
EFEITOS DA INFECÇÃO EXPERIMENTAL COM …livros01.livrosgratis.com.br/cp147325.pdf · Á amiga de...
58
i RÔMULO DIAS NOVAES EFEITOS DA INFECÇÃO EXPERIMENTAL COM Trypanosoma cruzi SOBRE A MORFOLOGIA DO MIOCÁRDIO, PROPRIEDADES MECÂNICAS DE CARDIOMIÓCITOS ISOLADOS E TOLERÂNCIA AO EXERCÍCIO FÍSICO DE RATOS WISTAR Dissertação apresentada à Universidade Federal de Viçosa, como parte das exigências do Programa de Pós-Graduação em Biologia Celular e Estrutural, para obtenção do título de “Magister Scientiae” VIÇOSA MINAS GERAIS – BRASIL 2010
Transcript of EFEITOS DA INFECÇÃO EXPERIMENTAL COM …livros01.livrosgratis.com.br/cp147325.pdf · Á amiga de...

i
RÔMULO DIAS NOVAES
EFEITOS DA INFECÇÃO EXPERIMENTAL COM Trypanosoma cruzi SOBRE A MORFOLOGIA DO MIOCÁRDIO, PROPRIEDADES MECÂNICAS DE
CARDIOMIÓCITOS ISOLADOS E TOLERÂNCIA AO EXERCÍCIO F ÍSICO DE RATOS WISTAR
Dissertação apresentada à Universidade Federal de Viçosa, como parte das exigências do Programa de Pós-Graduação em Biologia Celular e Estrutural, para obtenção do título de “Magister Scientiae”
VIÇOSA MINAS GERAIS – BRASIL
2010

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

i
RÔMULO DIAS NOVAES
EFEITOS DA INFECÇÃO EXPERIMENTAL COM Trypanosoma cruzi SOBRE A
MORFOLOGIA DO MIOCÁRDIO, PROPRIEDADES MECÂNICAS DE
CARDIOMIÓCITOS ISOLADOS E TOLERÂNCIA AO EXERCÍCIO F ÍSICO DE
RATOS WISTAR
Dissertação apresentada à Universidade Federal de Viçosa, como parte das exigências do Programa de Pós-Graduação em Biologia Celular e Estrutural, para obtenção do título de “Magister Scientiae”
APROVADO EM: ______________________________ ______________________________ Prof. André Talvani Pedrosa Prof. Cláudio César Fonseca Universidade Federal de Ouro Preto Universidade Federal de Viçosa
______________________________ _______________________________ Prof. Antônio José Natali Prof. Thales Nicolau Prímola Gomes Universidade Federal de Viçosa Universidade Federal de Viçosa Coorientador _______________________________________________
Profª. Izabel Regina dos Santos Costa Maldonado Universidade Federal de Viçosa
Orientadora

ii
A minha amada mãe Cláudia pela luta incessante, amor incondicional e pela representação de um modelo de inspiração.

iii
AGRADECIMENTOS À Universidade Federal de Viçosa e ao Programa de Pós-Graduação em Biologia Celular e Estrutural, por todo aprendizado. À FAPEMIG, pelo apoio financeiro. À minha orientadora Profª. Izabel Regina dos Santos Costa Maldonado, pelos ensinamentos, confiança e paciência. Aos meus Co-Orientadores Prof. Clóvis Andrade Neves e Prof. Antônio José Natali, pelos ensinamentos, paciência e disposição em ajudar. Ao Prof. André Talvani Pedrosa, pelo apóio na inoculação dos animais, disposição em ajudar e incentivo para continuar a caminhada. Ao Prof. Cláudio César Fonseca, pela disposição em participar da Banca Examinadora. À Profª. Sílvia Pompolo, por gentilmente permitir a utilização de seu laboratório para aquisição das imagens utilizadas. A todos os professores do Programa de Pós-Graduação em Biologia Celular e Estrutural, por todos os ensinamentos e incentivo. A todos os funcionários do Programa de Pós-Graduação em Biologia Celular e Estrutural, em especial Beth e Diana, pela ótima convivência, paciência e auxílio em todos os momentos necessários. Aos amigos do laboratório de Biologia Estrutural: Kener, Luíz Bozi, Marlí, Luíz Fernando, Daniel, Rafael, Daiane, Michele, Claudivânea, Lílian, Mônica, pela disposição em ajudar, pelas conversas e momentos de descontração, pela companhia na luta do dia-a-dia e pelo incentivo constante. Aos amigos do Biotério de Experimentação Animal: Márcia, Vítor, Lucas, Miguel, pelos pela companhia, pelos momentos de descontração e pela disponibilidade em ajudar. Ao companheiro de trabalho e amigo Alex Bhering por iluminar nossos caminhos em meio aos procedimentos e técnicas em histologia, pela paciência, pela companhia e pelos momentos de descontração. Á amiga de Viçosa Marisa, pelo acolhimento e apoio nas horas de necessidade.

iv
As amigas Arlete e Reggiani pela parceria. Aos amigos do coração que fazem o dia-a-dia mais feliz mesmo que a distância nos separe de alguns: Aline, Ana Izaura, Arlete, Eliziária, Kener e Reggiani. À minha mãe e meu pai por tudo que fizeram por mim e que me permitiu chegar até aqui. À Deus, pela minha vida e pela força para superar as dificuldades e continuar a caminhada.

v
ÍNDICE RESUMO ...............................................................................................................vi
ABSTRACT........................................... ...............................................................viii
1. INTRODUÇÃO.............................................................................................1
1.1. Alterações histopatológicas do miocárdio na doença de Chagas.........5
1.2. Alterações funcionais do coração na doença de Chagas......................7
1.3. Tolerância ao exercício e função cardíaca na doença de Chagas......12
2. OBJETIVOS....................................... .........................................................17
3. REFERÊNCIAS...........................................................................................18
4. ARTIGO...................................................................................................... 22
5. PERSPECTIVAS ....................................................................................... 45

vi
RESUMO NOVAES, Rômulo Dias, M.Sc., Universidade Federal de Viçosa, julho de 2010. Efeitos da infecção experimental com Trypanosoma cruzi sobre a morfologia do miocárdio, propriedades mecânicas de cardiomiócitos isolados e tolerância ao exercício físico de ratos Wistar . Orientadora: Izabel Regina dos Santos Costa Maldonado. Coorientadores: Antônio José Natali e Clóvis Andrade Neves.
O Trypanosoma cruzi (T. cruzi), um protozoário parasita intracelular, é o
agente causador da doença de Chagas. Esse parasito é capaz de induzir
modificações patológicas na morfologia e hemodinâmica do coração, alterações
que têm sido implicadas na redução da tolerância ao exercício em indivíduos
portadores da doença de Chagas. Modificações na mecânica de cardiomiócitos
nessa doença têm sido raramente relatadas e o papel dessas alterações na
tolerância ao exercício permanece desconhecido. Assim, o presente estudo
investigou os efeitos da infecção experimental com Trypanosoma cruzi sobre a
morfologia do miocárdio, propriedades mecânicas de cardiomiócitos isolados e
tolerância ao exercício físico de ratos Wistar. Vinte e oito ratos Wistar machos
foram divididos em um grupo não infectado (n=14) e um grupo infectado (n=14).
Após nove semanas da inoculação com T. cruzi, os animais foram submetidos a
um protocolo incremental de corrida para avaliação do desempenho físico. Em
seguida, os corações foram removidos para a análise histopatológica e
morfométrica e cardiomiócitos foram isolados por dispersão enzimática para
análise das propriedades mecânicas. Os resultados encontrados demonstraram
que a infecção com T. cruzi prejudicou a tolerância ao exercício resultando em
redução significativa da distância percorrida, tempo total até a fadiga e carga de
trabalho. Nos animais infectados observou-se a presença de infiltrado
inflamatório, hipertrofia dos ventrículos e de cardiomiócitos do ventrículo
esquerdo, além do aumento da quantidade de colágeno, vasos sanguíneos e

vii
prejuízos na mecânica celular. Miócitos atriais e ventriculares dos animais
infectados mostraram redução significativa da amplitude de contração, máxima
velocidade de contração e relaxamento, e aumento no tempo para metade do
relaxamento. Ainda, miócitos ventriculares desses animais infectados também
apresentaram aumento do tempo para o pico de contração. De acordo com esses
resultados, foi possível concluir que a disfunção da contratilidade dos
cardiomiócitos induzida pela infecção com T. cruzi ocorre em um ambiente
morfologicamente alterado e que essas alterações podem estar associadas com a
redução da tolerância ao exercício físico nos ratos infectados.

viii
ABSTRACT NOVAES, Rômulo Dias, M.Sc., Universidade Federal de Viçosa, july 2010. Effects of experimental Trypanosoma cruzi infection on myocardium morphology, mechanical properties of isolated cardi omyocytes and exercise tolerance in Wistar rats. Advisor: Izabel Regina dos Santos Costa Maldonado. Co-advisors: Antônio José Natali and Clóvis Andrade Neves.
Trypanosoma cruzi (T. cruzi), an intracellular protozoan parasite, is the
causative agent of Chagas’ disease. This parasite is able to alter the morphology
and hemodynamics of the heart, changes that have been implicated in the
reduction of exercise tolerance in individuals with Chagas’ disease. Changes in
cardiomyocyte mechanics in this disease have been rarely reported and the role of
these changes in exercise tolerance remains unknown. Thus, the present study
investigated the effects of experimental Trypanosoma cruzi infection on
myocardium morphology, mechanical properties of isolated cardiomyocytes and
exercise tolerance in Wistar rats. Twenty-eight male Wistar rats were divided into
an uninfected (n=14) and an infected group (n=14). After nine weeks of inoculation
with trypomastigote forms of T. cruzi, animals were subjected to an incremental
running protocol to evaluate the physical performance. Then, hearts were removed
for histopathological and morphometric analysis and cardiomyocytes were isolated
by enzymatic dispersion for analysis of mechanical properties. The results showed
that T. cruzi infection impaired exercise tolerance resulting in a significant
reduction in distance traveled, total time to fatigue and workload. In the heart of
infected animls it was observed the presence of inflammatory infiltrate,
hypertrophy of the ventricles and the left ventricular cardiomyocytes, increase of
collagen and blood vessels amount and impairments in cellular mechanics. Atrial
and ventricular myocytes from infected animals showed a significant reduction of
contraction amplitude, maximal rate of contraction and relaxation and increase in

ix
time to half relaxation. These ventricular myocytes also showed increased time to
peak of contraction. According to these results it was possible conclude that the
dysfunction of cardiomyocytes contractility induced by T. cruzi infection occurs in
an environment morphologically altered and these changes may be associated
with reduction of exercise tolerance in infected rats.

1
1. INTRODUÇÃO
A doença de Chagas ou Tripanossomíase Americana, desde a sua
descoberta por Carlos Chagas em 1909, tem se apresentado como uma doença
de difícil abordagem clínica devido a sua característica sistêmica e a variabilidade
de manifestações clínicas que se desenvolvem ao longo da evolução da doença
(WHO, 2002, BIOLO et al., 2010). Embora tenham sido desenvolvidos
medicamentos contendo os princípios ativos benzonidazol e nifurtimox para o
tratamento dessa doença, até o momento não existe cura após a disseminação e
instalação do parasito nos tecidos (RASSI-JR et al., 2010).
A doença de Chagas apresenta-se distribuída em diversos países do mundo
devido a presença de indivíduos infectados provenientes de países endêmicos e
mecanismos que não envolvem a forma vetorial de transmissão da doença como
transfusão de sangue e doação de órgãos infectados (Fig.1). A maior prevalência
da doença é encontrada no continente Americano, propagando-se desde o sul
dos Estados Unidos até o sul da Argentina (WHO, 2002). Além de ocorrer em
quase toda a América Latina, essa doença é apontada como uma das principais
causas de morbidade e mortalidade entre as populações carentes (WHO, 2002) e
ainda hoje constitui um problema médico-social grave no Brasil e em diversos
países Latino-americanos (SCHMUNIS, 1997; DUTHIE et al., 2005; WHO, 2002).
Estima-se uma prevalência de aproximadamente 15 milhões de indivíduos
infectados nas américas, causando a morte de 50.000 pessoas/ano e
contaminação de 400.000 pessoas/ano (WHO, 2007).

2
Figura 1: Distribuição geográfica da doença de Chag as no mundo. Os dados
de prevalência para países da América do norte e Europa, Japão e Austrália
referem-se a forma não vetorial de transmissão da doença de Chagas. Além
disso, esses dados incluem indivíduos provenientes de países endêmicos (Fonte:
RASSI-JR et al., 2010).
A doença de Chagas é causada pelo protozoário parasita Trypanosoma
cruzi (T. cruzi), sendo o homem o principal reservatório acidental da doença e a
transmissão vetorial que ocorre por meio de insetos hematófagos (triatomíneos) a
mais proeminente (80 a 90%) (CHAGAS, 1909, 1916; SCHMUNIS, 1997;
ANDRADE, 2000). Mediante a entrada do T. cruzi no organismo do hospedeiro
vertebrado, o parasito é capaz de migrar e infectar diversos órgãos como fígado,
rins, baço, medula óssea, pâncreas, coração e intestino (ANDRADE, 2000; LANA
e TAFURI, 2000) (Fig. 2). Nesses órgãos, as alterações causadas pelo T. cruzi se
agravam progressivamente e reduzem a capacidade do organismo doente de
manter a homeostase uma vez que altera a regulação de sua função motora,
3
secretora e metabólica (ALBUQUERQUE, et al., 1991; OLIVEIRA et al., 1993;
OSAER et al., 2000, WHO, 2002).
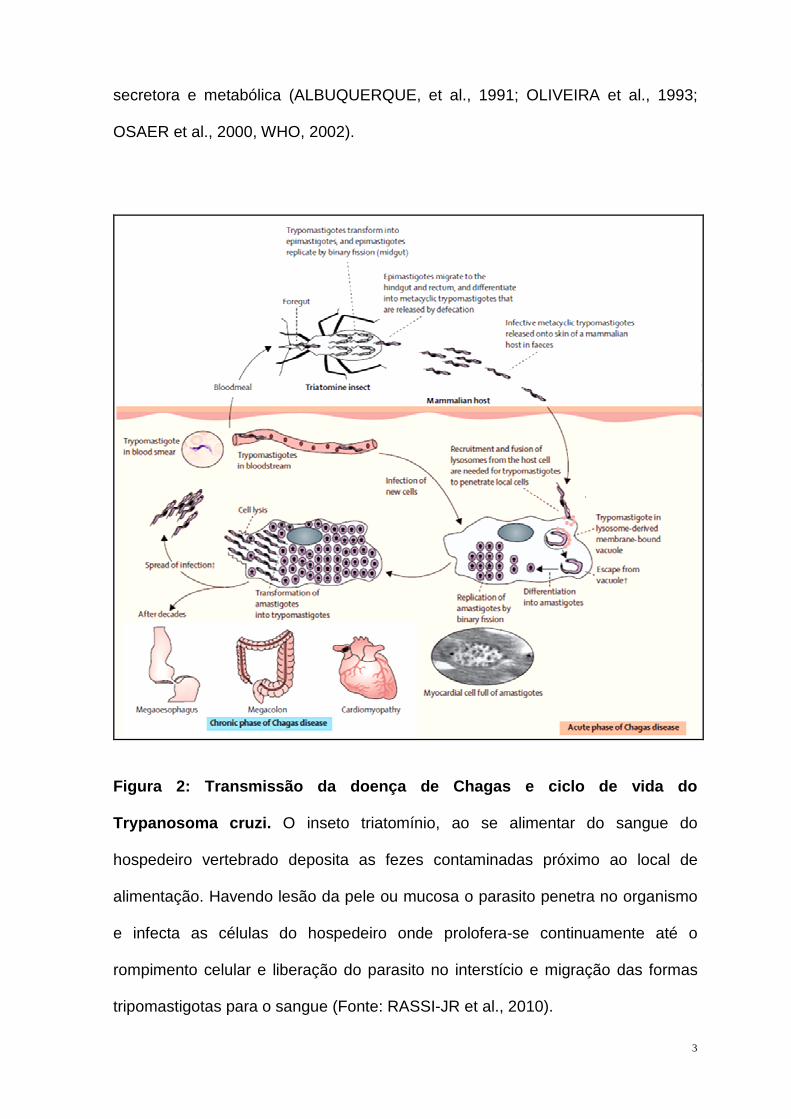
Figura 2: Transmissão da doença de Chagas e ciclo d e vida do
Trypanosoma cruzi . O inseto triatomínio, ao se alimentar do sangue do
hospedeiro vertebrado deposita as fezes contaminadas próximo ao local de
alimentação. Havendo lesão da pele ou mucosa o parasito penetra no organismo
e infecta as células do hospedeiro onde prolofera-se continuamente até o
rompimento celular e liberação do parasito no interstício e migração das formas
tripomastigotas para o sangue (Fonte: RASSI-JR et al., 2010).

4
A Doença de Chagas se desenvolve em dois diferentes estágios sucessivos:
o agudo e o crônico (ANDRADE, 2000; LANA e TAFURI, 2000). A fase aguda
inicia-se logo após a entrada do parasito no corpo do hospedeiro. Nessa fase,
parasitos de forma alongada (tripomastigotas) circulam nos vasos sangüíneos e
proliferam nas células do hospedeiro vertebrado, principalmente nas células do
sistema mononuclear fagocitário, para diferenciarem-se em formas arredondadas
ou ovóides denominadas amastigotas (Fig. 2) (ANDRADE, 2000; MARIN-NETO et
al., 2007; RASSI-JR et al., 2010; BIOLO et al., 2010). Essas formas amastigotas
podem se alojar em vários órgãos e provocar destruição celular associada a
processos inflamatórios (ANDRADE, 2000; HIGUCHI et al., 2003; MARIN-NETO
et al., 2007; BIOLO et al., 2010). Estudos demonstraram que os níveis de lesão
tecidual são intensos e evidentes nessa fase, com declínio progressivo e
recuperação parcial ou total dos tecidos parasitados entre a 4ª e 8ª semana após
a infecção (VIANNA, 1911; CHAGAS, 1916; WHO, 2002).
A fase crônica se inicia quando a parasitemia declina para níveis
indetectáveis e não são mais observados sinais de miocardite aguda. Essas
características são comumente encontradas entre a 4ª e 8ª semana após a
infecção, período coincidente com o término da fase aguda (CORBETT et al.,
2002; WHO, 2002; DUTHIE et al., 2005). Na fase crônica, podem ser encontradas
formas amastigotas alojadas nos tecidos e a presença de parasitos circulantes
geralmente não é evidenciada (ANDRADE, 2000). Entretanto, mesmo na
ausência de evidência histopatológica de infecção, a presença de proteínas e
ácidos nucléicos do parasito nos tecidos do hospedeiro pode ser detectada por
meio de testes imunológicos e bioquímicos (LANA e TAFURI, 2000; WHO, 2002).
Entretanto, em alguns casos pode ser observada intensa destruição celular,
infiltrados inflamatórios pericelulares e perivasculares e fibrose tecidual focal e/ou

5
difusa, processo patológico relacionado com a distribuição das formas
amastigotas nos tecidos (CORBETT et al., 2002; DUTHIE et al., 2005). Em
associação aos danos histopatológicos, é comum a ocorrência de alterações da
função gastrintestinal e cardíaca, caracterizando a forma digestiva e cardíaca da
doença de Chagas (ANDRADE, 2000; CAMARGOS et al., 2000).
Pesquisas realizadas sobre as manifestações clínicas da doença mostram
que a forma cardíaca é predominante e também a mais amplamente estudada
(MORRIS et al., 1990; ANDRADE et al., 2000; MARIN-NETO et al., 2007). O
comprometimento cardíaco é a principal causa de morbidade e mortalidade na
doença de Chagas (BIOLO et al., 2010; RASSI-JR et al., 2009, 2010). Existem
evidências de que a insuficiência cardíaca observada em muitos casos graves da
doença se desenvolve progressivamente ao longo da infecção cardíaca mediante
danos celulares, microvasculares e nervosos induzidos pelo parasito (HIGUCHI et
al., 2003; MARIN-NETO et al., 2007; RASSI-JR et al., 2009).
1.1. Alterações morfológicas do miocárdio na doença de C hagas
Ao longo da evolução da doença de Chagas podem ser observadas diversas
alterações anatomopatológicas cardíacas induzidas pelo parasito.
Macroscopicamente, na fase aguda da doença, pode ser observada a ocorrência
de graus variáveis de cardiomegalia associada a regiões de necrose tecidual e
áreas de hemorragia (VIANNA, 1911; HIGUCHI et al., 2003; RASSI-JR et al.,
2010). Na fase crônica, hipertrofia celular compensatória e extensa fibrose reativa
do miocárdio conduzem ao aumento da massa do coração e espessamento das
paredes das câmaras cardíacas, principalmente do ventrículo esquerdo. Além
disso, a ocorrência de trombose e fibrose apical no ventrículo esquerdo são

6
extremamente comuns e considerados eventos patognomônicos da
cardiomiopatia chagásica crônica (HIGUCHI et al., 2003; MARIN-NETO et al.,
2007). Focos de fibrose no trajeto do sistema de condução cardíaco também
podem ser evidenciados na fase crônica da doença e são responsáveis por parte
das alterações eletrofisiológicas desse órgão (MORRIS et al., 1990; HIGUCHI et
al., 2003).
A análise histológica do coração infectado por T. cruzi demonstra a
ocorrência de diferentes eventos nas duas fases da doença de chagas. Há relato
de que as lesões histológicas do miocárdio tornam-se evidentes em 50% dos
casos, dentro de quatro a dez dias após a inoculação do parasito no organismo
hospedeiro (LANA e TAFURI, 2000). Na fase aguda, o parasito induz a
desintegração da matriz extracelular (ME) nas áreas infectadas por meio da
expressão de metaloproteinases que degradam os componentes da ME,
principalmente as fibras de colágeno. Além disso, mediante o parasitismo dos
cardiomiócitos, a proliferação dos parasitos no interior dessas células conduz a
cardiomiocitólise, caracterizada por rompimento celular de origem mecânica
induzido pelo aumento progressivo da quantidade de parasitos intracelulares
(MORRIS et al., 1990; BONNEY e ENGMAN, 2008; RASSI-JR et al., 2010) (Fig.
2). Ambos os processos desencadeiam uma resposta inflamatória com a
migração de células de defesa para as regiões infectadas. Nessas regiões, a
liberação de substâncias citotóxicas como óxido nítrico, perforinas, granzima e
proteases pelas células de defesa com objetivo de combater os parasitos também
conduz a lesão de cardiomiócitos e eventualmente morte celular (HIGUCHI et al.,
2003; DUTHIE et al., 2005). Por meio desse mecanismo de combate ao parasito
na fase aguda da doença de Chagas, pode ser encontrado intenso infiltrado
inflamatório perivascular e pericelular e danos microvasculares (ANDRADE et al.,

7
2000; MARIN-NETO et al., 2007). Lesões vasculares decorrentes de dano
oxidativo às células endoteliais são observadas em associação com a ocorrência
de regiões de necrose tecidual, formação de trombos e oclusão vascular
(HIGUCHI et al., 2003; BONNEY e ENGMAN, 2008).
Alterações na inervação autonômica do coração também constituem uma
importante característica da fase aguda da doença de Chagas. Diversos estudos
têm demonstrado redução do número de neurônios no gânglio cardíaco
parassimpático e menor densidade de fibras nervosas parassimpáticas e
simpáticas em humanos e animais chagásicos (MACHADO et al., 1998;
CAMARGOS et al., 2000; MARIN-NETO et al., 2007; BONNEY e ENGMAN,
2008). Essas alterações podem persistir durante toda a fase aguda com
recuperação total ou parcial das lesões autonômicas na fase crônica da doença
(CAMARGOS et al., 2000, ANDRADE, 2000).
Durante a transição da fase aguda para a fase crônica da doença de
Chagas, ocorre o remodelamento progressivo da histoarquitetura do miocárdio.
Na fase crônica, pode ser observada a presença de infiltrado inflamatório e
persistência de antígenos do parasita em células fagocíticas do miocárdio
(HIGUCHI et al., 2003; BONNEY e ENGMAN, 2008; BIOLO et al., 2010) . Além
disso, formações císticas dos parasitos podem ser evidenciadas no interior dos
cardiomiócitos. A principal característica do remodelamento do miocárdio na fase
crônica é a reconstituição da matriz extracelular (ROSSI, 1998; HIGUCHI et al.,
2003). Nessa fase é comum a ocorrência de fibrose focal e/ou difusa com
deposição de fibras e feixes de colágeno de forma desordenada. O aumento da
quantidade e o padrão alterado de deposição de colágeno freqüentemente
conduz a interrupção parcial ou total da comunicação entre as células do sistema
de condução cardíaco, modificação da orientação dos cardiomiócitos, alteração

8
no trajeto dos vasos sanguíneos do miocárdio e colabamendo da parede de vasos
sanguíneos intramiocárdicos (HIGUCHI et al., 2003; MARIN-NETO et al., 2007).
1.2. Alterações funcionais do coração na doença de Chaga s
Além do remodelamento morfológico do coração na cardiopatia Chagásica,
alterações patológicas da função cardíaca também são comumente evidenciadas.
Alterações elétricas como disfunção do ritmo sinusal, bloqueios na condução do
potencial de ação e arritmias atriais e ventriculares têm sido amplamente
descritas (MORRIS et al., 1990; MARIN-NETO et al., 2007; BIOLO et al., 2010;
RASSI-JR et al., 2010). Em adição, distúrbios na mecânica e hemodinâmica
cardíaca como redução do volume diastólico final, força de contração muscular,
freqüência cardíaca, volume de ejeção ventricular e débito cardíaco também
estão envolvidos na disfunção cardíaca e contribuem para elevar o risco de óbito
na cardiopatia Chagásica (MORRIS et al., 1990; RASSI-JR et al., 2009). A figura
3 apresenta os principais eventos patológicos evidenciados na cardiopatia
Chagásica que conduzem a falha cardíaca e eventualmente ao óbito.

9
Figura 3: Representação esquemática das alterações patológicas
observadas na cardiopatia chagásica associadas à fa lha cardíaca e óbito. A
figura demonstra a relação entre a resposta inflamatória e alterações da
morfologia do miocárdio desencadeadas pela infecção com o T. cruzi e a
influência dessas alterações na patogênese das disfunções cardíacas que
potencialmente contribuem para o óbito (Fonte: RASSI-JR et al., 2009).
Embora parte dessas alterações seja determinada pelos danos celulares e à
matriz extracelular induzidos pelo parasito, mecanismos imunológicos têm sido
implicados na patogênese da disfunção cardíaca (BONNEY e ENGMAN et al.,
2008; HIGUCHI et al., 2003; MARIN-NETO et al., 2007; RASSI-JR et al., 2009).
Embora pouco se saiba a respeito dos mecanismos por meio dos quais o sistema
imunológico atua para modular a mecânica celular e cardíaca, alguns aspectos
patológicos associados a cardiopatias de origem não chagásica e que
apresentam similaridades com a cardiopatia chagásica podem explicar
parcialmente a disfunção contrátil do coração. Estudos prévios demonstraram que
citocinas comumente produzidas pelas células de defesa na fase aguda e/ou

10
crônica da doença de Chagas, como interferon (IFN)-γ, interleucina IL-1β, fator de
necrose tumoral (TNF)-α, IL-6, IL-10, IL-12, podem influenciar significativamente a
mecânica cardíaca e alterar a eficiência contrátil do miocárdio (DUNCAN et al.,
2007; ROMAN-CAMPOS et al., 2009).
Tem sido descrita uma correlação negativa entre os níveis de citocinas e o
desempenho cardíaco em cardiopatias de origem chagásica e não chagásica
(FERNANDEZ-VELASCO et al., 2007; ROMAN-CAMPOS et al., 2009). Existem
evidências de que a modulação da contração cardíaca por meio dessas citocinas
ocorre pela regulação diferencial de canais iônicos de membrana e proteínas
envolvidas na homeostase do cálcio e controle da maquinaria contrátil celular.
Acredita-se que essa regulação esteja relacionada à influência direta dessas
citocinas sobre a produção de óxido nítrico pelos cardiomiócitos, o qual é um
importante modulador de diferentes proteínas envolvidas na regulação da cinética
do cálcio intracelular (KROWN et al., 1995; GOLDHABER et al., 1996;
FERNANDEZ-VELASCO et al., 2007). O óxido nítrico apresenta reconhecido
papel na redução da contratilidade celular devido a inibição da ativação da
proteína Fosfoquinase A e da fosforilação dos canais de Ryanodina, canais de
cálcio tipo L, e da proteína Fosfolambam que modula a atividade da Ca2+-ATPase
do retículo sarcoplasmático. A inibição da fosforilação reduz a atividade desses
canais que controlam a mecânica contrátil Ca2+-dependente (AFANASYEVA et al.,
2004; FERNANDEZ-VELASCO et al., 2007). A figura 4 ilustra alguns mecanismos
por meio dos quais o sistema imunológico modula a contratilidade celular na
disfunção cardíaca.
11
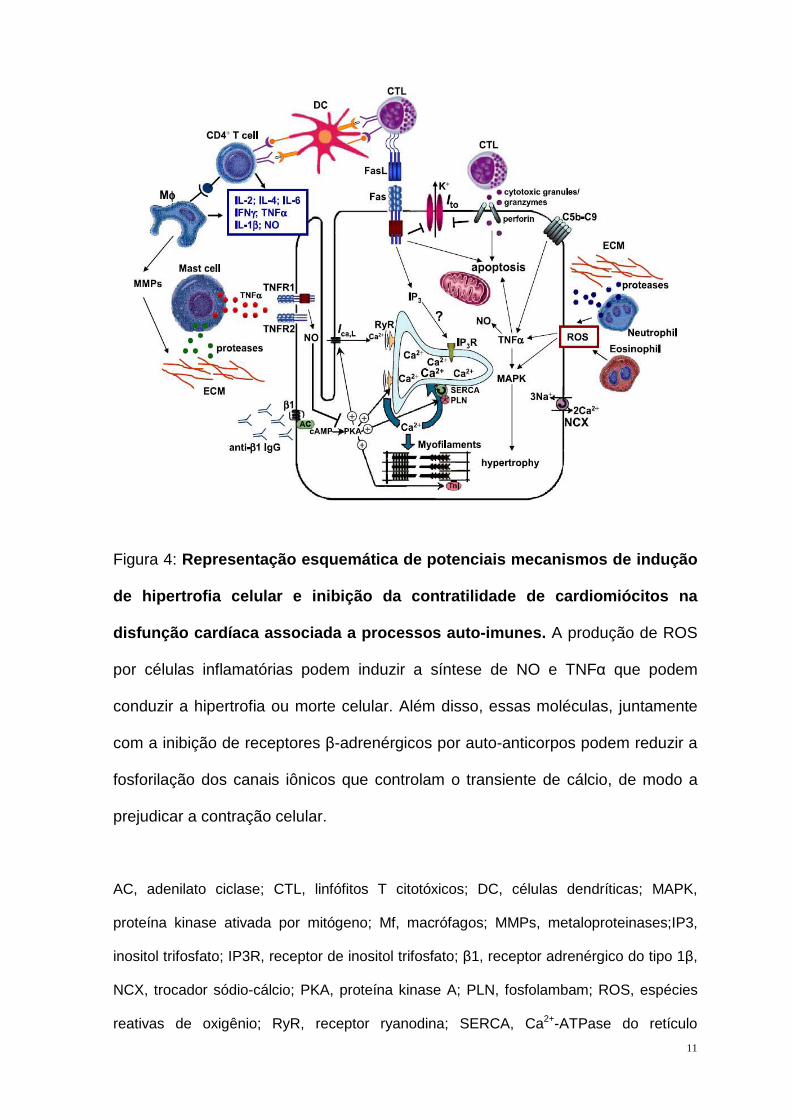
Figura 4: Representação esquemática de potenciais mecanismos de indução
de hipertrofia celular e inibição da contratilidade de cardiomiócitos na
disfunção cardíaca associada a processos auto-imune s. A produção de ROS
por células inflamatórias podem induzir a síntese de NO e TNFα que podem
conduzir a hipertrofia ou morte celular. Além disso, essas moléculas, juntamente
com a inibição de receptores β-adrenérgicos por auto-anticorpos podem reduzir a
fosforilação dos canais iônicos que controlam o transiente de cálcio, de modo a
prejudicar a contração celular.
AC, adenilato ciclase; CTL, linfófitos T citotóxicos; DC, células dendríticas; MAPK,
proteína kinase ativada por mitógeno; Mf, macrófagos; MMPs, metaloproteinases;IP3,
inositol trifosfato; IP3R, receptor de inositol trifosfato; β1, receptor adrenérgico do tipo 1β,
NCX, trocador sódio-cálcio; PKA, proteína kinase A; PLN, fosfolambam; ROS, espécies
reativas de oxigênio; RyR, receptor ryanodina; SERCA, Ca2+-ATPase do retículo

12
sarcoplasmático; TNFR1 e 2, receptor de fator de necrose tumoral 1 e 2; TnI, troponina I
(Fonte: AFANASYEVA et al., 2004).
Adicionalmente, auto-anticorpos anti-β-adrenorreceptor produzidos durante a
infecção contribuem para a disfunção cardíaca contrátil, principalmente na fase
crônica da doença de Chagas (BORDA et al., 1996; LABOVSKY et al., 2007). A
dessensibilização e/ou inibição alostérica desse receptor por auto-anticorpos anti-
β-adrenorreceptor reduz a eficiência contrátil celular e miocárdica uma vez que as
vias de regulação da inotropia e cronotropia celular são influenciadas
primariamente por esse receptor mediante fosforilação dos canais reguladores do
fluxo de cálcio celular (HUNTER e CHIEN, 1999; AFANASYEVA et al., 2004). A
figura 9 ilustra o papel dos receptores β-adrenérgicos no aumento da atividade do
canal Ca2+-ATPase do retículo sarcoplasmático que regula o relaxamento celular.
A ativação do receptor implica aumento da re-captação ativa de cálcio para o
retículo sarcoplasmático. A fosforilação da proteína fosfolambam por meio da
proteína fosfoquinase A dependente de AMP cíclico reduz o seu efeito tônico
inibitório sobre a bomba de cálcio do retículo sarcoplasmático favorecendo o
relaxamento celular. Anticorpos anti-β-adrenorreceptor podem influenciar
diretamente a mecânica celular por meio da inibição desse mecanismo, reduzindo
a eficiência contrátil celular e conseqüentemente do miocárdio.

13
Figura 5: Modulação da contração de cardiomiócitos mediada pela
estimulação de receptores β-adrenérgicos. A ativação do receptor leva a
aumento da re-captação ativa de cálcio para o retículo sarcoplasmático. A
fosforilação da proteína fosfolambam por meio da proteína quinase A dependente
de AMP cíclico reduz o seu efeito tônico inibitório sobre a bomba de cálcio do
retículo sarcoplasmático favorecendo o relaxamento celular. A presença de auto-
anticorpos anti-β1-adrenorreceptor inibe a ativação desse mecanismo (Fonte:
HUNTER e CHIEN, 1999).
1.3. Tolerância ao exercício e função cardíaca na doença de Chagas
A capacidade de realizar exercícios físicos é determinada por diferentes
sistemas corporais, principalmente pelo sistema nervoso central e periférico,
sistema musculoesquelético e cardiovascular (MCARDLE, et al., 1998; WILMORE
e COSTIL, 2001). Esses sistemas atuam sinergicamente no controle da
quantidade e qualidade de execução de exercícios físicos, e qualquer disfunção
em um deles é capaz de influenciar negativamente a tolerância ao exercício físico
(FITTS, 1994, MCCONELL et al., 1994). Na doença de Chagas, o T. cruzi é capaz
Anti-β-adrenergic
receptor

14
de induzir alterações morfológicas e funcionais nos três sistemas supracitados,
com repercussões negativas sobre a organização histológica e função dos
músculos esqueléticos, músculo cardíaco, nervos somáticos e autonômicos,
nestes últimos, destruindo as fibras nervosas autonômicas simpáticas e
parassimpáticas que modulam a inotropia e cronotropia celular e cardíaca
(MACHADO et al., 1998; CAMARGOS et al., 2000; HIGUCHI et al., 2003; MARIN-
NETO et al., 2007; BIOLO et al., 2010, RASSI-JR et al., 2009, 2010). Foi
previamente demonstrado que mesmo na ausência de cardiopatia chagásica
avançada e anormalidades eletrocardiográficas de repouso, alterações na função
cardíaca podem emergir quando maior ativação cardiovascular é requerida, como
durante a execução de exercícios físicos (MOLINA et al., 1981; MADY et al.,
2000).
Particularmente durante a execução de exercícios físicos, o aumento da
demanda energética pelos músculos recrutados exige um aumento do débito
cardíaco. Na ausência de doença, esse aumento ocorre por meio da elevação da
freqüência cardíaca e da força de contração do miocárdio, conseqüentemente, do
volume de ejeção. Esses ajustes contribuem para a manutenção do estado
homeostático quando o organismo encontra-se submetido a maior atividade
metabólica e permite a progressão do exercício (MCARDLE, et al., 1998;
WILMORE e COSTIL, 2001).
Na cardiopatia chagásica, as anormalidades elétricas e contráteis do coração
são freqüentemente correlacionadas a uma menor reserva funcional cardíaca que
se manifesta primordialmente como redução da capacidade do coração de se
adaptar a perturbações da homeostase corporal, como ocorre durante a
realização de exercícios físicos (GALLO et al., 1975; MOLINA et al., 1981; MADY
et al., 2000). Nesse contexto, estudos foram conduzidos para demonstrar que a

15
disfunção cardíaca e a inabilidade do coração em adaptar-se a altas cargas de
trabalho conduzem a redução da tolerância ao exercício físico (GALLO et al.,
1975; MOLINA et al., 1981, SOUSA et al., 2008, 2009; LIMA et al., 2010; NUNES
et al., 2010). Em geral, a redução da vascularização do miocárdio e o
desenvolvimento de arritmias atriais e ventriculares, a redução do volume
diastólico, pressão de enchimento e fração de ejeção do ventrículo esquerdo
(LIMA et al., 2010; NUNES et al., 2010) e a limitada capacidade de elevação da
freqüência cardíaca decorrente de denervação e/ ou dessensibilização
autonômica têm sido descritos como os principais parâmetros cardíacos
comprometidos na cardiomiopatia chagásica que estão associados com a
redução da tolerância ao exercício físico (GALLO et al., 1975; MOLINA et al.,
1981; SOUSA et al., 2008, 2009).
Embora existam estudos que demonstraram uma relação entre alterações da
contratilidade do coração com a redução da tolerância ao exercício físico em
indivíduos chagásicos, pouco se sabe a respeito dos fatores determinantes
dessas alterações na doença de Chagas. Até o presente momento, são escassas
as informações a respeito da influência da infecção com T. cruzi sobre as
propriedades mecânicas de cardiomiócitos isolados. Acredita-se que além das
alterações elétricas e morfológicas do miocárdio evidenciadas na doença de
Chagas, a infecção pelo T. cruzi também modifica as propriedades mecânicas
intrínsecas dos miócitos cardíacos, podendo ser um fator adicional na disfunção
da mecânica do coração. Além disso, ainda é pouco conhecido quais as
propriedades mecânicas dos cardiomiócitos são modificadas durante a infecção
por esse parasito, e se essas modificações podem interferir na capacidade de
realizar exercício físico. Elucidar esses mecanismos pode auxiliar a compreender
de forma mais ampla o impacto da doença de Chagas sobre o músculo estriado

16
cardíaco, além de definir o quanto às alterações nesses componentes são
capazes de contribuir para restringir a tolerância ao exercício físico.

17
OBJETIVOS
1) Investigar a influência da infecção experimental com T. cruzi sobre a
morfologia do miocárdio de ratos Wistar;
2) Investigar a influência da infecção experimental com T. cruzi sobre as
propriedades mecânicas de cardiomiócitos isolados de ratos Wistar;
3) Investigar influência da infecção experimental com T. cruzi sobre a
tolerância ao exercício físico de ratos Wistar.

18
REFERÊNCIAS
AFANASYEVA, M.; GEORGAKOPOULOS, D.; ROSE, N. R. Autoimmune
myocarditis: cellular mediators of cardiac dysfunction. Autoimmunity Reviews . v. 3, p. 476-486, 2004.
ALBUQUERQUE, S.; CARRARO, A. A.; RIBEIRO, R. D.; LOPES, R. A.;
PETENUSCI, S. O.; PETENUSCI, N. C. Doença de Chagas experimental em ratos: histopatologia do pâncreas e estudo de alguns constituintes metabólicos e fisiológicos. Revista de Ciências Farmacêuticas . v.13, p.201, 1991.
ANDRADE, A. A. Patologia da doença de Chagas. In: Trypanosoma cruzi e
doença de Chagas. Guanabara Koogan. Rio de Janeiro. 2ª ed., p. 201-230, 2000.
BIOLO, A.; RIBEIRO, A. L.; CLAUSELL, N. Chagas cardiomyopathy-where do we
stand after a hundred years? Progress in Cardiovascular Diseases . v. 52, p. 300-316, 2010.
BONNEY, K. M.; ENGMAN, D. M. Chagas Heart Disease Pathogenesis: One
Mechanism or Many? Current Molecular Medicine . v. 8, n. 6, p. 510-518, 2008.
BORDA, E. S.; STERIN-BORDA, L. Antiadrenergicmand muscarinic receptor antibodies in Chagas’ cardiomyopathy. International Journal Cardiology . v. 54, p. 149-156, 1996.
CAMARGOS, E. R. S.; FRANCO, D. J.; GARCIA, C. M. M. G.; DUTRA, A. P.;
TEIXEIRA-JÚNIOR, A. L.; CHIARI, E.; MACHADO, C. R. S. Infection with different Trypanosoma cruzi populations in rats: myocarditis, cardiac sympathetic denervation and involvement of disgestive organs. American Journal of Tropical Medicine and Hygiene . v.62, n.5, p. 604-12, 2000.
CHAGAS, C. Nova tripanozomiase humana. Estudos sobre a morfolojia e o ciclo
evolutivo de Schizotrypanum cruzi n. gen., n. sp., ajente etiolojico de nova entidade morbida do homen. Memórias do Instituto Oswaldo Cruz . v.1, p. 159-218, 1909.
CHAGAS, C. Tripanossomíase americana. Forma aguda da moléstia. Memórias
do Instituto Oswaldo Cruz . v.1, p. 159-218, 1916. CORBETT, C. E.; SCREMIN, L. H.; LOMBARDI, R. A.; GAMA-RODRIGUES, J. J.;
OKUMURA, M. Pancreatic lesions in acute experimental Chagas' disease. Revista do Hospital das Clinicas da Faculdade de Me dicina de São Paulo . v. 57, suppl.2, p. 63-6, 2002.
DUNCAN, D. J.; HOPKINS, P. M.; HARRISON, S. M. Negative inotropic effects of
tumour necrosis factor-alpha and interleukin-1beta are ameliorated by

19
alfentanil in rat ventricular myocytes. British Journal of Pharmacology . v. 150, p. 720-726, 2007.
DUTHIE, M. S.; KAHN, M.; WHITE, M.; KAPUR, R. P.; KAHN, S. J. Critical
proinflammatory and anti-inflammatory functions of different subsets of CD1d-restricted natural killer T cells during Trypanosoma cruzi infection. Infection and Immunity . v.73, suppl. 3, p. 181-92, 2005.
FERNANDEZ-VELASCO, M.; RUIZ-HURTADO, G.; HURTADO, O.; MORO, M. A.; DELGADO, C. TNF-alpha downregulates transient outward potassium current in rat ventricular myocytes through iNOS overexpression and oxidant species generation. American Journal of Physiology: Heart and Circulato ry Physiology . v. 293, p. 238-245, 2007.
FITTS, R. H. Cellular mechanisms of muscle fatigue. Physiology Review . v.74, p. 49-94, 1994.
GALLO, L. JR.; NETO, J. A.; MANCO, J. C.; RASSI, A.; AMORIM, D. S..
Abnormal heart rate responses during exercise in patients with Chagas’ disease. Cardiology. v. 60, p.147-162, 1975.
GOLDHABER, J. I.; KIM, K. H.; NATTERSON, P. D.; LAWRENCE, T.; YANG, P.;
WEISS, J. N. Effects of TNF-alpha on [Ca2+]i and contractility in isolated adult
rabbit ventricular myocytes. Am J Physiol Heart Circ Physiol. v. 271, p. 1449-1455, 1996.
HIGUCHI, M. L.; BENVENUTI, L. A.; REIS, M. M.; METZGER M. Pathophysiology
of the heart in Chagas’ disease: current status and new developments. Cardiovascular Research . v. 60, p. 96-107, 2003.
HUNTER, J. J.; CHIEN, K. R. Signaling pathways for cardiac hypertrophy and
failure. New England Journal of Medicine . v. 34, p. 1277-1283, 1999.
KROWN, K. A.; YASUI, K.; BROOKER, M. J.; DUBIN, A. E.; NGUYEN, C.; HARRIS, G. L.; MC-DONOUGH, P. M.; GLEMBOTSKI, C. C.; PALADE, P. T.; SABBADINI, R. A. TNF alpha receptor expression in rat cardiac myocytes: TNF alpha inhibition of L-type Ca2+ current and Ca2+ transients. FEBS Lett 376:24-30, 1995.
LABOVSKY, V.; SMULSKI, C. R.; GÓMEZ, K.; LEVY, G.; LEVIN, M. J. Anti-beta1-
adrenergic receptor autoantibodies in patients with chronic Chagas heart disease. Clinical and Experimental Immunology . v. 148,p. 440-449, 2007.
LANA, M.; TAFURI, W. L. Trypanosoma cruzi e doença de Chagas. In:
Parasitologia humana. Atheneu. São Paulo. 10ª ed., p. 73-96, 2000.
LIMA, M. M. O.; PEREIRA, M. C.; ROCHA, M. O. C.; BELOTI, F. R.; ALENCAR, M. C. N.; RIBEIRO, A. L. P. Left ventricular diastolic function and exercise capacity in patients with Chagas cardiomyopathy. Echocardiography . (2010). (in press)

20
MCARDLE, W. D.; KATCH, F. I.; KATCH, V. L. Fisiologia do Exercício. Energia,
Nutrição e Desempenho Humano. Guanabara Koogan. Rio de Janeiro. 4ª ed., 1998.
MACHADO, C. R. S.; CALIARI M. V.; LANA M.; TAFURI W. L. Heart autonomic
innervation during the acute phase of experimental American Trypanosomiasis in the dog. American Journal of Tropical Medicine and Hygiene . v. 59, n. 3, p. 492-496, 1998.
MADY, C.; IANNI, B. M.; ARTEAGA, E.; SALEMI, V. M. C.; FRIMM, C. C. Maximal
functional capacity in patients with chagas’ cardiomyopathy without congestive heart failure. Journal of Cardiac Failure . v.3, p. 220-224, 2000.
MCCONELL, G.; FABRIS, S.; PROIETTO, J.; HARGREAVES, M. Effect of
carbohydrate ingestion on glucose kinetics during exercise. Journal of Applied Physiology . v. 77, p. 1537-1541, 1994.
MARIN-NETO, J. A.; CUNHA-NETO, E.; MACIEL, B. C.; SIMÕES, M. V.
Pathogenesis of chronic Chagas’ heart disease. Circulation . v. 115, p. 1109-1123, 2007.
MOLINA, A. R.; CARRASCO, H. G.; MILANÉS, J.; MOLINA, C. A.; PACHECO, J.;
FUENMAYOR, A. P. LA prueba de esfuerzo en la miocardiopatía chagásica crónica. Su valor en el diagnóstico precoz. El comportamiento de las arritmias ventriculares y los transtornos de conducción al ejercicio en las fases más avanzadas de la enfermedad. Arquivos Brasileiros de Cardiologia . v. 36, p. 95-100, 1981.
MORRIS, S. A.; TANOWITZ, H. B.; WITTNER, M.; BILEZIKIAN, J. P.
Pathophysiological insights into the cardiomyopathy of Chagas' disease. Circulation . v. 82, p. 1900-1909, 1990.
NUNES, M. C. P.; BELOTI, F. R.; LIMA, M. M. O.; BARBOSA, M. M.; FILHO, M.
M. P.; BARROS, M. V. L.; ROCHA, M. O. C. Functional capacity and right ventricular function in patients with Chagas heart disease. European Journal of Echocardiography . (2010). (in press)
OLIVEIRA, L. C. M.; JULIANO, Y.; NOVO, N. F.; NEVES, M. M. Blood glucose
and insulin response to intravenous glucose by patients with cronic Chagas disease and alcholism. Brazilian Journal of Medical and Biological Research . v.26, p.1187-90, 1993.
OSAER, S.; AKINBAMIJO, O. O.; GOOSSENS, B. Some biochemical changes
following Trypanosoma congolense infection in Djallonké ewe lambs and breeding ewes fed on two levels of nutrition. Acta Tropical . v. 75, p.229-241, 2000.
RASSI-JR, A.; RASSI, A.; MARIN-NETO, J. A. Chagas heart disease:
pathophysiologic mechanisms, prognostic factors and risk stratification. Memórias do Instituto Oswaldo Cruz . v. 104, Suppl 1, p. 152-158, 2009.

21
RASSI-JR, A.; RASSI, A.; MARIN-NETO, J. A. Chagas disease. Lancet . v.375, p.
1388-1402, 2010. ROMAN-CAMPOS, D.; DUARTE, H. L. L.; SALES-JR, P. A.; NATALI, A. J.;
ROPERT, C.; GAZZINELLI, R. T.; CRUZ, J. S. Changes in cellular contractility and cytokines profile during Trypanosoma cruzi infection in mice. Basic Research in Cardiology. v. 104, p. 238-46, 2009.
ROSSI, M. A. Fibrosis and inflammatory cells in human chronic chagasic
myocarditis: scanning electron microscopy and immunohistochemical observations. International Journal of Cardiology . v. 66, p. 183-194, 1998.
SCHMUNIS, G. A. Tripanossomíase Americana: Seu impacto nas Américas e
perspectivas de eliminação. In: Clínica e Terapêutica da Doença de Chagas. Um manual prático para a Clínica Geral. Fiocruz. Rio de Janeiro. p. 11-24, 1997.
SOUSA, L. A. P.; BOTONI, F. A.; BRITTO, R. R.; ROCHA, M. O. C.; TEIXEIRA-
JÚNIOR, A. L.; TEIXEIRA, M. M.; REIS, A. M.; OLIVEIRA, B. M. R.; RIBEIRO, A. L. Six-minute walk test in Chagas cardiomyopathy. International Journal of Cardiology . v. 125, p. 139-141, 2008.
SOUSA, L. A. P.; ROCHA, M. O. C.; BRITTO, R. R.; LOMBARDI, F.; RIBEIRO, A.
L. Chagas disease alters the relationship between heart rate variability and daily physical activity. International Journal of Cardiology . v. 135, p. 257-259, 2009.
VIANNA, G. Contribuição para o estudo da anatomia patológica da “moléstia de
Chagas”. Memórias do Instituto Oswaldo Cruz . v. 3, p. 276-294, 1911. WILMORE, J. H.; COSTIL, D. L. Fisiologia do esporte e do exercício. 2º ed. Rio
de Janeiro: editora Manole, 2001. World Health Organization (2002). Control of Chagas disease. Second report of the WHO Expert Committee. Technical Report Series Nº. 905, Geneva, 96 pp.

22
Artigo
Effects of experimental Trypanosoma cruzi infection on myocardium
morphology, mechanical properties of isolated cardi omyocytes and exercise
tolerance in Wistar rats

23
Abstract
Background: Changes in the myocardium morphology and cardiomyocyte
contractile function impair cardiac function in Chagas’ disease, but the role of
these changes in exercise tolerance remains unknown. Thus, we investigated the
effects of experimental Trypanosoma cruzi infection on myocardium morphology,
mechanical properties of isolated cardiomyocytes and exercise tolerance in Wistar
rats. Methods and Results: Adult Wistar rats were randomized into control
(CG=14) and infected (IG=14) groups. Animals from IG were inoculated with T.
cruzi Y strain (300,000 trypomastigotes/50g wt.). After nine weeks, the animals
were subjected to treadmill running protocol. Then, the animals were euthanized
and cardiac right atrium (RA) and left ventricle (LV) were removed to morfological
evaluation and single-cells were isolated for contractility analysis. In exercise test,
animals of IG exhibited significant reduction in distance traveled, total time to
fatigue and workload compared to CG. In addition, IG animals presented
hypertrophy, inflammatory infiltrate and increased proportion of collagen and blood
vessels in the ventricular myocardium. RA and LV myocytes from IG showed
significant reduction of contraction amplitude and increase in time to half
relaxation. Ventricular cardiomyocytes also showed reduced time to peak of
contraction. In addition, IG animals presented reduced contractile response of RA
and LV myocytes to β-adrenergic stimulation compared to CG. Conclusion:
Dysfunction of cardiomyocyte contractility occurred in an environment
morphologically altered and these changes could constitute an additional
mechanism of cardiac contractile function impairment and reduced exercise
tolerance in T. cruzi infection.
Key-words: Chagas’ cardiomyopathy, cellular contractility, myocytes, physical
performance.

24
Introduction
Chagas’ disease (ChD) is a neglected illness caused by the intracellular
protozoan parasite Trypanosoma cruzi (T. cruzi) that remains as an important
health problem in 18 developing countries in South and Central Americas (Biolo et
al., 2010; Rassi-Jr et al., 2010). Its main clinical manifestations are cardiac and/or
digestive disturbances, with an prevalence of about 12-14 million cases worldwide,
been considered a major cause of cardiac infection disease in endemic countries
(WHO, 2005). Chagas’ cardiomyopathy (ChC) is the main cause of death and
occurs in approximately 30% of infected subjects (Marin-Neto et al., 2007; Rassi-
Jr et al., 2010). The clinical course of Chagas’ disease show great variability and
the mechanisms responsible for the development of this potentially lethal
cardiomyopathy remain not well understood (Biolo et al., 2010; Rassi-Jr et al.,
2010).
Cardiac denervation, interstitial mononuclear infiltrate, myocyte and vascular
degenerative changes, fibrosis accumulation and heart hypertrophy characterize
the main pathologic features of the ChC (Marin-Neto et al., 2007, Biolo et al.,
2010; Rassi-Jr et al., 2010). These morphological changes coexist and are
associated with abnormalities of the electrical and contractile cardiac activities
characterized mainly by conduction defects, frequent and complex ventricular
arrhythmias and systolic ventricular dysfunction (Marin-Neto et al., 2007; Biolo et
al., 2010). In addition, the chronotropic incompetence caused by changes in
sympathetic and parasympathetic tonus induced by immune-mediated process
has been recognized as one of the mechanisms capable of interfering with the
capacity of the heart to increase heart rate in response to different stimuli,
including physical exercise (Colucci et al., 1989; Talvani et al., 2006; Sousa et al.,
2009).
Few studies have evaluated exercise performance and the factors affecting
functional capacity and exercise tolerance in patients with ChD. These studies are
focused mainly on the electrophysiological (Molina et al., 1981) and hemodynamic
(Mady et al., 2000; Lima et al., 2010) abnormalities of atrial and ventricular
function. The reduction of exercise tolerance in individuals with Chagas’ disease is
involved with changes in mechanical and electrical activity of atria and ventricle
(Gallo et al., 1975; Colucci et al., 1989; Mady et al., 2000; Lima et al., 2010).

25
However, several aspects of the cellular and molecular basis of these changes
remain to be clarified.
Recently, our group showed for the first time changes in cellular mechanics of
cardiac myocytes isolated from the atrium and ventricle of C57BL/6 mice infected
with T. cruzi (Roman-Campos et al., 2009). We demonstrated decreased myocyte
contraction amplitude and prolonged contraction and relaxation time course which
were observed in the very beginning of the parasitism and remained until the
chronic phase of the disease. Data from our laboratory also showed that in normal
rats exercise performance is significantly influenced by the electromechanical
characteristics of cardiomyocytes (Prímola-Gomes et al., 2009). In this study,
cardiomyocytes isolated from rats with high-capacity running had greater calcium
transients, amplitude of cell contraction, maximum velocity of contraction and
relaxation compared with rats of the same progeny with standard-capacity running.
Given that T. cruzi infection modifies the morphology of the myocardium and the
mechanical properties of isolated cardiomyocytes which result in abnormalities of
the cardiac contractile function, the present study was designed to investigate the
Effects of experimental T. cruzi infection on myocardium morphology, mechanical
properties of isolated cardiomyocytes and exercise tolerance in Wistar rats. We
hypothesized that exercise tolerance in rats experimentally infected with T. cruzi is
associated with morphological changes in the myocardium and cardiomyocyte
contractile function.
Methods
Animals and Infection
Four-month-old male Wistar rats with initial weight of 366.25 ± 31.17g were
provided with rodent chow and water ad libitum and maintained in animal facilities
with a controlled temperature at 22 ºC and 12 hour light/dark inverted cycles.
Animals were randomly divided into control (CG=14) and infected (IG=14) groups.
Animals from IG were inoculated intraperitoneally with T. cruzi Y strain (300,000
trypomastigotes/50 g body weight in 1 mL of infected mice blood) (Martinelli et al.
2006). Infection was confirmed four days post-inoculation by the presence of
trypomastigotes in peripheral blood collected from the rat’s tail as described by

26
Brener (1962). All experimental procedure were conducted in accordance with the
Brazilian College of Animal Experimentation and approved by the Animal
Research Ethics Commission of the Veterinary Department at the Federal
University of Viçosa, Brazil (protocol 30/2009).
Measurement of Exercise Tolerance
Nine weeks after inoculation all animals were evaluated for exercise tolerance
by using a treadmill incremental running protocol adapted from Koch and Britton
(2001). Briefly, the rats were familiarized with the motor-driven treadmill (Insight
Instruments®, Ribeirão Preto, Brazil) by running at a speed of 10 m/min, at 5%
inclination, 5 min/day, for 7 consecutive days. Two days after familiarization, the
exercise trial was performed on three consecutive days at a constant slope of 5%
with the starting speed at 10 m/min. Treadmill velocity was increased by 1 m/min
every 2 min and each rat ran until fatigue. Fatigue was defined as the point at
which the animals were no longer able to keep pace with the treadmill (Lacerda et
al., 2006). Traveled distance (m), time until fatigue, and workload were used as
indexes of exercise tolerance (Lacerda et al., 2006). Workload (W; kgm) was
calculated using the equation W= body mass (kg) × TTF (min) × treadmill speed
(m/min) × sine θ (treadmill inclination), where TTF is time until fatigue (Brooks et
al., 1984). Due to variability in performance data, the mean of the indexes of
running performance were calculated for the three trials for each rat and
considered for analysis.
Heart Biometry and Myocardial Stereology
Forty-eight hours after the exercise test, five animals from each group were
sacrificed and hearts removed and weighed. The atria and ventricles were
dissected and weighed separately. The indexes of cardiac, atrial and ventricular
hypertrophy were calculated by the ratios of heart, atrium and ventricle weight to
tibia length, respectively (Bezerra et al., 2008). The atria and ventricles were put
into histological fixative for 48 hours (freshly prepared 10% w/v formaldehyde in
0.1 M phosphate buffer pH 7.2). The fragments of right atrium (RA) and left
ventricle (LV) were obtained through the Orthrip method for stereological study
(Mandarim-de-Lacerda, 2003). These fragments were dehydrated in ethanol,

27
cleared in xylol and embedded in paraffin. Blocks were cut into 4 µm sections and
stained by Masson’s trichromic or hematoxylin-eosin (H&E) and mounted on
histology slides. The slides were visualized and the images captured using a light
microscope (Olympus BX-60®, Tóquio, Japan) connected to a digital camera
(Olympus QColor-3®, Tóquio, Japan). Sixty fields from each Masson’s trichromic
(objective x20) and H&E (objective x40) were randomly chosen and a total of
4.37x106 µm2 and 1.41x106 µm2 myocardium area, respectively, were analyzed.
Sections stained with Masson’s trichromic were used for myocardial stereological
analysis. For the stereological analysis, a test system of 72 points was used in a
standard test area of 73x103 µ2 (Mandarim-de-Lacerda, 2003). All the stereological
analyses were performed according to Bezerra et al., (2008). The stereological
parameter of volume density (Vv) was estimated by point counting for
cardiomyocytes [cmy], collagen [col], and intramyocardial blood vessels [ibvs]:
Vv structure =
Where PP is the number of points that hit the structure and PT is the total test
points. The amount of intramyocardial vascularization was defined as
Vv[ibvs]/Vv[cmy] ratio. The mean cross-sectional area of cardiomyocytes was
estimated according to the following relationship:
A cmy = ; QA cmyn =
where QA [cmyn] is the cardiomyocyte nuclei profiles in the analyzed area (AT).
Overestimation of the measurements was avoided by the exclusion of the nuclei
profiles incident on two edges of the AT.
Kariometry and Myocardial Histopathology
Sections stained with H&E were used to assess the inflammatory process and
for the karyometric study of cardiomyocyte nuclei. In karyometric analysis, 50
longitudinally sectioned cardiomyocytes for each animal and cardiac segment (RA
PP [ structure ] ______________
PT
______________ Vv [cmy]
2.QA[cmyn]
___________ N [cmy]
AT

28
and LV) were analyzed. Longest axis, shortest axis, mean axis, longest axis to
shortest axis ratio, area, volume and volume to area ratio were determined (Sala
et al., 1994; Yan et al., 1999). The inflammatory process was evaluated by the
correlation index between the number of cells observed in myocardium from CG
and IG animals (Caldas et al., 2008). All morphological analysis was performed
using the software Image Pro-Plus 4.5® (Media Cybernetcs, Silver Spring, USA).
Cardiomyocytes Isolation
Nine animals from each group were used in this set of experiment. At sacrifice
the heart was rapidly removed and extraneous tissue dissected away. Then, the
heart was flushed with a modified Hepes-Tyrode solution of the following
composition (mM): 130 NaCl, 5.4 KCl, 1.4MgCl2, 0.4 NaH2PO4, 0.75 CaCl2, 5
Hepes, 10 glucose, 20 taurine and 10 creatine (pH set at 7.4). The heart was then
blotted and weighed before being mounted onto a Langendorff perfusion
apparatus for isolation of myocytes using a collagenase-protease dispersion
technique as previously described (Natali et al., 2001). Briefly, the heart was
perfused for 10-15 min with a solution containing 1 mg/ml collagenase type II
(Worthington, USA). Then, the digested heart was then removed from the cannula,
and the RA and LV were separated and cut into small pieces. Ventricular and atrial
cardiomyocytes cells were isolated by mechanical tritation during 5 minutes at
37ºC and single cells were separated from the non-dispersed tissue by filtration.
The resulting cell suspension was centrifuged at 2000 rpm for 15 s and
resuspended in Hepes-Tyrode and stored at 4ºC until analysis. Only calcium-
tolerant, quiescent, rod-shaped cadiomyocytes showing clear cross striations were
studied. The isolated cardiac myocytes were used within 4 h after isolation.
Measurements of Cell Contractile Function
Cellular contractile function was evaluated as described by Natali et al., (2001).
Isolated cells were placed in a chamber with a glass coverslip base mounted on
the stage of an inverted-type phase contrast video microscope (Eclipse-TS100®,
Nikon, Japan). The chamber was perfused with Tyrode’s solution (in mM): 140
NaCl, 5.4 KCl, 1 MgCl2, 1.8 CaCl2, 10 HEPES, 10 glucose (pH set at 7.4) at room

29
temperature (~ 28 ºC). Myocytes were stimulated via platinum bath electrodes with
voltage pulses with 5 ms duration and an intensity of 20 V at the stimulation
frequency of 3 Hz. Cells were visualized on a PC monitor with a NTSC camera
(Myo-Cam CCD100V®, Ionoptix, Milton, MA, USA) in partial scanning mode. This
image was used to measure cell shortening (index of contractile function) in
response to electrical stimulation, using a video motion edge detection system
(Ionoptix, Milton, MA, USA). The cell image was sampled at 240 Hz and cell
shortening was calculated from the output of the edge detector using an IonWizard
A/Dconverter (Ionoptix, Milton, MA, USA). From 8 to 16 consecutive contractions
were averaged and cell shortening (expressed as a percentage of resting cell
length), time to peak of shortening and time to half relaxation were calculated
(Roman-Campos et al., 1999).
β-adrenergic Stimulation
The contractile response of isolated cardiomyocytes to β-adrenergic stimulation
was assessed at the stimulation rate of 1 Hz by using the nonselective agonist
isoproterenol (ISO, 1, 2 and 3 mmol / L). After the baseline cell shortening
recording ISO was infused in the experimental chamber through a pipette and the
cells were electrically stimulated after 5 minutes of infusion (PRAHASH et al.,
2000) when cell shortening was recorded. This procedure was repeated for each
ISO concentration in different myocytes. Cell contractile function was analyzed
and the variation (∆) from the baseline to the larger stimulus (ISO, 3 mmol / L) was
used as an index of β-adrenergic sensitivity.
Statistical analysis
Data are presented as mean ± S.E.M. Normal distribution of data was verified
using the Kolmogorov-Smirnov test. Exercise tolerance data were compared using
the student t-test and morphological and cell contractile function data were
compared using the Wilcoxon´s test. A probability of P<0.05 was considered
statistically significant.
30
Results
Exercise Tolerance
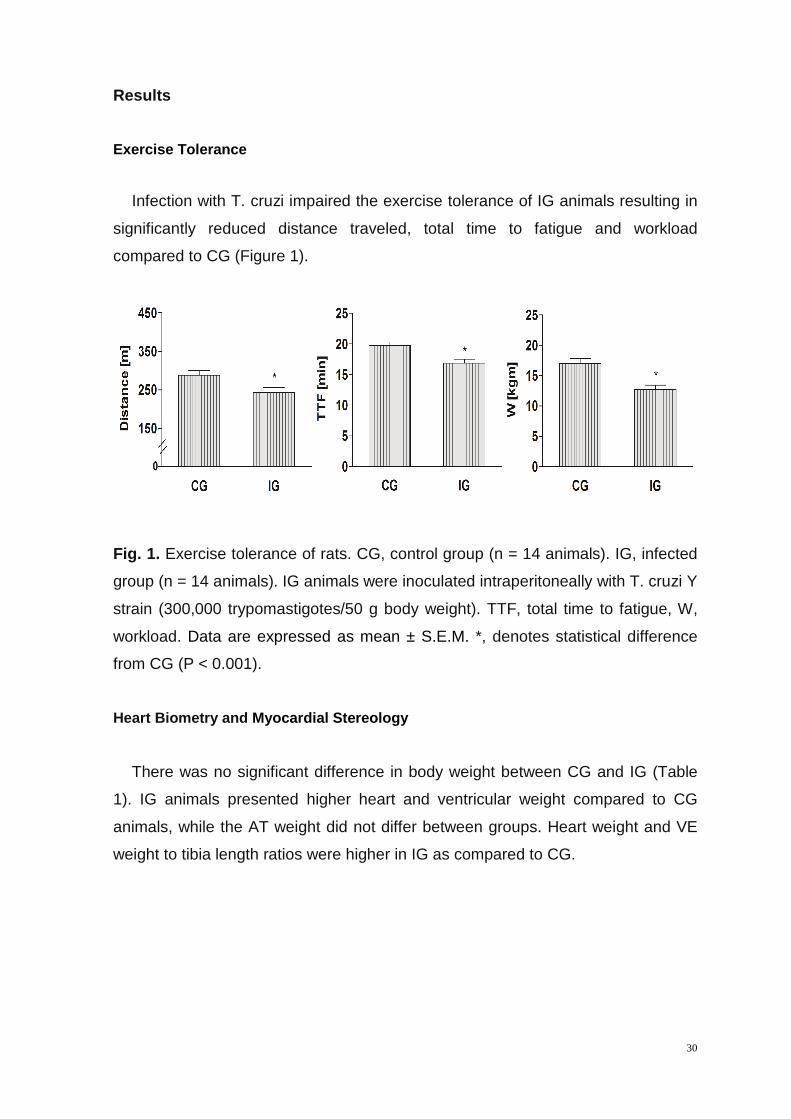
Infection with T. cruzi impaired the exercise tolerance of IG animals resulting in
significantly reduced distance traveled, total time to fatigue and workload
compared to CG (Figure 1).
Fig. 1. Exercise tolerance of rats. CG, control group (n = 14 animals). IG, infected
group (n = 14 animals). IG animals were inoculated intraperitoneally with T. cruzi Y
strain (300,000 trypomastigotes/50 g body weight). TTF, total time to fatigue, W,
workload. Data are expressed as mean ± S.E.M. *, denotes statistical difference
from CG (P < 0.001).
Heart Biometry and Myocardial Stereology
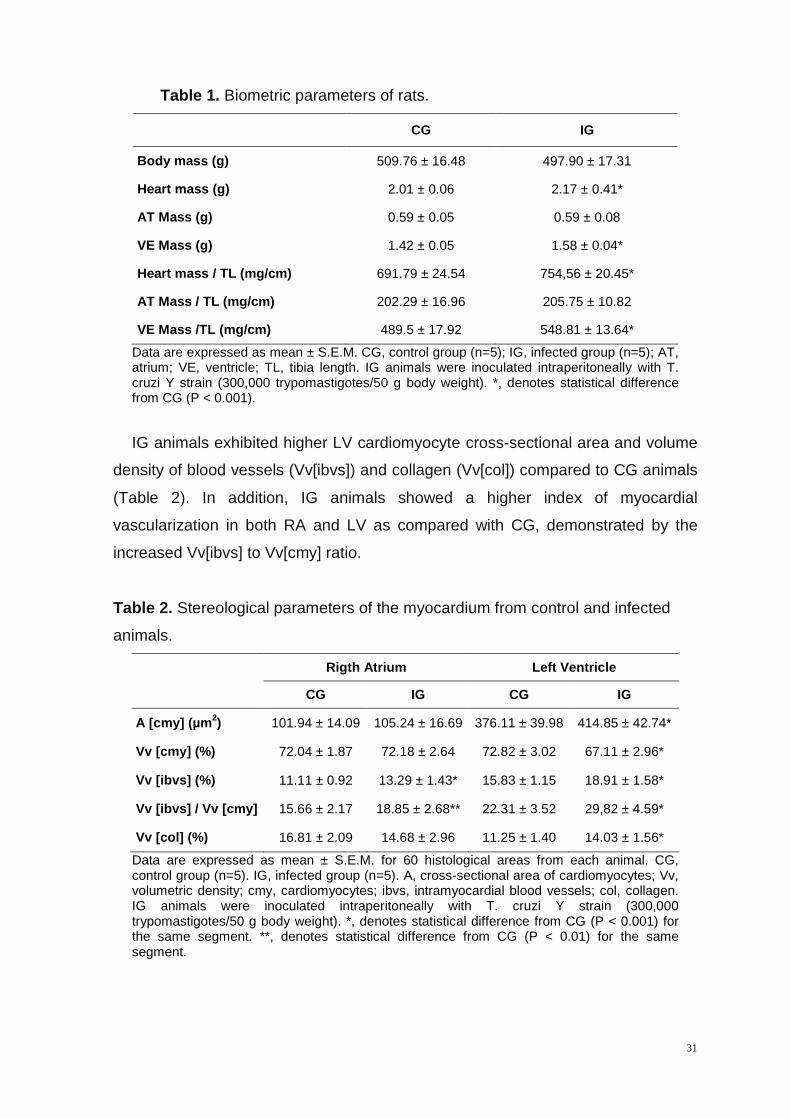
There was no significant difference in body weight between CG and IG (Table
1). IG animals presented higher heart and ventricular weight compared to CG
animals, while the AT weight did not differ between groups. Heart weight and VE
weight to tibia length ratios were higher in IG as compared to CG.
31
Table 1. Biometric parameters of rats.
CG IG
Body mass (g) 509.76 ± 16.48 497.90 ± 17.31
Heart mass (g) 2.01 ± 0.06 2.17 ± 0.41*
AT Mass (g) 0.59 ± 0.05 0.59 ± 0.08
VE Mass (g) 1.42 ± 0.05 1.58 ± 0.04*
Heart mass / TL (mg/cm) 691.79 ± 24.54 754,56 ± 20.45*
AT Mass / TL (mg/cm) 202.29 ± 16.96 205.75 ± 10.82
VE Mass /TL (mg/cm) 489.5 ± 17.92 548.81 ± 13.64*
Data are expressed as mean ± S.E.M. CG, control group (n=5); IG, infected group (n=5); AT, atrium; VE, ventricle; TL, tibia length. IG animals were inoculated intraperitoneally with T. cruzi Y strain (300,000 trypomastigotes/50 g body weight). *, denotes statistical difference from CG (P < 0.001).
IG animals exhibited higher LV cardiomyocyte cross-sectional area and volume
density of blood vessels (Vv[ibvs]) and collagen (Vv[col]) compared to CG animals
(Table 2). In addition, IG animals showed a higher index of myocardial
vascularization in both RA and LV as compared with CG, demonstrated by the
increased Vv[ibvs] to Vv[cmy] ratio.
Table 2. Stereological parameters of the myocardium from control and infected
animals.
Rigth Atrium Left Ventricle
CG IG CG IG
A [cmy ] (µm 2) 101.94 ± 14.09 105.24 ± 16.69 376.11 ± 39.98 414.85 ± 42.74*
Vv [cmy ] (%) 72.04 ± 1.87 72.18 ± 2.64 72.82 ± 3.02 67.11 ± 2.96 *
Vv [ ibvs ] (%) 11.11 ± 0.92 13.29 ± 1.43* 15.83 ± 1.15 18.91 ± 1.5 8*
Vv [ ibvs ] / Vv [ cmy ] 15.66 ± 2.17 18.85 ± 2.68** 22.31 ± 3.52 29,82 ± 4.59*
Vv [col ] (%) 16.81 ± 2.09 14.68 ± 2.96 11.25 ± 1.40 14.03 ± 1.56 *
Data are expressed as mean ± S.E.M. for 60 histological areas from each animal. CG, control group (n=5). IG, infected group (n=5). A, cross-sectional area of cardiomyocytes; Vv, volumetric density; cmy, cardiomyocytes; ibvs, intramyocardial blood vessels; col, collagen. IG animals were inoculated intraperitoneally with T. cruzi Y strain (300,000 trypomastigotes/50 g body weight). *, denotes statistical difference from CG (P < 0.001) for the same segment. **, denotes statistical difference from CG (P < 0.01) for the same segment.
32
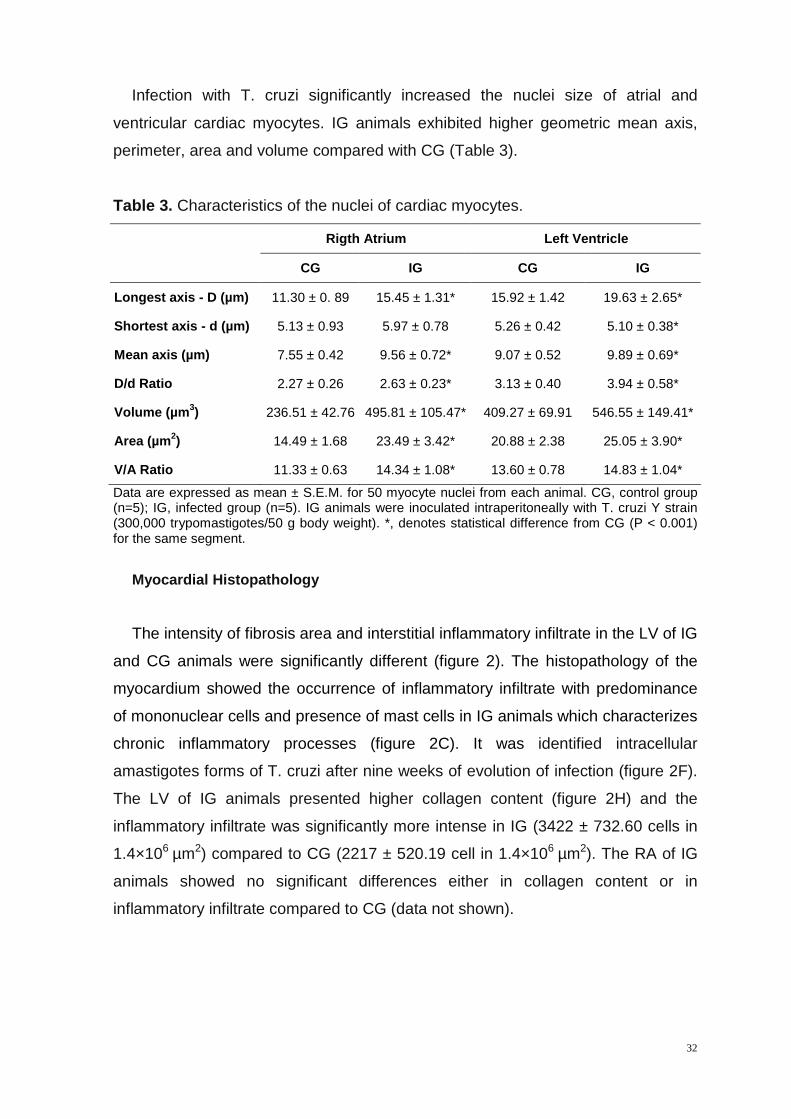
Infection with T. cruzi significantly increased the nuclei size of atrial and
ventricular cardiac myocytes. IG animals exhibited higher geometric mean axis,
perimeter, area and volume compared with CG (Table 3).
Table 3. Characteristics of the nuclei of cardiac myocytes.
Rigth Atrium Left Ventricle
CG IG CG IG
Longest axis - D (µm) 11.30 ± 0. 89 15.45 ± 1.31* 15.92 ± 1.42 19.63 ± 2. 65*
Shortest axis - d (µm) 5.13 ± 0.93 5.97 ± 0.78 5.26 ± 0.42 5.10 ± 0.38*
Mean axis (µm) 7.55 ± 0.42 9.56 ± 0.72* 9.07 ± 0.52 9.89 ± 0.69*
D/d Ratio 2.27 ± 0.26 2.63 ± 0.23* 3.13 ± 0.40 3.94 ± 0.58*
Volume (µm 3) 236.51 ± 42.76 495.81 ± 105.47* 409.27 ± 69.91 546.55 ± 149.41*
Area (µm 2) 14.49 ± 1.68 23.49 ± 3.42* 20.88 ± 2.38 25.05 ± 3.9 0*
V/A Ratio 11.33 ± 0.63 14.34 ± 1.08* 13.60 ± 0.78 14.83 ± 1.0 4*
Data are expressed as mean ± S.E.M. for 50 myocyte nuclei from each animal. CG, control group (n=5); IG, infected group (n=5). IG animals were inoculated intraperitoneally with T. cruzi Y strain (300,000 trypomastigotes/50 g body weight). *, denotes statistical difference from CG (P < 0.001) for the same segment.
Myocardial Histopathology
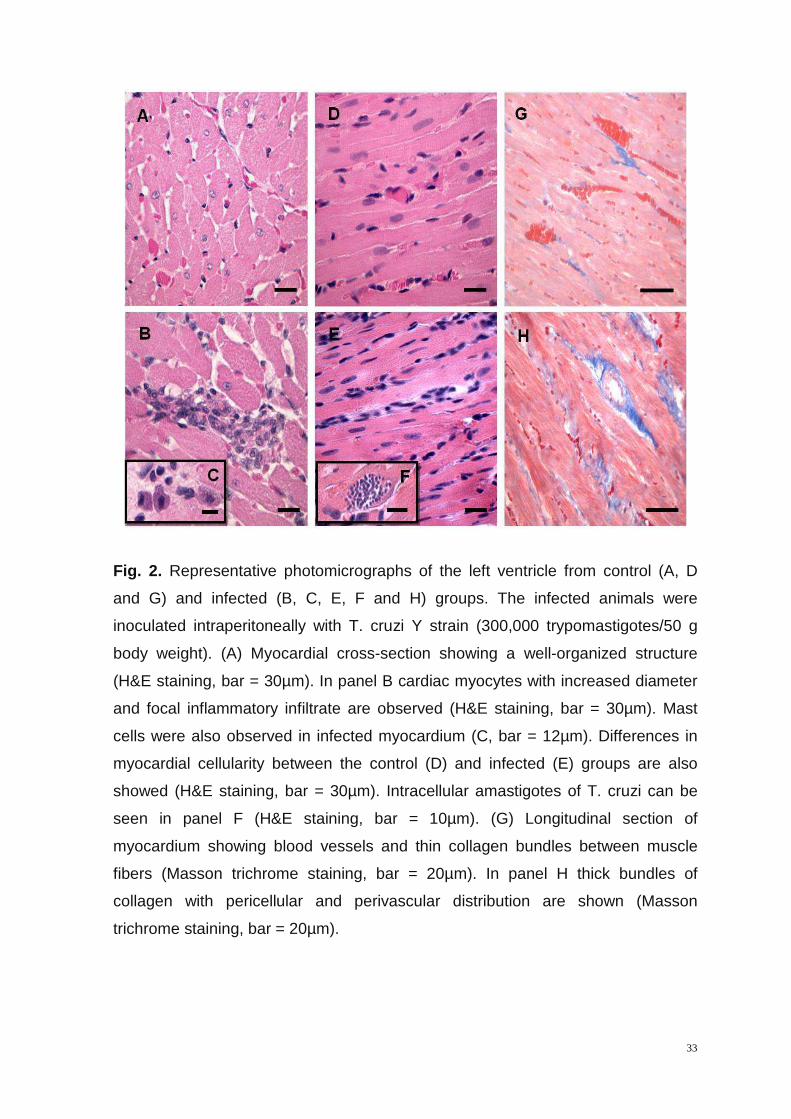
The intensity of fibrosis area and interstitial inflammatory infiltrate in the LV of IG
and CG animals were significantly different (figure 2). The histopathology of the
myocardium showed the occurrence of inflammatory infiltrate with predominance
of mononuclear cells and presence of mast cells in IG animals which characterizes
chronic inflammatory processes (figure 2C). It was identified intracellular
amastigotes forms of T. cruzi after nine weeks of evolution of infection (figure 2F).
The LV of IG animals presented higher collagen content (figure 2H) and the
inflammatory infiltrate was significantly more intense in IG (3422 ± 732.60 cells in
1.4×106 µm2) compared to CG (2217 ± 520.19 cell in 1.4×106 µm2). The RA of IG
animals showed no significant differences either in collagen content or in
inflammatory infiltrate compared to CG (data not shown).
33
Fig. 2. Representative photomicrographs of the left ventricle from control (A, D
and G) and infected (B, C, E, F and H) groups. The infected animals were
inoculated intraperitoneally with T. cruzi Y strain (300,000 trypomastigotes/50 g
body weight). (A) Myocardial cross-section showing a well-organized structure
(H&E staining, bar = 30µm). In panel B cardiac myocytes with increased diameter
and focal inflammatory infiltrate are observed (H&E staining, bar = 30µm). Mast
cells were also observed in infected myocardium (C, bar = 12µm). Differences in
myocardial cellularity between the control (D) and infected (E) groups are also
showed (H&E staining, bar = 30µm). Intracellular amastigotes of T. cruzi can be
seen in panel F (H&E staining, bar = 10µm). (G) Longitudinal section of
myocardium showing blood vessels and thin collagen bundles between muscle
fibers (Masson trichrome staining, bar = 20µm). In panel H thick bundles of
collagen with pericellular and perivascular distribution are shown (Masson
trichrome staining, bar = 20µm).
34
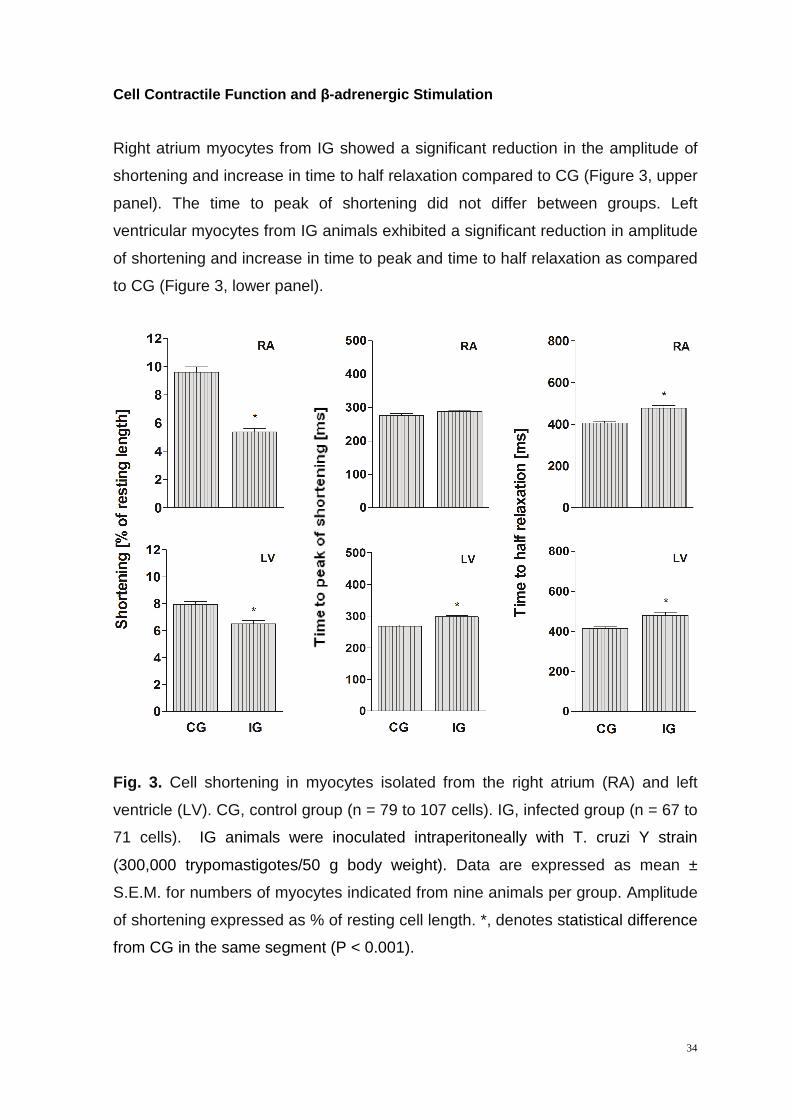
Cell Contractile Function and β-adrenergic Stimulation
Right atrium myocytes from IG showed a significant reduction in the amplitude of
shortening and increase in time to half relaxation compared to CG (Figure 3, upper
panel). The time to peak of shortening did not differ between groups. Left
ventricular myocytes from IG animals exhibited a significant reduction in amplitude
of shortening and increase in time to peak and time to half relaxation as compared
to CG (Figure 3, lower panel).
Fig. 3. Cell shortening in myocytes isolated from the right atrium (RA) and left
ventricle (LV). CG, control group (n = 79 to 107 cells). IG, infected group (n = 67 to
71 cells). IG animals were inoculated intraperitoneally with T. cruzi Y strain
(300,000 trypomastigotes/50 g body weight). Data are expressed as mean ±
S.E.M. for numbers of myocytes indicated from nine animals per group. Amplitude
of shortening expressed as % of resting cell length. *, denotes statistical difference
from CG in the same segment (P < 0.001).
35
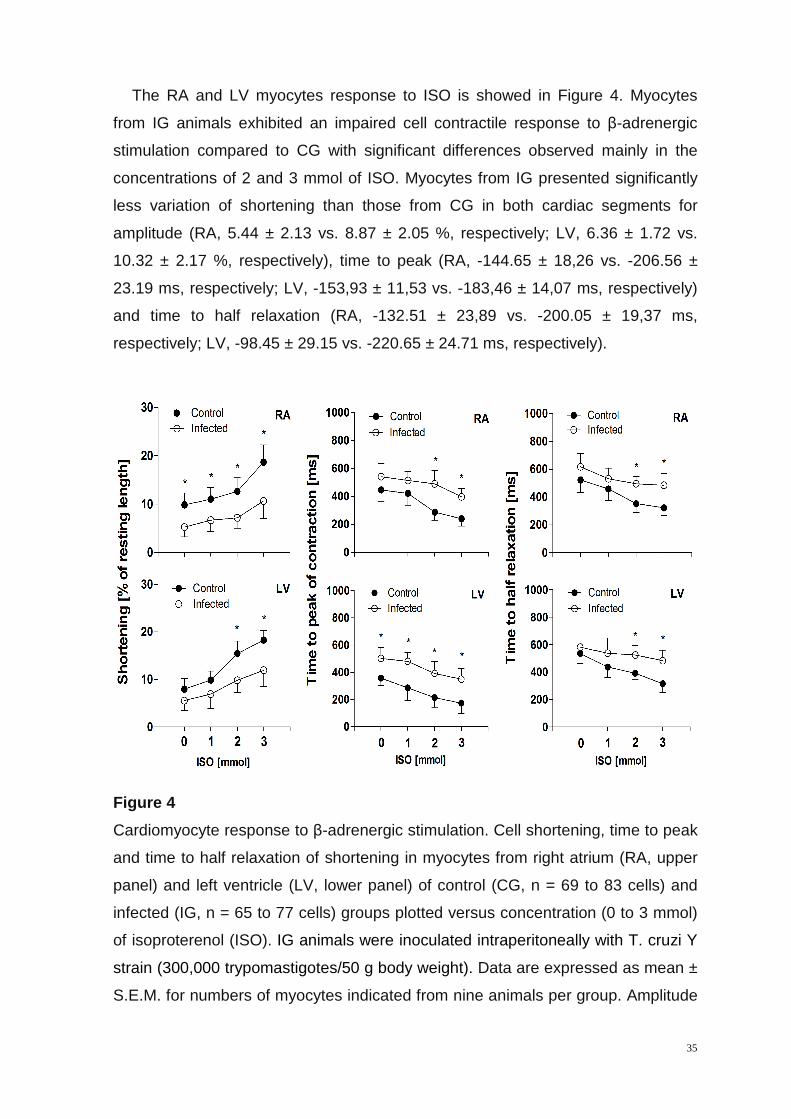
The RA and LV myocytes response to ISO is showed in Figure 4. Myocytes
from IG animals exhibited an impaired cell contractile response to β-adrenergic
stimulation compared to CG with significant differences observed mainly in the
concentrations of 2 and 3 mmol of ISO. Myocytes from IG presented significantly
less variation of shortening than those from CG in both cardiac segments for
amplitude (RA, 5.44 ± 2.13 vs. 8.87 ± 2.05 %, respectively; LV, 6.36 ± 1.72 vs.
10.32 ± 2.17 %, respectively), time to peak (RA, -144.65 ± 18,26 vs. -206.56 ±
23.19 ms, respectively; LV, -153,93 ± 11,53 vs. -183,46 ± 14,07 ms, respectively)
and time to half relaxation (RA, -132.51 ± 23,89 vs. -200.05 ± 19,37 ms,
respectively; LV, -98.45 ± 29.15 vs. -220.65 ± 24.71 ms, respectively).
Figure 4
Cardiomyocyte response to β-adrenergic stimulation. Cell shortening, time to peak
and time to half relaxation of shortening in myocytes from right atrium (RA, upper
panel) and left ventricle (LV, lower panel) of control (CG, n = 69 to 83 cells) and
infected (IG, n = 65 to 77 cells) groups plotted versus concentration (0 to 3 mmol)
of isoproterenol (ISO). IG animals were inoculated intraperitoneally with T. cruzi Y
strain (300,000 trypomastigotes/50 g body weight). Data are expressed as mean ±
S.E.M. for numbers of myocytes indicated from nine animals per group. Amplitude

36
of shortening expressed as % of resting cell length. *, denotes statistical difference
from CG in the same segment (P < 0.001).
Discussion
Our results showed that exercise tolerance was significantly impaired in animals
infected with T. cruzi. In addition, infected rats exhibited myocardial
histopathological damage and RA and LV cardiomyocyte contractile dysfunction.
Infected animals showed cardiac hypertrophy which was evidenced by the
presence of cellular hypertrophy and increased collagen amount in the
myocardium. The abnormal pattern of accumulation and organization of collagen
during the progression of the disease has been previously described in Chagas’
disease-induced pathological cardiac hypertrophy (Higuchi et al., 1999; Marin-
Neto et al., 2007; Rassi-Jr et al., 2010). This new organization of collagen fibers
may decrease the myocardial mechanical efficiency inasmuch as part of the force
used for pumping blood is diverted to correct the geometric distortion determined
by the abnormal organization of collagen and muscle bundles (Mady et al., 1999).
Moreover, the progressive accumulation of collagen reduces the myocardium
compliance and the efficiency of the regulatory mechanism of cellular and muscle
contraction force based on the length-tension relationship (Kitzman et al., 1991;
Higuchi et al., 1999).
The results of volume density of blood vessels and the relationship blood
vessel/cell did not indicate reduction in myocardial vascularization in the infected
animals. However, these findings do not exclude the possibility of vascular
dysfunction of vasomotor origin and inadequate balance in the blood flow
distribution. For example, our stereological data indicated the occurrence of
microvascular dilatation that may result of altered blood flow induced by diffuse
fibrosis and vascular derangement. The presence of inflammatory infiltrate nine
weeks after infection with T. cruzi and the occurrence of mast cells favor this
hypothesis. Indeed, mechanisms such as endothelial dysfunction, persistence of
T. cruzi antigens and release of nitric oxide associated with the chronic
inflammatory process have been additionally implicated in vascular dilatation and
dysfunction in Chagas’ disease (Higuchi et al., 1999; Marin-Neto et al., 2007). The
presence of vascular damage are not unusual in T. cruzi infection and the

37
reduction of myocardial vascularization has been considered as an important
component involved in deterioration of cardiac function (Higuchi et al., 1999; Biolo
et al., 2010) and exercise tolerance (Meiler et al., 1987). Previous studies have
shown that myocardial hypoperfusion significantly limits the exercise tolerance due
to the occurrence of abnormal heart rhythm with the onset of arrhythmias and
cardiac pump dysfunction (Verani et al., 1981; Meiler et al., 1987).
Our data showed that T. cruzi infection reduced the contractile function of RA
and LV myocyte (i.e. reduced cell shortening amplitude and increased time to
peak and time to half relaxation). Mechanisms such as down-regulation of ion
channels that modulate calcium flux and cell contraction and relaxation has been
implicated in the pathogenesis of cardiomyocyte mechanical dysfunction observed
in heart disease of different etiologies (Wisloff et al., 2002; Kemi and Wisloff,
2010). In myocardial infarction, diabetic cardiomyopathy and autoimmune
myocarditis, the reduced expression and / or inhibition of sodium and calcium
exchanger of the sarcolemma (NCX), ryanodine channel (RyR2), phospholamban
(PLB) and calcium ATPase (SERCA-2) of the sarcoplasmic reticulum has been
reported as important events associated with the chronotropic, inotropic and
lusotropic cardiomyocytes dysfunction (Wisloff et al., 2002; Afanasyeva et al.,
2004; Kemi and Wisloff, 2010). However, if these molecular changes are promoted
by T. cruzi infection warrants further investigations.
Previous studies have demonstrated positive relationship between improved
cardiomyocyte mechanical properties and parameters of exercise performance
such as higher maximal oxygen consumption (Wisloff et al., 2002; Kemi et al.,
2004) and intrinsic aerobic exercise capacity in normal/healthy rats (Prímola-
Gomes et al., 2009). There is evidence that in animals without disease (Kemi et
al., 2004, 2005; kemi and Wisloff, 2010) and in animal models of cardiovascular
disease, the improvement in mechanical properties of cardiomyocytes due to
chronic physical exercise programs occurs in association with increased density
and sensitivity of calcium ion channels of the sarcolemma and sarcoplasmic
reticulum. An important finding is that these contractile and molecular adaptations
of cardiomyocytes in response to physical exercise are accompanied by a
simultaneous improvement in physical performance (Wisloff et al., 2002; kemi and
Wisloff, 2010). It is believed that the results of high physical fitness are are due to
a better provision and use of oxygen to exercised tissues, and that part of this

38
adaptation is due to the high capacity of cells and myocardium to produce a
greater cardiac output (Kemi and Wisloff, 2010). In this context, it is not unrealistic
to assume that conditions that impair cellular contractility and consequently
myocardial function can potentially reduce physical capacity and exercise
tolerance. In Chagas’ disease, reduction in physical capacity has been widely
linked to disturbances in cardiac mechanics and hemodynamics (Gallo et al.,
1975; Lima et al., 2010).
We also observed that myocytes from T. cruzi infected animals presented
reduced sensibility to β-adrenergic stimulation. The inotropic and lusitropic
response to ISO proved to be dose dependent in both IG and CG groups,
however, all contractile parameters examined showed lower amplitude of variation
in animals from IG. In clinical and experimental studies of Chagas’ disease,
physical and pharmacological cardiac tests have showed a reduced ability of the
myocardium to respond to stimuli of progressive intensity, suggesting a lower
cellular functional reserve (Galo et al., 1975; Talvani et al., 2006; Sousa et al.,
2009; Lima et al., 2010). In this disease, changes in electrical and mechanical
cardiac function have been more pronounced in conditions of cardiac stress, as
occurs during exercise and the reduction of cardiac responsiveness to β-
adrenergic stimulation has been considered an important factor involved in
reducing exercise tolerance (Gallo et al., 1975; Molina et al., 1981; Colucci et al.,
1989). Data exist to support the role of the immune system in pathological
remodeling of cardiomyocyte contractility (Sterin-Borda et al., 1999; Chakraborti et
al., 2000; Afanasyeva et al., 2004), including in Chagas’ disease (Roman-Campos
et al., 2009). It has been demonstrated that in humans and experimental animals
with Chagas' disease anti-β1-adrenoreceptor antibodies produced during infection
by T. cruzi can inhibit the signaling pathway triggered by this receptor (Sterin-
Borda et al., 1999; Chakraborti et al., 2000). Under normal conditions, β-
Adrenergic pathways lead to the phosphorylation and inhibition of PLB, which
reduces its activity on SERCA-2 and hence improved inotropic, lusotropic and
chronotropic activity of cardiomyocites (Afanasyeva et al., 2004). However, direct
allosteric inhibition of β1-reptores by autoantibodies or desensitization mediated by
up-regulation of β-adrenergic-receptor kinase (βARK) may significantly impair
cardiomyocyte contractile function since this receptor is the main signaling
pathway that regulates cellular mechanics through adjustments in Ca2+ kinetic

39
(Afanasyeva et al., 2004). Furthermore, inhibition of β1 signaling reduces the
phosphorylation and activation of RyR2 and Ca2+ entry into the cell via the L-type
current mediated by Ca2+-induced Ca2+-release mechanism (Afanasyeva et al.,
2004).
In summary, we showed that experimental infection with T. cruzi significantly
reduced the exercise tolerance of rats and negatively influenced the myocardial
morphology and mechanical properties of single RA and LV myocytes. The results
of cell mechanics associated to β-adrenergic stimulation support the hypothesis
that disturbances of cardiomyocyte contractility in animals infected with T. cruzi
could constitute an additional mechanism of cardiac contractile function
impairment and reduced exercise tolerance. Little is known about the influence of
the parasite in the signaling pathways through which it acts to modulate the
cardiomyocyte mechanics. Thus, further studies are needed to clarify this issue.
References
Brener Z. Therapeutic activity and criterion of cure on mice experimentally
infected with Trypanosoma cruzi. Rev Inst Med Trop São Paulo 1962; 4:389-
96.
Brooks GA, Donovan CM, White TP. Estimation of anaerobic energy
production and efficiency in rats during exercise J Appl Physiol 1984;56:520-5.
Caldas IS, Talvani A, Caldas S, Carneiro CM, Lana M, Guedes PMM, et al.
Benznidazole therapy during acute phase of Chagas disease reduces parasite
load but does not prevent chronic cardiac lesions. Parasitol Res 2008;103:413-
21.
Koch LG, Britton SL. Artificial selection for intrinsic aerobic endurance running
capacity in rats. Physiol Genomics 2001;5:45-52.

40
Lacerda AR, Marubayashi U, Balthazar, CH, Coimbra CC. Evidence that brain
nitric oxide inhibition increases metabolic cost of exercise, reducing running
performance in rats. Neuroscience Letters 2006;393:260-3.
Martinelli PM, Camargos ERS, Azevedo AA, Chiari E, Morel G, Machado CRS.
Cardiac NGF and GDNF expression during Trypanosoma cruzi infection in
rats. Autonomic Neuroscience: Basic and Clinical 2006;130:32-40.
Natali AJ, Turner DL, Harrison SM, Wire E. Regional effects of voluntary
exercise on cell size and contraction-frequency responses in rat cardiac
myocyte. J Exp Biol 2001;204:1191-99.
Sala MA, Komesu MC, Lopes RA, Maia CG. Karyometric study of basal cell
carcinoma. Braz Dental J 1994;5:11-4.
Mandarim-de-Lacerda CA. Stereological tools in biomedical research. An Acad
Bras Ciênc 2003;75:469-86.
Bezerra DGA, Andrade LML, Cruz FOP, Mandarim-de-lacerda CA. Atorvastatin
attenuates cardiomyocyte loss in adult rats from protein-restricted dams. J
Card Fail 2008;14:151-60.
WHO, 2005 World Health Organization, 2005. Tropical Disease Research:
Progress 2003-2004 Seventeenth Programme Report of the
UNICEF/UNDP/World Bank/WHO. Special Programme for Research &
Training in Tropical Diseases (TRD). Programme Report, n. 17, Geneva, pp.
31–33.
Rassi-Jr A, Rassi A, Marin-Neto JÁ. Chagas disease. Lancet 2010;375:1388-
1402.
Talvani A, Rocha MOC, Ribeiro AL, Borda E, Sterin-Borda L, Teixeira MM.
Levels of anti-M2 and anti-β1 autoantibodies do not correlate with the degree

41
of heart dysfunction in Chagas’ heart disease. Microbes and Infection
2006;8:2459-64.
Mady C, Ianni BM, Arteaga E, Salemi VMC, Frimm CC. Maximal functional
capacity in patients with chagas’ cardiomyopathy without congestive heart
failure. J Card Fail 2000;3:220-4.
Lima MMO, Pereira MC, Rocha MOC, Beloti FR, Alencar MCN, Ribeiro ALP.
Left ventricular diastolic function and exercise capacity in patients with Chagas
cardiomyopathy. Echocardiography 2010;27:519-24.
Sousa LAP, Rocha MOC, Britto RR, Lombardi F, Ribeiro AL. Chagas disease
alters the relationship between heart rate variability and daily physical activity.
Int J Cardiol 2009;135:257-9.
Gallo L Jr, Neto JA, Manco JC, Rassi A, Amorim DS. Abnormal heart rate
responses during exercise in patients with Chagas’ disease. Cardiology
1975;60:147-62.
Molina AR, Carrasco HG, Milanés J, Molina CA, Pacheco J, Fuenmayor AP. La
prueba de esfuerzo en la miocardiopatía chagásica crónica. Su valor en el
diagnóstico precoz. El comportamiento de las arritmias ventriculares y los
transtornos de conducción al ejercicio en las fases más avanzadas de La
enfermedad. Arq Bras Cardiol 1981;36:95-100. (in spanish)
Roman-Campos D, Duarte HLL, Sales-Jr PA, Natali AJ, Ropert C, Gazzinelli
RT, et al. Changes in cellular contractility and cytokines profile during
Trypanosoma cruzi infection in mice. Basic Res Cardiol 2009; 104:238-46.
Prímola-Gomes TN, Campos LA, Lauton-Santos S, Balthazar CH, Guatimosim
S, Capettini LS A, et al. Exercise capacity is related to calcium transients in
ventricular cardiomyocytes. J Appl Physiol 2009;107:593-8.

42
Wisloff U, Najjar SM, Ellingsen O, Haram PM, Swoap S, Al-Share Q, et al.
Cardiovascular risk factors emerge after artificial selection for low aerobic
capacity. Science 2005;307:418-20.
Kemi OJ, Haram PM, Wisloff U, Ellingsen O. Aerobic fitness is associated with
cardiomyocyte contractile capacity and endothelial function in exercise training
and detraining. Circulation 2004;109:2897-904.
Wisloff U, Loennechen JP, Currie S, Smith GL, Ellingsen O. Aerobic exercise
reduces cardiomyocyte hypertrophy and increases contractility, Ca2+ sensitivity
and SERCA-2 in rat after myocardial infarction. Cardiovasc Res 2002;54:162-
74.
Kemi OJ, Wisloff U. Mechanisms of exercise-induced improvements in the
contractile apparatus of the mammalian myocardium. Acta Physiol
2010;199:425-39.
Colucci WS, Ribeiro JP, Rocco MB, Quigg RJ, Creager MA, Marsh JD, et al.
Impaired chronotropic response to exercise in patients with congestive heart
failure: role of postsynaptic beta-adrenergic desensitization. Circulation
1989;80:314-23.
Marin-Neto JA, Cunha-Neto E, Maciel BC, Simões MV. Pathogenesis of
chronic Chagas’ heart disease. Circulation 2007;115:1109-23.
Biolo A, Ribeiro AL, Clausell N. Chagas cardiomyopathy-where do we stand
after a hundred years? Prog Cardiovasc Dis 2010;52:300-16.
Higuchi ML, Fukasawa S, Brito T, Parzianello LC, Bellotti G, Ramires JAF.
Different microcirculatory and interstitial matrix patterns in idiopathic dilated
cardiomyopathy and Chagas’ disease: a three dimensional confocal
microscopy study. Heart 1999;82:279-85.

43
Mady C; Ianni BM, Arteaga E; Montes GS; Caldini EG, Andrade G, et al.
Relation between interstitial myocardial collagen and the degree of clinical
impairment in Chagas’ disease. Am J Cardiol 1999;84:354-6.
Afanasyeva M, Georgakopoulos D, Rose NR. Autoimmune myocarditis: cellular
mediators of cardiac dysfunction. Autoimmunity Reviews 2004;3:476-86.
Kitzman DW, Higginbotham MB, Cobb FR, Sheikh KH, Sullivan MJ. Exercise
intolerance in patients with heart failure and preserved left ventricular systolic
function: failure of the Frank-Starling mechanism. J Am Coll Cardiol
1991;17:1065-72.
Meiler SEL, Ashton JJ, Moeschberger ML, Unverferth DV, Leier CV. An
analysis of the determinants of exercise performance in congestive heart
failure. Am Heart J 1987;113:1207-17.
Verani MS, Hartung GH, Hoepfel-Harris J, Welton DE, Pratt CM, Miller RR.
Effects of exercise training on left ventricular performance and myocardial
perfusion in patients with coronary artery disease. Am J Cardiol 1981;47:797-
803.
Sterin-Borda L, Gorelik G, Postan M, Gonzalez Cappa S, Borda E. Alterations
in cardiac beta-adrenergic receptors in chagasic mice and their association
circulating beta-adrenoceptor-related autoantibodies. Cardiovasc Res
1999;41:116-25.
Chakraborti S, Chakraborti T, Shaw G. Beta-adrenergic mechanisms in cardiac
diseases: a perspective. Cell Signal 2000;12:499-513.
Yan SM, Finato N, Di Loreto C, Beltrami CA. Nuclear size of myocardial cells in
end-stage cardiomyopathies. Anal Quant Cytol Histol 1999;21:174-80.
Prahash AJC, Gupta S, Anand IS. Myocyte response to β-adrenergic
stimulation is preserved in the noninfarcted myocardium of globally

44
dysfunctional rat hearts after myocardial infarction. Circulation 2000;102:1840-
46.

45
Perspectivas
Alterações patológicas do miocárdio e deterioração das propriedades
contráteis de cardiomiócitos foram encontradas no presente estudo. Até o
momento, pouco se sabe a respeito dos mecanismos por meio dos quais o T.
cruzi atua para alterar a contratilidade. Existem dados na literatura que indicam
influência do sistema imunológico na modulação da contratilidade celular em
cardiopatias de origem não chagásica. Entretanto, a contribuição do sistema
imunológico na disfunção mecânica celular evidenciada na doença de Chagas
permanece pouco conhecida e requer novos estudos, principalmente em relação
ao papel das citocinas e quimiocinas na modulação da contratilidade celular.
Embora a investigação das alterações da morfologia cardíaca e mecânica
celular possam contribuir para o conhecimento atual da cardiopatia chagásica e
explicar parcialmente a redução da tolerância ao exercício físico, ainda existem
questionamentos a respeito dos fatores que limitam a capacidade de realizar
exercícios físicos na doença de Chagas, visto que trata-se de uma doença
sistêmica. Nesse contexto, elementos morfo-funcionais de outros sistemas que
são comprometidos na infecção com T. cruzi e que estão direta ou indiretamente
envolvidos nos ajustes requeridos à prática de exercícios físicos devem ser
adicionalmente investigados. Abordagem especial deve ser direcionada aos
músculos esqueléticos e pâncreas uma vez que apresentam relação direta com o
metabolismo energético e a conversão de energia em trabalho físico. Nesse
contexto, a análise da função pancreática, metabolismo de glicose e suas vias de
utilização pelos músculos exercitados podem fornecer novas informações sobre
os mecanismos por meio dos quais a infecção com T. cruzi modifica a tolerância
ao exercício físico.
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo











![parasito schistosoma[1]](https://static.fdocumentos.tips/doc/165x107/5571fb6f497959916994e04d/parasito-schistosoma1.jpg)






