
DESEMPENHO GERMINATIVO DE SEMENTES DE CINCO …
114
UNIVERSIDADE FEDERAL DE MATO GROSSO FACULDADE DE AGRONOMIA E MEDICINA VETERINÁRIA Programa de Pós-graduação em Agricultura Tropical DESEMPENHO GERMINATIVO DE SEMENTES DE CINCO ESPÉCIES NATIVAS DOS ECOSSISTEMAS DE MATO GROSSO ROSELI MUNIZ GIACHINI CUIABÁ – MT 2009
Transcript of DESEMPENHO GERMINATIVO DE SEMENTES DE CINCO …

1
UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE AGRONOMIA E MEDICINA VETERINÁRIA
Programa de Pós-graduação em Agricultura Tropical
DESEMPENHO GERMINATIVO DE SEMENTES DE CINCO
ESPÉCIES NATIVAS DOS ECOSSISTEMAS DE MATO
GROSSO
ROSELI MUNIZ GIACHINI
CUIABÁ – MT
2009

2
UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE AGRONOMIA E MEDICINA VETERINÁRIA
Programa de Pós-graduação em Agricultura Tropical
DESEMPENHO GERMINATIVO DE SEMENTES DE CINCO
ESPÉCIES NATIVAS DOS ECOSSISTEMAS DE MATO
GROSSO
ROSELI MUNIZ GIACHINI
Engenheira Agrônoma
Orientador: Prof. Dr. FRANCISCO DE ALMEIDA LOBO
Co-Orientadora: Profª. Drª. CARMEN EUGENIA R. ORTÍZ
Co-Orientadora: Profª. Drª. MARIA CRISTINA DE F.E ALBUQUERQUE
Dissertação apresentada à Faculdade de Agronomia e Medicina Veterinária da Universidade Federal de Mato Grosso, para obtenção do título de Mestre em Agricultura Tropical
CUIABÁ - MT
2009

3
Aos meus pais
Ladir Lau e Maria Leonor
Ao meu esposo
Vilmar
Aos meus filhos
Fernando e Fabíola
OFEREÇO
Ao Prof. Francisco de Almeida Lobo
À Profª. Maria Cristina de Figueiredo e Albuquerque
DEDICO

4
AGRADECIMENTOS
À Deus pela vida, saúde, família, proteção e pela oportunidade de tornar-me
mestre.
À Universidade Federal de Mato Grosso, pela oportunidade de realização
do curso.
À Capes pelo apoio financeiro.
Ao Prof. Dr. Francisco de Almeida Lobo, pela orientação, confiança e
amizade.
À Profa. Dra. Maria Cristina de Figueiredo e Albuquerque e à Profa. Dra.
Carmen Eugenia Rodriguez Ortíz, pelas sugestões para a melhoria deste
documento.
Aos amigos que fiz durante o curso que muito me ajudaram e que fazem
parte da minha vida: Diele, Islayne, Everton, Ramon, Lucas e Fábio.
Aos amigos Castrilon e Solange pela ajuda na coleta de dados nos dias de
feriado e nas altas horas da madrugada.
Aos amigos e colegas de pesquisa: Marcela, Renata, Amanda, Didi,
Carmen, Eliane, Elizandra, Leo, Jose, Jose Dida, Rene, Juliana, JU bióloga,
Carla, Profª. Patrícia, Prof. Sebastião, Prof. Sidnei, Nanda, João, Hugão,
Jaizon, Daniel, Carol, Ronncky, Alexandra, Dirce e Evaldo pela ajuda,
sugestões e troca de experiências.
A todos os professores, alunos, técnicos, mestrandos e doutorandos do
Programa de Pós-Graduação em Agricultura Tropical, pelos momentos de
suporte, críticas e descontração.
Às secretarias da Pós-graduação Denise e Maria pela amizade e
profissionalismo.
À amiga de toda hora, Sidnéia.
Aos alunos da Agronomia com os quais trocamos experiências e muito
aprendi.

5
DESEMPENHO GERMINATIVO DE SEMENTES DE CINCO ESPÉCIES
NATIVAS DOS ECOSSISTEMAS DE MATO GROSSO
RESUMO – Este trabalho foi conduzido com cinco espécies florestais:
Anadenanthera macrocarpa (angico vermelho), Amburana cearensis
(cerejeira), Samanea tubulosa (sete-cascas), Bowdichia virgilioides
(sucupira preta) e Buchenavia tomentosa (tarumarana), árvores típicas dos
biomas brasileiros dos Cerrados, Pantanal e Floresta Amazônica, com o
objetivo de avaliar o requerimento térmico para a germinação de sementes
e a formação plântulas. Foram utilizadas temperaturas de 10, 15, 20, 25, 30,
35 e 40ºC. A velocidade e o tempo para germinação e formação de
plântulas variaram entre as espécies e entre as temperaturas. A estimativa
das temperaturas cardinais obtidas pelo índice de velocidade de
germinação possibilitou identificar temperaturas máxima, mínima e ótima de
48,9°C; 6,2°C e 29,0°C para a cerejeira, de 42,4°C; 7,1°C e 33,2°C para a
sucupira preta e de 40,1°C; 10,4°C e 27,3°C para a tarumarana. A
estimativa das temperaturas cardinais obtidas pelo índice de velocidade de
formação de plântulas, por sua vez, permitiu identificar temperaturas
máxima, mínima e ótima de 43,4°C; 13,4°C e 33,1°C para o angico
vermelho, de 40,0°C; 12,0°C e 32,2°C para a cerejeira, de 40,0°C; 1,3°C e
34,0°C para a sete-cascas, de 40,0°C; 10,5°C e 33,0°C para a sucupira
preta e de 39,3°C; 14,5°C e 27,6°C para a tarumarana.
Palavras-chave: temperaturas cardinais, sementes florestais, áreas
degradadas.

6
SEED GERMINATION PERFORMANCE OF FIVE NATIVE SPECIES OF
MATO GROSSO´S ECOSSYSTEMS
ABSTRACT – This work was carried out with five species: Anadenanthera
macrocarpa (angico vermelho), Amburana cearensis (cerejeira), Samanea
tubulosa (sete-cascas), Bowdichia virgilioides (sucupira preta) and
Buchenavia tomentosa (tarumarana), typical trees species of Brazilian´s
savannah, woodland, and amazon forest, in order to evaluate the thermal
needs for seed germination and seedling formation. It was used
temperatures of 10.0, 15.0, 20.0, 25.0, 30.0, 35.0 and 40.0ºC. The speed
and the spent time for germination and seedling formation varied between
species and between temperatures. Estimates of cardinal temperatures
obtained by using germination speed made possible to identify maximum,
minimum and optimum temperature of 48.9°C; 6.2°C and 29.0°C for
cerejeira, 42.4°C; 7.1°C and 33.2°C for sucupira preta and 40.1°C; 10.4°C
and 27.3°C for tarumarana. Estimates of cardinal temperatures obtained by
using seedling formation speed made possible to identify maximum,
minimum and optimum temperature of 43.4°C; 13.4°C and 33.1°C for angico
vermelho, 40.0°C, 12.°C and 32.2°C for cerejeira, 40.0°C, 1.3°C and 34.0°C
for sete-cascas, 40.0°C, 10.5°C and 33.0°C for sucupira preta, and 39.3°C,
14.5°C and 27.6°C for tarumarana.
Keywords: cardinal temperatures, forest seeds, degraded areas.

7
LISTA DE FIGURAS
Página 1 Mapa de biomas do Brasil. ....................................................................
17
2 Processo de degradação progressiva no bioma Amazônico. ...............
21
3 Aspecto da planta com frutos de Anadenanthera macrocarpa, Cuiabá MT...........................................................................................................
25
4 Frutos (A) e sementes (B) de Anadenanthera macrocarpa (angico vermelho)...............................................................................................
25
5 Árvore de Amburana cearensis..............................................................
27
6 Frutos (A) e sementes (B) de cerejeira (Amburana cearensis)..................................................................................................
27
7 Aspecto da planta com flores de Samanea tubulosa.............................
28
8 Frutos (A) e sementes (B) de Samanea tubulosa.....................................................................................................
29
9 Árvores de Bowdichia virgilioides no campus da Faculdade de Agronomia e Medicina Veterinária – Cuiabá- MT. ..............................
30
10 Flores e frutos (A) e sementes (B), de Bowdichia virgilioides (sucupira preta....................................................................................
30
11 Árvore de Buchenavia tomentosa Eichler – tarumarana em Santo Antonio do Leverger- MT, Fazenda Experimental da Faculdade de Agronomia e Medicina Veterinária. .....................................................
31
12 Frutos (A) e sementes (B) de tarumarana (Buchenavia tomentosa)....
32
13 Germinação acumulada (%) de sementes de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à temperatura de 10°C. ...................................................
53
14 Germinação acumulada (%) de sementes de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à temperatura de 15°C. ...................................................
54
15 Germinação acumulada (%) de sementes de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana E), submetidas à temperatura de 20°C.....................................................
55

8
16 Germinação acumulada (%) de sementes de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à temperatura de 25°C.....................................................
56
17 Germinação acumulada (%) de sementes de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à temperatura de 30°C.....................................................
57
18 Germinação acumulada (%) de sementes de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à temperatura de 35°C.....................................................
58
19 Germinação acumulada (%) de sementes de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à temperatura de 40°C....................................................
59
20 Porcentagem acumulada da formação de plântulas de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à temperatura de 10°C..........................
60
21 Porcentagem acumulada da formação de plântulas de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à temperatura de 15°C..........................
61
22 Porcentagem acumulada da formação de plântulas de angico vermelho A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à temperatura de 20°C..........................
62
23 Porcentagem acumulada da formação de plântulas de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à temperatura de 25°C...........................
63
24 Porcentagem acumulada da formação de plântulas de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à temperatura de 30°C...........................
64
25 Porcentagem acumulada da formação de plântulas de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à temperatura de 35°C...........................
65
26 Porcentagem acumulada da formação de plântulas de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à temperatura de 40°C...........................
66
27 Plântulas de angico vermelho (Anadenanthera macrocarpa) formadas nas temperaturas de 10, 15, 20, 25, 30, 35 e 40°C............
68
28 Plântulas de cerejeira de oriundas sementes submetidas às

9
temperaturas de 20°C (A) e 40°C (B). ................................................
70
29 Plântulas de cerejeira formadas na temperatura de 25 e 30°C. .......
71
30 Plântulas de sete-cascas formadas a temperatura de 10°C. ..............
72
31 Plântulas de sete-cascas formadas na temperatura de 15°C. ............
73
32 Plântulas de sete-cascas formadas temperatura de 25°C, 30°C e 40°C. ..................................................................................................
73
33 Plântulas de sete-cascas formadas a 40°C. ........................................
74
34 Plântulas de sucupira preta (Bowdichia virgilioides) formadas a15°C (A) 20, 25 e 30°C (B) e semeadas em 9/12/07 e avaliadas em 04/01/08...............................................................................................
76
35 Plântulas formadas na temperatura de 15, 20 e 30°C aos 28 dias de semeadura. .........................................................................................
78
36. Plântulas de tarumarana formadas nas temperaturas de 25, 30 e 35°C. ...................................................................................................
78
37 Índice de velocidade de germinação de sementes de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à faixa de temperatura de 10 a 40°C. ....
81
38 Índice de velocidade para formação de plântulas de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à faixa de temperatura de 10 a 40°C. .......................
82
39 Tempo necessário para a obtenção de 50% de germinação de sementes de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à faixa de temperatura de 10 a 40°C....................................................................
84
40 Tempo necessário para a obtenção de 50% para formação de plântulas de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), submetidas à faixa de temperatura de 10 a 40°C.
85
41 Regressões empregadas nas estimativas das temperaturas cardinais do processo de germinação de sementes de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), empregando-se o índice de velocidade de germinação para expressar a taxa desse processo..........................
88
42 Regressões empregadas nas estimativas das temperaturas

10
cardinais do processo de formação de plântulas de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), empregando-se o índice de velocidade de germinação para expressar a taxa desse processo. ........................
90
43 Regressões empregadas nas estimativas das temperaturas cardinais do processo de germinação de sementes de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), empregando-se o inverso do tempo para alcançar 50% de germinação para expressar a taxa desse processo. ...........
93
44 Regressões empregadas nas estimativas das temperaturas cardinais do processo de formação de plântulas de angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E), empregando-se o inverso do tempo para alcançar 50% de germinação para expressar a taxa desse processo.............
95

11
LISTA DE TABELAS
Página
1 Valores obtidos do teor de água (%) e peso de mil sementes (g) de angico vermelho, cerejeira, sete-cascas, sucupira preta e tarumarana.........................................................................................
49
2 Regressões empregadas nas estimativas das temperaturas cardinais para a germinação das sementes das espécies em
estudo, com base no índice de velocidade do processo (50). .........
87
3 Regressões empregadas nas estimativas das temperatura cardinais para formação de plântulas das espécies em estudo, com base no
índice de velocidade do processo (50). ............................................
89
4 Temperaturas cardinais para o processo de germinação de sementes e de formação de plântulas obtidas a partir do índice de
velocidade desses processos (50) nas diferentes espécies em estudo. ..............................................................................................
91
5 Regressões empregadas nas estimativas das temperaturas cardinais para a germinação das sementes das espécies em estudo, com base nas taxas de desenvolvimento. ...........................
92
6 Regressões empregadas nas estimativas das temperaturas cardinais para formação de plântulas das espécies em estudo, com base nas taxas de desenvolvimento. ........................................
94
7 Temperaturas cardinais para o processo de germinação de sementes e formação de plântulas obtidas a partir das taxas de desenvolvimento nas diferentes espécies em estudo. ......................
96

12
SUMÁRIO
Página
1 INTRODUÇÃO…………………………………………………………… 13
2 REVISÃO DE LITERATURA…………………………………………… 16
3 MATERIAL E MÉTODOS………………………………………………. 43
3.1 Variáveis analisadas…………………………………………………...
45
3.2 Determinação das temperaturas cardinais para a germinação e formação de plântulas..........................................................................
48
4 RESULTADOS E DISCUSSÃO……………………………………….. 49
4.1 Teores de água e peso de mil sementes........................................
49
4.2 Avaliação da dinâmica da germinação e da formação de plântulas para os diferentes tratamentos térmicos ................................................
51
4.3 Índice da velocidade (50) de germinação das sementes e formação de plântulas para os diferentes tratamentos térmicos............
79
4.4 Tempo para obtenção de 50% de germinação das sementes e 50% do formação de plântulas para os diferentes tratamentos térmicos...................................................................................................
83
4.5 Estimativas das temperaturas cardinais para a germinação das sementes e formação de plântulas a partir do índice de velocidade desses processos (50)...................................................................................
86
4.6 Estimativas das temperaturas cardinais para a germinação das sementes e formação de plântulas a partir das taxas de desenvolvimento.....................................................................................
91
5 CONCLUSÃO …………………………………………………………….. 99
6 REFERÊNCIAS BIBLIOGRÁFICAS……………………………………. 101
7 ANEXO……………………………………………………………………... 113

13
1 INTRODUÇÃO
As mudanças no uso da terra que alteraram os ecossistemas
naturais no estado de Mato Grosso estão associadas, na sua maior
proporção, às atividades agrícolas, devido à implantação de cultivos e
pastagens e, em menor proporção, à atividade de mineração. Em todos
esses casos, a alteração da paisagem natural afeta diretamente a dinâmica
populacional das espécies originais, em um primeiro momento pela
substituição dessas espécies por outras e, em um segundo momento,
porque a sobre-exploração causa degradação ambiental de tal magnitude
que se esse ambiente degradado for deixado à própria sorte não
necessariamente voltará a ter a mesma composição florística original.
Nos biomas do estado de Mato Grosso vem ocorrendo uma
crescente degradação associada ao aumento da demanda por novas áreas
para serem incorporadas ao processo produtivo (agronegócio), como o
fornecimento de madeira, carne bovina e soja, realizado por meio da
eliminação da floresta e o estabelecimento de pastagem e cultivos. O
Cerrado, por exemplo, tem sido desmatado em ritmo duas vezes superior
ao da Amazônia (Pardini, 2008), pois ocorre desmatamento de 22 mil km2
por ano (Novaes, 2007). Os rios do Pantanal tem forte assoreamento

14
(Scherer, 2008) e suas matas ciliares originais tiveram desmatamento
situado entre 11,5% e 17%, dependendo da metodologia adotada (Girardi,
2008). Abdon et al. (2007) quantificaram 11,27% de áreas antropizadas até
2002 por desmatamento no bioma pantanal no estado de Mato Grosso.
Cerca de 18% da área total da floresta Amazônica já foi desmatada e
se estima que mais de 25% da área total desmatada nessa região
amazônica, em torno de 165.000 km2, encontra-se abandonada ou sub-
utilizada, muitas vezes em estado de degradação. Dados estimados em
2004 apontaram que mais da metade da área de pastagens cultivadas no
Cerrado se encontravam degradadas ou em degradação (Souza et al.,
2004). Nos últimos anos, Mato Grosso foi o estado em que se observou
maior degradação florestal; com uma área de 8.744 km2 em 2007 e de
12.534 km2 em 2008 (INPE, 2008).
Em áreas degradadas, a reconstituição da vegetação pode ocorrer
pelo crescimento das árvores remanescentes ou através do plantio de
novas mudas (Larson, 1992). No Brasil, de modo geral, a recomposição
dessas áreas tem sido efetuada por meio de novos plantios, o que exige o
conhecimento do comportamento germinativo das espécies usadas com
essa finalidade, e os requerimentos das sementes para formação de
plântulas com probabilidade real de se tornarem indivíduos adultos.
A recuperação de áreas degradadas, por meio do emprego de
sementes, pode não ter sucesso devido a incompatibilidade entre as
temperaturas apresentadas no substrato (solo) e as temperaturas cardinais
do processo germinativo e formação de plântulas.
Dentre os fatores determinantes do processo germinativo e de
estabelecimento de plântulas, a disponibilidade de água, o fotoperíodo e a
temperatura são reconhecidamente os que mais afetam. Nesse contexto, a
temperatura passa a ser o fator mais importante na regulação da
germinação de sementes não dormentes quando as condições de luz,
nutrientes e umidade não são limitantes (Garcia-Huidobro et al., 1982).
Por outro lado, a alteração da paisagem original causa igualmente
uma alteração no balanço energético da superfície do solo, porque muda o

15
albedo da superfície (Jackson e Idso, 1975; Sagan et al., 1979; Bonan,
2008) e expõe o solo sem proteção à demanda evaporativa da atmosfera, o
que altera seu conteúdo de água disponível e, por conseguinte, sua
capacidade tampão de manter a temperatura estável.
Todos esses aspectos foram decisivos na definição deste estudo,
porque somente conhecendo as temperaturas cardinais do processo de
germinação e formação de plântulas de espécies nativas é que se pode
levar a cabo projetos de recuperação de áreas degradadas dos
ecossistemas naturais com resultados mais efetivos.
Supõe-se que a faixa de temperatura entre 10 a 40°C seja suficiente
para estimar as temperaturas cardinais para os processos de germinação
das sementes e formação de plântulas das espécies angico vermelho,
cerejeira, sete-cascas, sucupira preta e tarumarana. Também que para
cada uma dessas espécies, as temperaturas cardinais são específicas para
cada estádio de desenvolvimento da plântula, e que as diferenças entre as
temperaturas cardinais para cada estádio de desenvolvimento da plântula
das espécies de um mesmo bioma são inferiores às observadas entre
espécies de diferentes biomas.
Assim, os objetivos neste estudo foram avaliar o requerimento
térmico para a germinação e formação de plântulas de cinco espécies de
ocorrência natural nos biomas do estado de Mato Grosso, avaliar a
qualidade física das sementes de angico vermelho (Anadenanthera
macrocarpa (Benth.) Brenan), cerejeira (Amburana cearensis (Alemão) A.C.
Smith), sete-cascas Samanea tubulosa (Benth.) Barneby & J.W. Grimes,
sucupira preta (Bowdichia virgilioides Kunt) e tarumarana (Buchenavia
tomentosa Eichler); avaliar a dinâmica do processo germinativo e da
formação de plântulas em função da temperatura; identificar quais as
temperaturas cardinais para o processo germinativo das sementes e
formação de plântulas; e identificar diferença nas temperaturas cardinais
para o processo germinativo e formação de plântulas entre espécies de um
mesmo bioma e de biomas distintos.

16
2 REVISÃO DE LITERATURA
2.1 Biomas do Estado de Mato Grosso e o Impacto na Mudança no Uso
da Terra
Bioma é caracterizado uma área do espaço geográfico, com
dimensões de até mais de um milhão de quilômetros quadrados, que tem
por características a uniformidade de um macroclima definido, de uma
determinada fitofisionomia ou formação vegetal, de uma fauna e outros
organismos vivos associados, e de outras condições ambientais, como a
altitude, o solo, alagamentos, o fogo, a salinidade, entre outros. Essas
características todas lhe conferem uma estrutura e uma funcionalidade
peculiares, uma ecologia própria (Coutinho, 2006).
No Estado de Mato Grosso existem três grandes biomas: Cerrado,
Pantanal e Floresta Amazônica (Figura 1).

17
FIGURA 1. Mapa de biomas do Brasil. Fonte: IBGE (2004)
A vegetação do bioma Cerrado apresenta fisionomias que englobam
formações florestais, savânicas e campestres. As formações florestais
típicas do Cerrado são as matas ciliares e as matas de galeria, que estão
associadas a cursos de água e as matas secas ou estacionais, que
ocorrem, nos interflúvios, em terrenos bem drenados e ricos em nutrientes
(Ribeiro e Walter, 1998).
O Cerrado que ocupa 25% do território brasileiro é o segundo maior
bioma da América do Sul, perdendo em tamanho somente para a Floresta
Amazônica. Sua flora riquíssima só agora começa a ser conhecida,
existindo cerca de 1000 espécies de árvores, 3000 espécies de ervas ou
arbustos e quase 500 trepadeiras (IBAMA, 2008).
Nos últimos 30 anos, a progressiva mecanização das lavouras e a
facilidade de preparar e adubar o solo contribuem para devastação
acelerada de vegetação nativa e se estima que cerca de 40% do bioma
Cerrado já tenha sido desmatado (Roesler et al., 2007), comprometendo a
conservação da sua biodiversidade. Analisando-se os resultados obtidos no
período de 2000/2001, nas regiões de cerrado, verificou-se que 19,2% da

18
área total foi convertida em culturas de soja, ou seja, a cultura contribuiu
fortemente para a mudança da cobertura da terra nesse bioma (Anderson et
al., 2003).
Entre as mais ricas savanas do mundo, a flora do cerrado brasileiro
apresenta espécies nativas que merecem especial atenção, pois esse
bioma foi considerado recentemente como um dos “hotspots” mundiais de
diversidade (Martins, 2004). Está entre os biomas de maior biodiversidade
brasileira. Maior reserva de diversidade biológica do mundo, a Amazônia é
também o maior bioma brasileiro em extensão e ocupa quase metade do
território nacional (49,29%). Sua área, de aproximadamente 6,5 milhões de
quilômetros quadrados, abriga a maior rede hidrográfica do planeta, que
escoa cerca de 1/5 do volume de água doce do mundo. Sessenta por cento
da bacia amazônica se encontra em território brasileiro, onde o Bioma
Amazônico ocupa a totalidade de cinco unidades da federação (Acre,
Amapá, Amazonas, Pará e Roraima), grande parte de Rondônia (98,8%),
mais da metade de Mato Grosso (54%), além de parte de Maranhão (34%)
e Tocantins (9%) (IBGE, 2004).
A floresta Amazônica é um ecossistema auto-sustentável. Ou seja, é
um sistema que se mantém com seus próprios nutrientes num ciclo
permanente. Os ecossistemas amazônicos são sorvedouros de carbono,
contribuindo para o equilíbrio climático global. Existe um delicado equilíbrio
nas relações das populações biológicas que são sensíveis a interferências
antrópicas. A grande diversidade geológica, aliada ao relevo diferenciado,
resultou na formação das mais variadas classes de solo, sob a influência
das grandes temperaturas e precipitações, características do clima
equatorial quente superúmido e úmido. Contudo, a fertilidade natural dos
solos é baixa, em contraste com a exuberância das florestas ombrófilas
(úmidas) que nelas se desenvolvem. A vegetação apresenta grande
variedade, destacando: matas de terra firmes, florestas inundadas, várzeas,
igapós, campos abertos e cerrados. Abriga uma infinidade de espécies
vegetais e animais (IBAMA, 2008). Entretanto, para o bioma Amazônia, a
pressão da soja para o desmatamento apresenta uma série de fatores de

19
pressão intrínsecos, atuando como agentes potenciais para a mudança do
uso e da cobertura da terra, como a intensificação da produção de soja, as
queimadas, o desmatamento, a abertura e pavimentação de estradas
(Anderson et. al., 2003).
No Brasil, o Pantanal ocupa 150.355 km², ou seja, 1,76% do território
nacional. A localização geográfica do bioma Pantanal é de particular
relevância, uma vez que representa a ligação entre o Cerrado, no Brasil
Central, o Chaco, na Bolívia, e a região Amazônica, ao Norte, identificando-
se, aproximadamente, com a bacia do alto Paraguai. Como área de
transição, a região do Pantanal ostenta um mosaico de ecossistemas
terrestres, com afinidades, sobretudo, com os Cerrados e, em parte, com a
floresta Amazônica, além de ecossistemas aquáticos e semi-aquáticos,
interdependentes em maior ou menor grau. Os planaltos e as terras altas da
bacia superior são formados por áreas escarpadas e testemunhos de
planaltos erodidos, conhecidos localmente como serras. São cobertos por
vegetações predominantemente abertas, tais como campos limpos, campos
sujos, cerrados e cerradões, determinadas, principalmente, por fatores de
solo (edáficos) e climáticos e, também, por florestas úmidas,
prolongamentos do ecossistema amazônico. Uma série de atividades de
impacto direto sobre o Pantanal pode ser observada, como garimpo de ouro
e diamantes, caça, pesca, turismo e agropecuária predatória, construção de
rodovias e hidrelétricas. Convém frisar a importância das atividades
extensivas nos planaltos circundantes como uma das principais fontes de
impactos ambientais negativos sobre o Pantanal (IBGE, 2004; EMBRAPA,
2008; IBAMA, 2008).
A supressão dos biomas vem ocorrendo em velocidade e intensidade
jamais vistas na história da humanidade, e se concentra nas áreas
remanescentes localizadas nos países do hemisfério Sul. Ela é promovida
para o desenvolvimento de atividades agrícolas extensivas, mineração,
infra-estruturas de transporte e energia, empreendimentos industriais e
assentamentos urbanos. Na realidade, a conversão desses biomas faz
parte de um processo de intensa exploração e disputa por todos os tipos de

20
recursos naturais, que vem ocorrendo no mundo inteiro e abrangem
matérias-primas, solos para a agricultura, minérios, petróleo, recursos
hídricos etc., processo esse determinado, também, pelo esgotamento das
reservas até então exploradas (Tagnin e Mattes, 2008).
Os biomas do estado de Mato Grosso têm sofrido o impacto das
atividades de exploração dos seus recursos naturais. E isso tem se
intensificado pela crescente demanda de grãos, fibras e carne para
exportação, que têm acarretado significativas mudanças no cenário agrícola
do território brasileiro. Áreas nativas estão sendo ocupadas por pastagem e
agricultura, terras utilizadas como pastagem estão sendo convertidas para
agricultura, e aquelas em uso agrícola há mais tempo estão sendo
submetidas a sistemas de manejo mais intensivos e tecnicamente mais
avançados. Os dois primeiros usos e mudanças de uso da terra vêm
ocorrendo em grandes proporções e de forma bastante acelerada na
Amazônia Legal e, em particular, na região sudoeste, abrangendo os
estados de Rondônia e Mato Grosso. As mudanças no uso e cobertura da
terra que vêm ocorrendo nos últimos 30 anos certamente estão causando
significativos impactos ambientais globais, sobretudo no que se refere ao
aquecimento da atmosfera terrestre. Além disso, a intensificação do uso
agrícola provoca degradação das terras em escalas regional e local. A
expansão agrícola nos ecossistemas ocasiona grande impacto ambiental,
pois incrementa a emissão de gases do efeito estufa (GEE) para a
atmosfera. Isso acontece devido ao desmatamento e à degradação do solo,
em particular da matéria orgânica, embora gere benefícios sócio-
econômicos importantes, pelo menos temporários, para a região Pantanal
(IBGE, 2004; EMBRAPA, 2008; IBAMA, 2008, INPE, 2008).
Na Figura 2 observam-se fotos do processo de degradação
progressiva onde o processo se inicia com a floresta intacta e termina com a
conversão completa da floresta original em outras coberturas. É prática
comum a introdução de capim nessas áreas degradadas na Amazônia. O
capim e a cobertura florestal remanescente são queimados e, quando o
capim rebrota, o gado é colocado para pastar na área assim degradada.

21
Queimadas subseqüentes destroem completamente o que restou da floresta
inicial e o processo de degradação pode durar anos (INPE, 2008).
FIGURA 2. Processo de degradação progressiva no bioma Amazônico.
Fonte: INPE (2008).
2.2 Aspectos Gerais da Multiplicação de Plantas nos Ecossistemas
Naturais e a Recuperação de Áreas Degradadas
As espécies vegetais multiplicam-se sexuada e assexuadamente. A
propagação (multiplicação assexuada) consiste na utilização de estruturas
vegetativas que apresentem capacidade de regeneração. Portanto, a
multiplicação assexuada é efetuada através de partes da planta aptas a
produzir indivíduos geneticamente idênticos à planta matriz, por meio de um
princípio denominado de totipotência. Como essas células reproduzidas são
somáticas, as plantas resultantes são denominadas clones (Graça e
Tavares, 2000; Marcos Filho, 2005). No entanto, há desvantagens, como as
maiores dificuldades para o manejo de materiais de propagação e o custo
relativamente elevado do transporte, devido ao volume do material

22
necessário para o plantio; as dificuldades ou a impossibilidade de
armazenamento dos propágulos em função da rápida perda de viabilidade;
o custo e o trabalho despendidos para a obtenção das mudas geralmente
são superiores aos verificados para produção de sementes; não há
variabilidade dos descendentes, além da menor capacidade de adaptação a
variações climáticas (Marcos Filho, 2005), porém tem a vantagem das
mudas obtidas por processo assexuado serem plantas com estabilidade
garantida, o que implicará na formação de plantas uniformes e mais
produtivas (Oliveira et al., 2003).
A reprodução (multiplicação sexuada) é baseada no processo
meiótico de divisão celular, em que o número de cromossomos das células
reprodutivas é reduzido à metade para formar os gametas: oosfera e o grão
de pólen. A divisão meiótica é de fundamental importância para a geração
da variabilidade por meio da divisão reducional e independente dos
cromossomos e Crossing over. Durante a divisão meiótica, os cromossomos
homólogos pareados trocam pares entre si, aumentando a variabilidade
genética (Borém, 1998). A propagação de plantas normalmente é realizada
por meio de sementes. No entanto, por esse método não se tem a certeza
de que os indivíduos formados, devido à recombinação gênica, mantenham
as mesmas características selecionadas das plantas parentais (Ono et al.,
1992). A propagação através de sementes retarda a produção, devido ao
longo período improdutivo ocasionado pela juvenilidade. Além disso, muitas
plantas originadas de sementes têm uma pequena produção (Hartmann et
al., 1997).
No processo de recuperação de áreas degradadas, um dos pontos
mais importantes e cruciais refere-se aos aspectos relacionados à
tecnologia de sementes e produção de mudas, já que a qualidade dos
reflorestamentos está intimamente ligada à qualidade dos indivíduos que o
compõem. Sendo assim, a propagação vegetativa (assexuada) é pouco
recomendada, principalmente devido ao fato de reduzir a variabilidade
genética das espécies, atuando na contramão dos princípios básicos na
implantação de florestas heterogêneas (Faria, 1999).

23
É imprescindível, então, que as mudas destinadas à recomposição
vegetal sejam produzidas, a partir de sementes (propagação sexuada),
provenientes de lotes que garantam a variabilidade genética das espécies e
para isso, diversos pontos devem ser contemplados (Davide et al., 1995;
Faria, 1999; Barbosa, 2000).
A realização de estudos que enfoquem aspectos da biologia
reprodutiva e fatores ambientais que possivelmente estão interferindo na
reprodução de espécies nativas é indispensável para estabelecer
estratégias para a conservação das mesmas (Velten e Garcia, 2005).
Dentre esses, o crescimento e propagação de espécies nativas tornam-se
necessários, uma vez que as técnicas de produção dessas espécies são
muito pouco conhecidas (Stringheta et al., 2005).
As diferenças fisiológicas na germinação de sementes nativas podem
ser reflexos de adaptações às condições ecológicas às quais essas
espécies estão sujeitas (Ranieri et al., 2003). A germinação de sementes é
um processo complexo, que envolve muitas reações e fases (Lopes e
Pereira, 2005). Para a análise de tais processos é utilizado teste de
germinação sob temperaturas controladas e fotoperíodo (Iossi et al., 2003).
A porcentagem de germinação varia em função da qualidade de luz, porém
na luz branca e na ausência de luz ocorrem variações em conseqüência das
temperaturas (Menezes et al., 2004) e esses percentuais podem estar
relacionados à preferência da espécie por ambientes com temperaturas
ideais (Fior et al., 2004). O comportamento germinativo em ampla faixa de
temperatura pode contribuir de forma significativa para o sucesso no
recrutamento de espécies em habitat natural, onde estão sujeitas a altas
irradiâncias e grandes variações de temperaturas (Garcia e Diniz, 2003;
Silva et al., 2006).

24
2.3 Descrição das Espécies
2.3.1 Anadenanthera macrocarpa (Benth.) Brenan – Angico Vermelho
Anadenanthera macrocarpa (Benth.) Brenan (Figuras 3 e 4 ) (sin.
Piptadenia macrocarpa Benth.), pertence a família Fabaceae e é
popularmente conhecida como angico, angico-vermelho e angico-preto. É
uma espécie arbórea com até 20 m de altura e tronco de 0,40-0,80 m de
diâmetro, com ocorrência bastante representativa nas caatingas, nos
estados do Maranhão e Nordeste do país até São Paulo, Minas Gerais e
Mato Grosso do Sul, principalmente na floresta latifoliada semidecídua. Tem
utilização muito diversificada como a extração de tanino da casca em
curtumes, fabricação de móveis, construção civil, forragens das folhas
fenadas, ornamentação e carvão, entre outras (Lorenzi, 1992). Planta
decídua, pioneira, heliófita, e seletiva xerófila, características das capoeiras,
florestas secundárias situadas em terrenos arenosos e cascalhentos, como
na mata primária densa em solos argilosos e férteis. É bastante freqüente
em Cerradões e matas de galeria de todo o Brasil Central, e ocorre
preferencialmente em terrenos altos e bem drenados. Árvore própria para
ornamentação e arborização de parques e praças e melífera. Por
apresentar rápido crescimento, pode ser aproveitada com sucesso para
reflorestamentos de áreas degradadas de preservação permanente
(Lorenzi, 1992) e muito usada para recomposição de matas ciliares
(Carvalho, 1994).

25
FIGURA 3. Aspecto da planta com frutos de Anadenanthera macrocarpa,
Cuiabá-MT.
FIGURA 4. Frutos (A) e sementes (B) de Anadenanthera macrocarpa.
A B

26
2.3.2 Amburana cearensis (Alemão) A.C.Smith – Cerejeira
Amburana cearensis (Alemão) A.C. Smith (Figura 5) (sin. Torresea
cearensis Fr. All), pertencente à família Fabaceae, também conhecida como
cumaru, amburana de cheiro e cumaru-do-Ceará, apresenta porte regular,
podendo atingir até 4-10 m de altura nas regiões de caatinga (Corrêa, 1978;
Lorenzi, 1992) e até 20 m na zona da mata pluvial e caducifólia, com tronco
marrom avermelhado de 0,40-0,80 m de diâmetro (Lorenzi, 1992). Ocorre
naturalmente do Nordeste ao Brasil Central, em regiões de caatinga e na
floresta pluvial de Minas Gerais, no vale do Rio Doce, nos afloramentos
calcários e matas decíduas do centro oeste e sudeste (Braga, 1976; Corrêa,
1978; Lorenzi, 1992). A espécie apresenta ampla distribuição geográfica na
América do Sul, sendo característica de floresta estacional. Portanto, ocorre
em floresta estacional semidecidual, restrita aos afloramentos rochosos ou
calcáreos; em floresta estacional decidual submontana (oeste da Bahia); em
floresta ombrófila densa (floresta atlântica), entrando até a caatinga/mata
seca, onde é freqüente. A sua ocorrência no Cerrado e no Pantanal
restringe-se às manchas de florestas estacionais de afloramento calcáreo e
suas zonas de transição com o Cerrado sensu stricto (Mendonça et al.
1998) em áreas bem drenadas e de moderada a elevada fertilidade. A
madeira é empregada em serviços de movelaria e marcenaria em geral e,
as sementes, em função do odor agradável exalado, são utilizadas para
perfumar roupas (Lorenzi, 1992). As sementes (Figura 6B) são utilizadas,
ainda, na medicina caseira como anti-espasmódica, emanagoga e para o
tratamento de doenças reumáticas (Tigre, 1968; Braga, 1976). A árvore é
muito ornamental, principalmente pelos ramos e tronco que são lisos de cor
vinho ou marrom avermelhado podendo ser usada no paisagismo em geral
(Lorenzi, 1992). Planta decídua, heliófila, seletiva xerófita, característica de
afloramentos rochosos ou calcários e terrenos secos em floresta decídua.
De acordo com Tigre (1968), a espécie pode ser recomendada para
recuperação de áreas degradadas.

27
FIGURA 5. Árvore de Amburana cearensis.
FIGURA 6. Frutos (A) e sementes (B) de cerejeira (Amburana cearensis).
A B

28
2.3.3 Samanea tubulosa (Benth.) Barneby & J.W. Grimes – Sete-Cascas
A Samanea tubulosa (Figura 7), popularmente conhecida como sete-
cascas, farinha seca, pertence à família Fabaceae. É uma árvore com porte
de 4 a 18 m, copa arredondada, tronco ereto, cilíndrico, casca grossa,
fissurada e muito suberosa, com folhas compostas bipinadas. Ocorre no
Mato Grosso do Sul, Mato Grosso no Pantanal Matogrossense, Chapada
dos Guimarães e Floresta Amazônica, Sul do Pará e Baixo Amazonas,
Bahia, Pará, Paraguai, Bolívia e Peru (Lorenzi, 2002). Ocorre
preferencialmente em capoeiras e em áreas abertas, como colonizadora em
várzeas aluviais e a beira de rios, onde o solo é suprido de água. De
crescimento rápido, é árvore ornamental e muito cultivada na arborização
rural. A madeira é empregada para marcenaria, moirões e para lenha. A
vagem dos frutos (Figura 8A) é forrageira para o gado. Encontrada em solos
organo-arenoso (Lorenzi, 2002). Planta caducifólia, heliófila, seletiva
hidrófita, pioneira, características da mata semidecídua do Pantanal
Matogrossense, da mata caducifólia do vale de São Francisco e das
savanas Amazônicas. Produz anualmente moderada quantidade de
sementes viáveis (Figura 8B) que são disseminadas por animais domésticos
(Lorenzi, 2002). Em solo com textura arenosa foi constatada nodulação
radicial com Rhyzobium em campo e em viveiro (Souza et al., 1994).
FIGURA 7. Aspecto da planta com flores de Samanea tubulosa.

29
FIGURA 8. Frutos (A) e sementes (B) de Samanea tubulosa.
2.3.4 Bowdichia virgilioides Kunt – Sucupira Preta
Bowdichia virgilioides (Figuras 9), conhecida vulgarmente como
sucupira preta, é uma espécie arbórea pertencente à família Fabaceae com
ampla dispersão pelo Brasil (Brandão e Ferreira, 1991; Lorenzi, 1992).
Planta decídua, heliófita, seletiva xerófita, característica do cerrado, floresce
de maio a setembro (Lorenzi, 1992; Silva Junior, 2004). É uma árvore típica
de áreas de savanas arborizadas. Essa espécie é encontrada nas regiões
nordeste e centro oeste brasileiras, onde é conhecida pelos nomes
populares de: sucupira-do-cerrado, sucupira-do-campo, angelim amargoso,
coração-de-negro (Almeida et al., 1998). Árvore de casca grossa e
fendilhada atinge até 20 m de altura, apresentando tronco com diâmetro
máximo de 0,60 m. Possue folhas compostas, pinadas, com folíolos
pubescentes (Lorenzi, 1992). As pequenas flores, com corola lilás conferem
um aspecto ornamental e apícola à árvore. Os frutos (Figura 10A) são
legumes, indeiscentes, achatados, contendo pequenas sementes (Figura
10B) com 3 a 5 mm de comprimento, apresentando coloração avermelhada
e dispersos pelo vento (Rizzini, 1990; Lorenzi, 1992). Planta pioneira e
adaptada a terrenos secos e pobres é ótima para plantios em programas de
reflorestamento e na recuperação de áreas degradadas de preservação
permanente e uso potencial em paisagismo, pela arquitetura e bela floração
(Lorenzi, 1992; Silva Junior, 2004).
A B

30
FIGURA 9. Árvores de Bowdichia virgilioides no campus da Faculdade de
Agronomia e Medicina Veterinária – Cuiabá- MT.
FIGURA 10. Flores e frutos (A), sementes (B) de Bowdichia virgilioides
(sucupira preta).
A B

31
2.3.5 Buchenavia tomentosa Eichler – Tarumarana
Buchenavia tomentosa (Figura 11), pertencente à família
Combretaceae, conhecida popularmente como tarumarana, cuiarana e
mirindiba, é uma árvore de grande porte com cerca de 5 a 12 metros de
altura, copa ampla e densa, tronco curto, folhas simples, com frutos de
setembro-outubro (Lorenzi, 2002), florescendo no mês de julho, perdendo
as flores durante a floração (Silva e Valente, 2005), com frutos em abril-
agosto (Pott e Pott, 1994). Ocorre predominantemente nos cerradões e
mata latifoliada semidecídua e nos estados de Tocantins, Goiás, Bahia,
Minas Gerais, Mato Grosso, Mato Grosso do Sul (Lorenzi, 2002),
encontrada em solos areno-argilosos (Lorenzi, 2002). Na Reserva Ducke é
uma árvore que vive em floresta de vertente, e cresce em áreas de solo
argiloso (Silva e Valente, 2005). Ocorre em campo cerrado, solos arenosos
e tolera fogo (Pott e Pott, 1994). Sua madeira moderadamente pesada,
dura, de boa resistência, é empregada na construção civil, tábuas e vigas,
como moirões, em cercas e também para lenha e carvão. Planta
semidecídua, heliófila, secundária, produz anualmente abundante
quantidade de sementes viáveis prontamente disseminadas pela fauna (Pott
e Pott, 1994; Lorenzi, 2002).
FIGURA 11. Árvore de Buchenavia tomentosa (tarumarana) em Santo
Antônio do Leverger-MT, Fazenda Experimental da
Faculdade de Agronomia e Medicina Veterinária.

32
É uma espécie que apresenta grande potencial apícola, sendo
recomendada para restauração florestal em áreas degradadas, devido à
grande procura dos seus frutos pela fauna regional, que conseqüentemente
irá promover sua disseminação (Azevedo, 2008). Os frutos (Figura 12) são
comestíveis e avidamente procurados por animais selvagens, tornando-a
muito recomendável para composição de reflorestamentos heterogêneos
destinados a preservação permanente (Lorenzi, 2002).
FIGURA 12. Frutos (A) e sementes (B) de tarumarana.
2.4 Germinação
A germinação é um evento fisiológico que depende da qualidade da
semente e das condições de germinação, como suprimento de água e de
oxigênio, temperatura, luz e substrato (Salomão et al., 2003). É necessário
que alguns fatores relacionados à semente e ao ambiente atuem de forma
favorável. Para tanto, é essencial que a semente esteja viva, sem
dormência e que haja disponibilidade de água, temperatura e oxigênio
(Bewley e Black, 1994).
A fase de germinação tem início com a embebição da água e com a
ativação do metabolismo do tecido embrionário. O primeiro passo é a
produção de energia via glicólise. Os fitormônios dão o sinal para a síntese
de enzimas, levando a mobilização de substâncias de reservas. Segue-se a
A B

33
síntese de hormônios, promovendo a divisão celular e o crescimento em
extensão, reorganização do protoplasma, intensificação da respiração
mitocondrial, síntese de proteínas e, mais tardiamente pelos processos de
crescimento que resultam no aparecimento da radícula (Larcher, 2000).
A temperatura é um fator determinante para a germinação das
sementes e está diretamente associada às características ecológicas das
espécies. Além disso, o processo germinativo compreende várias fases e
cada qual requer uma determinada temperatura (Mayer e Poljakoff-Mayer,
1989; Bewley e Black, 1994).
As espécies nativas do Brasil possuem sementes que germinam em
uma ampla faixa térmica, dependendo do bioma e da região, e parecem
apresentar relação positiva entre a temperatura ótima e o regime térmico da
região de sua ocorrência natural (Borghetti, 2005).
Entretanto uma germinação rápida e uniforme das sementes, seguida
por imediata emergência das plântulas são características altamente
desejáveis na formação de mudas, pois quanto mais tempo a plântula
permanecer nos estádios iniciais de desenvolvimento e demorar para
emergir do solo, mais vulnerável estará às condições adversas do meio
(Martins et al., 1999).
2.5 Temperatura
A temperatura, juntamente com a água e o oxigênio constituem os
principais fatores do ambiente que influenciam na germinação de uma
semente (Bewley e Black, 1982; Borges e Rena, 1993; Carvalho e
Nakagawa, 2000; Marcos Filho, 2005). Além desses, Bewley e Black (1982)
incluem a luz como fator determinante na germinação de sementes. As
sementes de uma mesma amostra podem variar nas exigências para
germinação devido a diferenças de maturidade entre elas, entre as matrizes
coletadas ou, ainda, na posição das sementes em uma mesma matriz
(Austin, 1972).

34
Os efeitos dos estresses ambientais, principalmente os causados
pela temperatura, ainda não são completamente conhecidos. O número
relativamente pequeno de pesquisas sobre o assunto, geralmente
conduzidas sob condições controladas, sugerem que a elevação da
temperatura, até atingir valores superiores a 30°C, pode causar prejuízos
severos à produção e à qualidade das sementes (Marcos Filho, 2005).
Para Bewley e Black (1982), a temperatura age na germinação de
três formas: determinando a capacidade e a porcentagem de germinação
das sementes, eliminando a dormência primária e secundária ou induzindo
a dormência secundária. A temperatura influencia a porcentagem final e a
velocidade da germinação, afetando tanto a absorção de água pela
semente quanto as reações bioquímicas que regulam o metabolismo
envolvido nesse processo (Bewley e Black, 1994).
Além de controlar a velocidade das reações metabólicas, a
temperatura é também determinante na estabilidade das membranas
celulares (Paiva e Oliveira, 2006). As membranas são compostas de
fosfolipídios e proteínas, formando um complexo que, em situação normal,
permanece em fase “cristalina liquida”, alterada para a de “gel”, em
circunstâncias especiais influenciadas pelas condições do ambiente
(Marcos Filho, 2005).
Quando submetidas a temperaturas constantes, sementes de
algumas espécies exibem modificações na composição e na estrutura da
camada de lipídios das membranas as quais passam da fase cristalina,
típica de elevada organização para a fase fluida ou desordenada. As
mudanças foram proeminentes na faixa de 30° a 35°C, com aumento de
efluxo de aminoácido da semente durante a germinação e com decréscimo
de germinação nas espécies que apresentaram essa alteração. Entretanto,
não se pode afirmar que seja a mudança na membrana o controle
determinante da germinação, sendo mais provável o efluxo de solutos por
meio de alterações metabólicas (Hendricks e Taylorson, 1976). Quando é
afetado o sistema de membranas, há liberação de solutos e isso implica que
muitos desses constituintes exsudados são essenciais para a germinação,

35
para a manutenção do potencial osmótico interno necessário para a
protrusão da raíz primária, além de que a liberação desses exsudados para
o meio externo estimula o desenvolvimento de microrganismos (Marcos
Filho, 2005), que poderá afetar a porcentagem de germinação.
A sensibilidade da semente à embebição é controlada pelo teor inicial
de água, temperatura ambiente e taxa de absorção de água (Pollock, 1969,
citado por Marcos Filho, 2005). A interação teor inicial de água, temperatura
e embebição podem alterar acentuadamente a germinação e tem efeitos
drásticos no vigor da plântula, sobretudo em sementes grandes (De Castro
et al., 2004; Marcos Filho, 2005). A embebição rápida em temperaturas
baixas causa evento conhecido como dano de embebição e se as sementes
estiverem demasiadamente secas quando colocadas em água, podem
sofrer danos irreparáveis no nível do sistema de membranas, o que leva a
lixiviação de conteúdos celulares, afetando negativamente a germinação
(Pollock e Toole, 1966; Wolk et al., 1989, citados por De Castro et al.,
2004). Entretanto, em temperaturas mais altas, o aquecimento das
sementes secas, antes da embebição permite que a transição do estado de
gel para o cristalino líquido aconteça antes que a água seja introduzida,
estando as membranas prontamente no estado cristalino e se a água for
introduzida, acontecerá pouca lixiviação ou dano à semente.
A embebição sob baixas temperaturas (próximas a mínima) pode
provocar redução do crescimento das plântulas, mesmo quando a
temperatura retornar a níveis favoráveis. Esse prejuízo geralmente é
proporcional ao período de exposição a essa temperatura. Quando
sementes com baixos teores de água são embebidas rapidamente a 25°C,
há um declínio pouco acentuado do vigor, mas as reduções são agravadas
quando essa embebição ocorre sob temperaturas inferiores a 10°C (Marcos
Filho, 2005).
Em temperaturas mais quentes, as membranas das sementes já se
encontram no estado cristalino líquido e, assim podem tolerar o influxo
rápido de água. O mesmo vale para sementes com conteúdos de água mais
elevados em temperaturas mais baixas (De Castro et al., 2004).

36
Temperaturas baixas reduzem a atividade cinética das moléculas,
reduzindo a velocidade das reações químicas. Por outro lado, temperaturas
elevadas danificam as enzimas (desnaturação), provocam excessiva fluidez
das membranas, podendo acarretar perda de seletividade e extravasamento
do conteúdo celular. O efeito de temperaturas elevadas prejudica
primeiramente por mudanças das propriedades das membranas e, em
segundo momento por perda da atividade de enzimas (Paiva e Oliveira,
2006).
Estudos sobre a influência da temperatura na germinação das
sementes são essenciais para entender os aspectos ecofisiológicos e
bioquímicos (Labouriau, 1983; Bewley e Black, 1994). Para compreender os
efeitos da temperatura sobre a germinação, podem ser avaliadas mudanças
ocasionadas na porcentagem e velocidade de germinação ao longo do
tempo (Labouriau e Pacheco, 1978), sendo que a faixa de temperatura
ótima é aquela onde ocorre a germinação máxima no menor tempo médio,
enquanto sob temperaturas máxima e mínima as sementes pouco
germinam (Labouriau, 1983; Bewley e Black, 1994). A temperatura em que
se observa a máxima velocidade é ligeiramente superior à correspondente à
máxima porcentagem (Marcos Filho, 2005). As sementes apresentam
comportamento variável frente a esse fator, não havendo uma temperatura
ótima e uniforme para todas as espécies (Borges e Rena, 1993). Sementes
de indivíduos dentro da população podem, portanto ter níveis diferentes de
tolerância (limites) tanto a altas como a baixas temperaturas. Para qualquer
semente de indivíduos na população, a taxa de germinação aumenta a
partir de uma temperatura base para uma temperatura ótima acima da qual
diminui quando se aproxima de uma temperatura limite, o que indica os
limites de sua tolerância (Finch-Savage, 2004).
A temperatura ótima de germinação de espécies tropicais encontra-
se entre 15ºC e 30ºC, a máxima entre 35ºC e 40ºC e a mínima geralmente
são inferiores à 15ºC; as espécies adaptadas a climas mais frios suportam
melhor as baixas temperaturas. A velocidade de germinação e uniformidade
de emergência diminuem com temperaturas abaixo da ótima e temperaturas

37
acima da ótima aumentam a velocidade de germinação, embora somente as
sementes mais vigorosas consigam germinar (Marcos Filho, 2005; Nassif et
al., 1998). Acima e abaixo desses limites máximo e mínimo,
respectivamente, pode ocorrer a morte das sementes (Mayer, e Poljakoff-
Mayer, 1989).
2.6 Temperaturas Cardinais
A temperatura é uma variável fundamental para os seres vivos
porque afeta especificamente as suas taxas de desenvolvimento, de
maneira que se as condições térmicas do ambiente encontram-se dentro
dos limites de requerimento térmico do organismo, geralmente a sua taxa
de desenvolvimento aumenta linearmente com o aumento da temperatura
(Trudgill et al., 2005).
Considerando esse aspecto, foi desenvolvido o conceito de unidades
térmicas de desenvolvimento – do inglês, “thermal time” – expressa em
graus-dia – do inglês, “growing degree-days” – que relaciona a taxa de
desenvolvimento linearmente com as temperaturas do ambiente que
ocorrem acima da temperatura mínima requerida pelo organismo,
denominada temperatura basal mínima (Tm) – do inglês, “base temperature”
(Yin et al., 1995). Para essa situação, a forma de se contabilizar a soma
térmica necessária para o organismo completar um determinado estádio de
desenvolvimento se faz com o emprego da Equação 1, calculando-se
diariamente e se totalizando os valores obtidos em cada dia ao final daquela
etapa de desenvolvimento em questão.
mminmax T
TTGDD
2 (Equação 1)
Em que: GDD = unidades térmicas de desenvolvimento diário
(graus-dia),
Tmax = temperatura máxima do dia (°C),

38
Tmin = temperatura mínima do dia (°C),
Tm = temperatura basal mínima do organismo (°C).
McMaster e Wilhelm (1997) atentaram para o fato de que a Equação
1 pode expressar diferentes situações que, quando não esclarecidas,
comete-se um erro conceitual. Isso ocorre quando a temperatura do
ambiente se encontra abaixo do valor de Tm; então duas possibilidades
podem ser consideradas para calcular as unidades térmicas de
desenvolvimento: (a) quando a temperatura média (a semi-soma das
extremas) do ambiente é menor que Tm e o seu valor é substituído pelo
próprio valor de Tm e (b) quando a temperatura máxima do dia (Tmax) ou a
temperatura mínima do dia (Tmin) é inferior a Tm, ambas são substituídas
pelo valor de Tm. A diferença entre os valores das unidades térmicas de
desenvolvimento diário obtidos, considerando essas duas possibilidades
para o milho, foi de 83%, o que, segundo os autores, sugerem claridade na
descrição do método empregado.
Há situações em que a temperatura ambiental excede os limites
suportáveis pelo organismo e, nesses casos, é preciso considerar que há
restrições ao desenvolvimento. Obviamente que esse fenômeno há tempos
também já havia sido diagnosticado pelos pesquisadores, mas dado ao fato
de que, do ponto de vista prático, tal situação manifesta-se com menor
probabilidade do que aquela em que a temperatura ambiental esteja dentro
dos limites suportáveis, o emprego da Equação 1 se generalizou. Um dos
primeiros exemplos da indicação de temperaturas extremas reportado na
literatura é o trabalho de Lehenbauer (1914), que demonstrou claramente o
rápido declínio na taxa de desenvolvimento quando as temperaturas
ambientais excediam um valor ótimo para a espécie – denominado de
temperatura basal ótima (To).
A partir do trabalho desenvolvido por Lehenbauer (1914), Gilmore e
Rogers (1958) propuseram uma correção na contabilidade da soma térmica,
subtraindo da mesma toda a porção do dia em que a temperatura do ar
excede a temperatura basal ótima.

39
Garcia-Hidobro et al. (1982) e Roberts e Summerfield (1987)
definiram como sub-ótimas as temperaturas ambientais entre a temperatura
basal mínima e a temperatura basal ótima para o desenvolvimento do
organismo e como supra-ótimas aquelas situadas entre a temperatura basal
ótima e a temperatura basal máxima (TM) para o desenvolvimento do
organismo, denominando esses requerimentos térmicos do organismo de
temperaturas cardinais mínima, ótima e máxima. A temperatura é um dos
fatores ambientais que assume o mais relevante papel no controle do
crescimento, desenvolvimento e produção dos vegetais, uma vez que todos
os processos biológicos respondem à temperatura e todas essas respostas
podem ser sumarizadas em termos das temperaturas cardinais Tm, To e TM
(Yan & Hunt, 1999).
Existem vários métodos para diagnosticar as temperaturas cardinais.
Dentre esses, existe o método interativo de estimativa da temperatura basal
mínima por meio da obtenção da menor variabilidade possível do número
de dias, ou da soma térmica, para que a etapa de desenvolvimento em
estudo se complete inteiramente, sendo esta variabilidade medida pelo
desvio padrão (Magoon e Culpepper, 1932; Stier, 1939; Kitchen, 1956), pelo
coeficiente de variação (Madriaga e Knott, 1951), ou por ambos (Nuttonson,
1948; Lindsey e Newman, 1956). Entretanto, de acordo com Arnold (1959),
a melhor medida da variabilidade é o coeficiente de variação do número de
dias para a obtenção da soma térmica.
Um segundo método, proposto por Hoover (1955), utiliza análise de
regressão linear, com a soma térmica como variável dependente e as
diferentes temperaturas médias a que o organismo foi submetido como
variável independente. Nesse segundo método, a escolha correta da
temperatura basal mínima determina a obtenção de um modelo não
significativo, ou seja, com coeficiente angular igual a zero, o que significa
que para qualquer temperatura a soma térmica é a mesma. De acordo com
Arnold (1959), esse método fornece resultados semelhantes ao do uso do
coeficiente de variação do número de dias para a obtenção da soma
térmica.

40
Um terceiro método, proposto por Arnold (1959), utiliza também a
técnica da análise de regressão linear, mas nesse caso, a análise de
regressão é utilizada para se determinar o valor da temperatura basal
mínima. O eixo das ordenadas é composto pela taxa de desenvolvimento
(expressa pelo inverso do tempo requerido para completar a etapa de
desenvolvimento em estudo) e o das abscissas pelas temperaturas médias
a que o organismo foi submetido. A equação é resolvida para a taxa de
desenvolvimento igual a zero e, desta forma, fornece o valor da temperatura
cardinal mínima. Segundo o autor, esse método também fornece resultados
semelhantes ao do uso do coeficiente de variação do número de dias para a
obtenção da soma térmica.
O método proposto por Arnold (1959), que originalmente foi
empregado para calcular a temperatura basal mínima, pode igualmente ser
utilizado para o cálculo da temperatura basal máxima, desde que fornecido
ao organismo em estudo temperaturas mais elevadas que a temperatura
basal ótima do mesmo, o que, em última análise, resulta no modelo bilinear,
adotado por vários pesquisadores, como Craufurd et al. (1998), Olsen et al.
(1993), Omaga et al. (1995, 1996), dentre outros.
A consideração de que o ambiente pode prover temperaturas fora da
faixa de requerimento térmico dos organismos de interesse é de
fundamental importância nas pesquisas que buscam identificar as
temperaturas cardinais de fenômenos fisiológicos para organismos que em
seu ambiente ficam sujeitos a uma ampla faixa de temperatura. Em se
tratando de germinação de sementes, esse é o caso típico de estudo, posto
que de acordo com as propriedades físicas do solo, a temperatura que esse
pode apresentar na profundidade de semeadura pode, em muitos casos,
exceder a temperatura cardinal máxima da semente. Exemplos do emprego
desse método em estudos com germinação de sementes podem ser
observados nos trabalhos de Steinmaus et al. (2000).
Seguindo o mesmo princípio da relação linear entre a temperatura e
a taxa de desenvolvimento, a partir da inclusão da temperatura basal
máxima, a soma térmica passou a ser descrita pela soma de todas as áreas

41
sob a curva diária de temperatura ambiental dentro da faixa das
temperaturas basais máxima e mínima próprias de um determinando
processo fisiológico do organismo em estudo, requerida para que essa
etapa de desenvolvimento específico se complete inteiramente (Ometto,
1981). Sempre e quando a temperatura ambiental não exceder a
temperatura basal máxima do organismo, a contabilidade dos graus dia é
definida pela Equação 1, sendo esse um caso particular dentro de todas as
possibilidades abrangidas quando a temperatura basal máxima é incluída
na análise. Neste caso, supõe-se que a taxa de desenvolvimento varia
linearmente desde Tm até TM, desconsiderando-se as observações de
Lehenbauer (1914) e a correção para a soma térmica proposta por Gilmore
e Rogers (1958) acerca do declínio da taxa de desenvolvimento quando a
temperatura ambiental supera To.
Considerando que a temperatura pode não ser o fator único que
explica as taxas de desenvolvimento, foram feitas sugestões como a da
inclusão do fotoperíodo, convertendo a soma térmica em unidades
fototérmicas, ou a de funções de resposta para outros fatores ambientais
que afetam a fenologia como a disponibilidade de água, nutrientes,
qualidade e quantidade de luz, concentração de CO2 (Nuttonson, 1948;
Masle et al., 1989; McMaster et al., 1992; Willelm e McMaster, 1995).
Uma observação mais crítica revela que a relação linear das taxas de
desenvolvimento do vegetal com a temperatura do ambiente só é
encontrada dentro de uma determinada faixa de taxa de desenvolvimento
(Summerfield e Roberts, 1987). Foi assim que Gao et al. (1992) propuseram
o denominado “Clock Rice Model” para descrever o desenvolvimento do
cultivar Indica de arroz, entretanto, devido ao número excessivo de
parâmetros do modelo e a restrição artificial atribuída ao mesmo a fim de
impor que a taxa máxima de desenvolvimento ocorra exatamente à
temperatura basal ótima do organismo, o modelo não foi bem aceito por
apresentar descontinuidade na relação não linear.
Tomando como base o trabalho de Gao et al. (1992), Yin et al. (1995)
propuseram um novo modelo com o emprego da função beta, amplamente

42
utilizada em estatística como uma função de densidade de probabilidade de
curtose (Abramowitz e Stegun, 1965), mas que ao mesmo tempo não
apresenta os problemas do “Clock Rice Model”, porque para esse novo
modelo o ajuste não linear não é abrupto e é mais próximo dos dados
originais. Por esse método, as três temperaturas cardinais são calculadas,
TM, To e Tm. A regressão possui cinco parâmetros: (a) TM, (b) Tm, (c) o
coeficiente que descreve a relação curvilinear entre Tm e To, (d) o
coeficiente e (e) o coeficiente que descreve a relação curvilinear entre To
e Tm. Esses cinco parâmetros são empregados para o cálculo do valor de
To. De acordo com os autores, o emprego da função beta descreve com
exatidão vários processos fisiológicos relacionados com o desenvolvimento,
sendo superior aos demais métodos de soma térmica para predizer a
floração do arroz.
Há, portanto, vários modelos que podem ser empregados nos
estudos relacionados com o efeito da temperatura sobre a taxa de
desenvolvimento. Há vantagens e desvantagens no emprego de cada um
deles, de modo que o melhor modelo não é sempre o que mais se aproxima
da realidade, que geralmente contém maior número de parâmetros, mas
sim aquele que, nas circunstâncias possíveis do trabalho executado, explica
suficientemente o fenômeno.

43
3 MATERIAL E MÉTODOS
O trabalho foi desenvolvido no Laboratório de Sementes da
Faculdade de Agronomia e Medicina Veterinária (FAMEV), vinculado à
Universidade Federal de Mato Grosso (UFMT), no período de setembro de
2007 a fevereiro de 2008, com as espécies vegetais angico vermelho
(Anadenanthera macrocarpa (Benth.) Brenan), cerejeira (Amburana
cearensis (Alemão) A.C. Smith), sete-cascas Samanea tubulosa (Benth.)
Barneby & J.W. Grimes, sucupira preta (Bowdichia virgilioides Kunt) e
tarumarana (Buchenavia tomentosa Eichler). O angico vermelho, a sucupira
preta e a tarumarana são características do bioma Cerrado no estado de
Mato Grosso, a cerejeira encontra-se distribuída no Estado de Mato Grosso
nos biomas Cerrado e Amazônia, e a sete-cascas tem ocorrência no
Pantanal e no bioma Amazônico.
As sementes de A. macrocarpa foram coletadas no Campus da
FAMEV, em julho de 2007. As sementes de A. cearensis foram obtidas de
quatro árvores matrizes do Campus da FAMEV, em setembro de 2007. As
sementes de S. tubulosa foram obtidas de frutos recém caídos de uma
árvore matriz do Parque Beripoconé, Poconé, Mato Grosso, em setembro
de 2007 e uma matriz do Campus da Universidade Federal de Mato Grosso.
As sementes de B. virgilioides foram obtidas de frutos recém caídos
de três árvores matrizes do Campus da FAMEV, em setembro de 2007.

44
Os frutos recém caídos de B. tomentosa foram obtidos de duas
árvores matrizes do Campus da FAMEV, em junho de 2007. Após a coleta,
as sementes foram levadas ao laboratório, onde a polpa dos frutos foi
removida por meio de atrito manual contra uma peneira. As sementes foram
lavadas em água corrente e secas à sombra durante 24 horas.
Para angico vermelho, cerejeira, sete-cascas e sucupira preta, após
a coleta, os frutos foram levados ao laboratório para retirada das sementes
das vagens e após as sementes foram homogeneizadas manualmente.
Em seguida as sementes de todas as espécies foram submetidas a
análise do teor de água(base do peso seco), pelo método de estufa a 105°C
± 3°C por 24 horas, sendo também determinada a massa de mil sementes,
utilizando-se oito subamostras de 100 sementes (Brasil, 1992) e após foram
acondicionadas em sacos de papel para serem armazenadas em câmara
refrigerada à temperatura de 18ºC ± 2ºC e 75% ± 4% de umidade relativa,
até o momento da instalação dos experimentos.
No momento da semeadura, para evitar incidência de fungos, as
sementes de todos os tratamentos foram previamente tratadas com imersão
em solução de hipoclorito de sódio (2%) durante cinco minutos.
As sementes de sete-cascas foram escarificadas com ácido sulfúrico
(98% p.a.) por 5 minutos e as de sucupira preta por 8 minutos (Albuquerque
et al., 2007), ambos tratamentos para quebra de dormência.
Para o teste de germinação de sete-cascas, sucupira preta e
cerejeira foi utilizado o substrato papel em forma de rolo (germitest),
utilizando quatro subamostras de 25 sementes e quatro repetições. Os
substratos foram umedecidos com água destilada a 2,5 vezes a massa do
papel seco (Brasil, 1992), exceto para o angico vermelho que foi utilizado
2,0 vezes a massa do papel. Os rolos de papel com as sementes foram
acondicionados em sacos de plásticos transparentes para manutenção da
umidade e mantidos em câmara de germinação até término do experimento.
Sempre que necessário foi feito o reumedecimento do papel com água
destilada.

45
Para tarumarana, foi utilizado o substrato vermiculita na forma entre
vermiculita em bandejas de plástico. A vermiculita foi previamente
esterilizada em autoclave a 121°C e pressão de 0,1MPa e umedecida com
água destilada na quantidade 2,5 vezes a massa do substrato (Dignart,
1998). Cada tratamento foi constituído de quatro subamostras de 20
sementes. As sementes foram previamente tratadas com fungicida
carboxina+tiram (200g/L + 200 g/L) na dosagem de 0,25 mL de produto por
grama de semente, antes da semeadura.
As sementes de todas as espécies foram colocadas para germinar
em câmaras de germinação reguladas nas temperaturas de 10, 15, 20, 25,
30, 35 e 40ºC e fotoperiodo de 12 horas até o máximo de 50 dias em
delineamento inteiramente casualizado.
3.1 Variáveis Analisadas
Foram analisadas as seguintes variáveis: (a) porcentagem de
germinação, (b) índice de velocidade de germinação, (c) tempo para
alcançar a mediana de germinação, (d) porcentagem de plântulas normais,
(e) índice de velocidade de formação de plântulas normais, (f) tempo para
alcançar a mediana da formação de plântulas normais.
As avaliações foram realizadas diariamente, após a semeadura das
espécies durante 50 dias.
A porcentagem de germinação das sementes foi obtida diariamente
(porcentagem acumulada) a partir da contagem do número de sementes
que germinaram em relação ao número total de sementes postas para
germinar (Equação 2). Considerou-se semente germinada aquela que
apresentou raiz com comprimento maior ou igual a 2 mm.
100N
nG (Equação 2)
Em que: G = porcentagem de germinação,

46
n =número de sementes germinadas,
N = número total de sementes postas a germinar.
O índice de velocidade de germinação foi obtido segundo o método
proposto por Timson (1965) e, nesse caso considerou-se um tempo de
observação de 50 dias (Equação 3).
100
... 5050221150
N
pnpnpn (Equação 3)
Em que: 50 = índice de velocidade de germinação,
n1 = número de sementes germinadas no primeiro dia,
p1 = valor atribuído às sementes germinadas no primeiro dia,
n2 = número de sementes germinadas no segundo dia,
p2 = valor atribuído às sementes germinadas no segundo dia,
n50 = número de sementes germinadas no quinquagésimo dia,
p50 = valor atribuído às sementes germinadas no
quinquagésimo dia,
N = número de sementes totais utilizadas.
Os valores atribuídos aos dias foram iguais a 50, para o primeiro dia,
49, para o segundo dia e assim consecutivamente até o qüinquagésimo dia
em que o seu valor foi igual a 1.
O tempo necessário para alcançar a mediana da germinação foi
obtido empregando-se duas aproximações, em função da dinâmica do
processo germinativo (Steinmaus et al., 2000). Quando os valores da
porcentagem de germinação acumulada diária apresentaram tendência
sigmóide significativa em função da temperatura, o valor do tempo
necessário para alcançar a mediana da germinação foi obtido pela solução
da Equação 4 para 50% de germinação. Quando pela análise da tendência
dos dados originais verificou-se que o ajuste da curva sigmóide mostrou-se
não adequado (analisado pelo R2, pela significância do teste de F do
modelo e pela significância dos parâmetros), o valor do tempo necessário
para alcançar a mediana da germinação foi obtido por interpolação linear

47
entre os valores imediatamente inferiores e superiores à porcentagem de
germinação de 50%.
c
bT
a%G
exp1
(Equação 4)
Em que: %G = porcentagem de germinação acumulada observada no tempo T (%), a = estimativa assintótica da porcentagem máxima de germinação alcançada (%), T = tempo (dias), b = constantes de ajustamento que se refere ao tempo em que ocorre a porcentagem de germinação igual ao valor de a (dias),
c = constante de ajustamento referente à concavidade da curva.
Ressalta-se que a escolha da Equação 4 recaiu no fato de ser a mais
simples equação em forma sigmóide que foi encontrada a partir de uma
análise de modelos empregando o programa TableCurve 2D v 2.03. As
equações de formato sigmóide expressam vários fenômenos biológicos
relacionados com o desenvolvimento (Motulsky e Christopoulos, 2004).
No caso de formação de plântulas normais, todas as variáveis foram
calculadas igualmente como explicadas para a germinação das sementes.
Nas contagens, foram consideradas plântulas normais, plântulas com
estruturas essenciais (raíz, epicótilo, hipocótilo) bem diferenciadas, sadias e
desenvolvidas e plântulas anormais, as que apresentavam qualquer uma
das suas estruturas essenciais ausentes, deformadas, muito danificadas ou
infectadas por patógenos da própria semente (Krzyzanowski et al. 1991;
Brasil, 1992).

48
3.2 Determinação das Temperaturas Cardinais para a Germinação e a
Formação de Plântulas
As temperaturas cardinais (temperatura mínima, temperatura ótima e
temperatura máxima) foram determinadas utilizando-se o modelo bilinear
(Steinmaus et al., 2000), após uma tentativa preliminar com emprego da
função beta que não se ajustou aos dados originais.
Essas temperaturas cardinais foram obtidas para o processo
germinativo e para formação de plântulas empregando-se duas
aproximações, a primeira considerando o inverso do tempo para alcançar
50% de germinação ou de formação de plântula (taxa de desenvolvimento)
e a segunda, considerando o índice de velocidade (50) de germinação ou
de formação de plântula.
As regressões foram obtidas utilizando-se a rotina “piecewise
polynomial”, selecionando-se a opção linear-linear, do programa NCSS
(Hintze, 2004). Todas as repetições de cada tratamento foram empregadas
no processo analítico.
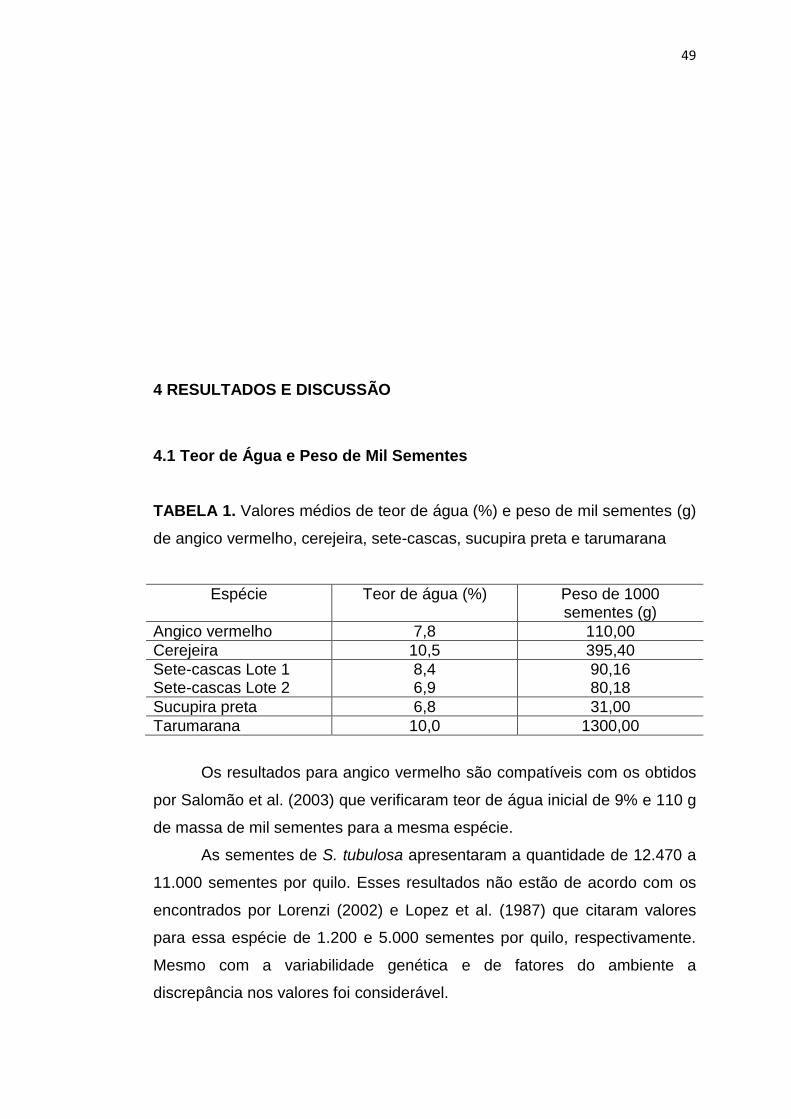
49
4 RESULTADOS E DISCUSSÃO
4.1 Teor de Água e Peso de Mil Sementes
TABELA 1. Valores médios de teor de água (%) e peso de mil sementes (g)
de angico vermelho, cerejeira, sete-cascas, sucupira preta e tarumarana
Espécie Teor de água (%) Peso de 1000 sementes (g)
Angico vermelho 7,8 110,00
Cerejeira 10,5 395,40
Sete-cascas Lote 1 Sete-cascas Lote 2
8,4 6,9
90,16 80,18
Sucupira preta 6,8 31,00
Tarumarana 10,0 1300,00
Os resultados para angico vermelho são compatíveis com os obtidos
por Salomão et al. (2003) que verificaram teor de água inicial de 9% e 110 g
de massa de mil sementes para a mesma espécie.
As sementes de S. tubulosa apresentaram a quantidade de 12.470 a
11.000 sementes por quilo. Esses resultados não estão de acordo com os
encontrados por Lorenzi (2002) e Lopez et al. (1987) que citaram valores
para essa espécie de 1.200 e 5.000 sementes por quilo, respectivamente.
Mesmo com a variabilidade genética e de fatores do ambiente a
discrepância nos valores foi considerável.

50
O resultado referente ao teor de água inicial das sementes de
sucupira preta diferiram dos estudos de Salomão et al. (2003), que
obtiveram 11% de teor de água inicial para a mesma espécie. Ferronato
(1999) verificou a massa de 25 g para a mesma espécie. Essa variabilidade
pode ocorrer em função das condições de temperatura e umidade do local
de coleta e tempo de armazenamento. Também Carvalho e Nakagawa
(2000) citaram que, as sementes, assim como outras partes da planta,
apresentam composição química bastante variável, em função da etapa de
desenvolvimento em que elas se encontram ou devido à influência de
fatores externos, o que pode explicar essa diferença no valores de teor de
água.
O grau de umidade inicial das sementes de tarumarana foi superior
ao valor encontrado no estudo de Salomão et al. (2003) que obtiveram teor
de água inicial de 5,4% para a mesma espécie. A massa mil sementes
diferiu dos estudos de Salomão et al. (2003) com a mesma espécie(2.800g).
Devido à rápida redução no teor de água durante a fase final da
maturação, a maioria das sementes apresenta de 5 a 20% de água da sua
massa total (Bradbeer, 1988). Neste trabalho, o teor de água das sementes
investigadas estava dentro dessa faixa, variando de 6,8 a 10,5%. De acordo
com Bewley e Black (1994), sementes ortodoxas mantêm baixo índice de
umidade a fim de permanecerem viáveis por longo período, germinando
apenas quando as condições ambientais forem favoráveis à sobrevivência
da plântula.
Não houve oscilação entre os teores iniciais de água e no momento
de montagem dos experimentos devido ao curto período de armazenamento
entre a coleta e a montagem do experimento.

51
4. 2 Avaliação da Dinâmica da Germinação e Formação de Plântulas
para os Diferentes Tratamentos Térmicos
As espécies apresentaram uma dinâmica particular de germinação
em função do tratamento térmico a que foram submetidas. Verificou-se o
efeito da temperatura tanto sobre o tempo em que se deu a protrusão da
raiz primária quanto sobre a variabilidade de resposta das repetições
(Figuras 13 a 26). Nas temperaturas mais baixas ocorreu o atraso do
processo germinativo, enquanto, a elevação da temperatura permitiu que as
sementes germinassem mais rapidamente. O processo de formação de
plântulas normais foi mais afetado pela temperatura que a germinação.
Pela análise de variância não paramétrica da dinâmica de
germinação, a porcentagem máxima de germinação não foi afetada pela
temperatura, entretanto as espécies tiveram efeito significativo (Tabela
A2.1). Para a porcentagem de formação de plântulas normais, a espécie e a
temperatura influenciaram significativamente a 6% e 4% de probabilidade,
respectivamente (Tabela A2.2).
A porcentagem máxima de germinação das sementes do angico
vermelho foi de 99%, sendo obtida no terceiro dia à temperatura de 15°C,
no segundo dia à 30°C e no primeiro dia quando as sementes foram
submetidas a 35°C. Para a cerejeira, esse valor máximo foi de 96% obtido
aos cinco dias sob temperatura de 30°C. Para a sete-cascas foi 99% obtida
aos dez dias para a temperatura de 10°C e aos seis dias para a de 15°C.
Para a sucupira preta, ocorreu 95% de germinação aos quatorze dias à
20°C e aos cinco dias a 30°C. Para tarumarana, ocorreu 84% de
germinação obtida aos vinte dias à 25°C. Observou-se que a tarumarana foi
a espécie mais sensível às variações de temperatura para germinação na
faixa de 10 a 40°C. As sementes de angico vermelho e da sete-cascas,
espécies de ocorrência em biomas diferentes, apresentaram uma notável
tolerância às baixas temperaturas em comparação com sucupira preta e
tarumarana. A cerejeira teve resposta intermediária para temperaturas
baixas com porcentagem de germinação de 48% após 30 dias na

52
temperatura de 10°C. As sementes de sucupira preta e de tarumarana não
germinaram na temperatura de 10°C (Figura 13).
Comparando a germinação das cinco espécies, nas temperaturas de
25, 30 e 35°C, verificou-se que o angico vermelho teve a germinação mais
homogênea com menor tempo em dias para emissão de raiz, e a
tarumarana foi a espécie com germinação mais heterogênea, com maior
tempo para protrusão de raiz primária (Figuras 16 a 18).
A temperatura influencia no tempo para o início da germinação, no
tempo médio e na porcentagem de germinação da maioria das sementes,
modificando a velocidade das reações químicas que irão acionar o
desdobramento, o transporte das reservas e a ressíntese de substâncias
para a plântula (Baskin e Baskin 1988; Bewley e Black 1994). Quanto maior
a faixa de temperatura para germinação, mais ampla é a distribuição
geográfica da espécie que está em estudo (Labouriau, 1983) e o tempo
médio de germinação é importante para avaliar a rapidez de ocupação de
uma espécie em determinado ambiente (Ferreira et al., 2001).
Nas temperaturas de 10, 20 e 25°C, germinaram 97% das sementes
de angico vermelho e 94% quando a 40°C (Figuras 13, 15,16 e 19). A
temperatura não afetou a porcentagem de germinação acumulada de
angico vermelho no período avaliado (Figuras 13 a 19), mas afetou a
porcentagem acumulada da formação de plântulas normais, nas
temperaturas de 10, 15 e 40°C (Figuras 20, 21 e 26).
Para cerejeira, a porcentagem de germinação acumulada aumentou
em função do aumento da temperatura. A 35°C verificou-se aumento e a
40°C, redução do tempo para atingir a germinação máxima, mas com
redução drástica da germinação nessa ultima temperatura. A 35°C foi
verificado aumento do tempo e redução para 69% a formação de plântulas
normais. Para sete-cascas, a porcentagem máxima de germinação (98%) e
o menor tempo ocorreu a 40°C, porém não houve formação de plântulas. Na
temperatura de 35°C ocorreu a porcentagem máxima de formação de
plântulas, entretanto não foi a que apresentou o melhor desempenho para
germinação. A porcentagem acumulada de germinação e a porcentagem de

53
formação de plântulas de sucupira preta tiveram melhor desempenho a 30°C
com 95% e 86%, reduzindo a 35°C para 76% e 71% respectivamente no
período de oito dias após a semeadura. Para tarumarana, a temperatura de
25°C foi onde se obteve a máxima germinabilidade (84%) em sete dias,
entretanto para formação de plântulas, a porcentagem máxima (81%)
ocorreu num período de 25 dias enquanto que a 30°C, verificou-se 80% num
período menor (21 dias).
0
20
40
60
80
100
120
Ge
rmin
açã
o a
cu
mu
lad
a
(%)
0
20
40
60
80
100
120
Tempo (dias)
0 10 20 30 40 50 60
0
20
40
60
80
100
120
0 10 20 30 40 50 60
A
B C
D E
FIGURA 13. Germinação acumulada (%) de sementes de angico vermelho
(A), cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E),
submetidas à temperatura de 10°C.

54
0
20
40
60
80
100
120
Ge
rmin
açã
o a
cu
mu
lad
a
(%)
0
20
40
60
80
100
120
Tempo (dias)
0 10 20 30 40 50 60
0
20
40
60
80
100
120
0 10 20 30 40 50 60
A
B C
D E
FIGURA 14. Germinação acumulada (%) de sementes de angico vermelho
(A), cerejeira (B), sete-cascas (C), sucupira preta (D) e
tarumarana (E), submetidas à temperatura de 15°C.
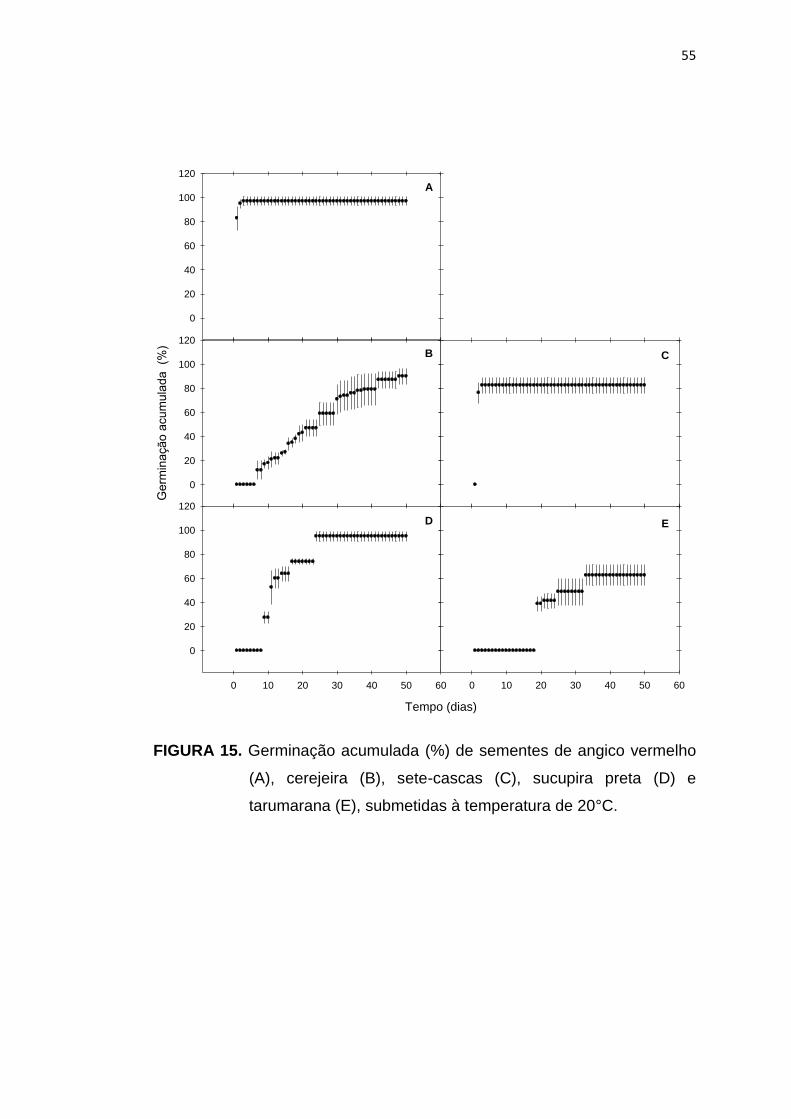
55
0
20
40
60
80
100
120
Ge
rmin
açã
o a
cu
mu
lad
a
(%)
0
20
40
60
80
100
120
Tempo (dias)
0 10 20 30 40 50 60
0
20
40
60
80
100
120
0 10 20 30 40 50 60
A
B C
D E
FIGURA 15. Germinação acumulada (%) de sementes de angico vermelho
(A), cerejeira (B), sete-cascas (C), sucupira preta (D) e
tarumarana (E), submetidas à temperatura de 20°C.

56
0
20
40
60
80
100
120
Ge
rmin
açã
o a
cu
mu
lad
a
(%)
0
20
40
60
80
100
120
Tempo (dias)
0 10 20 30 40 50 60
0
20
40
60
80
100
120
0 10 20 30 40 50 60
A
B C
D E
FIGURA 16. Germinação acumulada (%) de sementes de angico vermelho
(A), cerejeira (B), sete-cascas (C), sucupira preta (D) e
tarumarana (E), submetidas à temperatura de 25°C.
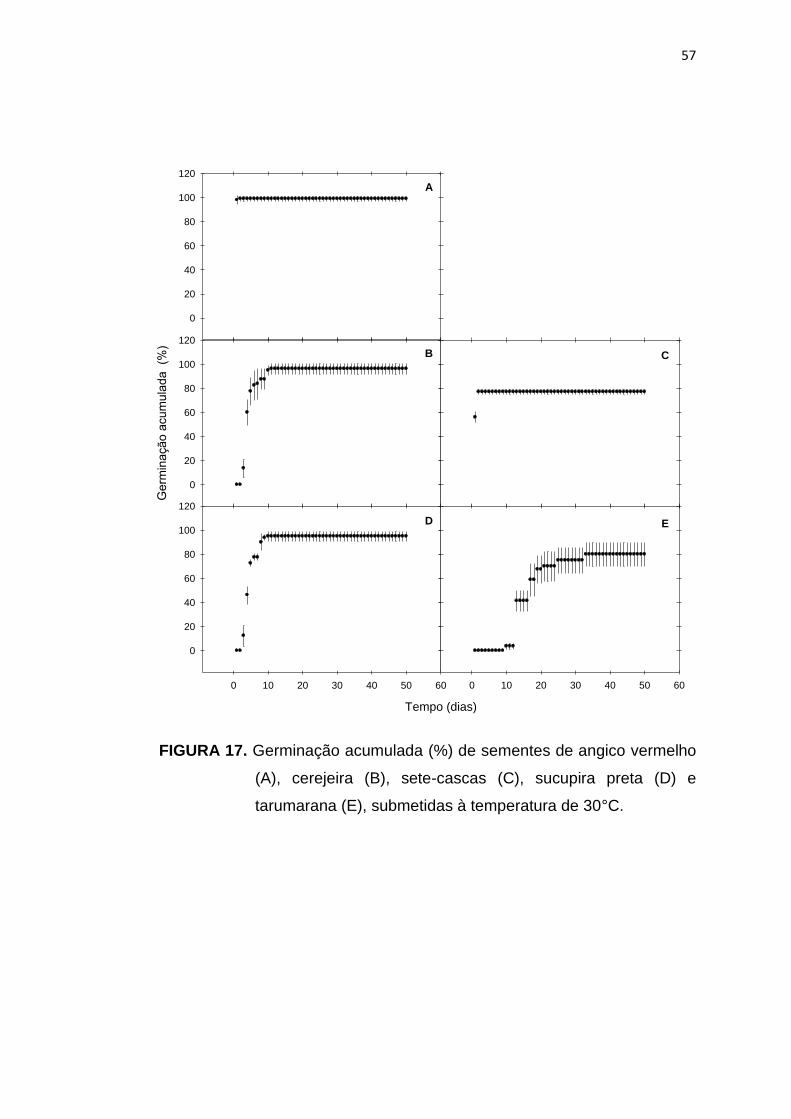
57
0
20
40
60
80
100
120
Ge
rmin
açã
o a
cu
mu
lad
a
(%)
0
20
40
60
80
100
120
Tempo (dias)
0 10 20 30 40 50 60
0
20
40
60
80
100
120
0 10 20 30 40 50 60
A
B C
D E
FIGURA 17. Germinação acumulada (%) de sementes de angico vermelho
(A), cerejeira (B), sete-cascas (C), sucupira preta (D) e
tarumarana (E), submetidas à temperatura de 30°C.
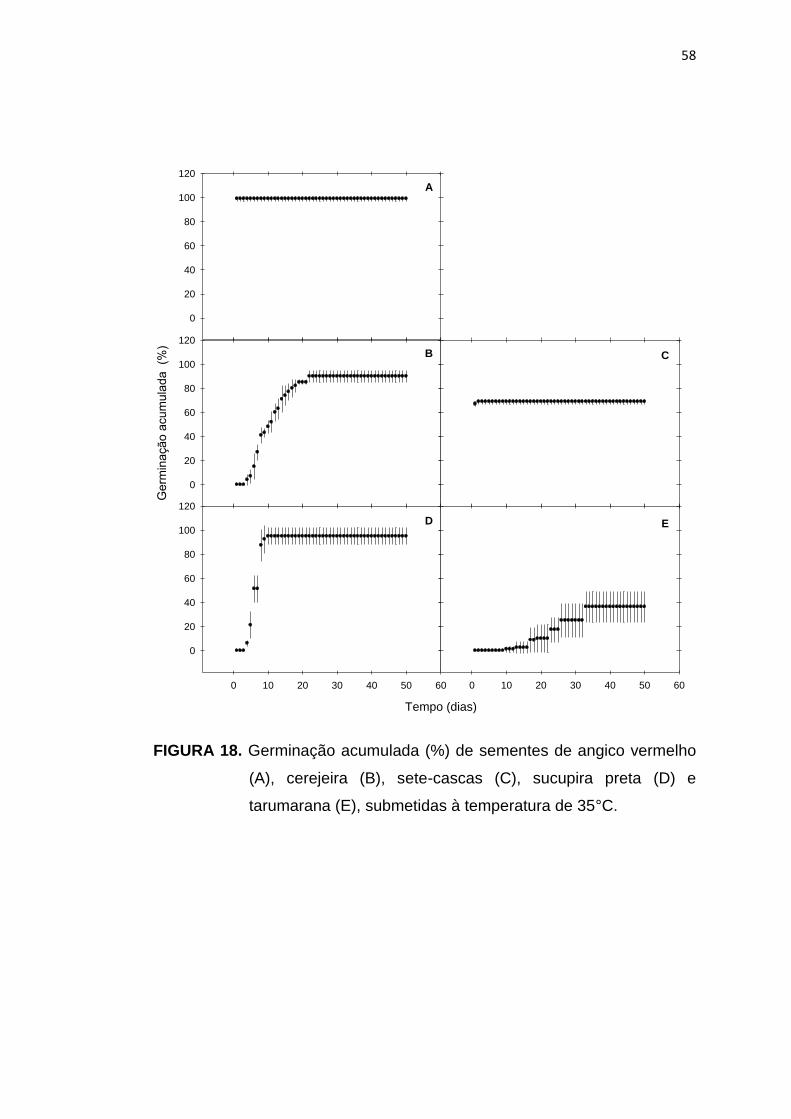
58
0
20
40
60
80
100
120
Ge
rmin
açã
o a
cu
mu
lad
a
(%)
0
20
40
60
80
100
120
Tempo (dias)
0 10 20 30 40 50 60
0
20
40
60
80
100
120
0 10 20 30 40 50 60
A
B C
D E
FIGURA 18. Germinação acumulada (%) de sementes de angico vermelho
(A), cerejeira (B), sete-cascas (C), sucupira preta (D) e
tarumarana (E), submetidas à temperatura de 35°C.
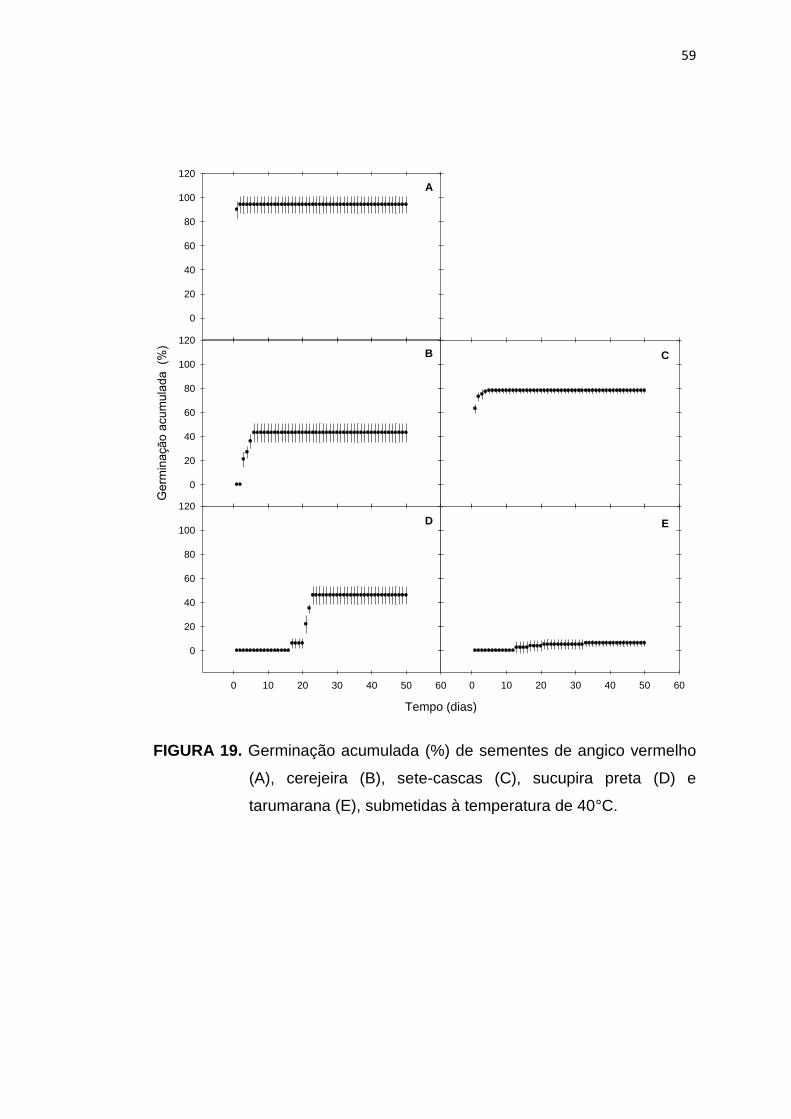
59
0
20
40
60
80
100
120
Ge
rmin
açã
o a
cu
mu
lad
a
(%)
0
20
40
60
80
100
120
Tempo (dias)
0 10 20 30 40 50 60
0
20
40
60
80
100
120
0 10 20 30 40 50 60
A
B C
D E
FIGURA 19. Germinação acumulada (%) de sementes de angico vermelho
(A), cerejeira (B), sete-cascas (C), sucupira preta (D) e
tarumarana (E), submetidas à temperatura de 40°C.

60
0
20
40
60
80
100
120
Po
rce
nta
ge
m a
cu
mu
lad
a d
e f
orm
açã
o d
e p
lân
tula
s
0
20
40
60
80
100
120
Tempo (dias)
0 10 20 30 40 50 60
0
20
40
60
80
100
120
0 10 20 30 40 50 60
A
B C
D E
FIGURA 20. Porcentagem acumulada de formação de plântulas normais de
angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira
preta (D) e tarumarana (E), submetidas à temperatura de
10°C.
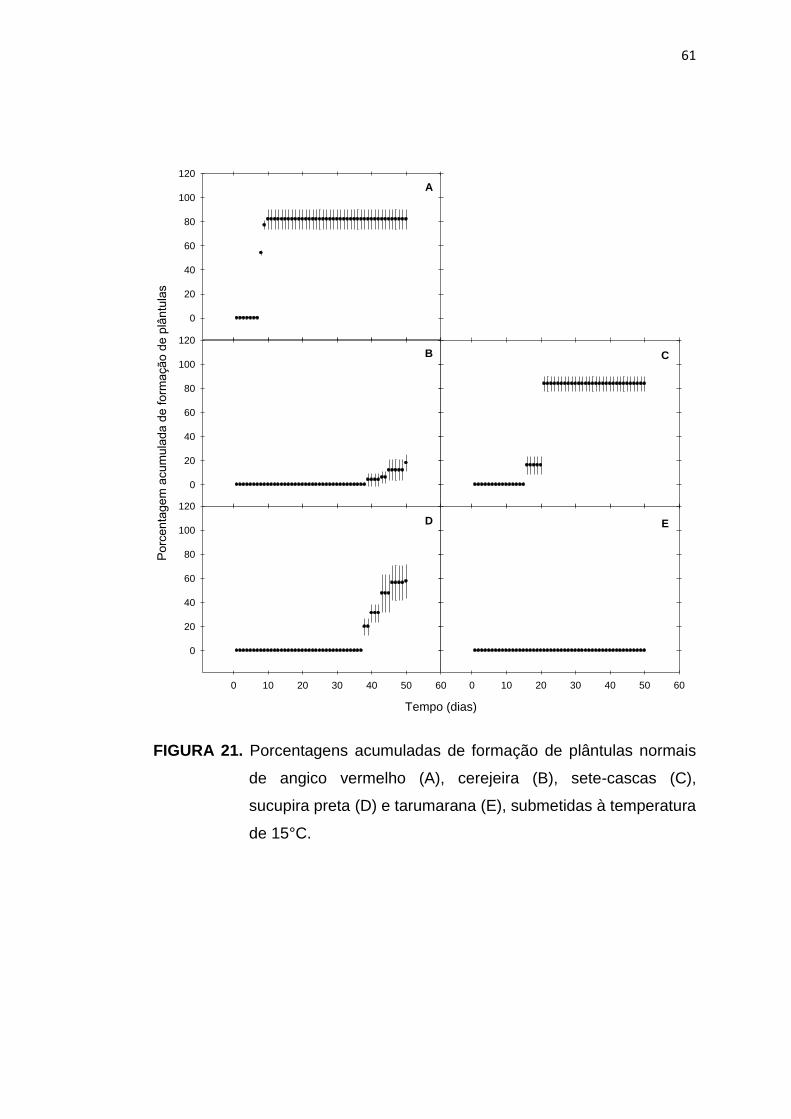
61
0
20
40
60
80
100
120
Po
rce
nta
ge
m a
cu
mu
lad
a d
e f
orm
açã
o d
e p
lân
tula
s
0
20
40
60
80
100
120
Tempo (dias)
0 10 20 30 40 50 60
0
20
40
60
80
100
120
0 10 20 30 40 50 60
A
B C
D E
FIGURA 21. Porcentagens acumuladas de formação de plântulas normais
de angico vermelho (A), cerejeira (B), sete-cascas (C),
sucupira preta (D) e tarumarana (E), submetidas à temperatura
de 15°C.

62
0
20
40
60
80
100
120
Po
rce
nta
ge
m a
cu
mu
lad
a d
e f
orm
açã
o d
e p
lân
tula
s
0
20
40
60
80
100
120
Tempo (dias)
0 10 20 30 40 50 60
0
20
40
60
80
100
120
0 10 20 30 40 50 60
A
B C
D E
FIGURA 22. Porcentagens acumuladas de formação de plântulas normais
de angico vermelho (A), cerejeira (B), sete-cascas (C),
sucupira preta (D) e tarumarana (E), submetidas à temperatura
de 20°C.
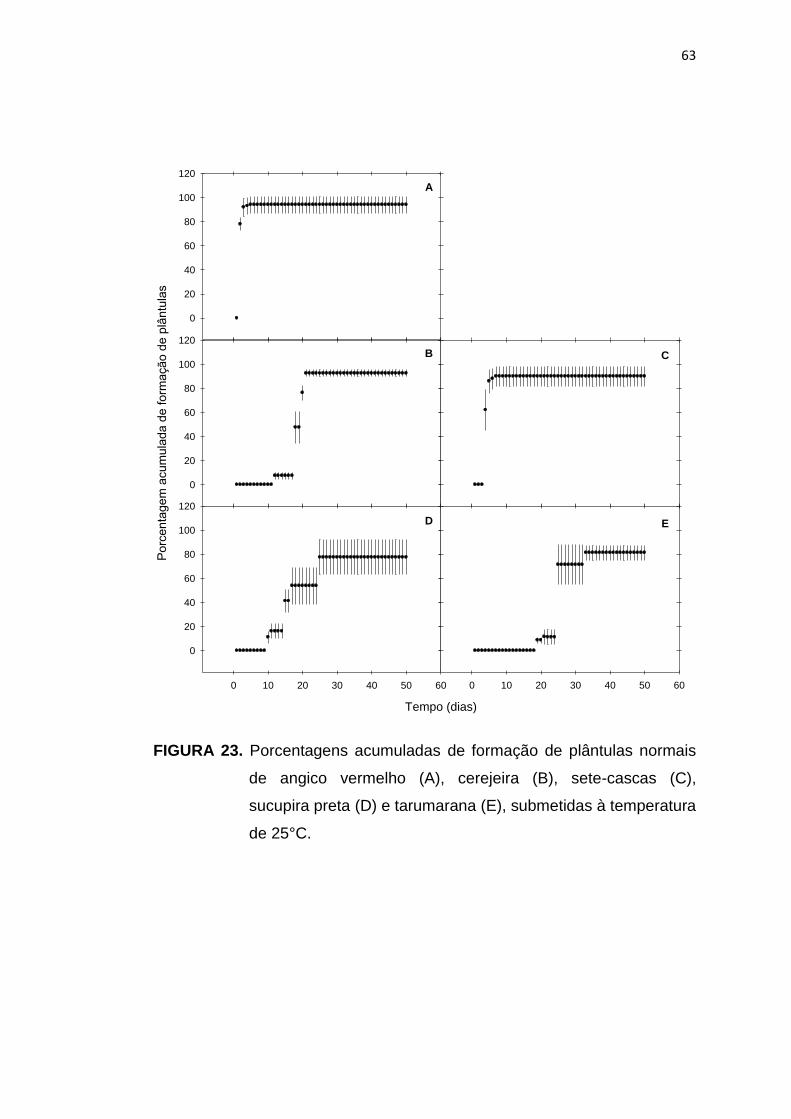
63
0
20
40
60
80
100
120
Po
rce
nta
ge
m a
cu
mu
lad
a d
e f
orm
açã
o d
e p
lân
tula
s
0
20
40
60
80
100
120
Tempo (dias)
0 10 20 30 40 50 60
0
20
40
60
80
100
120
0 10 20 30 40 50 60
A
B C
D E
FIGURA 23. Porcentagens acumuladas de formação de plântulas normais
de angico vermelho (A), cerejeira (B), sete-cascas (C),
sucupira preta (D) e tarumarana (E), submetidas à temperatura
de 25°C.
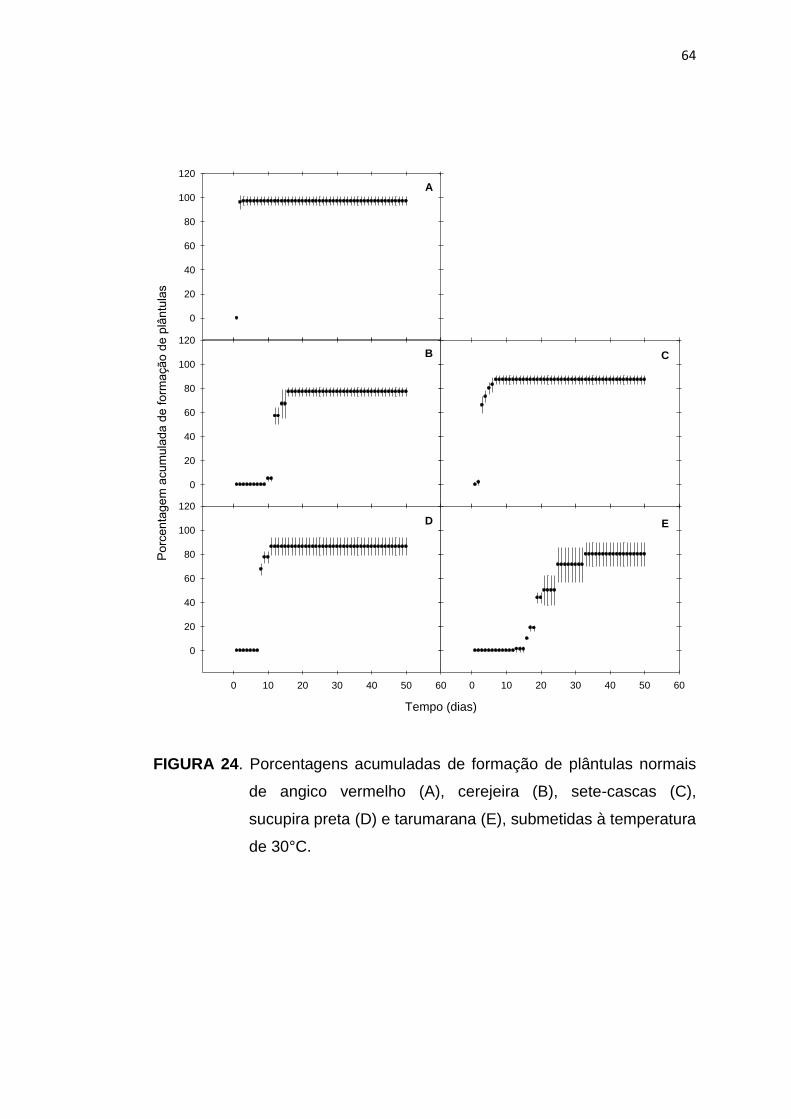
64
0
20
40
60
80
100
120
Po
rce
nta
ge
m a
cu
mu
lad
a d
e f
orm
açã
o d
e p
lân
tula
s
0
20
40
60
80
100
120
Tempo (dias)
0 10 20 30 40 50 60
0
20
40
60
80
100
120
0 10 20 30 40 50 60
A
B C
D E
FIGURA 24. Porcentagens acumuladas de formação de plântulas normais
de angico vermelho (A), cerejeira (B), sete-cascas (C),
sucupira preta (D) e tarumarana (E), submetidas à temperatura
de 30°C.
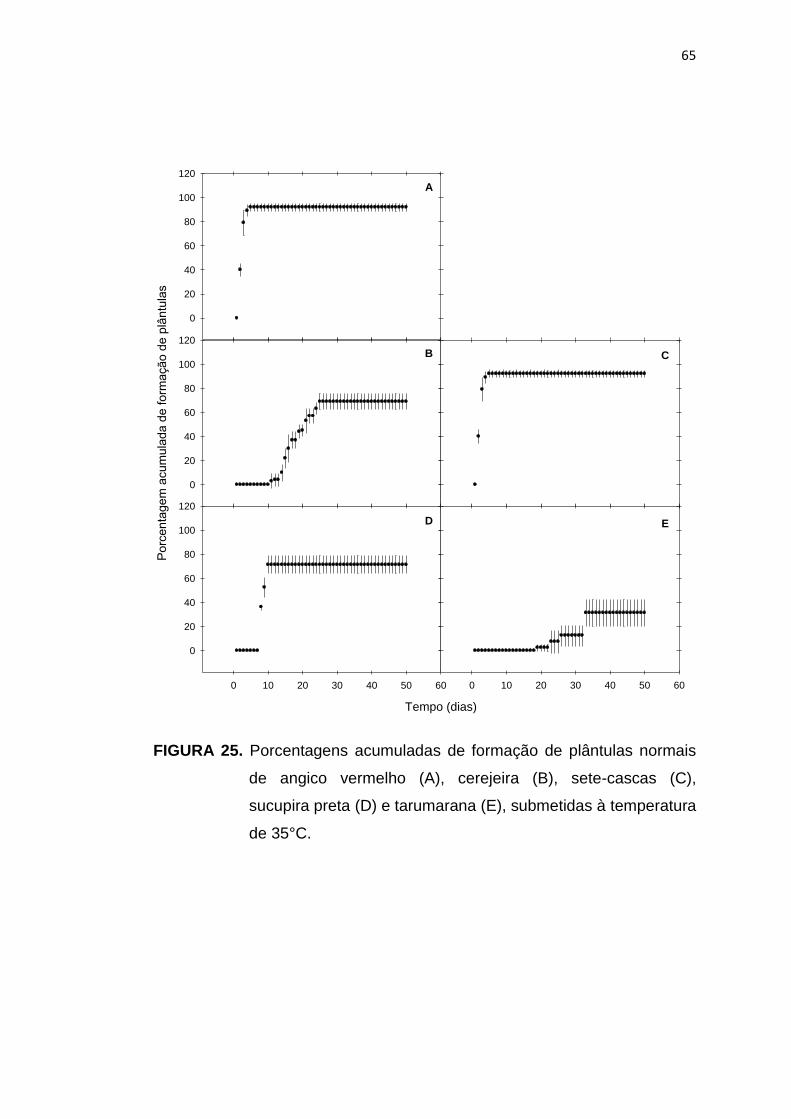
65
0
20
40
60
80
100
120
Po
rce
nta
ge
m a
cu
mu
lad
a d
e f
orm
açã
o d
e p
lân
tula
s
0
20
40
60
80
100
120
Tempo (dias)
0 10 20 30 40 50 60
0
20
40
60
80
100
120
0 10 20 30 40 50 60
A
B C
D E
FIGURA 25. Porcentagens acumuladas de formação de plântulas normais
de angico vermelho (A), cerejeira (B), sete-cascas (C),
sucupira preta (D) e tarumarana (E), submetidas à temperatura
de 35°C.
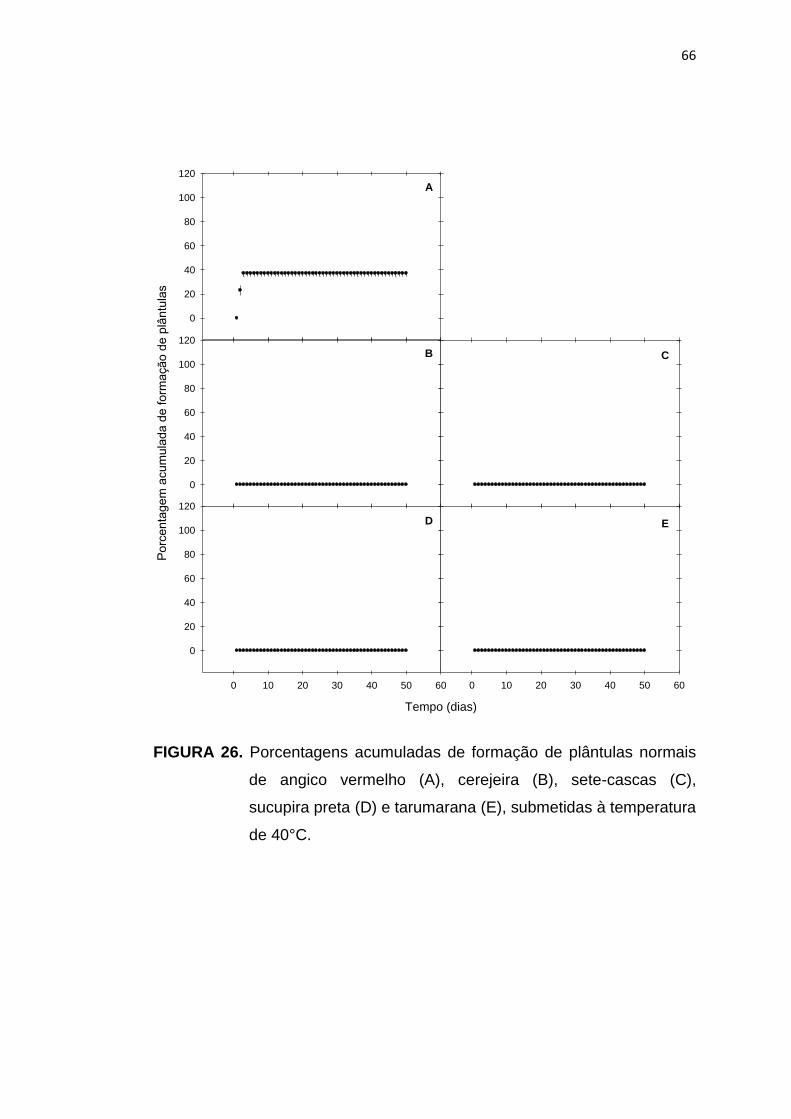
66
0
20
40
60
80
100
120
Po
rce
nta
ge
m a
cu
mu
lad
a d
e f
orm
açã
o d
e p
lân
tula
s
0
20
40
60
80
100
120
Tempo (dias)
0 10 20 30 40 50 60
0
20
40
60
80
100
120
0 10 20 30 40 50 60
A
B C
D E
FIGURA 26. Porcentagens acumuladas de formação de plântulas normais
de angico vermelho (A), cerejeira (B), sete-cascas (C),
sucupira preta (D) e tarumarana (E), submetidas à temperatura
de 40°C.

67
Foi observado 80% de plântulas normais fracas de angico vermelho
na temperatura de 10°C. Os restantes das plântulas, em fase de hipocótilo,
estavam mortas pelo efeito da baixa temperatura num tempo médio de 21
dias. Para 15°C, obteve-se 82% em nove dias, mas plântulas normais
fracas. Na temperatura de 20°C, formaram 93% de plântulas normais aos
cinco dias, porém com raízes menores e com desenvolvimento lento
quando comparado com o ocorrido na temperatura ótima (Figura 20A). O
maior desempenho de plântulas normais formadas (97%) ocorreu na
temperatura de 30°C num tempo médio de dois dias quando comparado
com as demais temperaturas (Figuras 20A e 24A).
Para o angico vermelho, as porcentagens de formação de plântulas
foram relativamente altas em todas as temperaturas exceto à temperatura
de 40°C com percentual de 37% de plântulas normais. Nessa temperatura
ocorreu a morte de plântulas e sementes de angico vermelho devido ao
longo tempo de exposição. A raiz apresentava sintoma de necrose nas
pontas (Figura 27). Nessa temperatura, a espécie apresentou a menor taxa
de germinação, devido ao efeito negativo de altas temperaturas que,
provavelmente, diminuiu o suprimento de aminoácidos livres, a síntese de
RNA e, conseqüentemente, de proteínas. A diminuição da produção de
enzimas ou mesmo, sua ausência conduz ao decréscimo ou inexistência de
reações metabólicas que podem retardar ou suprimir a germinação.
Alterações na membrana celular das sementes de várias espécies,
ocasionadas por diferentes temperaturas, foram relatadas por Hendricks e
Taylorson (1976) e foram provenientes de altas temperaturas, com o
aumento no efluxo de aminoácidos durante a germinação e decréscimo da
capacidade germinativa das sementes.
O angico vermelho é adaptado às condições de maior insolação e
baixa precipitação pluviométrica, com deficiência hídrica no inverno, comum
nessa região de distribuição do Cerrado. A alta porcentagem de germinação
e formação de plântulas na faixa ampla de temperaturas que a espécie foi
submetida refletiu a adaptação da espécie ao clima da região do Cerrado e

68
demonstrou a sua importância num programa de recuperação de áreas
degradadas que necessita de rápida colonização das espécies em
ambientes perturbados. No entanto esse rápido estabelecimento em função
de seus aspectos fisiológicos pode ser prejudicado se as condições do
ambiente não forem favoráveis devido a alta velocidade de germinação em
curto período de tempo. Por exemplo, condições de temperatura favorável
podem permitir a germinação se houver a disponibilidade de água,
entretanto, numa mudança súbita nas condições do ambiente como um
déficit hídrico, pode comprometer o estabelecimento da espécie que teve
germinação rápida e homogênea.
FIGURA 27. Plântulas de angico vermelho (Anadenanthera macrocarpa)
formadas nas temperaturas de 10, 15, 20, 25, 30, 35 e 40°C.
Colli et al. ( 2005) concluíram que a temperatura ótima para angico
vermelho é de 25 a 30°C, enquanto que Mondo et al. (2008) verificaram
para outra espécie de angico Parapiptadenia rígida, que o teste de
germinação deve ser conduzido na temperatura de 25ºC.
Segundo Carvalho (1994), das espécies de angico, a Anadenanthera
macrocarpa é a que apresenta maior abrangência geográfica, ocorrendo
desde o sul da Bolívia até o norte da Argentina; no Brasil, só não aparece
10°C 15°C 20°C 25°C 30°C 35°C 40°C
5 a 11/01/08

69
nos estados da Região Sul, sendo uma espécie comprovadamente
calcícola, de crescimento rápido e tolerante a solos arenosos e rasos e
muito usada para recomposição de matas ciliares.
A velocidade e a porcentagem acumulada para formação de
plântulas de angico vermelho foram mais afetadas pela temperatura que a
germinação, principalmente nas temperaturas sub e supra-ótimas.
O angico vermelho foi a espécie que apresentou a maior plasticidade
para germinação e formação de plântulas em resposta a variação de
temperatura. A plasticidade no desenvolvimento é a tendência dos
indivíduos de variarem com o tempo em resposta a diferentes condições
ambientais, ou a capacidade de responderem de modo distinto a diferentes
estímulos ambientais. Segundo Raven et al. (2001), as evidências de
adaptação ao ambiente físico podem ser vistas nas variações graduais que
acompanham a distribuição geográfica (cline) e nos grupos distintos de
fenótipos (ecotipos) das mesmas espécies que ocupam habitats diferentes.
Em geral, os angicos são encontrados tanto em formações das
capoeiras e florestas secundárias situadas em terrenos arenosos e
cascalhentos, como no interior da mata primária densa de solos argilosos e
férteis, porém sempre em terrenos altos e bem drenados (Lorenzi, 1992).
Algumas vezes a variação fenotípica dentro da mesma espécie acompanha
a distribuição geográfica e pode estar correlacionada com mudanças
graduais na temperatura, umidade ou alguma outra condição ambiental
(Raven et al., 2001). Se a amplitude térmica não foi um fator restritivo para a
germinação dessa espécie, isso indicou que a sua dispersão e ocorrência
foi uma adaptação da espécie ao longo dos anos, que favoreceu a sua
distribuição geográfica com plantas mais ajustadas aos diferentes
ambientes do bioma Cerrado. Essa espécie teve a maior velocidade para
germinação e formação de plântulas nas temperaturas estudadas,
característica típica de espécie pioneira que precisa se estabelecer
rapidamente em ambientes de variações sazonais de clima. De acordo com
Larcher (2000), as plantas também se ajustam à periodicidade sazonal da
radiação e duração do dia, além da temperatura e precipitação.

70
As sementes de cerejeira quando submetidas a temperatura de 10°C,
só atingiram 50% da germinação após 30 dias, mas não houve a formação
de plântulas normais (Figuras 13B e 20B).
Na temperatura de 15°C, houve redução significativa na formação de
plântulas em relação às temperaturas de 25 e 30°C (Figuras 21B, 23B e
24B). Houve formação de 18% de plântulas com as estruturas anormais. As
baixas temperaturas reduzem a velocidade de germinação, alteram a
uniformidade de emergência de plântulas (Carvalho e Nakagawa, 2000) e
afetam potencialmente a capacidade da semente em absorver água,
limitando a atividade de várias vias metabólicas, bem como a da própria
água, conduzindo a uma diminuição geral da velocidade do processo
germinativo (Bewley e Black, 1994). Na temperatura de 20°C, foi constatado
71% de plântulas pequenas, pouco desenvolvidas, com raízes curtas, com
aspecto atrofiado, consideradas normais fracas, quando comparadas com
as formadas na temperatura ótima (Figuras 22B e 28A). Na temperatura de
40°C, obteve-se 43% de germinação, porém as raízes morreram pela
exposição à alta temperatura. As sementes que não germinaram morreram
deterioradas (Figuras 26B e 28B) e não houve formação de plântulas
normais.
FIGURA 28. Plântulas de cerejeira de oriundas de sementes submetidas às
temperaturas de 20°C (A) e 40°C (B).
20°C
40°C
A B

71
Nas temperaturas de 25 e 35°C (Figuras 23B e 25B), ocorreram 93%
e 69% de germinação de sementes de cerejeira, no mesmo tempo de 19
dias, porém as plântulas normais apresentavam-se menores, em média, do
que as plântulas normais na temperatura de 30°C, temperatura essa em
que se obteve o máximo desempenho de formação de plântulas (96%) com
tempo médio de 13 dias (Figura 29). A formação de plântulas foi afetada na
temperatura de 35°C.
Silva Bello et al. (2008) verificaram que a temperatura de 30°C foi a
mais recomendada para condução de teste de germinação para Amburana
acreana. Entretanto, em estudo da mesma espécie, oriundas de Serra
Negra-RN, Guedes et al. (2008) verificaram que a temperatura de 35°C foi a
mais adequada para testes de germinação com essa espécie. Assim,
poderão haver ecotipos da mesma espécie que ao longo do tempo
adaptaram-se às temperaturas médias mais altas no seu local de
ocorrência, sugerindo novas pesquisas para a espécie.
FIGURA 29. Plântulas de cerejeira formadas a 25 e 30°C.
Para Virola surinamensis, espécie do bioma Amazônia, a
temperatura mais adequada para germinação de sementes foi 30°C (Limas
et al., 2007). Conforme Araujo Neto et al. (2003), sementes de monjoleiro
(Acassia polyphylla), tiveram melhor desempenho germinativo na
30°C 25°C

72
temperatura de 25°C. Assim, para espécies florestais subtropicais e
tropicais brasileiras a temperatura ótima de germinação situa-se entre 20 e
30°C, uma vez que essas são temperaturas encontradas em suas regiões
de origem, na época propícia para a germinação natural (Borges e Rena,
1993). Assim, a temperatura ótima para sementes de espécies florestais da
região amazônica como o pau-tanino, o jenipapo, a faveira preta, a
bacabinha e o jucá foi de 30ºC (Miranda e Ferraz, 1999; Nascimento et al.,
2000; Nascimento et al., 2003; Silva et al., 2006a; Lima et al., 2006), de 30 a
35ºC para o angelim pedra (Varela et al., 2005) e 25 a 35°C para o paricá
(Ramos et al., 2006).
Para sete-cascas, na temperatura de 10°C, as sementes atingiram
99% de germinação aos 30 dias, entretanto só formaram 41% de plântulas
normais fracas com tamanho reduzido, com plúmulas, mas com raízes
principais pequenas e sem raízes secundárias. O comprimento de plântulas
nessa temperatura, aos 30 dias, foi em média de 3,0 cm (Figura 30).
FIGURA 30. Plântulas de sete-cascas formadas na temperatura de 10°C.
Na temperatura de 15°C, aos 30 dias, houve 83% de formação de
plântulas normais, que apresentavam plúmulas, trifólios, raiz principal e
início de raízes secundárias, porém havia plântulas normais fracas com
10°C
08/12/07

73
hipocótilo menor que das plântulas normais. As plântulas anormais (15%)
apresentaram hipocótilo muito curto (Figura 31).
Nas temperaturas 25 e 30°C houve a maior taxa de crescimento das
plântulas quando comparadas as sub e supra- ótimas (Figura 32)
FIGURA 31. Plântulas de sete-cascas formadas na temperatura de 15°C.
FIGURA 32. Plântulas de sete-cascas formadas a temperatura de 25, 30°C
e 40°C.
40°C 25°
C
30°C
Plântulas normais
Plântulas normais fracas
Plântula anormal

74
Na temperatura de 40°C (Figura 33), para as sementes de sete-
cascas, houve emissão de raiz primária, mas não o desenvolvimento
subseqüente, impedindo a formação de plântulas normais e favorecendo a
deterioração das sementes. Ocorreu liberação de exsudados no meio
germinativo e desenvolvimento de fungos na superfície das sementes.
Houve atrofiamento da raiz, raiz principal pouco desenvolvida e
engrossamento de hipocótilo. Havia o odor característico de deterioração
pelo extravasamento de solutos da semente
FIGURA 33. Plântulas de sete-cascas formadas a 40°C.
O uso de altas temperaturas no processo germinativo de sementes
ocasiona estresse, proporcionando-lhes inibição térmica, dormência
secundária, ou a perda de viabilidade (Bewley e Black, 1982), e é provável
que a perda da viabilidade seja oriunda do efluxo de solutos, por meio de
alterações metabólicas (Hendricks e Taylorson, 1976).
Socolowski e Takaki (2007) verificaram que as temperaturas
cardinais para germinação das sementes de Tabebuia rosea estão entre 10
e 15°C a mínima e entre 40 e 45°C a temperatura máxima. Stockman et al.,
2007 observaram germinabilidade máxima a 25°C e 30°C para Tabebuia
roseo-alba. Estudos confirmados por Santos et al. (2005) verificaram que as
sementes das três espécies de Tabebuia são indiferentes à luz, com a faixa
de temperatura ótima entre 20°C e 30°C, a temperatura máxima entre 35 e
40°C e a temperatura mínima entre 10 e 15°C para Tabebuia chrysotricha e
Tabebuia roseo-alba e abaixo de 10°C para Tabebuia serratifolia.

75
Observou-se nas temperaturas 25, 30 e 35°C (Figuras 23C, 24C e
25C) maior desempenho na formação de plântulas normais. Confirmando
Wang e Engel, (1998) que afirmaram que a taxa máxima de
desenvolvimento ocorre na temperatura ótima. A máxima porcentagem na
formação de plântulas (92%) ocorreu na temperatura de 35°C com tempo de
três dias, quando comparado com 30°C (87%) e 25°C (90%).
Para sucupira preta, na temperatura 10°C não houve germinação
nem a formação de plântulas normais. Após 40 dias, as sementes foram
colocadas na temperatura de 30°C, e germinaram 90% das sementes,
porém formaram plântulas normais fracas e com menor comprimento,
quando comparadas com sementes sem período de exposição às baixas
temperaturas. Pelos resultados verificou-se que temperaturas mais baixas,
atuaram apenas bloqueando os processos metabólicos e a velocidade de
embebição, confirmando as observações de Hendricks e Taylorson (1976) e
Marcos Filho (2005).
Na temperatura de 15°C, as primeiras raízes emergiram após 15
dias, e ocorreram plântulas normais após 37 dias. As plântulas eram
normais, porém fracas, com desenvolvimento lento, sendo que as raízes e o
hipocótilo eram curtos quando comparados a essas estruturas nas plântulas
formadas em temperaturas ótimas (Figura 34). Observou-se aumento da
porcentagem acumulada de formação de plântulas normais com o aumento
da temperatura até 30°C (Figura 24D), com 86% de plântulas normais,
tendendo a decrescer a partir dessa temperatura. Segundo Albuquerque et
al. (2007a), as temperaturas mais adequadas para a germinação de
sucupira preta (emissão de raiz com 2mm) foram 25°C e alternadas 20-
30°C.
A porcentagem de formação de plântulas normais a 25°C foi de 77%
aos 20 dias da semeadura.

76
FIGURA 34. Plântulas de sucupira preta (Bowdichia virgilioides) formadas a 15°C (A) e 20, 25 e 30°C (B) com 26 dias após semeadura.
A germinação acumulada e a porcentagem de formação de plântulas
normais na temperatura de 35°C foram afetadas quando comparadas com a
temperatura de 30°C (Figuras 24D e 25D). A 35°C ocorreu 71% de plântulas
normais.
Na temperatura de 40°C, 21 dias após a semeadura, 46% das
sementes apresentaram início de protrusão da raiz primária, mas as
mesmas não se desenvolveram e morreram. Embora 3% das sementes
germinadas formaram plântulas, essas eram anormais com raiz e hipocótilo
curtos e engrossados.
Aos 30 dias nessa temperatura, todas as sementes com início de
protrusão de raiz primária e todas as plântulas morreram devido ao efeito da
alta temperatura. Essa temperatura foi letal para a espécie devido ao tempo
de exposição à mesma. Segundo Marcos Filho (2005), em temperaturas
acima de 40°C há inativação de proteínas e desorganização metabólica que
compromete a germinação. De acordo com Larcher (2000), as temperaturas
elevadas aceleram os movimentos das moléculas, tornando mais fracas as
A B
30°C
25°C
20°C
15°C

77
ligações químicas que associam os átomos que formam as moléculas, e as
camadas de lipídios das biomembranas tornam-se mais fluídas, o que
ocasiona um aumento no efluxo de material celular para o meio. Estes
resultados confirmam que as espécies tropicais apresentam tolerância às
temperaturas altas, com limite máximo igual ou superior a 35°C e limite
mínimo geralmente acima de 5°C (Moraes, 2002).
Martins et al. (2008) verificaram que, para sementes de
Stryphnodendron adstringens (barbatimão) as temperaturas constantes de
25, 30 ou 35°C ou alternadas de 20-30°C, foram as mais favoráveis para o
desempenho germinativo das sementes dessa espécie. Valadares e Paula
(2008) estudando a germinação de sementes de Poecilanthe parviflora
verificaram que nas temperaturas de 10°C e 40°C não houve germinação,
na temperatura de 15°C observou-se pequena porcentagem de sementes
com protrusão de raiz primária, não havendo, contudo, o desenvolvimento
de plântulas normais, e a melhor germinação ocorreu nas temperaturas
constantes de 25°C ou 30°C ou alternada de 25-35°C.
Não houve germinação na temperatura de 10°C para sementes de
tarumarana. Na temperatura de 15°C, após 28 dias da semeadura houve
formação de raízes, mas as mesmas eram curtas e apresentaram
desenvolvimento lento. Não houve formação de plântulas nas temperaturas
de 10 e 15°C. Na temperatura de 20°C germinaram 62% das sementes,
mas só formaram 56% de plântulas; essas plântulas formadas eram
normais, porém com raízes atrofiadas, curtas e ocorreu maior número de
plântulas anormais quando comparadas com 30°C (Figuras 15E, 22E e 35).
Na faixa infra-ótima de temperatura, o gradual congelamento dos
tecidos de reserva da semente impede a sua mobilização à medida que se
diminuiu a temperatura de incubação, sendo uma das prováveis causas da
redução ou inibição da germinação (Mayer e Pljakoff-Mayer, 1989).

78
FIGURA 35. Plântulas formadas na temperatura de 15, 20 e 30°C aos 28
dias de semeadura.
Na Figura 36 encontram-se as plântulas de tarumarana formadas sob
a faixa de temperatura de 25 a 35°C, e se observou o desempenho superior
das plântulas na temperatura de 30°C. A 30°C, verificou-se 80% de
formação de plântulas normais, 81% a 25°C e 31% a 35°C.
FIGURA 36. Plântulas de tarumarana formadas nas temperaturas de 25, 30
e 35°C.
De acordo com Godoi e Takaki (2005), a temperatura ótima para
germinação de sementes de Cecropia glaziovi (embaúba) é de 30°C, e pode
25°C 35°C
30°C 30°C
20°C
30°C
15°C

79
ser usada para recuperação de área degradada devido seu rápido
crescimento. Entretanto para Guazuma ulmifolia (mutamba), utilizada para
recuperação de áreas degradadas no Bioma Amazônia, Araújo Neto et
al.(2002) concluíram que as temperaturas observadas de 25 e 30°C
encontram-se dentro da faixa ótima para a germinação das sementes, a
temperatura máxima situa-se entre 35 e 45°C, e a mínima em torno de
10°C.
Na temperatura de 40°C, as sementes morreram, e as sementes
mais vigorosas que emitiram raiz (6%) morreram pelo efeito da alta
temperatura.
Temperaturas acima de 30°C, comuns na região de ocorrência das
espécies em Mato Grosso, ocasionaram redução na germinabilidade e na
formação de plântulas normais. Embora se espere que as melhores
temperaturas para a germinação de uma espécie sejam aquelas que
favorecem seu crescimento e desenvolvimento (Baskin e Baskin, 1998).
4.3 Índice de Velocidade (50) de Germinação das Sementes e da
Formação de Plântulas para os Diferentes Tratamentos Térmicos
Pelos resultados demonstrados anteriormente, acerca da dinâmica
dos processos germinativos das sementes e da formação de plântulas, era
de se esperar que o efeito da temperatura sobre a velocidade desses
processos diferisse entre as espécies em estudo, o que se confirma pela
interação significativa (p = 0,0001) entre esses fatores ( espécie e
temperatura) observada em ambos os casos (Tabelas A2.3 e A2.4).
Esses efeitos diferenciais da temperatura sobre as velocidades de
germinação e de formação de plântulas ficam mais evidentes quando se
observa o gráfico dessas variáveis (Figuras 37 e 38). Observou-se que as
diferentes espécies apresentaram diferença significativa para índices de
velocidade de germinação e de formação de plântulas. Isso em função das
diferenças na composição química, na estrutura do tegumento e no
tamanho das sementes.

80
Em espécies nativas, a germinação paulatina é um atributo
desejável, pois as sementes produzidas em uma só estação apresentam
maior probabilidade de sobrevivência em condições adversas de ambientes
intermitentes.
A sucupira preta é uma planta pioneira e adaptada a terrenos secos e
pobres, o que confere a espécie alta capacidade de se estabelecer em
ambientes degradados. Típica do bioma Cerrado e com desenvolvimento
rápido, foi a espécie que teve a maior heterogeneidade na velocidade de
formação de plântulas, na faixa de 25 a 35°C, assim como a tarumarana
(Figuras 38C e 38D), mas a maior velocidade tanto para protrusão de raiz
primária como para formação de plântula normal foi na temperatura de 30°C
(Figuras 37 e 38).
A tarumarana foi a espécie com menor velocidade de germinação e
formação de plântulas quando comparada com as outras espécies em
estudo ( Figura 37). De acordo com Labouriau (1983), na maioria dos casos
a velocidade de germinação aumenta com a temperatura dentro de certo
intervalo, passando a diminuir em outro, mas entre os intervalos ocorre a
faixa ótima ou, em alguns casos, a temperatura ótima. Para angico
vermelho, o índice de velocidade de germinação foi homogêneo na faixa de
temperatura estudada, porém para formação de plântulas a maior
velocidade foi a 30°C (Figuras 37A e 38A). O maior índice de velocidade de
germinação foi observado a 30°C para cerejeira, 25°C para sete-cascas, 30
e 35°C para sucupira preta e 25 e 30°C para tarumarana (Figuras 37 e 38).
Para a formação de plântulas, 30°C foi a temperatura que se observou a
maior velocidade para angico vermelho, cerejeira, sucupira preta e
tarumarana, exceto para sete-cascas que a 25 e 35°C os índices de
velocidade foram similares e maiores que na temperatura de 30°C.
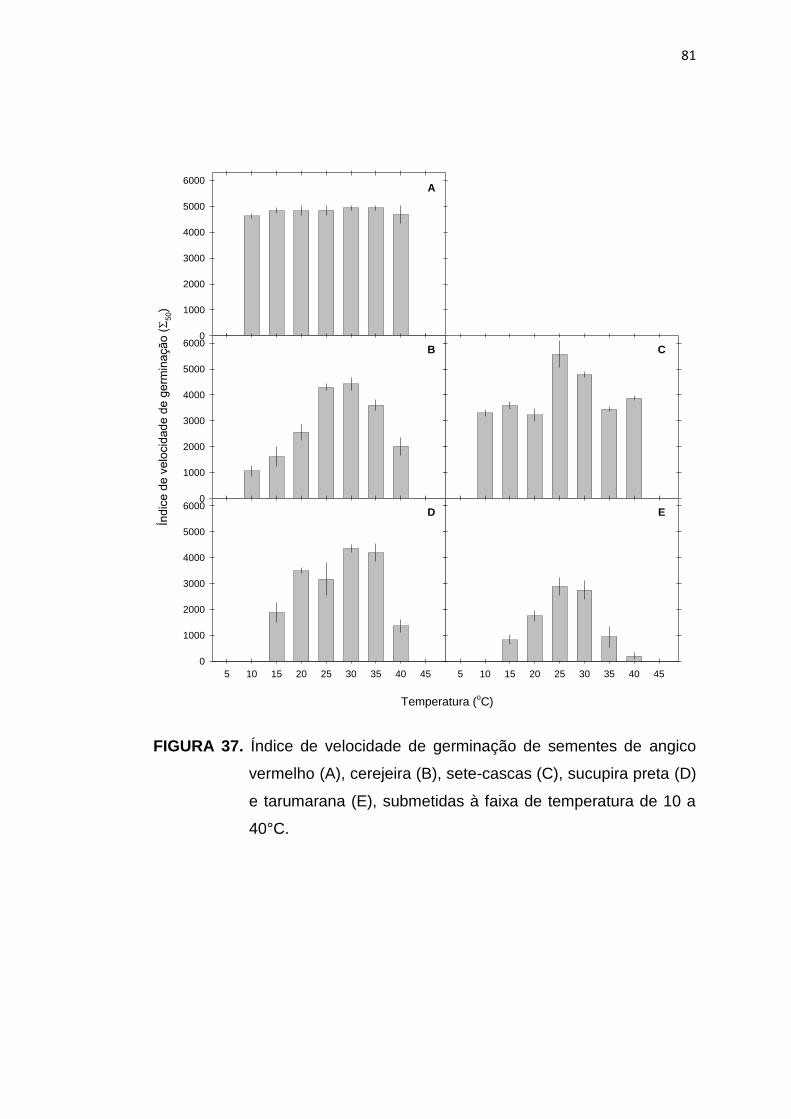
81
0
1000
2000
3000
4000
5000
6000
Índic
e d
e v
elo
cid
ade d
e g
erm
inação (
50)
0
1000
2000
3000
4000
5000
6000
Temperatura (oC)
5 10 15 20 25 30 35 40 45
0
1000
2000
3000
4000
5000
6000
5 10 15 20 25 30 35 40 45
A
B C
D E
FIGURA 37. Índice de velocidade de germinação de sementes de angico
vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta (D)
e tarumarana (E), submetidas à faixa de temperatura de 10 a
40°C.

82
0
1000
2000
3000
4000
5000
6000
Índic
e d
e v
elo
cid
ade d
e f
orm
ação d
e p
lântu
las (
50)
0
1000
2000
3000
4000
5000
6000
Temperatura (oC)
5 10 15 20 25 30 35 40 45
0
1000
2000
3000
4000
5000
6000
5 10 15 20 25 30 35 40 45
A
B C
D E
FIGURA 38. Índice de velocidade de formação de plântulas normais de
angico vermelho (A), cerejeira (B), sete-cascas (C), sucupira
preta (D) e tarumarana (E), submetidas à faixa de
temperatura de 10 a 40°C.
Observou-se que nas temperaturas mais elevadas ocorreu menor
velocidade de germinação. As altas temperaturas podem diminuir a
velocidade de germinação, provocando desorganização do processo sendo
que o número de sementes que conseguem completá-lo vai caindo

83
rapidamente, em decorrência, basicamente, dos efeitos sobre a atividade de
enzimas e das restrições ao acesso de oxigênio (Marcos Filho, 2005).
4.4 Tempo para Obtenção de 50% de Germinação das Sementes e 50%
de Formação de Plântulas para os Diferentes Tratamentos Térmicos
Ao analisar o tempo para obtenção de 50% de germinação das
sementes e 50% da formação de plântulas para os diferentes tratamentos
térmicos pode-se observar que a cerejeira foi a espécie que demorou maior
tempo para alcançar a mediana da germinação na temperatura de 15°C,
seguida de tarumarana a 20°C e sucupira preta a 15°C (Figura 39). Para
50% de formação de plântulas normais, foram observados maiores tempos
para alcançar 50% para as espécies sucupira preta a 15°C, cerejeira a 20°C
e tarumarana 20°C (Figura 40). Labouriau e Pacheco (1979) afirmaram que
a heterogeneidade das sementes em relação à sua taxa de germinação é
causada por uma minoria de sementes que germinam mais lentamente e
não por poucas que germinam mais rapidamente.

84
0
5
10
15
20
25
30
35
40
45
50
Tem
po p
ara
50%
de g
erm
inação (
d)
0
5
10
15
20
25
30
35
40
45
50
Temperatura (oC)
5 10 15 20 25 30 35 40 45
0
5
10
15
20
25
30
35
40
45
50
5 10 15 20 25 30 35 40 45
A
B C
D E
FIGURA 39. Tempo necessário para a obtenção de 50% de germinação de
sementes de angico vermelho (A), cerejeira (B), sete-cascas
(C), sucupira preta (D) e tarumarana (E), submetidas à faixa
de temperatura de 10 a 40°C.

85
0
5
10
15
20
25
30
35
40
45
50
Tem
po p
ara
50%
de form
ação d
e p
lântu
las (
d)
0
5
10
15
20
25
30
35
40
45
50
Temperatura (oC)
5 10 15 20 25 30 35 40 45
0
5
10
15
20
25
30
35
40
45
50
5 10 15 20 25 30 35 40 45
A
B C
D E
FIGURA 40. Tempo necessário para a obtenção de 50% de formação de
plântulas de angico vermelho (A), cerejeira (B), sete-cascas
(C), sucupira preta (D) e tarumarana (E), submetidas à faixa
de temperatura de 10 a 40°C.
A velocidade de germinação é bom índice para avaliar a rapidez de
ocupação de uma espécie em um determinado ambiente (Ferreira et al.,
2001). A germinação rápida é característica de espécies cuja estratégia é se
estabelecer no ambiente o mais rápido possível ou quando oportuno,
aproveitando condições ambientes favoráveis, como por exemplo, a

86
formação de clareiras ou ocorrência de chuvas. Em contrapartida a
germinação rápida pode ser imprópria ao estabelecimento de uma espécie
quando a germinação ocorrer, por exemplo, de forma errática e isolada na
estação seca (Borghetti e Ferreira, 2004). Segundo Ferreira et al. ( 2001),
quanto ao tempo de germinação as sementes podem ser classificadas
como rápidas (tempo médio <5 dias); intermediárias (tempo médio>5<10
dias) e lentas (tempo médio > 10 dias). Considerando esses critérios, o
angico vermelho e a sete-cascas foram classificadas como espécies rápidas
e, as outras espécies foram consideradas lentas.
4.5 Estimativas das Temperaturas Cardinais para a Germinação das
Sementes e para a Formação de Plântulas a partir do Índice de
Velocidade desses Processos (50)
As temperaturas cardinais para o processo germinativo das
sementes das espécies pode ser identificada com precisão somente para a
cerejeira, a sucupira preta e a tarumarana (Tabela 2), pois a espécie angico
vermelho e sete-cascas apresentaram baixo coeficiente de determinação da
regressão. Tal imprecisão pode ser devido ao fato de que as sementes
dessas espécies apresentaram um padrão de germinação, na qual não foi
possível identificar tendência significativa para a regressão (Figuras 41A e
41C) dentro da faixa de temperatura utilizada (entre 10 e 40ºC). Houve
situações em que não foi possível obter um ajuste bilinear, somente o ajuste
linear com a estimativa da temperatura basal mínima.
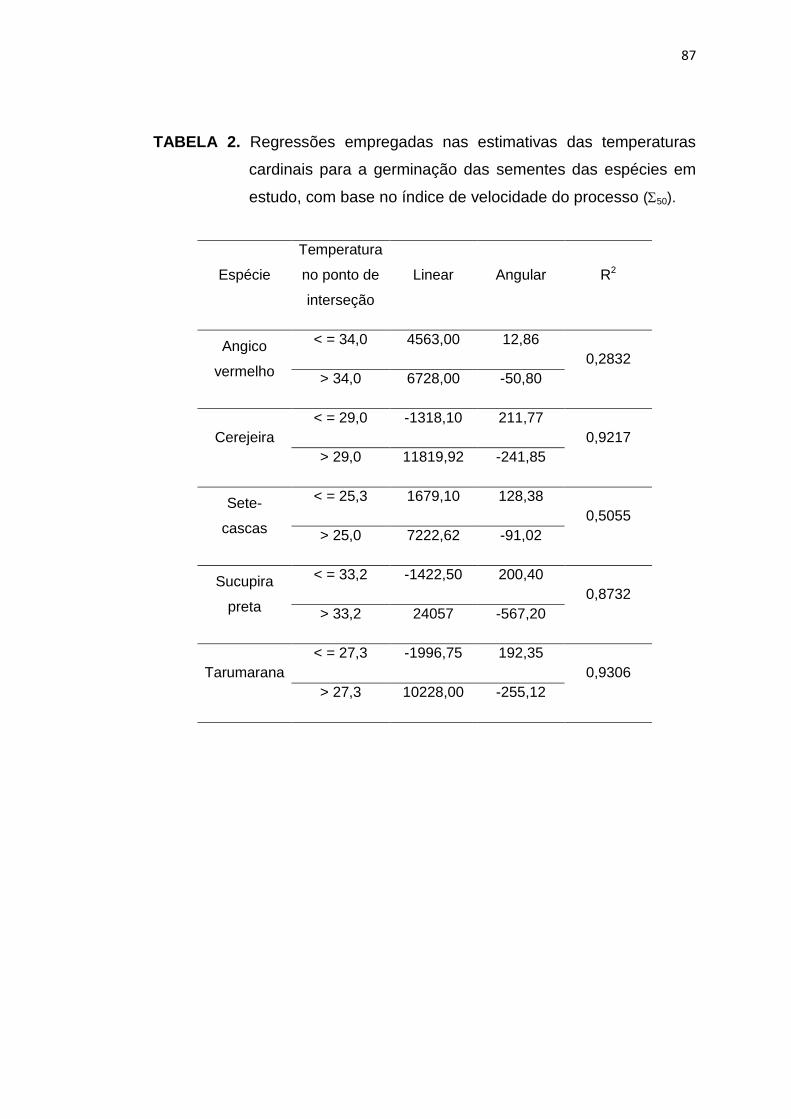
87
TABELA 2. Regressões empregadas nas estimativas das temperaturas
cardinais para a germinação das sementes das espécies em
estudo, com base no índice de velocidade do processo (50).
Espécie
Temperatura
no ponto de
interseção
Linear Angular R2
Angico
vermelho
< = 34,0 4563,00 12,86
0,2832
> 34,0 6728,00 -50,80
Cerejeira
< = 29,0 -1318,10 211,77
0,9217
> 29,0 11819,92 -241,85
Sete-
cascas
< = 25,3 1679,10 128,38
0,5055
> 25,0 7222,62 -91,02
Sucupira
preta
< = 33,2 -1422,50 200,40
0,8732
> 33,2 24057 -567,20
Tarumarana
< = 27,3 -1996,75 192,35
0,9306
> 27,3 10228,00 -255,12

88
Temperatura (oC)
5 10 15 20 25 30 35 40 45
0
1000
2000
3000
4000
5000
6000
Índic
e d
e v
elo
cid
ade d
e g
erm
inação ()
0
1000
2000
3000
4000
5000
6000
5 10 15 20 25 30 35 40 45
0
1000
2000
3000
4000
5000
6000
A
B C
D E
FIGURA 41. Regressões empregadas nas estimativas das temperaturas
cardinais do processo de germinação de sementes de angico
vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta
(D) e tarumarana (E), empregando-se o índice de velocidade
de germinação.
Já para todas as espécies foi possível estimar as temperaturas
cardinais para formação de plântulas de forma precisa (Tabela 3 e Figura
42).
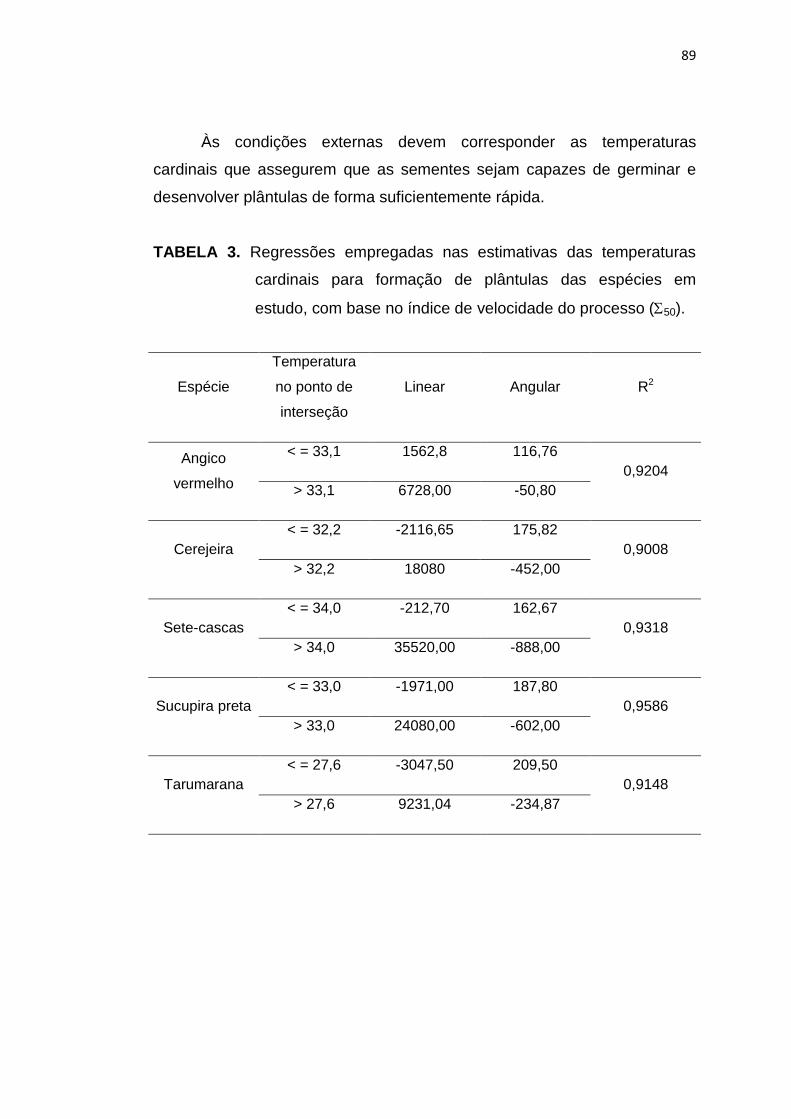
89
Às condições externas devem corresponder as temperaturas
cardinais que assegurem que as sementes sejam capazes de germinar e
desenvolver plântulas de forma suficientemente rápida.
TABELA 3. Regressões empregadas nas estimativas das temperaturas
cardinais para formação de plântulas das espécies em
estudo, com base no índice de velocidade do processo (50).
Espécie
Temperatura
no ponto de
interseção
Linear Angular R2
Angico
vermelho
< = 33,1 1562,8 116,76
0,9204
> 33,1 6728,00 -50,80
Cerejeira
< = 32,2 -2116,65 175,82
0,9008
> 32,2 18080 -452,00
Sete-cascas
< = 34,0 -212,70 162,67
0,9318
> 34,0 35520,00 -888,00
Sucupira preta
< = 33,0 -1971,00 187,80
0,9586
> 33,0 24080,00 -602,00
Tarumarana
< = 27,6 -3047,50 209,50
0,9148
> 27,6 9231,04 -234,87
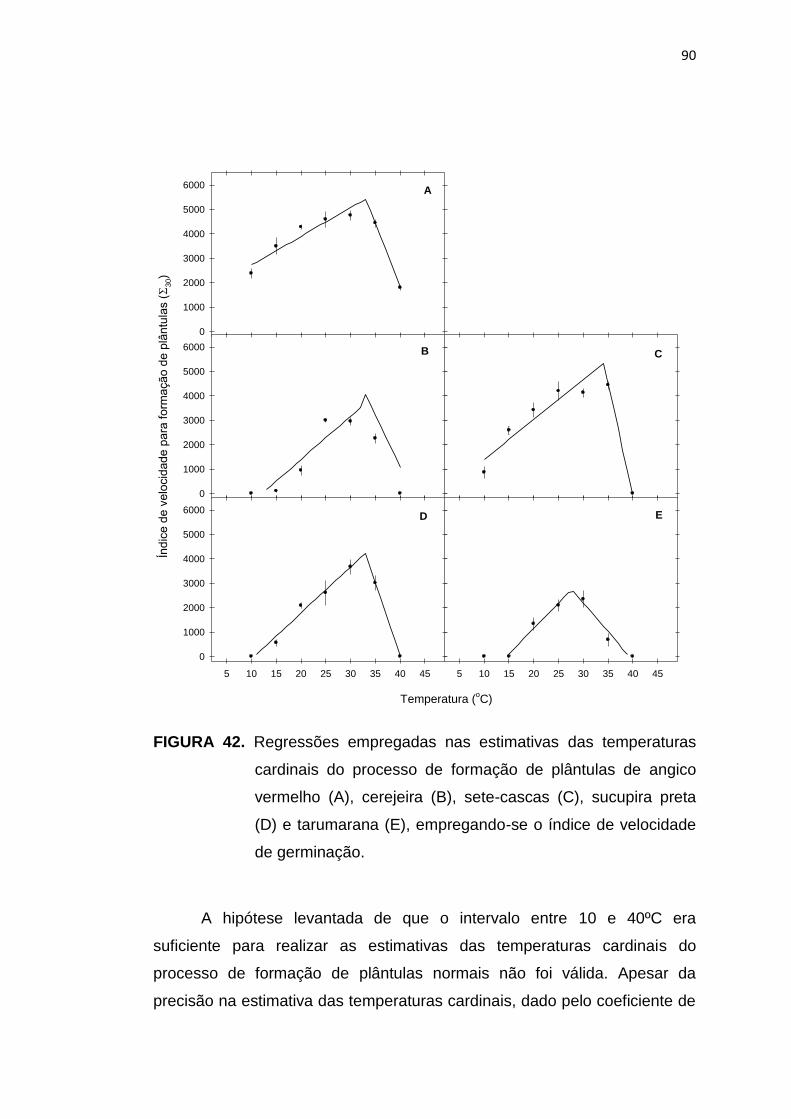
90
Temperatura (oC)
5 10 15 20 25 30 35 40 45
0
1000
2000
3000
4000
5000
6000
Índic
e d
e v
elo
cid
ade p
ara
form
ação d
e p
lântu
las (
30)
0
1000
2000
3000
4000
5000
6000
5 10 15 20 25 30 35 40 45
0
1000
2000
3000
4000
5000
6000
A
B C
D E
FIGURA 42. Regressões empregadas nas estimativas das temperaturas
cardinais do processo de formação de plântulas de angico
vermelho (A), cerejeira (B), sete-cascas (C), sucupira preta
(D) e tarumarana (E), empregando-se o índice de velocidade
de germinação.
A hipótese levantada de que o intervalo entre 10 e 40ºC era
suficiente para realizar as estimativas das temperaturas cardinais do
processo de formação de plântulas normais não foi válida. Apesar da
precisão na estimativa das temperaturas cardinais, dado pelo coeficiente de
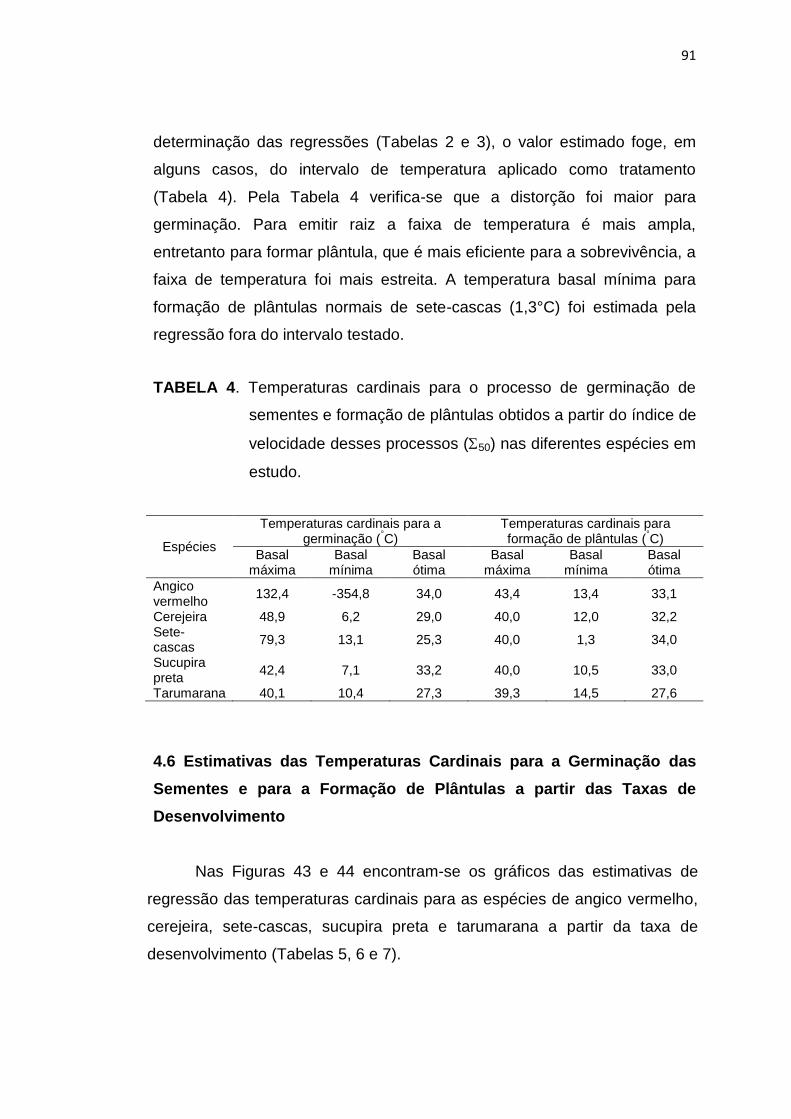
91
determinação das regressões (Tabelas 2 e 3), o valor estimado foge, em
alguns casos, do intervalo de temperatura aplicado como tratamento
(Tabela 4). Pela Tabela 4 verifica-se que a distorção foi maior para
germinação. Para emitir raiz a faixa de temperatura é mais ampla,
entretanto para formar plântula, que é mais eficiente para a sobrevivência, a
faixa de temperatura foi mais estreita. A temperatura basal mínima para
formação de plântulas normais de sete-cascas (1,3°C) foi estimada pela
regressão fora do intervalo testado.
TABELA 4. Temperaturas cardinais para o processo de germinação de
sementes e formação de plântulas obtidos a partir do índice de
velocidade desses processos (50) nas diferentes espécies em
estudo.
Espécies
Temperaturas cardinais para a germinação (
°C)
Temperaturas cardinais para formação de plântulas (
°C)
Basal máxima
Basal mínima
Basal ótima
Basal máxima
Basal mínima
Basal ótima
Angico vermelho
132,4 -354,8 34,0 43,4 13,4 33,1
Cerejeira 48,9 6,2 29,0 40,0 12,0 32,2 Sete-cascas
79,3 13,1 25,3 40,0 1,3 34,0
Sucupira preta
42,4 7,1 33,2 40,0 10,5 33,0
Tarumarana 40,1 10,4 27,3 39,3 14,5 27,6
4.6 Estimativas das Temperaturas Cardinais para a Germinação das
Sementes e para a Formação de Plântulas a partir das Taxas de
Desenvolvimento
Nas Figuras 43 e 44 encontram-se os gráficos das estimativas de
regressão das temperaturas cardinais para as espécies de angico vermelho,
cerejeira, sete-cascas, sucupira preta e tarumarana a partir da taxa de
desenvolvimento (Tabelas 5, 6 e 7).
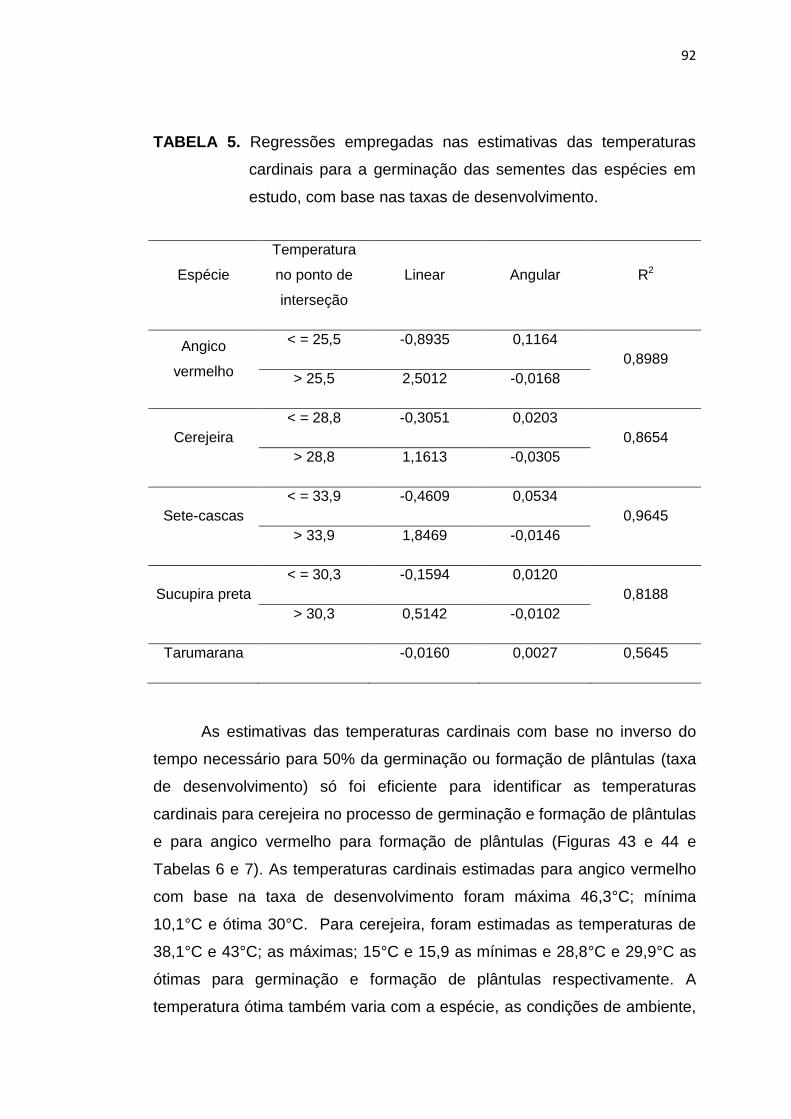
92
TABELA 5. Regressões empregadas nas estimativas das temperaturas
cardinais para a germinação das sementes das espécies em
estudo, com base nas taxas de desenvolvimento.
Espécie
Temperatura
no ponto de
interseção
Linear Angular R2
Angico
vermelho
< = 25,5 -0,8935 0,1164
0,8989
> 25,5 2,5012 -0,0168
Cerejeira
< = 28,8 -0,3051 0,0203
0,8654
> 28,8 1,1613 -0,0305
Sete-cascas
< = 33,9 -0,4609 0,0534
0,9645
> 33,9 1,8469 -0,0146
Sucupira preta
< = 30,3 -0,1594 0,0120
0,8188
> 30,3 0,5142 -0,0102
Tarumarana -0,0160 0,0027 0,5645
As estimativas das temperaturas cardinais com base no inverso do
tempo necessário para 50% da germinação ou formação de plântulas (taxa
de desenvolvimento) só foi eficiente para identificar as temperaturas
cardinais para cerejeira no processo de germinação e formação de plântulas
e para angico vermelho para formação de plântulas (Figuras 43 e 44 e
Tabelas 6 e 7). As temperaturas cardinais estimadas para angico vermelho
com base na taxa de desenvolvimento foram máxima 46,3°C; mínima
10,1°C e ótima 30°C. Para cerejeira, foram estimadas as temperaturas de
38,1°C e 43°C; as máximas; 15°C e 15,9 as mínimas e 28,8°C e 29,9°C as
ótimas para germinação e formação de plântulas respectivamente. A
temperatura ótima também varia com a espécie, as condições de ambiente,
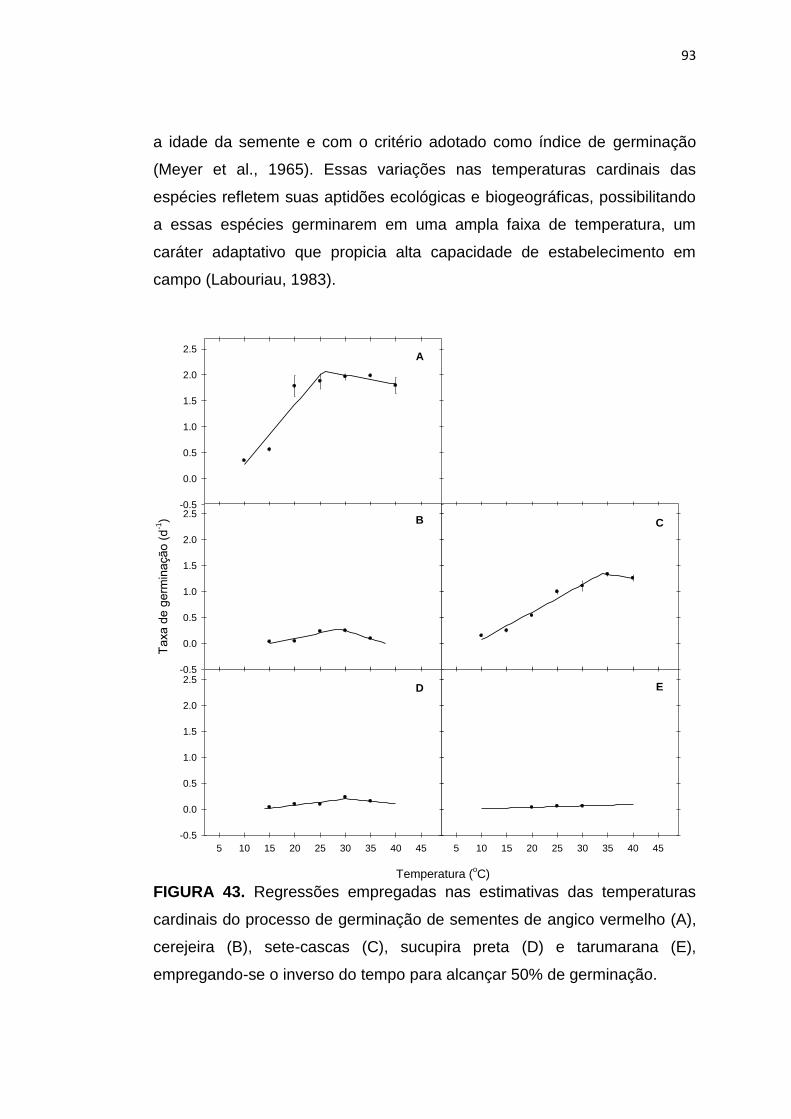
93
a idade da semente e com o critério adotado como índice de germinação
(Meyer et al., 1965). Essas variações nas temperaturas cardinais das
espécies refletem suas aptidões ecológicas e biogeográficas, possibilitando
a essas espécies germinarem em uma ampla faixa de temperatura, um
caráter adaptativo que propicia alta capacidade de estabelecimento em
campo (Labouriau, 1983).
Temperatura (oC)
5 10 15 20 25 30 35 40 45
-0.5
0.0
0.5
1.0
1.5
2.0
2.5
Taxa d
e g
erm
inação (
d-1
)
-0.5
0.0
0.5
1.0
1.5
2.0
2.5
5 10 15 20 25 30 35 40 45
-0.5
0.0
0.5
1.0
1.5
2.0
2.5
A
B C
D E
FIGURA 43. Regressões empregadas nas estimativas das temperaturas
cardinais do processo de germinação de sementes de angico vermelho (A),
cerejeira (B), sete-cascas (C), sucupira preta (D) e tarumarana (E),
empregando-se o inverso do tempo para alcançar 50% de germinação.
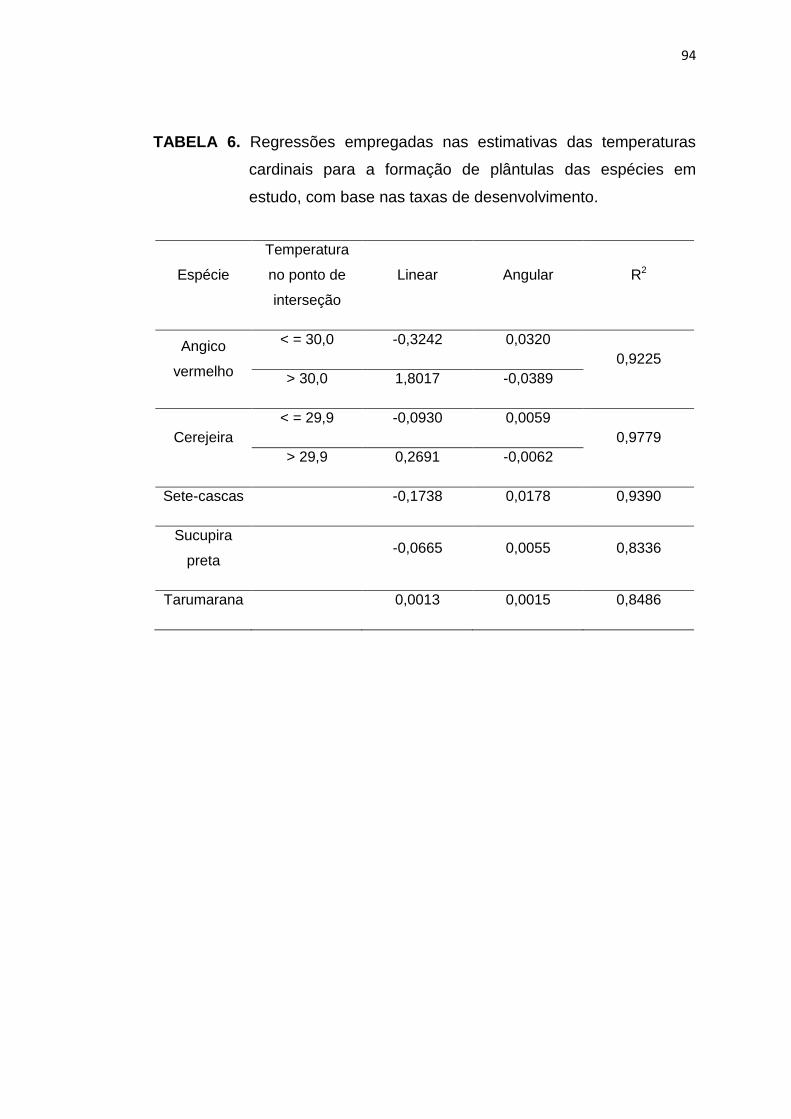
94
TABELA 6. Regressões empregadas nas estimativas das temperaturas
cardinais para a formação de plântulas das espécies em
estudo, com base nas taxas de desenvolvimento.
Espécie
Temperatura
no ponto de
interseção
Linear Angular R2
Angico
vermelho
< = 30,0 -0,3242 0,0320
0,9225
> 30,0 1,8017 -0,0389
Cerejeira
< = 29,9 -0,0930 0,0059
0,9779
> 29,9 0,2691 -0,0062
Sete-cascas -0,1738 0,0178 0,9390
Sucupira
preta -0,0665 0,0055 0,8336
Tarumarana 0,0013 0,0015 0,8486
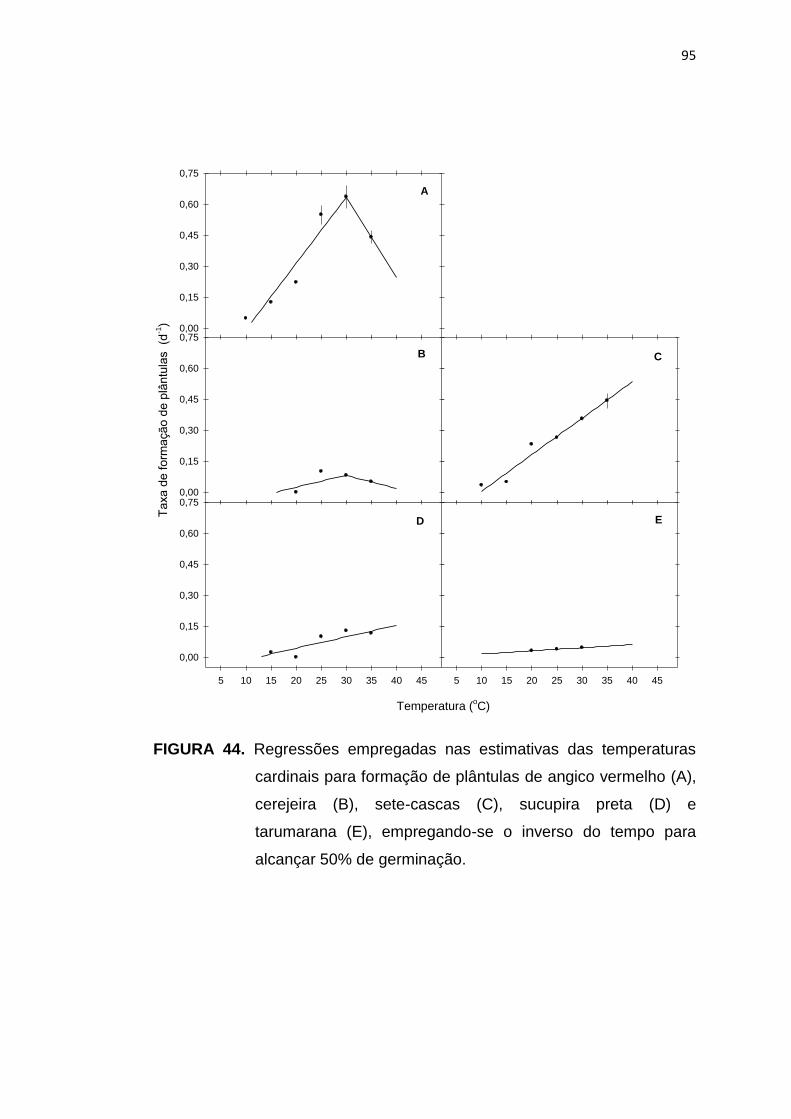
95
Temperatura (oC)
5 10 15 20 25 30 35 40 45
0,00
0,15
0,30
0,45
0,60
0,75T
axa
de f
orm
açã
o d
e p
lântu
las
(d-1
)
0,00
0,15
0,30
0,45
0,60
0,75
5 10 15 20 25 30 35 40 45
0,00
0,15
0,30
0,45
0,60
0,75
A
B C
D E
FIGURA 44. Regressões empregadas nas estimativas das temperaturas
cardinais para formação de plântulas de angico vermelho (A),
cerejeira (B), sete-cascas (C), sucupira preta (D) e
tarumarana (E), empregando-se o inverso do tempo para
alcançar 50% de germinação.
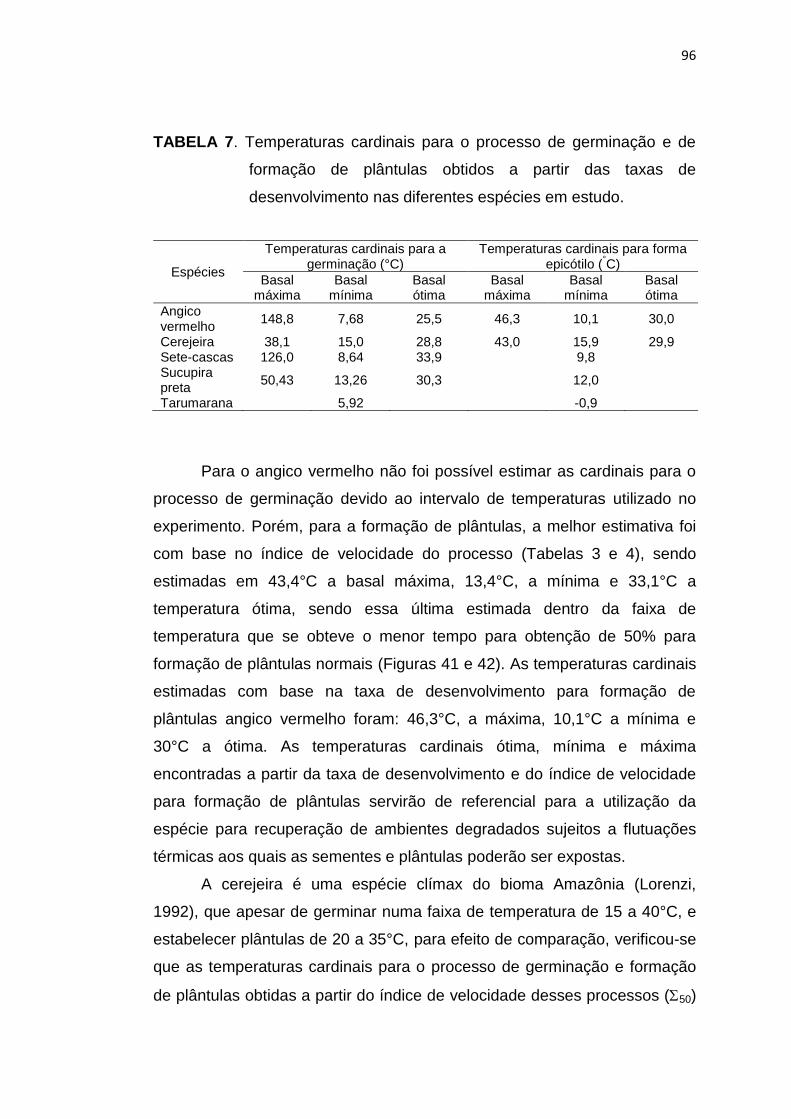
96
TABELA 7. Temperaturas cardinais para o processo de germinação e de
formação de plântulas obtidos a partir das taxas de
desenvolvimento nas diferentes espécies em estudo.
Espécies
Temperaturas cardinais para a germinação (°C)
Temperaturas cardinais para forma epicótilo (
°C)
Basal máxima
Basal mínima
Basal ótima
Basal máxima
Basal mínima
Basal ótima
Angico vermelho
148,8 7,68 25,5 46,3 10,1 30,0
Cerejeira 38,1 15,0 28,8 43,0 15,9 29,9 Sete-cascas 126,0 8,64 33,9 9,8 Sucupira preta
50,43 13,26 30,3 12,0
Tarumarana 5,92 -0,9
Para o angico vermelho não foi possível estimar as cardinais para o
processo de germinação devido ao intervalo de temperaturas utilizado no
experimento. Porém, para a formação de plântulas, a melhor estimativa foi
com base no índice de velocidade do processo (Tabelas 3 e 4), sendo
estimadas em 43,4°C a basal máxima, 13,4°C, a mínima e 33,1°C a
temperatura ótima, sendo essa última estimada dentro da faixa de
temperatura que se obteve o menor tempo para obtenção de 50% para
formação de plântulas normais (Figuras 41 e 42). As temperaturas cardinais
estimadas com base na taxa de desenvolvimento para formação de
plântulas angico vermelho foram: 46,3°C, a máxima, 10,1°C a mínima e
30°C a ótima. As temperaturas cardinais ótima, mínima e máxima
encontradas a partir da taxa de desenvolvimento e do índice de velocidade
para formação de plântulas servirão de referencial para a utilização da
espécie para recuperação de ambientes degradados sujeitos a flutuações
térmicas aos quais as sementes e plântulas poderão ser expostas.
A cerejeira é uma espécie clímax do bioma Amazônia (Lorenzi,
1992), que apesar de germinar numa faixa de temperatura de 15 a 40°C, e
estabelecer plântulas de 20 a 35°C, para efeito de comparação, verificou-se
que as temperaturas cardinais para o processo de germinação e formação
de plântulas obtidas a partir do índice de velocidade desses processos (50)

97
foram estimadas com maior precisão em comparação com as temperaturas
cardinais estimadas pela taxa de desenvolvimento que extrapolou, pela
estimativa da regressão o valor obtido em condições controladas do
experimento, a temperatura supra-ótima (Tabelas 4, 5, 6 e 7). Foram
estimadas para germinação e formação de plântulas de cerejeira as
temperaturas basais mínimas como sendo 6,2 e 12°C, as ótimas (29 e
32,2°C) e máximas (48,9 e 40°C, respectivamente).
As sementes de sete-cascas tiveram ampla adaptação às variações
de temperatura e, como o angico vermelho, suportou mínimas mais baixas
que as outras espécies estudadas. O amplo limite de temperatura no qual a
sete-cascas germinou possui um caráter adaptativo, pois segundo
Townsend e MacGinnies (1972), isto pode propiciar uma alta capacidade de
estabelecimento em campo, aumentando a chance de sobrevivência, em
comparação com espécies que apresentam limites estreitos de temperatura
para germinar. Os valores estimados das temperaturas cardinais para a
germinação no intervalo de 10 a 40°C não foi suficiente para realizar as
estimativas pelo índice de velocidade do processo (50), mas foi possível
estimar a basal máxima (40°C), a mínima (1,3°C) e a ótima (34°C) para
formação de plântulas (Tabela 4).
Para sucupira preta, os valores estimados das temperaturas
cardinais para germinação não foram precisos diferindo das respostas
obtidas nos tratamentos.
Os valores estimados com base no índice de velocidade do processo
(50) para formação de plântulas foram basal máxima (40°C), basal mínima
(10,5°C) e ótima (33°C) (Tabelas 4, 5 e 7).
A estimativa pela regressão linear com base no índice de velocidade
(50) estimou para tarumarana como temperaturas basais máximas (40,1°C
e 39,3°C), mínimas (10,4 e 14,5°C) e ótimas (27,3 e 27,6°C), para
germinação e formação de plântulas respectivamente. Entretanto o modelo
não foi eficiente para estimar as basais para germinação e formação de
plântulas com base na taxa de desenvolvimento (Figura 44 e Tabela 7)

98
porque houve temperaturas onde não se atingiu o tempo de 50% da
germinação ou da formação de plântulas.
Considerando-se que as espécies florestais nativas são, em sua
maioria, de crescimento lento, o fundamental é ter em mente que a
recuperação da cobertura florestal de uma área implica na formação de
novas populações com plantio de espécies florestais nativas com aspectos
ecofisiológicos conhecidos e que sejam utilizadas espécies de rápido
estabelecimento que sirvam como fontes de sementes adaptadas, com
ampla variabilidade genética, para possibilitar o contínuo processo de
sucessão em cada local.
Considerando os aspectos fisiológicos estudados das cinco espécies
dos três biomas do estado de Mato Grosso e as suas respostas a amplitude
térmica na germinação e na formação de plântulas;que dentro de cada um
desses biomas, ocorrem variações nas formas de cobertura vegetal, como
por exemplo, vegetações típicas do Cerrado encontradas dentro do bioma
Floresta Amazônica e no Pantanal; que além disso, ocorrem variações
ambientais marcantes, principalmente as associadas aos tipos de solo ao
clima e às variações fisiográficas, que propiciam a formação de populações
geneticamente distintas, com vantagens adaptativas em cada local,
confirmou-se assim a hipótese de que entre espécies desses biomas
haviam temperaturas cardinais diferentes para estabelecimento das
espécies foi confirmada, e assim várias espécies florestais têm ampla
distribuição natural, estendendo-se por mais de um bioma, o que sugere a
variabilidade genética das espécies do ecossistema matogrossense.

99
5 CONCLUSÕES
O teor de água das sementes das espécies estudadas varia de 6,8% a
10,5%.
Apenas para sete-cascas, a massa de mil sementes (de 80,18g a
90,96g) e o número de sementes por quilo (de 11.000 a 12.470) diferem dos
dados obtidos na literatura. Para as outras espécies, os valores são muito
próximos.
A cerejeira é a espécie dentre as estudadas que despende maior tempo
para obtenção de 50% da germinação na temperatura de 15°C, seguida de
tarumarana a 20°C e sucupira preta a 15°C. Os maiores tempos
necessários para 50% da formação de plântulas são observados na espécie
sucupira preta na temperatura de 15°C e na cerejeira a 20°C.
A faixa de temperatura de 10 a 40°C não é suficiente para estimar as
temperaturas cardinais do processo de germinação para o angico vermelho
e para a sete-cascas, com base no índice de velocidade de germinação.
Contudo, esse índice se presta para avaliar as temperaturas cardinais na
formação de plântulas. Na germinação das sementes, as temperaturas
cardinais máxima, mínima e ótima para a cerejeira são de 48,9°C; 6,2°C e
29,0°C. Para a sucupira preta, de 42,4°C; 7,1°C e 33,2°C. Para a
tarumarana, de 40,1°C; 10,4°C e 27,3°C. Na formação de plântulas normais,
as temperaturas cardinais máxima, mínima e ótima para o angico vermelho

100
são de 43,4°C; 13,4°C e 33,1°C, respectivamente. Para cerejeira, de
40,0°C; 12,0°C e 32,2°C. Para a sete-cascas, de 40,0°C; 1,3°C e 34,0°C.
Para a sucupira preta, de 40,0°C; 10,5°C e 33,0°C. Para a tarumarana, de
39,3°C; 14,5°C e 27,6°C.
As estimativas das temperaturas cardinais com base no inverso do
tempo necessário para obtenção de 50% da germinação ou 50% da
formação de plântulas não permite identificar as temperaturas cardinais dos
processos de germinação e de formação de plântulas na maioria dos casos.
Somente se identifica que, na germinação, as temperaturas cardinais
máxima, mínima e ótima para a cerejeira são de 38,1°C; 15,0°C e 28,8°C.
Na formação de plântulas, as temperaturas cardinais máxima, mínima e
ótima para o angico vermelho são de 46,3°C; 10,1°C e 30,0°C. Para
cerejeira, de 43,0°C; 15,9°C e 29,9°C.

101
6 REFERÊNCIAS BIBLIOGRÁFICAS
ABDON, M.M. et al. Desmatamento no bioma pantanal até o ano de 2002: relações com a fitofisionomia e limites municipais. Revista Brasileira de Cartografia, Rio de Janeiro, n.59, v.1, p.17-24, 2007.
ABRAMOWITZ, M.; STEGUN, I.A. Handbook of mathematical functions with formulas, graphs and mathematical tables. New York. Dover Publications, 1965.1045p.
ALBUQUERQUE, K.S.; GUIMARAES, R.M. Comportamento fisiológico de sementes de Bowdichia virgiliodes, Kunt. sob diferentes temperaturas e condições de luz. Cerne, Lavras, v.13, n.1, p.64-70, 2007a.
ALBUQUERQUE, K.S.; GUIMARAES, R.M.; ALMEIDA, Í.F. de e CLEMENTE, A. da C.S. Métodos para a superação da dormência em sementes de sucupira-preta (Bowdichia virgilioides Kunth.). Ciência e Agrotecnologia, Lavras, v.31, n.6, p.1716-1721, 2007.
ANDERSON, L.O.; ROJAS E.H.M.; SHIMABUKURO, Y.E. Avanço da soja sobre os ecossistemas Cerrado e Floresta no estado do Mato Grosso. In: XI Simpósio Brasileiro de Sensoriamento Remoto, 2003, Belo Horizonte. Anais... Belo Horizonte: INPE, 2003. p.19-25.
ARAÚJO NETO, J.C.; AGUIAR, I.B.; FERREIRA, V.M. Efeito da temperatura e da luz na germinação de sementes de Acácia polyphylla DC. Revista Brasileira de Botânica, São Paulo, v.26, n.2, p.249-256, 2003.
ARAÚJO NETO, J.C.; AGUIAR, I.B.; FERREIRA, V.M.; RODRIGUES, T.J.D. Temperaturas cardeais e efeito da luz na germinação de sementes de mutamba. Revista Brasileira de Engenharia Agrícola e Ambiental, Campina Grande, v.6, n.3, p.460-465, 2002.
ARNOLD, C.Y. The determination and significance of the base temperature in a linear heat unit system. American Society for Horticultural Science, Geneva, v.74, n.4, p.430-445, 1959.
AUSTIN, R.B. Effects of environmental before harvesting on viability. In: ROBERTS, E.H. (Ed.). Viability of seeds. Syracuse: Syracuse University Press, 1972. p.114-149.
AZEVEDO, M.I.R. Estrutura e restauração de cerradão em Palmas - TO e germinação de sementes de Buchenavia tomentosa Eichler, Hymenaea stigonocarpa Mart. ex Hayne, Guazuma ulmifolia Lam. e Enterolobium gummiferum (Mart.) J.F. Macbr. 2008. 139f. Tese (Doutorado em Ciência Florestal) - Universidade Federal de Viçosa, Minas Gerais, 2008.

102
BARBOSA, L. M. Manual sobre princípios da recuperação vegetal de áreas degradadas. São Paulo: SMA, 2000. 76p.
BASKIN, C.C.; BASKIN, J.M. Seeds: ecology, biogeography and evolution of dormancy and germination. San Diego: Academic Press, 1998. 666p.
BENECH-ARNOLD, R.L., SÁNCHEZ, R.A. (ed.): Handbook of seed physiology. Application to agriculture. - New York : The Haworth Press, 2004. 480p.
BEWLEY, J.D.; BLACK, M. Seeds: physiology of development and germination. New York: Plenum Press, 1994. 445p.
BEWLEY, J.P.; BLACK, M. Physiology and biochemistry of seeds. Berlin: Springer-Verlag, 1982. v.2, 375p.
BONAN, G.B. Forest and climate change: forcing, feedbacks, and climate benefits of forests. Science, v.320, n.5882, p.1444-1449, 2008.
BOREM, A. Melhoramento de plantas. 2. ed. São Paulo: UFV, 1998. 453p.
BORGES, E.E.L.; RENA, A.B. Germinação de sementes. In: AGUIAR, I.B.; PIÑA-RODRIGUES, F.C.M.; FIGLIOLIA, M.B. (Coord.) Sementes florestais tropicais. Brasília: ABRATES, 1993. p.83-136.
BORGHETTI, F. Temperaturas extremas e a germinação de sementes. In: NOGUEIRA, R.J.M.C.; ARAÚJO, E.L.; WILLADINO, L.G.; CAVALCANTE, U.M.T. (eds.) Estresses ambientais, danos e benefícios em plantas. Recife, PE: MXM Gráfica e Editora, 2005. p.207-218.
BORGHETTI, F.; FERREIRA, A.G. Interpretação de resultados de germinação. In: FERREIRA, A.G.; BORGHETTI, F. (orgs.). Germinação: do básico ao aplicado. Porto Alegre: Artmed, 2004. p.209-222.
BOTEZELLI, L. Estudo do armazenamento de sementes de quatro procedências de Baru, Dypteryx alata Vogel. Lavras: UFLA, 1998. 103p. Dissertação (Mestrado em Engenharia Florestal) - Universidade Federal de Lavras, 1998.
BRADBEER, J.W. Seed dormancy and germination. New York: Blackie Academic & Professional, Chapman and Hal, 1988. 146p.
BRAGA, R. Plantas do Nordeste, especialmente do Ceará. 3 ed. Fortaleza: ESAM, 1976. 510p.
BRANDÃO, M.; FERREIRA, P.B.D. Flora apícola do cerrado. Informe Agropecuário, Belo Horizonte, v.15, n.168, p.4-8, 1991.
BRASIL. Regras para análise de sementes. Brasília: Ministério da Agricultura, Departamento de Nacional de Produção Vegetal, 1992. 365p.

103
CARVALHO, N.M.; NAKAGAWA, J. Sementes: ciência, tecnologia e produção. Jaboticabal: Funep, 2000. 588p.
CARVALHO, P.E.R. Espécies florestais brasileiras: recomendações silviculturais, potencialidades e uso da madeira. Colombo: EMBRAPA – CNPF, Brasília: EMBRAPA – SPI, 1994. 640p.
COLLI, A.; TEODORO, A.S.; SILVEIRA FILHO, T.B.; RODRIGUES, D.F.; PINA-RODRIGUES, F.C.M. Influência da temperatura na germinação de sementes de angico-vermelho, Anadenanthera macrocarpa (FAM) Leg In: 57ª Reunião Anual da Sociedade Brasileira para Progresso da Ciência - Fortaleza, CE - Julho/2005. Anais... Fortaleza, 2005.
CORREA. M.P. Dicionário das plantas úteis do Brasil. Rio de Janeiro: IBDF, 1978. v.5, 687p.
COUTINHO, L.M. O conceito de bioma. Acta Botânica Brasilica, São Paulo, v.20, n.1, p.13-23, 2006.
CRAUFURD, P.Q.; QI, A.; ELLIS, R.H.; SUMMERFIELD, R.J.; ROBERTS, E.H.; MAHALAKSHMI, V. Effect of temperature on time to panicle initiation and leaf appearance in sorghum. Crop Science, Madison, v.38, n.4, p.942-947, 1998.
CUNHA, M. do C.L.; FERREIRA, R.A. Aspectos morfológicos da semente e do desenvolvimento da planta jovem de Amburana cearensis (Arr. Cam.) A.C. Smith - Cumaru - Leguminosae Papilionoideae. Revista Brasileira de Sementes, Brasília, v.25, n.2, p.89-96, 2003.
DAVIDE, A.C.; FARIA, J.M.R.; BOTELHO, S.A. Propagação de espécies florestais. Belo Horizonte: CEMIG/UFLA/FAEPE, 1995. 40p.
DE CASTRO, R.D.; BRADFORD, K.J; HILHORST, H.W.M. Desenvolvimento de sementes e conteúdo de água. In: FERREIRA, A.G.; BORGHETTI, F. (Org.). Germinação: do básico ao aplicado. 1ª ed. Porto Alegre: ARTMED, 2004. v.1, p.51-67.
DIGNART, S. Análise de sementes de jatobá do cerrado [Hymenaea stigonocarpa (Hayne) Mart.) ] e barbatimão [Stryphnodendron adstringens (Mart.) Cov.]. 1998, 58f. Dissertação (Mestrado em Agricultura Tropical) - Faculdade de Agronomia e Medicina Veterinária, Universidade Federal de Mato Grosso, Cuiabá- MT, 1998.
EMBRAPA, 2008. A Embrapa nos biomas brasileiros. 16p. Disponível em: <http://www.embrapa.br/publicacoes/institucionais/laminas-biomas.pdf.> Acesso em 06.01.2009.

104
FARIA, J.M.R. Propagação de espécies florestais. In: Simpósio Mata Ciliar: ciência e tecnologia, 1, 1999, Belo Horizonte. Anais... Lavras: UFLA/FAEPE/CEMIG, 1999. p.69-79.
FERREIRA, A.G.; CASSOL, B.; ROSA, S.G.T.; SILVEIRA, T.S.; STIVAL, A.L.; SILVA, A.A. Germinação de sementes de Asteraceae nativas no Rio Grande do Sul, Brasil. Acta Botânica Brasilica, São Paulo, v.15, n.2, p.231-242, 2001.
FERRONATO, A. Análises de sementes de Bowdichia virgilioides H.B.K. (sucupira preta) e Cybistax antisyphilitica M. (pé de anta). 1999. 80f - Dissertação (Mestrado em Agricultura Tropical) - Faculdade de Agronomia e Medicina Veterinária, Universidade Federal de Mato Grosso, Cuiabá, 1999.
FINCH-SAVAGE, W.E. The use of population-based threshold models to describe and predict the effects of seedbed environment on germination and seedling emergence of crops. In: BENECH-ARNOLD, R.; SANCHES, R.A. Seed physiology: Applications to Agriculture. New York: The Haworth Reference Press, 2004. p.51-95.
FIOR, S.S.; CALI, A.C.; LEONHARDT, C. Iheringia. Série Botânica, Porto Alegre, v. 59, n. 2, p. 207-210, 2004.
GAO, L.Z.; JIN, Z.Q.; HUANG, Y.; ZHANG, L.Z. Rice clock model – a computer simulation model of rice development. Agricultural and Forest Meteorology, Amsterdam, v.60, n.1, p.1-16, 1992.
GARCIA, Q.S.; DINIZ, I.S.S. Comportamento germinativo de três espécies de Vellozia da Serra do Cipó, MG. Acta Botânica Brasilica, São Paulo, v.17, n.4, p.487- 494, 2003.
GARCIA-HUIDOBRO, J.; MONTEITH, J.L.; SQUIRE, G.R. Time, temperature, and germination of pearl millet (Pennisetum typhoides S. & H.). Journal of Experimental Botany, Oxford, v.33, n.133, p.288-296, 1982.
GILMORE, E.C.; ROGERS, J.S. Heat units as a method of measuring maturity in corn. Agronomy Journal, Madison, v.50, n.10, p.611-615, 1958.
GIRARDI, G. Santuário interrompido. Página 22, n.18, abr. 2008. Disponível em: www.pagina22.com.br/index.cfm?fuseaction=homeReportagens.
GODOI, S; TAKAKI, M. Efeito da temperatura e a participação do fitocromo no controle da germinação de sementes de embaúba. Revista Brasileira de Sementes, Brasília, v.27, n.2, p.87-90, 2005.
GRAÇA, M.E.C.; TAVARES, F.R. Propagação vegetativa de espécies florestais. In: GALVÃO, A.P.M. Reflorestamento de propriedades rurais

105
para fins produtivos e ambientais. Brasília, DF: Embrapa, 2000. Cap. 9, p.175-197.
HARTMANN, H. T. et al. Plant propagation: principles and practices. 6. ed. New Jersey: Prentice-Hall, 1997, 770 p.
HENDRICKS, S.B.; TAYLORSON, R.B. Variation in germination and amino acid leakage of seeds with temperature related to membrane phase change. Plant Physiology, Maryland, v.58, n.1, p.7-11, 1976.
HINTZE, J. NCSS and PASS. Number Cruncher Statistical System.
Kaysville, UT, USA. 2004. www.ncss.com.
HOOVER, M.W. Some effects of temperature on the growth of southern peas. Proceedings of the American Society of Horticulture Science, v.66, p.308-414, 1955.
IBAMA. Amazônia. Disponível em: <http://www.ibama.gov.br/ecossistemas/amazonia.htm>. Acesso em: 18 dez. 2008.
IBAMA. Pantanal. Disponível em: <http://www.ibama.gov.br/ecossistemas/pantanal.htm>. Acesso em 18 dez. 2008.
IBGE. Diretoria de Geociências. Mapa de biomas do Brasil: primeira aproximação. Rio de Janeiro, 2004. 1 mapa; 110 cm x 92 cm. Escala 1:5. 000.000.
IBGE. Mapa de biomas do Brasil. Rio de Janeiro: IBGE/MMA, 2004.
INPE. Monitoramento da cobertura da florestal da Amazônia por satélites. SISTEMAS PRODES, DETER, DEGRAD E QUEIMADAS 2007-2008. São José dos Campos, 10-12-2008. 47p.
IOSSI, E.; SADER, R.; PIVETTA, K.F.L.; BARBOSA, J.C. Efeitos de substratos e temperaturas na germinação de sementes de tamareira-anã (Phoenix roebelenii O'Brien). Revista Brasileira de Sementes, Brasília, v.25, n.2, p.63-69, 2003.
JACKSON, R.D.; IDSO, S.B. Surface albedo and desertification. Science, v.189, n. 420, p.1012-1013, 1975.
KITCHEN, J.T. Occurrence and time of reproductive structure development and their relation to the yield of Rutgers tomatoes. 1956. Rutgers University (Ph. D. thesis). 1956.
LABOURIAU, L.G. A germinação das sementes. Washington D.C: Secretaria Geral das Organizações dos Estados Americanos, Programa Regional de Desenvolvimento Cientifico e Tecnológico. 1983.174p.

106
LABOURIAU, L.G.; PACHECO, A. Isothermal germination rates in seeds of Dolichos biflorus L. Separata del Boletin de la Sociedad Venezoelana de Ciências Naturales n.136 - Tomo XXXIV - p.73-112, Caracas, Venezuela, 1979.
LABOURIAU, L.G.; PACHECO, A. On the frequency of isothermal germination in seeds of Dolichos biflorus L. Plant and Cell Physiology, Oxford, v.19, n.3, p. 507-512 ,1978.
LARCHER, W. Ecofisiologia vegetal. São Carlos: Rima, 2000. 531p.
LARSON, B.C. Patways of development in mixed-species stands. In: KELTY, M.J.; LARSON, B.C.; OLIVER, C.D. (Ed.). The ecology and silviculture of mixed-species forest. Dordrecht: Kluwer Academic, 1992. p.3-10.
LEHENBAUER, P.A. Growth of maize seedlings in relation to temperature. Physiological Research, v.1, p.247-288, 1914.
LIMA, J.D.; ALMEIDA, C.C.; DANTAS, V.A.V.; SILVA, B.M.S., MORAES, W.S. Efeito da temperatura e do substrato na germinação de sementes de Caesalpinia ferrea Mart. ex Tul. (Leguminosae, Caesalpinoideae). Revista Árvore, Viçosa, v.30, n.4, 2006.
LIMAS, J.D.; SILVA, B.M.S.; MORAES, W.S. Germinação e armazenamento de sementes de Virola surinamensis (Rol.) Warb. (Myristicaceae). Revista Árvore, Viçosa, v.31, n.1, p.37-42, 2007.
LINDSEY, L.A.; NEWMAN, J.E. Use of official weather data in spring time – temperature analysis of an Indiana phenological record. Ecology, v.37, p.812-823, 1956.
LOPES, J.C.; PEREIRA, M.D. Germinação de sementes de cubiu em diferentes substratos e temperaturas. Revista Brasileira de Sementes, Brasília, v.27, n.2, p.146-150, 2005.
LOPEZ, J.A.; LITTLE, E.L., Jr.; RITZ, G.F.; ROMBOLD, J.S.; HAHN, W.J. Arboles comunes del Paraguay: ñande yvyra mata kuera. Washington: Cuerpo de Paz, 1987. 425 p.
LORENZI, H. Árvores brasileiras. Manual de identificação e cultivo de plantas arbóreas nativas do Brasil. Ed. Plantarum, Nova Odessa, SP, 1992. 368p.
LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. v.2. 2 ed., Ed. Plantarum, Nova Odessa, SP, 2002. 384p.

107
MADRIAGA, F.J.; KNOTT, J.E. Temperature summations in relation to lettuce growth. Proceedings of the American Society of Horticulture Science, v.58, p.147-152, 1951.
MAGOON, C.A.; CULPEPPER, C.W. Response of sweet corn to varying temperatures from time of planting to canning maturity. U.S. Department of Agriculture Technology Bulletin. 1932. 312p.
MAGUIRE, J.D. Speed of germination-aid in selection and evaluation for seedling emergence and vigor. Crop Science, Madison, v.2, n.1, p.176-177, 1962.
MARCOS FILHO, J. Fisiologia de sementes de plantas cultivadas. Piracicaba: FEALQ, 2005. 495p.
MARTINS, C.C.; MACHADO, C.G.; NAKAGAWA, J. Temperatura e substrato para o teste de germinação de sementes de barbatimão ((Stryphnodendron adstringens (Mart.) Coville (Leguminosae)). Revista Árvore, Viçosa, v.32, n.4, p.633-639, 2008.
MARTINS, C.C.; NAKAGAWA, J; BOVI, M.L.A. Efeito da posição da semente no substrato e no crescimento inicial das plântulas de palmito-vermelho (Euterpe espiritosantensis Fernandes – Palmae). Revista Brasileira de Sementes, Brasília, v.21, n.1, p.164-173, 1999.
MARTINS, R.C.C. Germinação e crescimento inicial de três espécies pioneiras do bioma Cerrado no Distrito Federal, Brasil. 2004. 141f. Tese (Doutorado em Ciência Florestal) - Universidade Federal de Viçosa, Viçosa-MG, 2004.
MASLE, J.; DOUSSINALUT, G.; FARQUHAR, G.D.; SUN, B. Foliar stage in wheat correlates better to photothermal time than to thermal time. Plant, Cell and Environment, Oxford, v.12, n.3, p.235-247, 1989.
MAYER, A.M.; POLJAKOFF-MAYER, A. The Germination of Seeds. 4th ed. Oxford: Pergamon Press, 1989. 270p.
MCMASTER, G.S.; WILHELM, W.W. Growing degree-days: one equation, two interpretations. Agricultural and Forest Meteorology, Amsterdam, v.87, n.4, p.291-300, 1997.
MCMASTER, G.S; WILHELM, W.W.; MORGAN, J.A. Simulating winter wheat shoot apex phenology. Journal of Agricultural Science, Cambridge, v.119, p.1-12, 1992.
MENDONÇA, R.; FELFILI, J.M.; WALTER, B.M.T.; SILVA JÚNIOR, M.C.; REZENDE, A.V.; FILGUEIRAS, T.S.; NOGUEIRA, P.E.N. Flora vascular do Cerrado. In: SANO, S.; ALMEIDA, S. (eds.). Cerrado: ambiente e flora. EMBRAPA-CPAC, Planaltina, 1998. p.287-556.

108
MENEZES, N.L. de; FRANZIN, S.M.; ROVERSI, T.; NUNES, P.E. Germinação de sementes de Salvia splendens Sellow em diferentes temperaturas e qualidades de luz. Revista Brasileira de Sementes, Brasília, v.26, n.1, p.32-37, 2004.
MEYER, B.; ANDERSON, O; BOHING, R. Introdução à Fisiologia Vegetal, 1.ª Ed., Fundação Calouste Gulbenkian, Lisboa, 564p. 1965.
MIRANDA, P.R.M.; FERRAZ, I.D.K. Efeito da temperatura na germinação de sementes e morfologia da plântula de Maquira sclerophylla (Ducke). Revista Brasileira Botânica, São Paulo, v.22, n.2, p.303-307, 1999.
MONDO, V.H. Vaz et al. Teste de germinação de sementes de Parapiptadenia rigida (Benth.) brenan (Fabaceae). Revista Brasileira de Sementes, Brasília, v.30, n.2, p.177-183, 2008.
MORAES, C.R.A.; MODOLO, V.A.; CASTRO, P.R.C. Fisiologia da germinação e dominância apical. In: CASTRO, P.R.C.; DE SENA, J.O.A.; KLUGE, R.A. (Ed.) Introdução à fisiologia do desenvolvimento vegetal. Maringá:Eduem, 2002. p.159-178.
MOTULSKY, H.; CHRISTOPOULOS, A. Fitting models to biological data using linear and nonlinear regression. Oxford University Press. Oxford. 2004, 351p.
NAKAGAWA, J. Testes de vigor baseados no desempenho de plântulas. In: KRZYZANOWSKI, F.C.; VIEIRA, R.D.; FRANÇA NETO, J. de B. Vigor de sementes: conceitos e testes. Londrina: ABRATES, 1999. p.2.1-2.24.
NASCIMENTO, W.M.O.; CARVALHO, J.E.U.E; CARVALHO, N.M. Germinação de sementes de jenipapo (Genipa americana L.) submetidas a diferentes temperaturas e substratos. Revista Brasileira Fruticultura, Jaboticabal, v.22, n.3, p.471-473, 2000.
NASCIMENTO, W.M.O.; RAMOS, N.P.; CARPI, V.A.F.; SCARPARE FILHO, J.A.; CRUZ, E.D. Temperatura e substrato para germinação de sementes de Parkia platycephala Benth. (Leguminosae-Caesalpinoideae). Revista Agricultura Tropical, Cuiabá, v.7, n.1, p.119-129, 2003.
NASSIF, S.M.L.; VIEIRA, I.G.; FERNADES, G.D. (LARGEA/). Fatores externos (ambientais) que influenciam na germinação de sementes. Piracicaba: IPEF/LCF/ESALQ/USP, Informativo Sementes IPEF, Abr-1998. Disponível em: <Http://www.ipef.br/sementes/>. Acesso em: 12/jan/2009.
NOVAES, W. Ruim com ele, pior sem ele. O Estado de São Paulo, 21 dez. 2007.
NOVEMBRE, A.D.; LUZ C. et al. Teste de germinação de sementes de sansão-do-campo Mimosa caesalpiniaefolia Benth. - Fabaceae-

109
mimosoideae). Revista Brasileira de Sementes, Brasília, v.29, n.3, p.47-51, 2007.
NUTTONSON, M.W. Some preliminary observations of phenological data as a tool in the study of photoperiodic and thermal requirements of various plant materials. In: Vernalization and photoperiodism. MURNEEK; A.E.; WHYTE, R.O. (ed.). Chronica Botanica. Waltham, Mass.1948. pp. 129-143.
OLIVEIRA, A.P.; NIENOW, A.A.; CALVETE, E.O. Capacidade de enraizamento de estacas semilenhosas de cultivares de pessegueiro tratados com AIB. Revista Brasileira Fruticultura, Jaboticabal, v.25, n.2, p.282- 285, 2003.
OLSEN, J.K.; MCMAHON, C.R.; HAMMER, G.L. Prediction of sweet maize phenology in subtropical environments. Agronomy Journal, Madison, v.85, n.2, p.410-415, 1993.
OMAGA, P.A.; SUMMERFIELD, R.J.; QI, A. Flowering of pigeonpea (Cajanus cajan) in Kenya: responses of early-maturing genotypes to location and date of sowing. Field Crops Research, Amsterdam, v.41, n.1, p.25-34, 1995.
OMAGA, P.A.; SUMMERFIELD, R.J.; QI, A. Flowering of pigeonpea (Cajanus cajan) in Kenya: responses of medium and late maturing genotypes to location and date of sowing. Experimental Agriculture, v.32, n.2, p.111-128, 1996.
OMETTO, J.C. Fluxo de calor sensível ou o calor destinado a aquecer o ar atmosférico. In: Bioclimatologia Vegetal. J.C. Ometto. Agronômica Ceres, São Paulo. p.129-155, 1981.
ONO, E.O.; RODRIGUES, J.D.; PINHO, S.Z. Interações entre auxinas e ácido bórico, no enraizamento de estacas caulinares de Coffea arabica L. cv. Mundo Novo. Scientia Agricola, Piracicaba, v.49, n.1, p.23-27, 1992.
PAIVA, R.; OLIVEIRA, L.M. Fisiologia e produção vegetal. Lavras: Ed. UFLA, 2006. 104p.
PARDINI, F. Além da Floresta. Página 22, n.18, abr.2008. Disponível em: <www.pagina22.com.br/index.cfm?fuseaction=homeReportagens.>
POTT, A.; POTT, V.J. Plantas do pantanal. Empresa Agropecuária, EMBRAPA. Centro de Pesquisa Agropecuária do Pantanal. Corumbá, MS, 1994, 320p.
RAMIN, A.A. The influence of temperature on germination of taree Irani (Allium ampeloprasum L. spp iranicum W.). Seed Science and Technology, Zurich, v.25, n.3, p.419-426, 1997.

110
RAMOS, M.B.P.; VARELA, V.P.; MELO, M.F.F. Influência da temperatura e da água sobre a germinação de sementes de Paricá (Schizolobium amazonicum Huber ex Ducke Leguminosae-Caesalpinioideae). Revista Brasileira Sementes, Pelotas, v.28, n.1, p.163-168, 2006.
RANIERI, B.D.; LANA, T.C. NEGREIROS, D.; ARAÚJO, L.M.; FERNANDES, G.W. Germinação de sementes de Lavoisiera cordata Cogn. e Lavoisiera francavillana Cogn. (Melastomataceae), espécies simpátricas da Serra do Cipó, Brasil. Acta Botânica Brasilica, São Paulo, v.17, n.4, p.523-530, 2003.
RAVEN, P.H.; EVERT, R.F.; EICHHORN, S.E. O processo de evolução. In: RAVEN, P.H.; EVERT, R. F.; EICHHORN, S.E. (orgs). Biologia Vegetal. 6ª ed. Rio de Janeiro: Guanabara Koogan, p.229-251. 2001.
RIBEIRO, F.J.; WALTER, B.M.T. Fitofisionomias do Bioma Cerrado. p. 89-166. In: SANO, S.M.; ALMEIDA, S.P. (eds.). Cerrado: Ambiente e Flora. Planaltina: EMBRAPA-CPAC, 1998.
RIZZINI, C.T. Árvores e madeiras úteis do Brasil. Manual de dendrologia brasileira. 2 ed.São Paulo: Edgard Blücher, 1990. 296p.
ROBERTS, E.H.; SUMMERFIELD, R.J. Measurement and prediction of flowering in annual crops. In: Manipulation of flowering. ATHERTON, J.G. (ed.). Butterworths, London.1987. pp.17-50.
ROESLER, R.; MALTA, L.G.; CARRASCO, L.C.; HOLANDA, R.B.; SOUSA, C.A.S.; PASTORE G.M. Atividade antioxidante de frutas do cerrado. Ciência e Tecnologia de Alimentos, Campinas, v.27, n.1, p.53-60, 2007.
SAGAN, C.; TOON, O.B.; POLLACK, J.B. Anthropogenic albedo changes and the earth’s climate. Science, v.206, n.4425, p.1363-1368, 1979.
SALOMÃO, A.N.; SOUSA-SILVA, J.C.; DAVIDE, A.C.; GONZÁLES, S.; TORRES, R.A.A.; WETZEL, M.M.V.S.; FIRETTI, F.; CALDAS, L.S. Germinação de sementes e produção de mudas de plantas do Cerrado. Rede de Sementes do Cerrado, Brasília. 2003. 96p.
SANTOS, D.L.; SUGAHARA, V.Y.; TAKAKI, M. Efeitos da luz e temperatura na germinação de sementes de Tabebuia serratifolia (Vahl) Nich, Tabebuia chrysotricha (Mart. Ex DC.) Standl. e Tabebuia róseo-alva (Ridl) Sand – Bignoniaceae. Ciência Florestal, Santa Maria, v.15, n.1, p.87-92, 2005.
SCHERER, M. Mensagem da garrafa. Página 22, n.18, abr. 2008. Disponível em: www.pagina22.com.br/index. cfm?fuseaction=homeReportagens.
SILVA JUNIOR, M.C. 100 Árvores do Cerrado: guia de campo. Ed. Rede de Sementes do Cerrado, Brasília, 2005. 278p.

111
SILVA, A.C.; ARAUJO, E.F. e FERREIRA, F.A. Períodos e temperaturas de pré-resfriamento na germinação de sementes de Leonotis nepetaefolia. Ciência Rural, Santa Maria, v.36, n.1, p.282-285, 2006.
SILVA, B.M.S.; CESARINO, F.; LIMA, J.D.; PANTOJA, T.F.; MORO, F.V. Germinação de sementes e emergência de plântulas de Oenocarpus minor Mart. (Arecaceae). Revista Brasileira de Fruticultura, Jaboticabal, v.28, n.2, p.289-292, 2006a.
SILVA, M.N.F.; VALENTE, M.C. Flora da Reserva Ducke, Amazonas, Brasil: Combretaceae. Rodriguésia, Rio de Janeiro, v.56, n.86, p.131-140. 2005.
SOCOLOWSKI, F.; TAKAKI, M. Germinação de sementes e emergência de plântulas de Tabebuia rosea (Bertoloni) a.p. de CANDOLLE (Bignoniaceae), uma espécie exótica com potencial invasor. Revista Árvore, Viçosa-MG, v.31, n.2, p.229-238, 2007.
SOUZA, D.M.G. et al. Manejo da adubação fosfatada em pastagens. In: SIMPÓSIO SOBRE MANEJO DA PASTAGEM, 21, 2004. Piracicaba. Anais... Piracicaba: Fealq, 2004. p. 101-138.
SOUZA, L.A.G.; SILVA, M.F.; MOREIRA, F.W. Capacidade de nodulação de cem leguminosas da Amazônia. Acta Amazônica, Manaus, v.24, n.1/2, p.9-18, 1994.
STEINMAUS, S.J.; PRATHER, T.S.; HOLT, J.S. Estimation of base temperatures for nine weed species. Journal of Experimental Botany, Oxford, v.51, n.343, p.275-286, 2000.
STIER, H.S. A physiological study of growth and fruiting in the tomato (Lycopersicon esculentum) with reference to the effect of climatic and edaphic conditions. 1939. University of Md. (Ph.D. thesis).1939.
STOCKMAN, A.L.; BRANCALION, P.H.S.; NOVEMBRE, A.D.L.C.; CHAMMA, H.M.C.P. Sementes de ipê-branco (Tabebuia roseo-alba (Ridl.) Sand. - Bignoniaceae): temperatura e substrato para o teste de germinação. Revista Brasileira de Sementes, Brasília, v.29, n.3, p.139-143, 2007.
STRINGHETA, A.C.O.; SILVA, D.J.H.; CARDOSO, A.A. FONTES, L.E.F.; BARBOSA, G.B. Germinação de sementes e sobrevivência das plântulas de Tillandsia geminiflora Brongn, em diferentes substratos. Acta Scientiarum Agronomy, Maringá, v. 27, n.1, p.165-170, 2005.
SUMMERFIELD, R.J.; ROBERTS, E.H. Effects of illuminance on flowering in long- and short-day grain legumes: a reappraisal and unifying model. In: Manipulation of Flowering. J.G. Atherton (ed). Butterworths, London.1987. p.203-223.

112
TAGNIN, R; MATTES, D. Conversão dos biomas remanescentes em agrocombustíveis e alimentos: queimar ou comer? INTERFACEHS – Revista de Gestão Integrada em Saúde do Trabalho e Meio Ambiente, Santo Amaro, v.3, n.2, p.1-10, 2008. Disponível em: www.interfacehs.sp.senac.br/br/artigos. Acesso em 12.12.2008.
TIGRE, C.B. Silvicultura para as matas xerófilas. Fortaleza: DNOCS, 1968. 175p.
TIMSON, J. New methods of recording germination data. Nature, v.207, p.216-217.
TOWNSEND, C.E.; McGINNIES, W.J. Mechanical scarification of cicer milkvetch (Astragabus cicer L.) seed. Crop Science, Madison, v.12, p.392-394, 1972.
TRUDGILL, D.L.; HONEK, A.; LI, D.; VAN STRAALEN, N.M. Thermal time – concepts and utility. Annals of Applied Biology, London, v.146, n.1, p.1-14, 2005.
VALADARES, J; PAULA, R.C.. Temperaturas para germinação de sementes de Poecilanthe parviflora Bentham (Fabaceae - Faboideae). Revista Brasileira de Sementes, Brasília, v.30, n.2, p.164-170, 2008.
VARELA, V.P.; BROCKI, E.E.; SÁ, S.T.V. Tratamentos pré-germinativos de sementes de espécies florestais da Amazônia: IV. Faveira camuzê - Stryphnodendron pulcherrimum (Willd.) Hochr Leguminosae. Revista Brasileira Sementes, Brasília, v.13, n.2, p.87-90, 1991.
VELTEN, S.; GARCIA, Q. Efeitos da luz e da temperatura na germinação de sementes de Eremanthus (Asteraceae), ocorrentes na Serra do Cipó, MG, Brasil. Acta Botânica Brasilica, São Paulo, v.19, n.4, p.753-761, 2005.
WILHELM, W.W.; MCMASTER, G.S. The importance of phyllochron in studying the development of grasses. Crop Science, Madison, v.35, n.1, p.1-3, 1995.
YAN, W.; HUNT, L.A. An equation for modelling the temperature response of plants using only the cardinal temperatures. Annals of Botany, Oxford, v.84, n.5, p.607-614, 1999.
YIN, X.; KROPFF, M.J.; MCLAREN, G.; VISPERAS, R.M. A nonlinear model for crop development as a function of temperature. Agricultural and Forest Meteorology, Amsterdam, v.77, n.5, p.1-16, 1995.
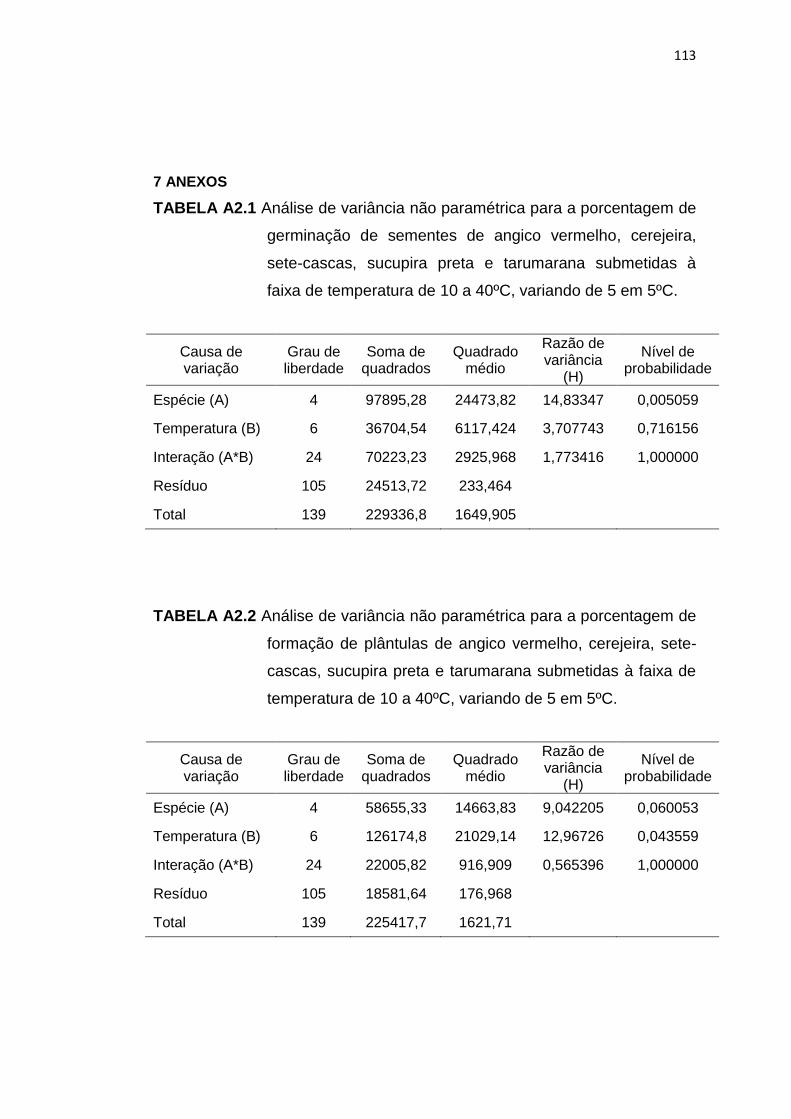
113
7 ANEXOS
TABELA A2.1 Análise de variância não paramétrica para a porcentagem de
germinação de sementes de angico vermelho, cerejeira,
sete-cascas, sucupira preta e tarumarana submetidas à
faixa de temperatura de 10 a 40ºC, variando de 5 em 5ºC.
Causa de variação
Grau de liberdade
Soma de quadrados
Quadrado médio
Razão de variância
(H)
Nível de probabilidade
Espécie (A) 4 97895,28 24473,82 14,83347 0,005059
Temperatura (B) 6 36704,54 6117,424 3,707743 0,716156
Interação (A*B) 24 70223,23 2925,968 1,773416 1,000000
Resíduo 105 24513,72 233,464
Total 139 229336,8 1649,905
TABELA A2.2 Análise de variância não paramétrica para a porcentagem de
formação de plântulas de angico vermelho, cerejeira, sete-
cascas, sucupira preta e tarumarana submetidas à faixa de
temperatura de 10 a 40ºC, variando de 5 em 5ºC.
Causa de variação
Grau de liberdade
Soma de quadrados
Quadrado médio
Razão de variância
(H)
Nível de probabilidade
Espécie (A) 4 58655,33 14663,83 9,042205 0,060053
Temperatura (B) 6 126174,8 21029,14 12,96726 0,043559
Interação (A*B) 24 22005,82 916,909 0,565396 1,000000
Resíduo 105 18581,64 176,968
Total 139 225417,7 1621,71
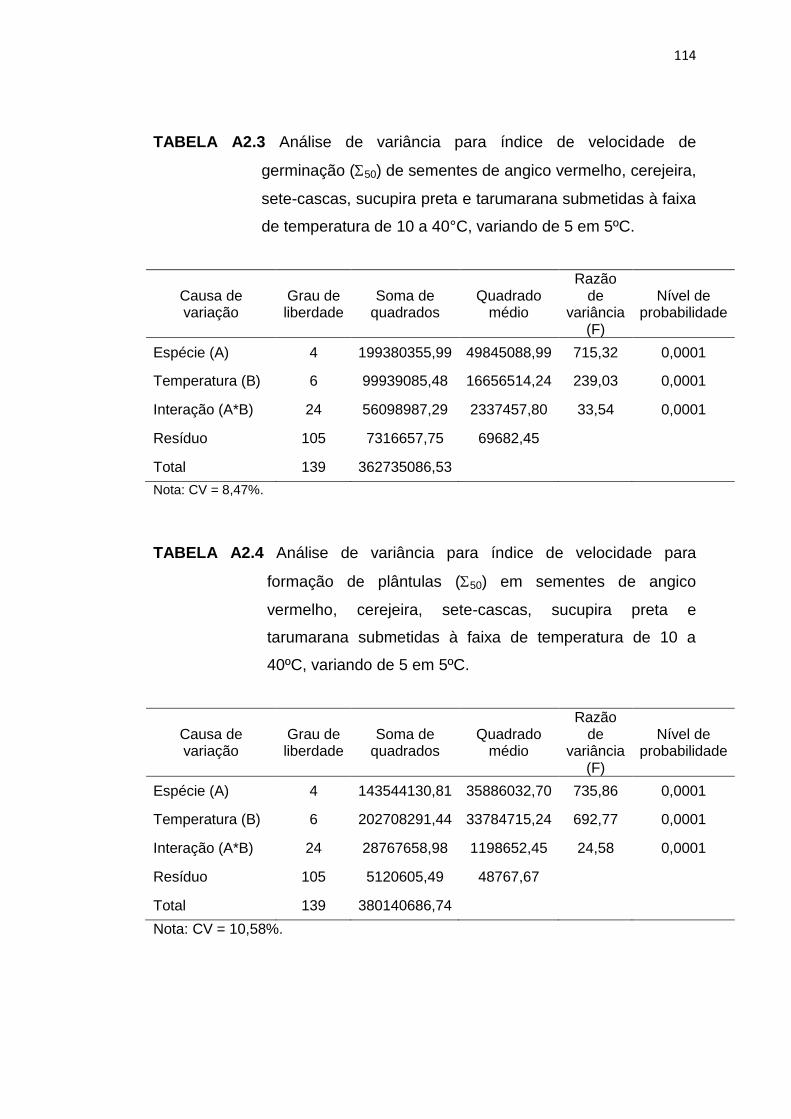
114
TABELA A2.3 Análise de variância para índice de velocidade de
germinação (50) de sementes de angico vermelho, cerejeira,
sete-cascas, sucupira preta e tarumarana submetidas à faixa
de temperatura de 10 a 40°C, variando de 5 em 5ºC.
Causa de variação
Grau de liberdade
Soma de quadrados
Quadrado médio
Razão de
variância (F)
Nível de probabilidade
Espécie (A) 4 199380355,99 49845088,99 715,32 0,0001
Temperatura (B) 6 99939085,48 16656514,24 239,03 0,0001
Interação (A*B) 24 56098987,29 2337457,80 33,54 0,0001
Resíduo 105 7316657,75 69682,45
Total 139 362735086,53
Nota: CV = 8,47%.
TABELA A2.4 Análise de variância para índice de velocidade para
formação de plântulas (50) em sementes de angico
vermelho, cerejeira, sete-cascas, sucupira preta e
tarumarana submetidas à faixa de temperatura de 10 a
40ºC, variando de 5 em 5ºC.
Causa de variação
Grau de liberdade
Soma de quadrados
Quadrado médio
Razão de
variância (F)
Nível de probabilidade
Espécie (A) 4 143544130,81 35886032,70 735,86 0,0001
Temperatura (B) 6 202708291,44 33784715,24 692,77 0,0001
Interação (A*B) 24 28767658,98 1198652,45 24,58 0,0001
Resíduo 105 5120605,49 48767,67
Total 139 380140686,74
Nota: CV = 10,58%.


















