
COMPOSIÇÃO FLORÍSTICA E ESTRUTURA DA VEGETAÇÃO … · A Sidrô nio Bastos, Ivan Amorim e Carla...
92
Alessandra Terezinha Chaves Cotrim Reis COMPOSIÇÃO FLORÍSTICA E ESTRUTURA DA VEGETAÇÃO DE VEREDAS DO CERRADO NO OESTE DA BAHIA, BRASIL FEIRA DE SANTANA –BAHIA 2008
Transcript of COMPOSIÇÃO FLORÍSTICA E ESTRUTURA DA VEGETAÇÃO … · A Sidrô nio Bastos, Ivan Amorim e Carla...

Alessandra Terezinha Chaves Cotr im Reis
COMPOSIÇÃO FLORÍSTICA E ESTRUTURA DA VEGETAÇÃO DE VEREDAS DO CERRADO NO
OESTE DA BAHIA, BRASIL
FEIRA DE SANTANA – BAHIA
2008

10
UNIVERSIDADE ESTADUAL DE FEIRA DE SANTANA
DEPARTAMENTO DE C IÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓSGRADUAÇÃO EM BOTÂNICA
COMPOSIÇÃO FLORÍSTICA E ESTRUTURA DA VEGETAÇÃO DE VEREDAS DO CERRADO NO
OESTE DA BAHIA, BRASIL
Alessandra Terezinha Chaves Cotr im Reis
Dissertação apresentada ao Programa de Pós
Graduação em Botânica da Universidade
Estadual de Feira de Santana como parte dos
requisitos para a obtenção do título de Mestre
em Botânica.
ORIENTADOR: DR. FLÁVIO FRANÇA (UEFS)
COORIENTADOR: DR. ABEL CONCEIÇÃO (UEFS)
FEIRA DE SANTANA – BAHIA
2008

11
Ficha Catalográfica: Biblioteca Central Julieta Carteado

12
BANCA EXAMINADORA
_____________________________________________
Prof. Dr. Eduardo Van den Berg
_____________________________________________
Prof(a). Dr(a). Carolin Proença
_____________________________________________
Prof. Dr . Flávio França
Orientador e Presidente da Banca
Feira de Santana – BA
2008

13
Aos meus queridos pais Humberto e Marlene, minha irmã Fabíola, minha sobrinha Sophia, a Saul meu esposo e a Manoelito meu filho.
DEDICO

14
Agradecimentos
A Deus por me permitir vencer mais uma etapa em minha vida.
A Universidade Estadual de Feira de Santana (UEFS), especialmente ao Departamento
de Ciências Biológicas, através do Programa de Pósgraduação em Botânica, pela
oportunidade na realização deste curso.
Ao Orientador Prof. Dr. Flavio França do Departamento de Ciências Biológicas da
Universidade Estadual de Feira de Santana (UEFS), pela orientação, apoio, paciência e
sobretudo pela amizade demonstrada ao longo desta jornada.
Ao Coorientador Prof. Dr. Abel Conceição, do Departamento de Ciências Biológicas
da Universidade Estadual de Feira de Santana (UEFS), pelas sugestões, críticas e palavras de
incentivo.
À Profa. Dra. Efigênia Melo, do Laboratório de Taxonomia da UEFS, pela atenção e
apoio durante a realização do trabalho.
Ao MsC. Floriano Pastore pela presteza e atenção que sempre me dispensou, durante a
identificação das plantas. Seu auxilio foi fundamental, muito obrigada.
À Prof. Lenise pela atenção, presteza, alegria e disposição dispensada durante a minha
visita ao Herbário Alexandre Leal Costa (ALCB) UFBA.
À Prof. Dra. Carolin Proença pela atenção dispensada durante a minha visita ao
Herbário UN da UnB.
A Sidrônio Bastos, Ivan Amorim e Carla Fabíola do Instituto de Meio Ambiente da
Bahia, por acreditarem, permitirem e incentivarem a realização deste trabalho.
Aos meus amigos da Universidade do Estado da Bahia (UNEB), Campus IX,
Barreiras, Glauciana Araújo, Roxana Nascimento, Perla Matos, Bruna Thais e Vinicius Melo,
pelo apoio na realização dos trabalhos de campo, muito obrigada, nunca vou me esquecer de
vocês.
A Jackeline Miclos, Fernanda Paz, Valmir Filho e Renata Damaso, Valmir e Lourdes
Damaso, Cristiana e Jorge Costa obrigada pela força e palavras de incentivo durante a
realização do trabalho.
Aos Professores da Universidade do Estado da Bahia (UNEB), Campus IX/Barreiras,
especialmente a MsC. Rita Sanches, MsC. Charles Leonel e Dr. Joaquim Neto pelo apoio e
incentivo na realização dos trabalhos, principalmente os de campo.

15
Aos meus queridos e inesquecíveis Mestres do CIRCEA em especial Lourdes Souto,
Genivaldo Cardoso, Padre Carlos, Valdemberg Trindade e Irmã Luna.
Aos funcionários da Universidade do Estado da Bahia (UNEB), Campus IX, Barreiras,
em especial a Luza e a Domingas Calistro, Sr. Raimundo, Sr. Carlos, Lauri, que sempre me
incentivaram mesmo com a correria do dia a dia ... não vou me esquecer de vocês.
Aos especialistas pela identificação das espécies e a atenção dispensada: MsC.
Floriano Pastore (UEFS); Dra. Efigênia Melo (UEFS); Dra. Alexa Oliveira Paes Coelho
(UEFS); MsC. Cristiano Menezes (UCSal); Dra. Adilva Souza Conceição (UNEB); Maria
Lenise Guedes (UFBA); MsC. Luisa Senna (UEFS/UNEB); MsC. Andreia Karla Santos
(UEFS/UFBA); MsC. Cosme Correia (UEFS); Dra. Nadia Roque (UFBA); Jacqueline
Miranda Gonçalves (UEFS); Dr. Marcos Sobral (UFMG); Dra. Ana Maria Giulietti (UEFS);
Dra. Tânia Silva (UEFS); Dr. Raymond Herley; Dr. Luciano P. Queiroz (UEFS); Dra. Inês
Cordeiro; Dr. Antônio Luís Toscano de Brito (UEFS); MsC. Daniela CarneiroTorres
(UEFS).
A Adriana de Olinda Moraes da Universidade Estadual de Feira de Santana (UEFS),
pela dedicação, incentivo e atenção dispensada durante toda realização deste trabalho.
Aos colegas da UEFS Aigara Alves, Ana Paula Couto, Círio Santana, Ivania Oliveira,
João Rodrigo, Patrícia Cristina, Paulo Ricardo, Tatiana Cerqueira, muito obrigada pela
atenção que me foi dispensada durante este período.
Aos queridos colegas do Laboratório de Taxonomia (TAXON) da UEFS, Alexa
Oliveira, Claudia Bastos, Jamile Lima, Reinaldo Alves, Samia Neves, Carla de Lima, Leiton
Silva e Fabio Peixoto pela paciência, apoio e incentivo durante a realização de todo o
trabalho.
À Adriana Estrela, secretária da PósGraduação em Botânica/UEFS, pela atenção,
paciência e preocupação em atender todas às nossas solicitações.
A Josimara e Simony Reis pela elaboração dos mapas das áreas trabalhadas.
A Gardênia Maria Araújo Aires e Helton Ricardo Carneiro Júnior, secretário da Pós
Graduação de Biotecnologia/UEFS, por serem sempre prestativos.
Aos funcionários da Universidade Estadual de Feira de Santana (UEFS), em especial
aos que trabalham no LABIO.
Aos funcionários do Herbário HUEFS (UEFS), Msc. Teonildes Sacramento N. Sena,
Maria José e Silvia Carla pela atenção que sempre me dispensaram.
À Maria Clara Ferreira do Instituto Brasileiro de Geografia e Estatística (IBGE) pela
preocupação, incentivo e atenção. Obrigada pela força amiga!

16
À minha família, meu pai Humberto Wagner Pinto Cotrim e minha mãe Marlene
Chaves Cotrim, a minha irmã Fabíola Diana, a minha sobrinha Sophia Maria, que sempre
tiveram do meu lado em todos os momentos, torcendo por mim, me incentivando, e
confortando nos momentos mais difíceis e me mostrando o quanto família e importante para
estrutura de uma pessoa. Muito obrigada família por toda ajuda e apoio! Eu amo vocês muito!
Aos meus avós Abdias e Odete Cotrim (in memoriam), João e Dulce Chaves (in
memoriam); aos meus tios e tias em especial a Dalva Chaves e Epaminondas Guimarães, Maria
da Conceição e Eugênio Ramos, Moises e Celeste Pinto; aos meus padrinhos Lédima e Artur
Matias; aos primos e primas em especial a Virginia Ramos, Andréa e Douglas Guimarães; a
minha sogra Maria Norma e aos meus cunhados; a Maria de Lourdes minha irmã querida; a
Gerolina Aranha, Helena Santana e Ana Ribeiro, queridas companheiras, muito obrigada pelo
incentivo, força, confiança, amor e compreensão, que me tem dispensado ao longo dos anos.
A Saul Reis que ao longo destes 20 anos juntos vem sempre me incentivando em todos
os momentos... E nestes dois últimos anos então..... tenho certeza que sem ele não
conseguiria, pois com tantas dificuldades e correria, ele foi o meu grande companheiro.
Obrigada por estar sempre por perto, nas dificuldades, nas horas de estresse, no nervosismo e
nas alegrias. Pois o seu profissionalismo e dedicação durante as coletas de campo foi
preponderante para conseguirmos realizar este trabalho... acordando de madrugada para
chegar nas áreas de coletas... como são difíceis acessos.. coletou e prensou material, tomou
chuva e dirigiu pelo cerrado em busca das veredas. Muito obrigada por tudo. Obrigada pela
vida que temos juntos e pelo nosso lindo e querido filho Manoelito, que acaba de chegar...eu
sou muito feliz e te amo muito!
A todos aqueles que contribuíram direta ou indiretamente para eu conseguir
concretizar este trabalho, fica o meu sincero agradecimento.
Muito Obrigada!

17
SUMÁRIO
RELAÇÃO DE TABELAS E FIGURAS ................................... i
RELAÇÃO DE SÍMBOLOS E ABREVIAÇÕES ...................... v
1. INTRODUÇÃO ............................................................................. 1
2.
3.
OBJETIVOS ..................................................................................
MATERIAL E MÉTODOS ..........................................................
7
8
3.1. Levantamento Bibliográfico ................................................... 8
3.2. Área de Estudo ....................................................................... 8
3.3. Delineamento do Trabalho de Campo .................................... 11
3.4. Coleta do material e Identificação do Material Botânico ....... 14
3.5. Análises dos dados ................................................................. 17
4. RESULTADOS E DISCUSSÃO .................................................. 19
4.1. Composição Florística e Diversidade ..................................... 19
4.2 Similariadade .......................................................................... 42
4.3 Cobertura ................................................................................ 52
4.4 Hábito e Formas de Vida ....................................................... 54
5. CONSIDERAÇÕES FINAIS ........................................................ 57
6. RESUMO ....................................................................................... 58
7. ABSTRACT ................................................................................... 59
8. REFERÊNCIAS BIBLIOGRÁFICAS ........................................ 66

18
RELAÇÃO DE TABELAS
Tabela 1.
Tabela 2.
Localização das áreas estudadas, onde: Rio de janeiro (RJ), Rio
Branco (RB), Cerrado (CE), Borda (B), Meio (M) e Fundo (F). Oeste
da Bahia, Brasil.
Criptógamas ocorrentes nas parcelas de 10 x 10 m nas veredas Rio de
Janeiro (RJ) e Rio Branco (RB) e em áreas de cerrado circundantes,
Oeste da Bahia, Brasil.
12
22
Tabela 3. Fanerógamas ocorrentes nas parcelas de 10 x 10 m nas veredas Rio
de Janeiro (RJ) e Rio Branco (RB) e em áreas de cerrado
circundantes, Oeste da Bahia, Brasil.
22
Tabela 3. Análise da estrutura da vegetação das áreas de Cerrado e Veredas. 37
RELAÇÃO DE FIGURAS
Figura 1. Localização das áreas de estudo no Nordeste do Brasil (a), com
detalhe da região Oeste do Estado da Bahia, APA Bacia do Rio
de Janeiro, na Bacia do Rio Grande, Bahia, Brasil (b). ................... 9
Figura 2. Mapa de localização da vereda do Rio Branco na Bacia do Rio
Grande, região Oeste da Bahia, Brasil. ............................................ 10
Figura 3. Mapa de localização da vereda do Rio de Janeiro na Bacia do Rio
Grande, região Oeste da Bahia, Brasil. ............................................ 10
Figura 4. Delimitação das áreas de coletas no cerrado e veredas (zonas de
borda, meio e fundo) dos Rios de Janeiro e Branco, Janeiro na
Bacia do Rio Grande, região Oeste da Bahia, Brasil. ...................... 11
Figura 5. Áreas de coletas na vereda do Rio Branco, APA RJ, Bacia do Rio
Grande, Oeste da Bahia, Brasil. ....................................................... 15
Figura 6. Áreas de coletas na vereda do Rio de Janeiro, APA RJ, Bacia do

19
Rio Grande, Oeste da Bahia, Brasil. ................................................. 16
Figura 7. Famílias com maior número de espécies na vereda do Rio Branco,
Bahia, Brasil. .................................................................................... 19
Figura 8. Famílias com maior número de espécies na vereda do Rio de
Janeiro, Bahia, Brasil. ...................................................................... 20
Figura 9. Distribuição de H’ nas áreas de coletas do Rio Branco e Rio de
Janeiro, Bacia do Rio Grande, Bahia, Brasil. ................................... 38
Figura 10. Resultado da Análise de variâncias (Two Way ANOVA), obtido
nas três zonas das veredas e do cerrado adjacente no Rio Branco e
do Rio de Janeiro, Bacia do Rio Grande, Bahia, Brasil. .................. 39
Figura 11. Valores médios de H’, obtidos a partir da analise de variância
(Two way ANOVA) nas áreas de solos mais secos e/ou úmidos,
obtido nas três zonas das veredas e do cerrado adjacente no Rio
Branco e do Rio de Janeiro, Bacia do Rio Grande, Bahia, Brasil. ... 39
Figura 12. Distribuição da variável riqueza obtida a partir do numero de
espécies em cada subquadricula de 2 x 2 m na vereda do Rio
Branco, Bacia do Rio Grande, Bahia, Brasil. ................................... 40
Figura 13. Valores médios da riqueza, obtidos a partir da analise de variância
nas áreas de solos mais secos e/ou úmidos, obtido nas três zonas
das veredas e do cerrado adjacente no Rio Branco, Bacia do Rio
Grande, Bahia, Brasil. ...................................................................... 41
Figura 14. Dendrograma de similaridade florística das áreas de cerrado e
veredas, obtido por uma matriz de presença/ausência das espécies
utilizando UPGMA e o quociente de Jaccard. ................................. 44
Figura 15. Dendrograma de similaridade florística das áreas de cerrado e
veredas obtido por uma matriz de presença/ausência das espécies
utilizando UPGMA e o quociente de Sørensen. ............................... 45
Figura 16. Dendrograma de similaridade florística obtido por uma matriz de
presença/ausência das espécies utilizando UPGMA e o quociente
de Jaccard, do Rio Branco, Bacia do Rio Grande, Oeste da Bahia,
Brasil. ............................................................................................... 47
Figura 17. Dendrograma de similaridade florística obtido por uma matriz de

20
presença/ausência das espécies utilizando UPGMA e o quociente
de Sørensen, do Rio Branco, Bacia do Rio Grande, Oeste da
Bahia, Brasil. ....................................................................................
48
Figura 18. Dendrograma de similaridade florística obtido por uma matriz de
presença/ausência das espécies utilizando UPGMA e o quociente
de Jaccard, do Rio de Janeiro, Bacia do Rio Grande, Oeste da
Bahia, Brasil. .................................................................................... 49
Figura 19. Dendrograma de similaridade florística obtido por uma matriz de
presença/ausência das espécies utilizando UPGMA e o quociente
de Sørensen, do Rio de Janeiro, Bacia do Rio Grande, Oeste da
Bahia, Brasil. .................................................................................... 50
Figura 20. Total de espécimes coletadas e respectivas coberturas, nas veredas
do Rio de Janeiro e do Rio Branco, Bacia do Rio Grande, Oeste da
Bahia, Brasil. .................................................................................... 53
Figura 21. Hábito das plantas encontradas na vereda do Rio de Janeiro, Bacia
do Rio Grande, Oeste da Bahia, Brasil. ............................................ 57
Figura 22. Hábito das plantas encontradas na vereda do Rio do Rio Branco,
Bacia do Rio Grande, Oeste da Bahia, Brasil. ................................. 57
Figura 23. Formas de vida encontradas nas veredas dos Rios de Janeiro e
Branco, Bacia do Rio Grande, Oeste da Bahia, Brasil. .................... 58
Figura 24. Formas de vida encontradas na vereda do Rio de Janeiro, Bacia do
Rio Grande, Oeste da Bahia, Brasil. .............................................. 59
Figura 25. Formas de vida encontradas na vereda do Rio Branco, Bacia do
Rio Grande, Oeste da Bahia, Brasil. ............................................... 59
Figura 26. Formas de vida encontradas nas áreas de coleta (cerrado e vereda)
na nascente do Rio de Janeiro, Bacia do Rio Grande, Oeste da
Bahia, Brasil. .................................................................................... 60
Figura 27. Formas de vida encontradas nas áreas de coleta (cerrado e vereda)
na nascente no Rio Branco, Bacia do Rio Grande, Oeste da Bahia,
Brasil. ............................................................................................... 61
Figura 28. Formas de vida encontradas nas áreas de coleta (cerrado e vereda)
na nascente no Rio Branco e Rio de Janeiro, Bacia do Rio Grande,
Oeste da Bahia, Brasil......................................................................
62

21
RELAÇÃO DE SÍMBOLOS E ABREVIAÇÕES
cm centrímetro (10 2 m). ha Hectare. km kilômetro (10 3 m). km 2 1 000 000 metros quadrados (m 2 ).
º grau(s): medida do ângulo em relação ao equador ou a um meridiano de referência.
ºC grau(s) Celsius. ’ horas(s). ’’ Minuto(s). % percentagem ou porcentagem. S ponto cardeal sul. W ponto cardeal oeste.
CAPES Coordenação de Aperfeiçoamento de Pessoal de nível Superior. EMBRAPA Empresa Brasileira de Pesquisa Agropecuária. APA RJ Área de Proteção Ambiental Bacia do Rio de Janeiro. GPS Sistema de Posicionamento Global. SAD South American Datum (sistema geodésico regional para a América do
Sul/ 1969). APG Angiosperm Phylogeny Group (Grupo para a Filogenia das
Angiospermas). H’ Índice de diversidade de Shannon.
ALCB Herbário Alexandre Leal Costa da Universidade Federal da Bahia (UFBA).
UB Herbário da Universidade de Brasília (UnB). HUEFS Herbário da Universidade Estadual de Feira de Santana (UEFS). Fig. Figura ou figuras consecutivas. Figs. Figuras não consecutivas. RB Rio Branco. RJ Rio de Janeiro. CE Cerrado. M Meio. F Fundo. B Borda. s.l. sensu lato: em sentido amplo. s.s. sensu sricto: em sentido estrito. sp. espécie. spp espécies. Fn Fanerófita Nf Nanofanerófitas Mf Microfanerófita Hp Hemicriptófita Cm Caméfita Tr Terófita Hd Hidrófita Sf Saprófita

22
Reis, Alessandra Terezinha Chaves Cotrim R298c Composição floristica e estrutura da vegetação de veredas do cerrado no
oeste da Bahia, Brasil / Alessandra Terezinha Chaves Cotrim Reis, 2009. 79f. : il.
Orientador: Flavio França Coorientador: Abel Conceição Dissertação (Mestrado em Botânica) – Universidade Estadual de Feira
de Santana, 2008.
1. Veredas. 2. Vegetação – Cerrado – Bahia. 3.Cerrado Bahia. 4. Floristia – Bahia. I. Universidade Estadual de Feira de Santana. II. França, Flavio. III. Conceição, Abel. IV. Título.
CDU : 582

23
1 INTRODUÇÃO
O cerrado (sensu lato) ocorre predominantemente na região CentroOeste do Brasil. Caracterizase por ser um complexo vegetacional que apresenta relações ecológicas e
fisionômicas com outras savanas da América Tropical e de continentes como África e
Austrália (Beard 1953; Cole 1958; Eiten 1972, 1994; Allem & Valls 1987 apud Sano &
Almeida 1998). Suas fisionomias são resultados de um longo desenvolvimento evolutivo,
durante o qual as plantas se adaptaram às condições ambientais impostas (Pinto et al. 1990). Esta vegetação apresenta variações fisionômicoflorísticas consideráveis, desde formações
florestais a campos puramente herbáceos (Coutinho 1978; Ferreira 1980; Eiten 1983, 1990,
1992; Klink et al. 1993). A sua flora é bem característica e diferenciada dos biomas
adjacentes, embora muitas fisionomias compartilhem espécies com outros biomas (Ribeiro et al. 2001). Considerando que em sua maioria é constituído por fisionomias savânicas, sua maior riqueza florística pode se encontrar na flora herbáceosubarbustiva (Castro et al. 1999). Estudos sobre este componente indicam a ocorrência de grandes mudanças na composição
florística em diferentes regiões, demonstrando tratarse de uma flora sensível a variações de
clima, solo e intensidade da antropização (Loefgren 1980; Mantovani & Martins 1993; Felfili
et al. 1998; Filgueiras 2002). Do ponto de vista da diversidade, estudos recentes demonstram que a riqueza florística
encontrada neste bioma pode superar, em muito, o que inicialmente se supunha (Castro et al. 1999); estimativas pouco precisas da riqueza da flora têm sugerido cerca de 2.000 a 5.000
espécies (Ratter et al. 1997; Mendonça et al. 1998; Castro et al. 1999). No entanto, a maioria dos estudos florísticos realizados com base em levantamentos fitossociológicos, considera
principalmente espécies arbustivoarbóreas (Mantovani & Martins 1993). Desta forma, se por
um lado a flora arbustivoarbórea deste bioma é relativamente bem estudada, por outro, o
reduzido número de levantamentos da flora herbáceosubarbustiva restringe seu conhecimento
a poucas localidades (Ratter et al. 1997; Castro et al. 1999; Araújo et al. 2002; Guimarães et al. 2002; Tannus & Assis 2004).

24
Assim, diante da escassez de estudos para esse componente florístico, os dados são
pouco conclusivos (Ratter et al. 1997; Castro et al. 1999; Filgueiras 2002 apud Tannus & Assis 2004).
Entre as fitocenoses consideradas próprias do Cerrado (sensu latu) (cerradão, cerrado senso stricto, campo cerrado, campo sujo e campo limpo), outras fisionomias, como as matas de galeria, veredas e campos úmidos, ocorrem freqüentemente no contexto deste bioma, porém
de forma menos expressiva (Rizzini 1979; Oliveira Filho et al. 1989; Eiten 1992; Ratter et al. 1997; Pivello et al. 1999).
As veredas ocorrem em geral em áreas de nascentes, com elevado nível de umidade do
solo, representando um ecossistema de grande relevância para a região do cerrado, pois
participam do controle do lençol freático desempenhando importante papel no equilíbrio dos
cursos d’água, funcionando como ecossistema ribeirinho do bioma cerrado, importante para
perenização de córregos, ribeirões e rios as jusantes destes sistemas (Araújo et al. 2002; Ramos et al. 2006).
A água presente o ano todo nestes ambientes, representa um importante recurso para
as populações locais, além de ser fator de grande relevância à biodiversidade, constituindo
sítios pontuais de distribuição de diversas espécies da flora e da fauna do cerrado (Castro
1980; Boaventura 1988; Eiten 1994; Resende et al. 1998; Araújo et al. 2002; Pinto et al. 2005). De acordo Hickman (1990), estas áreas se destacam por apresentar importantes
funções ecológicas, entre estas a manutenção da produtividade e diversidade, contribuição
com os processos globais de transferência de gases do efeito estufa (Ewel 1991 apud Oliveira 2005), além dos consideráveis efeitos positivos sobre a redução da contaminação da água,
pois, a flora de áreas de nascentes é extremamente importante, não apenas por fornecer abrigo
e alimento para a fauna silvestre, mas também pela autodepuração das águas, por assimilação
de nutrientes e retenção de sedimentos (Josephon 1992; Hamilton 1993).
Fonseca & Silva (1998), acrescentam ainda, que estes ambientes apresentam valor
paisagístico e papel social, uma vez que pequenas comunidades de agricultores exploram
sustentavelmente os buritis para diversas finalidades. Assim, estas áreas apresentam
significado ecológico, sócioeconômico e estético paisagístico, o que lhe conferem
importância regional, principalmente por se tratarem de ambientes nascedouros das fontes
hídricas do Planalto Central Brasileiro, que abastece as três principais bacias hidrográficas do
Brasil (Ferreira 2005).
A presença marcante da vegetação herbácea, a qual se estabelece em solos temporária
ou permanentemente encharcados, conferem a estas áreas uma fisionomia campestre (Eiten

25
1963, 1992; Couto et al. 1985; Ratter et al. 1997; Pivello et al. 1999; Ribeiro & Walter 1998;
Resende et al. 1998; Araújo et al. 2002). A existência de comunidades vegetais hidrófilas é caracterizada por dois tipos de vegetação: uma herbáceograminosa que ocupa a maior parte
de sua área e a outra arbóreaarbustiva, onde se destaca a palmeira arbórea Mauritia flexuosa (buriti) (Carvalho 1991). De acordo com Ramirez & Brito (1990), os buritizais definem áreas
tropicais brejosas, uma vez que diferem facilmente da vegetação do cerrado. Quando os vales
tornamse mais encaixados e o lençol freático é mais rebaixado, podem ser encontradas nas
veredas junto aos buritizais, ilhas de vegetação lenhosa caracterizando as matas de galeria
(Guimarães et al. 2002; Felfili et al. 2005). De acordo com Castro (1980) & Carvalho (1991)
a ocorrência dessas comunidades vegetais esta condicionada ao afloramento do lençol
freático, decorrente, entre outros fatores, da alternância de permeabilidades das camadas do
solo, em áreas sedimentares do Terciário. Carvalho (1991) ressalta que as veredas apresentam
quatro estágios de desenvolvimento: No estágio um, a área de nascente é ocupada por densa
vegetação herbácea, com predomínio de espécies graminoides e buritis jovens. No segundo
estágio, além da vegetação herbácea surgem os primeiros, arbustos, subarbustos e plantas
vasculares sem sementes. No terceiro estágio surgem às primeiras trepadeiras e algumas
espécies arbóreas começam a se estabelecer ao longo do canal de drenagem; finalmente no
quarto estágio, com o canal de drenagem mais profundo, desenvolvese uma mata de galeria.
As veredas estão localizadas principalmente no Brasil Central (EMBRAPA 1982; Lima
& Queiróz Neto 1996; Guimarães et al. 2002), nas formações areníticas do Chapadão das Gerais (Boaventura 1988); abrangendo as bacias dos rios Paranaíba, São Francisco, Grande,
Tocantins, Araguaia, Paraná, compreendendo as regiões do Triângulo Mineiro, Alto
Paranaíba, Alto São Francisco e Paracatu (Carvalho 1991).
Vários trabalhos procuram relatar sobre estes ambientes (Walter 2006; Almeida & Sano
1998; Ferreira 2005). Alguns se restringem ao estudo dos solos (Couto et al., 1985; Corrêa, 1989; Amaral 1999; Guimarães 2001; EMBRAPA (1982, 1986, 1999); outros procuram
caracterizar solos, água e seus usos (Ramos et al. 2006); Lima & Queiroz Neto (1996), retrata a evolução da paisagem; já Melo (1992) e Ramos (2004) caracterizaram veredas no contexto
geomorfológico. Grande parte dos trabalhos realizados em áreas de veredas referese
estritamente à descrição da paisagem sem detalhar a composição florística (Magalhães 1966;
Boaventura 1978; Ferreira 1980; Carvalho 1991; Eiten 1994), outros trazem abordagens sobre
a fisionomia e estrutura da vegetação (Ribeiro et al. 1983; Almeida et al. 1983; Schiavini & Araújo 1989, Ribeiro & Walter 1998); já Araújo et al. (2002) ressaltam a estrutura das veredas, caracterizando a composição florística, similaridades e a zonação presente nestes

26
ambientes; Guimarães et al. (2002) compara diversidade florística encontrada em veredas antropizada e conservada; Pinto et al. (2005) estudaram diversidade e similaridades entre áreas de nascentes em Minas Gerais; Oliveira (2005) estudou a vegetação em função do gradiente de
umidade do solo, nas zonas de borda meio e fundo em veredas do Triangulo Mineiro; Costa
(2007) relatou sobre zonação no gradiente vegetacional em áreas de cerrado típico, campo sujo
e veredas. No entanto, poucas informações encontramse sistematizadas.
Algumas coletas botânicas realizadas em áreas de veredas estão incluídas em listagens
da flora do bioma Cerrado (sensu lato) (Almeida & Sano 1998; Felfilli 2005; Mendonça et al. 1998); outros estudos florísticos abrangem vários tipos fisionômicos de vegetação de cerrado,
incluindo parcialmente áreas de vereda, também foram realizados por Pereira et al. (1990), Silva Júnior & Felfili (1996) e Mendonça et al. (1998) no Distrito Federal e Brandão & Gavilanes (1994) no Triângulo Mineiro. Muitas vezes, os resultados quanto à composição
florística tem sido incluído em outras fisionomias similares às veredas, como brejo e campo
úmido (Mendonça et al. 1998; Silva Júnior & Felfili 1998; Felfili et al. 2001). Alguns estudos procuram relatar sobre áreas de campos úmidos como fisionomias similares a veredas (Eiten
1963, 1992; Couto et al. 1985; Schiavini & Araújo 1989; Ratter et al. 1997; Pivello et al. 1998; Ribeiro & Walter 1998 apud Tannus & Assis, 2004). Estudos florísticos detalhados sobre as veredas estão disponíveis apenas em algumas localidades (Araújo et al. 2002; Guimarães et al. 2002; Pinto et al. 2005; Rocha Filho & Lomônaco 2006), o que dificulta o reconhecimento da biodiversidade nestes ambientes, uma vez que as informações ainda
encontramse fragmentadas.
Tendo em vista a importância destes ambientes estas áreas são considerados Áreas de
Preservação Permanente (APP), por legislações ambientais federal nº 4.771/65 e estadual nº
6.569/94 (Brasil 1992, 2002, 2006; Bahia 1997). No entanto, mesmo sendo protegidas por lei,
estas áreas vêm sofrendo alterações de natureza antrópica, que em muitos casos se tornam
irreversíveis (Carvalho 1991), devido principalmente a sua baixa resiliência (Boaventura
1988; Felfili et al. 2005). De acordo com Meirelles et al. (2004), as áreas úmidas têm sido ameaçadas por intervenções antrópicas diretas, como desmatamentos e uso abusivo do lençol
freático, o que pode levar a exclusão de espécies naturalmente estabelecidas nestes ambientes.
De acordo com Odum & Sarmento (1998) áreas úmidas em longo prazo serão mais
economicamente valiosa em seu estado natural do que quando drenadas ou modificadas para
outros usos. Conforme Wilson e Keddy (1985), Bertness e Ellison (1987) apud Costa (2007), em ambientes ribeirinhos a freqüência e a magnitude dos distúrbios físicos determinam o
padrão de sucessão e a taxa de substituição de espécies em escala local.

27
Desta forma, uma das ações mais básicas à conservação destes ambientes é o
reconhecimento da diversidade, que pode ser medida pelo número de espécies (riqueza), que
dá mesmo peso a qualquer espécie, seja rara ou comum (Krebs 1986), ou pela abundância
relativa, na tentativa de atribuir pesos diferentes conforme a representatividade das espécies
(Peet 1974), visto que a vegetação não varia somente pela riqueza de espécies, mas também
quanto às composições florísticas, estruturas e predominâncias de cada espécie. A estrutura de
uma comunidade diz respeito aos padrões de composição, riqueza e abundância de espécies e
das forças evolutivas que moldam estes padrões (Cody & Mooney 1978). Assim, para
caracterizar níveis de organizações ecológicas, a fisionomia da vegetação é fator
determinante, pois a fitofisionomia expressa condições do meio abiótico e biótico presente
num dado local. Desta maneira, uma comunidade vegetal deverá apresentar características que
refletem condições do meio físico e químico local. A mudança progressiva nestas
características constitui gradiente ambiental, a uma faixa de transição relacionada a fatores
ambientais como altitude, temperatura, luminosidade, umidade e nutrientes do solo; assim, a
variação da abundância das espécies e o estabelecimento das comunidades é uma resposta a
esses fatores (Mata & Quevedo 1992).
Em ambientes de veredas, o gradiente ambiental é visível podendo ser separado em
três partes do centro à borda: 1) zona central (de fundo) que fica sempre alagada, com solo
permanentemente saturado com água e essencialmente orgânico; 2) zona do meio que fica
parcialmente alagada durante determinado período do ano, com solo mais escuro; 3) zona de
borda, parcialmente em contado com a vegetação do entorno (cerrado s.l.), com solos mais claros e com melhor drenagem (EMBRAPA 1982; Almeida et al. 1983; Araújo et al. 2002).
Na Bahia, estas áreas se encontram na região Oeste do estado e, assim como outras
regiões de domínio do cerrado, são vistas como áreas de expansão agropecuária, fato que
acelera o processo de antropização, comprometendo a diversidade e interferindo o fluxo
hídrico regional, uma vez que se trata de áreas de nascentes de rios fomentadores do Rio
Grande, e afluentes do Rio São Francisco.
Considerando as peculiaridades encontradas nestas áreas e sua importância para o
bioma cerrado, foi realizado um estudo florístico e estrutural comparativo entre duas áreas de
cerrado e veredas no Oeste da Bahia, onde foram realizadas análises de diversidade. Esperase
que os resultados, deste estudo possam subsidiar políticas públicas de preservação ambiental e
projetos de recomposição de áreas degradadas.

28
2 OBJETIVOS
GERAL
Determinar e comparar a diversidade da vegetação em duas veredas na Bacia do Rio Grande,
no município de Barreiras, Bahia.
ESPECÍFICOS
Caracterizar a composição florística destes ambientes;
Avaliar a similaridade florística entre as áreas;
Determinar a estrutura da vegetação.

29
3 METODOLOGIA
3.1. Levantamento Bibliográfico
Previamente às visitas de campo, foi realizada revisão bibliográfica relacionada ao
ambiente de veredas, existente em áreas de domínio de cerrado. A coleta do material que
subsidiou a revisão bibliográfica foi feita em bibliotecas, acervos botânicos, herbários,
dissertações de mestrado e teses doutorado, artigos de revistas, periódicos e acesso à internet
em sites especializados, tais como o portal da CAPES. Foi realizado levantamento dados, para
verificação da distribuição geográfica das espécies.
3.2. Áreas de Estudo
O trabalho de campo foi realizado em duas veredas do Oeste da Bahia, no município de
Barreiras, na Área de Proteção Ambiental Bacia do Rio de Janeiro, a qual abrange os
municípios de Barreiras e Luiz Eduardo Magalhães, com uma área total de 351.300 ha.
(Bahia 2001) (Fig. 1). A vereda do Rio de Janeiro dista cerca de 130 km da sede municipal,
nas coordenadas 11 o 52’49,6”S e 45º49’32,9”W, a 750 m s.n.m, abrangendo uma área com
aproximadamente 6 km 2 ; a vereda do Rio Branco, a cerca de 200 km da sede municipal, nas
coordenadas 11 o 37’12,5”S e 46º02’18,9”W, a 785 m s.n.m., com aproximadamente 5 km 2 . O
clima da região segundo a classificação de Köppen (1948) é Aw (Clima de Savana tropical)
com duas estações bem definidas, com período chuvoso entre os meses outubro a março e
período seco entre os meses de abril a setembro, com médias pluviométricas variando de 1200
mm em regiões do vale a 1700 mm em regiões de cerrado de planalto (EBDA 2006). O solo é
hidromórfico, com transição para Latossolo na medida em que se aproxima da borda. No local
existem grandes empreendimentos agrícolas, sendo que as áreas de coletas encontramse
inseridas em propriedades particulares, nas Fazendas Sete Belo (Vereda do Rio Branco) (Fig.
2) e Bananal (Vereda do Rio de Janeiro) (Fig. 3), respectivamente, com produção de grãos.
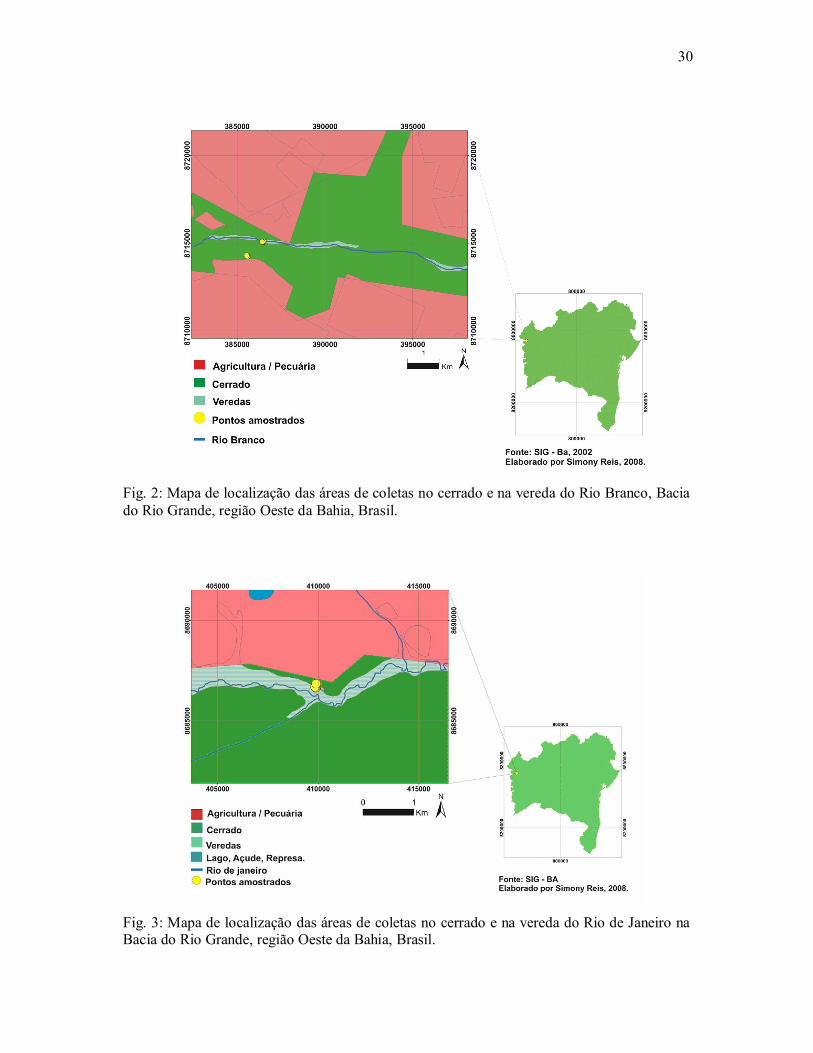
30
Fig. 2: Mapa de localização das áreas de coletas no cerrado e na vereda do Rio Branco, Bacia do Rio Grande, região Oeste da Bahia, Brasil.
Fig. 3: Mapa de localização das áreas de coletas no cerrado e na vereda do Rio de Janeiro na Bacia do Rio Grande, região Oeste da Bahia, Brasil.

3.3. Delineamento do trabalho de campo
Parcelas foram sorteadas no cerrado e em três zonas de cada vereda: borda,
parcialmente em contado com a vegetação do entorno (cerrado sensu lato), meio, que fica parcialmente alagada durante determinado período do ano e fundo, que fica sempre
alagada, em solo essencialmente orgânico permanentemente saturado com água (Fig. 4)
(EMBRAPA 1982; Almeida et al. 1983; Araújo et al. 2002). Inicialmente uma área de 100 m x 100 m foi delimitada em cada vereda utilizandose o mapa da APA Bacia do Rio de
Janeiro, foram incluídas não só a vereda propriamente dita, mas também uma parte da
vegetação do seu entorno (cerrado). A área foi dividida em 10 faixas de 10 m x 100 m e
três dessas faixas foram sorteadas para a realização da amostragem da vegetação. Em cada
uma das faixas sorteadas foram delimitadas quatro parcelas de 10 m x 10 m, perfazendo
um total de 12 em cada vereda: a) três parcelas na vegetação do entorno da vereda (cerrado s.l.); b) três parcelas na área de transição entre a vegetação do entorno e a vegetação da vereda propriamente dita (borda); c) três parcelas dentro da vegetação da vereda onde há
afloramento do lençol freático apenas esporadicamente (meio) e d) três parcelas no centro
da vereda, onde o lençol freático é permanentemente aflorado (fundo) (Tabela 1).
Fig. 4: Delimitação das áreas de coletas no cerrado e veredas (zonas de borda, meio e fundo) do Rio de Janeiro e Rio Branco, na Bacia do Rio Grande, região Oeste da Bahia, Brasil. Fonte: Google Earth 2008.
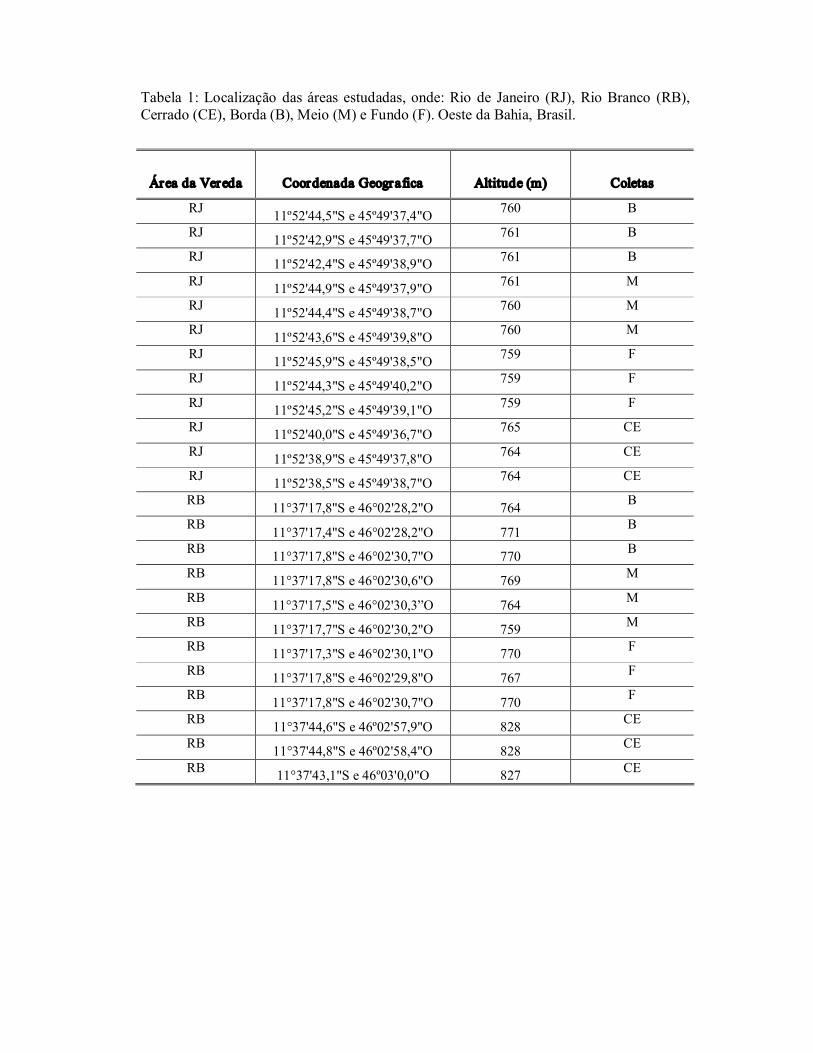
Tabela 1: Localização das áreas estudadas, onde: Rio de Janeiro (RJ), Rio Branco (RB), Cerrado (CE), Borda (B), Meio (M) e Fundo (F). Oeste da Bahia, Brasil.
Área da Vereda Coordenada Geografica Altitude (m) Coletas
RJ 11º52'44,5"S e 45º49'37,4"O 760 B
RJ 11º52'42,9"S e 45º49'37,7"O 761 B
RJ 11º52'42,4"S e 45º49'38,9"O 761 B
RJ 11º52'44,9"S e 45º49'37,9"O 761 M
RJ 11º52'44,4"S e 45º49'38,7"O 760 M
RJ 11º52'43,6"S e 45º49'39,8"O 760 M
RJ 11º52'45,9"S e 45º49'38,5"O 759 F
RJ 11º52'44,3"S e 45º49'40,2"O 759 F
RJ 11º52'45,2"S e 45º49'39,1"O 759 F
RJ 11º52'40,0"S e 45º49'36,7"O 765 CE
RJ 11º52'38,9"S e 45º49'37,8"O 764 CE
RJ 11º52'38,5"S e 45º49'38,7"O 764 CE
RB 11°37'17,8"S e 46°02'28,2"O 764 B
RB 11°37'17,4"S e 46°02'28,2"O 771 B
RB 11°37'17,8"S e 46°02'30,7"O 770 B
RB 11°37'17,8"S e 46°02'30,6"O 769 M
RB 11°37'17,5"S e 46°02'30,3”O 764 M
RB 11°37'17,7"S e 46°02'30,2"O 759 M
RB 11°37'17,3"S e 46°02'30,1"O 770 F
RB 11°37'17,8"S e 46°02'29,8"O 767 F
RB 11°37'17,8"S e 46°02'30,7"O 770 F
RB 11°37'44,6"S e 46º02'57,9"O 828 CE
RB 11°37'44,8"S e 46º02'58,4"O 828 CE
RB 11°37'43,1"S e 46º03'0,0"O 827 CE

As parcelas de 10 m x 10 m sorteadas tiveram as espécies ocorrentes determinadas,
sendo utilizadas para determinação das composições florísticas, riquezas, similariade,
formas de vida e cobertura. As parcelas de 10 m x 10 m foram subdivididas em
subparcelas de 2 m x 2 m (Conceição 2003; Conceição & Pirani 2005), onde foram
sorteadas cinco dessas (Fig. 5 e 6).
Nas subparcelas 2 x 2 m cada cada espécies coletada, se procedeu: a) contagem do
número de espécies; b) identificação da forma de vida de acordo com o sistema de
Raunkiaer (1934), conforme MuellerDombois & Ellenberg (1974), sendo consideradas:
Fanerófitas (Nanofanerófitas (< 2m) e Microfanerófitas (25m) (plantas perenes de gemas
expostas acima de 30 cm do solo), Caméfitas (gemas acima do nível do chão mas apenas
alguns centímetros (130 cm), Hemicriptófitas (mantêm suas gemas rentes ao chão ou seja,
com crescimento prostado no solo), Geófitas (apresentam estrutura vegetativa em baixo do
solo), Terófitas (plantas de ciclo anual) e Hidrófitas (matem suas gemas submersas em
água); sendo que as caméfitas, geófitas, hemicriptófitas e terófitas foram consideradas
como pertencentes ao componente herbáceosubarbustivo, enquanto as fanerófitas foram
consideradas tanto no componente herbáceosubarbustivo como no arbustivoarbóreo
adaptado Tannus & Assis (2006); c) estimativa da cobertura em classes a saber: 010%;
1020%; 2040%; 4060%; 6080% e 80100%, conforme em França et al., (2006) e d) quantificação do número de indivíduos.
Algumas espécies com indivíduos de tamanho reduzido e difícil individualização, em
geral das famílias Droseraceae, Lentibulariaceae, Eriocaulaceae, Xyridaceae, Poaceae e
Cyperaceae, foi estimado a partir de cinco quadrículas de 50 x 50 cm sorteadas em cada
uma das cinco subparcelas de cada subparcela de 2 x 2m.
Nos casos de indivíduos ainda mais reduzidos, uma subquadrícula de 10 x 10 cm foi
sorteada em cada quadrícula. Os indivíduos quantificados nas quadrículas e subquadrículas
foram estimados para cada subparcela considerando que as proporções de indivíduos das
quadrículas e subquadrículas se mantêm nas subparcelas. Apenas nas famílias Poaceae e
Cyperaceae considerouse cada tufo isolável como um indivíduo (França et al. 2006). Expedições mensais foram realizadas para complementação de dados e coleta de
material fértil.

3.4. Coleta e identificação do mater ial botânico
O material botânico foi coletado seguindo as normas gerais do manejo do material
fanerogâmico herborizado (Mori et al. 1989). As coletas das plantas vasculares foram realizadas durante um ano e os testemunhos encontramse depositados no Herbário da
Universidade Estadual de Feira de Santana (HUEFS). Para fins de comparações entre as
parcelas, foram coletadas amostras de ramos foliares, jovem e maduro, de espécies estéreis
(morfotipos). As identificações foram feitas utilizandose chaves analíticas, consultas a
especialistas e/ou através de comparação com outras exsicatas depositadas nos Herbários
da Universidade Estadual de Feira de Santana – HUEFS, Alexandre Leal Costa – HALC e
da Universidade de Brasília UnB – UB. Para organização das espécies foi adotado o
sistema de classificação de APG II (2003), exceto para plantas vasculares sem sementes
que considerou o sistema de Smith et al. (2006). Todas as parcelas foram
georreferenciadas através de GPS, datum SAD 69, em graus.

Fig. 5: Áreas de coletas no cerrado e na vereda do Rio Branco, APA RJ, Bacia do Rio Grande, Oeste da Bahia, Brasil. Onde: a) área de cerrado; b) vereda do Rio Branco; c) zona de borda; d) zona do meio; e) e f) zona de fundo.
e f
a b
c d

Fig. 6: Áreas de coletas na vereda do Rio de Janeiro, APA RJ, Bacia do Rio Grande, Oeste da Bahia, Brasil. Onde: a) área de cerrado; b) vereda do Rio de Janeiro; c) zona de borda; d) zona do meio; e) e f) vereda zona de fundo.
b a
c d
e f

3.5. Análises dos dados
A similaridade florística entre as veredas e suas respectivas zonas foi calculada a
partir da matriz de presença e ausência das espécies nas parcelas de 10 x 10 m, utilizando
se o índice de Jaccard e Sørensen e método UPGMA (Unweighte Pair Group Method with Arithmetic Mean), calculados pelos programas NTSYSPC (Rohlf 1997), SIMQUAL
(Similarity for qualitative Data) e PAST (Hammer et al. 2001). Utilizouse o procedimento de bootstrap (Efron & Tibshirani 1986), a fim de estimar
o coeficiente de variação (CV) dos índices de Similariade de Jaccard e Sørensen nos
dendogramas de similaridade, utilizandose de 1.000 combinações, de acordo com o
procedimento de Tiving et al. (1994).
Os parâmetros de diversidade utilizados foram: riqueza (número de espécies), índice
de ShannonWiener (H’) e a equitabilidade (Krebs 1989; PintoCoelho 2000). Calculado
para cada parcela 10 x 10 m e a partir das quadrículas 2 x 2 m (Conceição, 2003;
Conceição & Pirani 2005).
Os índices de ShannonWiener, assim como testes de diferenças entre os índices
foram calculados pelo programa SDR IV (Seaby & Henderson 2006). Foram feitas
também análises de variância para as áreas comparandose a riqueza e índice de Shannon
Wiener (H’), calculadas pelo Programa Statistica 6.0.
Inicialmente as variáveis foram testadas quanto a normalidade (Kolmogorov
Smirnov, p < 0,05 e homocedastacidade (Levene, p < 0,05) A hipótese testada na ANOVA
foi a da igualdade entre as médias em cada local H0: µRB = µRJ α = 0,05, sendo RB = vereda
do Rio Branco e RJ = vereda do Rio de Janeiro e em cada habitat H0: µCE = µBR = µM = µF α = 0,05, sendo CE = Cerrado, BR = Borda, M = Meio e F = Fundo). No caso de rejeição
da igualdade admitiuse diferença e aplicouse o teste de Tukey a 5% para discernir as
diferenças entre os locais e habitats (α = 0,05). Os resultados estão apresentados em
gráficos, também elaborados pelo Statistica 6.0 (One way ANOVA e Two way ANOVA)
(CallegariJacques 2003).
Quanto não ocoreu a normalidade das variáveis analisadas para as duas veredas
estudadas, processouse o teste para cada veredas em separado. Com relação ao H’ usouse
o índice calculado em cada parcela 10 x 10 m e no caso da riqueza usouse o número o
número de espécies contido nas quadrículas 2 x 2 m. Para a abundância das espécies foi
considerado apenas número de indivíduos encontrados nas quadriculas 2 x 2m.

Foram considerados para o cerrado e as veredas diferentes gradientes ambientais,
onde: as áreas de cerrado e zonas de borda de cada vereda foram considerados ambientes
secos e as zonas do meio e do fundo como ambientes úmidos. Sendo considerados para as
analises do gradiente ambiental ambientes secos e úmidos.
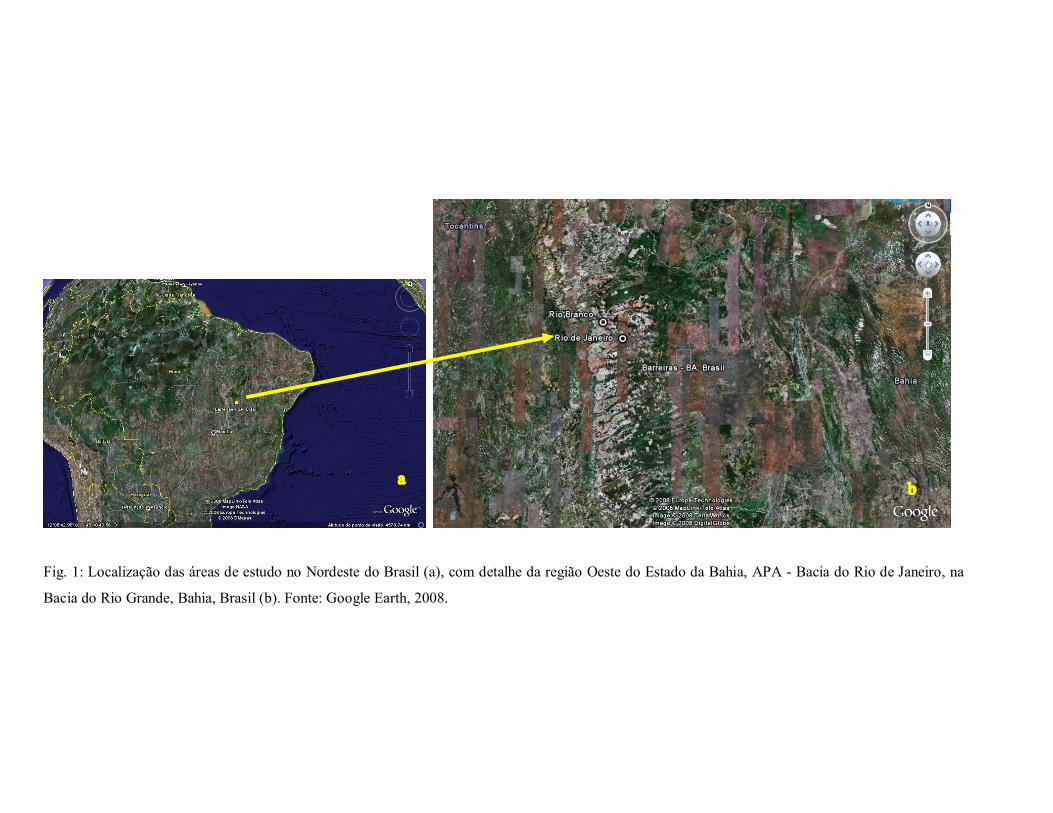
Fig. 1: Localização das áreas de estudo no Nordeste do Brasil (a), com detalhe da região Oeste do Estado da Bahia, APA Bacia do Rio de Janeiro, na
Bacia do Rio Grande, Bahia, Brasil (b). Fonte: Google Earth, 2008.
b a

4 RESULTADOS E DISCUSSÃO
4.1. Composição flor ística e Diversidade
As áreas de veredas do Rio de Janeiro e do Rio Branco apresentam alta
heterogeneidade ambiental e grande diversidade floristica. Foram coletadas nestas áreas 323
espécies, distribuídas em 83 famílias, sendo 3 espécies Criptógamas e 320 Angiospermas, e
apenas 64 comuns às duas veredas. Na vereda do Rio de Janeiro (abreviada por RJ) foram
encontradas 167 espécies distribuídas em 57 famílias; já na vereda do Rio Branco (abreviada
por RB) foram coletadas 226 espécies distribuídas em 58 famílias (Tabelas 2 e 3). As famílias
com maior número de espécies na vereda RB são Fabaceae (37), Poaceae (21), Myrtaceae
(12), Asteraceae (11), Rubiaceae (10), Euphorbiaceae (10) e Cyperaceae (9) (Fig. 7); já na
vereda RJ, as famílias com maior número de espécies são Fabaceae (23), Poaceae (16),
Asteraceae (10), Rubiaceae (10) e Cyperaceae (9) (Fig. 8). Em quatro veredas de Minas
Gerais, as famílias mais ricas em espécies foram Poaceae (64), Asteraceae (63), Cyperaceae
(54), Melastomataceae (27) e Fabaceae (23) (Araújo et al. 2002); já Guimarães et al. (2002) estudando duas veredas em Minas Gerais encontrou maior representatividade para as áreas
úmidas: Poaceae (12), Cyperaceae (20), Lentibulariaceae (9), Eriocaulaceae (8) e Xyridaceae
(7), reforçando a grande importância das famílias Poaceae e Cyperaceae, em ambientes
similares também em Minas Gerais.
23%
13%
7% 7% 6%
6% 6%
32% Fabaceae Poaceae
Myrtaceae
Asteraceae Euphorbiaceae
Rubiaceae
Cyperaceae outras
Fig. 7: Famílias com maior número de espécies na vereda do Rio Branco, Bahia, Brasil.

19%
13%
8% 8% 8%
44%
Fabaceae
Poaceae Asteraceae
Rubiaceae Cyperaceae
Outras
Fig.8: Famílias com maior número de espécies na vereda do Rio de Janeiro, Bahia, Brasil.
Quando se comparam as duas veredas, verificouse que as famílias Alismataceae,
Commelinaceae, Clusiaceae, Salicaceae e Velloziaceae acontecem apenas na vereda RB; já as
famílias Aristolochiaceae, Iridaceae, Sapindaceae, Solanaceae, Urticaceae e Gleicheniaceae,
apresentaram registros apenas na vereda RJ (Tabelas 2 e 3).
Analisando as distintas zonas das veredas, constatase maior riqueza florística entre as
parcelas na zona da borda, onde foram encontradas 219 espécies, sendo Fabaceae a família
mais rica, seguida de Asteraceae, Rubiaceae, Myrtaceae, Poaceae e Cyperaceae. Tais famílias
também foram importantes nessa zona em veredas de Minas Gerais (Araújo et al. 2002),
demonstrando certo padrão florístico. Estudos realizados em bordas de veredas e no cerrado
propriamente dito salientam a importância da família Fabaceae, com grande número de
espécies arbustivas e arbóreas do cerrado (Goodland 1970 apud Araújo et al. 2002; Mantovani & Martins 1993; Castro et al. 1999), sendo importante nas duas veredas estudadas: Bauhinia acuruana, Galactia martii, Vigna firmula, entre outras.
Algumas espécies são restritas a vereda RB: Sagittaria guianensis e Sagittaria rhombifolia, Commelina cf. elegans, Kielmeyera coriacea, Casearia cf. arborea e Vellozia froesii. Outras são restritas a vereda RJ: Aristolochia sp., Sisyrinchium sp. 1, Serjania sp.1), Schwenckia sp. 1, Cecropia pachystachya e Dicranopteris cf. flexuosa.
Nas áreas de borda são comuns as espécies Anacardium humile, Tabebuia ochraceae, Davilla elliptica, Croton didrichsenii e Manihot caerulescens. Nas zonas do meio foram amostradas 132 espécies, merecendo destaque Syngonanthus cf. nitens; já nas áreas de fundo foram amostradas 63 espécies, com destaque à Mauritia flexuosa, uma vez que estas espécies apresentam grande número de indivíduos. Nas áreas de cerrado ocorreram 127 espécies, onde
houve maior representatividade as espécies Annona crassiflora e Annona coriacea. Quando as zonas são individualmente avaliadas, borda apresenta maior número de espécies: 99 em RJ,

sendo exclusivas deste ambiente Annona tomentosa e Baccharis leptocephala; 120 em RB, com destaque à Annona montana, Hymatanthus obovatus, Calea harleyi, Gochnatia oligocephala, Ichthyothere terminalis, Commelina cf. elegans, Curatella americana e Chamaecrista desvauxii, entre outras. Ainda na vereda RB, algumas espécies são comuns às áreas de borda e cerrado, como Calliandra dysantha, Dimorphandra mollis, Copaifera
langsdorffii, Sclerolobium paniculatum e Pterodon cf. emarginatus. As áreas de fundo apresentaram menor número de espécies: 36 na vereda RB e 27 na RJ (Tabela 4).
Do total de plantas coletadas nas zonas do meio e do fundo das duas veredas, 102
espécies (45%) foram coletadas em RB e 93 (55,6%) em RJ, sendo que as demais espécies
foram coletadas nas áreas de cerrado e borda de cada vereda.
Os parâmetros de diversidade nas veredas e respectivas zonas estão na Tabela 4. O
índice de diversidade de Shannon (H’) variou de 1,55 a 3,45 nats/indivíduos; sendo que a área
de borda da vereda do Rio de Janeiro apresentou 3,459 nats/indivíduos e a do Rio de Branco
3,21 nats/indivíduos. Tais áreas correspondem à área de transição com o cerrado, o que
possibilita a ocorrência de algumas espécies de cerrado nessa zona.
Tabela 2: Espécies de Criptógamas de ocorrência nas parcelas de 10 x 10 m nas veredas do Rio de Janeiro (RJ) e do Rio Branco (RB) e em áreas de cerrado circundantes. Oeste da Bahia, Brasil. * A. Cotrim et al.
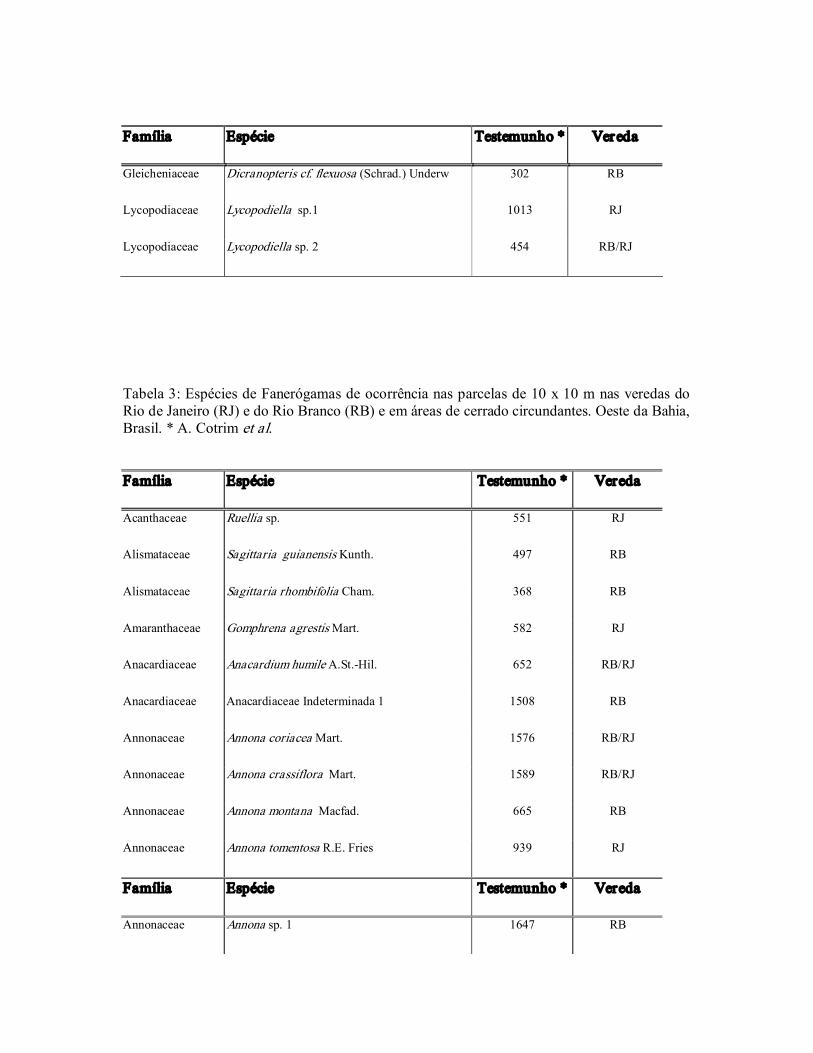
Família Espécie Testemunho * Vereda
Gleicheniaceae Dicranopteris cf. flexuosa (Schrad.) Underw 302 RB
Lycopodiaceae Lycopodiella sp.1 1013 RJ
Lycopodiaceae Lycopodiella sp. 2 454 RB/RJ
Tabela 3: Espécies de Fanerógamas de ocorrência nas parcelas de 10 x 10 m nas veredas do Rio de Janeiro (RJ) e do Rio Branco (RB) e em áreas de cerrado circundantes. Oeste da Bahia, Brasil. * A. Cotrim et al.
Família Espécie Testemunho * Vereda
Acanthaceae Ruellia sp. 551 RJ
Alismataceae Sagittaria guianensis Kunth. 497 RB
Alismataceae Sagittaria rhombifolia Cham. 368 RB
Amaranthaceae Gomphrena agrestis Mart. 582 RJ
Anacardiaceae Anacardium humile A.St.Hil. 652 RB/RJ
Anacardiaceae Anacardiaceae Indeterminada 1 1508 RB
Annonaceae Annona coriacea Mart. 1576 RB/RJ
Annonaceae Annona crassiflora Mart. 1589 RB/RJ
Annonaceae Annona montana Macfad. 665 RB
Annonaceae Annona tomentosa R.E. Fries 939 RJ
Família Espécie Testemunho * Vereda
Annonaceae Annona sp. 1 1647 RB
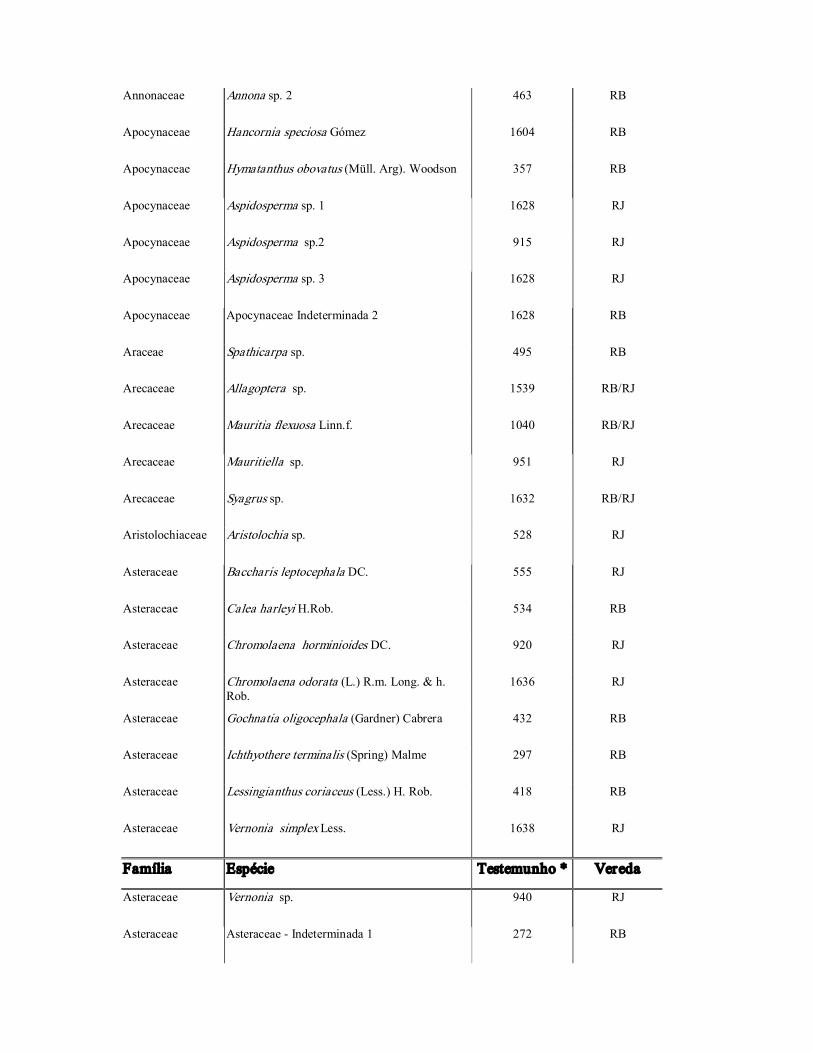
Annonaceae Annona sp. 2 463 RB
Apocynaceae Hancornia speciosa Gómez 1604 RB
Apocynaceae Hymatanthus obovatus (Müll. Arg). Woodson 357 RB
Apocynaceae Aspidosperma sp. 1 1628 RJ
Apocynaceae Aspidosperma sp.2 915 RJ
Apocynaceae Aspidosperma sp. 3 1628 RJ
Apocynaceae Apocynaceae Indeterminada 2 1628 RB
Araceae Spathicarpa sp. 495 RB
Arecaceae Allagoptera sp. 1539 RB/RJ
Arecaceae Mauritia flexuosa Linn.f. 1040 RB/RJ
Arecaceae Mauritiella sp. 951 RJ
Arecaceae Syagrus sp. 1632 RB/RJ
Aristolochiaceae Aristolochia sp. 528 RJ
Asteraceae Baccharis leptocephala DC. 555 RJ
Asteraceae Calea harleyi H.Rob. 534 RB
Asteraceae Chromolaena horminioides DC. 920 RJ
Asteraceae Chromolaena odorata (L.) R.m. Long. & h. Rob.
1636 RJ
Asteraceae Gochnatia oligocephala (Gardner) Cabrera 432 RB
Asteraceae Ichthyothere terminalis (Spring) Malme 297 RB
Asteraceae Lessingianthus coriaceus (Less.) H. Rob. 418 RB
Asteraceae Vernonia simplex Less. 1638 RJ
Família Espécie Testemunho * Vereda
Asteraceae Vernonia sp. 940 RJ
Asteraceae Asteraceae Indeterminada 1 272 RB
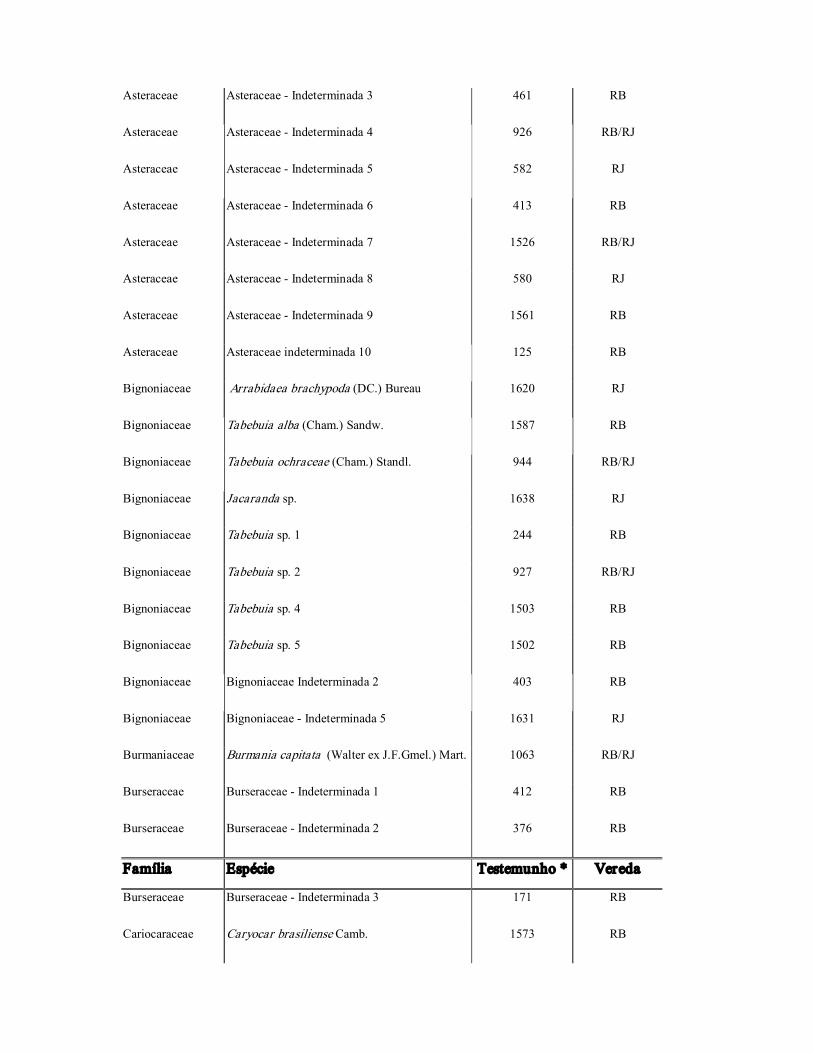
Asteraceae Asteraceae Indeterminada 3 461 RB
Asteraceae Asteraceae Indeterminada 4 926 RB/RJ
Asteraceae Asteraceae Indeterminada 5 582 RJ
Asteraceae Asteraceae Indeterminada 6 413 RB
Asteraceae Asteraceae Indeterminada 7 1526 RB/RJ
Asteraceae Asteraceae Indeterminada 8 580 RJ
Asteraceae Asteraceae Indeterminada 9 1561 RB
Asteraceae Asteraceae indeterminada 10 125 RB
Bignoniaceae Arrabidaea brachypoda (DC.) Bureau 1620 RJ
Bignoniaceae Tabebuia alba (Cham.) Sandw. 1587 RB
Bignoniaceae Tabebuia ochraceae (Cham.) Standl. 944 RB/RJ
Bignoniaceae Jacaranda sp. 1638 RJ
Bignoniaceae Tabebuia sp. 1 244 RB
Bignoniaceae Tabebuia sp. 2 927 RB/RJ
Bignoniaceae Tabebuia sp. 4 1503 RB
Bignoniaceae Tabebuia sp. 5 1502 RB
Bignoniaceae Bignoniaceae Indeterminada 2 403 RB
Bignoniaceae Bignoniaceae Indeterminada 5 1631 RJ
Burmaniaceae Burmania capitata (Walter ex J.F.Gmel.) Mart. 1063 RB/RJ
Burseraceae Burseraceae Indeterminada 1 412 RB
Burseraceae Burseraceae Indeterminada 2 376 RB
Família Espécie Testemunho * Vereda
Burseraceae Burseraceae Indeterminada 3 171 RB
Cariocaraceae Caryocar brasiliense Camb. 1573 RB

Celastraceae Celastraceae Indeterminada 1 423 RB
Celastraceae Maytenus sp. 230 RB
Chrysobalanaceae Hirtella ciliata Mart. ex Zucc. 341 RB
Chrysobalanaceae Licania sp. 957 RJ
Chrysobalanaceae Hirtella sp. 312 RB
Chrysobalanaceae Chrysobalanaceae Indeterminada 1 381 RB
Clusiaceae Kielmeyera coriacea (Spreng.) Mart. 305 RB
Combretaceae Terminalia sp. 1534 RB
Commelinaceae Commelina cf. elegans Kunth. 224 RB
Connaraceae Connaraceae Indeterminada 1 938 RB/RJ
Convolvulaceae Merremia digitata (Sprengel) Hall. f. 1516 RB/RJ
Convolvulaceae Convolvulaceae Indeterminada 1 948 RJ
Convolvulaceae Convolvulaceae Indeterminada 2 918 RJ
Cyperaceae Bulbostylis capillaris (L.) C.B. Clarke 1552 RJ
Cyperaceae Bulbostylis lanata (Kunth) C.B. 1010 RJ
Cyperaceae Cyperus cf. sesquifloru s(Torr.) Mattf. 406 RB
Cyperaceae Eleocharis cf. acutangula (Roxb.) Steud. 209 RB
Cyperaceae Fimbristylis sp. 1 239 RB
Cyperaceae Fimbristylis sp. 2 238 RB
Cyperaceae Lagenocarpus rigidus Nees 1055 RB/RJ
Cyperaceae Rhynchospora filiformes Vahl 380 RB
Família Espécie Testemunho * Vereda
Cyperaceae Rhynchospora sp 6 1046 RJ

Cyperaceae Rhynchospora sp. 7 1008 RJ
Cyperaceae Rhynchospora sp. 8 1059 RJ
Cyperaceae Rhynchospora sp. 9 1038 RJ
Cyperaceae Rhynchospora sp. 10 484 RB
Cyperaceae Cyperaceae Indeterminada 2 1029 RB/RJ
Cyperaceae Cyperaceae Indeterminada 3 195 RB
Cyperaceae Cyperaceae Indeterminada 5 1026 RJ
Dilleniaceae Curatella americana L. 420 RB
Dilleniaceae Davilla elliptica A.St.Hil. 537 RB/RJ
Dilleniaceae Davilla sp. 1640 RB/RJ
Droseraceae Drosera comunis A.H.Hil. 1003 RB/RJ
Ebenaceae Dyospyrus sp. 664 RB
Ericaceae Gaylussacia brasiliensis Meisn. 1068 RJ
Ericaceae Ericaceae Indeterminada 1 357 RB
Eriocaulaceae Actinocephalus sp. 1042 RJ
Eriocaulaceae Eriocaulon longifolium Nees ex Kunth. 1056 RB/RJ
Eriocaulaceae Syngonanthus cf. nitens (Bong.) Ruhl. 1017 RB
Eriocaulaceae Syngonanthus xeranthemoides Ruhland 1021 RB
Eriocaulaceae Syngonanthus sp. 501 RB
Eriocaulaceae Eriocaulaceae Indeterminada 1 1033 RB/RJ
Eriocaulaceae Eriocaulaceae Indeterminada 2 448 RB
Família Espécie Testemunho * Vereda
Erythroxylaceae Erythroxylum betulaceae Mart. 352 RB

Erythroxylaceae Erythroxylum cf. affine A.St.Hil. 430 RB
Erythroxylaceae Erythroxylum sp. 1 584 RJ
Erythroxylaceae Erythroxylum sp. 2 539 RJ
Euphorbiaceae Amanoa sp. 329 RB
Euphorbiaceae Croton agoensis Baill. 921 RJ
Euphorbiaceae Croton cf. antisyphiliticus Mart. 410 RB
Euphorbiaceae Croton didrichsenii G.L Webster 577 RB/RJ
Euphorbiaceae Euphorbia potentilloides Boiss. 1558 RB/RJ
Euphorbiaceae Euphorbia sp. 1 521 RJ
Euphorbiaceae Manihot caerulescens (Pohl) Rog. 943 RB/RJ
Euphorbiaceae Manihot sp. 1 400 RB
Euphorbiaceae Sebastiania bidentata (Mart. & Zucc.) Pax 196 RB
Euphorbiaceae Sebastiania daphnoides Muell. Arg. 389 RB
Euphorbiaceae Euphorbiaceae Indeterminada 7 1067 RB/RJ
Fabaceae Aeschynomene sp 1. 542 RB/RJ
Fabaceae Aeschynomene sp. 2 919 RJ
Fabaceae Bauhinia acuruana Moric 1618 RB/RJ
Fabaceae Bauhinia sp. 1 1593 RB
Fabaceae Bauhinia sp.2 1505 RB
Fabaceae Bauhinia sp. 3 1505 RJ
Fabaceae Calliandra dysantha Benth. 1548 RB
Família Espécie Testemunho * Vereda
Fabaceae Camptosema sp. 1572 RB/RJ
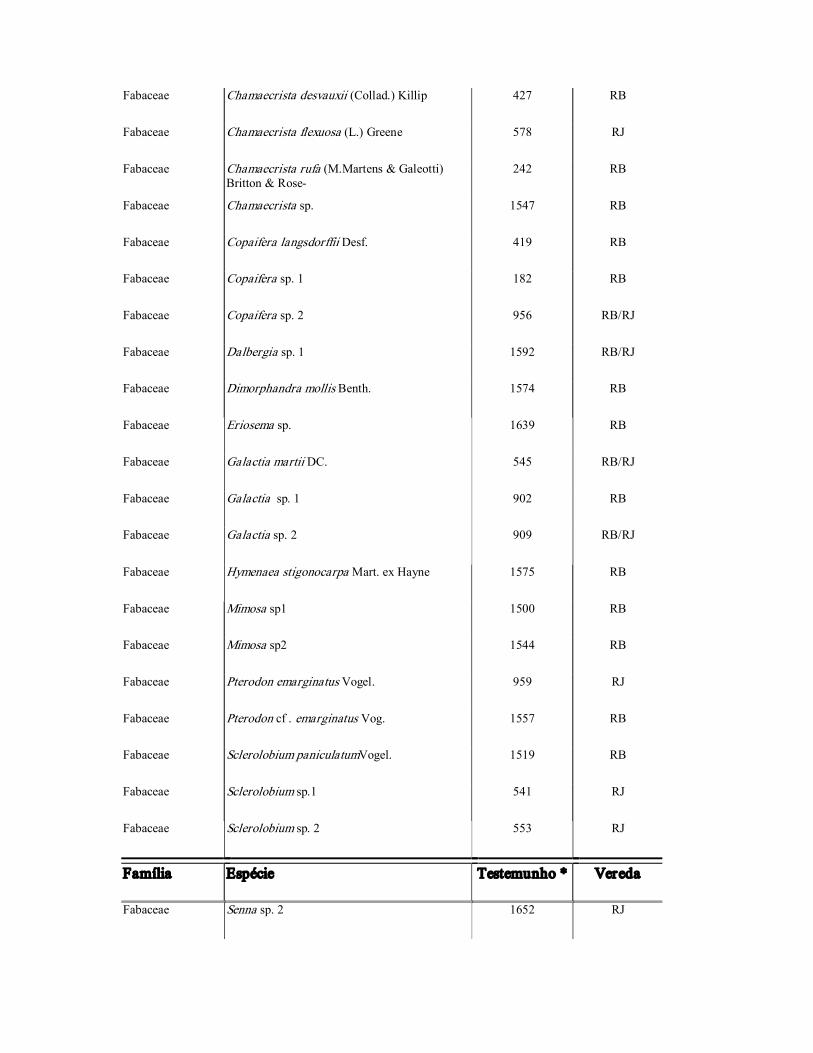
Fabaceae Chamaecrista desvauxii (Collad.) Killip 427 RB
Fabaceae Chamaecrista flexuosa (L.) Greene 578 RJ
Fabaceae Chamaecrista rufa (M.Martens & Galeotti) Britton & Rose
242 RB
Fabaceae Chamaecrista sp. 1547 RB
Fabaceae Copaifera langsdorffii Desf. 419 RB
Fabaceae Copaifera sp. 1 182 RB
Fabaceae Copaifera sp. 2 956 RB/RJ
Fabaceae Dalbergia sp. 1 1592 RB/RJ
Fabaceae Dimorphandra mollis Benth. 1574 RB
Fabaceae Eriosema sp. 1639 RB
Fabaceae Galactia martii DC. 545 RB/RJ
Fabaceae Galactia sp. 1 902 RB
Fabaceae Galactia sp. 2 909 RB/RJ
Fabaceae Hymenaea stigonocarpa Mart. ex Hayne 1575 RB
Fabaceae Mimosa sp1 1500 RB
Fabaceae Mimosa sp2 1544 RB
Fabaceae Pterodon emarginatus Vogel. 959 RJ
Fabaceae Pterodon cf . emarginatus Vog. 1557 RB
Fabaceae Sclerolobium paniculatumVogel. 1519 RB
Fabaceae Sclerolobium sp.1 541 RJ
Fabaceae Sclerolobium sp. 2 553 RJ
Família Espécie Testemunho * Vereda
Fabaceae Senna sp. 2 1652 RJ
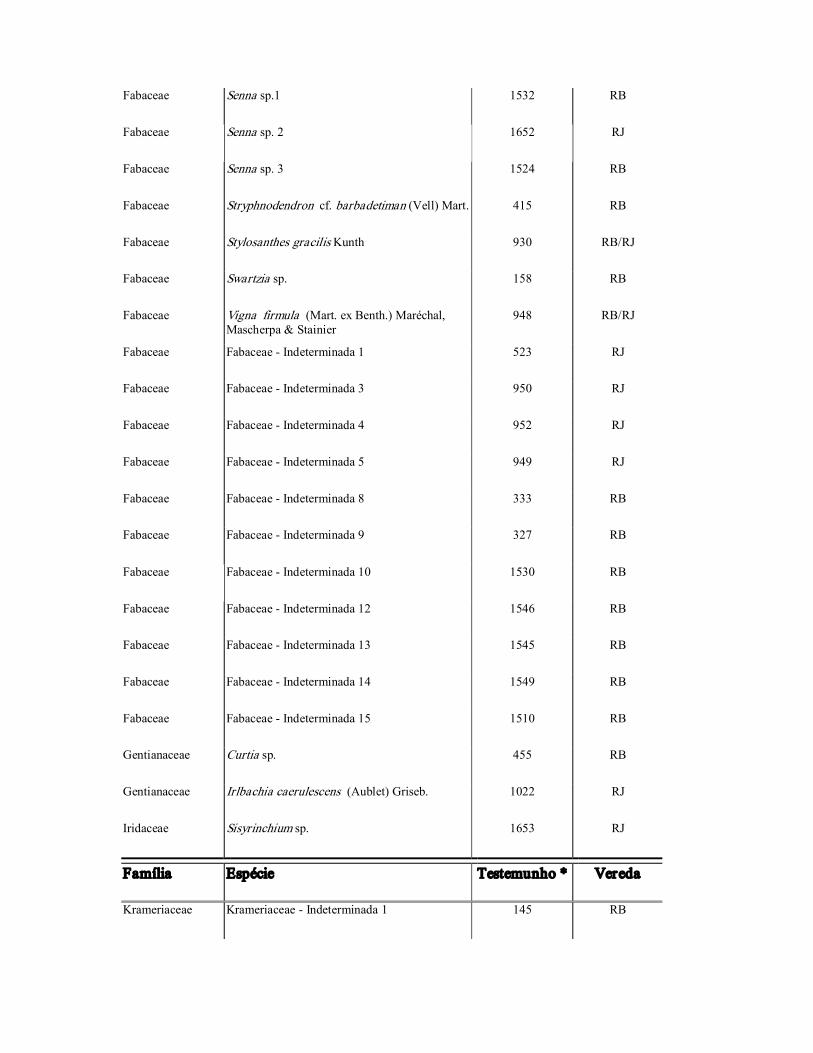
Fabaceae Senna sp.1 1532 RB
Fabaceae Senna sp. 2 1652 RJ
Fabaceae Senna sp. 3 1524 RB
Fabaceae Stryphnodendron cf. barbadetiman (Vell) Mart. 415 RB
Fabaceae Stylosanthes gracilis Kunth 930 RB/RJ
Fabaceae Swartzia sp. 158 RB
Fabaceae Vigna firmula (Mart. ex Benth.) Maréchal, Mascherpa & Stainier
948 RB/RJ
Fabaceae Fabaceae Indeterminada 1 523 RJ
Fabaceae Fabaceae Indeterminada 3 950 RJ
Fabaceae Fabaceae Indeterminada 4 952 RJ
Fabaceae Fabaceae Indeterminada 5 949 RJ
Fabaceae Fabaceae Indeterminada 8 333 RB
Fabaceae Fabaceae Indeterminada 9 327 RB
Fabaceae Fabaceae Indeterminada 10 1530 RB
Fabaceae Fabaceae Indeterminada 12 1546 RB
Fabaceae Fabaceae Indeterminada 13 1545 RB
Fabaceae Fabaceae Indeterminada 14 1549 RB
Fabaceae Fabaceae Indeterminada 15 1510 RB
Gentianaceae Curtia sp. 455 RB
Gentianaceae Irlbachia caerulescens (Aublet) Griseb. 1022 RJ
Iridaceae Sisyrinchium sp. 1653 RJ
Família Espécie Testemunho * Vereda
Krameriaceae Krameriaceae Indeterminada 1 145 RB

Lamiaceae Amasonia campestris (Abdul.) Moldenke 1658 RB
Lamiaceae Eriope sp. 666 RB
Lamiaceae Hiptis sp. 953 RJ
Lamiaceae Peltodon tomentosus Pohl 221 RB
Lamiaceae Vitex rufescens A.Juss. 958 RJ
Lamiaceae Lamiaceae Indeterminada 1 382 RB
Lauraceae Lauraceae Indeterminada 1 1614 RB
Lauraceae Lauraceae Indeterminada 2 259 RB
Lentibulariaceae Utricularia cf. amethystina A. St. Hil. 1061 RB/RJ
Lentibulariaceae Utricularia gibba L. 1031 RB/RJ
Lythraceae Cuphea antisyphilitica H.B.K. 1062 RB/RJ
Lythraceae Diplusodon sp. 398 RB
Lythraceae Lythraceae Indeterminada 1 1507 RB
Malpighiaceae Banisteriopsis cf. malifolia (Nees & Mart.) B. Gates
247 RB
Malpighiaceae Byrsonima basiloba A.Juss. 1656 RB
Malpighiaceae Byrsonima crassifolia (L.) Kunth 935 RJ
Malpighiaceae Byrsonima stannardii W. R. Anderson 562 RJ
Malpighiaceae Byrsonima umbellata Mart. 1065 RJ
Malpighiaceae Byrsonima verbascifolia (L.) Rich. 650 RB
Malpighiaceae Byrsonima sp. 1 1596 RB
Malpighiaceae Byrsonima sp. 2 960 RJ
Família Espécie Testemunho * Vereda
Malpighiaceae Camarea cf. affinis St. Hil. 154 RB

Malpighiaceae Peixotoa sp. 519 RB/RJ
Malpighiaceae Malpighiaceae Indeterminada 1 1623 RJ
Malpighiaceae Malpighiaceae Indeterminada 2 1623 RB
Malvaceae Helicteres velutina K.Schum. 443 RB
Malvaceae Pavonia cf. longitricha Fryxell 257 RB
Malvaceae Pavonia sp. 157 RB
Malvaceae Malvaceae Indeterminada 1 665 RB
Marantaceae Maranta sp. 660 RB
Melastomataceae Cambessedesia hilariana (Kunth) DC. 471 RB
Melastomataceae Desmocelis villosa (Aubl.) Naud. 462 RB
Melastomataceae Macairea radula (Bonpl.) DC. 488 RB
Melastomataceae Mouriri cf. pusa Gardner 961 RJ
Melastomataceae Mouriri sp. 367 RB
Melastomataceae Microlicia sp. 1064 RJ
Melastomataceae Melastomataceae Indeterminada 7 490 RB
Menispermaceae Cissampelos ovalifolia DC. 544 RJ
Moraceae Brosmium cf. gaudichaudii Trécul. 191 RB
Moraceae Moraceae Indeterminada 4 1581 RB/RJ
Moraceae Moraceae Indeterminada 5 1616 RB
Myrtaceae Eugenia disynterica DC. 1559 RB
Myrtaceae Eugenia punicifolia (Kunth.) DC 176 RB
Família Espécie Testemunho * Vereda
Myrtaceae Eugenia sp. 173 RB
Myrtaceae Myrcia laricina (O. Berg) Burret ex Luetzellb. 442 RB
Myrtaceae Myrcia guianensis (Aubl.) DC. 576 RJ
Myrtaceae Myrcia cf. mollis (Kunth) DC. 960 RJ
Myrtaceae Myrcia sp. 1 1512 RB
Myrtaceae Myrcia sp. 10 1613 RB
Myrtaceae Myrtaceae Indeterminada 1 152 RB
Myrtaceae Myrtaceae Indeterminada 2 133 RB
Myrtaceae Myrtaceae Indeterminada 3 331 RB
Myrtaceae Myrtaceae Indeterminada 5 354 RB
Myrtaceae Myrtaceae Indeterminada 8 917 RJ
Myrtaceae Myrtaceae Indeterminada 9 569 RJ
Myrtaceae Myrtaceae Indeterminada 11 1582 RB
Myrtaceae Myrtaceae Indeterminada 12 1512 RB
Nyctaginaceae Guapira cf. Noxia (Netto) Lundell 579 RJ
Ochnaceae Sauvagesia cf. sprengelli A. St. Hil. 270 RB
Ochnaceae Sauvagesia linearifolia A. St. Hil. 1023 RB/RJ
Ochnaceae Ouratea cf. spectabilis (Mart.) Engl. 409 RB
Olacaceae Ximenia sp. 1651 RB
Opiliaceae Agonandra brasiliensis Miers 1633 RB
Orchidaceae Cyrtopodium sp. 561 RB/RJ
Orchidaceae Epistephium sclerophylum Lind. 1671 RB
Família Espécie Testemunho * Vereda
Orchidaceae Habenaria cf. secundiflora Barb. Rodr. 1009 RB/RJ
Orchidaceae Habenaria sp. 376 RB
Orchidaceae Habenaria josephensis Barb. Rodr. 1672 RB
Orobanchaceae Buchnera sp. 1002 RB/RJ
Phyllantaceae Phyllanthus orbiculatus LC Rich. 347 RB
Poaceae Aristida riparia Trin. 934 RJ
Poaceae Aristida sp. 924 RJ
Poaceae Axonopus grandifoliumRev. 558 RB/RJ
Poaceae Echinolaena inflexa (Poir.) Chase 547 RJ
Poaceae Luziola bahiensis (Steud.) Hitchc. 485 RB
Poaceae Luziola fragilis Swallen 239 RB
Poaceae Luziola sp. 203 RB
Poaceae Panicum rupestre Trin 489 RB
Poaceae Panicum sp. 1 1624 RJ
Poaceae Panicum sp. 2 1801 RB/RJ
Poaceae Paspalum foveolatum Sdend. 416 RB/RJ
Poaceae Paspalum sp. 2 1551 RB/RJ
Poaceae Trachypogon spicatus (L.f/Kuntze) 552 RB/RJ
Poaceae Poaceae – Indeterminada 3 916 RJ
Poaceae Poaceae – Indeterminada 4 928 RJ
Poaceae Poaceae – Indeterminada 5 933 RJ
Poaceae Poaceae – Indeterminada 6 1020 RJ
Família Espécie Testemunho * Vereda
Poaceae Poaceae – Indeterminada 7 1070 RJ
Poaceae Poaceae – Indeterminada 8 910 RJ
Poaceae Poaceae – Indeterminada 9 498 RB
Poaceae Poaceae – Indeterminada 12 314 RB
Poaceae Poaceae – Indeterminada 15 452 RB
Poaceae Poaceae – Indeterminada 19 1522 RB
Poaceae Poaceae – Indeterminada 20 1531 RB
Poaceae Poaceae – Indeterminada 21 1535 RB
Poaceae Poaceae – Indeterminada 23 1542 RB
Poaceae Poaceae – Indeterminada 24 1553 RB
Poaceae Poaceae – Indeterminada 25 1550 RB
Poaceae Poaceae – Indeterminada 26 1800 RB
Poaceae Poaceae – Indeterminada 28 1802 RB/RJ
Poaceae Poaceae – Indeterminada 29 1803 RB
Polygalaceae Polygala adenosphora DC. 1005 RB/RJ
Polygalaceae Polygala cf. equisetoides A.St.Hil & Moq. 568 RJ
Polygalaceae Polygala rhodoptera Mart. ex A.W. Benn. 1014 RB/RJ
Polygalaceae Polygala sp. 1012 RJ
Polygalaceae Polygalaceae Indeterminada 1 1617 RB/RJ
Rapateaceae Cephalostemon ridelianus Koen. 1048 RB/RJ
Rubiaceae Alibertia edulis (Rich.) A. Rich ex DC. 571 RJ
Rubiaceae Borreria cf. crispata (K. Schum) E.L. Cobral et Bacigalupo
1528 RB
Família Espécie Testemunho * Vereda
Rubiaceae Borreria sp. 2 1635 RJ
Rubiaceae Chomelia anisomeris Mull. Arg. 178 RB
Rubiaceae Galianthe sp. 1 1655 RJ
Rubiaceae Declieuxia fruticosa (Wild. ex Ruiz & Pav.) Kuntze
906 RB/RJ
Rubiaceae Diodia sp. 1 1537 RB/RJ
Rubiaceae Emmeorrhizia umbellata (Spreng.) K. Shum. 583 RJ
Rubiaceae Ladenbergia sp. 1066 RJ
Rubiaceae Palicouria rígida Kunth 954 RB/RJ
Rubiaceae Spermacoce capitata Moc. & Sesse 147 RB
Rubiaceae Spermacoce sp. 580 RB/RJ
Rubiaceae Staelia sp. 431 RB
Rubiaceae Tocoyena formosa (Cham. & Schltdl.) K.Schum.
408 RB
Rubiaceae Rubiaceae Indeterminada 3 936 RJ
Rubiaceae Rubiaceae Indeterminada 6 190 RB
Sabiaceae Casearia cf. arborea (Rich.) Urb. 142 RB
Sapindaceae Serjania sp. 904 RJ
Simaroubaceae Simarauba sp. 1 961 RJ
Simaroubaceae Simarauba sp. 2 910 RJ
Smilacaceae Smilax guianensis Vitman 930 RB/RJ
Solanaceae Schwenckia sp. 1637 RJ
Urticaceae Cecropia pachystachya Trec. 1041 RJ
Velloziaceae Vellozia froesii L.B. Sm. 405 RB
Família Espécie Testemunho * Vereda
Verbenaceae Verbenaceae Indeterminada 4 1504 RB

Verbenaceae Lippia sp. 1 546 RJ
Verbenaceae Lippia sp. 2 581 RJ
Vochysiaceae Qualea grandiflora Mart. 1605 RB
Vochysiaceae Qualea parviflora Mart. 340 RB/RJ
Vochysiaceae Vochysia gardneri Warm. 441 RB
Vochysiaceae Vochysiaceae Indeterminada 4 941 RJ
Xyridaceae Abolboda sp. 1037 RJ
Xyridaceae Xyris blanchetiana Malme 1006 RJ
Xyridaceae Xyris jupicai L.C. Rich. 1051 RB/RJ
Xyridaceae Xyris cf. goyazensis Malme 1015 RB/RJ
Xyridaceae Xyris savanensis Miquel 1025 RB/RJ
Família Indeterminada 1
Família Indeterminada 1 519 RJ
Família Indeterminada 2
Família Indeterminada 2 536 RJ
Família Indeterminada 4
Família Indeterminada 4 910 RJ
Família Indeterminada 5
Família Indeterminada 5 905 RJ
Família Indeterminada 6
Família Indeterminada 6 911 RJ
Família Indeterminada 10
Família Indeterminada 10 946 RJ
Família Indeterminada 11
Família Indeterminada 11 949 RJ
Família Indeterminada 17
Família Indeterminada 17 232 RB
Família Indeterminada 19
Família Indeterminada 19 220 RB
Família Indeterminada 25
Família Indeterminada 25 1511 RB/RJ
Tabela 4: Parâmetros de diversidade obtido a partir das parcelas de 10 x 10 m (riqueza) e subparcelas de 2 x 2m (H’ e número de indivíduos) na vegetação das veredas Rio de Janeiro (RJ) e Rio Branco (RB), incluindo o cerrado circundante e as zonas da borda (B), meio (M) e fundo (F). Oeste da Bahia, Brasil.
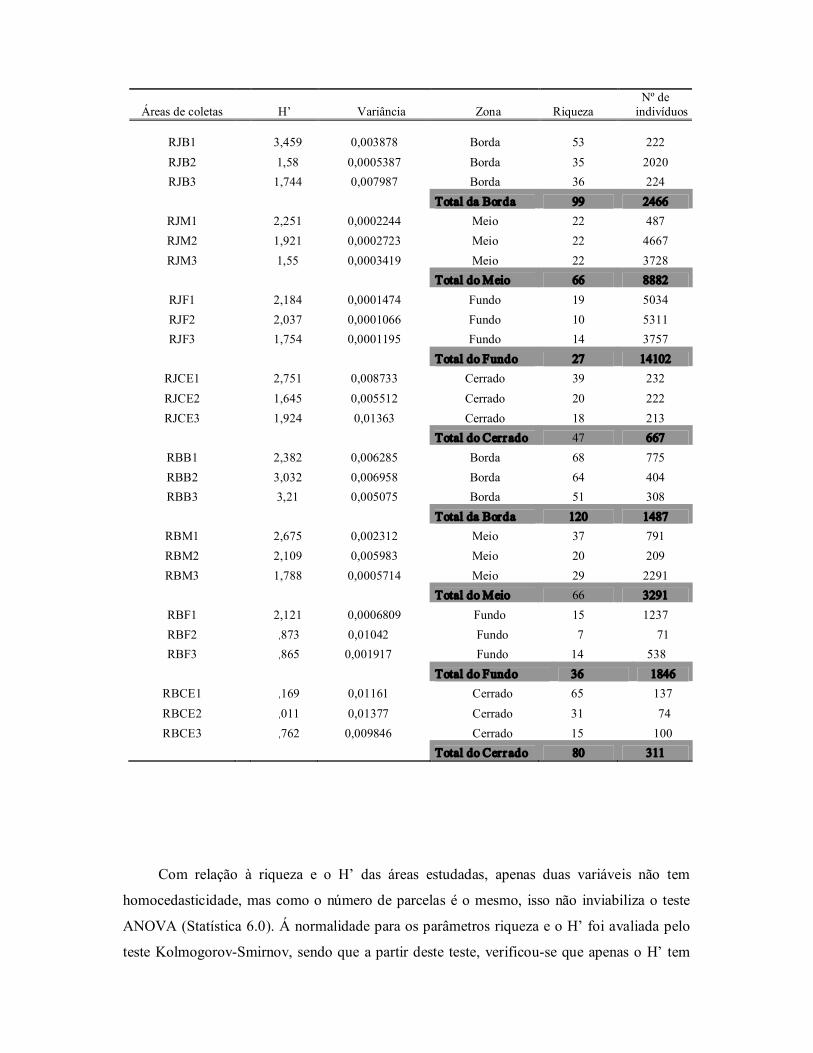
Áreas de coletas H’ Variância Zona Riqueza Nº de indivíduos
RJB1 3,459 0,003878 Borda 53 222 RJB2 1,58 0,0005387 Borda 35 2020 RJB3 1,744 0,007987 Borda 36 224
Total da Borda 99 2466 RJM1 2,251 0,0002244 Meio 22 487 RJM2 1,921 0,0002723 Meio 22 4667 RJM3 1,55 0,0003419 Meio 22 3728
Total do Meio 66 8882 RJF1 2,184 0,0001474 Fundo 19 5034 RJF2 2,037 0,0001066 Fundo 10 5311 RJF3 1,754 0,0001195 Fundo 14 3757
Total do Fundo 27 14102 RJCE1 2,751 0,008733 Cerrado 39 232 RJCE2 1,645 0,005512 Cerrado 20 222 RJCE3 1,924 0,01363 Cerrado 18 213
Total do Cerr ado 47 667 RBB1 2,382 0,006285 Borda 68 775 RBB2 3,032 0,006958 Borda 64 404 RBB3 3,21 0,005075 Borda 51 308
Total da Borda 120 1487 RBM1 2,675 0,002312 Meio 37 791 RBM2 2,109 0,005983 Meio 20 209 RBM3 1,788 0,0005714 Meio 29 2291
Total do Meio 66 3291 RBF1 2,121 0,0006809 Fundo 15 1237 RBF2 1,873 0,01042 Fundo 7 71 RBF3 1,865 0,001917 Fundo 14 538
Total do Fundo 36 1846 RBCE1 3,169 0,01161 Cerrado 65 137 RBCE2 3,011 0,01377 Cerrado 31 74 RBCE3 2,762 0,009846 Cerrado 15 100
Total do Cerr ado 80 311
Com relação à riqueza e o H’ das áreas estudadas, apenas duas variáveis não tem
homocedasticidade, mas como o número de parcelas é o mesmo, isso não inviabiliza o teste
ANOVA (Statística 6.0). Á normalidade para os parâmetros riqueza e o H’ foi avaliada pelo
teste KolmogorovSmirnov, sendo que a partir deste teste, verificouse que apenas o H’ tem
distribuição normal, não apresentando normalidade para a riqueza. Utilizandose deste mesmo
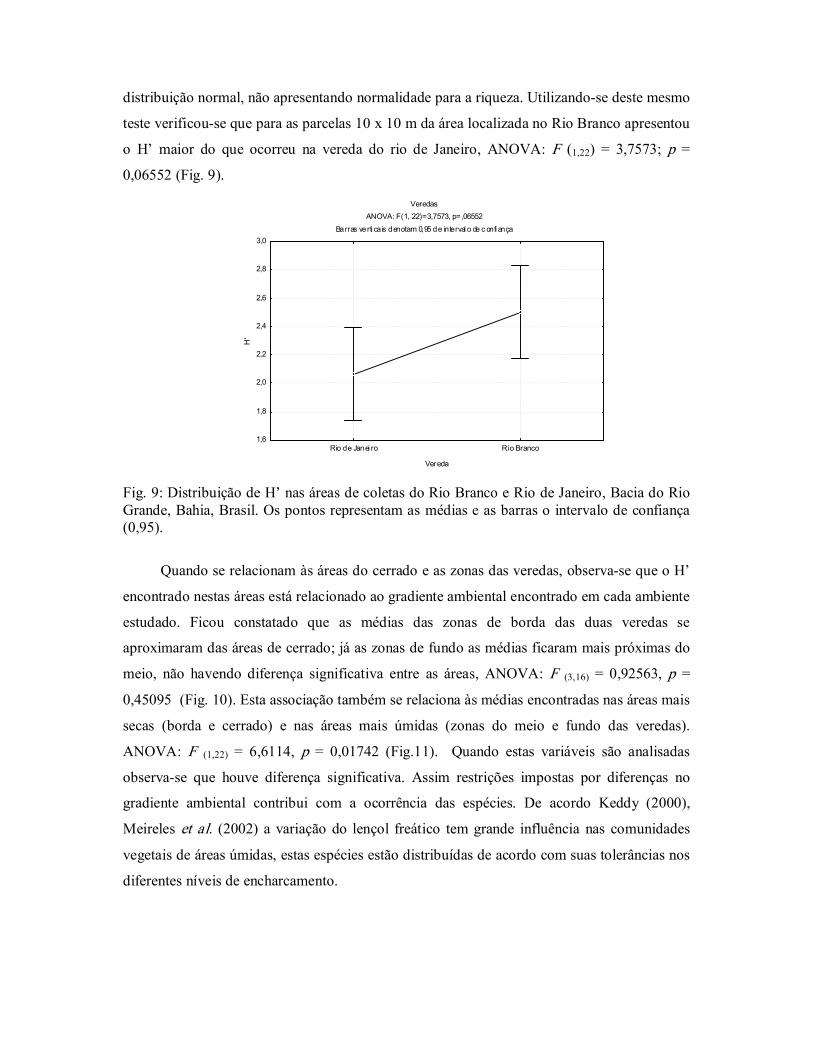
teste verificouse que para as parcelas 10 x 10 m da área localizada no Rio Branco apresentou
o H’ maior do que ocorreu na vereda do rio de Janeiro, ANOVA: F (1,22) = 3,7573; p = 0,06552 (Fig. 9).
Veredas ANOVA: F(1, 22)=3,7573, p=,06552
Barras verti cais denotam 0,95 de interval o de c onfi ança
Rio de Janei ro Rio Branco
Vereda
1,6
1,8
2,0
2,2
2,4
2,6
2,8
3,0 H'
Fig. 9: Distribuição de H’ nas áreas de coletas do Rio Branco e Rio de Janeiro, Bacia do Rio Grande, Bahia, Brasil. Os pontos representam as médias e as barras o intervalo de confiança (0,95).
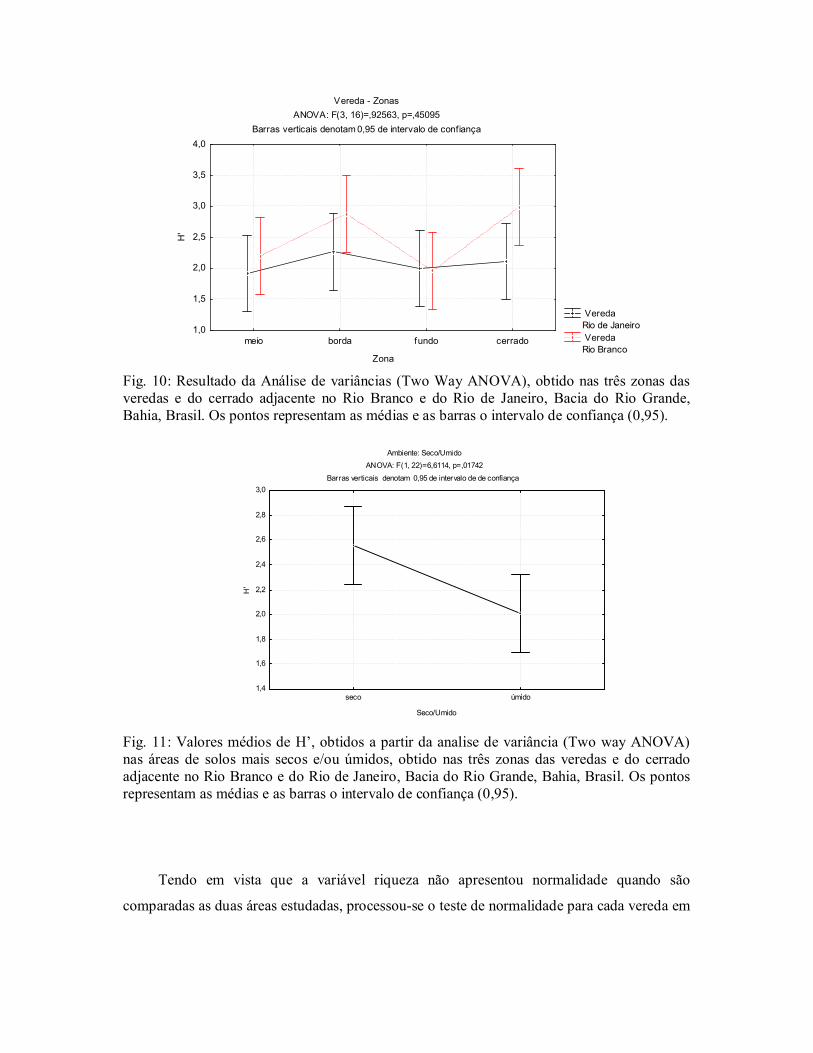
Quando se relacionam às áreas do cerrado e as zonas das veredas, observase que o H’
encontrado nestas áreas está relacionado ao gradiente ambiental encontrado em cada ambiente
estudado. Ficou constatado que as médias das zonas de borda das duas veredas se
aproximaram das áreas de cerrado; já as zonas de fundo as médias ficaram mais próximas do
meio, não havendo diferença significativa entre as áreas, ANOVA: F (3,16) = 0,92563, p =
0,45095 (Fig. 10). Esta associação também se relaciona às médias encontradas nas áreas mais
secas (borda e cerrado) e nas áreas mais úmidas (zonas do meio e fundo das veredas).
ANOVA: F (1,22) = 6,6114, p = 0,01742 (Fig.11). Quando estas variáveis são analisadas observase que houve diferença significativa. Assim restrições impostas por diferenças no
gradiente ambiental contribui com a ocorrência das espécies. De acordo Keddy (2000),
Meireles et al. (2002) a variação do lençol freático tem grande influência nas comunidades
vegetais de áreas úmidas, estas espécies estão distribuídas de acordo com suas tolerâncias nos
diferentes níveis de encharcamento.
Vereda Zonas ANOVA: F(3, 16)=,92563, p=,45095
Barras verticais denotam 0,95 de intervalo de confiança
Vereda Rio de Janeiro Vereda Rio Branco
meio borda fundo cerrado
Zona
1,0
1,5
2,0
2,5
3,0
3,5
4,0
H'
Fig. 10: Resultado da Análise de variâncias (Two Way ANOVA), obtido nas três zonas das veredas e do cerrado adjacente no Rio Branco e do Rio de Janeiro, Bacia do Rio Grande, Bahia, Brasil. Os pontos representam as médias e as barras o intervalo de confiança (0,95).
Ambiente: Seco/Umido ANOVA: F(1, 22)=6,6114, p=,01742
Barras verticais denotam 0,95 de intervalo de de confiança
seco úmido
Seco/Umido
1,4
1,6
1,8
2,0
2,2
2,4
2,6
2,8
3,0
H'
Fig. 11: Valores médios de H’, obtidos a partir da analise de variância (Two way ANOVA) nas áreas de solos mais secos e/ou úmidos, obtido nas três zonas das veredas e do cerrado adjacente no Rio Branco e do Rio de Janeiro, Bacia do Rio Grande, Bahia, Brasil. Os pontos representam as médias e as barras o intervalo de confiança (0,95).
Tendo em vista que a variável riqueza não apresentou normalidade quando são
comparadas as duas áreas estudadas, processouse o teste de normalidade para cada vereda em
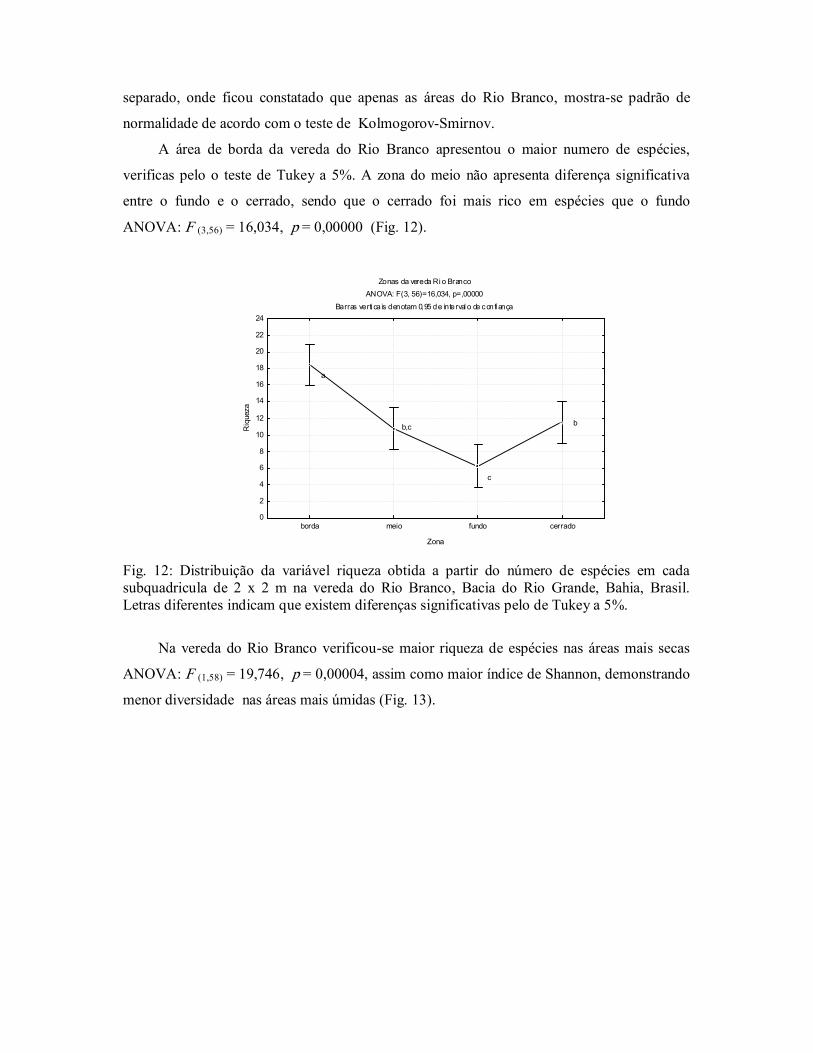
separado, onde ficou constatado que apenas as áreas do Rio Branco, mostrase padrão de
normalidade de acordo com o teste de KolmogorovSmirnov.
A área de borda da vereda do Rio Branco apresentou o maior numero de espécies,
verificas pelo o teste de Tukey a 5%. A zona do meio não apresenta diferença significativa
entre o fundo e o cerrado, sendo que o cerrado foi mais rico em espécies que o fundo
ANOVA: F (3,56) = 16,034, p = 0,00000 (Fig. 12).
Zonas da vereda Ri o Branco ANOVA: F(3, 56)=16,034, p=,00000
Barras verti ca is denotam 0,95 de interval o de confi ança
borda meio fundo cerrado
Zona
0
2
4
6
8
10
12
14
16
18
20
22
24
Riqueza
a
c
b,c b
Fig. 12: Distribuição da variável riqueza obtida a partir do número de espécies em cada subquadricula de 2 x 2 m na vereda do Rio Branco, Bacia do Rio Grande, Bahia, Brasil. Letras diferentes indicam que existem diferenças significativas pelo de Tukey a 5%.
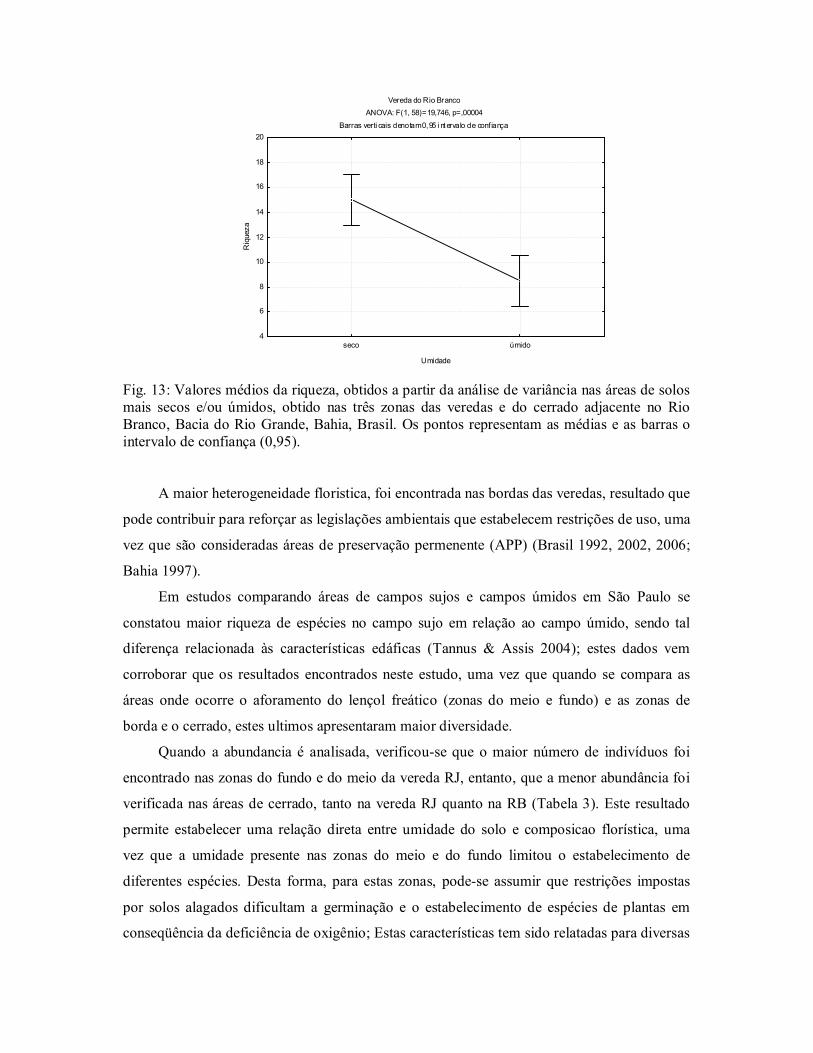
Na vereda do Rio Branco verificouse maior riqueza de espécies nas áreas mais secas
ANOVA: F (1,58) = 19,746, p = 0,00004, assim como maior índice de Shannon, demonstrando menor diversidade nas áreas mais úmidas (Fig. 13).
Vereda do Rio Branco ANOVA: F(1, 58)=19,746, p=,00004
Barras verti cais denotam 0,95 i nt ervalo de conf iança
seco úmido
Umidade
4
6
8
10
12
14
16
18
20
Riqueza
Fig. 13: Valores médios da riqueza, obtidos a partir da análise de variância nas áreas de solos mais secos e/ou úmidos, obtido nas três zonas das veredas e do cerrado adjacente no Rio Branco, Bacia do Rio Grande, Bahia, Brasil. Os pontos representam as médias e as barras o intervalo de confiança (0,95).
A maior heterogeneidade floristica, foi encontrada nas bordas das veredas, resultado que
pode contribuir para reforçar as legislações ambientais que estabelecem restrições de uso, uma
vez que são consideradas áreas de preservação permenente (APP) (Brasil 1992, 2002, 2006;
Bahia 1997).
Em estudos comparando áreas de campos sujos e campos úmidos em São Paulo se
constatou maior riqueza de espécies no campo sujo em relação ao campo úmido, sendo tal
diferença relacionada às características edáficas (Tannus & Assis 2004); estes dados vem
corroborar que os resultados encontrados neste estudo, uma vez que quando se compara as
áreas onde ocorre o aforamento do lençol freático (zonas do meio e fundo) e as zonas de
borda e o cerrado, estes ultimos apresentaram maior diversidade.
Quando a abundancia é analisada, verificouse que o maior número de indivíduos foi
encontrado nas zonas do fundo e do meio da vereda RJ, entanto, que a menor abundância foi
verificada nas áreas de cerrado, tanto na vereda RJ quanto na RB (Tabela 3). Este resultado
permite estabelecer uma relação direta entre umidade do solo e composicao florística, uma
vez que a umidade presente nas zonas do meio e do fundo limitou o estabelecimento de
diferentes espécies. Desta forma, para estas zonas, podese assumir que restrições impostas
por solos alagados dificultam a germinação e o estabelecimento de espécies de plantas em
conseqüência da deficiência de oxigênio; Estas características tem sido relatadas para diversas

formações vegetais que usualmente apresentam baixa riqueza e diversidade florística
(Ivanauskas et al. 1997; Penning & Moore 2001; Araújo et al. 2002; Cattanio et al. 2002). Ainda, de acordo com Araújo et al. (2000), densas touceiras de espécies filiformes podem dificultar o estabelecimento de outras espécies nas zonas mais úmidas de veredas, fato que
vem contribuir com a abundância de poucas espécies. Este estudo corrobora com esta
afirmação, uma vez que as famílias de maior abundancia encontradas nestas zonas foram:
Poaceae, Cyperaceae, Eriocaulaceae e Xyridaceae.
Kent & Coker (1994), Pandey & Singh (1991) relataram que o gradiente ambiental é
responsável pela variação da abundância das espécies e o estabelecimento de comunidades
está diretamente relacionado a fatores como umidade e altitude. Estes mesmos autores
estabelecem que quando algumas espécies crescem juntas num ambiente em particular,
geralmente estas apresentam as mesmas necessidades de luz, temperatura, umidade e talvez
compartilhem habilidades para tolerar estresse de atividades antrópicas e ambientais.
Os afloramentos do lençol freático nas zonas de fundo (ano todo) e nas zonas do meio
(período do ano) nas duas veredas estudadas não acontecem de forma pontual e sim de forma
difusa. A forma pontual é quando o fluxo d’água ocorre em um único local do terreno,
enquanto a difusa apresenta vários olhos d’água (Pinto et al. 2005). Na vereda RB, a zona de fundo permanece alagada com pelo menos 1,5 m de água durante todo o ano, enquanto que na
vereda RJ e no mesmo período essa mesma zona não ultrapassa 30 cm de água. O nível de
água presente durante o ano todo na vereda RB devese possivelmente à construção da BA
825 que atravessou essa vereda impedindo o fluxo natural da água, o que pode ter influência
na tendência de maior riqueza constatada nesta vereda, já que as perturbações podem
aumentar a heterogeneidade ambiental (Ricklefs 1996; Guimarães et al. 2002). Outro fator importante que também deve ser considerado é a característica do regime de água que afeta a
distribuição das espécies (OliveiraFilho et al. 1994; Pinto et al. 2005).
4.2. Similaridade
A análise de similaridade florística (Jaccard e Sørensen) entre as áreas de cerrado e
veredas revelou a grande importância do gradiente ambiental na estruturação da área estudada
(Fig. 14 e 15).

A ordenação encontrada nas zonas das veredas estudadas (Fig. 14 e 15) mostrou maior
similaridade floristica entre o meio e fundo, do que com as bordas. Este fato já foi relatado
para as veredas do Triangulo Mineiro (Ramos 2000; Araujo et al. 2002; Guimarães et al. 2002; Oliveira 2005). Para este estudo foram agregadas as áreas de cerrado, desta forma
verificouse que as zonas de borda se aproximaram mais das áreas de cerrado.
As parcelas nas zonas de bordas e no cerrado se mostram bem distintas das parcelas nas
zonas do meio e do fundo, com apenas 4% de similaridade, sendo que grupos relacionados à
zona de cada vereda são formados a partir de 10%, de acordo com a análise de Jaccard.
Quando as duas veredas são analisadas em conjunto, verificouse que as áreas de bordas das
duas veredas analisadas se agrupam com 12% de similaridade, sendo as áreas de borda da
vereda do RB, apresentou maior similaridade com 34% (Fig. 16 e 17); as áreas de cerrado
agrupamse com 24% na vereda do RB e 34% na vereda RJ. As zonas do meio da vereda RB
se agrupam com 34% de similaridade, enquanto que nas áreas de fundo apresentam
similaridade de 40%. Na vereda RJ há um grupo incluindo todas as parcelas do meio e do
fundo com 24% de similaridade, sendo as zonas do meio mais similares com 65%, enquanto
que as do fundo apresentou 34%. As parcelas das faixas 1 e 2 da zona do meio no RJ são as
mais similares (50%) (Fig. 18 e 19).
A índice de Sørensen apresentou o mesmo padrão de distribuição de Jaccard, no
entanto, este índice apresentou maior similaridade, uma vez que atribui maior peso as
espécies comuns, ou seja, as espécies que apresentam ocorrência em mais de uma área.
Assim, o dendograma mostra que as parcelas das zonas de borda e no cerrado quando são
comparadas com as zonas do meio e do fundo, apresentaram 8% de similaridade; o primeiro
grupo foi formado a partir de 13% os quais correspondem às áreas de borda e cerrado; o
segundo grupo formado a partir de 24% de similaridade incluem as zonas de meio e fundo das
duas veredas. As zonas de meio e fundo da vereda RB apresentaram 24% enquanto as
mesmas áreas, na vereda RJ apresentou similaridade de 38%. (Fig. 16 e 17). Os agrupamentos
formados entre as bordas da vereda RB e as bordas RJ apresentaram similaridade entre 30%
para as duas veredas. Já as áreas de cerrado se agruparam a partir de 45%. Quando são
consideradas apenas as zonas de meio observase que para a RJ, a similaridade apresentou
60%, com confiança de 99%, sendo as faixas 1 e 2 mais similares, com 75% de similaridade,
quando comparadas as demais áreas. As zonas do meio da vereda RB apresentou 40% de
similaridade, já as zonas do fundo, se agrupam em 50% de similaridade para as duas veredas
(Fig. 18 e 19).
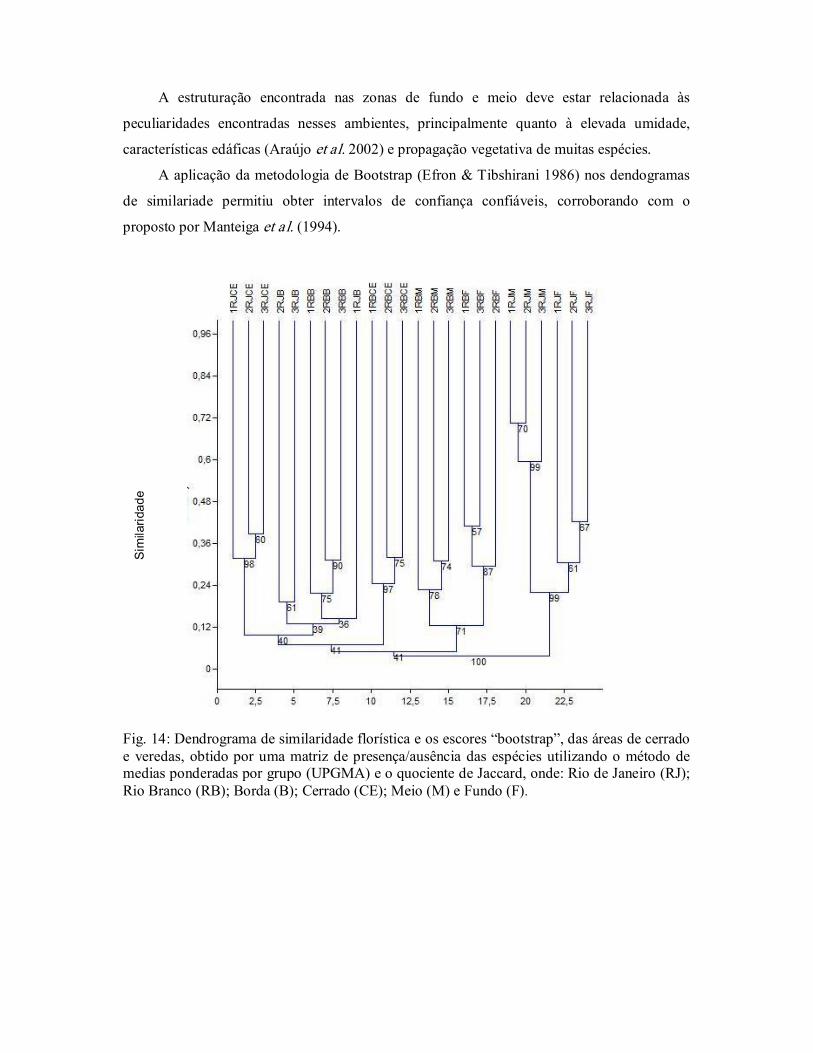
A estruturação encontrada nas zonas de fundo e meio deve estar relacionada às
peculiaridades encontradas nesses ambientes, principalmente quanto à elevada umidade,
características edáficas (Araújo et al. 2002) e propagação vegetativa de muitas espécies. A aplicação da metodologia de Bootstrap (Efron & Tibshirani 1986) nos dendogramas
de similariade permitiu obter intervalos de confiança confiáveis, corroborando com o
proposto por Manteiga et al. (1994).
Fig. 14: Dendrograma de similaridade florística e os escores “bootstrap”, das áreas de cerrado e veredas, obtido por uma matriz de presença/ausência das espécies utilizando o método de medias ponderadas por grupo (UPGMA) e o quociente de Jaccard, onde: Rio de Janeiro (RJ); Rio Branco (RB); Borda (B); Cerrado (CE); Meio (M) e Fundo (F).
Similaridade
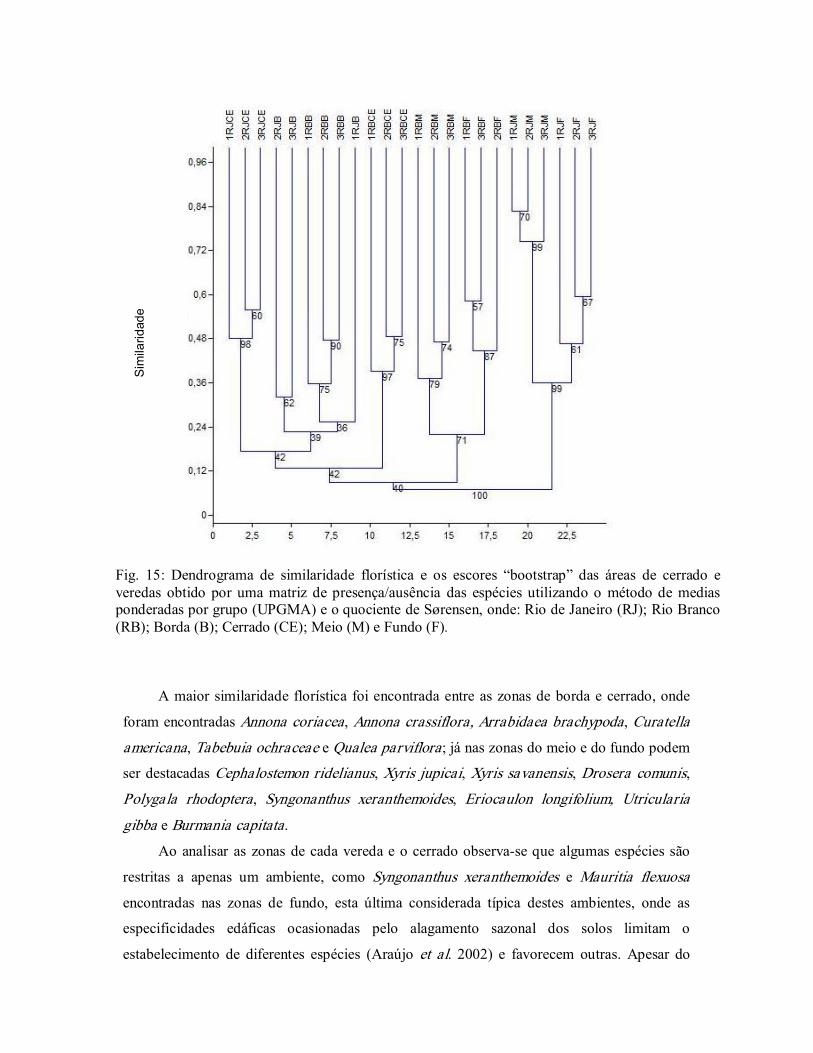
Fig. 15: Dendrograma de similaridade florística e os escores “bootstrap” das áreas de cerrado e veredas obtido por uma matriz de presença/ausência das espécies utilizando o método de medias ponderadas por grupo (UPGMA) e o quociente de Sørensen, onde: Rio de Janeiro (RJ); Rio Branco (RB); Borda (B); Cerrado (CE); Meio (M) e Fundo (F).
A maior similaridade florística foi encontrada entre as zonas de borda e cerrado, onde
foram encontradas Annona coriacea, Annona crassiflora, Arrabidaea brachypoda, Curatella americana, Tabebuia ochraceae e Qualea parviflora; já nas zonas do meio e do fundo podem ser destacadas Cephalostemon ridelianus, Xyris jupicai, Xyris savanensis, Drosera comunis, Polygala rhodoptera, Syngonanthus xeranthemoides, Eriocaulon longifolium, Utricularia
gibba e Burmania capitata. Ao analisar as zonas de cada vereda e o cerrado observase que algumas espécies são
restritas a apenas um ambiente, como Syngonanthus xeranthemoides e Mauritia flexuosa encontradas nas zonas de fundo, esta última considerada típica destes ambientes, onde as
especificidades edáficas ocasionadas pelo alagamento sazonal dos solos limitam o
estabelecimento de diferentes espécies (Araújo et al. 2002) e favorecem outras. Apesar do
Similaridade

ambiente peculiar destes ambientes, Lorenzi et al. (2004) classificou a M. flexuosa como a palmeira mais abundante do país. A espécie Sauvagesia linearifolia foi encontrada apenas nas zonas do meio, apesar de ter sido coletada em outras áreas de cerrado onde as condições dos
solos são semelhantes, como nas florestas ribeirinhas.
Algumas espécies coletadas nas veredas do Oeste da Bahia apresentam registros para as
veredas do Triangulo Mineiro e áreas de campos úmidos, como Xyris jupicai, Xyris savanensis e Xyris blanchetiana (Araújo et al. 2002; Oliveira 2005).
Estudos realizados em áreas de veredas (borda, meio e fundo) de Minas Gerais também
relataram Xyris savanensisis e Xyris jupicai; com Declieuxia fruticosa e Palicuria rigida nas zonas de borda (Araújo et al. 2002).
De acordo com a análise de agrupamento da vereda RB, as zonas de fundo apresentam
maior similaridade florística (Fig. 16 e 17), onde acontece o alagamento total do terreno.
Entre as espécies que são exclusivas destes ambientes, merecem destaques Sagittaria rhombifolia e Sagittaria guianensis, coletadas apenas nestes ambientes. Sagittaria rhombifolia também ocorre em zona de fundo de veredas em Minas Gerais (Araújo et al. 2002; Oliveira 2005), assim como em igarapés na Amazônia (Tolleto & Bosh 2008). Foram
encontradas ainda nesta zona da vereda RB Macairea radula e Panicum rupestre.
De acordo com Mendonça et al. (1993, 2001), Meireles et al. (2002) e Silva (2005), a variação do lençol freático tem grande influência nas comunidades vegetais de áreas úmidas,
com espécies distribuídas de acordo com suas tolerâncias aos diferentes níveis de
encharcamentos. Para Costa (2007) os padrões de zonação das espécies tem relação direta
com o nível do lençol freático, que por sua vez esta associado a fatores como topografia,
precipitação e nível de antropização. Keddy (2000), ressalta que os padrões de zonação em
áreas úmidas são resultados de diferentes níveis de altura da lamina d’ água, duração de
submersão, grau de competição e das condições edáficas.
Guimarães et al. (2002) e Ramos (2004), ressaltam ainda, que a vegetação encontrada em áreas de veredas (borda, meio e fundo), estão bem ajustadas ao percentual de água no solo,
sugerindo a interferência da profundidade do lençol frático na distribuição da maioria da
vegetação, podendo apresentar espécies de ocorrência restrita, e espécies generalistas, com
ampla distribuição nos extremos do gradiente de umidade do solo.
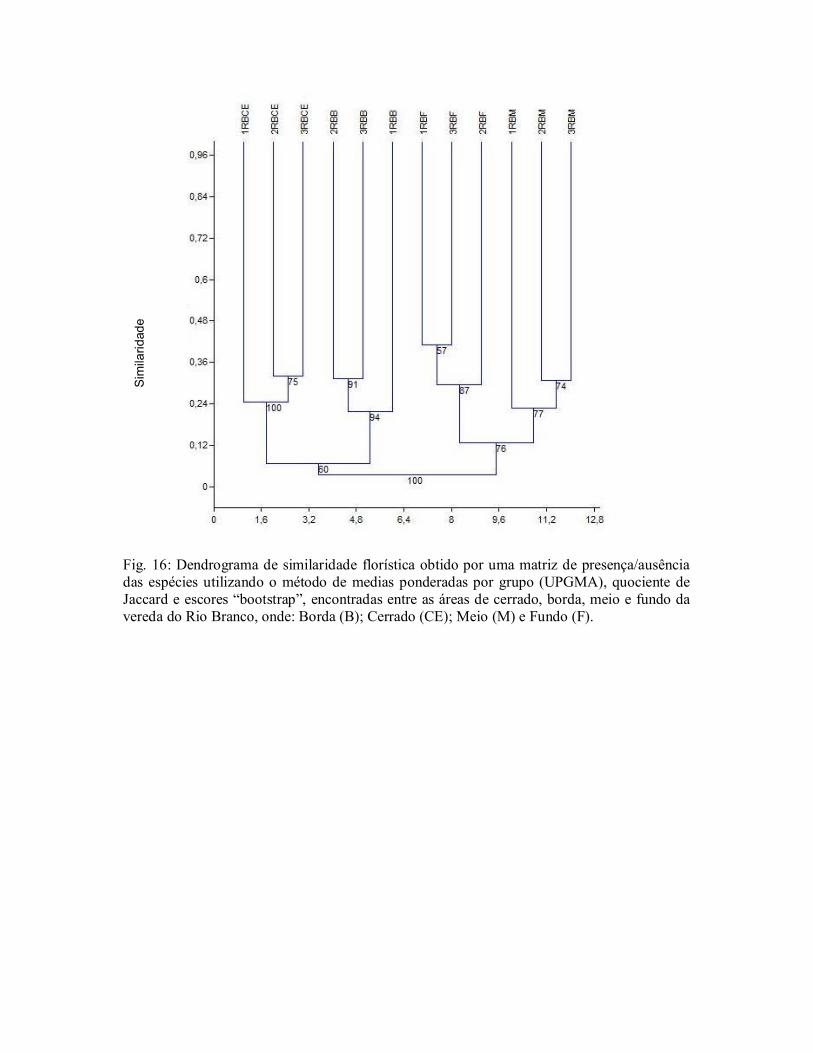
Fig. 16: Dendrograma de similaridade florística obtido por uma matriz de presença/ausência das espécies utilizando o método de medias ponderadas por grupo (UPGMA), quociente de Jaccard e escores “bootstrap”, encontradas entre as áreas de cerrado, borda, meio e fundo da vereda do Rio Branco, onde: Borda (B); Cerrado (CE); Meio (M) e Fundo (F).
Similaridade
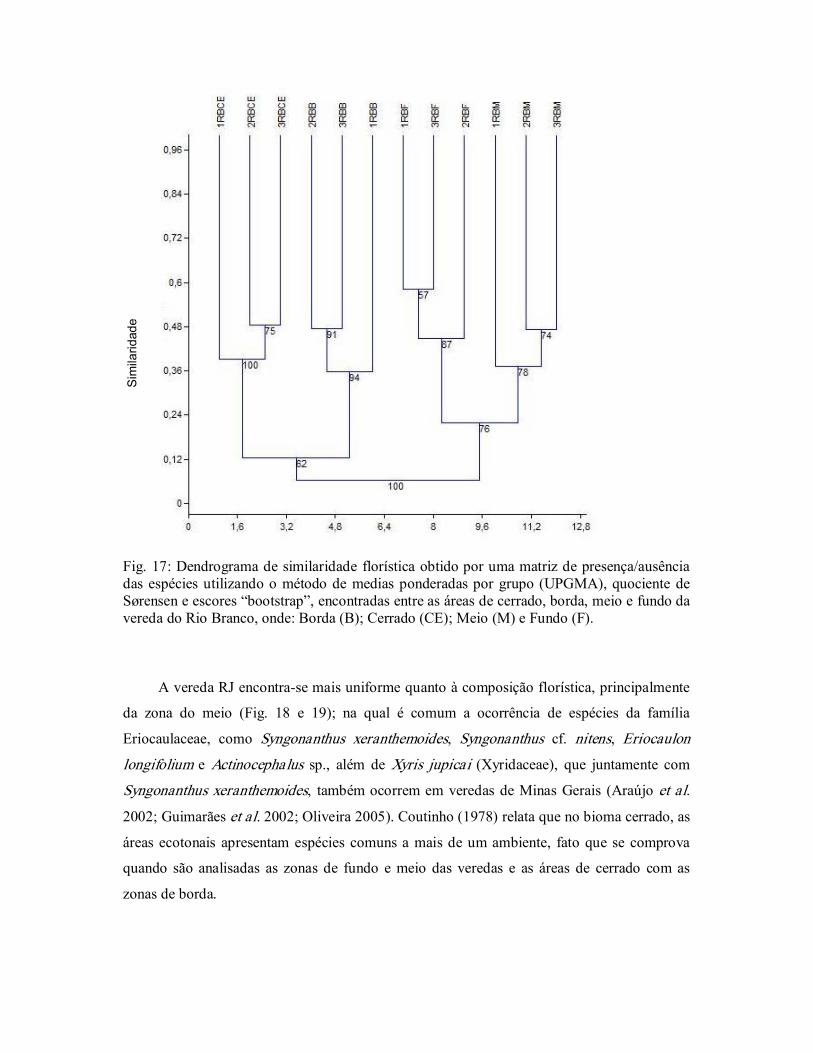
Fig. 17: Dendrograma de similaridade florística obtido por uma matriz de presença/ausência das espécies utilizando o método de medias ponderadas por grupo (UPGMA), quociente de Sørensen e escores “bootstrap”, encontradas entre as áreas de cerrado, borda, meio e fundo da vereda do Rio Branco, onde: Borda (B); Cerrado (CE); Meio (M) e Fundo (F).
A vereda RJ encontrase mais uniforme quanto à composição florística, principalmente
da zona do meio (Fig. 18 e 19); na qual é comum a ocorrência de espécies da família
Eriocaulaceae, como Syngonanthus xeranthemoides, Syngonanthus cf. nitens, Eriocaulon longifolium e Actinocephalus sp., além de Xyris jupicai (Xyridaceae), que juntamente com Syngonanthus xeranthemoides, também ocorrem em veredas de Minas Gerais (Araújo et al.
2002; Guimarães et al. 2002; Oliveira 2005). Coutinho (1978) relata que no bioma cerrado, as áreas ecotonais apresentam espécies comuns a mais de um ambiente, fato que se comprova
quando são analisadas as zonas de fundo e meio das veredas e as áreas de cerrado com as
zonas de borda.
Similaridade
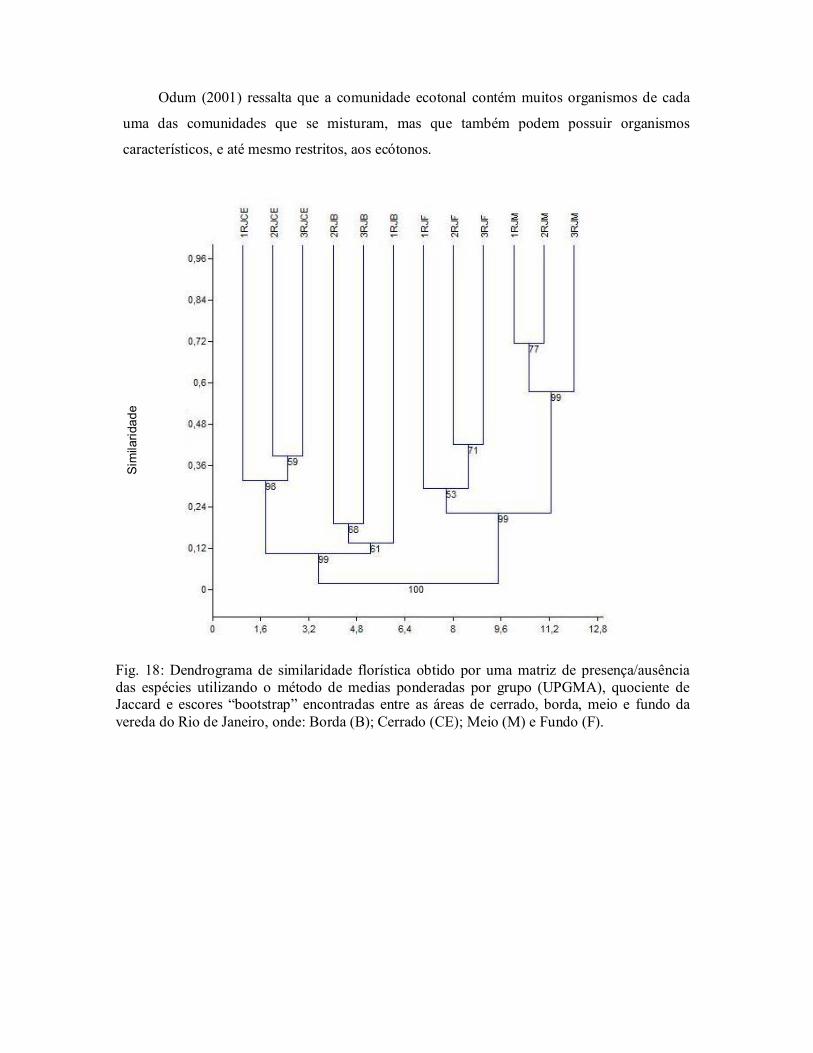
Odum (2001) ressalta que a comunidade ecotonal contém muitos organismos de cada
uma das comunidades que se misturam, mas que também podem possuir organismos
característicos, e até mesmo restritos, aos ecótonos.
Fig. 18: Dendrograma de similaridade florística obtido por uma matriz de presença/ausência das espécies utilizando o método de medias ponderadas por grupo (UPGMA), quociente de Jaccard e escores “bootstrap” encontradas entre as áreas de cerrado, borda, meio e fundo da vereda do Rio de Janeiro, onde: Borda (B); Cerrado (CE); Meio (M) e Fundo (F).
Similaridade
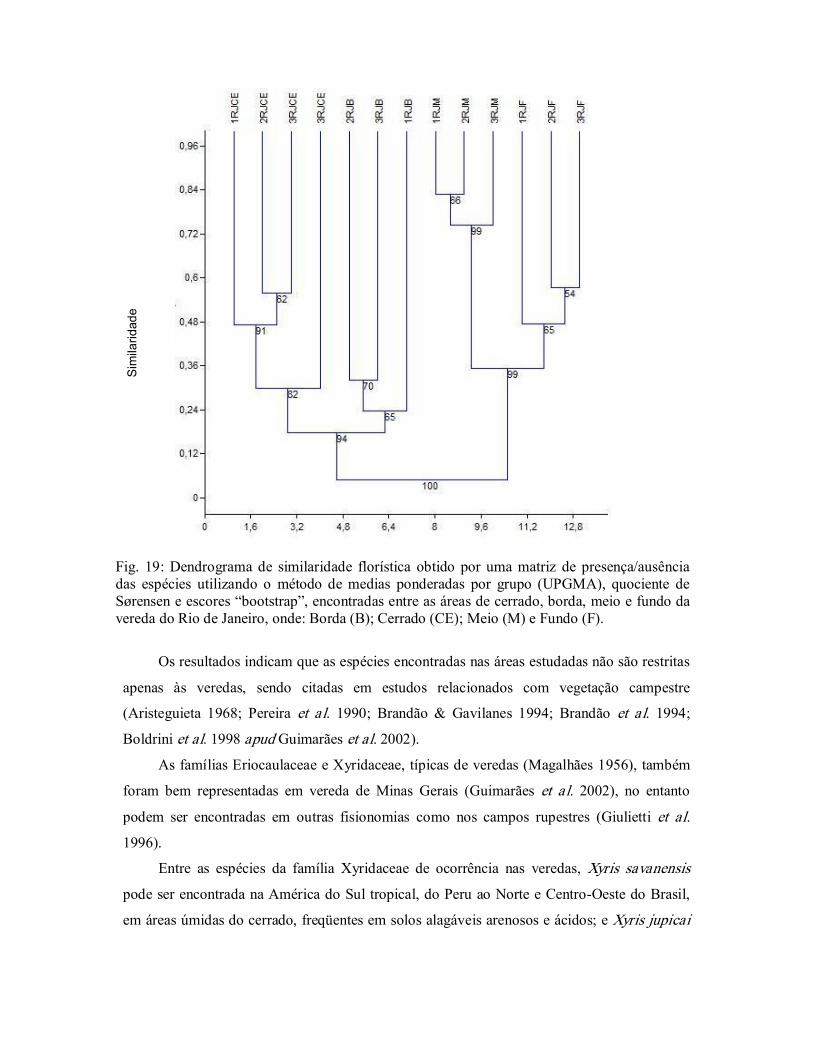
Fig. 19: Dendrograma de similaridade florística obtido por uma matriz de presença/ausência das espécies utilizando o método de medias ponderadas por grupo (UPGMA), quociente de Sørensen e escores “bootstrap”, encontradas entre as áreas de cerrado, borda, meio e fundo da vereda do Rio de Janeiro, onde: Borda (B); Cerrado (CE); Meio (M) e Fundo (F).
Os resultados indicam que as espécies encontradas nas áreas estudadas não são restritas
apenas às veredas, sendo citadas em estudos relacionados com vegetação campestre
(Aristeguieta 1968; Pereira et al. 1990; Brandão & Gavilanes 1994; Brandão et al. 1994; Boldrini et al. 1998 apud Guimarães et al. 2002).
As famílias Eriocaulaceae e Xyridaceae, típicas de veredas (Magalhães 1956), também
foram bem representadas em vereda de Minas Gerais (Guimarães et al. 2002), no entanto
podem ser encontradas em outras fisionomias como nos campos rupestres (Giulietti et al. 1996).
Entre as espécies da família Xyridaceae de ocorrência nas veredas, Xyris savanensis pode ser encontrada na América do Sul tropical, do Peru ao Norte e CentroOeste do Brasil,
em áreas úmidas do cerrado, freqüentes em solos alagáveis arenosos e ácidos; e Xyris jupicai
Similaridade

apresenta ampla distribuição nas Américas, de ocorrência em todo o Brasil, freqüente em
áreas úmidas perturbadas, preferencialmente em superfície orgânica (Pott & Pott 2000).
As famílias Burmanniaceae, Droseraceae, Eriocaulaceae, Rapateaceae, Cyperaceae,
Lentibulariaceae, Orchidaceae e Xyridaceae podem ser encontradas desde áreas com
vegetação campestres, abertas e brejosas, de solos com maior umidade, até em ambientes
onde os solos são mais secos, de difícil drenagem como em campos rupestres (Andrade et al. 1986; Brandão et al. 1994; Giulietti et al. 1996; Araújo et al. 2002).
Tannus & Assis (2004), estudando áreas de campo úmido em São Paulo, constatou
maior riqueza para as famílias Cyperaceae, Poaceae, Lentibulariaceae, Eriocaulaceae e
Xyridaceae, sendo que estas últimas têm registros para ecossistemas semelhantes às veredas
como os “morichales” venezuelanos (Aristeguieta 1968).
A espécie Utricularia gibba (Lentibulariaceae), encontrada nas duas veredas, principalmente nas zonas de meio e fundo, apresenta distribuição pantropical, com ampla
ocorrência no Brasil, em todas as regiões do Pantanal, em águas rasas, solos argilosos ou
arenosos (Pott & Pott 2000); a espécie Epistephium sclerophylum (Orchidaceae), encontrada na vereda RB, apresenta ampla distribuição no Brasil, com registros para a Chapada
Diamantina, podendo ser encontrada também na Venezuela, Peru e Bolívia (Toscano de Brito
2005).
A família Polygalaceae encontrada nas zonas do meio das duas veredas estudadas é
representada pelo gênero Polygala. Esta família apresenta ampla distribuição no globo e para o Brasil existem registros para as regiões Sul, Sudeste e Nordeste (Paiva 1998 apud Silva 2001).
As espécies Annona coriacea e Annona crassiflora (Annonaceae), Arrabidaea brachypoda e Tabebuia ochraceae (Bignoniaceae), Caryocar brasiliense (Caryocaraceae), Curatella americana (Dilleniaceae) e Qualea parviflora (Vochysiaceae) são classificadas por Lorenzi (2002a) como características de áreas de cerrado.
Algumas espécies apresentam registros para outras regiões na Bahia, tendo seu registro
pela primeira vez para a região Oeste do estado da Bahia. Entre estas espécies temse: Gomphrena agrestis (Amaranthaceae) registrada para áreas de campos rupestres em Rio de
Contas na Chapada Diamantina; Burmania capitata (Burmaniaceae) com registros para áreas de caatinga, restinga e campos rupestres; Trachypogon spicatus (Poaceae) encontrado em áreas de cerrado e campo rupestre da Chapada Diamantina; Cambessedesia hilariana (Melastomataceae) encontrada na serra do Sincorá (Harley & Mayo 1980).

Algumas espécies coletadas nas veredas estudadas são classificadas como daninhas: Bulbostylis capillaris e Spermacoce capitata (Lorenzi 2000). A espécie Sagittaria guianensis, coletada apenas na vereda RB, de acordo com Pott & Pott (2000), apresenta ampla
distribuição na América do Sul Tropical, freqüente no Pantanal, em água rasa ou na beira de
lagos, sendo considerada como daninha e infestante de canais de irrigação e drenagem
(Lorenzi 2000).
Nas áreas estudadas não existem registros de outros levantamentos. Muitas espécies
coletadas encontramse apenas ao nível de gênero, o que suscita a ocorrência de novas
espécies e impossibilita a afirmação de registros de espécies endêmicas. Pois regiões pouco
estudadas botanicamente, à medida que se amplia os levantamentos resultaram em novos
registros.
4.3. Cober tura das Espécies
Ao que diz respeito à cobertura das espécies no estrato herbáceo nas áreas estudadas,
apenas as famílias Poaceae, Cyperaceae e Eriocaulaceae apresentaram cobertura superior a
40%, para as áreas estudadas; na vereda do Rio de Janeiro nas zonas do meio e de fundo a
espécie Syngonanthus cf. nitens, apresentou cobertura entre 6080% e 80100%,
respectivamente; já a espécie Cyperus sp. 3 apresentou cobertura entre 80100% apenas para as zonas do meio. A espécie Cephalostemon ridelianus, foi registrada cobertura de 6080%, apenas nas zonas de fundo na vereda do Rio de Janeiro; já na vereda do Rio Branco na mesma
zona a cobertura desta espécie não ultrapassou 20%. A espécie Sagittaria rhombifolia de ocorrência apenas nas zonas de fundo da vereda do Rio Branco apresentou cobertura entre 10
20%, sendo esta espécie comum apenas nas áreas de fundo desta vereda.
Algumas espécies de habito arbustivo e arbóreo apresentaram cobertura superior a 40%,
entre elas merecem destaque: Annona crassiflora, Tabebuia ochraceae, Qualea parviflora,
Qualea grandiflora, Sclerolobium paniculatum e Dimorphandra mollis. A maioria das espécies coletas nas áreas de cerrado e veredas (zonas de borda, meio e
fundo) do Rio de Janeiro e do Rio Branco apresentavam cobertura entre 010%.
Do total de espécimes coletadas 4 apresentaram coberetura entre 80100%, as demais
apresentaram a seguinte distribuição: 8 espécimes entre 6080%, 23 entre 4060%, 31 entre
4060%, 56 entre 2010% e entre 010% (Fig. 20).
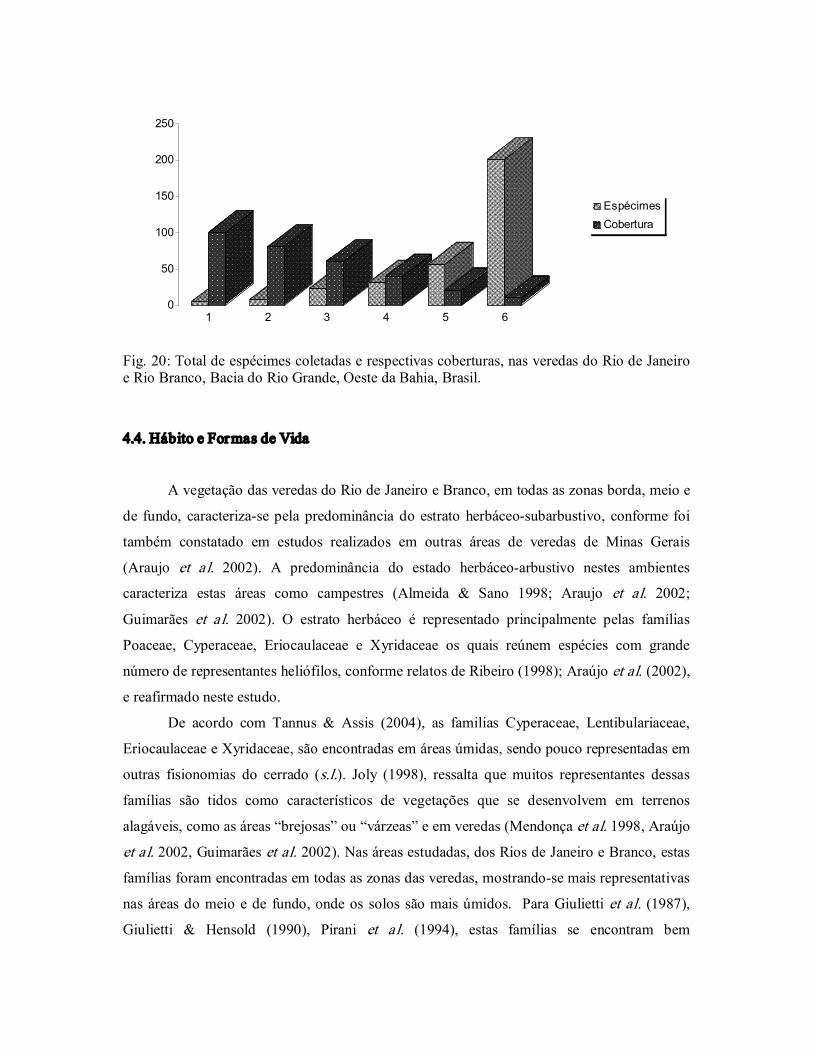
0
50
100
150
200
250
1 2 3 4 5 6
Espécimes Cobertura
Fig. 20: Total de espécimes coletadas e respectivas coberturas, nas veredas do Rio de Janeiro e Rio Branco, Bacia do Rio Grande, Oeste da Bahia, Brasil.
4.4. Hábito e Formas de Vida
A vegetação das veredas do Rio de Janeiro e Branco, em todas as zonas borda, meio e
de fundo, caracterizase pela predominância do estrato herbáceosubarbustivo, conforme foi
também constatado em estudos realizados em outras áreas de veredas de Minas Gerais
(Araujo et al. 2002). A predominância do estado herbáceoarbustivo nestes ambientes caracteriza estas áreas como campestres (Almeida & Sano 1998; Araujo et al. 2002;
Guimarães et al. 2002). O estrato herbáceo é representado principalmente pelas famílias Poaceae, Cyperaceae, Eriocaulaceae e Xyridaceae os quais reúnem espécies com grande
número de representantes heliófilos, conforme relatos de Ribeiro (1998); Araújo et al. (2002), e reafirmado neste estudo.
De acordo com Tannus & Assis (2004), as familias Cyperaceae, Lentibulariaceae,
Eriocaulaceae e Xyridaceae, são encontradas em áreas úmidas, sendo pouco representadas em
outras fisionomias do cerrado (s.l.). Joly (1998), ressalta que muitos representantes dessas famílias são tidos como característicos de vegetações que se desenvolvem em terrenos
alagáveis, como as áreas “brejosas” ou “várzeas” e em veredas (Mendonça et al. 1998, Araújo et al. 2002, Guimarães et al. 2002). Nas áreas estudadas, dos Rios de Janeiro e Branco, estas famílias foram encontradas em todas as zonas das veredas, mostrandose mais representativas
nas áreas do meio e de fundo, onde os solos são mais úmidos. Para Giulietti et al. (1987), Giulietti & Hensold (1990), Pirani et al. (1994), estas famílias se encontram bem

representadas na flora dos campos rupestres, no entanto, o número de espécies em comum
com áreas de campos úmidos é reduzido.
4.4.1. Estrato herbáceo
O estrado herbáceo mostrase mais rico para as famílias Poaceae, Cyperaceae e
Eriocaulaceae. Entre os representantes destas famílias, merecem destaque: Poaceae
representada pelos gêneros: Aristida, Paspalum e Trachypogon; Cyperaceae com
predominância dos gêneros Cyperus, Bulbostylis e Rhinchospora e Euriocaulaceae tendo como maiores representantes os gêneros Paepalanthus e Syngonanthus. A família Poaceae, também apresentou alta riqueza em outras comunidades campestres de Minas Gerais
(Andrade et al. 1986), no Rio Grande do Sul (Zocche & Porto 1992; Boldrini & Egers 1996; Boldrini et al. 1998) e em áreas de cerrado no estado de São Paulo (Montovani & Martins
1993); também mostraramse representativas em áreas campestres no Distrito Federal (Silva
Junior & Felfili 1996) e em Alto Paraíso de Goiás – GO (Munhoz & Felfili 2006). A espécie
Echinolena inflexa (Poaceae) coletada apenas nas zonas de borda da vereda do Rio de Janeiro apresentou menor freqüência em área antropizada que em áreas conservada de acordo com
estudos realizados por Guimarães et al. (2002), este dado corrobora com os resultados encontrados neste estudo, uma vez que esta espécie não apresentou registros na vereda do Rio
Branco, a qual é mais antropizada.
Para Araújo et al. (2002) em estudos realizados nas veredas do Triangulo Mineiro,
existe uma semelhança entre a flora encontrada nas veredas e as fisionomias campestres uma
vez que foram encontradas espécies com folhas filiformes principalmente das famílias
Poaceae e Cyperaceae, em todas as zonas das veredas estudadas. Este estudo mostra que estas
famílias estiveram presentes em todas as áreas estudadas, cerrado e veredas do Oeste da
Bahia. Estudos realizados por Costa (2007), mostraram a grande representatividade destas
famílias principalmente em áreas alagadas. Coutinho (1978) ressalta que estas famílias
reúnem gêneros e espécies que se desenvolvem em ambientes bem iluminados, desta forma,
estes dados complementam os resultados obtidos neste levantamento, uma vez que as veredas
apresentam solos heteromórficos, com elevando teor de umidade e se tratam de áreas abertas.
4.4.1. Estratos subarbustivo e arbustivo

Nos estratos subarbustivos e arbustivos, encontrouse maior representatividade das
famílias Fabaceae, Asteraceae, Rubiaceae, Myrtaceae e Malpighiacea para todas as áreas
estutadas. De acordo Magalhães (1966), AcháPanoso (1978), Carvalho (1991) apud Araújo et al. (2002) o estrato arbustivo e subarbustivo presentes em áreas úmidas das veredas, é caracterizado principalmente por espécies das famílias Melastomataceae e Rubiaceae; no
entanto, nas áreas de coletas do Oeste da Bahia, a família Melastomataceae mostrouse maior
abundancia apenas nas zonas de fundo da vereda do Rio Branco, nas demais área mostrouse
pouco representativa.
4.4.2. Estrato arbóreo
As espécies que constituem o estrato arbóreo presentes principalmente nas áreas de
borda e do cerrado das duas áreas estudadas mostraramse de pequeno porte, atingindo cerca
de 3m de altura. Entre os representados deste estrato, merecem destaque: Fabaceae
(Hymenaea stigonocarpa, Copaifera langsdorffii, Sclerolobium paniculatum e Dimorphandra
mollis.), Vochysiaceae (Qualea parviflora, Qualea grandiflora e Vochysia gardneri) e Annonaceae (Annona coriacea e Annona crassiflora). Estas espécies foram encontradas nas áreas de solos mais secos. Ribeiro & Walter (1998), ressaltam que nas áreas próximas a mata
de galeria, tende a ocorrer o rebaixamento do lençol freático, o que favorece o
estabelecimento de espécies arbóreas, podendo ser encontradas representantes das famílias
Apocynaceae, Fabaceae, Myrtaceae e Annonaceae.
De acordo com Goodland (1970), Mantovani & Martins (1993), Castro et al. (1999) e Araújo et al. (2002), entre as famílias que apresentam grande número de espécies arbustivas e arbóreas de ampla distribuição no cerrado, a família Fabaceae mostrase mais representativa
para diferentes fisionomias. Nas áreas estudadas, dos Rios de Janeiro e Branco, a ocorrência
desta família está associada principalmente às áreas de cerrado e zonas de bordas das veredas.
A grande maioria dos representantes desta família, apresentaram hábito subarbustivo e
arbustivo, demonstrando que o ambiente pode estar limitando o estabelecimento de outras
formas de vida com maior porte. Entre as espécies coletadas nas duas áreas estudadas
merecem destaque: Bauhinia acuruana, Galactia martii, Stylosanthes gracilis e Vigna firmula.
As espécies herbáceas foram encontradas nas áreas de cerrado e nas veredas (zonas de
borda, meio e fundo), apresentando maior abundância nas áreas do meio e do fundo; as
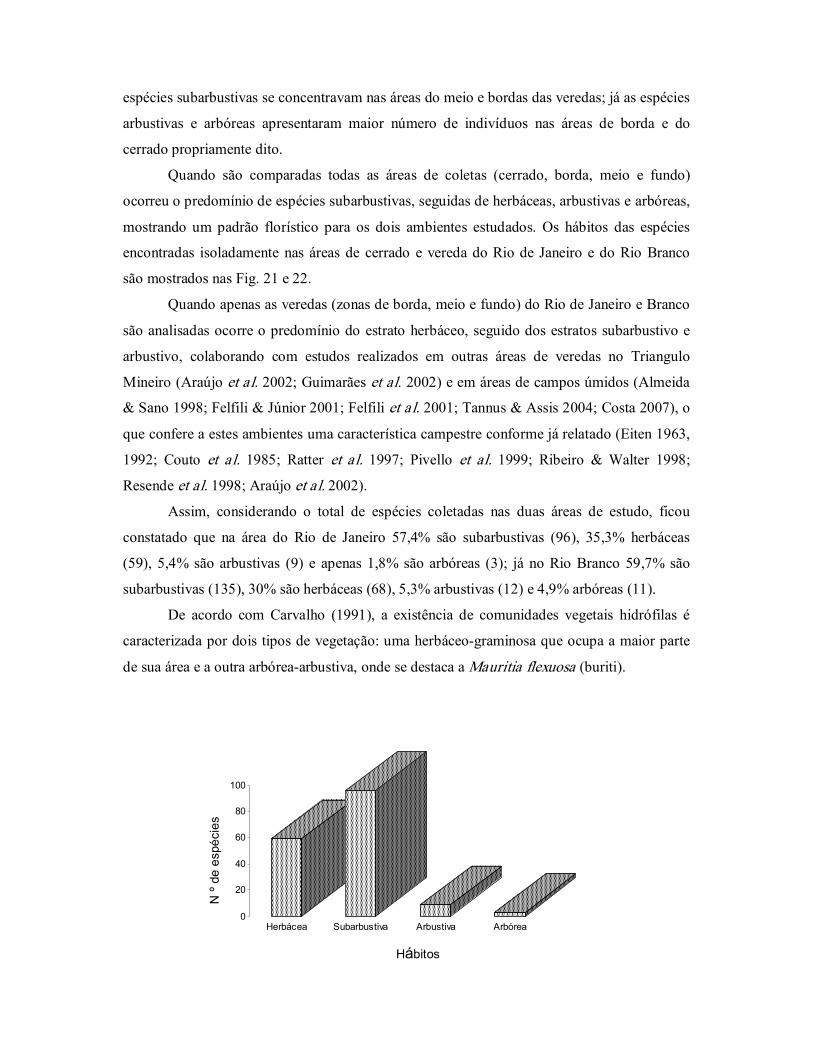
espécies subarbustivas se concentravam nas áreas do meio e bordas das veredas; já as espécies
arbustivas e arbóreas apresentaram maior número de indivíduos nas áreas de borda e do
cerrado propriamente dito.
Quando são comparadas todas as áreas de coletas (cerrado, borda, meio e fundo)
ocorreu o predomínio de espécies subarbustivas, seguidas de herbáceas, arbustivas e arbóreas,
mostrando um padrão florístico para os dois ambientes estudados. Os hábitos das espécies
encontradas isoladamente nas áreas de cerrado e vereda do Rio de Janeiro e do Rio Branco
são mostrados nas Fig. 21 e 22.
Quando apenas as veredas (zonas de borda, meio e fundo) do Rio de Janeiro e Branco
são analisadas ocorre o predomínio do estrato herbáceo, seguido dos estratos subarbustivo e
arbustivo, colaborando com estudos realizados em outras áreas de veredas no Triangulo
Mineiro (Araújo et al. 2002; Guimarães et al. 2002) e em áreas de campos úmidos (Almeida & Sano 1998; Felfili & Júnior 2001; Felfili et al. 2001; Tannus & Assis 2004; Costa 2007), o
que confere a estes ambientes uma característica campestre conforme já relatado (Eiten 1963,
1992; Couto et al. 1985; Ratter et al. 1997; Pivello et al. 1999; Ribeiro & Walter 1998;
Resende et al. 1998; Araújo et al. 2002). Assim, considerando o total de espécies coletadas nas duas áreas de estudo, ficou
constatado que na área do Rio de Janeiro 57,4% são subarbustivas (96), 35,3% herbáceas
(59), 5,4% são arbustivas (9) e apenas 1,8% são arbóreas (3); já no Rio Branco 59,7% são
subarbustivas (135), 30% são herbáceas (68), 5,3% arbustivas (12) e 4,9% arbóreas (11).
De acordo com Carvalho (1991), a existência de comunidades vegetais hidrófilas é
caracterizada por dois tipos de vegetação: uma herbáceograminosa que ocupa a maior parte
de sua área e a outra arbóreaarbustiva, onde se destaca a Mauritia flexuosa (buriti).
0
20
40
60
80
100
Herbácea Subarbustiva Arbustiva Arbórea
N º de espécies
Hábitos
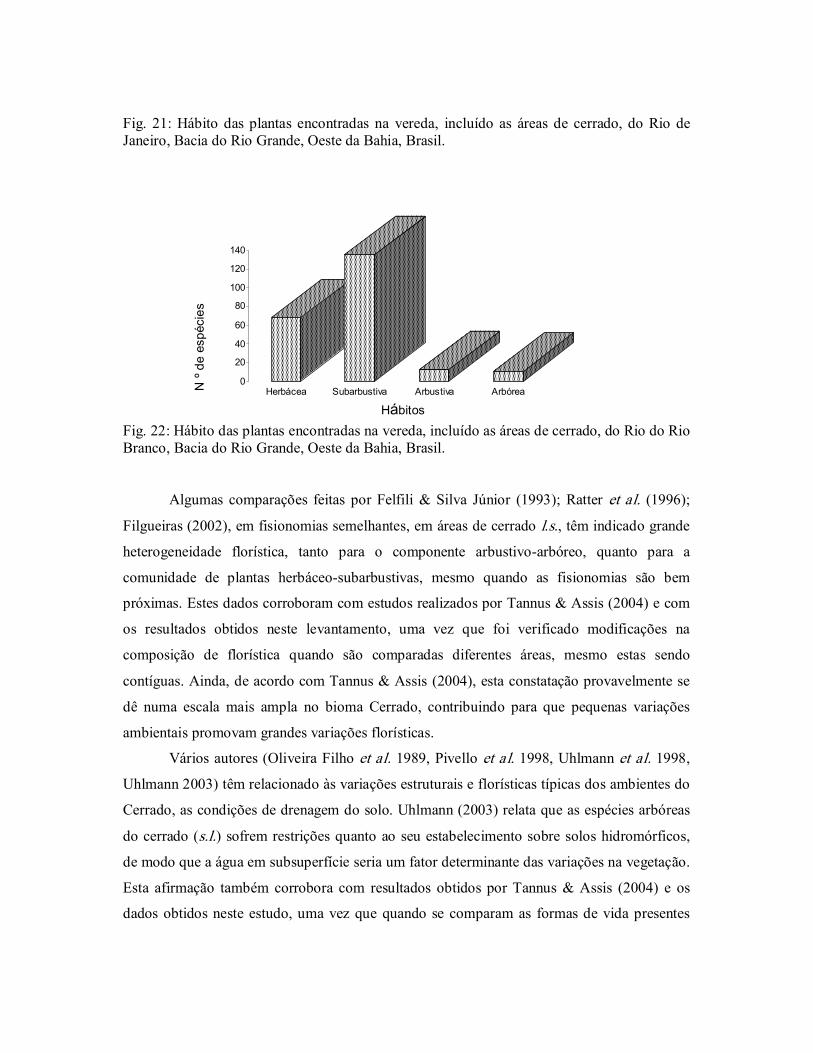
Fig. 21: Hábito das plantas encontradas na vereda, incluído as áreas de cerrado, do Rio de Janeiro, Bacia do Rio Grande, Oeste da Bahia, Brasil.
0
20
40
60
80
100
120
140
Herbácea Subarbustiva Arbustiva Arbórea
Fig. 22: Hábito das plantas encontradas na vereda, incluído as áreas de cerrado, do Rio do Rio Branco, Bacia do Rio Grande, Oeste da Bahia, Brasil.
Algumas comparações feitas por Felfili & Silva Júnior (1993); Ratter et al. (1996);
Filgueiras (2002), em fisionomias semelhantes, em áreas de cerrado l.s., têm indicado grande heterogeneidade florística, tanto para o componente arbustivoarbóreo, quanto para a
comunidade de plantas herbáceosubarbustivas, mesmo quando as fisionomias são bem
próximas. Estes dados corroboram com estudos realizados por Tannus & Assis (2004) e com
os resultados obtidos neste levantamento, uma vez que foi verificado modificações na
composição de florística quando são comparadas diferentes áreas, mesmo estas sendo
contíguas. Ainda, de acordo com Tannus & Assis (2004), esta constatação provavelmente se
dê numa escala mais ampla no bioma Cerrado, contribuindo para que pequenas variações
ambientais promovam grandes variações florísticas.
Vários autores (Oliveira Filho et al. 1989, Pivello et al. 1998, Uhlmann et al. 1998, Uhlmann 2003) têm relacionado às variações estruturais e florísticas típicas dos ambientes do
Cerrado, as condições de drenagem do solo. Uhlmann (2003) relata que as espécies arbóreas
do cerrado (s.l.) sofrem restrições quanto ao seu estabelecimento sobre solos hidromórficos, de modo que a água em subsuperfície seria um fator determinante das variações na vegetação.
Esta afirmação também corrobora com resultados obtidos por Tannus & Assis (2004) e os
dados obtidos neste estudo, uma vez que quando se comparam as formas de vida presentes
N º de espécies
Hábitos
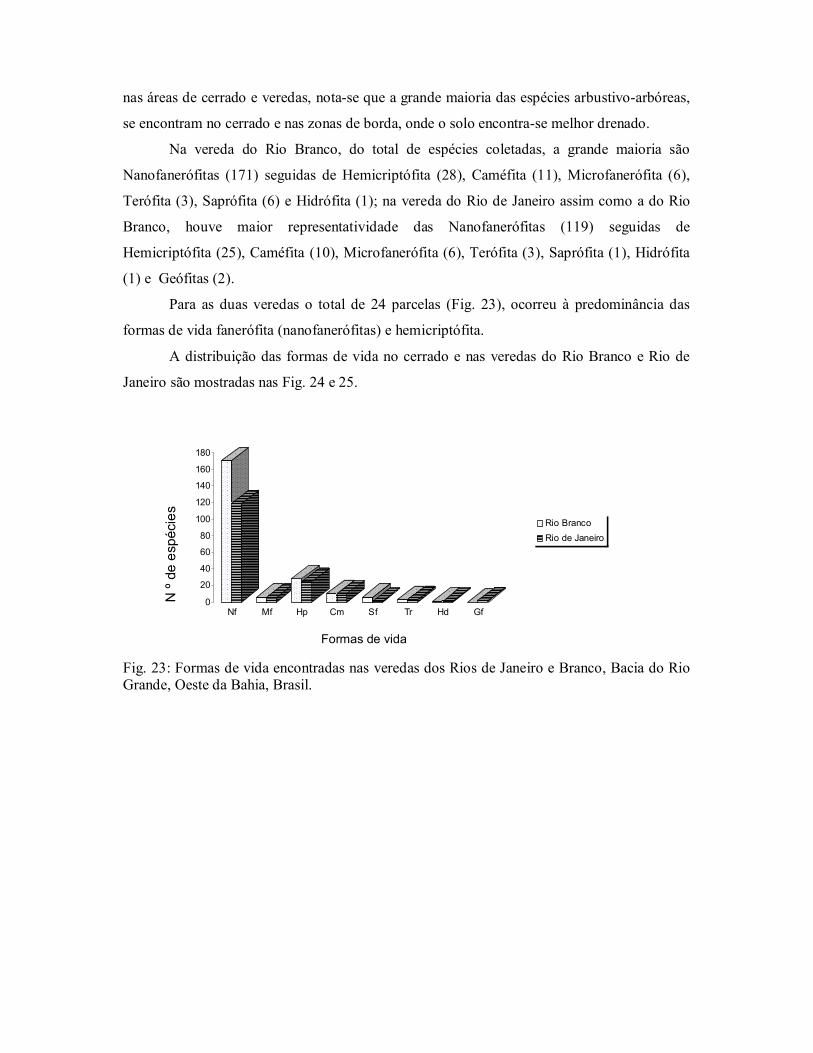
nas áreas de cerrado e veredas, notase que a grande maioria das espécies arbustivoarbóreas,
se encontram no cerrado e nas zonas de borda, onde o solo encontrase melhor drenado.
Na vereda do Rio Branco, do total de espécies coletadas, a grande maioria são
Nanofanerófitas (171) seguidas de Hemicriptófita (28), Caméfita (11), Microfanerófita (6),
Terófita (3), Saprófita (6) e Hidrófita (1); na vereda do Rio de Janeiro assim como a do Rio
Branco, houve maior representatividade das Nanofanerófitas (119) seguidas de
Hemicriptófita (25), Caméfita (10), Microfanerófita (6), Terófita (3), Saprófita (1), Hidrófita
(1) e Geófitas (2).
Para as duas veredas o total de 24 parcelas (Fig. 23), ocorreu à predominância das
formas de vida fanerófita (nanofanerófitas) e hemicriptófita.
A distribuição das formas de vida no cerrado e nas veredas do Rio Branco e Rio de
Janeiro são mostradas nas Fig. 24 e 25.
0
20
40
60
80
100
120
140
160
180
Nf Mf Hp Cm Sf Tr Hd Gf
Rio Branco Rio de Janeiro
Fig. 23: Formas de vida encontradas nas veredas dos Rios de Janeiro e Branco, Bacia do Rio Grande, Oeste da Bahia, Brasil.
N º de espécies
Formas de vida

70%
1%1% 2% 4% 6%
15%
1% NfHp Cm Mf TrGfHd Sf
Fig. 24: Formas de vida encontradas na vereda do Rio de Janeiro, Bacia do Rio Grande, Oeste da Bahia, Brasil. Onde: Nanofanerófitas (Nf), Hemicriptófita (Hp), Caméfita (Cm), Microfanerófita (Mf), Terófita (Tr), Geófita (Gf), Hidrófita (Hd) e Saprófita (Sf).
75%
12%
5% 3% 3% 1%1%
NfHp Cm SfMf TrHd
Fig. 25: Formas de vida encontradas na vereda do Rio Branco, Bacia do Rio Grande, Oeste da Bahia, Brasil. Onde: Nanofanerófitas (Nf), Hemicriptófita (Hp), Caméfita (Cm), Microfanerófita (Mf), Terófita (Tr), Hidrófita (Hd) e Saprófita (Sf).
Do total de plantas encontradas no cerrado e na vereda (zona de borda, meio e fundo)
no Rio de Janeiro estão expressos na Fig. 26, no entanto em todas as áreas houve a maior
representatividade da forma fanerófitica.

0
10
20
30
40
50
60
70
80
90
Nf Mf Hp Cm Tr Sf Hd Gf
Borda Meio Fundo Cerrado
Fig. 26: Formas de vida encontradas nas áreas de coleta (cerrado e vereda) na nascente do Rio de Janeiro, Bacia do Rio Grande, Oeste da Bahia, Brasil. Onde: Nanofanerófitas (Nf), Hemicriptófita (Hp), Caméfita (Cm), Microfanerófita (Mf), Terófita (Tr), Geófita (Gf), Hidrófita (Hd) e Saprófita (Sf).
Na vereda do Rio de Janeiro, foram encontrados exemplares geófitos apenas nas áreas
de borda e no meio da vereda. A presença de microfanerófitas foram restritas apenas para as
áreas de borda e cerrado propriamente dito. Nesta vereda exemplares terófitos e saprófitos
foram encontrados apenas em áreas intermediárias e alagadas.
Na vereda do Rio Branco (Fig. 27), assim como na vereda do Rio de Janeiro, as
fanerófitas (nanofanerófitas) predominaram em todas as áreas, já as microfanerófitos fezse
presente nas áreas de borda, cerrado e de fundo; as hemicriptófitas mostrouse presente além
das 3 zonas encontras nas veredas: borda, meio e no fundo, tendo também registros no
cerrado. Exemplares caméfitos foram registrados nas 3 zonas (borda, meio e fundo) das duas
veredas, no entanto, mostrouse pouco representativo (11). Verificouse que a grande maioria
das plantas coletadas não são restritas a apenas uma zona.
Formas de vida
N º de espécies

0 10 20
30 40
50 60
70 80
90 100
Nf Mf Hp Cm Tr Sf Hd
Borda Meio Fundo Cerrado
Fig. 27: Formas de vida encontradas nas áreas de coleta na nascente (cerrado e vereda) no Rio Branco, Bacia do Rio Grande, Oeste da Bahia, Brasil. Onde: Nanofanerófitas (Nf), Hemicriptófita (Hp), Caméfita (Cm), Microfanerófita (Mf), Terófita (Tr), Hidrófita (Hd) e Saprófita (Sf).
Quando são avaliadas as duas veredas, entre as espécies fanerofitas merecem destaque: Anacardium humile (Anacardiaceae), Annona coriacea e Annona crassiflora (Annonaceae), Tabebuia ochraceae (Bignoniaceae), Merremia digitata (Convolvulaceae), Davilla elliptica (Dilleniaceae), Bauhinia acuruana, Galactia martii, Stylosanthes gracilis, Vigna firmula (Fabaceae), Sauvagesia linearifolia (Ochnaceae) e Declieuxia fruticosa (Rubiaceae), espécies encontradas principalmente nas zonas de borda e do cerrado. Entre as hemicriptófitas as
famílias Cyperaceae (Rhynchospora spp) e Poaceae (Panicum rupestre, Paspalum foveolatum, Paspalum sp. 2, Trachypogon spicatus, merecem destaque. Entre as caméfitas
foram coletadas Drosera comunis (Droseraceae) e Eriocaulon longifolium (Eriocaulaceae); as saprofitas Burmania capitata (Burmaniaceae), Lagenocarpus rigidus (Cyperaceae) e Habenaria cf. secundiflora (Orchidaceae); já as terófitas foram representadas por Utricularia cf. amethystina (Lentibulariaceae).
A predominância de espécies nanofanerófitas está relaciona a formação campestre
encontradas nas duas áreas estudadas, uma vez que foram incluídas as áreas de cerrado
encontrada no entorno de cada vereda. No entanto, quando são avaliadas apenas as zonas de
borda, meio e fundo das veredas acontece a mesma distribuição com maior representatividade
de espécies nanofanerófitas (Fig.28).
N º de espécies
Formas de vida

0
20 40
60
80 100
120 140 160
180 200
Nf Mf Hp Cm Tr Sf Hd Gf
Borda Meio Fundo
Fig. 28: Formas de vida encontradas nas áreas de coleta nas zonas de borda, meio e fundo, nas veredas do Rio Branco e Rio de Janeiro, Bacia do Rio Grande, Oeste da Bahia, Brasil. Onde: Nanofanerófitas (Nf), Hemicriptófita (Hp), Caméfita (Cm), Microfanerófita (Mf), Terófita (Tr), Hidrófita (Hd) e Saprófita (Sf).
Estudos realizados por Tannus & Assis (2004) em áreas de campo úmido em São
Paulo, o único componente florístico verificado foi o herbáceosubarbustivo, com predomínio
de espécies hemicriptófitas; entre as famílias mais ricas foram encontradas Cyperaceae e
Poaceae. Corroborando com os levantamentos encontrados neste estudo. No Foram
encontradas neste ambiente, também formas de vida geófitas representados pelas famílias
Cyperaceae, Araceae e Lycopodiaceae; terófitas por Orchidaceae, Polygalaceae e Rubiaceae;
sendo que as caméfitas foi representada apenas pela família Melastomataceae.
Para alguns autores Huntchinnchon (1975), Spence (1985) apud Costa (2007), a
profundidade da água produz variações nas formas de vida. Nos ambientes úmidos o padrão
de distribuição espacial das plantas é normalmente relacionado á topografia e ao nível de água
subterrânea, os quais determinam a freqüência de alagamento, a umidade do solo e a
cobertura vegetal associada (Mendonça & Castellani 1993). Fato que se comprova quando se
analisa a ocorrência das espécies em diferentes zonas das veredas.

5 CONSIDERAÇÕES FINAIS
As áreas estudadas de cerrado s.s. e veredas apresentam similaridade fisionômica, as quais possivelmente esta relacionada ao gradiente de umidade encontrado no ambiente. Quando são
comparadas as veredas (zonas de borda, meio e fundo) e o cerrado, há maior afinidade
florística entre o cerrado e a borda, e entre as zonas do meio e de fundo nas duas áreas
estudadas. As zonas de bordas das veredas apresentam maior diversidade quando comparadas
com as demais zonas e cerrado s.s., sendo constatada maior diversidade no ambiente mais seco. A maior abundância foi verificada nas zonas de fundo das veredas, apesar das condições
de umidade limitar a adaptação de diferentes espécies, tal fato está relacionado à dominância
de poucas espécies neste ambiente. A posição do lençol freático e a variação da umidade
edáfica devem ser os fatores preponderantes na distribuição das espécies. A forma de vida
fanerófítica (nanofanerófita) e o estrato herbáceoarbustivo se mostraram mais representativos
para as duas áreas analisadas. Em geral, a maioria das espécies coletadas nestas áreas não é
restrita a estes ambientes, podendo ser encontradas em outras formações de gradiante
ambiental semelhante. Tendo em vista que a maior diversidade vegetal foi encontrada nas
zonas de bordas das veredas tornase importante a ampliação das áreas de preservação
permantente (APP) destes ambientes, a fim conservar a biodiversidade.

6 RESUMO
(Composição Florística e Estrutura da vegetação de veredas do Cerrado no Oeste da Bahia,
Brasil). As veredas são sistemas úmidos com vegetação herbáceasubarbustiva e presença
típica de Mauritia flexuosa. Esses ambientes são Áreas de Preservação Permanente e participam do controle do lençol freático desempenhando importante papel no equilíbrio dos
cursos d’água, além de contribuírem para manutenção da fauna. Este trabalho apresenta
estudo florístico e estrutural comparativo entre duas veredas localizadas nas nascentes do Rio
Branco e Rio de Janeiro no Oeste da Bahia. A vegetação foi amostrada em parcelas de 10 x 10
m sorteadas em distintas zonas (borda, meio e fundo) das duas veredas e nos cerrados
circundantes. Foi realizado o levantamento das espécies e processadas análises da
similaridade florística e cálculos de parâmetros de diversidade. Foram reconhecidas 323
espécies, sendo apenas 64 comuns às duas veredas. A forma de vida fanerofítica
(nanofanerófita) e o estrato herbáceoarbustivo se mostraram mais representativo para todas
as áreas analisadas. A maioria das espécies coletadas não ultrapassaram cobertura de 10%. O
índice de diversidade de ShannonWienner variou de 1,55 a 3,45 nats/indivíduos, sendo que
as bordas das veredas foram significativamente mais diversas que as demais zonas estudadas.
Constatouse que as zonas mais úmidas das veredas (fundo e meio) tendem a possuir um
padrão florístico diferente das partes mais secas (zona da borda e cerrado), demonstrando a
importância do ambiente nas veredas. A formação de grupos das mesmas zonas e veredas
demonstra a importância das áreas geográficas na composição das espécies. Na vereda Rio de
Janeiro, as zonas do meio apresentaramse mais similares entre si, enquanto que na vereda
Rio Branco a maior similaridade foi entre as zonas de fundo.

7 ABSTRACT
(Floristic composition and structure of vegetation of the “veredas” of savanna in the west of Bahia,
Brazil). The “veredas” are swamp systems with vegetation herbaceous shrub and typical presence
of Mauritia flexuosa. These environment are Permanent Preservation Area and participate of control the ground water playing important function in the equilibrium of watercourses, beyond to
contribute for maintenance the fauna. This work presents floristic and comparative structural
studies between two “veredas” localized in nascent the rivers Branco and Rio de Janeiro in the west
of Bahia. The vegetation was sampled in plots of 10 x 10 m in the distinct areas (edge, middle and
background) of the two “veredas” and surrounding savanna. It was realized survey of species and to
processed analysis of the floristic similarity and calculations to diversity parameters. Were
recognized 323 species, being only 64 commor of the two “veredas”. The life form phanerophytes
(nanophanerophytes) and the stratum herbaceousshrub were the more representative for all the
analyzed areas. The most of species collected not exceeded the coverage of the 10%. The Shannon
Wienner index diversity ranged between of 1,55 to 3,45 nats/individual, being that the of “veredas”
edge were significantly more diverse than the other studied areas. The areas more swamp of the
“veredas” (middle and background) tend to have a different floristic pattern a the more drying
areas (edge and savanna), showing the importance of “veredas” areas. The formation of groups of
the same areas and “veredas” show the importance of the geograpghic areas in the species
composition. In the “veredas” of Rio de Janeiro, the areas of middle showed up more similars
together, while in the “veredas” of the Rio Branco the more similarity was backgroud areas.

8 REFERÊNCIAS
ALMEIDA, J.R., BARUQUI, F.M., BARUQUI, A.M. & MOTTA, P.E.F. 1983. Principais
solos de várzeas do Estado de Minas Gerais e suas potencialidades agrícolas. Informe
Agropecuár io 9:7078.
ALMEIDA, S.P.; Proença, C.E.B.; Sano, S.M.; Ribeiro, J.F. 1998. Cer rado: espécies
vegetais úteis. Planaltina: EMBRAPACPAC, 160 p.il..
AMARAL, A.F. 2002. Caracterização fenológica, e aspectos do solo em áreas queimada e
desbastada de uma vereda em Uberlândia, MG. Disser tação de Mestrado, Universidade
Federal de Uberlândia, Uberlândia.
ANDRADE, P.M., GONTIJO, T.A. & GRANDI, T.S.M. 1986. Composição florística e
aspectos estruturais de uma área de “campo rupestre” do morro do Chapéu, Nova Lima,
Minas Gerais. Revista Brasileira de Botânica. v.9, p.1321.
APG II. 2003. An update of the Angiosperm Phylogeny Group classification for the orders
and families of flowering plants: APG II. Botanical Journal of the Linnean Society 41:399
439.
ARAÚJO, G.M.; BARBOSA, A.A.A.; ARANTES, A. A. & AMARAL A.F. 2002.
Composição florística de veredas no Município de Uberlândia, MG. Revista Brasileira de
Botânica. v.25, n.4., p.475493.
ARISTEGUIETA, L. 1968. Consideraciones sobre la flora de los Morichales Llaneros al
Norte del Orinoco. Acta Botanica Venezuelana. 3: 3 22.

BAHIA. Secretaria de Agricultura, Irrigação e Reforma Agrária. 1997. Política Florestal do
Estado da Bahia: Lei nº 6.569 de 17 de janeiro de 1994 e Decreto nº 6.785 de 23 de
setembro de 1997. 62p. Publicado em D.O.E. em 18.01.1994.
BAHIA. Centro de Centro Recursos Ambientais/CRA. 2001. Bacia do Rio São Francisco:
Sér ie Áreas de Proteção Ambiental. v.2, Salvador, Bahia. 143p.il.
BRASIL. 1992. Resoluções CONAMA de 1984 a 1991. SEMAM / IBAMA. Brasília, DF. 4ª
ed. 245 p.
BRASIL. Congresso. Senado. Resolução. 2002. Resolução CONAMA n. º 303, de
20/03/2002. Dispõe sobre parâmetros, definições e limites de áreas de Preservação
Permanente. Disponível em: <http:// www.mma.gov.br/conama/>. Acesso em: 02.05.2007.
BRASIL. Congresso. Senado. Resolução. 2006. Resolução CONAMA nº 369, de
28/03/2006. Dispõe sobre parâmetros, definições e limites de áreas de Preservação
Permanente. Disponível em: <http:// www.mma.gov.br/conama/>. Acesso em: 02.05.2007.
BRANDÃO, M. & GAVILANES, M.L. 1994. Cobertura Vegetal da Microrregião 178
(Uberaba), Minas Gerais, Brasil. Daphne.v.4, n.2, p. 2957.
BRANDÃO, M., GAVILANES, M.L. & ARAÚJO, M.G. 1994. Aspectos físicos e botânicos
de campos rupestres do Estado de Minas GeraisI. Daphne. v.4, n.1, p.1738.
BOAVENTURA, R. S. 1988. Preservação das veredas – síntese. 109–122p. In: Anais do 2º
Encontro latinoamer icano: r elação ser humanoambiente, Chapter. Belo Horizonte.
BOLDRINI, I.I., EGGERS, L. 1996. Vegetação campestre do Sul do Brasil: dinâmica de
espécies à exclusão do gado. Acta Botanica Brasílica. v.10, p.3758.
BOLDRINI, I.I., MIOTTO, S.T.S., LONGHIWAGNER, H.M., PILLAR, V.P. &
MARZALL, K. 1998. Aspectos florísticos e ecológicos da vegetação campestre do Morro da
Polícia, Porto Alegre, RS. Acta Botânica Brasílica. v.12, p.89100.

CAIAFA, A. N. & SILVA, A. F. 2005. Composição florística e espectro biológico de um
campo de altitude no Parque Estadual da Serra do Brigadeiro, Minas Gerais, Brasil.
Rodr iguésia. v.56, n.87, p.163173.
CALEGARIJACQUES, S.M. 2003. Bioestatística: princípios e aplicações. Artmed. Porto
Alegre.
CARVALHO, P.G.S. 1991. As Veredas e sua Importância no Domínio dos Cerrados.
Informe Agropecuár io: v.168, p.4754.
CASTRO, A.A.J.F., MARTINS, F.R., TAMASHIRO, J.Y. & SHEPHERD, G.J. 1999. How
rich is flora of Brazilian cerrados? Annals of Missour i Botanical Garden. v.86, p.192224.
CASTRO, J.P.C. 1980. As veredas e a sua proteção jurídica. Análise e Conjuntura. v.10,
p.321331.
CATTANIO, J.H., ANDERSON, A.B. & CARVALHO, M.S. 2002. Floristic composition
and topographic variation in a tidal floodplain Forest in the Amazon Estuary. Revista
Brasileira de Botânica. v.25, p. 419430.
CODY, M.L.; MOONEY, H.A. 1978. Convergencecversus nonconvergence in
Mediterranenclimate ecosystems. Annul review of ecology and systematics. Stanford. v.9 p.
265321.
CONCEIÇÃO, A. A. & GIULIETTI, A. M. 2002. Composição florística e aspectos
estruturais de campo rupestre em dois platôs do Morro do Pai Inácio, Chapada Diamantina,
Bahia. Hoehnea. v.29, n.1, p. 3748p.
CONCEIÇÃO, A. A. 2003. Ecologia da vegetação em afloramentos rochosos na Chapada
Diamantina, Bahia, Brasil. Tese de Doutorado. Instituto de Biociências da Universidade de
São Paulo. Departamento de Botânica. 262 p.

CONCEIÇÃO, A. A. & PIRANI, J. R. 2005. Delimitação de habitats em Campos Rupestres
na Chapada Diamantina, Bahia: substratos, composição florística e aspectos estruturais.
Revista Brasileira de Botânica. Univ. São Paulo. v.23, n.1, p.85111.
COSTA, F. da, A. 2007. Zonação no gradiente vegetacional Cerrado Típico – Cerrado Sujo –
Vereda, na Estação Ecológica de Águas Emendadas, Brasília DF. Disser tação de
Mestrado. Departamento de Ecologia, 65p.
COUTINHO, L.M. 1978. O conceito de Cerrado. Revista Brasileira de Botânica. v.1. p.17
23.
COUTO, E.G., RESENDE, M.B. & REZENDE, S.B. 1985. Terra ardendo. Ciência Hoje.
16:4857.
CUNHA, A.S.; MUELLER, C.C.; ALVES, E.R.A.; SILVA, J.E. 1993. Uma avaliação da
sustentabilidade da agricultura nos cerrados. Brasília, IPEA, n.2.
EBDA. 2006. Empresa Baiana de Desenvolvimento Agropecuário. Boletim Informativo.
Barreiras, BA.
EFRON, B.; TIBSHIRANI, R. 1986. Bootstrap methods for standard errors, confidence intervals, and other measures of statistical accuracy, Statistical Science. v. 1, n. 1, p. 55–77.
EITEN, G. 1963. Habitat flora of Fazenda Campininha, São Paulo, Brasil. In Simpósio sobre o Cerrado (M.G. Fer ri, ed.). Editora da Universidade de São Paulo, São Paulo, p.179231.
EITEN, G. 1983. Classificação da vegetação do Brasil. CNPq, Brasília.
EITEN, G. Vegetação do Cerrado. In: Pinto, M.N. (org.). 1990. Cerrado: caracter ização,
ocupação e perspectivas. Editora Universidade de Brasília UNB, Brasília, 0965p.
EITEN, G. 1992. Natural Brazilian vegetation types and their causes. Anais da Academia
Brasileira de Ciências. v.64, p.3565.
EITEN, G. 1994. Cerrado: carater ização, ocupacão e per spectivas (2ed)., Chapter
Vegetacão do Cerrado. Brasília (DF). 1773p.

EMBRAPA. 1982. Levantamento de reconhecimento de média intensidade dos solos e
avaliação da aptidão agr ícola das terras do Tr iângulo Mineiro. Serviço nacional de
levantamento e conservação de solos. Epamig – DRNR, Rio de Janeiro.
EMBRAPA. 1999. Centro Nacional de Pesquisas de Solos. Sistema Brasileiro de
Classificação de Solos. Rio de Janeiro: Embrapa, CNPS, 412p.
FELFILI, J.M.; SILVA JÚNIOR. M.C.; FILGUEIRAS, T.S. & NOGUEIRA, P.E. 1998.
Comparison of cerrado (sensu stricto) vegetation in central Brazil. Ciência e Cultura. v.50, p.237243.
FELFILI, J.M.; CARVALHO, F.A.; HAIDAR, R.F. 2005. Manual de monitoramento de
parcelas permanentes no Bioma Cer rado e Pantanal. UNB, Departamento de Engenharia
Florestal. Brasília, DF. 55p.il. .
FELFILI, J.M. & JÚNIOR, M.C.S. 2001. Biogeografia do Bioma Cer rado: Estudo
Fisionômico do da Chapada do Espigão Mestre do São Francisco. UNB, Departamento de
Engenharia Florestal. Brasília, DF. 152p.il.
FERREIRA, M.B. 1980. O cerrado em Minas Gerais gradações e composição florística.
Informe Agropecuár io. v.61, p.48.
FERREIRA, I.M. 2005. Bioma Cerrado: caracterização do subsistema de vereda. IX
EREGEO – Encontro terr itorialidades – integração e redefinição regional. Porto
Nacional.
FILGUEIRAS, T.S. 2002. Herbaceous plant communities. In The Cerrados of Brazil:
Ecology and natural history of a neotropical savanna (P.S. Oliveira & J.R. Marquis, eds.).
Columbia University Press, New York, p.121139.
FRANÇA, F.; MELO, E.; GONÇALVES, J.M. 2006. Aspectos da diversidade da vegetação
no topo de um inselbergue no semiárido da Bahia, Brasil. Sitientibus Sér ie Ciências
Biológicas. v.6, n.1, p.3035.

FONSCECA, V.S. & SILVA, I.M. 1998. Etnobotânica: base para conservação. Workshop
Brasileiro de Etnobiologia, 136p.
GIULIETTI, A.M., MENEZES, N.L., PIRANI, J.R., MEGURO, M. & WANDERLEY,
M.G.L. 1987. Flora da Serra do Cipó, Minas Gerais: caracterização e lista de espécies.
Boletim de Botânica da Universidade de São Paulo. v.9, p.1151.
GIULIETTI, A.M. & HENSOLD, N. 1990. Padrões de distribuição geográfica dos gêneros de
Eriocaulaceae. Acta Botanica Brasílica. v.4, p.133158.
GIULIETTI, A. M.; QUEIROZ, L. P. & HARLEY, R. M. 1996. Vegetação e flora da
Chapada Diamantina, Bahia. Pp. 144156. In: Anais 4ª r eunião especial da SBPC, Feira de
Santana, Bahia.
GUIMARÃES, A.J.M.; ARAÚJO, G.M. de; CORRÊA, G.F. 2002. Estrutura fitossociológica
em área natural e antropizada de uma vereda em Uberlândia, MG. Acta Botânica Brasílica.
v. 16, n.3, p.317329.
HALL, P. 1988. Theorical comparison of bootstrap confidence intervals, The Annals of Statistics. v.16, n.3, p.987–953.
HAMILTON, S.K. 1993. Características limnologicas de importância para plantas aquáticas
no Pantanal. II Encontro de Botânicos do Centro Oeste CEUC/UEMS, Anais... Brasília,
SBB. p.14.
HAMMER, Ø.; HARPER, D.A.T.; AND P. D. RYAN. 2001. PAST: Paleontological
Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica
4(1): 9pp. http://palaeoelectronica.org/2001_1/past/issue1_01.htm
HARLEY, R.M. & MAYO, S.J. 1980. Towards a checklist of the flora of Bahia. Royal
Botanic Gardens, Kew, Richmond, Surrey, England. 250p.
HICKMAN, C.A. 1990. Forest – wetland trends in the United Satate: an economic
perspective. Forest Ecological Management. v.3334, p.227238.

IVANAUSKAS, N.M., RODRIGUES, R.R. & NAVE, A.G. 1997. Aspectos ecológicos de
um trecho de floresta de brejo em Itatinga, SP: florística, fitossociologia e seletividade de
espécies. Revista Brasileira de Botânica. v.20, p.139154.
IRGANG, B. & GASTAL Jr., C.V. 1996. Macrófitas aquáticas da planície costeira do Rio
Grande do Sul. Porto Alegre, UFRS.
JOLY, A.B. 1979. Botânica: introdução à taxonomia vegetal. Companhia Editora Nacional,
São Paulo. p.il. 739.
JOSEPHON, J. 1992. Status of wetland. Environmental Science & Technology. V.26, p.
422.
KLINK, C.A., MOREIRA, A.G. & SOLBRIG, O.T. 1993. Ecological impact of agr icultural
development in the Brazilian cer rados. In The World’s savanna: economic driving forces,
ecological constraints and policy options for sustainable land use (M.D. Young & O.T.
Solbrig, eds.). Man and biosphere series. Unesco/Parthenon, Paris, v.12, p.259282.
KÖPPEN, W. 1948. Climatologia. Fondo de Cultura Económica, México.
LIMA, W. P. 1989. Função hidrológica da mata ciliar. In: SIMPÓSIO SOBRE MATA
CILIAR, Anais... Campinas: Fundação Cargil. p. 2542.
LIMA, S.C. & QUEIROZ NETO, J.P. 1996. As veredas e a evolução do relevo. Sociedade e
Natureza. v.15, p.481488.
LOEFGREN, A. 1980. Contribuição para a botânica paulista. Região Campestre. Boletim da
Comissão Geográfica e Geológica do Estado de São Paulo. v.5, p. 351.
LORENZI, H. 2000. Plantas Daninhas do Brasil Terrestres, Aquáticas, Parasitas e
Tóxicas. Nova Odessa, Instituto Plantarum.
LORENZI, H. 2002. Árvores Brasileiras: manual de identificação e cultivo de plantas
arbóreas do Brasil, vol.1, SP: Instituto Plantarum.

LORENZI, H. 2002. Árvores Brasileiras: manual de identificação e cultivo de plantas
arbóreas do Brasil, vol.2, SP: Instituto Plantarum.
LORENZI, H; SOUZA, H.M de; COSTA, T.J.de M.; CERQUEIRA, L.S.C.de; FERREIRA,
E. 2004. Palmeiras Brasileiras e Exóticas Cultivadas. Nova Odessa, SP: Instituto
Plantantarum. 415p.l.
KEDDY, P.A. 2000. Wetland Ecology: pr inciples and conservation. Combridge:
Combridge University Press. 614p.
KENT, M.; COKER, P. 1994. Vegetation descr iption and analysis. A Practical Approach. J.
Wiley, Chichester, 2d. 363p.
KREBS, C. J. 1986. Ecologiaanálisis exper imental de la distr ibuición y abundancia.
Pirámide, S. A. 782p.
KREBS, C. J. 1989. Ecological methodology. New York: Harper & Row, Publishers.
MAGALHÃES, G.M. 1966. Características de alguns tipos florísticos de Minas Gerais.
Revista Brasileira de Biologia. v.1, p.7692.
MANTOVANI, W. & MARTINS, F.R. 1993. Florística do cerrado na Reserva Biológica de
MojiGuaçu SP. Acta Botanica Brasilica. v.7, p.3359.
MATA, A; QUEVEDO, F. 1992. Diccionar io ditactico de la ecologia. Editorial de la
Universidad de la Costa Rica. São José, Costa Rica. 396p.
MEIRELES, M.L.; OLIVEIRA, R.C. de; RIBEIRO, J.F.; VIVELDI, L.J.; RODRIGUES,
L.A.; SILVA, J.P. 2002. Espécies do estrato herbáceo e altura do lençol freático em áreas
úmidas do cerrado (Planaltina – DF). Boletim de Pesquisa e Desenvolvimento. Planaltina –
DF, Embrapa Cerrados, pp. 4168.
MEIRELES, M.L.; GUIMARÃES, A.J.M; OLIVEIRA, R.C.de; ARAÚJO, G.M.; RIBEIRO,
J.F. 2004. Impactos sobre o estrato herbáceo de áreas úmidas do cerrado. In: AGUIAR,

L.M.S.; CAMARGO, A. J.A. (Ed.). Cerrado: Ecologia e caracterização. Planaltina: DF.
Embrapa Cerrado. Pp4168.
MELO, D.R. 1992. As veredas nos planaltos do noroeste mineiro; caracterizações
pedológicas e os aspectos morfológicos e evolutivos. Disser tação de Mestrado. UNESP, Rio
Claro.
MENDONÇA, R.C., FELFILI, J.M., WALTER, B.M.T., SILVA JÚNIOR, M.C., REZENDE,
A.V., FILGUEIRAS, T.S. & NOGUEIRA, P.E. 1998. Flora Vascular do Cerrado. In:
Cer rado ambiente e flora. EMBRAPACPAC. 556p.
MENDONÇA, E.N.; CASTELLANI, T.T. 1993. Aspecto da ecologia populacional de
Drosera brevifolia Pursh em um trecho de baixada úmida de dunas. Biotemas. v 6. n 1p. 31
48.
MORI, S.A. L. MATTOSSILVA; G. LISBOA; L. CORADIN. 1989. Manual de Manejo do
Herbár io Fanerogâmico. 2 ª ed. Ilhéus: CEPEC.
MUELLERDOMBOIS, D.; ELLENBERG, H. 1974. Aims and Methods of Vegetation
Ecology. New York: John Wiley & Sons.
MUNHOZ, C. B.R; FELFILI, J.M. 2006. Flaristic of the herbaceus and subshorub layer of a
moist grassland in the cerrado Biosphere Reserve (Alto Pariso de Goias) Brazil. Endiburgh
Journal of Botany. v.63. n.2 & 3 p.0343354.
ODUM, E.P. & SARMIENTO, F.O. 1998. Ecología: el puente entre ciencia y sociedad.
México: Mc GrawInteramericana, 343p.
ODUM, E.P. 2001. Fundamentos de ecologia. Fundação Calouste Gulbekian, Lisboa.
OLIVEIRA, G.C. 2005. Perfil floristico e distribuição das espécies vegetais, em relação ao
gradiente de umidade do solo, em seis veredas no triangulo mineiro. Disser tação
Universidade Federal de Uberlândia, Instituto de Biologia.

OLIVEIRA FILHO, A.T., SHEPHERD, G.J., MARTINS, F.R. & STUBBLEBINE, W.H.
1989. Environmental factors affecting physiognomic and floristic variation in an area of
cerrado in central Brazil. Journal of Tropical Ecology. v.5, p.413431.
OLIVEIRAFILHO, A. T. 1994. Estudos ecológicos da vegetação como subsídios para
programas de revegetação com espécies nativas: uma proposta metodológica. Cerne, v.1, n.1,
p. 64 72.
PANDEY, C.B. & SINGH, J.S. 1991. Influence of grazing and soil conditions on secondary
savanna vegetation in India. Journal Vegetation Science. v.2, p.95102.
PEET, R.K. 1974. The measurement of species diversity. Annual review of Ecology and
Systematis. v.5, p. 285307.
PENNING,S.C.; MOORE, D.J. 2001. Zontion of shrubs in western Atlantic salt marshes.
Oecologia. v. 126, p. 587594.
PEREIRA, B.A.S., MENDONÇA, R.C., FILGUEIRAS, T.S., PAULA, J.E. & HERINGER,
E.P. 1990. Levantamento florístico da Área de Proteção Ambiental (APA) da bacia do Rio
São Bartolomeu, DF. Anais do XXXVI Congresso Brasileiro de Botânica 1: 419492.
PINTO, M.N. (org.) 1990. Cer rado. Caracterização, ocupação e perspectivas. Editora
Universidade de Brasília UNB, Brasília, 657p.
PINTOCOELHO, R. 2000. Fundamentos em Ecologia. Porto Alegre: Artes Médicas Sul,
252p.
PINTO, L.V.A.; BOTELHO, S.A.; OLIVEIRAFILHO, A.; TEIXEIRA et al. 2005. Estudo
da vegetação como subsídios para propostas de recuperação das nascentes da bacia
hidrográfica do ribeirão Santa Cruz, Lavras, MG. Revista Árvore. v.29, n.5, p.775793.
PIRANI, J.R., GIULIETTI, A.M., MELLOSILVA, R. & MEGURO, M. 1994. Checklist and
patterns of geographic distribuition of the vegetation of Serra do Ambrósio, Minas Gerais,
Brazil. Revista Brasileira de Botânica. v.17, p.133147.

PIVELLO, V.R., BARBIERI, R.F., RUGGIERO, P.G.C. & OLIVEIRA FILHO, F. 1998.
Análise da variação fisionômica na Arie Cerrado PédeGigante (Santa Rita do Passa Quatro
SP) em relação às características pedológicas locais. In: IV Simpósio de Ecossistemas
Brasileiros. Aciesp, São Paulo, v.3, p.729.
PIVELLO, V.R., CARVALHO, V.M.C., LOPES, P.F., PECCININI, A.A. & ROSSO, S.
1999. Abundance and distribution of native and alien grasses in a “cerrado” (Brazilian
savanna) Biological Reserve. Biotropica. v.31, p.7182.
POTT, V. J. & POTT, A. 2000. Plantas Aquáticas do Pantanal (Corumbá, MS).Brasília:
Embrapa Comunicação para Transferência de Tecnologia. 404p.il.
RAMIREZ, N. & BRITO, Y. 1990. Reproductive of a tropical palm swamp community in
Venezuelan illanos. Amer ican Journal of Botany. v.77, p. 12601271.
RAMOS, M.V.V. 2000. Veredas do Triangulo Mineiro: solos, água e uso. Lavras: UFLA,
Dissertação de Mestrado.127 p.il.
RAMOS, M.V.V. 2004. Caracterização dos solos e da estrutura fitossociológica e do estado
nutricional da vegetação de veredas em diferentes superfícies geomorfológicas do Triangulo
Mineiro. Tese de Doutorado. UnB, Brasília – DF, 128p.il.
RATTER, J.A.; RIBEIRO, J.F. & BRIDGEWATER, S. 1997. The Brazilian cerrado
vegetation and threats to its biodiversity. Annals of Botany. v.80, p.223230.
REZENDE, A. V. 1998. Importância das matas de galeria: manutenção e recuperação. In:
RIBEIRO, J. F. ed. Cerrado: matas de galerias. Planaltina: EMBRAPA CPAC, 164p.
RIBEIRO, J.F.; WALTER, B.M.T. 1998. Fitofisionomias do Bioma Cerrado. In: Sano, S.M.;
Almeida, S.P. de (orgs.). Cer rado: ambiente e flora. PlanaltinaDF: Embrapa, 89166.
RIBEIRO, J.F.; FONSCECA, C.E.L.; SILVA, J.C.S. 2001. Cerrado: caracter ização e
recuperação de matas de galeria. PlanaltinaDF: Embrapa Cerrados. 899 p.il.

RICKLEFS, R.E. 1996. A Economia da Natureza. 3 ª ed. Ed. Guanabara Koogan S.A. Rio
de Janeiro. 470 p.
RIZZINI, C.T. 1979. Tratado de Fitogeografia do Brasil. v.2. Aspectos sociológicos e
florísticos. Edusp, São Paulo.
ROCHA FILHO, L.C.da; LOMÔNACO, C. 2006. Variações fenotípicas em subpopulações
de Davilla elliptica A. St.Hil. (Dilleniaceae) e Byrsonima intermedia A. Juss.
(Malpighiaceae) em uma área de transição cerradovereda. Acta Botanica Brasílica. 20 (3):
719725.
RODAL, M.J.; SAMPAIO, E.; FIGUEREDO, M. 1992. Manual sobre métodos de estudos
flor ísticos de e fitossociológico: Ecossistema Caatinga. 24p.
RODRIGUES, R. R. & SHEPHERD, G. J. 2000. Fatores condicionantes da vegetação ciliar.
Pp. 101107. In: R. R. Rodrigues & H. F. LeitãoFilho (eds.). Matas ciliares: conservação e
recuperação. EDUSP, São Paulo.
ROHLF, F. 1997. NTSYSpc: Numerical taxonomy and multivariate analysis system.
Setauket (New York): Exeter Software, 176p.
SANO, S. M.; ALMEIDA, S.P. 1998. Cer rado: ambiente e flora. Planaltina: EMBRAPA
CPAC. 556p.il..
SCHIAVINI, I. & ARAÚJO, G.M. 1989. Considerações sobre a vegetação da Reserva
Ecológica do Panga (Uberlândia). Sociedade e Natureza. n.1, p.6166.
SILVA JÚNIOR, M.C.; FELFILI, J.M. 1996. A vegetação Ecológica da Estação Ecológica
de Águas Emendadas. Instituto de Ecologia e Meio Ambiente do Distrito Federal, Brasília
DF. 43p.
SILVA, E.B.M. 2001. A família Polygalaceae na região de Catolés, Chapada Diamantina,
Bahia, Brasil. Dissertação de Mestrado. Departamento de Ciências Biológicas. 97p.il.

SILVA, M.A. da. 2005. Mundança temporal da flórula subarbustivaherbácea em
comunidades de campo sujo de cerrado submetidas a queimadas preescritas no Distrito
Federal, Brasil. Dissertação de Mestrado. Universidade de Brasília, Brasile, DF. 84 p.
SMITH, A.R.; PRYER, K.M., SCHUETTPELZ, E.; KORALL, P.; SCNEIDER, H. & WOLF,
P.G. 2006. A Classification for extant ferns. Taxon. v.55, n.3, p.705731.
SOUZA, V. C. & LORENZI, H. 2005. Botânica Sistemática: guia ilustrado para
identificação das famílias de Angiospermas da flora Brasileira, baseado no APG II. Nova
Odessa, SP: Instituto Plantarum, 640 p.il.
TIVING, J.G.; NIENHIUS, J.;SMITH, O.S. 1994. Estimation of sampling variance of
molecular marker data using the bootstrap procedure. Theorical Apllied Genetics. Berlin,
v.89, n.2/3, p.259264.
TANNUS, J. L.S.; ASSIS, M.A. 2004. Composição de espécies vasculares de campo sujo e
campo úmido em área de cerrado, Itirapina – SP, Brasil. Revista Brasileira Botânica. v.27,
n.3, p.489506.
TOLEDO, M.B. de; BUSH, M.B. 2008. Vegetation and hydrology changes in Eastern
Amazonia inferred from a pollen record. Anais da Academia Brasileira de Ciências
[online]. v. 80, n.1, p.191203.
TOSCANO DE BRITO, A.L V. & CRIBB, PHILLIP. 2005. Orquídeas da Chapada
Diamantina. Nova Fronteira. 399.p.il.
UHLMANN, A., GALVÃO, F. & SILVA, S.M. 1998. Análise da estrutura de duas unidades
fitofisionômicas de savana (Cerrado) no Sul do Brasil. Acta Botanica Brasílica. n.12, p.231
247.
UHLMANN, A. 2003. Análise estrutural de duas áreas de vegetação savânica (Cerrado) sob
influência de gradientes ambientais complexos. Tese de Doutorado, Universidade Estadual
de Campinas, Campinas SP.

WALTER, B.M.T. 2006. Fisionomias do bioma cerrado: síntese terminológica e relações
florísticas. Tese de Doutorado. Departamento de Ecologia do Instituto de Ciências
Biológicas da Universidade de Brasília – UnB, Brasília DF.
WILSON, S. D.; KEDDY, P.A. Plant zonation on a shoreline gradient: physiological response
curves of component species. J ournal of Ecology. v. 73, p. 851860.
ZOCCHE, J.J. & PORTO, M.L. 1992. Florística e fitossociologia de campo natural sobre
banco de carvão em áreas mineradas, Rio Grande do Sul, Brasil. Acta Botanica Brasílica.
n.6, p.4784.


















