
DO PRODUTOR AO CONSUMIDOR: INFLUÊNCIA DA … · isômeros de CLA, em particular o C18:2 cis-9,...
31
I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte DO PRODUTOR AO CONSUMIDOR: INFLUÊNCIA DA NUTRIÇÃO SOBRE A QUALIDADE DA CARNE BOVINA Marcio Machado Ladeira 1 , Mario Luiz Chizzotti 1 , Otávio Rodrigues Machado Neto 1 , Dalton Mendes de Oliveira 1 , Leandro Samia Lopes 1 , Eduardo Mendes Ramos 2 1 Departamento de Zootecnia da Universidade Federal de Lavras – UFLA; 2 Departamento de Ciência de Alimentos da Universidade Federal de Lavras - UFLA 1- Introdução Atualmente, em várias regiões do mundo, os consumidores estão cada vez mais exigentes quanto à qualidade dos alimentos e informados da relação entre dieta e saúde. Portanto, a Zootecnia moderna deve se preocupar ainda mais com a qualidade dos alimentos produzidos, dando preferência por aqueles mais saudáveis. Devido à sua composição lipídica, constituída em grande parte pelos ácidos graxos saturados (AGS), a carne bovina é considerada uma das carnes com maior efeito prejudicial à saúde humana. Entretanto, tem sido amplamente demonstrado que ácidos graxos insaturados AGI de cadeia longa participam de vários processos metabólicos benéficos à saúde humana (Cook et al., 2001; Varela et al., 2004) e que as gorduras da carne de animais ruminantes são fontes naturais de alguns desses ácidos graxos, como o ácido oleico e os isômeros de CLA, em particular o C18:2 cis-9, trans-11 (French et al., 2000). No entanto, apenas os aspectos negativos são ressaltados por grande parte dos profissionais de saúde e pela mídia de massa, ignorando-se a importância da carne bovina como um dos componentes de uma dieta saudável. A estratégia dos pesquisadores para aumentar a concentração dos ácidos graxos benéficos na carne bovina está em tentar manipular a biohidrogenação ruminal dos AGI, com o intuito de reduzir a concentração dos AGS e alguns ácidos graxos trans. A literatura cita que isto pode ser alcançado por meio da utilização de fontes de lipídeos na dieta. Entretanto, quais as melhores fontes para esta manipulação ainda não estão bem descritas. Neste artigo serão abordados os principais fatores ligados ao metabolismo de lipídeos no ruminante e como estes podem influenciar a qualidade da carne bovina. Para tanto, serão apresentados alguns resultados
Transcript of DO PRODUTOR AO CONSUMIDOR: INFLUÊNCIA DA … · isômeros de CLA, em particular o C18:2 cis-9,...

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
DO PRODUTOR AO CONSUMIDOR: INFLUÊNCIA DA NUTRIÇÃO SOBRE A
QUALIDADE DA CARNE BOVINA
Marcio Machado Ladeira1, Mario Luiz Chizzotti1, Otávio Rodrigues Machado
Neto1, Dalton Mendes de Oliveira1, Leandro Samia Lopes1, Eduardo Mendes
Ramos2
1 Departamento de Zootecnia da Universidade Federal de Lavras – UFLA; 2 Departamento de Ciência de Alimentos da Universidade Federal de Lavras - UFLA 1- Introdução
Atualmente, em várias regiões do mundo, os consumidores estão cada
vez mais exigentes quanto à qualidade dos alimentos e informados da relação
entre dieta e saúde. Portanto, a Zootecnia moderna deve se preocupar ainda
mais com a qualidade dos alimentos produzidos, dando preferência por
aqueles mais saudáveis.
Devido à sua composição lipídica, constituída em grande parte pelos
ácidos graxos saturados (AGS), a carne bovina é considerada uma das carnes
com maior efeito prejudicial à saúde humana. Entretanto, tem sido amplamente
demonstrado que ácidos graxos insaturados AGI de cadeia longa participam de
vários processos metabólicos benéficos à saúde humana (Cook et al., 2001;
Varela et al., 2004) e que as gorduras da carne de animais ruminantes são
fontes naturais de alguns desses ácidos graxos, como o ácido oleico e os
isômeros de CLA, em particular o C18:2 cis-9, trans-11 (French et al., 2000).
No entanto, apenas os aspectos negativos são ressaltados por grande parte
dos profissionais de saúde e pela mídia de massa, ignorando-se a importância
da carne bovina como um dos componentes de uma dieta saudável.
A estratégia dos pesquisadores para aumentar a concentração dos
ácidos graxos benéficos na carne bovina está em tentar manipular a
biohidrogenação ruminal dos AGI, com o intuito de reduzir a concentração dos
AGS e alguns ácidos graxos trans. A literatura cita que isto pode ser alcançado
por meio da utilização de fontes de lipídeos na dieta. Entretanto, quais as
melhores fontes para esta manipulação ainda não estão bem descritas.
Neste artigo serão abordados os principais fatores ligados ao
metabolismo de lipídeos no ruminante e como estes podem influenciar a
qualidade da carne bovina. Para tanto, serão apresentados alguns resultados

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
de pesquisas desenvolvidas na Universidade Federal de Lavras sobre o tema.
Por fim, também será abordado como o conteúdo de gordura intramuscular da
carne também pode ser influenciado pela nutrição do animal.
2- Biohidrogenação ruminal
Animais ruminantes alimentados com dietas convencionais normalmente
consomem limitadas quantidades de lipídeos. No entanto, a utilização de
lipídeos em dietas de ruminantes permite o aumento da concentração
energética da dieta dos animais. Ácidos graxos produzem mais energia que os
demais nutrientes orgânicos, quando metabolizados pelo animal. Segundo o
NRC (2000) o valor energético de fontes lipídicas é no mínimo duas vezes
superior aos observados para grãos de cereais.
A gordura dietética afeta direta e indiretamente a produção ruminal de
ácidos graxos voláteis, a digestibilidade da dieta e o suprimento de ácidos
graxos para o animal. As ligações éster existentes entre ácidos graxos e o
glicerol são hidrolisadas de maneira rápida e extensiva pelas enzimas
microbianas lipolíticas no rúmen, tendo como consequência a formação de
ácidos graxos livres e glicerol (Jenkins, 1993). O glicerol oriundo desta hidrólise
pode então ser metabolizado pelos micro-organismos ruminais para produzir
ácidos graxos voláteis (Nagaraja et al., 1997).
Os AGI livres, liberados como consequência da hidrólise microbiana,
podem exercer efeitos antimicrobianos no ambiente ruminal (Palmquist &
Jenkins, 1980), o que resultaria em alterações da proporção molar dos ácidos
graxos voláteis (Doreau & Chilliard, 1997). Para reduzir os efeitos tóxicos dos
AGI os micro-organismos ruminais utilizam a biohidrogenação ruminal (Figura
1), sendo que o ácido esteárico (C18:0) é formado ao término das reações
(Jenkins, 1993). Todavia, o acúmulo de ácido linoleico no ambiente ruminal
pode inibir a biohidrogenação completa (Jenkins & Adams, 2002).
Dessa forma, a suplementação da dieta de bovinos com ácidos graxos
insaturados pode aumentar a passagem destes para o intestino delgado, o que
possibilita maior absorção e possibilidade de alteração do perfil de ácidos
graxos da carne destes animais. Os ácidos graxos de cadeia longa não são
utilizados como fonte de energia pelos micro-organismos ruminais, sendo que
os ácidos graxos oriundos da dieta apenas sofrem transformação
(biohidrogenação) pela ação destes. A quantificação dos ácidos graxos que

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
atingem o duodeno para posterior absorção é de grande importância para que
se possa estimar a transferência destes para os tecidos (Merchen et al., 1997).
As taxas de lipólise e biohidrogenação irão depender da quantidade e do
tipo de fonte lipídica fornecida aos animais (Beam et al., 2000) e do pH ruminal
(Van Nevel & Demeyer, 1996). No entanto, o grau médio de biohidrogenação
ruminal é de 70%, podendo variar de 60 a 90% (Zinn et al., 2000).
O ácido linoleico conjugado (C18:2 cis-9, trans-11), produzido como
consequência da biohidrogenação incompleta do ácido linoleico, tem sido alvo
de muitos estudos e efeitos benéficos do seu uso têm sido relatados. Entre
estes, podemos destacar a prevenção de câncer, redução de aterosclerose,
melhoria da resposta imune além de modificações do metabolismo protéico e
energético (Whigham et al., 2000; Belury, 2002; Pariza, 2004; Palmquist et al.,
2005).
cis-9, cis-12, cis-15 – 18:3
cis-9, trans-11, cis-15 – 18:3
trans-11, cis-15 – 18:2
cis-15 e trans-15 18:2 trans-11 18:1
18:0
cis-9, cis-12 18:2
cis-9, trans-11 18:2
trans-11 18:1
Figura 1. Rotas de biohidrogenação dos ácidos α-linolênico (A), linoleico (B) e
oleico (C). Adaptado de Harfoot & Hazlewood (1988).
(A)
(B) (C)
cis-9 – 18:1 trans- 18:1
18:0

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
O C18:2 cis-9, trans-11, está presente em maiores concentrações nos
produtos oriundos de ruminantes, sendo que em produtos oriundos de não-
ruminantes sua concentração é irrisória. Este fenômeno ocorre devido ao fato
de que para a sua síntese, há necessidade de ação das bactérias ruminais
sobre o ácido linoleico. O C18:2 cis-9, trans-11 é considerado o principal
isômero de CLA com benefícios sobre a saúde humana (Pariza, 2004). Além
disso, o ácido graxo C18:1 trans-11 (ácido vaccênico), que também é um
intermediário da biohidrogenação dos ácidos linoleico e linolênico, é substrato
para a formação de C18:2 cis-9, trans-11 no tecido adiposo do animal. Desta
forma, estratégias que possibilitem o aumento no fluxo de ácido vaccênico para
o intestino delgado e sua posterior absorção colaboram de forma decisiva para
o aumento da concentração de CLA na carne bovina.
2.1- Principais micro-organismos envolvidos na biohidrogenação.
A bactéria ruminal Butyrivibrio fibrisolvens foi identificada como um
importante micro-organismo envolvido na biohidrogenação de ácidos graxos e
síntese de C18:2 cis-9, trans-11 e C18:1 trans-11, durante o processo de
biohidrogenação do ácido linoleico (Polan et al., 1964; Kepler et al., 1966).
Entretanto, esta bactéria não está envolvida na formação de ácido esteárico a
partir do ácido linoleico. A maioria das bactérias isoladas do rúmen por Kemp et
al. (1975) convertiam ácido linoleico até C18:1, principalmente trans-11, tendo
sido então denominadas como “Grupo A” por este autor. Bactérias que tinham
a capacidade de produzir ácido esteárico foram denominadas de bactérias do
“Grupo B”, sendo que algumas espécies envolvidas nesta etapa da
biohidrogenação foram identificadas por Kemp et al. (1975) como sendo
Fusocillus spp. Mais recentemente, van de Vossenberg & Joblin (2003)
isolaram de uma vaca em sistema de pastejo, uma bactéria que também
poderia formar ácido esteárico a partir de ácido linoleico, sendo que esta era
fenotipicamente semelhante ao gênero Fusocillus e filogeneticamente próxima
de Butyrivibrio hungatei.
Maia et al. (2006) realizaram experimentos com o objetivo de avaliar a
importância quantitativa de diferentes espécies microbianas na
biohidrogenação ruminal e também para identificar as bactérias mais
importantes na conversão de ácido vaccênico à ácido esteárico. Onze das 26
espécies de bactérias ruminais predominantes no meio, metabolizaram o ácido

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
linoleico de forma substancial, aparentemente através de várias rotas
possíveis. O produto oriundo da biohidrogenação do ácido linoleico mais
comumente observado foi o ácido vaccênico, que foi produzido por três
linhagens de Butyrivibrio e 2 linhagens de Clostridium proteoclasticum. No
entanto, somente C. proteoclasticum, produziu C18:0. Uma vez que o ácido
vaccênico é formado a partir do C18:2 cis-9, trans-11, bactérias que produzem
ácido vaccênico, devem também ser consideradas produtoras deste isômero
de CLA. Apesar de sua nomenclatura, o C. proteoclasticum tem muitas
características semelhantes ao gênero Butyrivibrio.
Wallace et al. (2006) avaliaram a capacidade de bactérias em
metabolizar o ácido linoleico, tendo verificado que apenas um pequeno grupo
de bactérias que produzem CLA e acido vaccênico são capazes de produzir
ácido esteárico. Neste trabalho os autores verificaram que as bactérias
produtoras de ácido esteárico eram morfologicamente semelhantes ao gênero
Fusocillus além também de apresentar propriedades metabólicas próximas das
observadas neste gênero. Através de análises filogenéticas, essas bactérias
foram identificadas como sendo Clostridium proteoclasticum.
Em relação aos protozoários, pode-se afirmar que cerca de metade da
biomassa microbiana ruminal é composta por estes micro-organismos (Williams
& Coleman, 1992) e que aproximadamente 75% dos ácidos graxos microbianos
ruminais estão contidos neles. Diante disso, pode-se inferir que estes micro-
organismos representam uma importante fonte de ácidos graxos poli-
insaturados, ácido linoleico conjugado e ácido vaccênico.
Os fungos anaeróbicos ruminais são os micro-organismos em menor
concentração no ambiente ruminal. No entanto, estes têm grande importância
no metabolismo ruminal devido à sua grande atividade celulolítica (Wilson &
Wood, 1992). Segundo Kemp et al. (1984), o fungo Pyromices communis tem a
capacidade de converter ácido esteárico em ácido oleico. Estes autores
incubaram P. communis em um meio com os ácidos linoleico e oleico e
observaram a formação de ácidos conjugados. Maia et al. (2006) cultivaram
duas espécies de fungos ruminais: P. communis e Neocallimastix frontalis em
um meio com 50 μg/mL de ácido linoleico e observou que não houve
crescimento de P. communis no meio, no entanto o fungo N. frontalis
metabolizou cerca de 50% do ácido linoleico presente no meio, produzindo
C18:2 cis-9, trans-11. No entanto, a capacidade deste fungo em formar CLA é

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
muito baixa quando comparada à atividade observada para a bactéria B.
fibrisolvens.
2.2- Digestibilidade ruminal dos nutrientes
O efeito da suplementação com lipídeos sobre a digestibilidade ruminal
dos nutrientes tem sido extensivamente relatado na literatura, principalmente
no que diz respeito ao seu efeito sobre a digestibilidade ruminal da fibra.
Doreau & Chilliard (1997) verificaram que o decréscimo da relação
acetato/propionato no rúmen dos animais suplementados com gordura foi
acompanhada por redução na digestibilidade ruminal da matéria orgânica,
principalmente da fração fibrosa. Segundo Hess (2008), em experimento
realizado com a suplementação de óleo de soja em dietas com alta proporção
de volumoso, a inclusão de mais que 6% deste alimento na MS da dieta
resultou em decréscimo da digestão da fibra. A fonte de lipídeos tem marcante
efeito sobre a fermentação ruminal. Fontes de gordura com baixa proporção de
ácidos graxos insaturados, como o sebo hidrogenado, muito utilizado em dietas
de vacas leiteiras e bovinos em crescimento nos Estados Unidos parecem
afetar pouco a fermentação ruminal.
Bateman & Jenkins (1998) avaliaram o efeito da inclusão de niveis
crescentes de óleo de soja (0, 2, 4, 6 ou 8% da MS da dieta) sobre a
digestibilidade total dos nutrientes dietéticos. Não foi verificado efeito do nível
de suplementação sobre a digestibilidade da MS, MO, PB e FDN. Segundo os
autores, a fermentação nos intestinos pode ter compensado a redução na
digestibilidade ruminal destes nutrientes. Em situações onde a suplementação
lipídica afeta a digestibilidade ruminal dos nutrientes mas não afeta a
digestibilidade total, tem-se um ganho em termos de eficiência energética, uma
vez que a produção de metano poderá ser suprimida.
Montgomery et al. (2008) avaliou o efeito de diferentes fontes de óleo
(sebo bovino, gérmen de milho, óleo de milho e óleo de canola) além de um
tratamento sem lipídio suplementar sobre a digestibilidade ruminal da fibra em
bovinos em terminação submetidos a dieta com 90% de concentrado. Todas
dietas continham 6,7% de lipídeos na MS, exceto a dieta sem lipídeo
suplementar, com 3,7% de lipídeos. Não foi observado efeito dos tratamentos
sobre a digestibilidade da FDN.

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
Palmquist (1988) afirmou que o acréscimo de forragem na dieta dilui os
efeitos negativos da gordura insaturada. De forma geral, os efeitos da inclusão
de gordura em dietas de bovinos têm sido inconsistentes. Em alguns
experimentos (Zinn, 1988; Zinn, 1992, Doreau et al., 1991) a gordura
suplementar não teve efeito sobre a digestão da fibra. No entanto, em outras
pesquisas (Zinn, 1989, Zinn, 1994; Zinn & Plascencia, 1992, Zinn & Plascencia,
1993; Elliot et al., 1997) a inclusão de gordura afetou fortemente a digestão da
fibra dietética. Segundo Plascencia et al. (1999) a variabilidade de respostas à
suplementação lipídica pode estar relacionada ao pH ruminal. Zinn (1988)
avaliou o efeito da inclusão de 4% de óleo de fritura residual em dieta com 88%
de concentrado para novilhos cruzados e não observou efeito da dieta sobre a
digestibilidade ruminal do amido, FDA e proteína, quando comparado ao
tratamento controle, sem gordura suplementar.
Duas teorias para explicar o efeito inibitório dos ácidos graxos têm sido
propostas (Jenkins, 1993). A primeira afirma que o lipídeo envolve o micro-
organismo com um filme hidrofóbico, impedindo o metabolismo e também
interferindo na aderência da bactéria à celulose. Outro efeito inibitório tem sido
atribuído ao efeito dos ácidos graxos insaturados sobre a fluidez das
membranas citoplasmáticas, além de poder afetar a sua permeabilidade.
Segundo Hess (2008), para ruminantes alimentados com dietas de alto
grão, a suplementação da gordura em 6% da MS dietética causa poucos
efeitos negativos sobre a utilização dos outros nutrientes dietéticos. Todavia,
em dietas com alta inclusão de forragem, este autor afirma que o nível ótimo de
inclusão de lipídeos seria de 4% da MS dietética.
O uso de lipídeos em dietas de ruminantes também pode produzir
efeitos benéficos sobre o metabolismo do nitrogênio no rúmen. De acordo com
Doreau & Ferlay (1995), a redução na concentração de amônia no rúmen é
uma das principais características da defaunação ruminal. Ikwuegbu & Sutton
(1982) realizaram a infusão de óleo de linhaça diretamente no rúmen de ovinos
e observaram redução na digestão ruminal da proteína além de redução da
concentração ruminal de amônia e aumento no fluxo de nitrogênio para o
duodeno. Decréscimos na população de protozoários no rúmen são
frequentemente associados à redução na concentração ruminal de amônia,
primariamente como um resultado de um decréscimo na proteólise de proteína
bacteriana por protozoários ruminais (Broderick, 1991). A magnitude da

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
atividade anti-protozoários dos ácidos graxos depende basicamente do seu
grau de insaturação (Hristov et al., 2005), sendo assim, é de se esperar que
fontes ricas em ácido linolênico (óleo de linhaça) tenham grande capacidade
em deprimir a população ruminal de protozoários.
Hristov et al. (2009) realizaram a suplementação de ácidos graxos
saturados de cadeia média (ácido láurico e mirístico) na dieta de vacas
holandesas em lactação e verificaram que estes ácidos foram efetivos em
reduzir a concentração ruminal de amônia, quando comparada à dieta controle.
Assim como observado quando ocorre a suplementação com ácidos graxos
insaturados, a inclusão destes ácidos de cadeia média têm efeito defaunatório,
reduzindo a proteólise da proteína bacteriana por protozoários.
As fontes de lipídeos suplementar utilizadas em dietas de ruminantes
parecem afetar pouco a digestibilidade de carboidratos não fibrosos (CNF).
(Zinn, 1994; Zinn & Plascencia, 1996). Entretanto, Montgomery et al. (2008)
utilizando diferentes fontes de óleo (sebo, gérmen de milho, óleo de canola e
óleo de milho) observaram queda na digestão ruminal aparente do amido
quando comparado ao tratamento controle. Porém, a digestibilidade total do
amido não foi alterada pelos tratamentos. Zinn & Shen (1996) avaliando o
efeito de dois níveis de cálcio (0,45 e 0,90%) e dois níveis de óleo de fritura
residual (0 ou 5% da MS) não observaram efeito dos tratamentos sobre a
digestibilidade ruminal de amido, em dietas com 88% de concentrado.
3- Perfil de ácidos graxos e qualidade da carne bovina
Nos últimos anos tem aumentado o interesse em manipular a
composição do perfil de ácidos graxos da carne (Wood et al., 2003), com o
intuito de aumentar a proporção de ácidos graxos considerados saudáveis
(Beaulieu et al., 2002), pois, de acordo com Mills et al. (1992) os esforços
realizados no sentido de melhorar a produtividade através de cruzamentos ou
de mudanças na nutrição somente serão completos se forem consideradas a
composição e a palatabilidade da carne.
Sinclair et al. (1982) ao avaliarem o perfil de ácidos graxos na carne de
bovinos encontraram os teores de 49,0; 42,2 e 8,8% para os AGS, AGMI e
AGPI, respectivamente. Sendo que os ácidos graxos mais representativos na
carne bovina são o oleico, o palmítico e o esteárico (Cutrignelli et al., 1996).

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
As gorduras contendo AGS, em geral, elevam as lipoproteínas de baixa
densidade (LDL). Considera-se nesta situação o efeito hipercolesterolêmico
dos AGS: láurico, mirístico e palmítico, por diminuírem a atividade dos
receptores hepáticos da LDL (Givens, 2005), aumentando sua concentração no
plasma.
Por outro lado a razão ω6/ω3 é particularmente benéfica na carne de
ruminates que consomem gramíneas ou fontes oleaginosas com elevado teor
de C18:3 (Enser, 2001). De acordo com recomendações nutricionais, o
aumento no consumo de AGPI da série ω-3 tem sido recomendado na tentativa
de não exceder essa razão, que segundo a World Health Organization (2003)
deve ser entre 4:1 a 5:1.
O ácido oleico presente na carne de ruminantes é oriundo da
biohidrogenação ruminal (Lanna et al., 2001), bem como da dessaturação
endógena do ácido esteárico (Corl et al., 1998), e tem a vantagem de não
reduzir as lipoproteínas de alta densidade (HDL), atuando contra o surgimento
de doenças coronarianas (Molketin, 2000).
Felton & Kerley (2004) avaliaram o perfil de ácidos graxos de bovinos
alimentados com dietas tradicionais, à base de farelo de soja e milho, e dietas
com altos níveis de lipídeos. Os autores encontraram que o músculo dos
animais que receberam maiores teores de lipídeos apresentou menores
concentrações dos ácidos mirístico e palmítico. Da mesma forma, Scollan et al.
(2001) conseguiram aumentar os teores de AGPI na carne de novilhos
recebendo gordura protegida de linhaça e óleo de peixe.
Medeiros et al. (2005), estudando bovinos de três grupos genéticos,
confinados com dieta controle (3,0% EE), ou dieta rica em óleo (6,0% EE) à
base de caroço de algodão, na proporção de 9,5% da matéria seca (MS), não
encontraram diferenças significativas para a maioria dos ácidos graxos
presentes no tecido adiposo dos animais. Os pesquisadores concluíram que a
adição de caroço de algodão nesses níveis não foi suficiente para alterar o
perfil de ácidos graxos da carcaça.
Huerta-Leidenz et al. (1991) também não detectaram diferenças na
composição dos ácidos graxos da gordura da carcaça de novilhos de corte
alimentados com dietas contendo 30% de caroço de algodão, durante 54 dias
de confinamento.

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
Por outro lado, estudos citados por Duckett (2001) relataram redução na
porcentagem de AGS na carne de novilhos alimentados com 5% de óleo de
girassol ou semente de canola. Todavia, a adição de semente de algodão à
dieta de terminação não alterou a pontuação de marmorização nem a
composição da gordura subcutânea.
Madron et al. (2002) avaliaram o perfil de ácidos graxos na carne de
bovinos alimentados com diferentes quantidades de soja extrusada. Com os
resultados encontrados foram mostrados maiores teores de CLA na carne dos
animais que consumiram a dieta com alta quantidade de soja extrusada. Em
outra pesquisa, com o intuito, também, de determinar o teor de CLA, Mir et al.
(2004) forneceram dietas com e sem óleo de girassol para diferentes raças
bovinas e conseguiram elevar de forma bem significativa, aproximadamente,
em quatro vezes, o teor de CLA. De acordo com o autor, a adição de óleo pode
ser utilizada para alterar a composição do perfil de ácidos graxos da gordura
intramuscular de bovinos, mas não alteraria o seu conteúdo. Embora a adição
de óleo possa aumentar o conteúdo de CLA da gordura muscular, esse
aumento relativo poderia, também, ser afetado por outros componentes da
dieta.
Avaliando o efeito de diferentes fontes de lipídeos sobre o teor de CLA e
o perfil de ácidos graxos no músculo e na capa de gordura em novilhos
bubalinos, Oliveira et al. (2008) observaram que o fornecimento de óleo de soja
resultou em maior concentração de CLA e menores concentrações de AGS,
principalmente, os ácidos mirístico e palmítico no músculo e na capa de
gordura. Os animais que receberam a dieta com grão de soja integral
apresentaram teor de CLA semelhante à dieta sem lipídeo adicional.
Entretanto, houve redução nas concentrações de mirístico e palmítico.
3.1- Diferentes fontes lipídicas usadas na terminação de bovinos de corte:
influência sobre a qualidade de carne
Três experimentos (Oliveira, 2010; Machado Neto, não publicado;
Santarosa, não publicado) foram conduzidos no Departamento de Zootecnia da
UFLA entre 2008 e 2010 com o objetivo de estudar o efeito de diferentes fontes
lipídicas sobre a qualidade da carne de bovinos terminados em sistema de
confinamento.
I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
EXPERIMENTO 1: Qualidade da carne de novilhos Zebuínos recebendo
diferentes grãos de oleaginosas (Oliveira, 2010)
Metodologia
Foram utilizados 31 novilhos Zebuínos castrados, com idade inicial
média de 23 meses e peso vivo inicial de 365 +37,5 kg. Os animais foram
confinados durante 112 dias, ente os meses de agosto a novembro de 2008.
As dietas tiveram a silagem de milho como volumoso e quatro diferentes
tipos de concentrados foram utilizados, contendo diferentes oleaginosas
moídas. Assim, cada tipo de concentrado representou um determinado
tratamento: sem lipídeo adicional, grão de soja, caroço de algodão e semente
de linhaça (Tabela 1).
TABELA 1: Composição percentual de ingredientes e química das dietas experimentais: sem lipídeo adicional (SLA), grão de soja (GS), caroço de algodão (CA) e semente de linhaça (SL)
Ingredientes Composição (%MS)
SLA GS CA SL
Silagem de Milho 40,0 40,0 40,0 40,0 Milho Integral 49,2 43,8 37,2 43,8 Farelo de Soja 9,0 - 3,0 7,8 Soja Grão - 14,4 - - Caroço de Algodão - - 18,0 - Linhaça - - - 6,6 Núcleo Mineral 1,8 1,8 1,8 1,8
Nutrientes
MS¹ 64,2 64,7 64,7 64,4 PB² 13,2 13,3 13,3 13,5 FDN² 33,3 32,1 41,2 34,5 CNF² 45,0 43,2 34,2 42,3 NDT² 74,9 76,1 75,4 75,2 EE² 3,5 6,1 6,7 5,8
¹ - base da matéria natural, ² - base da matéria seca
As oleaginosas foram moídas em peneira de malha de 5 mm com a
finalidade de aumentar a disponibilidade ruminal dos lipídeos. O diâmetro
Geométrico Médio (DGM) foi de 823, 446 e 849 µm, respectivamente, para o
grão de soja, caroço de algodão e semente de linhaça.
O abate dos animais foi realizado em frigorífico comercial, utilizando a
técnica de concussão cerebral e secção da veia jugular. As amostras para
análises foram extraídas do músculo Longissimus dorsi da meia-carcaça
esquerda, a partir da 13a costela, em direção cranial.
I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
Resultados
O teor de extrato etéreo ou a inclusão de grãos de oleaginosas na dieta
não influenciaram a composição centesimal do músculo Longissimus dorsi e as
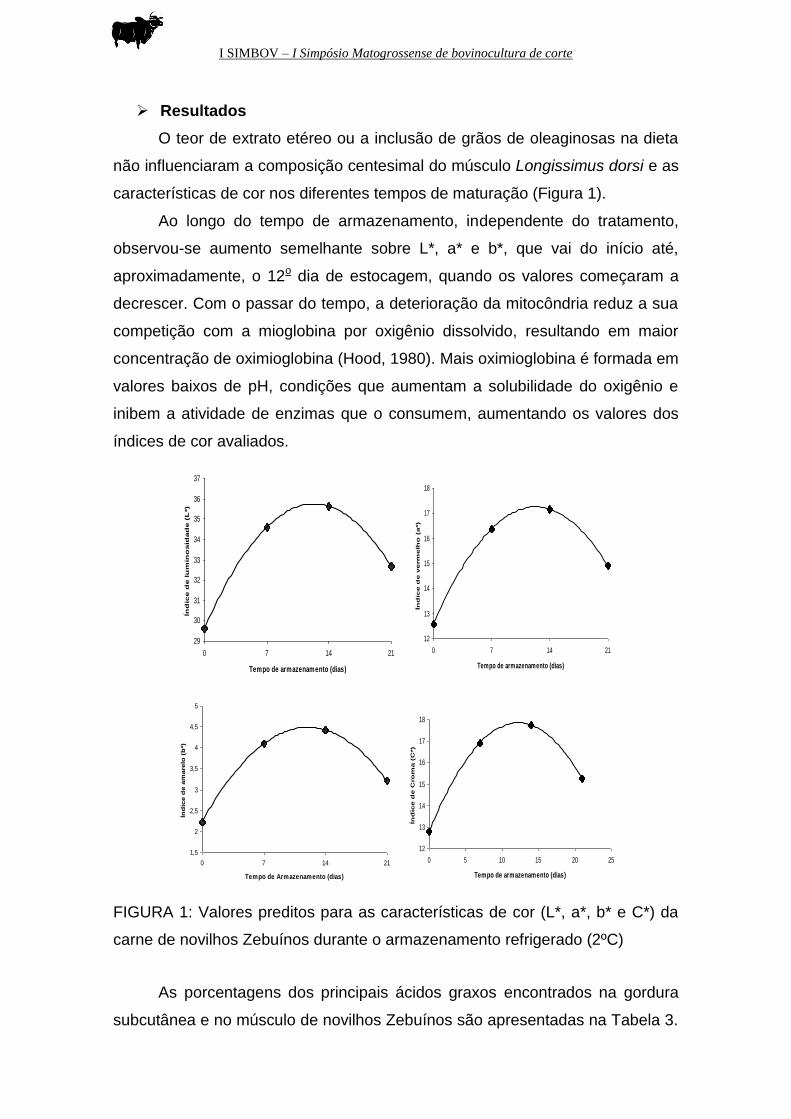
características de cor nos diferentes tempos de maturação (Figura 1).
Ao longo do tempo de armazenamento, independente do tratamento,
observou-se aumento semelhante sobre L*, a* e b*, que vai do início até,
aproximadamente, o 12o dia de estocagem, quando os valores começaram a
decrescer. Com o passar do tempo, a deterioração da mitocôndria reduz a sua
competição com a mioglobina por oxigênio dissolvido, resultando em maior
concentração de oximioglobina (Hood, 1980). Mais oximioglobina é formada em
valores baixos de pH, condições que aumentam a solubilidade do oxigênio e
inibem a atividade de enzimas que o consumem, aumentando os valores dos
índices de cor avaliados.
FIGURA 1: Valores preditos para as características de cor (L*, a*, b* e C*) da
carne de novilhos Zebuínos durante o armazenamento refrigerado (2ºC)
As porcentagens dos principais ácidos graxos encontrados na gordura
subcutânea e no músculo de novilhos Zebuínos são apresentadas na Tabela 3.
29
30
31
32
33
34
35
36
37
0 7 14 21
Tempo de armazenamento (dias)
Índ
ice d
e lu
min
osid
ad
e (
L*)
12
13
14
15
16
17
18
0 7 14 21
Tempo de armazenamento (dias)
Índ
ice d
e v
erm
elh
o (
a*)
1,5
2
2,5
3
3,5
4
4,5
5
0 7 14 21
Tempo de Armazenamento (dias)
Índ
ice d
e a
mare
lo (
b*)
12
13
14
15
16
17
18
0 5 10 15 20 25
Tempo de armazenamento (dias)
Índ
ice d
e C
ro
ma (
C*)
I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
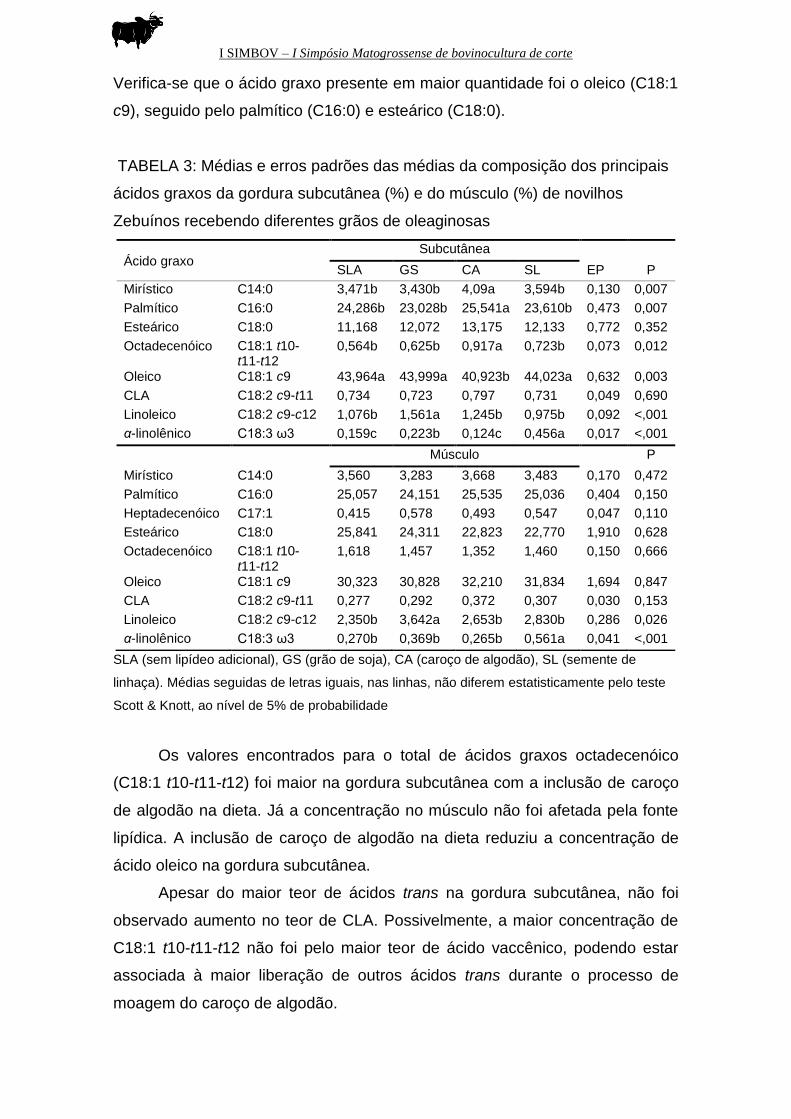
Verifica-se que o ácido graxo presente em maior quantidade foi o oleico (C18:1
c9), seguido pelo palmítico (C16:0) e esteárico (C18:0).
TABELA 3: Médias e erros padrões das médias da composição dos principais
ácidos graxos da gordura subcutânea (%) e do músculo (%) de novilhos
Zebuínos recebendo diferentes grãos de oleaginosas
SLA (sem lipídeo adicional), GS (grão de soja), CA (caroço de algodão), SL (semente de
linhaça). Médias seguidas de letras iguais, nas linhas, não diferem estatisticamente pelo teste
Scott & Knott, ao nível de 5% de probabilidade
Os valores encontrados para o total de ácidos graxos octadecenóico
(C18:1 t10-t11-t12) foi maior na gordura subcutânea com a inclusão de caroço
de algodão na dieta. Já a concentração no músculo não foi afetada pela fonte
lipídica. A inclusão de caroço de algodão na dieta reduziu a concentração de
ácido oleico na gordura subcutânea.
Apesar do maior teor de ácidos trans na gordura subcutânea, não foi
observado aumento no teor de CLA. Possivelmente, a maior concentração de
C18:1 t10-t11-t12 não foi pelo maior teor de ácido vaccênico, podendo estar
associada à maior liberação de outros ácidos trans durante o processo de
moagem do caroço de algodão.
Ácido graxo Subcutânea
SLA GS CA SL EP P
Mirístico C14:0 3,471b 3,430b 4,09a 3,594b 0,130 0,007
Palmítico C16:0 24,286b 23,028b 25,541a 23,610b 0,473 0,007
Esteárico C18:0 11,168 12,072 13,175 12,133 0,772 0,352
Octadecenóico C18:1 t10-t11-t12
0,564b 0,625b 0,917a 0,723b 0,073 0,012
Oleico C18:1 c9 43,964a 43,999a 40,923b 44,023a 0,632 0,003
CLA C18:2 c9-t11 0,734 0,723 0,797 0,731 0,049 0,690
Linoleico C18:2 c9-c12 1,076b 1,561a 1,245b 0,975b 0,092 <,001
α-linolênico C18:3 ω3 0,159c 0,223b 0,124c 0,456a 0,017 <,001
Músculo P
Mirístico C14:0 3,560 3,283 3,668 3,483 0,170 0,472
Palmítico C16:0 25,057 24,151 25,535 25,036 0,404 0,150
Heptadecenóico C17:1 0,415 0,578 0,493 0,547 0,047 0,110
Esteárico C18:0 25,841 24,311 22,823 22,770 1,910 0,628
Octadecenóico C18:1 t10-t11-t12
1,618 1,457 1,352 1,460 0,150 0,666
Oleico C18:1 c9 30,323 30,828 32,210 31,834 1,694 0,847
CLA C18:2 c9-t11 0,277 0,292 0,372 0,307 0,030 0,153
Linoleico C18:2 c9-c12 2,350b 3,642a 2,653b 2,830b 0,286 0,026
α-linolênico C18:3 ω3 0,270b 0,369b 0,265b 0,561a 0,041 <,001
I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
As dietas com grão de soja e semente de linhaça apresentaram os
maiores valores de linoleico e α-linolênico, respectivamente. A diferença no teor
destes ácidos graxos entre os alimentos explica a maior concentração no
músculo e na gordura subcutânea, pois, provavelmente ocorreu maior escape
ruminal.
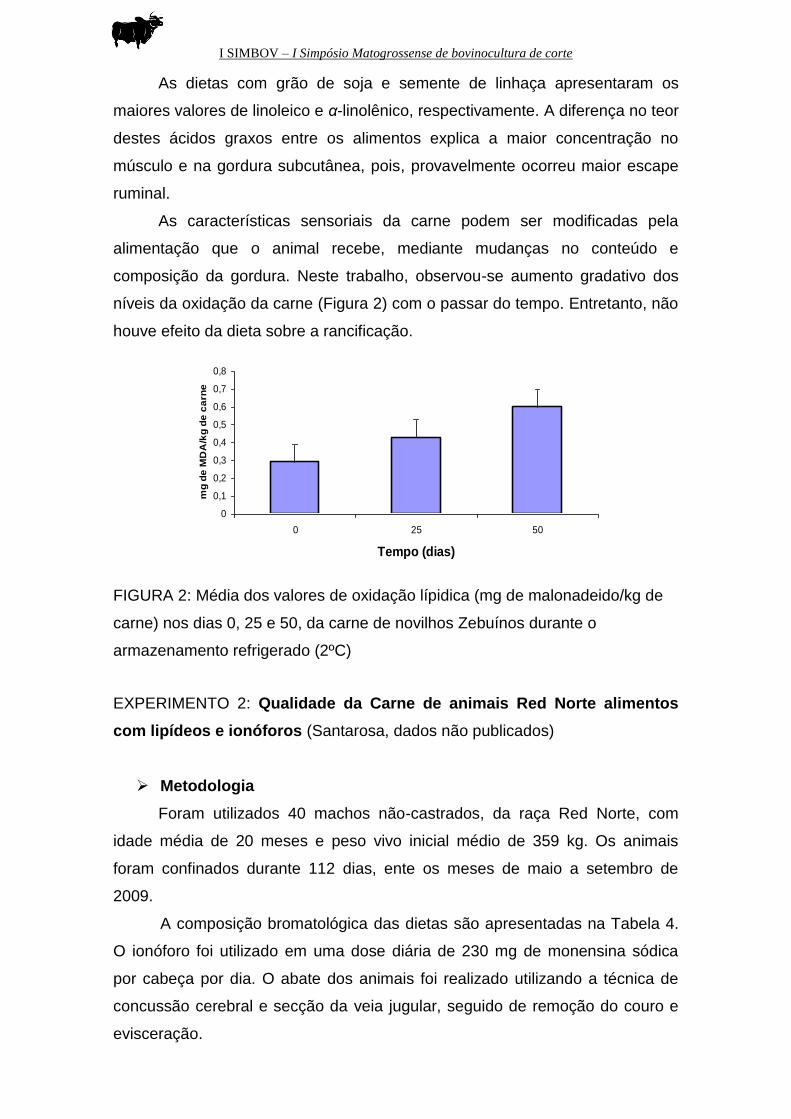
As características sensoriais da carne podem ser modificadas pela
alimentação que o animal recebe, mediante mudanças no conteúdo e
composição da gordura. Neste trabalho, observou-se aumento gradativo dos
níveis da oxidação da carne (Figura 2) com o passar do tempo. Entretanto, não
houve efeito da dieta sobre a rancificação.
FIGURA 2: Média dos valores de oxidação lípidica (mg de malonadeido/kg de
carne) nos dias 0, 25 e 50, da carne de novilhos Zebuínos durante o
armazenamento refrigerado (2ºC)
EXPERIMENTO 2: Qualidade da Carne de animais Red Norte alimentos
com lipídeos e ionóforos (Santarosa, dados não publicados)
Metodologia
Foram utilizados 40 machos não-castrados, da raça Red Norte, com
idade média de 20 meses e peso vivo inicial médio de 359 kg. Os animais
foram confinados durante 112 dias, ente os meses de maio a setembro de
2009.
A composição bromatológica das dietas são apresentadas na Tabela 4.
O ionóforo foi utilizado em uma dose diária de 230 mg de monensina sódica
por cabeça por dia. O abate dos animais foi realizado utilizando a técnica de
concussão cerebral e secção da veia jugular, seguido de remoção do couro e
evisceração.
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0 25 50
Tempo (dias)
mg
de
MD
A/k
g d
e c
arn
e
I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
TABELA 4: Composição percentual de ingredientes e química das dietas
experimentais: grão de soja sem monensina (GS), grão de soja + monensina
(GSM), gordura protegida sem monensina (GP) e gordura protegida +
monensina (GPM)
Ingredientes Composição (%MS)
GS GSM GP GPM
Silagem de milho
40,0 40,0 40,0 40,0
Milho grão moído
41,1 41,1 40,2 40,2
Farelo de soja - - 13,8 13,8 Grão de soja 17,1 17,1 - - Núcleo mineral 1,8 1,8 1,8 1,8 Megalac (GP) Monensina*
- -
- 230 mg/dia
4,2 -
4,2 230 mg/dia
Nutrientes
MS¹ 67,67 67,67 64,85 64,85 Proteína Bruta² 12,88 12,88 12,48 12,48 FDN² 30,88 30,88 28,02 28,02 CNF² 45,74 45,74 48,47 48,47 Extrato Étereo² 6,89 6,89 7,41 7,41
¹ - base da matéria natural, ² - base da matéria seca
Resultados
A monensina sódica praticamente não alterou o perfil de ácidos graxos
na gordura subcutânea dos principais ácidos graxos estudados (Tabela 4).
Apenas o ácido linolênico foi influenciado, em que a gordura subcutânea dos
animais recebendo ionóforos apresentou maior concentração.
A dieta com grão de soja fez com que houvesse menor concentração
dos ácidos mirístico e palmítico. Por outro lado o teor de C18:0 se elevou.
Estes resultados demonstram que a alimentação com grão de soja reduziu na
gordura subcutânea a concentração dos ácidos graxos saturados
hipercolesterolêmicos.
A inclusão de gordura protegida aumentou a concentração de ácido
oleico e CLA na gordura subcutânea. No entanto, as concentrações dos ácidos
graxos essenciais (linoleico e linolênico) foram superiores na gordura dos
animais alimentados com grão de soja, o que provavelmente ocorreu devido ao
maior teor destes ácidos na soja.
I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
A monensina sódica, na quantidade utilizada, também não alterou o
perfil de ácidos graxos no músculo dos animais alimentados com grão de soja
ou gordura protegida. Portanto, apesar deste aditivo alterar a microbiota
ruminal, a dose utilizada neste experimento pode não ter alterado de forma
significativa a biohidrogenação ruminal.
As fontes de lipídeos não afetaram a concentração de ácido mirístico e
palmítico no músculo dos animais, mesmo tendo a gordura protegida maior
concentração destes ácidos graxos. Todavia, os animais que receberam a
gordura protegida apresentaram maior teor de ácido oleico no músculo, que
pode ser explicado pela proteção parcial desta fonte de lipídeo à
biohidrogenação ruminal. O teor de CLA (C18:2 cis-9 trans-11) foi um pouco
superior no músculo de animais alimentados com grão de soja e pode ser
justificado pela maior exposição dos ácidos graxos da soja à biohidrogenação.
Esta explicação também justifica a maior concentração de ácido esteárico
quando a dieta com grão de soja foi utilizada.
TABELA 4: Médias e erros padrões dos valores das concentrações dos
principais ácidos graxos presentes na gordura subcutânea e no músculo de
animais Red Norte alimentados com grão de soja (GS), grão de soja +
monensina (GSM), gordura protegida (GP) e gordura protegida + monensina
(GPM)
Ácido graxo Subcutânea
EP
Probabilidade
GS GSM GP GPM L M LxM
Mirístico C14:0 3,15 3,09 3,86 3,54 0,2
1
<0,0
1
0,38 0,54
Palmítico C16:0 22,7
3
23,6
3
25,2
1
24,4
6
0,6
3
0,01 0,90 0,18
Esteárico C18:0 22,8
8
19,3
8
17,7
6
18,7
9
1,2
5
0,02 0,31 0,06
Oleico C18:1 c9 37,3
5
39,2
2
41,0
5
41,5
0
1,4
8
0,04 0,42 0,62
CLA C18:2 c9-
t11
0,41 0,33 0,91 0,77 0,0
6
<0,0
1
0,10 0,62
Linoleico C18:2c9-
c12
3,39 3,27 1,22 1,46 0,2
1
<0,0
1
0,80 0,39
Linolênico C18:3 n3 0,64 0,92 0,27 0,40
b
0,0
7
<0,0
1
<0,01 0,27
I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
L: efeito para a fonte de lipídeo; M: efeito para o uso da monensina; LxM: interação entre
lipídeo e monensina
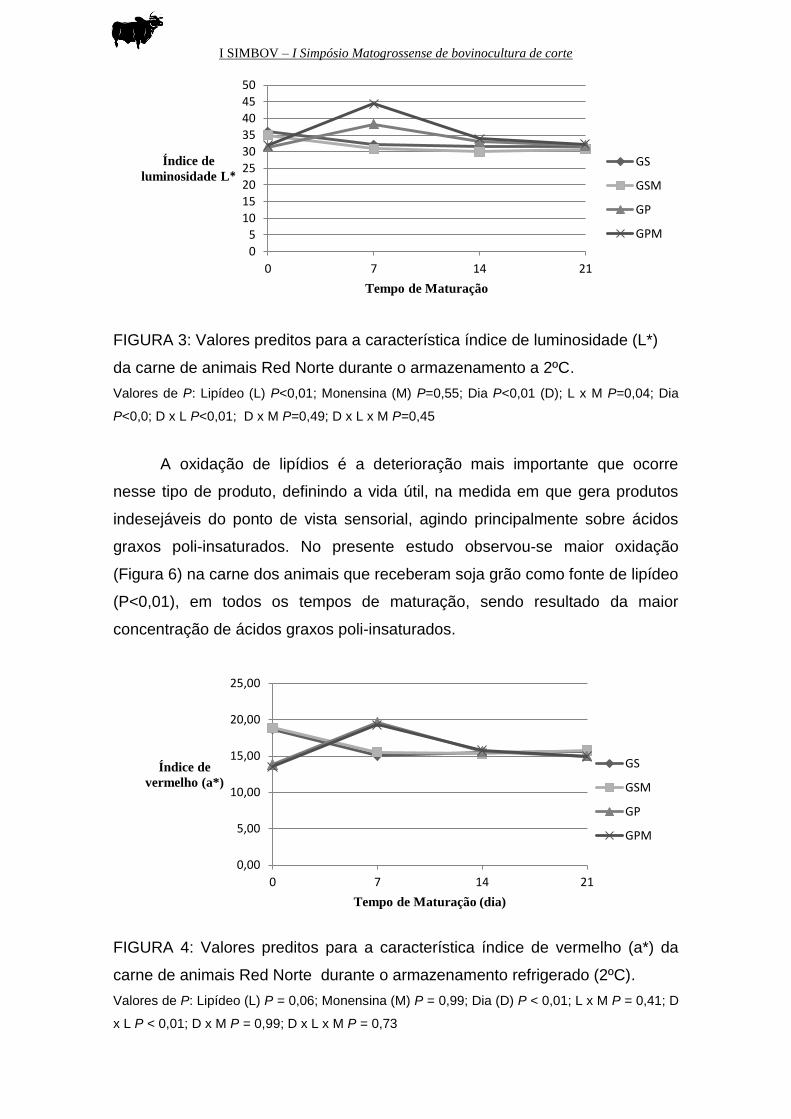
Os dados de coloração (Figuras 3, 4 e 5) são discutidos levando-se em
consideração os efeitos de fonte de lipídeo, pois a adição ou não do ionóforo
monensina, isoladamente, não influenciou tais características.
Ao longo do tempo de maturação (de 0 a 14 dias), os animais
alimentados com grão de soja apresentaram carne mais escura,
provavelmente, devido à uma maior oxidação da mioglobina. Como a carne
desses animais apresentaram maior concentração de ácidos graxos poli-
insaturados que são mais oxidativos, os radicais livres gerados durante a
oxidação lipídica podem ter oxidado o pigmento heme, bem como provocar a
desnaturação da parte protéica, levando a mudanças de cor indesejáveis.
Em relação ao índice a*, que representa a intensidade de vermelho da
carne (Figura 4), foi observado o mesmo comportamento de L*, ou seja, a
carne dos animais que consumiram grão de soja se apresentou mais vermelha
com 0 dia de maturação. No entanto, de 0 a 7 dias de maturação, esta carne se
apresentou com um vermelho menos intenso.
Músculo Probabilidade
Mirístico C14:0 2,02 2,36 2,55 2,27 0,1
8
0,23 0,89 0,09
Palmítico C16:0 20,9
2
23,6
3
23,7
3
22,6
8
1,0
2
0,35 0,40 0,06
Esteárico C18:0 16,4
0
14,8
7
12,4
2
13,0
6
0,6
3
<0,0
1
0,47 0,08
Oleico C18:1 c9 33,9
7
34,9
8
37,6
8
38,4
8
1,3
3
<0,0
1
0,48 0,93
CLA C18:2 c9-
t11
0,76 0,68 0,60 0,61 0,0
5
0,03 0,42 0,40
Linoleico C18:2c9-
c12
10,7
7
9,54 7,32 7,70 1,0
2
0,01 0,66 0,42
Linolênico C18:3 n3 0,31 0,32 0,49 0,47 0,0
2
<0,0
1
0,83 0,56
I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
FIGURA 3: Valores preditos para a característica índice de luminosidade (L*)
da carne de animais Red Norte durante o armazenamento a 2ºC.
Valores de P: Lipídeo (L) P<0,01; Monensina (M) P=0,55; Dia P<0,01 (D); L x M P=0,04; Dia
P<0,0; D x L P<0,01; D x M P=0,49; D x L x M P=0,45
A oxidação de lipídios é a deterioração mais importante que ocorre
nesse tipo de produto, definindo a vida útil, na medida em que gera produtos
indesejáveis do ponto de vista sensorial, agindo principalmente sobre ácidos
graxos poli-insaturados. No presente estudo observou-se maior oxidação
(Figura 6) na carne dos animais que receberam soja grão como fonte de lipídeo
(P<0,01), em todos os tempos de maturação, sendo resultado da maior
concentração de ácidos graxos poli-insaturados.
FIGURA 4: Valores preditos para a característica índice de vermelho (a*) da
carne de animais Red Norte durante o armazenamento refrigerado (2ºC).
Valores de P: Lipídeo (L) P = 0,06; Monensina (M) P = 0,99; Dia (D) P < 0,01; L x M P = 0,41; D
x L P < 0,01; D x M P = 0,99; D x L x M P = 0,73
0
5
10
15
20
25
30
35
40
45
50
0 7 14 21
Índice de
luminosidade L*
Tempo de Maturação
GS
GSM
GP
GPM
0,00
5,00
10,00
15,00
20,00
25,00
0 7 14 21
Índice de
vermelho (a*)
Tempo de Maturação (dia)
GS
GSM
GP
GPM
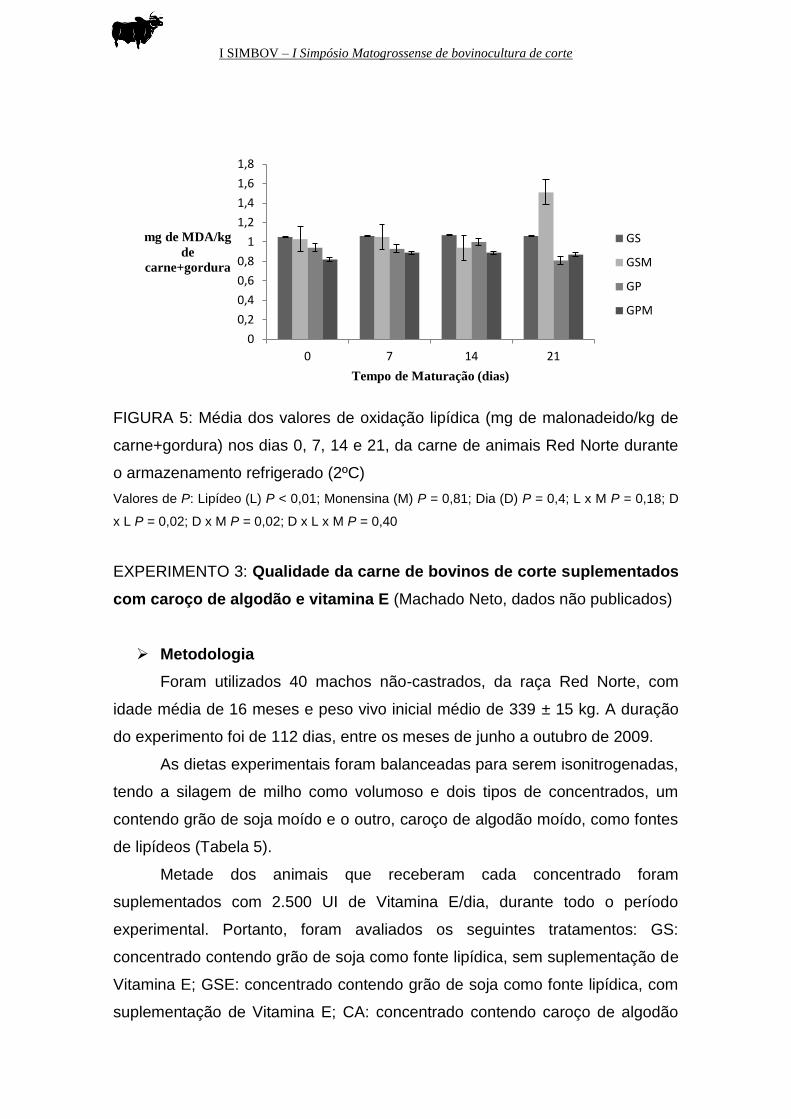
I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
FIGURA 5: Média dos valores de oxidação lipídica (mg de malonadeido/kg de
carne+gordura) nos dias 0, 7, 14 e 21, da carne de animais Red Norte durante
o armazenamento refrigerado (2ºC)
Valores de P: Lipídeo (L) P < 0,01; Monensina (M) P = 0,81; Dia (D) P = 0,4; L x M P = 0,18; D
x L P = 0,02; D x M P = 0,02; D x L x M P = 0,40
EXPERIMENTO 3: Qualidade da carne de bovinos de corte suplementados
com caroço de algodão e vitamina E (Machado Neto, dados não publicados)
Metodologia
Foram utilizados 40 machos não-castrados, da raça Red Norte, com
idade média de 16 meses e peso vivo inicial médio de 339 ± 15 kg. A duração
do experimento foi de 112 dias, entre os meses de junho a outubro de 2009.
As dietas experimentais foram balanceadas para serem isonitrogenadas,
tendo a silagem de milho como volumoso e dois tipos de concentrados, um
contendo grão de soja moído e o outro, caroço de algodão moído, como fontes
de lipídeos (Tabela 5).
Metade dos animais que receberam cada concentrado foram
suplementados com 2.500 UI de Vitamina E/dia, durante todo o período
experimental. Portanto, foram avaliados os seguintes tratamentos: GS:
concentrado contendo grão de soja como fonte lipídica, sem suplementação de
Vitamina E; GSE: concentrado contendo grão de soja como fonte lipídica, com
suplementação de Vitamina E; CA: concentrado contendo caroço de algodão
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
0 7 14 21
mg de MDA/kg
de
carne+gordura
Tempo de Maturação (dias)
GS
GSM
GP
GPM
I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
como fonte lipídica, sem suplementação de Vitamina E; CAE: concentrado
contendo caroço de algodão como fonte lipídica, com suplementação de
Vitamina E.
TABELA 5: Ingredientes e composição bromatológica das dietas experimentais
Ingredientes
Grão de Soja Caroço de Algodão
Silagem de milho 40,0% 40,0% Milho grão moído 38,2% 31,2% Grão de soja moído 20,0% - Farelo de soja - 3,00% Caroço de algodão - 24,0% Núcleo mineral1 1,80% 1,80%
Composição Química
PB 12,9% 12,7% EE 6,48% 6,56% FDN 27,0% 36,0% CNF1 46,0% 40,0% NDT1 82,0% 77,0% 1Níveis de garantia por kg do produto: Ca: 170 g; Co: 15 mg; Cu: 396 mg; P: 31 g; I: 29 mg; Mg:
15 g; Mn: 515 mg; Se: 5,4 mg; Na: 155 g; Zn: 2.000 mg. 1Calculado segundo o NRC (2001)
Resultados
Foi verificado que a gordura subcutânea dos animais alimentados com a
dieta com caroço de algodão apresentou maior concentração de C16:0 do que
aqueles alimentados com grão de soja, o que pode ser justificado pela maior
concentração deste ácido graxo no caroço de algodão. Resultado semelhante
foi observado para o C18:0, em que o maior teor de C18:0 ocorreu
provavelmente devido à inibição da ação da enzima Δ9 dessaturase pelo ácido
estercúlico presente no caroço de algodão. Como consequência ao aumento
da concentração destes ácidos graxos saturados, verificou-se redução da
concentração de ácidos graxos insaturados na gordura subcutânea, como o
C18:1 e o C18:3. Além disso, a suplementação da dieta com caroço de algodão
deprimiu a concentração de CLA (C18:2 cis-9 trans-11) na gordura subcutânea.
Os animais suplementados com caroço de algodão apresentaram
concentração superiores de ácido vaccênico, o que também se justifica pela
presença do inibidor da Δ9 dessaturase neste alimento.
TABELA 6: Médias e erros padrões dos valores das concentrações dos ácidos
graxos presentes na gordura subcutânea e no músculo de animais Red Norte
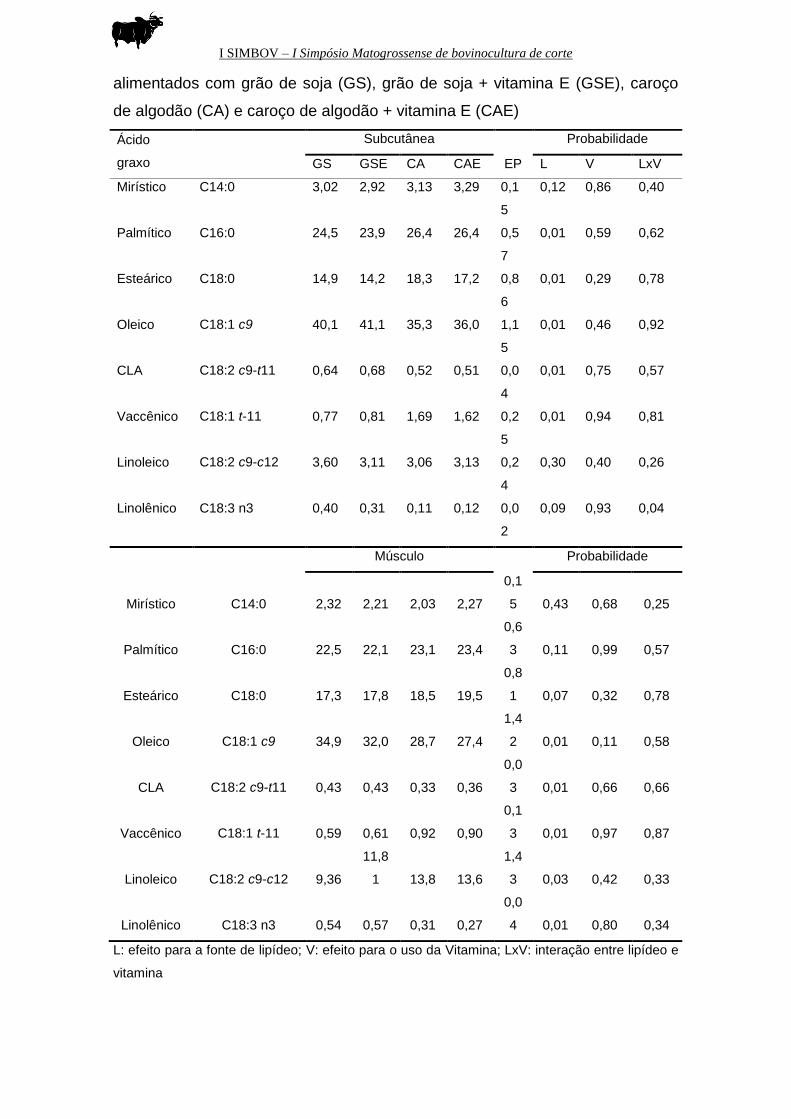
I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
alimentados com grão de soja (GS), grão de soja + vitamina E (GSE), caroço
de algodão (CA) e caroço de algodão + vitamina E (CAE)
L: efeito para a fonte de lipídeo; V: efeito para o uso da Vitamina; LxV: interação entre lipídeo e
vitamina
Ácido
graxo
Subcutânea
EP
Probabilidade
GS GSE CA CAE L V LxV
Mirístico C14:0 3,02 2,92 3,13 3,29 0,1
5
0,12 0,86 0,40
Palmítico C16:0 24,5 23,9 26,4 26,4 0,5
7
0,01 0,59 0,62
Esteárico C18:0 14,9 14,2 18,3 17,2 0,8
6
0,01 0,29 0,78
Oleico C18:1 c9 40,1 41,1 35,3 36,0 1,1
5
0,01 0,46 0,92
CLA C18:2 c9-t11 0,64 0,68 0,52 0,51 0,0
4
0,01 0,75 0,57
Vaccênico C18:1 t-11 0,77 0,81 1,69 1,62 0,2
5
0,01 0,94 0,81
Linoleico C18:2 c9-c12 3,60 3,11 3,06 3,13 0,2
4
0,30 0,40 0,26
Linolênico C18:3 n3 0,40 0,31 0,11 0,12 0,0
2
0,09 0,93 0,04
Músculo Probabilidade
Mirístico C14:0 2,32 2,21 2,03 2,27
0,1
5 0,43 0,68 0,25
Palmítico C16:0 22,5 22,1 23,1 23,4
0,6
3 0,11 0,99 0,57
Esteárico C18:0 17,3 17,8 18,5 19,5
0,8
1 0,07 0,32 0,78
Oleico C18:1 c9 34,9 32,0 28,7 27,4
1,4
2 0,01 0,11 0,58
CLA C18:2 c9-t11 0,43 0,43 0,33 0,36
0,0
3 0,01 0,66 0,66
Vaccênico C18:1 t-11 0,59 0,61 0,92 0,90
0,1
3 0,01 0,97 0,87
Linoleico C18:2 c9-c12 9,36
11,8
1 13,8 13,6
1,4
3 0,03 0,42 0,33
Linolênico C18:3 n3 0,54 0,57 0,31 0,27
0,0
4 0,01 0,80 0,34

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
Em relação ao perfil de ácidos graxos da gordura intramuscular, foi
verificada tendência de aumento da concentração de C16:0 (P=0,11) e também
do C18:0 (P=0,07). Animais alimentados com caroço de algodão apresentaram
menor concentração de C18:2 e CLA.
4- Gordura intramuscular
O conteúdo de gordura intramuscular da carne é um importante atributo
na qualidade. Estudos envolvendo consumidores e painéis treinados revelaram
que a gordura intramuscular é uma das mais importantes características que
influenciam alguns aspectos, como: maciez, suculência e sabor (Shackelford et
al., 1991).
Morfologicamente, a gordura intramuscular é o total de lipídios
associados com todas as células presentes na amostra de carne,
principalmente miócitos e adipócitos, excluindo os adipócitos depositados na
gordura intermuscular. Quimicamente, esses lipídios podem ser divididos em
fosfolipídios, triglicerídios, mono e diglicerídios, colesterol, ésteres de colesterol
e ácidos graxos livres, sendo as frações fosfolipídios e triglicerídios as mais
importantes para os estudos de gordura intramuscular (Gerbens, 2004).
Nos estados Unidos, com a crescente utilização de milho e outras
culturas para a produção de biocombustíveis, há a tendência de aumentar a
utilização de resíduos de grãos de destilaria nos confinamentos, o que segundo
Smith et al. (2008), aumentaria a dureza da carne devido à menor síntese de
ácidos graxos monoinsaturados no tecido adiposo, tornando a carne americana
menos competitiva em mercados específicos, como o japonês e o coreano.
No Brasil, a pecuária de corte se baseia principalmente na utilização de
pastagens ao longo do ciclo produtivo e o consumo de alimentos fibrosos,
como as pastagens, leva à absorção de determinados ácidos graxos ou outros
nutrientes que promovem a depressão da síntese da gordura intramuscular
(Smith et al., 2008) promovendo baixo grau de marmoreio na carne.
Os preços dos alimentos apresentam a maior parte do custo de
produção de animais em confinamento. Porém, na safra de grãos no Brasil, é
possível encontrar menores preços de grãos, como o milho. Este ingrediente é
importante quando se quer satisfazer a exigência de mercados que pagam por
qualidade, pois o tecido adiposo intramuscular parece ser sensível à utilização
de grãos, principalmente em animais jovens em terminação.

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
4.1- Síntese de tecido adiposo intramuscular
De acordo com Gilbert et al. (2003), o tecido adiposo intramuscular
usa alta proporção de glicose para a síntese de ácidos graxos, diferente do
tecido adiposo subcutâneo que utiliza acetato para a deposição de lipídios.
Segundo os mesmos autores, aproximadamente 61 a 76% do milho moído
consumido é degradado no rúmen, onde é convertido em ácidos graxos
voláteis, e o restante sofre digestão pós-ruminal, principalmente no intestino
delgado, o que resulta em glicose livre para a absorção. Sendo assim, a maior
passagem de amido para o intestino delgado é uma alternativa para tentar
aumentar o grau de marmoreio no tecido intramuscular (Miller et al., 1991).
A insulina estimula a captação de glicose pelos tecidos periféricos
aumentando a lipogênese ou reduzindo a lipólise, sendo as concentrações
plasmáticas de insulina positivamente correlacionadas com a adiposidade na
carcaça do animal. A variação na sensibilidade à insulina pode afetar a partição
energética entre os tecidos e entre tecidos em desenvolvimento (Rhoades et
al., 2007).
Gilbert et al. (2003) relataram que o tecido adiposo intramuscular é
mais sensível à insulina, quando comparado ao tecido adiposo subcutâneo.
Sendo assim, os autores sugeriram que alimentos que aumentam a produção
de propionato, como o milho e outros grãos, têm maior capacidade glicogênica
e insulinogênica, o que poderia aumentar a deposição de gordura
intramuscular.
Durante muitos anos foi demonstrado que a glicose contribuía com
maior proporção de unidades de acetil para a síntese de novo de ácidos graxos
no tecido adiposo intramuscular, quando comparada ao tecido adiposo
subcutâneo, sendo responsável por aproximadamente 70% do fornecimento de
acetil necessários para a síntese, enquanto que o acetato e o lactato
contribuíam com apenas 20% (Smith & Crouse, 1984).
Em um trabalho realizado por Chung et al. (2007), um grupo de
novilhos foi alimentado com uma dieta de terminação baseada em milho moído
(48%) para atingir uma taxa de crescimento de 1,36 kg/dia durante oito meses
(alimentação curta, baseada em milho), ou foram alimentados com feno e
suplementados para obtenção de ganhos de 0,9 kg/dia durante 12 meses
(alimentação curta, baseada em feno). Nestas duas dietas objetivou-se um
I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
peso de abate de aproximadamente 525 kg. Um segundo conjunto de animais
foi alimentado com as mesmas dietas baseadas em milho e feno durante 16 e
20 meses respectivamente objetivando um abate com peso de
aproximadamente 650 Kg.
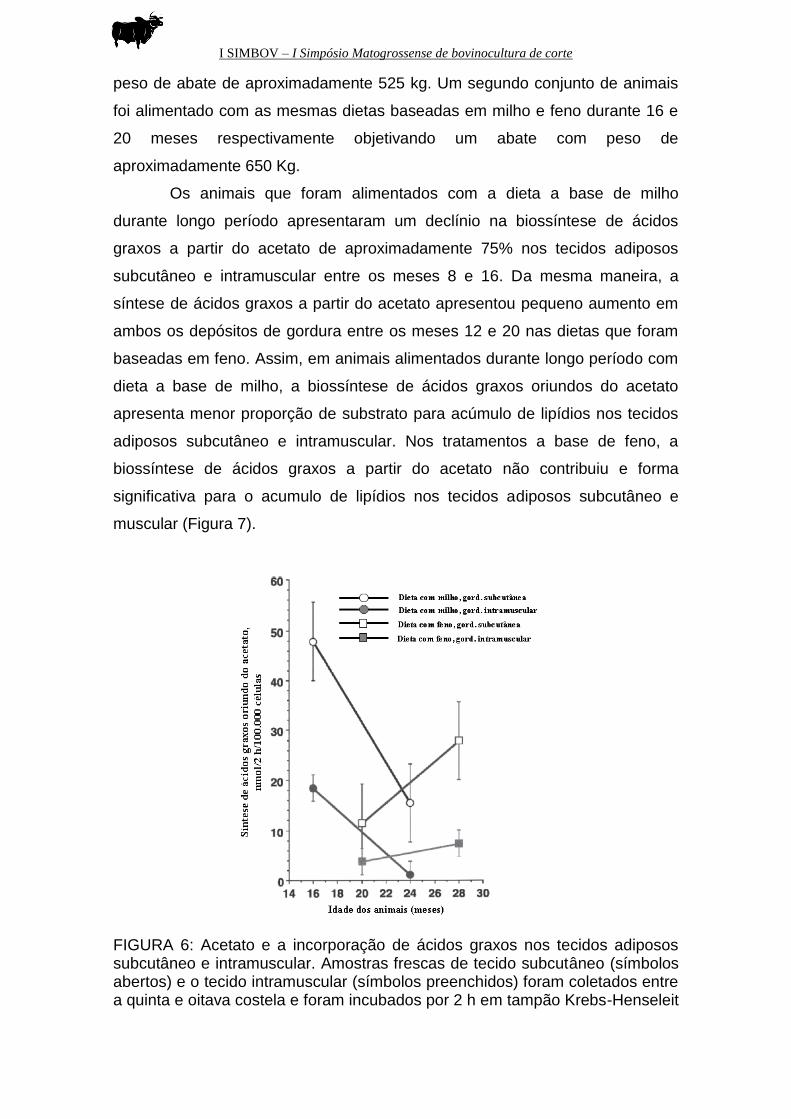
Os animais que foram alimentados com a dieta a base de milho
durante longo período apresentaram um declínio na biossíntese de ácidos
graxos a partir do acetato de aproximadamente 75% nos tecidos adiposos
subcutâneo e intramuscular entre os meses 8 e 16. Da mesma maneira, a
síntese de ácidos graxos a partir do acetato apresentou pequeno aumento em
ambos os depósitos de gordura entre os meses 12 e 20 nas dietas que foram
baseadas em feno. Assim, em animais alimentados durante longo período com
dieta a base de milho, a biossíntese de ácidos graxos oriundos do acetato
apresenta menor proporção de substrato para acúmulo de lipídios nos tecidos
adiposos subcutâneo e intramuscular. Nos tratamentos a base de feno, a
biossíntese de ácidos graxos a partir do acetato não contribuiu e forma
significativa para o acumulo de lipídios nos tecidos adiposos subcutâneo e
muscular (Figura 7).
FIGURA 6: Acetato e a incorporação de ácidos graxos nos tecidos adiposos subcutâneo e intramuscular. Amostras frescas de tecido subcutâneo (símbolos abertos) e o tecido intramuscular (símbolos preenchidos) foram coletados entre a quinta e oitava costela e foram incubados por 2 h em tampão Krebs-Henseleit
I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
contendo 10nM de tampão HEPES (pH 7,4), 10 mM de acetato, 10mM de glicose e acetato U-14C (Adaptado de Chung et al., 2007).
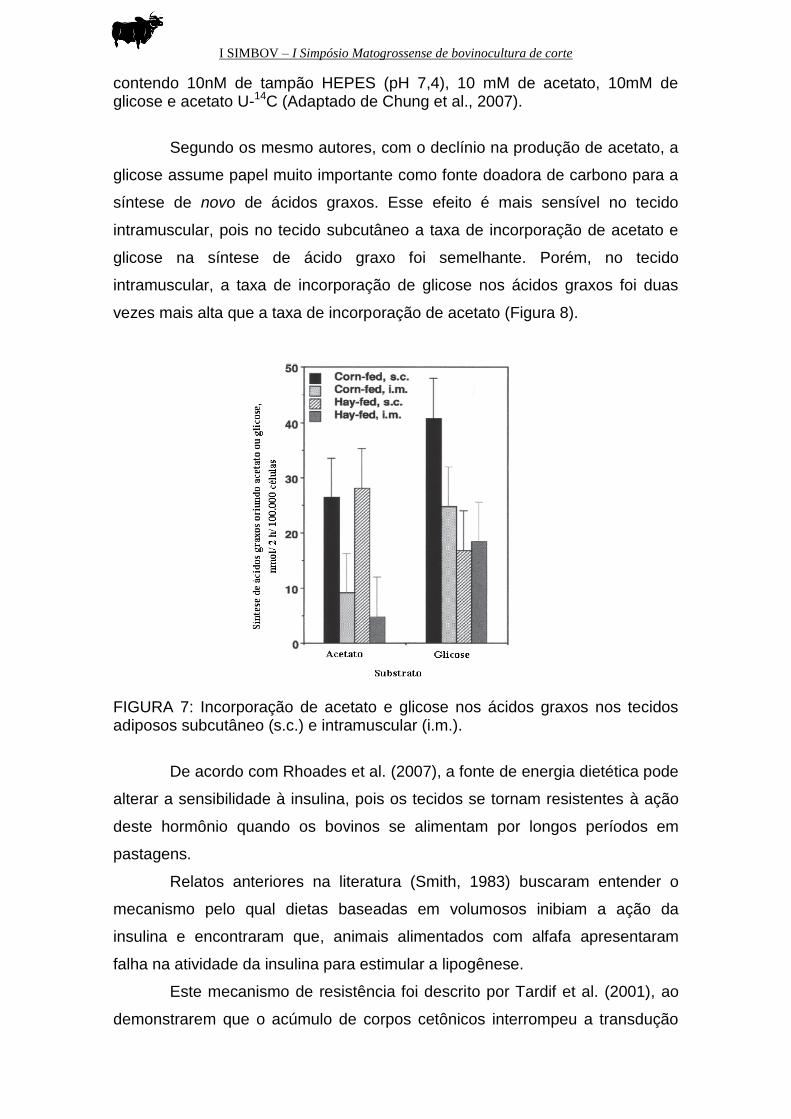
Segundo os mesmo autores, com o declínio na produção de acetato, a
glicose assume papel muito importante como fonte doadora de carbono para a
síntese de novo de ácidos graxos. Esse efeito é mais sensível no tecido
intramuscular, pois no tecido subcutâneo a taxa de incorporação de acetato e
glicose na síntese de ácido graxo foi semelhante. Porém, no tecido
intramuscular, a taxa de incorporação de glicose nos ácidos graxos foi duas
vezes mais alta que a taxa de incorporação de acetato (Figura 8).
FIGURA 7: Incorporação de acetato e glicose nos ácidos graxos nos tecidos adiposos subcutâneo (s.c.) e intramuscular (i.m.).
De acordo com Rhoades et al. (2007), a fonte de energia dietética pode
alterar a sensibilidade à insulina, pois os tecidos se tornam resistentes à ação
deste hormônio quando os bovinos se alimentam por longos períodos em
pastagens.
Relatos anteriores na literatura (Smith, 1983) buscaram entender o
mecanismo pelo qual dietas baseadas em volumosos inibiam a ação da
insulina e encontraram que, animais alimentados com alfafa apresentaram
falha na atividade da insulina para estimular a lipogênese.
Este mecanismo de resistência foi descrito por Tardif et al. (2001), ao
demonstrarem que o acúmulo de corpos cetônicos interrompeu a transdução

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
do sinal da insulina e reduziu sua migração para a superfície da célula. Esta
redução no transportador reduziria a entrada de glicose estimulada pela
insulina, e consequentemente limitaria a taxa de utilização de glicose. Para
Herdt et al. (1981), os corpos cetônicos podem acumular-se como acetato,
principalmente quando a glicose está limitada, e as maiores concentrações de
acetato quando os animais estão sob regime de pastejo podem afetar mais a
resistência de utilização da glicose.
Schoonmaker et al. (2003) encontraram maiores concentrações de
insulina em novilhos alimentados com dietas de alto concentrado em relação
aos novilhos alimentos com dietas de alta forragem. Os autores relataram que
o músculo Longissimus dorsi e o tecido adiposo de animais alimentados com
alto concentrado, não somente apresentaram maiores concentrações de
insulina circulante, mas também estavam mais sensíveis aos seus efeitos na
absorção de glicose e subsequente utilização.
Devido à aparente menor utilização de acetato pelo tecido adiposo
subcutâneo como substrato para a síntese de novo de ácidos graxos, e a
semelhança entre a utilização de glicose pelos tecidos intramuscular e
subcutâneo para a incorporação de lipídios, pode-se inferir que a limitação no
fornecimento ou absorção teria mais efeito na taxa de lipogênese no tecido
adiposo intramuscular do que no tecido adiposo subcutâneo. Choat et al.
(2003) encontraram aumento na deposição de tecido adiposo intramuscular em
novilhos alimentados com dietas de alto concentrado, que geraram 39,3% a
mais de propionato, quando comparado aos novilhos que alimentaram dietas
volumosas.
Dessa forma, além das raças utilizadas na bovinocultura de corte,
estratégias de alimentação devem ser levadas em conta, como a
suplementação animal nas distintas fases do ciclo de produção, principalmente
quando se objetiva produzir animais com qualidade superior, pois mercados
exigentes como o americano e o japonês além de uma carne bem marmorizada
(padrão choice), exigem também animais com elevada espessura de gordura
subcutânea (12 mm).

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
Considerações finais
Ficou demonstrado no presente artigo que a nutrição influencia
diretamente a qualidade da carne, tornando-se portanto, uma ferramenta
importante para agregar valor ao produto.

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
Referências bibliográficas
BAUMAN, D. E.; BAUMGARD, L. H.; CORL, B. A. Biosynthesis of conjugated linoleic acid in ruminants. In: AMERICAN SOCIETY OF ANIMAL SCIENCE, 1999, Ithaca. Proceedings… Ithaca: Cornel University, 1999. p. 1-15.
BEAM, T. M., JENKINS, T.C.;. MOATE, P.J.. et al. 2000. Effects of amount and source of fat on the rates of lipolysis and biohydrogenation of fatty acids in ruminal contents. Journal of Dairy Science v. 83 p.2564–2573.
BELURY, M. A. 2002. Dietary conjugated linoleic acid in health: Physiological effects and mechanisms of action. Annual Review Nutrition v. 22. p. 505–531
CHOAT, W.T.; KREHBIEL, C.R.; DUFF, G.C. et al. Influence of grazing dormant native range or winter wheat pasture on subsequent finishing cattle performance, carcass characteristics, and ruminal metabolism. Journal of Animal Science, n.81, 3191-3201, 2003.
CHUNG, K.Y.; LUNT, D.K.; KAWACHI, H. et al. Lipogenesis and steroyl-Coa desaturase gene expression and enzyme activity in adipose tissue of short- and long-fed Angus and Wagyu steers fed corn- or hay-based diets. Journal of Animal Science, n.85, p.380-387, 2007.
DAWSON, R. M. C.; KEMP, P. The effect of defaunation on the phospholipids and on the hydrogenation of unsaturated fatty acids in the rumen. Biochemestry Journal v.115 p.351–352, 1969.
DOREAU, M.; CHILLIARD, Y. Digestion and metabolism of dietary fat in farm animals. British Journal of Nutrition. v.78 p.15–35, 1997
FERLAY, A., J. CHABROT, Y. ELMEDDAH, AND M. DOREAU. Ruminal lipid balance and intestinal digestion by dairy cows fed calcium salts of rapeseed oil fatty acids or rapeseed oil. Journal of Animal Science. v.71, p. 2237, 1992.
GERBENS, F. Genetic control of intramuscular fat accretion. In: PAS, M.F.W.; EVERTS, M.E.; HAAGSMAN, H.P. (Eds.) Muscle Development of Livestock Production – Physiology, Genetics and Meat Quality, CABI Publishing, Oxfordshire, UK, pp.342-361.
GILBERT, C.D.; LUNT, D.K.; MILLER, R.K.; SMITH, S.B. Carcass, sensory, and adipose tissue traits of brangus steers fed casein-formaldehyde-protected starch and/or canola lipid. Journal of Animal Science, n.81, p.2457-2468, 2003.
HERDT, T.H.; STEVENS, J.B.; LINN, J.; LARSON, V. Influence of ration composition and energy balance on blood β-hydroxybutyrat (ketone) and plasma glucose concentration of dairy cows in early lactation. American Journal of Veterinary Research, n. 42. P.1177-1180, 1981.
HESS, B.W.; MOSS, G.E.; RULE, D.C. A decade of developments in the area of fat supplementation research with beef cattle and sheep. Journal of Animal Science. 2007.
HOOD, D.E. Factors affecting the rate of metmyoglobin accumulation in pre-packaged beef. Meat Science, Barking, v. 4, n. 4, p. 247-265, Aug. 1980.
JENKINS, T.C. Lipid metabolism in the rumen. Journal of Dairy Science v. 76, p.3851–3863, 1993.
JENKINS, T.C. Regulation of lipid metabolism in the rumen. Journal Nutrition v. 124, p.1372S–1376, 1994.

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
JENKINS, T.C.; A.A. ABUGHAZALEH; S. FREEMAN; E.J. THIES The production of 10-hydroxystearic acid and 10-ketostearic acids is an alternate route of oleic acid transformation by the ruminal microbiota in cattle. Journal Nutrition v. 136, p.926-931, 2006.
JENKINS, T.C.; ADAMS, C.S. The biohydrogenation of linoleamide in vitro and its effects on linoleic acid concentration in duodenal contents of sheep. Journal of Animal Science. v. 80, p.533–540, 2002.
KEMP, P.; R.W. WHITE; D.J. LANDER. The hydrogenation of unsaturated fatty acids by five bacterial isolates from the sheep rumen, including a new species. Journal of Genetics and Microbiology. v. 90, p.100–114, 1975.
KRYSL, L.J.; M.B. JUDKINS; V.R. BOHMAN. Influence of ruminal or duodenal soybean oil infusion on intake, ruminal fermentation, site and extent of digestion, and microbial protein synthesis in beef heifers consuming grass hay. Journal of Animal Science. v.69, p.2585–2590, 1991.
KEPLER, C.R.; K.P. HIRONS; J.J. MCNEILL; S.B. TOVE Intermediates and products of the biohydrogenation of linoleic acid by Butyrivibrio fibrisolvens. Journal of Biology Chemestry. v.241, p.1350–1354, 1966.
LEE, K.T.; YOON, C.S. Quality changes and shelf-life of imported vacuum-packaged beef chuck during storage at 0ºC. Meat Science, Barking, v. 59, n. 1, p. 71-77, Sept. 2001.
MAIA, M.R.G.; L.C. CHAUDHARY; L. FIGUERES; R.J. WALLACE. Metabolism of polyunsaturated fatty acids and their toxicity to the microflora of the rumen. Antonie Leeuwenhoek v. 91 p.303–314, 2006.
MERCHEN, N.R.; J.C. ELIZALDE; J.K. DRACKLEY Current perspective on assessing site of digestion in ruminants. Journal of Animal Science. v.75, p.2223–2234, 1997.
MILLER, M.F.; CROSS, H.R.; LUNT, D.K.; SMITH, S.B. Lipogenesis in acute and 48-hour cultures of bovine intramuscular and subcutaneous bovine adipose tissue explants. Journal of Animal Science, n.69, p.162-170.
MONTGOMERY, S.P.; DROUILLARD, J.S.; NAGARAJA, T.G.; TITGEMEYER, E.C.; SINDT, J.J. Effects of supplemental fat source on nutrient digestion and ruminal fermentation in steers. Journal of Animal Science v. 86. p.640-650. 2008.
NAGARAJA, T.G.; C.J. NEWBOLD; C.J. VEN NEVEL; D.I. DEMEYER Manipulation of ruminal fermentation. Pages 523–632 in The Rumen Microbial Ecosystem, P. N. Hubson and C. S. Stewart, ed. Blackie Acad. and Prof., an imprint of Chapman and Hall, London, UK, 1997.
PALMQUIST, D. L.; A.L. LOCK; K.J. SHINGFIELD; D.E. BAUMAN Biosynthesis of conjugated linoleic acid in ruminants and humans. Advances in Food Nutrition Research. v. 50, p.179–217, 2005.
PALMQUIST, D.L.; T.C. JENKINS Fat in lactation rations. Journal of Dairy Science. v.63. p.1–14, 1980.
PARIZA, M.W. Perspective on the safety and effectiveness of conjugated linoleic acid. American Journal of Clinical Nutrition. v.79, p.1132–1136, 2004.

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
POLAN, C. E.; J.J. MCNEILL; S.B. TOVE Biohydrogenation of unsaturated fatty acids by rumen bacteria. Journal of Bacteriology. v.88, p.1056–1064, 1964.
RHOADES, R.D.; SAWYER, J.E.; CHUNG, K.Y.; SCHELL, M.L.; LUNT, D.K.; SMITH, S.B. Effect of dietary energy on in vitro substrate utilization and insulin sensitivity of muscle and adipose tissue of Angus and Wagyu steers. Journal of Animal Science, n.85, 1719-1726, 2007.
SCHOONMAKER, J.P.; CECAVA, M.J.; FAULKNER, D.B.; FLUHARTY, F.L.; ZERBY, H.N.; LOERCH, S.C. Effects of source of energy and rate of growth on performance, carcass characteristics, ruminal fermentation, and serum glucose and insulin of early-weaned steers. Journal of Animal Science, n.81, p.843-855, 2003.
SHACKELFORD, S.D.; KOOHMARAIE, M.; MILLER, M.F.; CROUSE, J.D.; REAGAN, J.O. An evaluation of tenderness of the longissimus muscle of Angus by Hereford versus Brahman crossbred heifers. Journal of Animal Science, n.69, p.171-177, 1991.
SMITH, S.B.; KAWACHI, H.; CHOI, C.B.; CHOI, C.W.; WU, G.; SAWYER, J.E. Cellular regulation of bovine intramuscular adipose tissue development and composition. Journal of Animal Science, n.87, p.72-82, 2007.
SMITH, S.B.; CROUSE, J.D. Relative contributions of acetate, lactate and glucose to lipogenesis in bovine intramuscular and subcutaneous adipose tissue. Journal of Nutrition, n.114, p.792-800, 1984.
SMITH, S.B. Contribution of pentose cycle to lipogenesis in bovine adipose tissue. Archives of Biochemistry and Biophysics, n.221, p.46-56, 1983.
SHORLAND, F. B.; R.O. WEENINK; H. GOLDFINE Effect of the rumen on dietary fat. Nature. v.175, p.1129–1130, 1955.
VAN DE VOSSENBERG, J. L.; K.N. JOBLIN. Biohydrogenation of C18 unsaturated fatty acids to stearic acid by a strain of Butyrivibrio hungatei from the bovine rumen. Letters in Applied Microbiology. v.37, p.424–428, 2003.
VAN NEVEL, C.J.; DEMEYER, D.I. Influence of pH on lipolysis and biohydrogenation of soybean oil by rumen contents in vitro. Animal Reproduction. v.36, p.53–63, 1996.
TARDIF, A.; JULIEN, N.; PELLETEIR, A.; THIBAULT, G.; SRIVASTAVA, A.; CHIASSON, J.L.; CODERRE, L. Chronic exposure to β-hydroxybutyrate impairs insulin action in primary cultures of adult cardiomyocytes. American Journal of Physiology, n.28, p.1205-1212, 2001.
WALLACE, R.J.; L.C. CHAUDHARY; N. MCKAIN et al. Clostridium proteoclasticum: A ruminal bacterium that forms stearic acid from linoleic acid. FEMS Microbiol. Lett. 265:195 201, 2006.
WARD, P.F.V.; T.W. SCOTT; R.M.C. DAWSON The hydrogenation of unsaturated fatty acids in the ovine digestive tract. Biochemestry Journal, v.92, p.60–68, 1964.
WHIGHAM, L.D.; M.E. COOK; R.L. ATKINSON Conjugated linoleic acid: Implications for human health. Pharmacology Research, v.42, p.503–510, 2000.

I SIMBOV – I Simpósio Matogrossense de bovinocultura de corte
WILLIAMS, A.G.; COLEMAN, A.G. The Rumen Protozoa. Springer-Verlag, New York, NY. 1992.
WILSON, C.A.; WOOD, T.M. The anaerobic fungus Neocallimastix frontalis: Isolation and properties of a cellulosome-type enzyme fraction with the capacity to solubilise hydrogen-bondordered cellulose. Applied Microbiology Biotechnology, v.37, p.125–129, 1992.
WOOD, R.D.; M.C. BELL; R.B. GRAINGER; R.A. Teekel Metabolism of labeled linoleic-1-14C acid in the sheep rumen. Journal of Nutrition, v.79, p.62–68, 1963.
ZINN, R.A.; S.K. GULATI; A. PLASCENCIA; J. SALINAS Influence of ruminal biohydrogenation on the feeding value of fat in finishing diets for feedlot cattle. Journal of Animal Science, v.78, p.1738–1746, 2000.


















