição controle
(teste t de Student para uma amostra, p = 0,32) como após privação de sono
por 72 h (teste t de Student para uma amostra, p = 0,55) (Figura 25). Por outro
lado, a comparação do tempo total de exploração (Figura 26) mostrou que os
animais na condição controle apresentaram mais comportamento exploratório
do que após privação de sono (teste t de Student pareado, p < 0,05).
Sessão 2: os dados estão apresentados nas Figuras 27 e 28. Na condição
controle, os animais exploraram mais o objeto novo em relação ao acaso (teste
t de Student para uma amostra de 50%, p < 0,05). Após a privação de sono por
72 h, no entanto, não houve diferença de exploração do objeto novo em
relação ao objeto familiar (teste t de Student para uma amostra, p = 0,95). A
comparação do tempo total de exploração nas duas situações (Figura 28)
mostrou que os animais na condição controle apresentaram mais
comportamento exploratório do que na condição após privação de sono (teste t
de Student pareado, p < 0,05).

76
Figura 25: Tempo relativo de exploração de objetos idênticos durante a sessão 1 da tarefa de
reconhecimento de objetos, em condição controle (A) e após privação de sono por 72 h (B).
Figura 26: Tempo total de exploração dos objetos durante a sessão 1 da tarefa em condição
controle (CT) e após privação de sono por 72 h (PS). * p < 0,05 vs. condição controle (teste t de
Student pareado).
*
CT PS

77
Figura 27: Tempo relativo de exploração de objetos durante a sessão 2 da tarefa de
reconhecimento de objetos, em condição controle (A) e após privação de sono por 72 h (B). * p
< 0,05 vs. 50% (teste t de Student para uma amostra).
Figura 28: Tempo total de exploração dos objetos durante a sessão 2 da tarefa em condição
controle (CT) e após privação de sono por 72 horas (PS). * p < 0,05 vs. condição CT (teste t de
Student pareado).
*
*
CT PS

78
Alterações de Estados do Ciclo Sono-Vigília após Privação de Sono
Duração das fases do ciclo sono-vigília: os resultados estão apresentados na
figura 29. Foi encontrada uma diferença significativa na duração de sono
paradoxal entre as condições controle e privado de sono: a condição PS
demonstrou aumento de sono paradoxal em relação à condição CT (teste t de
Student pareado, p < 0,05). Com relação à duração de sono de ondas lentas,
foi encontrada uma redução significativa das condição PS em comparação à
condição CT (teste t de Student pareado, p < 0,05). Não foi encontrada
diferença significativa com relação à duração total da vigília (teste t de Student
pareado, p > 0,5).
Episódios de Sono Paradoxal: os resultados estão apresentados nas figuras
30A e 30B. Não foram encontradas diferenças significativas no número de
episódios e na duração média de episódio de sono paradoxal (teste t de
Student pareado, p > 0,05).
Episódios de Sono de Ondas Lentas: os resultados estão apresentados nas
figuras 30C e 30D. Não foram encontradas diferenças significativas no número
de episódios ou na duração média de episódio de sono de ondas lentas entre
as condições PS e CT (teste t de Student pareado, p > 0,05).

79
Figura 29: Efeitos da privação de sono no ciclo sono-vigília durante intervalo de 4 horas entre
sessões da tarefa de reconhecimento de objetos. CT = condição controle; PS = privação de
sono por 72 horas antes da tarefa. * p < 0,05 vs. condição CT (teste de Student pareado).
0
25
50
75
100
125
150
sono paradoxal sono de ondas lentas vigília
Du
raçã
o t
ota
l (m
in)
duração dos estados do ciclo sono-vigília
CT
PS
*
*

80
Figura 30: Efeitos da privação de sono em parâmetros específicos dos estados de sono. A:
número de episódios de sono paradoxal (SP); B: duração média de episódio de SP; C: número
de episódios de sono de ondas lentas (SOL); D: duração média de episódio de SOL. CT =
condição controle; PS = privação de sono por 72 horas antes da tarefa. Nenhuma diferença
significativa foi encontrada (teste t de Student pareado).
A B
C D

81
5.2.2. Experimento 2 – Análise Da Potência De Ritmo Teta
Hipocampal Durante Uma Tarefa De Reconhecimento De
Objetos Em Ratos Privados De Sono
Potência de Teta Total (4-12 Hz): os resultados estão apresentados na figura
32A. Na condição CT, foi encontrada redução significativa na potência de teta
na sessão 2 em relação à sessão 1 (teste t de Student pareado, p > 0,05). Foi
encontrada aumento na potência de teta na sessão 2 na condição PS em
relação à condição CT (teste t de Student pareado, p > 0,05).
Potência de Teta tipo 1 (8-12 Hz): os resultados estão apresentados na figura
32B. Foi encontrado aumento na potência de teta tipo 1 na sessão 2 na
condição PS em relação à condição CT (teste t de Student pareado, p > 0,05).
Potência de Teta tipo 2 (4-8 Hz): os resultados estão apresentados na figura
32C. Na condição PS, foi encontrado aumento significativo na potência de teta
tipo 2 na sessão 2 em relação à sessão 1 (teste t de Student pareado, p >
0,05). Foi encontrada diminuição na potência de teta tipo 2 na sessão 1 na
condição PS em relação à condição CT (teste t de Student pareado, p > 0,05).
Pico de frequência de teta: os resultados estão apresentados na figura 33. Não
foram encontradas diferenças significativas entre as condições ou entre as
sessões. (teste t de Student pareado, p > 0,05).

82
Figura 31: Espectro de potência normalizado do registro hipocampal durante as duas sessões
da tarefa de reconhecimento de objetos. CT = condição controle; PS = privação de sono por 72
horas. Valores em média ± erro padrão.

83
Figura 32: Efeitos da privação de sono sobre potência normalizada de teta total (4-12 Hz, A),
teta 1 (8-12 Hz, B), e teta 2 (4-8 Hz, C). CT = condição controle; PS = privação de sono por 72
horas. * p < 0,05 vs. condição CT na sessão 1; # p < 0,05 vs. condição CT na mesma sessão;
@ p < 0,05 vs. condição PS na sessão 1 (teste de Student pareado). Valores em média ± erro
padrão.

84
Figura 33: Pico de frequência de ritmo teta hipocampal nas condições controle (CT) e privação
de sono por 72 horas (PS) na tarefa de reconhecimento de objetos. Nenhuma diferença
significativa foi encontrada (teste t de Student pareado). Valores em média ± erro padrão.
4
5
6
7
8
9
10
11
12
Sessão 1 Sessão 2
Pic
o d
e F
rqu
ên
cia
(Hz)
CT
PS

85
5.3. DISCUSSÃO PARCIAL
Os resultados obtidos no Experimento 1 mostram que, quando ratos são
privados de sono por 72 h, estes apresentam prejuízo significativo no
desempenho da tarefa de reconhecimento de objetos. Estudos prévios têm
demonstrado que a privação de sono entre as sessões da tarefa induz piora no
desempenho da mesma (Palchykova et. al, 2006; Rolls et al., 2011; Binder et
al., 2011), ao passo que nosso trabalho descreve pela primeira vez os efeitos
da privação de sono prévia à apresentação da TRO. Nossos dados mostram
ainda que, na condição de privação de sono, há redução da atividade
exploratória tanto durante a primeira quanto na segunda sessão da tarefa.
Com relação ao intervalo entre sessões da tarefa, nossos dados
demonstraram que a prévia privação de sono por 72 h induziu aumento de
sono paradoxal e redução de sono de ondas lentas, enquanto que a duração
da vigília não foi alterada. Esses dados estão de acordo com os obtidos por
Ravassard et al. (2009), que aplicaram o mesmo procedimento de privação de
sono e avaliaram seu rebote durante 150 minutos. Não foram encontradas
diferenças significativas no número de episódios ou duração média por
episódio, tanto do sono paradoxal como do sono de ondas lentas.
Provavelmente, o pequeno número amostral de nosso experimento não
permitiu analisar com precisão a qual fator se deveu o aumento e a diminuição
destas fases. Por outro lado, uma explicação alternativa seria a de que houve
uma combinação de aumentos graduais tanto no número de episódios quanto
na duração por episódio de sono paradoxal, ao passo que o mesmo ocorreu
com a diminuição do sono de ondas lentas.

86
Um aspecto interessante quanto ao rebote de sono paradoxal,
especialmente entre sessões de uma tarefa de memória, é que ele pode estar
promovendo, ou sustentando, uma redução de sono de ondas lentas. Moreira
(2011) apresentou um argumento interessante quanto aos efeitos deletérios da
privação de sono em tarefas de memória, relacionando o rebote de sono – e
sua consequente desorganização dos estados do ciclo sono-vigília – como um
fator a ser considerado na consolidação de uma versão da tarefa de esquiva
inibitória (esquiva inibitória por múltiplas tentativa). Em sua tese, Moreira (2011)
demonstrou que ratos apresentam aumento na duração média de episódio de
sono de ondas lentas nas primeiras horas após serem treinados na tarefa de
esquiva inibitória. Por outro lado, ratos privados de sono por 96 h, treinados ou
não nessa tarefa, apresentam redução de sono de ondas lentas e aumento de
sono paradoxal durante o rebote (Machado et al., 2004, Moreira, 2011). De
forma similar, Binder et al. (2011) encontraram aumento da atividade do ritmo
delta durante o sono de ondas lentas de ratos treinados numa versão espacial
da tarefa de reconhecimento de objetos, reforçando o envolvimento dessa fase
do sono na consolidação de tarefas hipocampo dependentes. Tomados em
conjunto, esses dados sugerem que a redução do sono de ondas lentas,
verificada entre as sessões da tarefa, pode estar relacionada ao prejuízo
subsequente dos animais na discriminação do objeto novo em relação ao
antigo.
Além das alterações observadas na quantidade de exploração dos
objetos entre as condições controle e privação de sono, buscamos analisar a
atividade hipocampal durante o comportamento exploratório, a fim de estimar
se o processamento de informações encontra-se reduzido após a privação de

87
sono. Assim, os resultados do Experimento 2 mostram que a potência do ritmo
teta hipocampal não difere na primeira sessão entre as condições, ao passo
que é verificada redução do ritmo na condição controle em relação a condição
de privação de sono. Por outro lado, uma análise mais detalhada mostra que,
durante a sessão 1, a condição de PS apresenta uma redução do ritmo teta
tipo 2 em relação à condição controle. Por outro lado, foi verificado aumento do
ritmo teta tipo 1 na condição de PS em relação ao controle na sessão 2. Com
relação a alterações entre sessões, foi verificado que o ritmo teta total é
reduzido na condição controle da sessão 1 para sessão 2, ao passo que foi
observado aumento do ritmo teta tipo 2 na condição de privação de sono.
A redução do ritmo teta tipo 2 durante a primeira sessão da TRO sugere
uma relação causal entre os efeitos da PS na atividade hipocampal, associada
com traços de memória. Nesse sentido, Yang et al. (2010) demonstraram que
ratos apresentam aumento do ritmo teta tipo 2 concomitantemente ao
treinamento da versão espacial do labirinto aquático de Morris, ao passo que
ambos os efeitos são abolidos com a privação de sono. Dessa forma, é
plausível que os prejuízos de aprendizagem decorrentes da PS, tais como os
baixos índices de exploração na sessão 1 da TRO (Experimento 1), estejam
associados com uma perturbação dos mecanismos eletrofisiológicos
relacionados à plasticidade na formação hipocampal, como a redução da LTP
(Ravassard et al., 2009). Por outro lado, a ausência de diferenças no ritmo teta
tipo 2 durante a segunda sessão da TRO sugere que a perturbação induzida
pela PS é transitória, e que pode ser revertida com a oportunidade de dormir
após a privação de sono. Novamente, esta observação está de acordo com
Ravassard et al., (2009), que demonstraram que a redução da LTP hipocampal

88
induzida pela PS pode ser revertida com uma recuperação breve de 150
minutos de sono.
Os efeitos da PS relacionados ao ritmo teta tipo 1 são menos evidentes,
uma vez que este ritmo está mais envolvido com movimentos voluntários e
exploração ativa (Buzsaki et al., 2002). Em razão das análises terem sido
realizadas com registros obtidos apenas durante atividade exploratória, é
curioso constatar que há uma diferença desse ritmo na segunda sessão da
tarefa entre as condições experimentais. Uma possível explicação reside num
estudo realizado por Nerad e Bilkey (2005), que demonstraram a presença de
uma faixa específica de oscilações (10-12 Hz), denominada flutter, que está
envolvida no reconhecimento de padrões contextuais familiares. O ritmo flutter
pode ocorrer simultaneamente ao ritmo teta hipocampal, de forma a confundir a
mensuração do ritmo teta tipo 1 (Nerad e Bilkey, 2005). Dessa forma, é
plausível que a diferença encontrada entre as condições na segunda sessão
seja devida a modificações do ritmo flutter. Assim, animais na condição
controle tenderiam a apresentar um flutter reduzido na segunda sessão de
TRO, em função deste ritmo diminuir na presença de pistas contextuais novas
(i.e. a presença do objeto novo, Nerad e Bilkey, 2005). Por outro lado, quando
os animais estavam privados de sono, não puderam se habituar de forma
eficiente às pistas contextuais disponíveis na primeira sessão de TRO, devido à
baixa atividade exploratória combinada com a potência reduzida do teta tipo 2.
Dessa forma, o ritmo flutter e, consequentemente, o ritmo teta tipo 1, não
estavam modificados na segunda sessão. Mais estudos serão necessários
para investigar o envolvimento deste ritmo nos efeitos induzidos pela privação
de sono.

89
6. DISCUSSÃO GERAL
Um dos objetivos do presente estudo foi avaliar o envolvimento dos
receptores AMPA no prejuízo de desempenho de tarefas dependentes do
hipocampo. Assim, na primeira etapa foram investigados os efeitos da privação
de sono sobre aspectos neuroquímicos e moleculares dos receptores AMPA,
bem como a ação de fármacos que atuam nestes receptores visando a
prevenção do prejuízo de desempenho em tarefas de memória emocional
aversiva. Por sua vez, a segunda etapa teve como objetivo analisar a atividade
eletrofisiológica da formação hipocampal no momento em que ratos privados
de sono são expostos a uma tarefa de reconhecimento de objetos, em busca
de elementos que auxiliem a compreensão dos efeitos da privação de sono em
tarefas hipocampo dependentes.
Na primeira etapa, foi revelada a redução seletiva da marcação do
receptor AMPA na formação hipocampal após a privação de sono por 96 h, e
que tal efeito desapareceu com a oportunidade de recuperação de sono por 24
h. Estes achados estão alinhados com dados obtidos por Ravassard et al.
(2009) e parcialmente com o estudo de Hagewoud et al. (2009). É sabido que
os receptores AMPA estão envolvidos com a formação de memórias, e que
neuropatologias ou intervenções farmacológicas que resultam numa redução
de sua função estão associados com prejuízos cognitivos, seja em humanos ou
em animais. Outrossim, a evidência de redução de receptores AMPA após a
privação de sono fornece uma explicação plausível para os prejuízos de
memória nessa condição.

90
Nossas análises de hibridização in situ não revelaram alteração da
transcrição da subunidade GluR1 nas subáreas hipocampais de ratos privados
de sono. Tal ausência de efeito distingue os efeitos da privação de sono sobre
os receptores AMPA em relação ao mal de Alzheimer e a isquemia cerebral,
patologias em que são identificadas redução da transcrição das subunidades
que compõe o receptor. Ravassard et al. (2009) e Hagewoud et al. (2009)
argumentam que a redução da função dos receptores após a privação de sono
é causada por processos pós transcricionais, tais como ancoramento de
receptores AMPA à membrana do terminal pós sináptico e a fosforilação de
aminoácidos que modulam a atividade do receptor.
A evidência da redução de receptores AMPA hipocampais após PS
direcionou a busca por estratégias farmacológicas que pudessem prevenir os
prejuízos induzidos por essa condição na tarefa de esquiva inibitória. Dessa
forma, duas abordagens principais foram propostas: aumento da função dos
receptores AMPA pelo tratamento com um potenciador (aniracetam), e a
prevenção da redução dos receptores por meio do tratamento crônico com um
antagonista (GYKI 52466). Dentre elas, apenas o tratamento sistêmico com
aniracetam surtiu o efeito esperado, e apenas na maior dose utilizada. Estes
resultados encontram consonância com um estudo realizado em humanos
privados de sono, cujos prejuízos cognitivos foram revertidos com uma dose
alta de potenciador de receptores AMPA (Boyle et al., 2011).
Os resultados obtidos pelo tratamento com aniracetam motivaram a
realização de um experimento complementar que conjugasse testes de
memória hipocampal vs. não hipocampal e um tratamento local no hipocampo
de ratos privados de sono. Ao injetar aniracetam diretamente no hipocampo de

91
ratos privados de sono, logo antes de serem treinados numa tarefa de
condicionamento de medo, esperávamos encontrar melhora de desempenho
no teste de condicionamento de medo ao contexto. Ao contrário de nossas
expectativas, não encontramos efeitos do fármaco, o que nos sugere que
talvez seja necessário o tratamento de todas as subáreas da formação
hipocampal para que ratos privados de sono possam desempenhar bem o
módulo contextual do condicionamento de medo, haja vista que a
administração da droga se restringiu à porção dorsal de CA1. Por sua vez, o
teste de condicionamento de medo ao som confirmou a especificidade do
prejuízo de memória dos ratos privados de sono, ao não ser encontradas
diferenças de desempenho dos animais controles e privados.
Os dados obtidos na primeira etapa fornecem as bases para uma
expansão da reversão de efeitos deletérios da PS sobre a atividade
hipocampal. Por exemplo, diversos estudos de potenciação sináptica de longo
prazo (LTP) mostram que esta encontra-se reduzida no hipocampo de ratos
privados de sono (Campbell et al., 2002; Mcdermott et al., 2003, 2006).
Considerando as evidências de que a aplicação de aniracetam facilita a
potenciação sináptica das subáreas hipocampais (Rao et al. 2001; Satoh et al.
1986; Arai e Lynch 1992), e de que a tarefa de esquiva inibitória compartilha
efeitos similares a uma LTP hipocampal (Whitlock et al., 2006), seria
interessante verificar se o mesmo tratamento com aniracetam, aplicado no
experimento 2-1 da primeira etapa, poderia ser replicado num modelo
eletrofisiológico putativo de formação de memória em ratos privados de sono.
Um aspecto importante da atividade hipocampal é seu ritmo teta,
geralmente associado com processamento de informações do ambiente e

92
desempenho cognitivo. O ritmo teta hipocampal está relacionado tanto a
tarefas de memórias como a potenciação sináptica de longo prazo, haja vista
que uma das formas de indução de LTP é a estimulação sob a mesma faixa do
ritmo teta tipo 2 (Greenstein et al., 1988; Larson et al., 1986). No entanto, há
poucos dados na literatura relacionando os prejuízos de memória decorrentes
da privação de sono com modificações do ritmo teta hipocampal (Yang et al.,
2011). Em vista disso, realizamos uma série de experimentos na segunda
etapa, no intuito de investigar o ritmo teta de animais privados de sono durante
a aprendizagem e evocação de uma tarefa de reconhecimento de objetos.
Como esperado, pudemos constatar que a privação de sono por 72 h induziu
prejuízo no desempenho da tarefa, tanto durante a sessão de treino como na
sessão de teste, sendo observada uma redução do comportamento
exploratório. Além disso, verificamos que o rebote de sono entre as sessões da
tarefa induziu aumento de sono paradoxal e redução do sono de ondas lentas.
Este último efeito pode estar implicado diretamente com o desempenho dos
animais na tarefa, uma vez que evidências recentes apontam o envolvimento
do sono de ondas lentas na consolidação de tarefas hipocampo dependentes,
como a esquiva inibitória (Moreira, 2011) e a versão espacial de
reconhecimento de objetos (Binder et al., 2011).
Com relação aos efeitos da privação de sono por 72 h sobre o ritmo teta
hipocampal na TRO, verificamos que a potência do teta tipo 2 está reduzida
durante a atividade exploratória da primeira sessão da tarefa. Uma vez que o
protocolo de privação de sono de nosso estudo foi idêntico ao de Ravassard et
al. (2009), e ao considerar os efeitos de antagonistas de receptores AMPA na
redução da potência do ritmo teta hipocampal, é plausível supor que haja uma

93
associação entre a redução da função de receptores AMPA após a privação de
sono e a diminuição da potência do ritmo teta tipo 2. Esta hipótese é fortalecida
com a evidência de que não foi encontrada redução do ritmo teta tipo 2 na
atividade exploratória durante a segunda sessão da tarefa em animais privados
de sono. Provavelmente, a oportunidade de rebote de sono entre as sessões
da tarefa promoveu a reversão dos efeitos da PS sobre os receptores AMPA,
de forma a normalizar o ritmo teta tipo 2 na segunda sessão.
Interessantemente, há evidências de que a injeção intrahipocampal de
aniracetam promove um aumento da potência do ritmo teta tipo 2, bem como
melhora de desempenho em uma tarefa de memória operacional espacial, em
ratos tratados com antagonista histaminérgico (Masuoka et al, 2007).
Os dados referentes ao ritmo teta tipo 1 mostram que a condição de PS
induz um aumento de potência desta faixa de frequência na segunda sessão
da tarefa. Em um primeiro momento, tais resultados eram inesperados, haja
vista que este ritmo está envolvido com atividade exploratória. Por outro lado,
nossos dados parecem estar mesclados a um outro tipo de ritmo, denominado
flutter (Nerad e Bilker, 2005). Este ritmo, ainda pouco estudado, diminui de
intensidade conforme o animal encontra elementos novos em seu ambiente. Ao
considerar que os animais privados de sono apresentam um prejuízo de
exploração na primeira sessão da tarefa, associado com uma redução tanto de
receptores AMPA como da potência do ritmo teta tipo 2, parece coerente supor
que não seja detectada uma modificação de elementos contextuais na
condição de PS. Animais na condição controle, por outro lado, estariam mais
habituados aos elementos familiares da primeira sessão, de forma a explorar

94
mais o objeto novo na segunda sessão e, por conseguinte, detectar o novo
elemento do ambiente.
Tomados em conjunto, os dados da primeira e segunda etapas mostram
que a privação de sono induz efeitos seletivos e transitórias em características
neuroquímicas e eletrofisiológicas da formação hipocampal. Tais efeitos estão
intrinsecamente associados com prejuízos em tarefas de memória hipocampo
dependentes, e sugerem uma possível terapia para reverter tais efeitos por
meio de potenciadores de receptores AMPA. Experimentos futuros serão
necessários para agregar estas evidências e aprofundar as implicações do
presente estudo.

95
7. CONCLUSÕES
1. A privação de sono por 96 h induz redução da marcação de receptores
AMPA em subáreas da formação hipocampal, mas não em outras áreas
relacionadas com a formação de memórias.
2. A recuperação de 24 h reverte os efeitos da privação de sono sobre os
receptores AMPA hipocampais.
3. A privação de sono por 96 h não modifica a transcrição da subunidade
GluR1.
4. O tratamento agudo e sistêmico com o potenciador de receptores AMPA
aniracetam promove reversão dos efeitos deletérios da privação de sono
sobre a tarefa de esquiva inibitória.
5. Os tratamentos agudo e crônico com o antagonista de receptores AMPA
GYKI 52466 não interferem com o prejuízo de desempenho na tarefa de
esquiva inibitória de ratos privados de sono.
6. A privação de sono por 72 h induz prejuízo de desempenho da tarefa de
reconhecimento de objetos, associado com alterações do ritmo teta
hipocampal e seus componentes (tipos 1 e 2).

96
8. REFERÊNCIAS BIBLIOGRÁFICAS
Albensi, B. C.; Oliver, D.R.; Toupin, J.; Odero, G.(2007). Electrical stimulation protocols for
hippocampal synaptic plasticity and neuronal hyper-excitability: Are they effective or relevant?
Exp Neurol, 204( 1), 1-13.
Arai, A.; Lynch, G. (1992). Factors regulating the magnitude of long-term potentiation induced
by theta pattern stimulation. Brain Res, 598( 1-2), 173-84.
Aserinsky, E.; Kleitman, N. (1953). Regularly occurring periods of eye motility, and concomitant
phenomena, during sleep. Science, 118 ( 3062), 273-4.
Bartolini, L.; Casamenti, F.; Pepeu, G. (1996). Aniracetam restores object recognition impaired
by age, scopolamine, and nucleus basalis lesions. Pharmacol Biochem Behav, 53(2), 277-83.
Bear, M. F.; Malenka, R. C. (1994). Synaptic plasticity: LTP and LTD. Curr Opin Neurobiol, 4,
(3), 389-99.
Berger, H. (1929). Über das Elektrenkephalogramm des Menschen. Archiv für Psychiatrie und
Nervenkrankheiten, 87, 44.
Berry, S. D.; Swain, R. A. (1989). Water deprivation optimizes hippocampal activity and
facilitates nictitating membrane conditioning. Behav Neurosci, 103( 1), 71-6.
Bettendorff, L.; Sallanon-Moulin, M.; Touret, M.; Wins, P.; Margineanu I.; Schoffeniels, E.
(1996). Paradoxical sleep deprivation increases the content of glutamate and glutamine in rat
cerebral cortex. Sleep, 19( 1), 65-71.
Binder, S.; Baier, P.C.; Mölle, M.; Inostroza, M.; Born, J.;, Marshall, L. (2011). Sleep enhances
memory consolidation in the hippocampus-dependent object-place recognition task in rats.
Neurobiol Learn Mem.
Bittencourt, L.R.; Santos-Silva, R.; Taddei, J.A.; Andersen, M.L.; de Mello, M.T.; Tufik, S.
(2009). Sleep complaints in the adult Brazilian population: a national survey based on
screening questions. J Clin Sleep Med, 5(5), 459-63.
Bliss, T. V.; Collingridge, G. L. (1993).A synaptic model of memory: long-term potentiation in the
hippocampus. Nature, 361( 6407), 31-9.
Block, F.; Schwarz, M. (1997). Correlation between hippocampal neuronal damage and spatial
learning deficit due to global ischemia. Pharmacol Biochem Behav, 56(4), 755-61.

97
Bódizs, R.; Kántor, S.; Szabó, G.; Szûcs, A.; Erõss, L.; Halász, P. (2001). Rhythmic
hippocampal slow oscillation characterizes REM sleep in humans. Hippocampus, 11(6), 747-53.
Bonini, J.S.; Rodrigues, L.; Kerr, D.S.; Bevilaqua, L.R.; Cammarota, M.; Izquierdo, I. (2003).
AMPA/kainate and group-I metabotropic receptor antagonists infused into different brain areas
impair memory formation of inhibitory avoidance in rats. Behav Pharmacol, 14(2), 161-6.
Bonnet, M. H.; Arand, D. L. (2003). Clinical effects of sleep fragmentation versus sleep
deprivation. Sleep Med Rev, 7( 4), 297-310.
Boyle, J.; Stanley, N.; James, L.M.; Wright, N.; Johnsen, S.; Arbon, E.L.; Dijk, D.J.(2011).
Acute sleep deprivation: the effects of the AMPAKINE compound CX717 on human cognitive
performance, alertness and recovery sleep. J Psychopharmacol.
Bueno, O. F.; Lobo, L.L.; Oliveira, M.G.; Gugliano, E.B.; Pomarico, A.C.; Tufik, S. (1994).
Dissociated paradoxical sleep deprivation effects on inhibitory avoidance and conditioned fear.
Physiol Behav, 56(4), 775-9.
Buzsaki, G. (2002). Theta oscillations in the hippocampus. Neuron, 33(3), 325-40.
Buzsaki, G.; Leung, L. W.; Vanderwolf, C. H. (1983) Cellular bases of hippocampal EEG in the
behaving rat. Brain Res, 287(2), 139-71.
Cammarota, M.; Bernabeu, R.; Izquierdo, I.; Medina, J.H. (1996). Reversible changes in
hippocampal 3H-AMPA binding following inhibitory avoidance training in the rat. Neurobiol
Learn Mem, 66(1), 85-8.
Cammarota, M.; Izquierdo, I.; Wolfman, C.; Levi de Stein, M.; Bernabeu, R.; Jerusalinsky, D.;
Medina, J.H. (1995). Inhibitory avoidance training induces rapid and selective changes in
3[H]AMPA receptor binding in the rat hippocampal formation. Neurobiol Learn Mem, 64(3), 257-
64.
Campbell, I. G.; Guinan, M. J.; Horowitz, J. M. (2002). Sleep deprivation impairs long-term
potentiation in rat hippocampal slices. J Neurophysiol, 88(2), 1073-6.
Cohen, H. B.; Dement, W. C. (1965). Sleep: changes in threshold to electroconvulsive shock in
rats after deprivation of "paradoxical" phase. Science, 150(3701), 1318-9.
Corsi-Cabrera, M.; Ponce-De-Leon, M.; Juarez, J.; Ramos, J. (1994). Effects of paradoxical
sleep deprivation and stress on the waking EEG of the rat. Physiol Behav, 55(6), 1021-7.

98
Csicsvari, J.; Hirase, H.; Czurkó, A.; Mamiya, A.; Buzsáki, G. (1999). Fast network oscillations in
the hippocampal CA1 region of the behaving rat. J Neurosci, 19(16), RC20.
Cumin, R.; Bandle, E.F.; Gamzu, E.; Haefely, W.E. (1982). Effects of the novel compound
aniracetam (Ro 13-5057) upon impaired learning and memory in rodents. Psychopharmacology
(Berl), 78(2), 104-11.
Dametto, M.; Suchecki, D.; Bueno, O.F.; Moreira, K.M.; Tufik, S.; Oliveira, M.G. (2002). Social
stress does not interact with paradoxical sleep deprivation-induced memory impairment. Behav
Brain Res, 129( 1-2), 171-8.
Danysz, W.; Essmann, U.; Bresink, I.; Wilke, R. (1994). Glutamate antagonists have different
effects on spontaneous locomotor activity in rats. Pharmacol Biochem Behav, 48(1), 111-8.
Dere, E.; Huston, J. P.; De Souza Silva, M. A. (2007). The pharmacology, neuroanatomy and
neurogenetics of one-trial object recognition in rodents. Neurosci Biobehav Rev, 31(5), 673-704.
Dewar, D.; Chalmers, D.T.; Graham, D.I.; McCulloch, J. (1991). Glutamate metabotropic and
AMPA binding sites are reduced in Alzheimer's disease: an autoradiographic study of the
hippocampus. Brain Res, 553(1), 58-64.
Dickey, C. A.; Loring, J.F.; Montgomery, J.; Gordon, M.N.; Eastman, P.S.; Morgan, D.
Selectively reduced expression of synaptic plasticity-related genes in amyloid precursor protein
+ presenilin-1 transgenic mice. J Neurosci, 23(12), 5219-26.
Drummond, S.P.; Brown, G.G.; Gillin, J.C.; Stricker, J.L.; Wong, E.C.; Buxton, R.B. (2000).
Altered brain response to verbal learning following sleep deprivation. Nature, 403(6770), 655-7.
Dubiela, F.P.; Oliveira, M.G.; Moreira, K.M.; Nobrega, J.N.; Tufik, S.; Hipólide, D.C. (2005).
Learning deficits induced by sleep deprivation and recovery are not associated with altered
[(3)H]muscimol and [(3)H]flunitrazepam binding. Brain Res, 1037(1-2), 157-63.
Dubiela, F.P.; Messias, M.F.; Moreira, K.D.; Zanlorenci, L.H.; Grassl, C.; Filho, R.F.; Nobrega,
J.N.; Tufik, S.; Hipólide, D.C. Reciprocal interactions between MK-801, sleep deprivation and
recovery in modulating rat behaviour. Behav Brain Res, 216(1), 180-5.
Dubiela, F.P.; Oliveira, M.G.; Moreira, K.M.; Nobrega, J.N.; Tufik, S.; Hipólide, D.C.(2010)
.Inverse benzodiazepine agonist beta-CCM does not reverse learning deficit induced by sleep
deprivation. Neurosci Lett, 469(1), 169-73.

99
Eichenbaum, H. (2000). A cortical-hippocampal system for declarative memory. Nat Rev
Neurosci, 1(1), 41-50.
Elger, B.; Huth, A.; Neuhaus, R.; Ottow, E.; Schneider, H.; Seilheimer, B.; Turski, L. (2005).
Novel alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionate (AMPA) receptor antagonists of
2,3-benzodiazepine type: chemical synthesis, in vitro characterization, and in vivo prevention of
acute neurodegeneration. J Med Chem, 48(14), 4618-27.
Esteban, J.A.; Shi, S.H.; Wilson, C.; Nuriya, M.; Huganir, R.L.; Malinow, R. (2003). PKA
phosphorylation of AMPA receptor subunits controls synaptic trafficking underlying plasticity.
Nat Neurosci, 6(2), 136-43.
Gervasoni, D.; Lin, S.C.; Ribeiro, S.; Soares, E.S.; Pantoja, J.; Nicolelis, M.A. (2004). Global
forebrain dynamics predict rat behavioral states and their transitions. J Neurosci, 24(49), 11137-
47.
Givens, B.; Olton, D. S. (1995). Bidirectional modulation of scopolamine-induced working
memory impairments by muscarinic activation of the medial septal area. Neurobiol Learn Mem,
63(3), 269-76.
Goosens, K. A.; Maren, S. (2001). Contextual and auditory fear conditioning are mediated by
the lateral, basal, and central amygdaloid nuclei in rats. Learn Mem, 8(3), 148-55.
Gouliaev, A. H.; Senning, A. (1994). Piracetam and other structurally related nootropics. Brain
Res Brain Res Rev, 19(2), 180-222.
Grahnstedt, S.; Ursin, R. (1985). Platform sleep deprivation affects deep slow wave sleep in
addition to REM sleep. Behav Brain Res, 18(3), 233-9.
Greenstein, Y. J.; Pavlides, C.; Winson, J. (1988). Long-term potentiation in the dentate gyrus is
preferentially induced at theta rhythm periodicity. Brain Res, 438(1-2), 331-4.
Gyertyan, I.; Gigler, G.; Simo, A. (1999). The neu roprotective and hypothermic effect of
GYKI-52466, a non-competitive alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid-
antagonist on histological and behavioural variables in the gerbil global ischemia model. Brain
Res Bull, 50(3), 179-86.
Hagewoud, R.; Havekes, R.; Novati, A.; Keijser, J.N.; Van der Zee, E.A.; Meerlo, P. (2010).
Sleep deprivation impairs spatial working memory and reduces hippocampal AMPA receptor
phosphorylation. J Sleep Res, 19(2), 280-8.

100
Harrison, Y.; Horne, J. A. (2000). Sleep loss and temporal memory. Q J Exp Psychol A, 53(1),
271-9.
Hayashi, Y.; Shi, S.H.; Esteban, J.A.; Piccini, A.; Poncer, J.C.; Malinow, R. (2000).Driving
AMPA receptors into synapses by LTP and CaMKII: requirement for GluR1 and PDZ domain
interaction. Science, 287(5461), 2262-7.
Izquierdo, I.; Medina, J. H. Memory formation: the sequence of biochemical events in the
hippocampus and its connection to activity in other brain structures. Neurobiol Learn Mem,
68(3), 285-316.
Izquierdo, I.; Medina, J.H.; Bianchin, M.; Walz, R.; Zanatta, M.S.; Da Silva, R.C.; Bueno e Silva,
M.; Ruschel, A.C.; Paczko, N. (1993). Memory processing by the limbic system: role of specific
neurotransmitter systems. Behav Brain Res, 58(1-2), 91-8.
Jin, R.; Clark, S.; Weeks, A.M.; Dudman, J.T.; Gouaux, E.; Partin, K.M. (2005). Mechanism of
positive allosteric modulators acting on AMPA receptors. J Neurosci, 25(39), 9027-36.
Jouvet, D.; Vimont, P.; Delorme, F. (1964). Study Of Selective Deprivation Of The Paradoxal
Phase Of Sleep In The Cat. J Physiol (Paris), 56(381).
Jouvet, M. (1969). Biogenic amines and the states of sleep. Science, 163(3862), 32-41.
Kandel, E. R. ; Schwartz, J. H. ; Jessell, T. M. Principles of Neural Science, quarta edição,
Editora McGraw-HillMichigan, Michigan, 2008, 1414 p.
Kim, J. J.; Fanselow, M. S. (1992). Modality-specific retrograde amnesia of fear. Science,
256(5057), 675-7.
Kostandy, B. B. (2011). The role of glutamate in neuronal ischemic injury: the role of spark in
fire. Neurol Sci.
Kralik, J.D.; Dimitrov, D.F.; Krupa, D.J.; Katz, D.B.; Cohen, D.; Nicolelis, M.A.
(2001).Techniques for long-term multisite neuronal ensemble recordings in behaving animals.
Methods, 25(2), 121-50.
Kramis, R.; Vanderwolf, C. H.; Bland, B. H. (1975). Two types of hippocampal rhythmical slow
activity in both the rabbit and the rat: relations to behavior and effects of atropine, diethyl ether,
urethane, and pentobarbital. Exp Neurol, 49(1 Pt 1), 58-85.
Larson, J.; Wong, D.; Lynch, G. (1986). Patterned stimulation at the theta frequency is optimal
for the induction of hippocampal long-term potentiation. Brain Res, 368(2), 347-50.

101
Leung, L. S.; Shen, B. (2004). Glutamatergic synaptic transmission participates in generating
the hippocampal EEG. Hippocampus, 14(4), 510-25.
Lu, Y.; Wehner, J. M. (1997). Enhancement of contextual fear-conditioning by putative (+/-)-
alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA) receptor modulators and N-
methyl-D-aspartate (NMDA) receptor antagonists in DBA/2J mice. Brain Res, 768(1-2), 197-
207.
Machado, R.B.; Hipólide, D.C.; Benedito-Silva, A.A.; Tufik, S. (2004). Sleep deprivation induced
by the modified multiple platform technique: quantification of sleep loss and recovery. Brain
Res, 1004(1-2), 45-51.
Macrides, F.; Eichenbaum, H. B.; Forbes, W. B. Temporal relationship between sniffing and the
limbic theta rhythm during odor discrimination reversal learning. J Neurosci, 2(12), 1705-17.
Maren, S.; Tocco, G.; Standley, S.; Baudry, M.; Thompson, R.F. (1993). Postsynaptic factors in
the expression of long-term potentiation (LTP): increased glutamate receptor binding following
LTP induction in vivo. Proc Natl Acad Sci U S A, 90(20), 9654-8.
Marques, N.; Menna-Barreto, L. Cronobiologia: princípios e aplicações, EDUSP/Fiocruz, São
Paulo/Rio de Janeiro , 1997, 320 p.
Masuoka, T.; Mikami, A.; Yasuda, M.; Shinomiya, K.; Kamei, C.(2007). Effects of histamine H(1)
receptor antagonists on hippocampal theta rhythm during spatial memory performance in rats.
Eur J Pharmacol, 576(1-3), 77-82.
McDermott, C.M.; Hardy, M.N.; Bazan, N.G.; Magee, J.C. (2006). Sleep deprivation-induced
alterations in excitatory synaptic transmission in the CA1 region of the rat hippocampus. J
Physiol, 570(Pt 3), 553-65.
McDermott, C.M.; LaHoste, G.J.; Chen, C.; Musto, A.; Bazan, N.G.; Magee, J.C. (2003).Sleep
deprivation causes behavioral, synaptic, and membrane excitability alterations in hippocampal
neurons. J Neurosci, 23(29), 9687-95.
McGaugh, J.L.; Introini-Collison, I.B.; Nagahara, A.H.; Cahill, L.; Brioni, J.D.; Castellano, C.
(1990). Involvement of the amygdaloid complex in neuromodulatory influences on memory
storage. Neurosci Biobehav Rev, 14(4), 425-31.
Mednick, S.C.; Nakayama, K.; Cantero, J.L.; Atienza, M.; Levin, A.A.; Pathak, N.; Stickgold, R.
(2002). The restorative effect of naps on perceptual deterioration. Nat Neurosci, 5(7), 677-81.

102
Mohammed, H.S.; Aboul Ezz, H.S.; Khadrawy, Y.A.; Noor, N.A. (2011).Neurochemical and
electrophysiological changes induced by paradoxical sleep deprivation in rats. Behav Brain Res,
225(1), 39-46.
Monaghan, D.T.; Wenthold, R.J. The Ionotropic Glutamate Receptors, Humana Press, Totowa,
1997, 378 p.
Montgomery, S. M.; Betancur, M. I.; Buzsaki, G. (2009). Behavior-dependent coordination of
multiple theta dipoles in the hippocampus. J Neurosci, 29(5), 1381-94.
Montori, S.; Dos Anjos, S.; Ríos-Granja, M.A.; Pérez-García, C.C.; Fernández-López, A.;
Martínez-Villayandre, B.(2010). AMPA receptor downregulation induced by
ischaemia/reperfusion is attenuated by age and blocked by meloxicam. Neuropathol Appl
Neurobiol, 36(5), 436-47.
Moreira, K.M.; Ferreira, T.L.; Hipolide, D.C.; Fornari, R.V.; Tufik, S.; Oliveira, M.G. (2010)
Modafinil prevents inhibitory avoidance memory deficit induced by sleep deprivation in rats.
Sleep, 33(7), 990-3.
Moreira KM; Hipólide DC; Nobrega JN; Bueno OF; Tufik S; Oliveira MG.(2003). Deficits in
avoidance responding after paradoxical sleep deprivation are not associated with altered
[3H]pirenzepine binding to M1 muscarinic receptors in rat brain. Brain Res, 977(1), 31-7.
Moreira KM. Relação do desempenho de ratos em uma tarefa de esquiva inibitória com o
padrão polissonográfico, [tese de doutorado] Universidade Federal de São Paulo, São Paulo:
2011.
Morris, G. O.; Williams, H. L.; Lubin, A.(1960). Misperception and Disorientation During Sleep-
Deprivation. Archives of General Psychiatry, 2(3), 247-254.
National Sleep Foundation. (2002). “Sleep in America” poll. 1–52. URL:
http://www.sleepfoundation.org.
Nerad, L.; Bilkey, D. K. (2005). Ten- to 12-Hz EEG oscillation in the rat hippocampus and rhinal
cortex that is modulated by environmental familiarity. J Neurophysiol, 93(3), 1246-54.
Nicolelis, M.A.; Ghazanfar, A.A.; Faggin, B.M.; Votaw, .S; Oliveira, L.M. (1997).Reconstructing
the engram: simultaneous, multisite, many single neuron recordings. Neuron, 18(4).
Nunes, J.R.; G.P.; TUFIK, S. (1994). Validation of the modified multiple platform method (MPM)
of paradoxical sleep deprivation in rats. Sleep Research, 22, 339.

103
O'Keefe, J. (1990). A computational theory of the hippocampal cognitive map. Prog Brain Res,
83), 301-12.
O'Neill, M.J.; Bleakman, D.; Zimmerman, D.M.; Nisenbaum, E.S. (2004). AMPA receptor
potentiators for the treatment of CNS disorders. Curr Drug Targets CNS Neurol Disord, 3(3),
181-94.
Onen, S.H.; Alloui, A.; Eschalier, A.; Dubray, C. (2000). Vocalization thresholds related to
noxious paw pressure are decreased by paradoxical sleep deprivation and increased after sleep
recovery in rat. Neurosci Lett. 291, 25-8.
Palchykova, S.; Winsky-Sommerer, R.; Meerlo, P.; Dürr, R.; Tobler, I. (2006). Sleep deprivation
impairs object recognition in mice. Neurobiol Learn Mem, 85(3), 263-71.
Pan, W. X.; Mcnaughton, N.(1997). The medial supramammillary nucleus, spatial learning and
the frequency of hippocampal theta activity. Brain Res, 764(1-2), 101-8.
Pape, H.C.; Narayanan, R.T.; Smid, J.; Stork, O.; Seidenbecher, T. (2005).Theta activity in
neurons and networks of the amygdala related to long-term fear memory. Hippocampus, 15(7),
874-80.
Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, fourth edition, Academic
Press, New York, 1998, 256 p.
Pearlman, C. A. (1979). Rem-Sleep and Information-Processing - Evidence from Animal
Studies. Neuroscience and Biobehavioral Reviews, 3(2), 57-68.
Pellegrini-Giampietro, D. E.; Bennett, M. V.; Zukin, R. S. (1994). AMPA/kainate receptor gene
expression in normal and Alzheimer's disease hippocampus. Neuroscience, 61(1), 41-9.
Phillips, R. G.; Ledoux, J. E. (1992). Differential contribution of amygdala and hippocampus to
cued and contextual fear conditioning. Behav Neurosci, 106(2), 274-85.
Phillips, R. G.; Ledoux, J. E. (1994). Lesions of the dorsal hippocampal formation interfere with
background but not foreground contextual fear conditioning. Learn Mem, 1(1), 34-44.
Platt, S.R. (2007). The role of glutamate in central nervous system health and disease: A
review. The Veterinary Journal, 173, 278-286.
Porrino, L.J.; Daunais, J.B.; Rogers, G.A.; Hampson, R.E.; Deadwyler, S.A.(2005). Facilitation
of task performance and removal of the effects of sleep deprivation by an ampakine (CX717) in
nonhuman primates. PLoS Biol, 3(9), e299.

104
Puma, C.; Bizot, J. C. (1999). Hippocampal theta rhythm in anesthetized rats: role of AMPA
glutamate receptors. Neuroreport, 10(11), 2297-300.
Rao, Y.; Xiao, P.; Xu, S. (2001). Effects of intrahippocampal aniracetam treatment on Y-maze
avoidance learning performance and behavioral long-term potentiation in dentate gyrus in rat.
Neurosci Lett, 298(3), 183-6.
Ravassard, P.; Pachoud, B.; Comte, J.C.; Mejia-Perez, C.; Scoté-Blachon, C.; Gay, N.;
Claustrat, B.; Touret, M.; Luppi, P.H.; Salin, P.A. (2009). Paradoxical (REM) sleep deprivation
causes a large and rapidly reversible decrease in long-term potentiation, synaptic transmission,
glutamate receptor protein levels, and ERK/MAPK activation in the dorsal hippocampus. Sleep,
32(2), 227-40.
Ribeiro, S.; Goyal, V.; Mello, C.V.; Pavlides, C. (1999). Brain gene expression during REM
sleep depends on prior waking experience. Learning & Memory, 6(5), 500-508.
Ribeiro, S.; Mello, C.V.; Velho, T.; Gardner, T.J.; Jarvis; E.D.; Pavlides, C.(2002). Induction of
hippocampal long-term potentiation during waking leads to increased extrahippocampal zif-268
expression during ensuing rapid-eye-movement sleep. Journal of Neuroscience, 22(24), 10914-
10923.
Ribeiro, S.; Nicolelis, M. A. L. (2004). Reverberation, storage, and postsynaptic propagation of
memories during sleep. Learning & Memory, 11(6), 686-696.
Rolls, A.; Colas, D.; Adamantidis, A.; Carter, M.; Lanre-Amos, T.; Heller, H.C.; de Lecea, L.
(2011). Optogenetic disruption of sleep continuity impairs memory consolidation. Proc Natl Acad
Sci U S A, 108(32), 13305-10.
Ruel, J.; Guitton, M. J.; Puell, J. L. (2002). Negative allosteric modulation of AMPA-preferring
receptors by the selective isomer GYKI 53784 (LY303070), a specific non-competitive AMPA
antagonist. CNS Drug Rev, 8(3), 235-54.
Ruskin, D.N.; Liu, C.; Dunn, K.E.; Bazan, N.G.; LaHoste, G.J. (2004).Sleep deprivation impairs
hippocampus-mediated contextual learning but not amygdala-mediated cued learning in rats.
Eur J Neurosci, 19(11), 3121-4.
Santos-Silva, R.; Bittencourt, L.R.; Pires, M.L.; de Mello, M.T.; Taddei, J.A.; Benedito-Silva,
A.A.; Pompeia, C.; Tufik, S. (2010).Increasing trends of sleep complaints in the city of Sao
Paulo, Brazil. Sleep Med, 11(6), 520-4.

105
Satoh, M.; Ishihara, K.; Iwama, T.; Takagi, H. (1986). Aniracetam augments, and midazolam
inhibits, the long-term potentiation in guinea-pig hippocampal slices. Neurosci Lett, 68(2), 216-
20.
Seidenbecher, T.; Laxmi, T.R.; Stork, O.; Pape, H.C. (2003). Amygdalar and hippocampal theta
rhythm synchronization during fear memory retrieval. Science, 301(5634), 846-50.
Shi, S.; Hayashi, Y.; Esteban, J.A.; Malinow, R. (2001). Subunit-specific rules governing AMPA
receptor trafficking to synapses in hippocampal pyramidal neurons. Cell, 105(3), 331-43.
Siegel, J. M. Do all animals sleep? Trends Neurosci, 31(4), 208-13.
Sinha, A.K.; Azevedo, R.; Chi, O.Z.; Weiss, H.R. (2004). Down-regulation of AMPA glutamate
receptors reduces cerebrocortical metabolic response to stimulation. Neurochem Res, 29(7),
1425-30.
Sinha, A.K.; Mirza, A.H.; Liu, X.; Chi, O.Z.; Weiss, H.R.(1999). Effect of upregulation of AMPA
glutamate receptors on cerebral O(2) consumption and blood flow in rat. Brain Res, 842(1), 230-
2.
Smith, C. (1995). Sleep states and memory processes. Behav Brain Res, 69(1-2), 137-45.
Smith, C.; Lapp, L. (1991). Increases in number of REMS and REM density in humans following
an intensive learning period. Sleep, 14(4), 325-30.
Smith, C.; Rose, G. M. (1997). Posttraining paradoxical sleep in rats is increased after spatial
learning in the Morris water maze. Behavioral Neuroscience, 111(6), 1197-1204.
Sommer, C.; Fahrner, A.; Kiessling, M. (2002). [3H]muscimol binding to gamma-aminobutyric
acid(A) receptors is upregulated in CA1 neurons of the gerbil hippocampus in the ischemia-
tolerant state. Stroke, 33(6), 1698-705.
Squire, L. R. (1992). Memory and the hippocampus: a synthesis from findings with rats,
monkeys, and humans. Psychol Rev, 99(2), 195-231.
Squire, L. R.; Zola-Morgan, S. (1991). The medial temporal lobe memory system. Science,
253(5026), 1380-6.
Suchecki, D.; Lobo, L.L.; Hipólide, D.C.; Tufik, S. (1998). Increased ACTH and corticosterone
secretion induced by different methods of paradoxical sleep deprivation. J Sleep Res, 7(4), 276-
81.

106
Suchecki, D.; Tufik, S. (2000) Social stability attenuates the stress in the modified multiple
platform method for paradoxical sleep deprivation in the rat. Physiol Behav, 68(3), 309-16.
Timo-Iaria, C.; Negrão, N.; Schmidek, W.R.; Hoshino, K.; Lobato de Menezes, C.E.; Leme da
Rocha, T. (1970). Phases and states of sleep in the rat. Physiol Behav, 5(9), 1057-62.
Tocco, G.; Annala, A.J.; Baudry, M.; Thompson, R.F.(1992). Learning of a hippocampal-
dependent conditioning task changes the binding properties of AMPA receptors in rabbit
hippocampus. Behav Neural Biol, 58(3), 222-31.
Ueki, A.; Miwa, C.; Miyoshi, K. (1994). Impairment in the acquisition of passive and active
avoidance learning tasks due to bilateral entorhinal cortex lesions. J Neurol Sci, 125(1), 14-21.
Van Hulzen, Z. J.; Coenen, A. M. (1981). Paradoxical sleep deprivation and locomotor activity in
rats. Physiol Behav, 27(4), 741-4.
Van Hulzen, Z. J.; Coenen, A. M. (1984).Photically evoked potentials in the visual cortex
following paradoxical sleep deprivation in rats. Physiol Behav, 32(4), 557-63.
Vanderwolf, C. H. (1969). Hippocampal electrical activity and voluntary movement in the rat.
Electroencephalogr Clin Neurophysiol, 26(4), 407-18.
Vyazovskiy, V.V.; Cirelli, C.; Pfister-Genskow, M.; Faraguna, U.; Tononi, G. (2008).Molecular
and electrophysiological evidence for net synaptic potentiation in wake and depression in sleep.
Nat Neurosci, 11(2), 200-8.
Vyazovskiy, V. V.; Tobler, I. (2005). Theta activity in the waking EEG is a marker of sleep
propensity in the rat. Brain Res, 1050(1-2), 64-71.
Walker, M. P.; Stickgold, R. (2006). Sleep, memory, and plasticity. Annu Rev Psychol, 57), 139-
66.
Westerberg, E.; Monaghan, D.T.; Cotman, C.W.; Wieloch, T.(1987). Excitatory amino acid
receptors and ischemic brain damage in the rat. Neurosci Lett, 73(2), 119-24.
Whitlock, J.R.; Heynen, A.J.; Shuler, M.G.; Bear, M.F. (2006). Learning induces long-term
potentiation in the hippocampus. Science, 313(5790), 1093-7.
Xavier GF. Alguns fatores relevantes na interpretação do teste de esquiva passiva. Implicações
para estudos de aprendizagem. [dissertação de mestrado] Universidade Federal de São Paulo:
São Paulo, 1981.

107
Yang, R.H.; Hou, X.H.; Xu, X.N.; Zhang, L.; Shi, J.N.; Wang, F.; Hu, S.J.; Chen, J.Y. (2011).
Sleep deprivation impairs spatial learning and modifies the hippocampal theta rhythm in rats.
Neuroscience, 173), 116-23.
Youngblood, B.D.; Smagin, G.N.; Elkins, P.D.; Ryan, D.H.; Harris, R.B. (1999). The effects of
paradoxical sleep deprivation and valine on spatial learning and brain 5-HT metabolism. Physiol
Behav, 67(5), 643-9.
Youngblood, B.D.; Zhou, J.; Smagin, G.N.; Ryan, D.H.; Harris, R.B. (1997). Sleep deprivation by
the "flower pot" technique and spatial reference memory. Physiol Behav, 61(2), 249-56.

108
9. ANEXO

Rua Botucatu, 572 - 1º andar – conj. 14 - CEP 04023-062 - São Paulo / Brasil Tel.: (011) 5571-1062 - 5539.7162
1
São Paulo, 11 de janeiro de 2008.
CEP 2075/07 IImo(a). Sr(a). Pesquisador(a) DÉBORA CRISTINA HIPÓLIDE Co-Investigadores: Francisco Paulino dubiela e Diva Maria Lima Disciplina/Departamento: Psicobiologia da Universidade Federal de São Paulo/Hospital São Paulo Patrocinador: AFIP, CNPq.
PARECER DO COMITÊ DE ÉTICA INSTITUCIONAL Ref: Projeto de pesquisa intitulado: “Efeitos da privação de sono sobre a plasticidade hipocampal a importância
dos receptores AMPA”.
CARACTERÍSTICA PRINCIPAL DO ESTUDO: EXPERIMENTAL - CATEGORIA D - ESTUDO CRÕNICO.
RISCOS ADICIONAIS PARA O PACIENTE: NÃO SE APLICA.
OBJETIVOS: Avaliar o papel dos receptores AMPA na modulação dos prejuízos de memória dependente de
hipocampo após a privação de sono..
RESUMO: Serão utilizados ratos Wistar machos provenientes do CEDEME da UNIFESP. Os animais serão
mantidos em condições controladas de temperatura e ciclo de claro e escuro com livre acesso ao alimento e água.
Para o procedimento de privação de sono será utilizado o método da plataforma múltipla modificado que será
utilizado para produzir uma privação de sono em animais provenientes da mesma gaiola. Esse modelo tem a
vantagem de ser mais seletivo para os efeitos da privação de sono, por reduzir o envolvimento de variáveis como o
isolamento, a instabilidade social e a imobilização presentes no método da plataforma única..
FUNDAMENTOS E RACIONAL: Diversos estudos mostram que a falta de sono interfere com a formação de novas
memórias, especialmente as dependentes do hipocampo..
MATERIAL E MÉTODO: materiais e métodos adequadamente descritos.
CRONOGRAMA: Adequado.
OBJETIVO ACADÊMICO: Doutorado.
ENTREGA DE RELATÓRIOS PARCIAIS AO CEP PREVISTOS PARA: 10/1/2009 e 10/1/2010.
O Comitê de Ética em Pesquisa da Universidade Federal de São Paulo/Hospital São Paulo ANALISOU e APROVOU
o projeto de pesquisa referenciado.
1. Comunicar toda e qualquer alteração do projeto.
2. Comunicar imediatamente ao Comitê qualquer evento adverso ocorrido durante o desenvolvimento do estudo.

Rua Botucatu, 572 - 1º andar – conj. 14 - CEP 04023-062 - São Paulo / Brasil Tel.: (011) 5571-1062 - 5539.7162
2
3. Os dados individuais de todas as etapas da pesquisa devem ser mantidos em local seguro por 5 anos para
possível auditoria dos órgãos competentes.
Atenciosamente,
Prof. Dr. José Osmar Medina Pestana Coordenador do Comitê de Ética em Pesquisa da Universidade Federal de São Paulo/ Hospital São Paulo CEP 2075/07


















