
BIOLOGIA DE AVES CAPTURADAS EM UM FRAGMENTO DE MATA ...€¦ · 4.3. Bionomia e Biometria ......
82
UNIVERSIDADE FEDERAL DE PERNAMBUCO CENTRO DE CIÊNCIAS BIOLÓGICAS DEPARTAMENTO DE ZOOLOGIA MESTRADO EM BIOLOGIA ANIMAL BIOLOGIA DE AVES CAPTURADAS EM UM FRAGMENTO DE MATA ATLÂNTICA, IGARASSU – PE VIVYANNE SANTIAGO MAGALHÃES RECIFE 2005
Transcript of BIOLOGIA DE AVES CAPTURADAS EM UM FRAGMENTO DE MATA ...€¦ · 4.3. Bionomia e Biometria ......

i
UNIVERSIDADE FEDERAL DE PERNAMBUCO
CENTRO DE CIÊNCIAS BIOLÓGICAS
DEPARTAMENTO DE ZOOLOGIA
MESTRADO EM BIOLOGIA ANIMAL
BIOLOGIA DE AVES CAPTURADAS EM UM FRAGMENTO DE MATA
ATLÂNTICA, IGARASSU – PE
VIVYANNE SANTIAGO MAGALHÃES
RECIFE
2005

ii
UNIVERSIDADE FEDERAL DE PERNAMBUCO
CENTRO DE CIÊNCIAS BIOLÓGICAS
DEPARTAMENTO DE ZOOLOGIA
MESTRADO EM BIOLOGIA ANIMAL
BIOLOGIA DE AVES CAPTURADAS EM UM FRAGMENTO DE MATA
ATLÂNTICA, IGARASSU – PE
VIVYANNE SANTIAGO MAGALHÃES
RECIFE
2005

iii
VIVYANNE SANTIAGO MAGALHÃES
BIOLOGIA DE AVES CAPTURADAS EM UM FRAGMENTO DE MATA
ATLÂNTICA, IGARASSU – PE
Dissertação apresentada ao Mestrado em Biologia
Animal da Universidade Federal de Pernambuco,
como parte dos requisitos para a obtenção do grau de
Mestre em Ciências Biológicas na área de Biologia
Animal.
Orientador: Dr. Severino Mendes de Azevedo Júnior
RECIFE
2005

iv

v

vi
“É muito melhor arriscar coisas grandiosas, alcançar
triunfo e glória, mesmo expondo-se a derrota, do que
formar fila com os pobres de espírito que nem gozam
muito nem sofrem muito, porque vivem nessa
penumbra cinzenta que não conhece vitória nem
derrota”.
Theodore Roosevelt

vii
SUMÁRIO
DEDICATÓRIA............................................................................................................... i
AGRADECIMENTOS....................................................................................................... ii
LISTA DE FIGURAS........................................................................................................ v
LISTA DE TABELAS........................................................................................................ vii
RESUMO........................................................................................................................ viii
ABSTRACT.................................................................................................................... ix
1. INTRODUÇÃO....................................................................................................... 1
2. OBJETIVOS............................................................................................................ 5
2.1. Objetivo Geral......................................................................................... 5
2.2. Objetivos Específicos.............................................................................. 5
3. MATERIAL E MÉTODOS..................................................................................... 6
3.1. Área de estudo......................................................................................... 6
3.2. Métodos................................................................................................... 9
3.3. Tratamento Estatístico............................................................................. 13
4. RESULTADOS....................................................................................................... 15
4.1. Biologia das Espécies.............................................................................. 15
4.2. Temperatura e Índices Pluviométricos.................................................... 16
4.3. Bionomia e Biometria.............................................................................. 18
5. DISCUSSÃO........................................................................................................... 33
6. CONSIDERAÇÕES FINAIS.................................................................................. 43
7. REFERÊNCIAS BIBLIOGRÁFICAS.................................................................... 45
8. APÊNDICES........................................................................................................... 53

i
Aos meus avozinhos, Dona Leó e Seu Gigi, in
memorian, por terem construído o amor e
companheirismo através de uma família tão
maravilhosa e deixado valores que nos guiarão
como seres viventes conscientes e destinados
ao bem. Por terem sonhado com esse momento.
Que estejam vendo o resultado do que
plantaram!

ii
Agradecimentos
Hoje agradeço, sobretudo, a Deus, pela minha existência e por toda a confiança que Ele
depositou em mim, mostrando que cada minuto trilhado da vida, por mais difícil que seja
(e foram anos difíceis), vale a pena!
A minha mãezinha querida, Terezinha, que com muito carinho esteve eternamente
presente e atenta às minhas necessidades, principalmente nas noites solitárias em que me
encontrava acordada escrevendo, com seus lanchinhos providenciais. Fez tudo isso de
coração aberto, muitas vezes sequer imaginando a dimensão dos meus projetos de vida.
Ao meu pai, Mariceu, que este ano de 2004 vivenciou tantas mudanças em sua vida, mas
mesmo assim não se rendeu às adversidades, estando pronto aos meus chamados sem
pestanejar e providenciou – orgulhoso – toda a logística para facilitar a escrita da minha
dissertação.
Aos meus irmãos, Polyanna e “Manoel”, que passam atualmente pelo mesmo momento
“literário” que eu, mas que, ainda assim, encontraram a palavra certa para me ajudar e
depositar confiança e estímulo a continuar, desde o campo, do qual sem eles esta
empreitada não teria sido concluída.
A Maria José e a Rose e os meninos, Camila e Tiago, que ajudaram da forma que
puderam e trouxeram momentos de descontração tão necessários nessa fase.
A família como um todo, pela paciência e compreensão da minha ausência (para alguns,
uma ausência aliviada – rs). Foi apenas uma etapa minha gente. Estou de volta!!!
Ao orientador, Severino Mendes de Azevedo Júnior, por aceitar ceder seu nome para
que eu pudesse seguir adiante com meu projeto. Agradeço as oportunidades profissionais
que me ofereceu ao longo desses anos.
A CAPES pelo apoio financeiro concedido durante parte do período do mestrado.

iii
A Michele e Roberto Siqueira, que permitiram a realização desta pesquisa no RECD e
ajudaram no que puderam, tornando possível este desafio; não esquecendo àqueles que
fazem parte íntima do Refúgio: Daniel, Henrique, Branco e Manoel.
Aos queridos, Rachel e Wallace, que começaram tudo isso, através de um convite que
inicialmente pensei que seria apenas um sonho. Este sonho está em fase de conclusão,
meninos. Obrigada!
Àqueles que participaram desse processo nos bastidores: Ana Elizabeth, da Secretaria do
Mestrado; Mário, experiente taxidermista e “professor de campo” do Laboratório de
Ornitologia da UFPE com quem aprendi bastante; Ana Katarina da biblioteca da
UFRPE, sempre disposta a ajudar “para ontem” e Gustavo da geologia da UFPE, com
seu profissionalismo e simpatia atuantes.
Aos professores da UFPE que se dispuseram e auxiliaram este trabalho com grande
empenho, cada um dentro da sua especialidade: José Roberto Botelho, Miriam Camargo
Guarnieri, Maria Eduarda de Larrazábal, Sigrid Leitão e Waldir Luna.
A Marina Anciães, que se mostrou sempre receptiva, mesmo virtualmente.
A Daniela Pedrosa, vulgo Dani Bananinha, por todo o apoio desde o campo ao trabalho
braçal no computador e pelo grande carinho que construímos durante esse período de
convivência intensa. Uma pessoa que nunca me faltou quando mais precisei. Obrigada!
Aos amigos: Carmem, Samanta, Luiza, Emerson, Weber, Mércia, Cláudio Cazal, Sheila
(Popoca), Viviane, Tininha, Milena, Arnaldo, juntamente com todos do Laboratório de
Répteis da UFPE, pela força.
A Karina Linhares: abro um espaço especial a você amiga que muito me ensinou em tão
pouco tempo e sempre esteve por perto. Que bom essa disciplina ter existido para nos
aproximar ainda mais!
A Marco Antônio, eita, vixe, oxe, Marco Antônio... Por todo carinho, atenção, paciência
(sobretudo), ajuda, leitura crítica, apoio moral, madrugadas virtuais de estudo,
momentos de relaxamento e, principalmente, pela confiança ilimitada na minha
capacidade e no meu trabalho, qualquer que seja ele. E ainda, agradeça por mim aos

iv
amigos em comum que fizemos ou adquirimos um do outro nessa sua terrinha e que
também desejaram o melhor. Valeu!
A ADAGRO, nas pessoas de Dr. Jair Virgínio e Fernando Góes de Miranda, que
compreenderam esse momento importante da minha carreira profissional e deram
oportunidade para que eu concluísse com tranqüilidade essa etapa, me resguardando das
demais tarefas que me competiam. Sem essa ajuda, talvez eu não tivesse conseguido.
Virgínia, Edjailson e Tereza pela força, compreensão e cuidados sempre presentes e aos
demais companheiros da ADAGRO, pela torcida incondicional.
A Maria Luiza Santos, pelos dias (principalmente seus sábados) que dedicou em me
ajudar com a parte estatística, até tarde da noite, com olhos vermelhos de cansaço, mas
sem pestanejar no apoio irrestrito. Uma nova amizade construída! E a Rodrigo, pelo bom
humor sempre presente e pelo empréstimo do quarto e do computador.
Ao professor Edmilson Mazza que nessa reta final foi fundamental para a conclusão do
meu estudo, trabalhando em tempo recorde em pleno carnaval.
A todos que porventura não citei, mas que ajudaram de alguma forma.
Ao Gordo, Sérgio eu agradeço... Ele sabe por quê! E aos seus pais, Cristina Alcantara e
José Elizeu e Silva, que sempre estiveram presentes, ajudando e torcendo, sobretudo.
Meu muito obrigada!
E a minha eterna Kelly, em nome de todos os animais.

v
LISTA DE FIGURAS
Figura 1. Planta Diretora de Igarassu (Escala 1:760.000). A. Localização do
município de Igarassu. B. Posição geográfica do Refúgio Ecológico
Charles Darwin (RECD). C. Localização georeferenciada das
redes. Fonte: SINTAXE Consultoria.................................................. 6
Figura 2. Aspectos da vegetação a partir de campo aberto. Em destaque,
notar proporção da altura da mata em relação à estatura de um
adulto. Foto da autora......................................................................... 8
Figura 3. Medição do Turdus leucomelas (Sabiá-barranco): A. Medição da
asa; B. Medição do diâmetro do tarso, no terço distal; C.
Comprimento do tarso. Fotos: Marco A. B. de Almeida.................... 11
Figura 4. Placas de Incubação: A. Placa em estágio 2, entrando na fase de
postura; B. Placa em estágio 3, correspondente ao período de
incubação dos ovos propriamente dito (IBAMA 1994). Fotos da
autora.................................................................................................. 12
Figura 5. Índice de Precipitação Pluviométrica, distribuído ao longo dos
meses de coleta................................................................................... 16
Figura 6. Temperatura média distribuída ao longo dos meses de coleta........... 17
Figura 7. Freqüência da presença e ausência de Placa de Incubação ao longo
dos meses de estudo............................................................................ 24
Figura 8. Freqüência de placa e muda simultâneos (sobreposição) por tipo,
segundo o mês de ocorrência.............................................................. 26
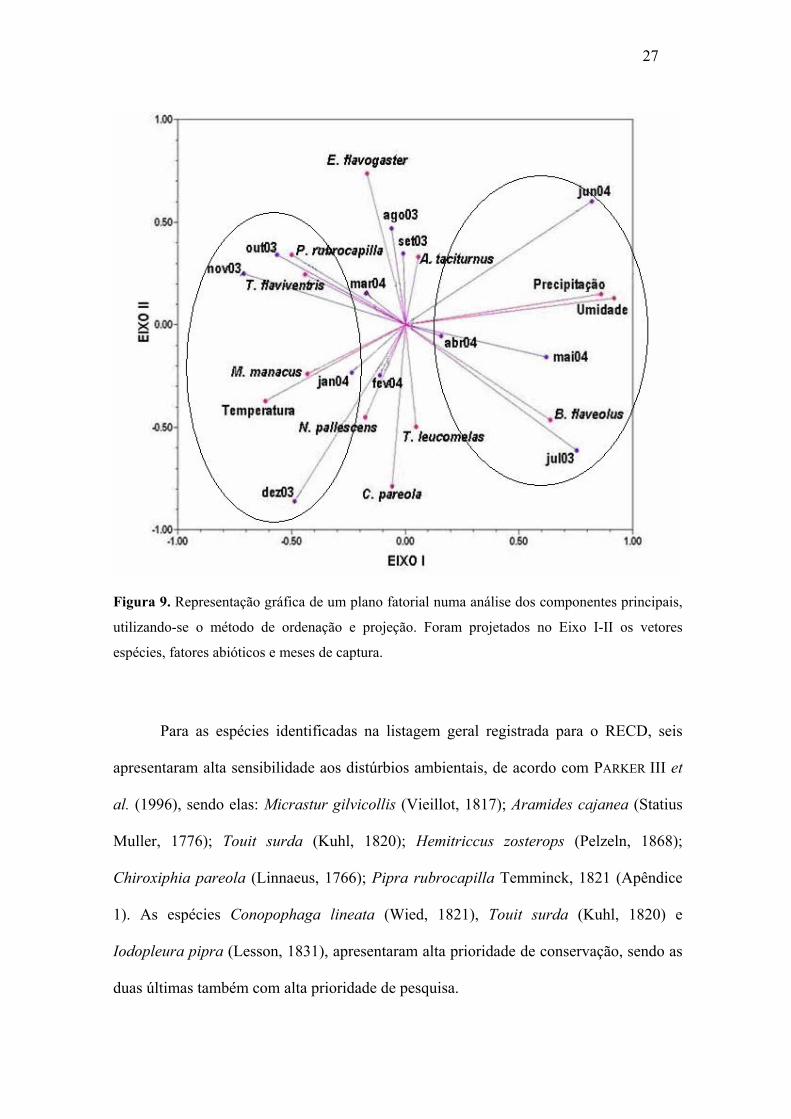
Figura 9. Representação gráfica de um plano fatorial numa análise dos
componentes principais, utilizando-se o método de ordenação e
projeção. Foram projetados no Eixo I-II os vetores espécies, fatores
abióticos e meses de captura............................................................... 27
Figura 10. Espécies capturadas no RECD entre julho de 2003 e junho de 2004.
A. Conopophaga lineata, B. Xenops minutus, C. Touit surda, D.
Ornithion inerme. Fotos da autora...................................................... 28

vi
Figura 11. A. Ninho ocupado em dezembro de 2003 pelo Trogon curucui
(Surucuá-de-barriga-vermelha). B. Ninho destruído em fevereiro de
2004, com ovo em seu interior. C. Detalhe dos outros ovos
destruídos fora do ninho. Fotos da autora........................................... 31

vii
LISTA DE TABELAS
Tabela I. Tempo de anilha das espécies recuperadas no RECD entre 1996/1997 e
2003/2004. A = Adulto, J = Jovem, M = Macho, F = Fêmea, I =
Indeterminado, Id = Idade, Sx = Sexo, TA = Tempo de Anilha.................. 15
Tabela II. Freqüência de ocorrência, médias e desvio padrão das massas corpóreas
registradas para as espécies de aves capturadas no RECD entre julho de
2003 e junho de 2004. Ordem de evolução de acordo com SICK (1997).
FO = freqüência de ocorrência, <MC(g) = menor massa corporal
encontrada, >MC(g) = maior massa corporal encontrada, dp = desvio
padrão e (n) = nº de indivíduos analisados quanto ao peso.......................... 19
Tabela III. Média e desvio padrão da massa corpórea de três espécies de aves
segundo o sexo dos animais. (n) = número de indivíduos analisados por
espécie e sexo, dp = desvio padrão............................................................... 20
Tabela IV. Média e desvio padrão da massa corpórea de três espécies de aves
segundo a estação seca e chuvosa. (n) = número de indivíduos analisados
por espécie e sexo, dp = desvio padrão........................................................ 21
Tabela V. Média e desvio padrão das medidas: asa, tarso e diâmetro do tarso de três
espécies de aves segundo o sexo dos animais. (n) = número de indivíduos
analisados por espécie e sexo, dp = desvio padrão, Ø do tarso = diâmetro
do tarso......................................................................................................... 22
Tabela VI. Avaliação da ocorrência de cada tipo de muda segundo a estação da
captura.......................................................................................................... 23
Tabela VII. Avaliação da ocorrência de cada tipo de muda segundo a ocorrência de
placa de incubação........................................................................................ 25
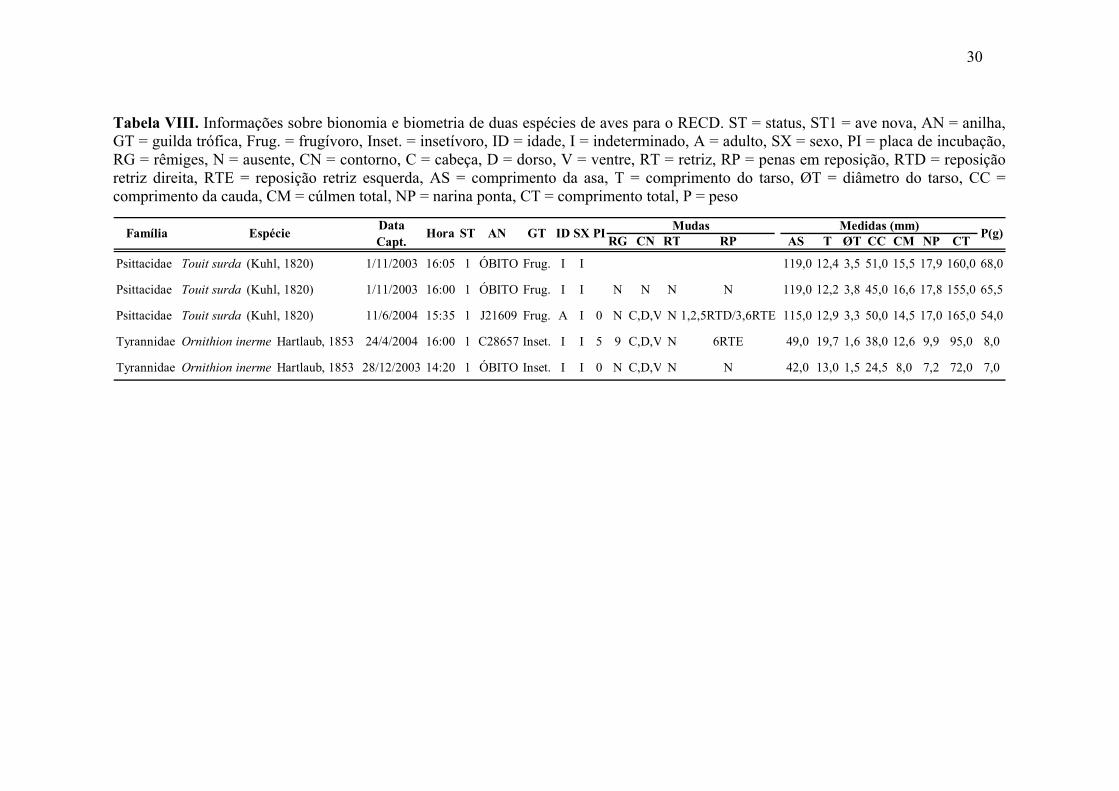
Tabela VIII. Informações sobre bionomia e biometria de duas espécies de aves para o
RECD. ST = status, ST1 = ave nova, AN = anilha, GT = guilda trófica,
Frug. = frugívoro, Inset. = insetívoro, ID = idade, I = indeterminado, A =
adulto, SX = sexo, PI = placa de incubação, RG = rêmiges, N = ausente,
CN = contorno, C = cabeça, D = dorso, V = ventre, RT = retriz, RP =
penas em reposição, RTD = reposição retriz direita, RTE = reposição
retriz esquerda, AS = comprimento da asa, T = comprimento do tarso, ØT
= diâmetro do tarso, CC = comprimento da cauda, CM = cúlmen total,
NP = narina ponta, CT = comprimento total, P = peso................................ 30

viii
RESUMO
Este trabalho estudou a biologia da avifauna do Refúgio Ecológico Charles
Darwin, fragmento de 60 ha de Mata Atlântica, no município de Igarassu, Pernambuco.
Objetivando obter informações acerca das espécies desse bioma, foram realizadas
observações entre agosto de 1996 e julho de 1997 e capturas mensais utilizando redes de
neblina, de julho de 2003 a junho de 2004. Entre observações, capturas, recapturas e
recuperações, foram registradas 151 espécies (31 famílias) para a área, onde 456 aves de
53 espécies e 25 famílias foram capturadas com redes ornitológicas. Foram recuperadas
10 espécies (tempo de anilha de seis a oito anos). O número de capturas foi maior nos
meses mais quentes. A maioria das espécies capturadas (52,8%) teve freqüência de
ocorrência menor que 25%, sendo Manacus manacus (Linnaeus, 1766), Arremon
taciturnus (Hermann, 1783), Neopelma pallescens (Lafresnaye, 1853) e Turdus
leucomelas Vieillot 1818 as mais freqüentes. Houve correlação significativa entre
médias de massa corpórea e comprimento da asa e tarso e diâmetro do tarso relacionado
ao sexo do indivíduo, e de mudas com estação do ano. O maior período com muda
associada à placa de incubação foi de março a maio (pico em maio). Os achados
fortalecem a imprevisibilidade dos efeitos das alterações ambientais na estrutura da
comunidade de aves em longo prazo. Mantém-se afirmativo que os desequilíbrios
populacionais possam vir a aumentar as chances de extinção, sendo necessárias novas
alternativas para a proteção da biodiversidade, sobretudo em fragmentos florestais.
PALAVRAS CHAVE. Avifauna, ecologia, fragmentação.

ix
ABSTRACT
Biology of birds captured in an Atlantic Forest fragment, at Igarassu, Pernambuco
A study was carried out, about biology of the avifauna of the Refúgio Ecológico
Charles Darwin, a 60 ha fragment of Atlantic Forest, in the town of Igarassu,
Pernambuco. Planning obtain information about the species of that bioma, were carried
out observations between August – 1996 and July – 1997 and monthly captures utilizing
mist nets, between July – 2003 and June – 2004. Among observations, captures,
recaptures and recovers, 151 species (31 families) were registered in the study area, and
456 birds (from 53 species and 25 families) were trapped in the nets. Ten species had
been recovered (time of banding from six to eight years). The amount of captures it was
bigger in hottest months. The majority of captured species (52,8%) had frequency of
occurrence less than 25%. Manacus manacus (Linnaeus, 1766), Arremon taciturnus
(Hermann, 1783), Neopelma pallescens (Lafresnaye, 1853) and Turdus leucomelas
Vieillot 1818 was the most frequent. It had significant correlation between averages of
corporal mass and length of the wing and tarsus and diameter of tarsus related with sex,
and of molt with seasons. The main period with molt associated to the brood patches was
from March to May (with a peak in May). The findings fortify the unpredictability of the
effects of alterations in the structure of bird community in long stated period. It is
remained affirmative that the population unbalance can come to increase the
extinguishing possibilities, being necessary new alternatives for the protection of
biodiversity, over all in forest fragments. KEY WORDS. Bird community, ecology,
fragmentation.

1
1. INTRODUÇÃO
A rapidez com que os ambientes naturais estão sendo destruídos não tem
precedentes na evolução histórica da floresta tropical úmida e certamente pode causar
profundos efeitos sobre o ecossistema (BIERREGAARD et al. 1992). Neste contexto, a
Mata Atlântica é considerada um dos ecossistemas florestais mais ameaçados e
devastados pelo homem, um dos “hotspots” da biodiversidade, com seus remanescentes
florestais sofrendo ainda forte pressão antrópica (FONSECA 1985; RANTA et al. 1998,
MYERS et al. 2000), restringindo-se, atualmente, a apenas 7,3% de sua cobertura original
(IBAMA 2003).
A região Nordeste possuía 36,8% de sua área coberta com este ecossistema no
início do século XX e o Estado de Pernambuco 34,14% no mesmo período (GONZAGA
DE CAMPOS 1912). Atualmente mantém apenas 2% da sua cobertura original, em
fragmentos rodeados por monoculturas canavieiras e aglomerados urbanos (VIANA et al.
1997; CHIARELLO 1999; SILVA & TABARELLI 2000). Sendo assim, o que resta de Mata
Atlântica em Pernambuco está distribuído principalmente em pequenos fragmentos, uma
vez que aproximadamente 48% dos trechos restantes possuem menos de 10 hectares e
apenas 7% destas “ilhas” são maiores que 100 ha com o tamanho médio de 34 ha por
fragmento (RANTA et al. 1998).
Apesar de tão explorada, pouco se conhece da Floresta Atlântica, mas sabe-se
que este complexo ecossistema contém uma diversidade de espécies maior do que a
Floresta Amazônica e é caracterizada por altos níveis de endemismo entre plantas e
animais (FONSECA 1985; BROWN & BROWN 1992, TERBORGH 1992).

2
A destruição da floresta tem resultado na eliminação de muitas populações e
potencialmente, na erosão da diversidade genética de várias espécies (BROWN & BROWN
1992, MORELLATO & HADDAD 2000). Inicia-se assim um processo contínuo de extinção
local, onde a composição e diversidade biológica da comunidade passam por estágios de
transição até a fase de equilíbrio (TABARELLI 2000). WILLIS (1979) e BIERREGAARD et
al. (1992) estão de acordo com TABARELLI (2000) quando relatam a diminuição da área
florestal associada a um decréscimo no número de espécies de aves. Por isso entende-se
que a presença ou ausência de certas espécies em áreas estudadas da Mata Atlântica ou
de outros ecossistemas, determinam a situação ecológica de ecossistemas específicos
(TELINO JÚNIOR et al. 2000).
MARINI (1998), em estudos acerca dos efeitos da fragmentação florestal sobre as
aves, relata a importância das pequenas áreas fragmentadas de Mata Atlântica, bem
como a necessidade de preservação e desenvolvimento de novas pesquisas nestes
refúgios.
Para LAURANCE (1991) e TABARELLI (2000), a fragmentação apresenta efeitos
em longo prazo sobre as populações através de mudanças nos principais processos
ecológicos como a polinização, predação, comportamento territorial e hábitos
alimentares. Esses efeitos podem surgir como resultado indireto de conseqüências
microclimáticas da fragmentação, como mudança na radiação solar, umidade e padrões
de vento, que são importantes para muitos organismos.
Toda essa fragmentação aumenta a proporção das áreas de borda nesta região,
que se apresenta fisicamente diferente do interior do habitat, estando inversamente
relacionado ao tamanho do fragmento, o que resulta em falha por estes fragmentos em
suportar espécies geralmente encontradas em ecossistemas intactos (BIERREGAARD et al.

3
1992; RANTA et al. 1998). Para a avifauna, a fragmentação florestal geralmente atua
favorecendo as aves de ambientes abertos e de borda, atuando negativamente, na maior
parte dos casos, sobre espécies florestais (WILLIS 1979; BIERREGAARD et al. 1992;
ALEIXO & VIELLIARD 1995).
Os distúrbios antropogênicos no hábitat natural, aliados aos fatores supracitados,
levam à necessidade de acompanhamento do tamanho e qualidade das populações
silvestres, o que pode ser feito através do desenvolvimento de métodos simples,
aplicadas como índice para monitoramento do grau de conservação de populações de
aves em fragmentos da Mata Atlântica (ANCIÃES & MARINI 2000 a e b).
Segundo AZEVEDO-JÚNIOR et al. (2001), diversos estudos já demonstraram a
importância da marcação de aves, principalmente no que concerne a dados sobre a
biologia, longevidade, deslocamentos e migrações. SICK (1997) afirma que o
anilhamento é útil, se não indispensável para estudo de populações locais, incluindo
aquelas que não migram, trazendo informações importantes como subsídios das atitudes
e políticas de conservação, empreendidas pelo órgão federal responsável (Instituto
Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis - IBAMA), bem
como pela difusão do uso da técnica de anilhamento em aspectos de pesquisa básica ou
aplicada (IBAMA 1994).
Embora as aves, de um modo geral, já se apresentem como um grupo
amplamente estudado, informações relacionadas à bionomia e biometria são escassas
(CLARK JÚNIOR 1979, FOSTER 1975, REINERT et al. 1996). Estudos prolongados sobre
mudas de aves neotropicais são raros (MARINI & DURÃES 2001). Aqueles autores que
trazem informações sobre massa corpórea, em geral apresentam informações pontuais,
bem como aqueles que fazem inferência acerca de dados biométricos (BALDWIN &

4
KENDEIGH 1938, MCNEIL 1971, CLARK JÚNIOR 1979, ONIKI 1981, ONIKI & WILLIS 1993,
REINERT et al. 1996).
A despeito disto, poucas são as pesquisas realizadas que mostram como as
espécies respondem ao atual estado de fragmentação de seus habitats na região
nordestina. Alguns trabalhos ornitológicos com anilhamento e biologia das espécies
foram desenvolvidos entre os anos de 1996 e 1997, na mesma área onde foi realizado o
presente trabalho (LYRA-NEVES et al. 2000; TELINO-JÚNIOR et al. 2000).
Desta forma, objetiva-se apresentar novas informações acerca da biologia das
espécies de aves em uma área fragmentada de Mata Atlântica secundária no nordeste do
Brasil, através de informações obtidas por observação, captura, recaptura e recuperação,
visando à obtenção de dados que possam ser utilizados em estratégias de conservação da
avifauna.

5
2. OBJETIVO
2.1. OBJETIVO GERAL
Esta pesquisa teve como objetivo apresentar novos conhecimentos acerca da
biologia das espécies de aves em um fragmento de Mata Atlântica, que possam ser
utilizadas para subsidiar estratégias de conservação da avifauna.
2.2. OBJETIVOS ESPECÍFICOS
► Gerar informações sobre a biologia das espécies de aves capturadas em um fragmento
de Mata Atlântica, pela avaliação da idade, sexo, tempo de anilha, período reprodutivo e
mudas;
► Inferir se há ou não diferenças e associações significativas entre fatores bióticos e
abióticos encontrados.
6
3. MATERIAL E MÉTODOS
3.1. ÁREA DE ESTUDO
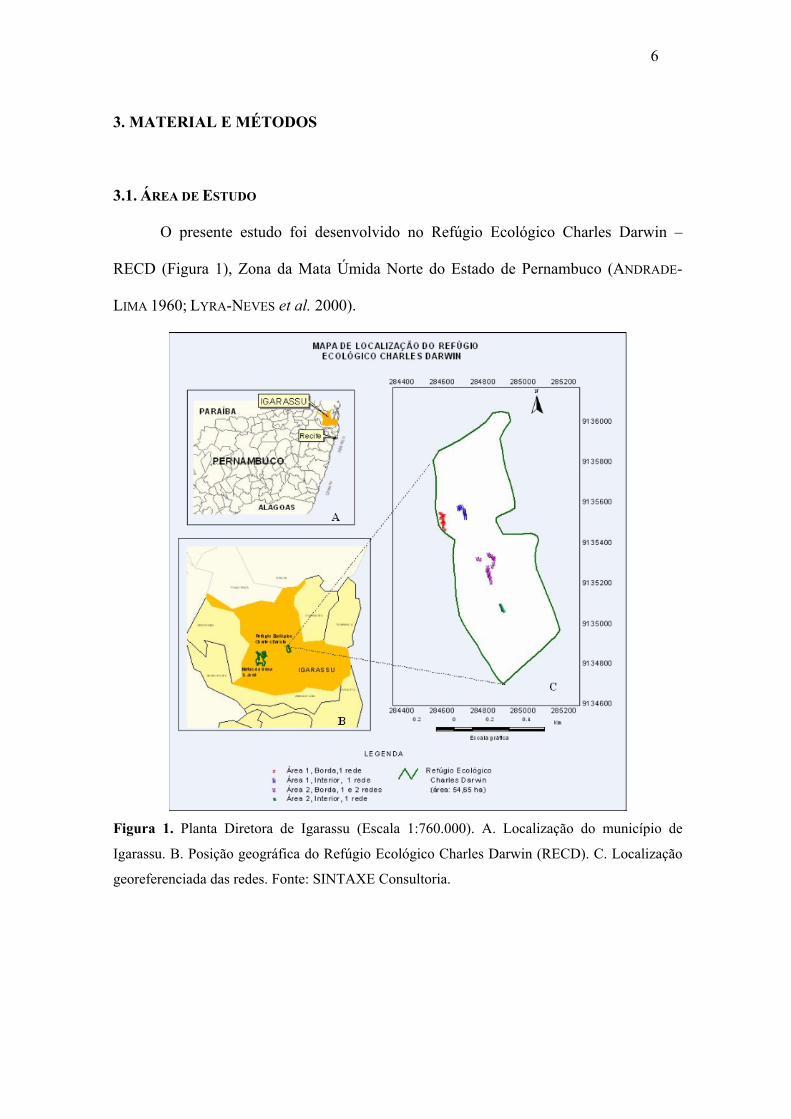
O presente estudo foi desenvolvido no Refúgio Ecológico Charles Darwin –
RECD (Figura 1), Zona da Mata Úmida Norte do Estado de Pernambuco (ANDRADE-
LIMA 1960; LYRA-NEVES et al. 2000).
Figura 1. Planta Diretora de Igarassu (Escala 1:760.000). A. Localização do município de
Igarassu. B. Posição geográfica do Refúgio Ecológico Charles Darwin (RECD). C. Localização
georeferenciada das redes. Fonte: SINTAXE Consultoria.

7
O RECD é um fragmento florestal de Mata Atlântica localizado no Município de
Igarassu entre as coordenadas 7º48’ e 7º49’ Sul e 34º56’ e 34º57’ Oeste, distando 34 km
da capital pernambucana, fazendo parte do Complexo Litorâneo da Reserva da Biosfera
da Mata Atlântica. Propriedade de domínio privado ocupa uma área de
aproximadamente 60 ha, totalmente ilhada por monocultura de capim-elefante
(Pennisetum purpureum Schum.). Criado há cerca de 40 anos, somente no final da
década de 80 teve início à sistematização de suas atividades como Centro de Pesquisas
(LIMA 1998).
A floresta que compõe este fragmento sofreu ação antrópica no passado, sendo
atualmente representada por um remanescente de Mata Atlântica com formação vegetal
secundária em fase de recomposição, embora as áreas vizinhas já estejam desmatadas
servindo à agropecuária. Conforme descrito em trabalhos anteriores (FIDEM 1993),
nesta zona da mata a vegetação encontra-se dividida em capoeirão, capoeira rala,
vegetação arbustiva e campos, assim descritos: capoeirão, de formação secundária,
originária da Floresta Atlântica primitiva, apresenta vegetação arbórea, onde predomina
espécies de porte médio e alto, demonstrando recomposição; a capoeira rala é
semelhante ao capoeirão, porém, de porte menor e com uma maior densidade de
espécies; a vegetação arbustiva é constituída por uma faixa de transição, com extrato
arbóreo inferior a três metros de altura, composta por arbustos finos e gramíneas; nos
campos predominam as gramíneas, com diversas espécies arbóreas (frutíferas em sua
maioria, a exemplo de: sapotizeiro [Achras sapota L.], pitangueira [Eugenia uniflora L.]
e cajueiro [Anacardium ocidentalis L.]), sendo distribuídas espaçadamente ao longo da
área. A borda da mata é composta predominantemente por vegetação arbustiva e o seu
interior composto principalmente por capoeirão (Figura 2).

8
ANDRADE-LIMA (1961) relata que causas diversas condicionam os tipos de
floresta, sendo a mais importante delas o clima. O autor afirma que em Pernambuco
ocorre significativa variação de condições climáticas, sendo a distribuição das chuvas
desiguais em todas as áreas do Estado. Nos municípios mais próximos da costa há
chuvas entre abril e agosto, podendo prolongar-se até setembro, em um total anual
variando de 700 a 1.300 mm/ano nos municípios mais ao norte e de 1.500 a 2.300
mm/ano naqueles municípios costeiros e do sul do Estado, em Recife chegando a 1.700
mm/ano (RANTA et al. 1998), com os períodos de estiagem descritos por AZEVEDO-
JÚNIOR (1990) ocorrendo entre os meses de setembro a março.
Figura 2. Aspectos da vegetação a partir de campo aberto. Em destaque, notar proporção da
altura da mata em relação à estatura de um adulto. Foto da autora.
O clima de Igarassu é considerado quente e úmido (LYRA-NEVES 1998). Quanto
à hidrografia, o RECD é cortado pelo Rio Tabatinga, também chamado de Jacoca, cujo

9
leito é perene e possui uma largura variando de dois a oito metros e sua profundidade
entre 30 centímetros a dois metros (LYRA-NEVES op cit.).
3.2. MÉTODOS
No RECD, dois ambientes foram escolhidos, sendo denominados Área I (AI) e
Área II (AII). Ambos tiveram redes posicionadas na borda e no interior da mata. Cada
área foi amostrada mensalmente durante um período de três dias, entre julho de 2003 e
junho de 2004. A AII não foi amostrada no mês de novembro, sendo as duas expedições
realizadas na AI. Para algumas análises, este mês foi removido ou, quando possível, a
média de espécies capturadas naquele mês foi utilizada para os cálculos.
O registro das espécies presentes no RECD (Apêndice 1) ocorreu entre os
períodos de agosto de 1996 a julho de 1997 (apenas por meio de observação visual) e de
julho de 2003 a junho de 2004 através de um esforço padronizado de captura pelo uso de
redes de neblina (mist-net), em número médio de 12, com 36 mm de malha, 12 m de
comprimento e aproximadamente 3 m de altura. As redes foram dispostas em
transecções lineares no sub-bosque, com localização georeferenciada para as redes
colocadas entre 2003 e 2004, que foram abertas entre 5:00h e 10:00h e fechadas no
intervalo mais quente do dia, quando as aves estão em descanso. As mesmas foram
reabertas das 15:00h às 17:00h, quando retornam a atividade mais intensa antes do
período de repouso. As redes foram vistoriadas a cada hora.
Todos os dados referentes à biometria e bionomia (idade, sexo, mudas, placa de
incubação, guildas alimentares, recapturas e recuperações) foram anotados em planilha
de campo (Apêndice 2) para cada indivíduo capturado.

10
Para fins desse trabalho, entende-se por captura toda ave marcada pela primeira
vez. Por recaptura as aves que são marcadas e capturadas novamente pelo mesmo
pesquisador, naquela localidade. E recuperação, quando são capturadas aves marcadas
por outros anilhadores.
Dados de temperatura e pluviometria para o período de 2003 e 2004 foram
obtidos junto ao Instituto Nacional de Meteorologia (INMET), pela Seção de
Observação e Meteorologia Aplicada (SEOMA).
Todos os indivíduos foram identificados ao nível específico, de acordo com
DUNNING (1987), RIDGELY & TUDOR (1994 a-b), SICK (1997) e SOUZA (2002). A
nomenclatura científica e os nomes vulgares utilizados são aqueles propostos por SICK
(1997). As aves capturadas receberam anilhas metálicas fornecidas pelo Centro Nacional
de Pesquisa para a Conservação de Aves Silvestres do Instituto Brasileiro do Meio
Ambiente e dos Recursos Naturais Renováveis (CEMAVE/IBAMA) seguindo as
orientações contidas no manual de anilhamento deste centro (IBAMA 1994).
Para os dados biométricos, todos os indivíduos capturados foram mensurados,
utilizando-se: paquímetro com precisão de 0,02 mm e régua metálica milimetrada, onde
foram aferidos comprimento das asas e tarsos e diâmetro dos tarsos. As asas foram
medidas pelas penas primárias em sua extensão plena, mantidas o mais retilínea
possível. A décima primária não foi considerada na medição por ser rudimentar em
alguns grupos (SICK 1997). Os tarsos foram mensurados a partir da articulação tíbio-
tarso até o final da última escama completa do tarso e o seu diâmetro medido
longitudinalmente na porção mais distal do tarso, próximo à articulação tarso-metatarso
(Figura 3). Aves em muda de rêmiges primárias ou com anomalias nos tarsos não foram

11
medidas. A massa corpórea foi obtida através de balanças Pesola tipo dinamômetro
com precisão de 1 e 2g.
Figura 3. Medição do Turdus leucomelas (Sabiá-barranco): A. Medição da asa; B. Medição do
diâmetro do tarso, no terço distal; C. Comprimento do tarso. Fotos: Marco A. B. de Almeida.
A determinação da idade e sexo, quando possível, foi baseada na plumagem.
Quanto às mudas, foram observadas: penas de contorno (penas que recobrem o
corpo), penas da asa ou rêmiges e penas da cauda ou retrizes. Foi estabelecida a presença
de muda de contorno quando havia penas em crescimento (canhão) na região da cabeça,
dorso e ventre, num número mínimo de três penas em quaisquer dessas regiões. Nas
penas de vôo da asa, apenas as mudas das rêmiges primárias foram observadas, critério
amplamente utilizado nos trópicos (MILLER 1961), aconselhado por este fenômeno
ocorrer em um curto espaço de tempo, substituindo praticamente toda a plumagem no
período de muda (AVERY 1985). Em acordo com o trabalho de MARINI & DURÃES
(2001), nesta pesquisa as rêmiges e retrizes foram consideradas em muda somente
12
quando apresentavam reposição simétrica e para esta análise foram excluídas as aves
imaturas ou àquelas com algumas penas perdidas acidentalmente durante a captura e
processamento. Para a análise das mudas, àquelas aves recapturadas ou recuperadas em
diferentes meses foram tratadas como novos registros, mas somente no primeiro registro
individual do mês amostrado (MARINI & DURÃES op cit.).
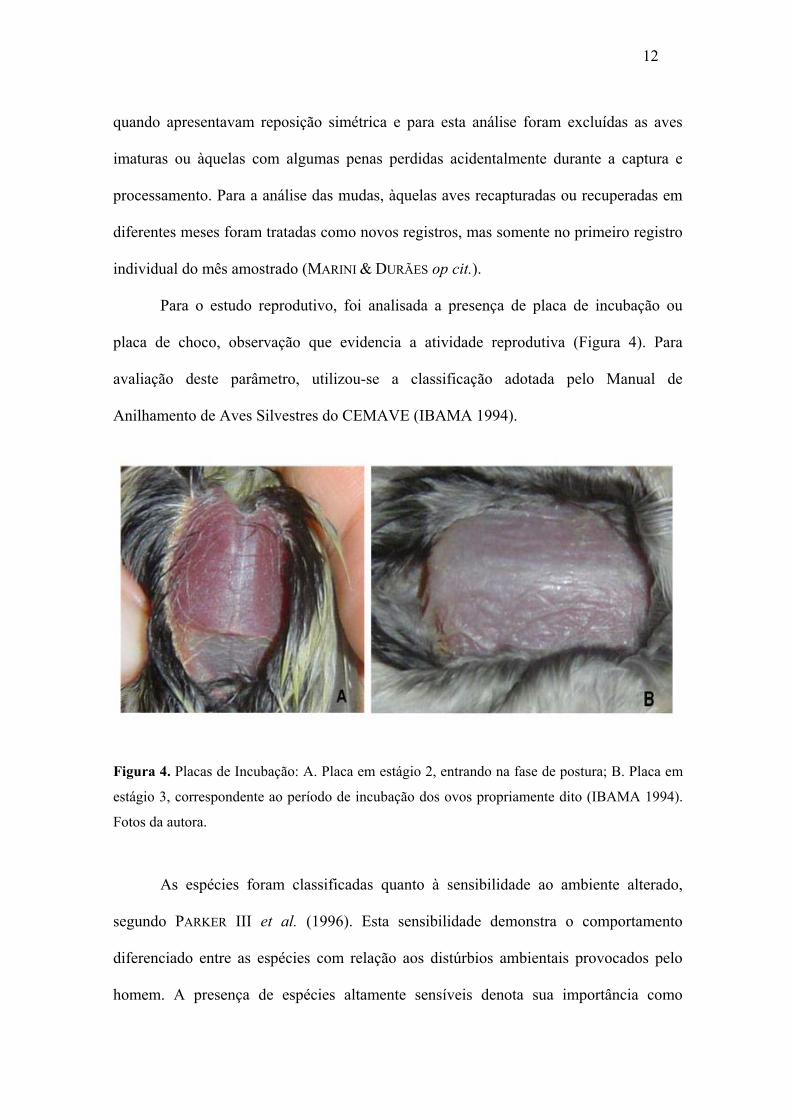
Para o estudo reprodutivo, foi analisada a presença de placa de incubação ou
placa de choco, observação que evidencia a atividade reprodutiva (Figura 4). Para
avaliação deste parâmetro, utilizou-se a classificação adotada pelo Manual de
Anilhamento de Aves Silvestres do CEMAVE (IBAMA 1994).
Figura 4. Placas de Incubação: A. Placa em estágio 2, entrando na fase de postura; B. Placa em
estágio 3, correspondente ao período de incubação dos ovos propriamente dito (IBAMA 1994).
Fotos da autora.
As espécies foram classificadas quanto à sensibilidade ao ambiente alterado,
segundo PARKER III et al. (1996). Esta sensibilidade demonstra o comportamento
diferenciado entre as espécies com relação aos distúrbios ambientais provocados pelo
homem. A presença de espécies altamente sensíveis denota sua importância como

13
bioindicadores de ambientes antropizados. PARKER III et al. (1996) determinam ainda
aquelas espécies com prioridade de conservação ou com prioridade de pesquisa
(Apêndice 1). A primeira é estimada através do estado de conservação do habitat, local
de distribuição da espécie, abundância relativa, tolerância ao habitat e sensibilidade aos
distúrbios. A segunda baseia-se no conhecimento da história natural, distribuição e
taxonomia de cada espécie. Poucas espécies neotropicais já foram estudadas
detalhadamente. Muitas dessas com prioridade de pesquisa ocorrem especificamente em
vários locais e são de grande importância para a conservação.
As espécies endêmicas e ameaçadas de extinção foram identificadas segundo
SICK (1997) e através da Lista Nacional das Espécies da Fauna Brasileira Ameaçadas de
Extinção (IBAMA 2003).
As guildas alimentares foram agrupadas em quatro categorias tróficas: Onívoras
(O), Granívoras (G), Insetívoras (I) e Frugívoras (F), seguindo WILLIS (1979),
TERBORGH et al. (1990) e SICK (1997).
3.3. TRATAMENTO ESTATÍSTICO
Quanto ao tratamento estatístico dos dados, foi calculada a freqüência de
ocorrência (FO) das espécies de aves capturadas. Estas interpretam, em porcentagem, o
número de visitas em que uma espécie foi encontrada, avaliando a regularidade com que
a mesma foi achada na área analisada. Foram obtidas distribuições absolutas, percentuais
das variáveis nominais ou categóricas e as medidas: média e desvio padrão para as
variáveis numéricas (estatística descritiva) e foi utilizado o teste t-Student para amostras
independentes com variâncias iguais ou desiguais e teste Qui-quadrado de independência
(estatística inferencial). A verificação da hipótese de igualdade de variâncias foi
realizada através do teste F.

14
Objetivando proporcionar informações sobre as semelhanças ecológicas das
amostras, foi empregado o Método de Ordenação através da Análise de Componentes
Principais (ACP) para espécies com FO superior a 58%, através do índice de correlação
de Pearson, utilizando-se o Software NTSYS pc 2.1. Para este método houve
cruzamento dos dados de FO com fatores abióticos, como temperatura e pluviometria.
O nível de significância utilizado na decisão dos testes estatísticos foi de 5%
(0,05). Os dados foram digitados na planilha Excel e o “software” utilizado para a
obtenção dos cálculos estatísticos foi o SAS (Statistical Analysis System) na versão 8.
15
4. RESULTADOS
4.1. BIOLOGIA DAS ESPÉCIES
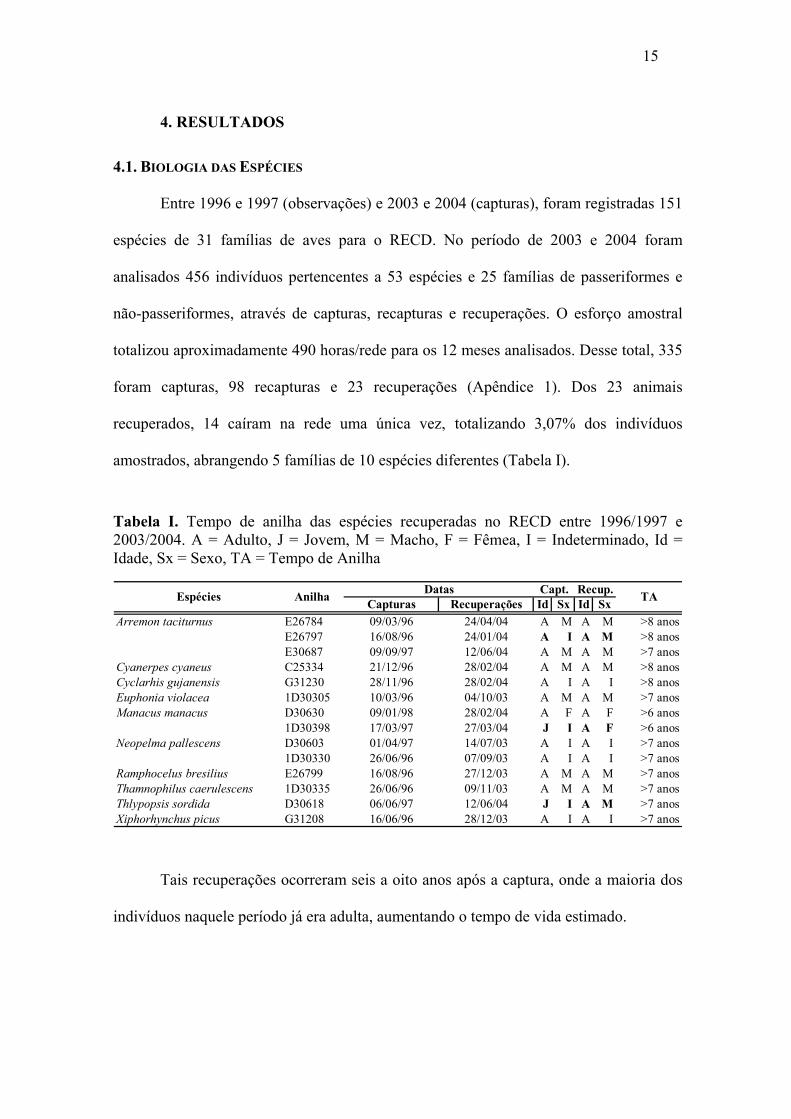
Entre 1996 e 1997 (observações) e 2003 e 2004 (capturas), foram registradas 151
espécies de 31 famílias de aves para o RECD. No período de 2003 e 2004 foram
analisados 456 indivíduos pertencentes a 53 espécies e 25 famílias de passeriformes e
não-passeriformes, através de capturas, recapturas e recuperações. O esforço amostral
totalizou aproximadamente 490 horas/rede para os 12 meses analisados. Desse total, 335
foram capturas, 98 recapturas e 23 recuperações (Apêndice 1). Dos 23 animais
recuperados, 14 caíram na rede uma única vez, totalizando 3,07% dos indivíduos
amostrados, abrangendo 5 famílias de 10 espécies diferentes (Tabela I).
Tabela I. Tempo de anilha das espécies recuperadas no RECD entre 1996/1997 e 2003/2004. A = Adulto, J = Jovem, M = Macho, F = Fêmea, I = Indeterminado, Id = Idade, Sx = Sexo, TA = Tempo de Anilha
Capturas Recuperações Id Sx Id SxArremon taciturnus E26784 09/03/96 24/04/04 A M A M >8 anos
E26797 16/08/96 24/01/04 A I A M >8 anosE30687 09/09/97 12/06/04 A M A M >7 anos
Cyanerpes cyaneus C25334 21/12/96 28/02/04 A M A M >8 anosCyclarhis gujanensis G31230 28/11/96 28/02/04 A I A I >8 anosEuphonia violacea 1D30305 10/03/96 04/10/03 A M A M >7 anosManacus manacus D30630 09/01/98 28/02/04 A F A F >6 anos
1D30398 17/03/97 27/03/04 J I A F >6 anosNeopelma pallescens D30603 01/04/97 14/07/03 A I A I >7 anos
1D30330 26/06/96 07/09/03 A I A I >7 anosRamphocelus bresilius E26799 16/08/96 27/12/03 A M A M >7 anosThamnophilus caerulescens 1D30335 26/06/96 09/11/03 A M A M >7 anosThlypopsis sordida D30618 06/06/97 12/06/04 J I A M >7 anosXiphorhynchus picus G31208 16/06/96 28/12/03 A I A I >7 anos
Espécies Capt.Datas TAAnilha Recup.
Tais recuperações ocorreram seis a oito anos após a captura, onde a maioria dos
indivíduos naquele período já era adulta, aumentando o tempo de vida estimado.
16
4.2. TEMPERATURA E ÍNDICES PLUVIOMÉTRICOS
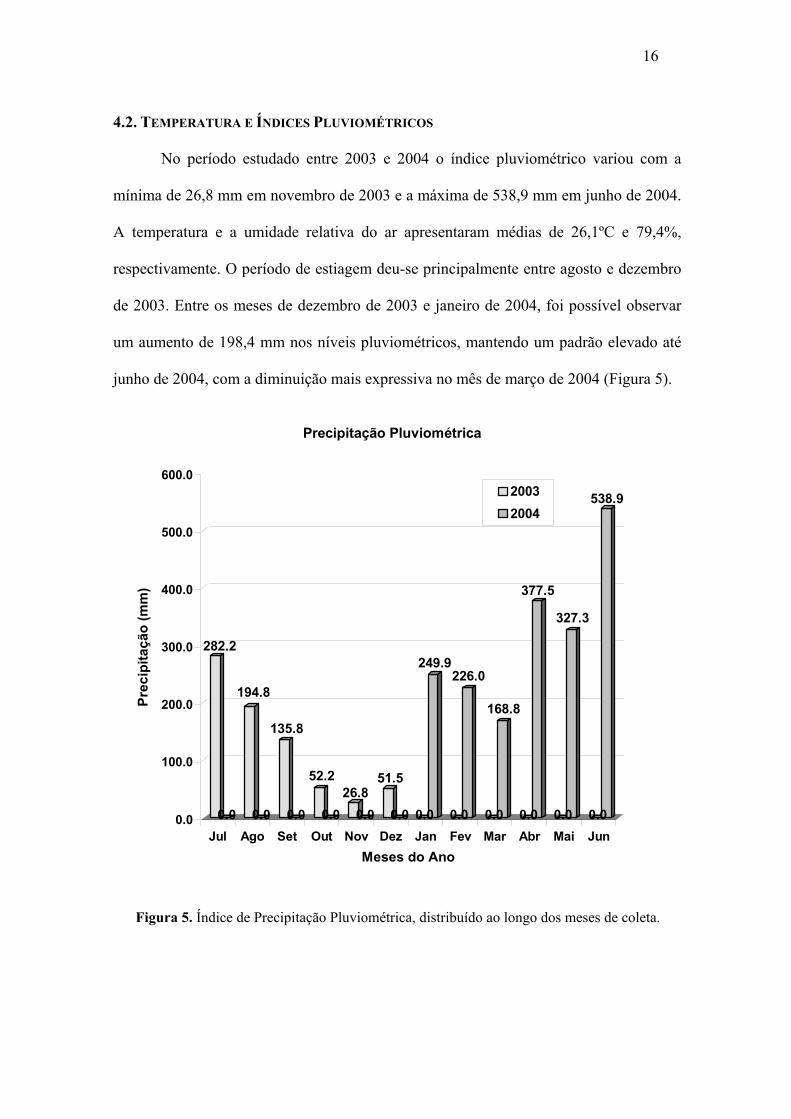
No período estudado entre 2003 e 2004 o índice pluviométrico variou com a
mínima de 26,8 mm em novembro de 2003 e a máxima de 538,9 mm em junho de 2004.
A temperatura e a umidade relativa do ar apresentaram médias de 26,1ºC e 79,4%,
respectivamente. O período de estiagem deu-se principalmente entre agosto e dezembro
de 2003. Entre os meses de dezembro de 2003 e janeiro de 2004, foi possível observar
um aumento de 198,4 mm nos níveis pluviométricos, mantendo um padrão elevado até
junho de 2004, com a diminuição mais expressiva no mês de março de 2004 (Figura 5).
282.2
0.0
194.8
0.0
135.8
0.0
52.2
0.026.8
0.0
51.5
0.0 0.0
249.9
0.0
226.0
0.0
168.8
0.0
377.5
0.0
327.3
0.0
538.9
0.0
100.0
200.0
300.0
400.0
500.0
600.0
Prec
ipita
ção
(mm
)
Jul Ago Set Out Nov Dez Jan Fev Mar Abr Mai JunMeses do Ano
Precipitação Pluviométrica
20032004
Figura 5. Índice de Precipitação Pluviométrica, distribuído ao longo dos meses de coleta.
17
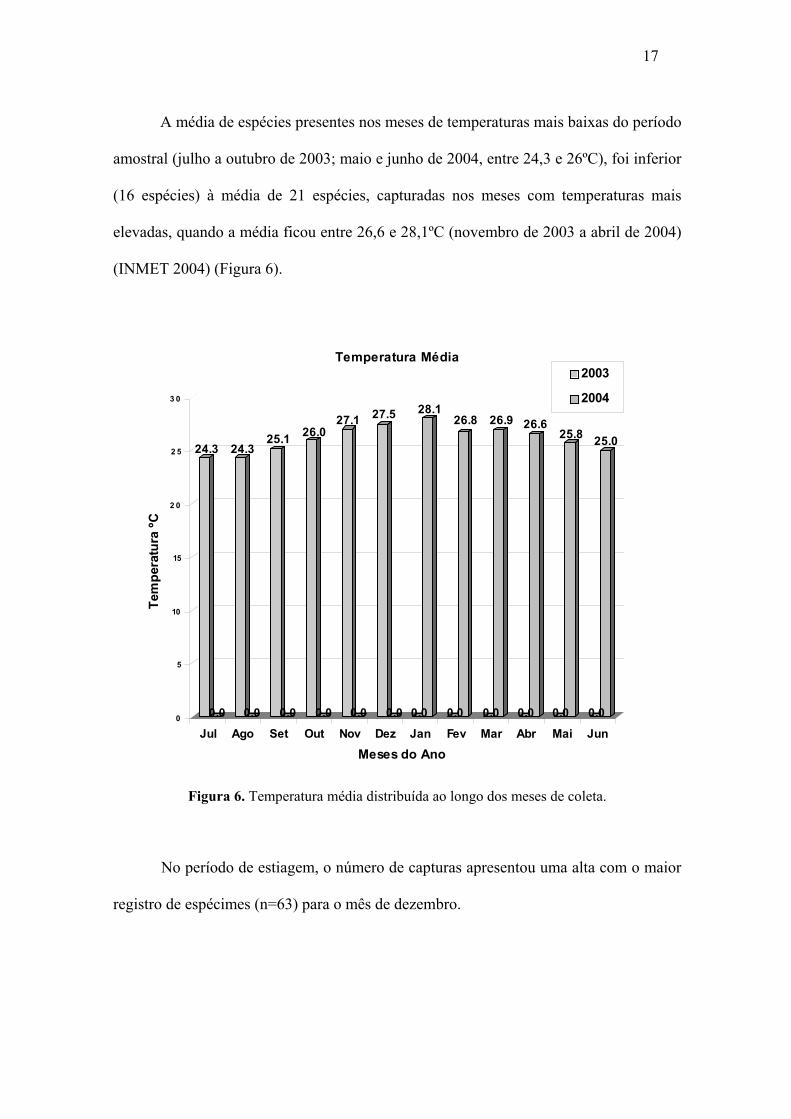
A média de espécies presentes nos meses de temperaturas mais baixas do período
amostral (julho a outubro de 2003; maio e junho de 2004, entre 24,3 e 26ºC), foi inferior
(16 espécies) à média de 21 espécies, capturadas nos meses com temperaturas mais
elevadas, quando a média ficou entre 26,6 e 28,1ºC (novembro de 2003 a abril de 2004)
(INMET 2004) (Figura 6).
24.3
0.0
24.3
0.0
25.1
0.0
26.0
0.0
27.1
0.0
27.5
0.0 0.0
28.1
0.0
26.8
0.0
26.9
0.0
26.6
0.0
25.8
0.0
25.0
0
5
10
15
2 0
2 5
3 0
Tem
pera
tura
ºC
Jul Ago Set Out Nov Dez Jan Fev Mar Abr Mai Jun
Meses do Ano
Temperatura Média2003
2004
Figura 6. Temperatura média distribuída ao longo dos meses de coleta.
No período de estiagem, o número de capturas apresentou uma alta com o maior
registro de espécimes (n=63) para o mês de dezembro.

18
4.3. BIONOMIA E BIOMETRIA
A maior parte (52,8%) da avifauna capturada no RECD apresentou FO inferior a
25%, ou seja, presentes em menos de quatro do total de 12 visitas (Tabela II). Manacus
manacus (Linnaeus, 1766), Arremon taciturnus (Hermann, 1783), Neopelma pallescens
(Lafresnaye, 1853) e Turdus leucomelas Vieillot 1818, foram as espécies mais
freqüentes, apresentando FO acima de 75%, com M. manacus representando 91,67% e
os demais, 83,33% das amostras. As três famílias mais representativas foram: Pipridae
(26,1%), seguida de Emberizidae (15,1%) e Tyrannidae (13,8%), totalizando 55% das
aves capturadas, com 71,3% dessas capturas ocorridas na AI.
Informações referentes à massa corpórea das espécies capturadas constam na
Tabela II, com suas médias, valores máximo e mínimo, bem como o desvio padrão para
aquelas que apresentaram um número de captura acima de dois indivíduos. Doze
espécies com representatividade única tiveram esse dado registrado, como segue:
Família Caprimulgidae, com Nyctidromus albicollis (Gmelin, 1789): 62,0g; Família
Trogonidae, com Trogon curucui (Linnaeus, 1766): 49,0g; Família Picidae, com
Colaptes melanochloros (Gmelin, 1788): 85,5g e Veniliornis passerinus (Linnaeus,
1766): 27,5g; Família Tyrannidae, com Elaenia cristata (Pelzeln, 1868): 14,0g, Pitangus
sulphuratus (Linnaeus, 1766): 51,0g e Pachyramphus polychopterus (Vieillot, 1818):
17,0g; Família Vireonidae, com Cyclarhis gujanensis (Gmelin, 1789): 28,0g e, Família
Emberizidae, com Thlypopsis sordida (D’Orbigny & Lafresnaye, 1837): 15g, Sporophila
nigricollis (Vieillot, 1823): 9,5g, Icterus cayanensis (Linnaeus, 1766): 32,5g e
Molothrus bonariensis (Gmelin, 1789): 42,0g.
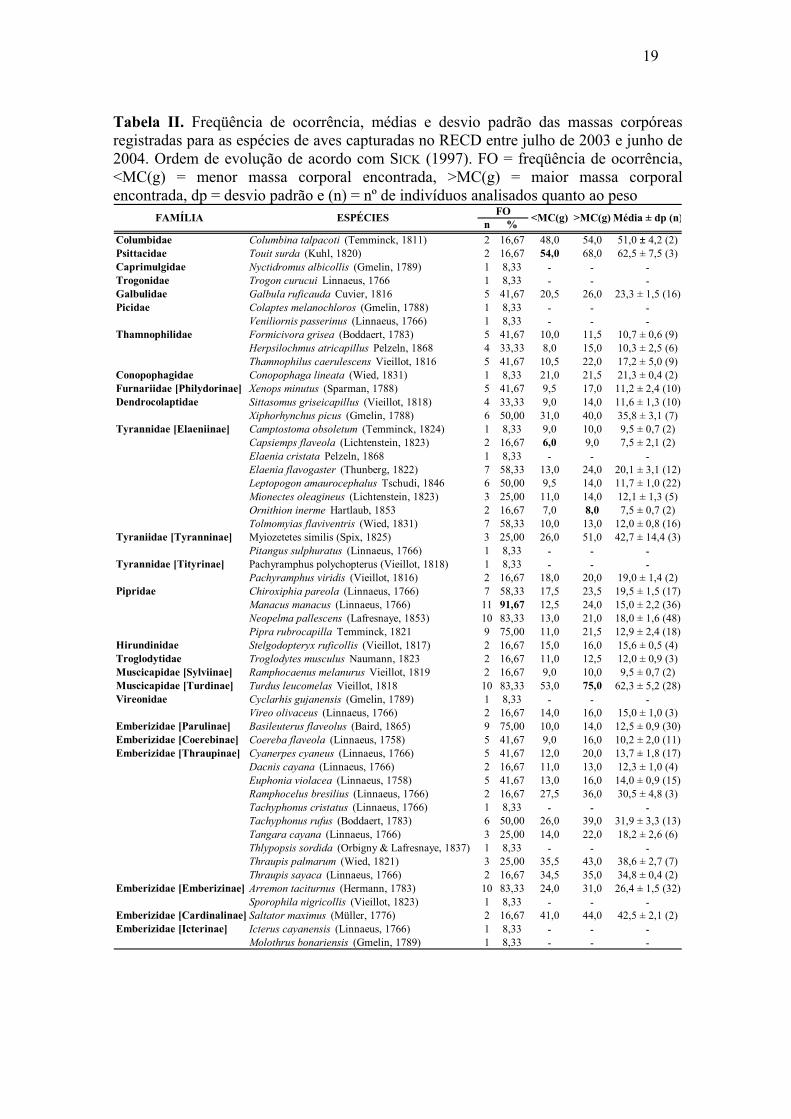
19
Tabela II. Freqüência de ocorrência, médias e desvio padrão das massas corpóreas registradas para as espécies de aves capturadas no RECD entre julho de 2003 e junho de 2004. Ordem de evolução de acordo com SICK (1997). FO = freqüência de ocorrência, <MC(g) = menor massa corporal encontrada, >MC(g) = maior massa corporal encontrada, dp = desvio padrão e (n) = nº de indivíduos analisados quanto ao peso
n %Columbidae Columbina talpacoti (Temminck, 1811) 2 16,67 48,0 54,0 51,0 ± 4,2 (2)Psittacidae Touit surda (Kuhl, 1820) 2 16,67 54,0 68,0 62,5 ± 7,5 (3)Caprimulgidae Nyctidromus albicollis (Gmelin, 1789) 1 8,33 ـ ـ ـTrogonidae Trogon curucui Linnaeus, 1766 1 8,33 ـ ـ ـGalbulidae Galbula ruficauda Cuvier, 1816 5 41,67 20,5 26,0 23,3 ± 1,5 (16)Picidae Colaptes melanochloros (Gmelin, 1788) 1 8,33 ـ ـ ـ
Veniliornis passerinus (Linnaeus, 1766) 1 8,33 ـ ـ ـThamnophilidae Formicivora grisea (Boddaert, 1783) 5 41,67 10,0 11,5 10,7 ± 0,6 (9)
Herpsilochmus atricapillus Pelzeln, 1868 4 33,33 8,0 15,0 10,3 ± 2,5 (6)Thamnophilus caerulescens Vieillot, 1816 5 41,67 10,5 22,0 17,2 ± 5,0 (9)
Conopophagidae Conopophaga lineata (Wied, 1831) 1 8,33 21,0 21,5 21,3 ± 0,4 (2)Furnariidae [Philydorinae] Xenops minutus (Sparman, 1788) 5 41,67 9,5 17,0 11,2 ± 2,4 (10)Dendrocolaptidae Sittasomus griseicapillus (Vieillot, 1818) 4 33,33 9,0 14,0 11,6 ± 1,3 (10)
Xiphorhynchus picus (Gmelin, 1788) 6 50,00 31,0 40,0 35,8 ± 3,1 (7)Tyrannidae [Elaeniinae] Camptostoma obsoletum (Temminck, 1824) 1 8,33 9,0 10,0 9,5 ± 0,7 (2)
Capsiemps flaveola (Lichtenstein, 1823) 2 16,67 6,0 9,0 7,5 ± 2,1 (2)Elaenia cristata Pelzeln, 1868 1 8,33 ـ ـ ـElaenia flavogaster (Thunberg, 1822) 7 58,33 13,0 24,0 20,1 ± 3,1 (12)Leptopogon amaurocephalus Tschudi, 1846 6 50,00 9,5 14,0 11,7 ± 1,0 (22)Mionectes oleagineus (Lichtenstein, 1823) 3 25,00 11,0 14,0 12,1 ± 1,3 (5)Ornithion inerme Hartlaub, 1853 2 16,67 7,0 8,0 7,5 ± 0,7 (2)Tolmomyias flaviventris (Wied, 1831) 7 58,33 10,0 13,0 12,0 ± 0,8 (16)
Tyraniidae [Tyranninae] Myiozetetes similis (Spix, 1825) 3 25,00 26,0 51,0 42,7 ± 14,4 (3)Pitangus sulphuratus (Linnaeus, 1766) 1 8,33 ـ ـ ـ
Tyrannidae [Tityrinae] Pachyramphus polychopterus (Vieillot, 1818) 1 8,33 ـ ـ ـPachyramphus viridis (Vieillot, 1816) 2 16,67 18,0 20,0 19,0 ± 1,4 (2)
Pipridae Chiroxiphia pareola (Linnaeus, 1766) 7 58,33 17,5 23,5 19,5 ± 1,5 (17)Manacus manacus (Linnaeus, 1766) 11 91,67 12,5 24,0 15,0 ± 2,2 (36)Neopelma pallescens (Lafresnaye, 1853) 10 83,33 13,0 21,0 18,0 ± 1,6 (48)Pipra rubrocapilla Temminck, 1821 9 75,00 11,0 21,5 12,9 ± 2,4 (18)
Hirundinidae Stelgodopteryx ruficollis (Vieillot, 1817) 2 16,67 15,0 16,0 15,6 ± 0,5 (4)Troglodytidae Troglodytes musculus Naumann, 1823 2 16,67 11,0 12,5 12,0 ± 0,9 (3)Muscicapidae [Sylviinae] Ramphocaenus melanurus Vieillot, 1819 2 16,67 9,0 10,0 9,5 ± 0,7 (2)Muscicapidae [Turdinae] Turdus leucomelas Vieillot, 1818 10 83,33 53,0 75,0 62,3 ± 5,2 (28)Vireonidae Cyclarhis gujanensis (Gmelin, 1789) 1 8,33 ـ ـ ـ
Vireo olivaceus (Linnaeus, 1766) 2 16,67 14,0 16,0 15,0 ± 1,0 (3)Emberizidae [Parulinae] Basileuterus flaveolus (Baird, 1865) 9 75,00 10,0 14,0 12,5 ± 0,9 (30)Emberizidae [Coerebinae] Coereba flaveola (Linnaeus, 1758) 5 41,67 9,0 16,0 10,2 ± 2,0 (11)Emberizidae [Thraupinae] Cyanerpes cyaneus (Linnaeus, 1766) 5 41,67 12,0 20,0 13,7 ± 1,8 (17)
Dacnis cayana (Linnaeus, 1766) 2 16,67 11,0 13,0 12,3 ± 1,0 (4)Euphonia violacea (Linnaeus, 1758) 5 41,67 13,0 16,0 14,0 ± 0,9 (15)Ramphocelus bresilius (Linnaeus, 1766) 2 16,67 27,5 36,0 30,5 ± 4,8 (3)Tachyphonus cristatus (Linnaeus, 1766) 1 8,33 ـ ـ ـTachyphonus rufus (Boddaert, 1783) 6 50,00 26,0 39,0 31,9 ± 3,3 (13)Tangara cayana (Linnaeus, 1766) 3 25,00 14,0 22,0 18,2 ± 2,6 (6)Thlypopsis sordida (Orbigny & Lafresnaye, 1837) 1 8,33 ـ ـ ـThraupis palmarum (Wied, 1821) 3 25,00 35,5 43,0 38,6 ± 2,7 (7)Thraupis sayaca (Linnaeus, 1766) 2 16,67 34,5 35,0 34,8 ± 0,4 (2)
Emberizidae [Emberizinae] Arremon taciturnus (Hermann, 1783) 10 83,33 24,0 31,0 26,4 ± 1,5 (32)Sporophila nigricollis (Vieillot, 1823) 1 8,33 ـ ـ ـ
Emberizidae [Cardinalinae] Saltator maximus (Müller, 1776) 2 16,67 41,0 44,0 42,5 ± 2,1 (2)Emberizidae [Icterinae] Icterus cayanensis (Linnaeus, 1766) 1 8,33 ـ ـ ـ
Molothrus bonariensis (Gmelin, 1789) 1 8,33 ـ ـ ـ
>MC(g) Média ± dp (n)FOFAMÍLIA ESPÉCIES <MC(g)

20
Foram realizados cálculos de média, desvio padrão e os resultados do teste
comparativo entre sexo e peso médio das aves, entre peso médio e estação do ano
(chuvosa – de abril a agosto e seca – de setembro a março) e entre sexo e medidas do
tamanho da asa e tarso e diâmetro do tarso (Tabelas III, IV e V). Com isso, objetivou-se
verificar se existe diferença significativa entre as variáveis para as espécies com pelo
menos cinco animais capturados em cada subgrupo (excluídos não adultos, sexo
indeterminado ou recapturas).
Foi possível destacar que os valores médios da massa corpórea foram mais
elevados entre os machos do que entre as fêmeas para a espécie Arremon taciturnus e o
contrário ocorreu com as espécies Cyanerpes cyaneus (Linnaeus, 1766) e Manacus
manacus. Entretanto A. taciturnus foi a única espécie que apresentou diferença
significante entre sexos (p < 0,05) (Tabela III).
Tabela III. Média e desvio padrão da massa corpórea de três espécies de aves segundo o sexo dos animais. (n) = número de indivíduos analisados por espécie e sexo, dp = desvio padrão Sexo Espécie Fêmea Macho Valor de p Média ± dp (n) Média ± dp (n)
Arremon taciturnus 25,20 ± 1,10 (5) 26,69 ± 1,23 (13) p(1) = 0,0312* Cyanerpes cyaneus 14,58 ± 2,94 (6) 13,25 ± 0,68 (10) p(2) = 0,3211 Manacus manacus 15,03 ± 2,78 (17) 14,86 ± 0,69 (7) p(1) = 0,8742
(*) – Diferença significante ao nível de 5,0%. (1) – Através do teste t-Student com variâncias iguais. (2) – Através do teste t-Student com variâncias desiguais.
Verificou-se que os valores médios foram mais elevados entre os animais
capturados na estação chuvosa do que na estação seca para as espécies: M. manacus e
Pipra rubrocapilla Temminck, 1821 e o contrário ocorreu com a espécie A. taciturnus.
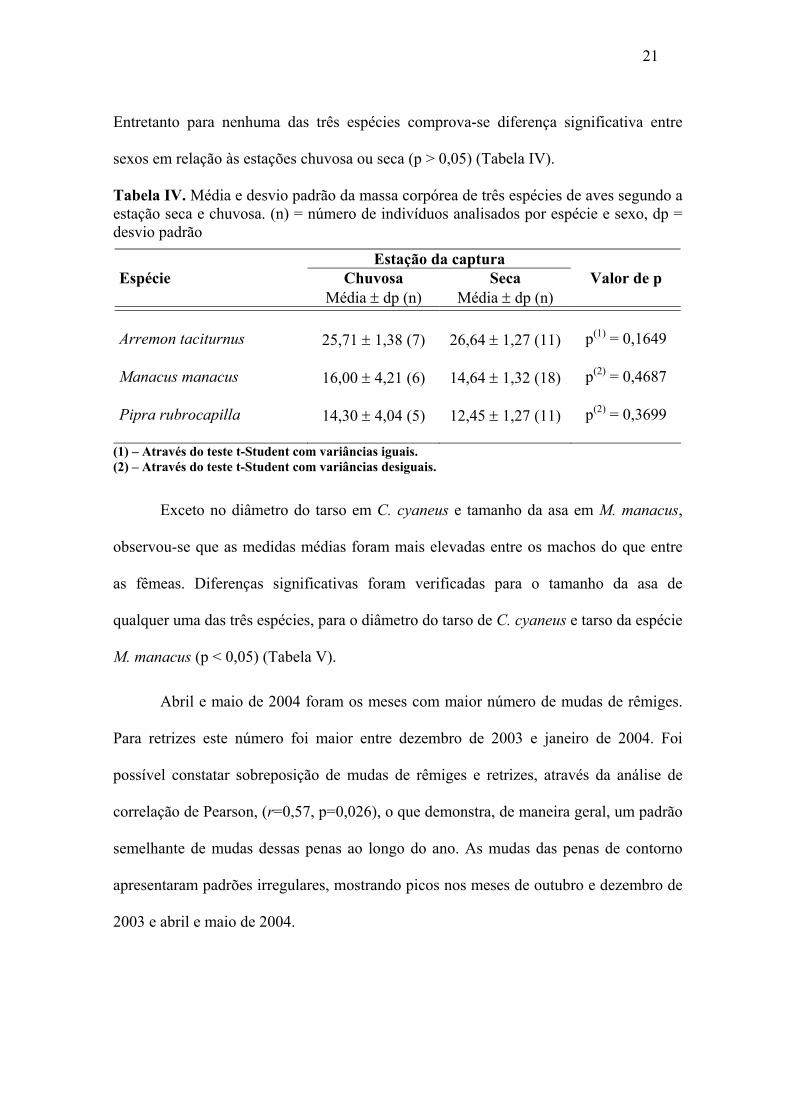
21
Entretanto para nenhuma das três espécies comprova-se diferença significativa entre
sexos em relação às estações chuvosa ou seca (p > 0,05) (Tabela IV).
Tabela IV. Média e desvio padrão da massa corpórea de três espécies de aves segundo a estação seca e chuvosa. (n) = número de indivíduos analisados por espécie e sexo, dp = desvio padrão
Estação da captura Espécie Chuvosa Seca Valor de p Média ± dp (n) Média ± dp (n)
Arremon taciturnus 25,71 ± 1,38 (7) 26,64 ± 1,27 (11) p(1) = 0,1649 Manacus manacus 16,00 ± 4,21 (6) 14,64 ± 1,32 (18) p(2) = 0,4687 Pipra rubrocapilla 14,30 ± 4,04 (5) 12,45 ± 1,27 (11) p(2) = 0,3699
(1) – Através do teste t-Student com variâncias iguais. (2) – Através do teste t-Student com variâncias desiguais. Exceto no diâmetro do tarso em C. cyaneus e tamanho da asa em M. manacus,
observou-se que as medidas médias foram mais elevadas entre os machos do que entre
as fêmeas. Diferenças significativas foram verificadas para o tamanho da asa de
qualquer uma das três espécies, para o diâmetro do tarso de C. cyaneus e tarso da espécie
M. manacus (p < 0,05) (Tabela V).
Abril e maio de 2004 foram os meses com maior número de mudas de rêmiges.
Para retrizes este número foi maior entre dezembro de 2003 e janeiro de 2004. Foi
possível constatar sobreposição de mudas de rêmiges e retrizes, através da análise de
correlação de Pearson, (r=0,57, p=0,026), o que demonstra, de maneira geral, um padrão
semelhante de mudas dessas penas ao longo do ano. As mudas das penas de contorno
apresentaram padrões irregulares, mostrando picos nos meses de outubro e dezembro de
2003 e abril e maio de 2004.
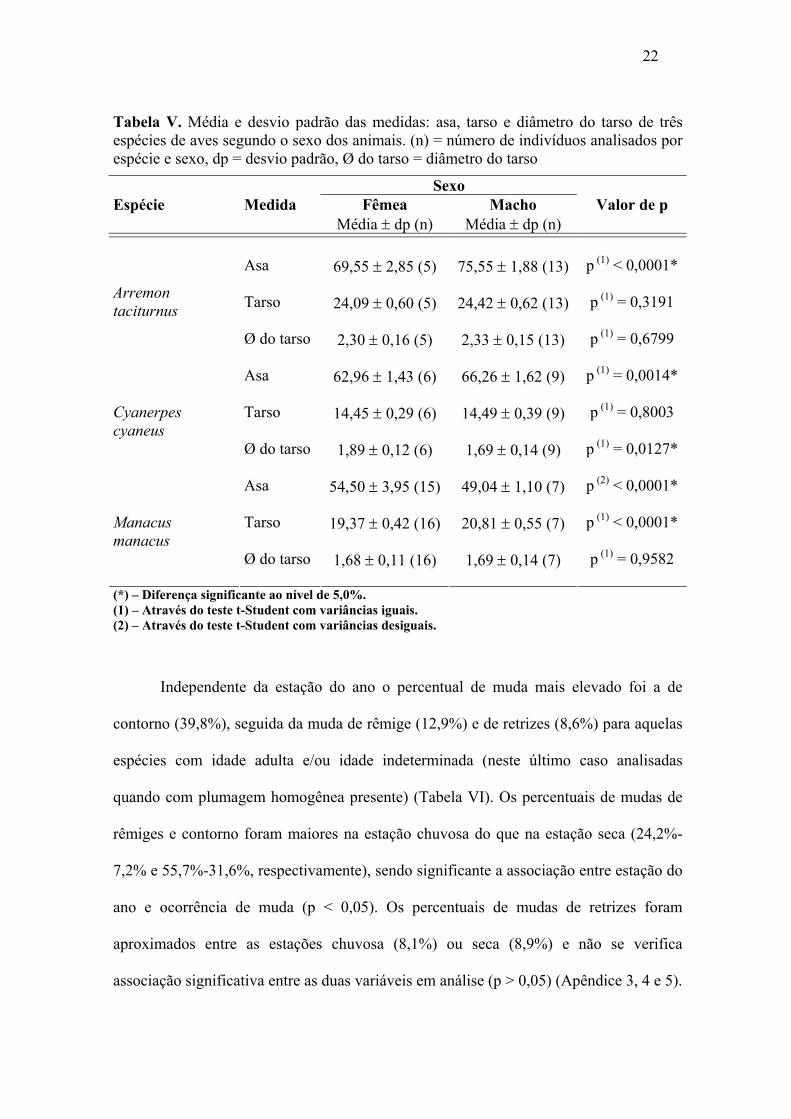
22
Tabela V. Média e desvio padrão das medidas: asa, tarso e diâmetro do tarso de três espécies de aves segundo o sexo dos animais. (n) = número de indivíduos analisados por espécie e sexo, dp = desvio padrão, Ø do tarso = diâmetro do tarso
Sexo Espécie Medida Fêmea Macho Valor de p Média ± dp (n) Média ± dp (n)
Asa
69,55 ± 2,85 (5) 75,55 ± 1,88 (13) p (1) < 0,0001*
Tarso
24,09 ± 0,60 (5) 24,42 ± 0,62 (13) p (1) = 0,3191 Arremon taciturnus
Ø do tarso
2,30 ± 0,16 (5) 2,33 ± 0,15 (13) p (1) = 0,6799
Asa
62,96 ± 1,43 (6) 66,26 ± 1,62 (9) p (1) = 0,0014*
Tarso
14,45 ± 0,29 (6) 14,49 ± 0,39 (9) p (1) = 0,8003 Cyanerpes cyaneus
Ø do tarso
1,89 ± 0,12 (6) 1,69 ± 0,14 (9) p (1) = 0,0127*
Asa
54,50 ± 3,95 (15) 49,04 ± 1,10 (7) p (2) < 0,0001*
Tarso
19,37 ± 0,42 (16) 20,81 ± 0,55 (7) p (1) < 0,0001* Manacus manacus
Ø do tarso
1,68 ± 0,11 (16) 1,69 ± 0,14 (7) p (1) = 0,9582
(*) – Diferença significante ao nível de 5,0%. (1) – Através do teste t-Student com variâncias iguais. (2) – Através do teste t-Student com variâncias desiguais.
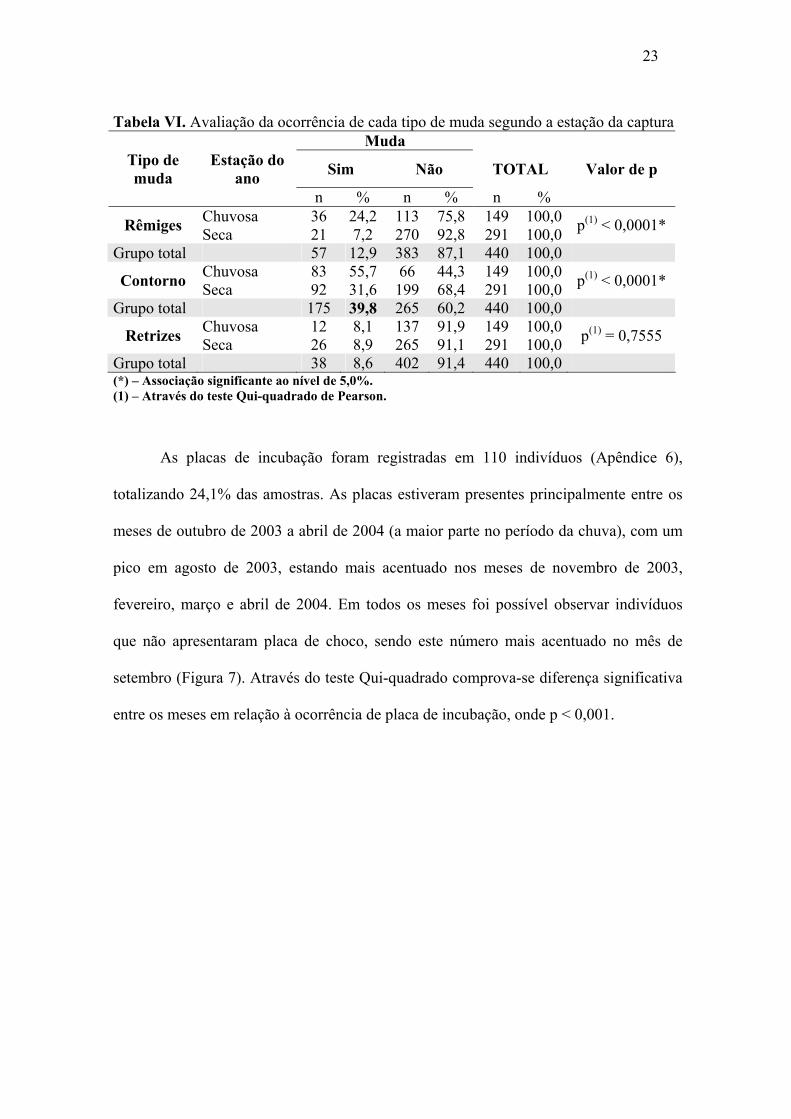
Independente da estação do ano o percentual de muda mais elevado foi a de
contorno (39,8%), seguida da muda de rêmige (12,9%) e de retrizes (8,6%) para aquelas
espécies com idade adulta e/ou idade indeterminada (neste último caso analisadas
quando com plumagem homogênea presente) (Tabela VI). Os percentuais de mudas de
rêmiges e contorno foram maiores na estação chuvosa do que na estação seca (24,2%-
7,2% e 55,7%-31,6%, respectivamente), sendo significante a associação entre estação do
ano e ocorrência de muda (p < 0,05). Os percentuais de mudas de retrizes foram
aproximados entre as estações chuvosa (8,1%) ou seca (8,9%) e não se verifica
associação significativa entre as duas variáveis em análise (p > 0,05) (Apêndice 3, 4 e 5).
23
Tabela VI. Avaliação da ocorrência de cada tipo de muda segundo a estação da captura Muda
Tipo de muda
Estação do ano Sim Não TOTAL Valor de p
n % n % n % Chuvosa 36 24,2 113 75,8 149 100,0 Rêmiges Seca 21 7,2 270 92,8 291 100,0 p(1) < 0,0001*
Grupo total 57 12,9 383 87,1 440 100,0 Chuvosa 83 55,7 66 44,3 149 100,0 Contorno Seca 92 31,6 199 68,4 291 100,0 p(1) < 0,0001*
Grupo total 175 39,8 265 60,2 440 100,0 Chuvosa 12 8,1 137 91,9 149 100,0 Retrizes Seca 26 8,9 265 91,1 291 100,0 p(1) = 0,7555
Grupo total 38 8,6 402 91,4 440 100,0 (*) – Associação significante ao nível de 5,0%. (1) – Através do teste Qui-quadrado de Pearson.
As placas de incubação foram registradas em 110 indivíduos (Apêndice 6),
totalizando 24,1% das amostras. As placas estiveram presentes principalmente entre os
meses de outubro de 2003 a abril de 2004 (a maior parte no período da chuva), com um
pico em agosto de 2003, estando mais acentuado nos meses de novembro de 2003,
fevereiro, março e abril de 2004. Em todos os meses foi possível observar indivíduos
que não apresentaram placa de choco, sendo este número mais acentuado no mês de
setembro (Figura 7). Através do teste Qui-quadrado comprova-se diferença significativa
entre os meses em relação à ocorrência de placa de incubação, onde p < 0,001.
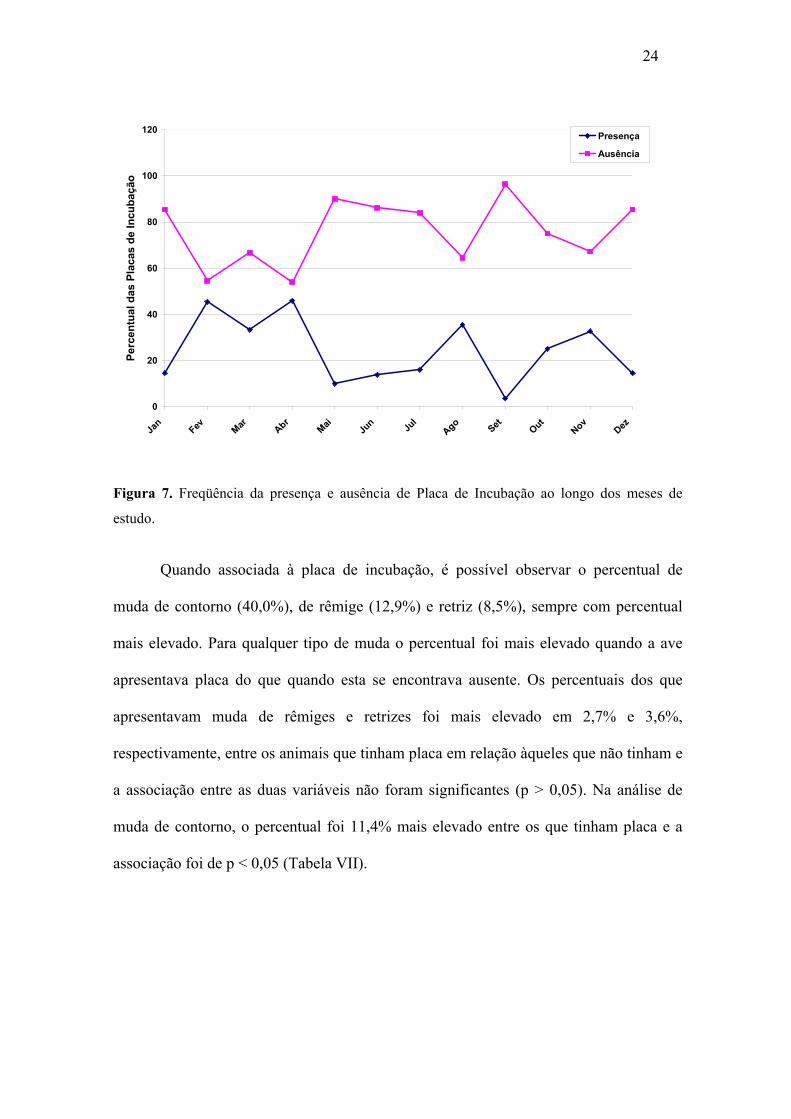
24
0
20
40
60
80
100
120
Jan
Fev Mar AbrMai Ju
n Jul
Ago Set OutNov
Dez
Perc
entu
al d
as P
laca
s de
Incu
baçã
oPresença
Ausência
Figura 7. Freqüência da presença e ausência de Placa de Incubação ao longo dos meses de
estudo.
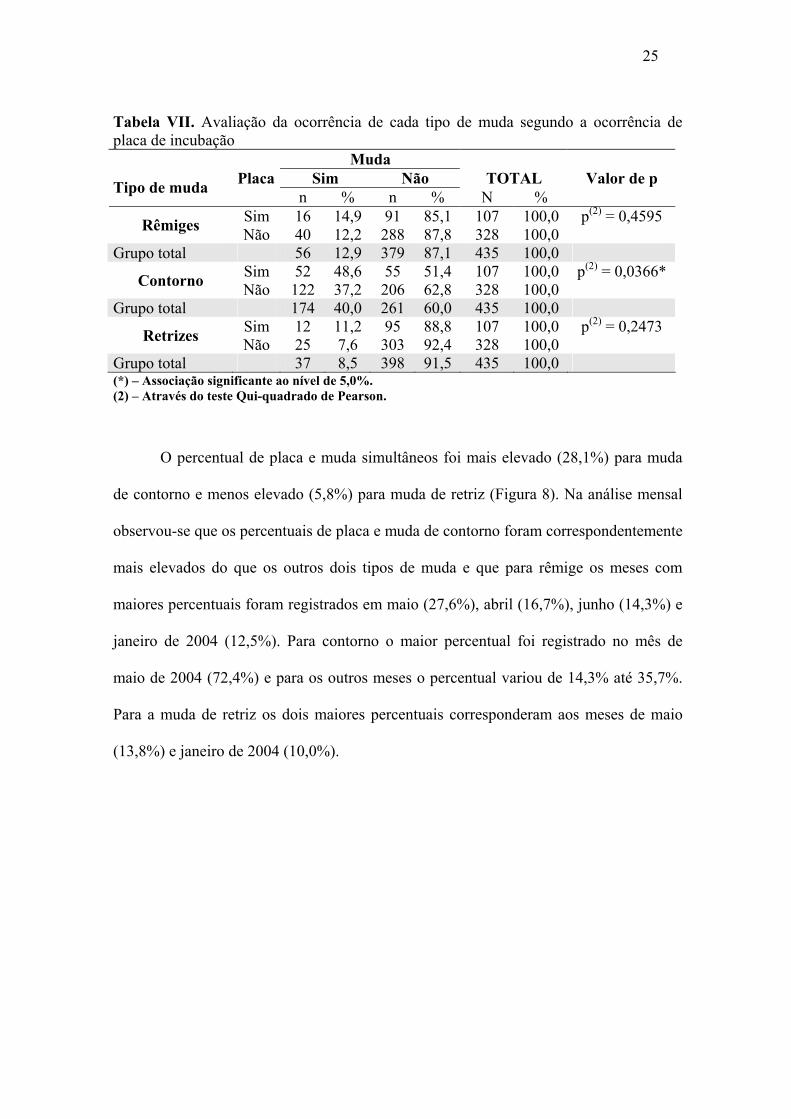
Quando associada à placa de incubação, é possível observar o percentual de
muda de contorno (40,0%), de rêmige (12,9%) e retriz (8,5%), sempre com percentual
mais elevado. Para qualquer tipo de muda o percentual foi mais elevado quando a ave
apresentava placa do que quando esta se encontrava ausente. Os percentuais dos que
apresentavam muda de rêmiges e retrizes foi mais elevado em 2,7% e 3,6%,
respectivamente, entre os animais que tinham placa em relação àqueles que não tinham e
a associação entre as duas variáveis não foram significantes (p > 0,05). Na análise de
muda de contorno, o percentual foi 11,4% mais elevado entre os que tinham placa e a
associação foi de p < 0,05 (Tabela VII).
25
Tabela VII. Avaliação da ocorrência de cada tipo de muda segundo a ocorrência de placa de incubação Muda
Placa Sim Não TOTAL Valor de p Tipo de muda n % n % N % Sim 16 14,9 91 85,1 107 100,0 p(2) = 0,4595 Rêmiges Não 40 12,2 288 87,8 328 100,0
Grupo total 56 12,9 379 87,1 435 100,0 Sim 52 48,6 55 51,4 107 100,0 p(2) = 0,0366* Contorno Não 122 37,2 206 62,8 328 100,0
Grupo total 174 40,0 261 60,0 435 100,0 Sim 12 11,2 95 88,8 107 100,0 p(2) = 0,2473 Retrizes Não 25 7,6 303 92,4 328 100,0
Grupo total 37 8,5 398 91,5 435 100,0 (*) – Associação significante ao nível de 5,0%. (2) – Através do teste Qui-quadrado de Pearson.
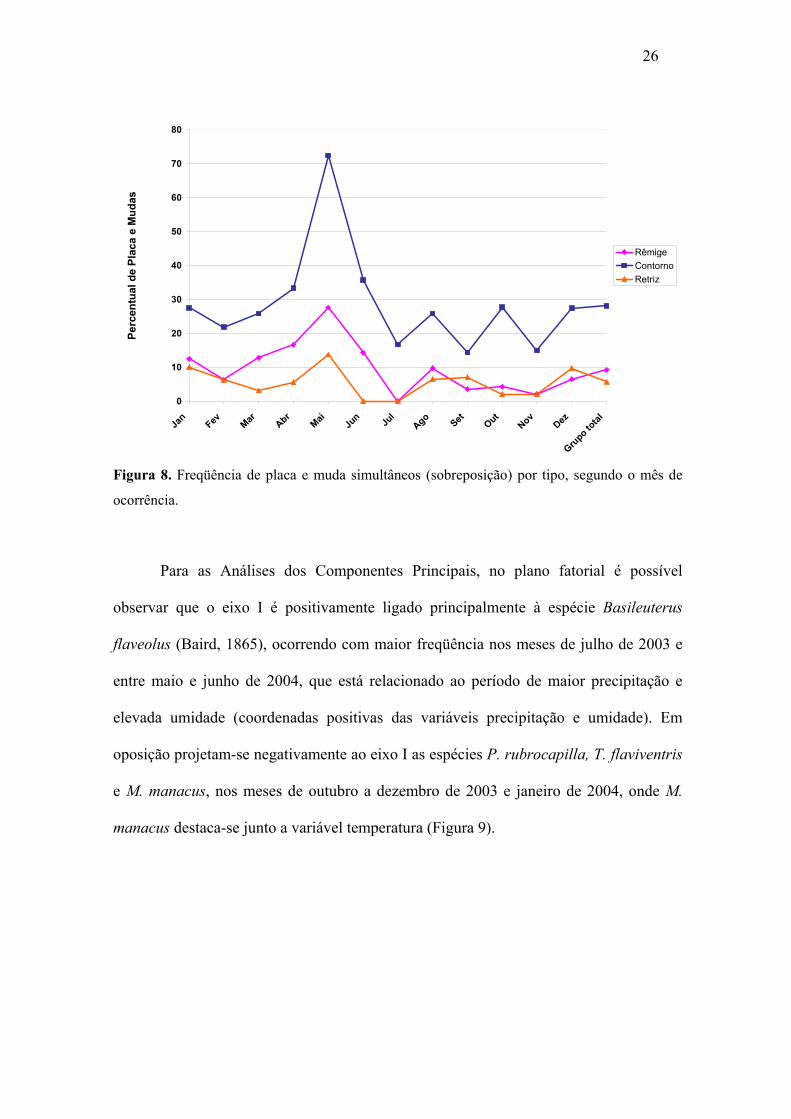
O percentual de placa e muda simultâneos foi mais elevado (28,1%) para muda
de contorno e menos elevado (5,8%) para muda de retriz (Figura 8). Na análise mensal
observou-se que os percentuais de placa e muda de contorno foram correspondentemente
mais elevados do que os outros dois tipos de muda e que para rêmige os meses com
maiores percentuais foram registrados em maio (27,6%), abril (16,7%), junho (14,3%) e
janeiro de 2004 (12,5%). Para contorno o maior percentual foi registrado no mês de
maio de 2004 (72,4%) e para os outros meses o percentual variou de 14,3% até 35,7%.
Para a muda de retriz os dois maiores percentuais corresponderam aos meses de maio
(13,8%) e janeiro de 2004 (10,0%).
26
0
10
20
30
40
50
60
70
80
Jan
Fev Mar AbrMai Ju
n Jul
Ago Set OutNov
Dez
Grupo to
tal
Perc
entu
al d
e Pl
aca
e M
udas
RêmigeContornoRetriz
Figura 8. Freqüência de placa e muda simultâneos (sobreposição) por tipo, segundo o mês de
ocorrência.
Para as Análises dos Componentes Principais, no plano fatorial é possível
observar que o eixo I é positivamente ligado principalmente à espécie Basileuterus
flaveolus (Baird, 1865), ocorrendo com maior freqüência nos meses de julho de 2003 e
entre maio e junho de 2004, que está relacionado ao período de maior precipitação e
elevada umidade (coordenadas positivas das variáveis precipitação e umidade). Em
oposição projetam-se negativamente ao eixo I as espécies P. rubrocapilla, T. flaviventris
e M. manacus, nos meses de outubro a dezembro de 2003 e janeiro de 2004, onde M.
manacus destaca-se junto a variável temperatura (Figura 9).
27
Figura 9. Representação gráfica de um plano fatorial numa análise dos componentes principais,
utilizando-se o método de ordenação e projeção. Foram projetados no Eixo I-II os vetores
espécies, fatores abióticos e meses de captura.
Para as espécies identificadas na listagem geral registrada para o RECD, seis
apresentaram alta sensibilidade aos distúrbios ambientais, de acordo com PARKER III et
al. (1996), sendo elas: Micrastur gilvicollis (Vieillot, 1817); Aramides cajanea (Statius
Muller, 1776); Touit surda (Kuhl, 1820); Hemitriccus zosterops (Pelzeln, 1868);
Chiroxiphia pareola (Linnaeus, 1766); Pipra rubrocapilla Temminck, 1821 (Apêndice
1). As espécies Conopophaga lineata (Wied, 1821), Touit surda (Kuhl, 1820) e
Iodopleura pipra (Lesson, 1831), apresentaram alta prioridade de conservação, sendo as
duas últimas também com alta prioridade de pesquisa.

28
Destas 151 espécies, quatro - Conopophaga lineata (Wied, 1831), Xenops
minutus (Sparrman, 1788), Picumnus exilis (Lichtenstein, 1823) e Thamnophilus
caerulescens Vieillot, 1816 - encontram-se na categoria de ameaçada de extinção e três -
Touit surda (Kuhl, 1820), Iodopleura pipra (Lesson, 1831) e Ramphocelus bresilius
(Linnaeus, 1766) - foram consideradas endêmicas para a região (Figura 10).
Figura 10. Espécies capturadas no RECD entre julho de 2003 e junho de 2004. A. Conopophaga
lineata, B. Xenops minutus, C. Touit surda, D. Ornithion inerme. Fotos da autora.
As espécies onívoras apresentaram um percentual mais elevado do que as demais
guildas alimentares identificadas para estas amostras, com 183 indivíduos (40,1%) dos
representantes, acompanhados dos insetívoros (36%). Os granívoros tiveram o menor
registro (0,7%) para este método de captura.
No RECD, três indivíduos de Touit surda foram capturados, sendo um casal em
01/11/2003 e o terceiro em 11/06/2004, todos na AI e na mesma rede que se encontrava

29
disposta entre dois sapotizeiros (Achras sapota L.), de altura inferior a quatro metros. Os
dois primeiros foram depositados na coleção da Universidade Federal de Pernambuco
(UFPE) sob número de tombo 4024 para o macho e 4025 para a fêmea. Ambos
encontravam-se forrageando nos sapotizeiros uma vez que apresentavam polpa da fruta
aderida ao bico (Tabela VIII).
Os dois indivíduos de Ornithion inerme foram capturados nos dias 28/12/03 e
24/04/04, ambos na AI. O primeiro teve sua pele encaminhada para a coleção da UFPE,
sob número de tombo 3993. Salienta-se que este é o primeiro registro de captura da
espécie para o Estado. O segundo exemplar foi anilhado, com anilha metálica C28657 e
dados de bionomia e biometria foram coletados. Ambos apresentavam muda de
contorno, mas apenas o indivíduo anilhado substituía sua 9a rêmige e indicava placa de
choco em estágio final de sucessão, sugerindo que o período de reprodução deveria estar
terminando ou já concluído. Pesavam 7 e 8g, com comprimento total de 7,2 e 9,5cm,
respectivamente, onde aquele indivíduo anilhado sempre apresentou as maiores medidas
(Tabela VIII).
30
Tabela VIII. Informações sobre bionomia e biometria de duas espécies de aves para o RECD. ST = status, ST1 = ave nova, AN = anilha, GT = guilda trófica, Frug. = frugívoro, Inset. = insetívoro, ID = idade, I = indeterminado, A = adulto, SX = sexo, PI = placa de incubação, RG = rêmiges, N = ausente, CN = contorno, C = cabeça, D = dorso, V = ventre, RT = retriz, RP = penas em reposição, RTD = reposição retriz direita, RTE = reposição retriz esquerda, AS = comprimento da asa, T = comprimento do tarso, ØT = diâmetro do tarso, CC = comprimento da cauda, CM = cúlmen total, NP = narina ponta, CT = comprimento total, P = peso
RG CN RT RP AS T ØT CC CM NP CT
Psittacidae Touit surda (Kuhl, 1820) 1/11/2003 16:05 1 ÓBITO Frug. I I 119,0 12,4 3,5 51,0 15,5 17,9 160,0 68,0
Psittacidae Touit surda (Kuhl, 1820) 1/11/2003 16:00 1 ÓBITO Frug. I I N N N N 119,0 12,2 3,8 45,0 16,6 17,8 155,0 65,5
Psittacidae Touit surda (Kuhl, 1820) 11/6/2004 15:35 1 J21609 Frug. A I 0 N C,D,V N 1,2,5RTD/3,6RTE 115,0 12,9 3,3 50,0 14,5 17,0 165,0 54,0
Tyrannidae Ornithion inerme Hartlaub, 1853 24/4/2004 16:00 1 C28657 Inset. I I 5 9 C,D,V N 6RTE 49,0 19,7 1,6 38,0 12,6 9,9 95,0 8,0
Tyrannidae Ornithion inerme Hartlaub, 1853 28/12/2003 14:20 1 ÓBITO Inset. I I 0 N C,D,V N N 42,0 13,0 1,5 24,5 8,0 7,2 72,0 7,0
PI Mudas Medidas (mm) P(g)AN GT ID SXFamília Espécie Hora STData Capt.

31
Em 27 de dezembro de 2003, na trilha de acesso ao RECD, foi observada a
ocupação de um ninho (provavelmente de cupim) pela espécie Trogon curucui Linnaeus,
1766 (Surucuá-de-barriga-vermelha), que o utiliza para nidificar (Figura 11),
confirmando o início do período de atividade reprodutiva demonstrado pela presença da
placa de incubação nas demais espécies.
Figura 11. A. Ninho ocupado em dezembro de 2003 pelo Trogon curucui (Surucuá-de-barriga-
vermelha). B. Ninho destruído em fevereiro de 2004, com ovo em seu interior. C. Detalhe dos
outros ovos destruídos fora do ninho. Fotos da autora.
Em fevereiro de 2004 o mesmo ninho foi encontrado no solo, destruído,
apresentando três ovos, dos quais um ainda estava no seu interior e os demais se

32
encontravam ao lado, externamente, com a casca parcialmente destruída e seu conteúdo
totalmente removido, sugerindo o ataque por animais de pequeno porte.

33
5. DISCUSSÃO
Modelos estatísticos podem ser empregados para determinar a estimativa de
sobrevivência das aves através de capturas e recapturas de indivíduos marcados, embora
em geral esses estudos necessitem de um acompanhamento contínuo dessas recapturas
ou recuperações ao longo dos anos (ELDER & ZIMMERMAN 1983, PAYNE & PAYNE 1990).
As recuperações ocorridas demonstram a fidelidade de algumas aves ao ambiente
e reforçam a importância dos pequenos fragmentos para espécies residentes e ocasionais.
Com isso, foi possível registrar o tempo de anilha para determinadas espécies, conforme
apresentado por SNOW & LILL (1974), que relatam a idade para Manacus manacus em
mais de 14 anos e LOPES et al. (1980) que fornecem o tempo de anilha para várias aves
que serviram para estudos sobre possíveis vírus presentes neste grupo silvestre. Desses
dados, apenas duas espécies (M. manacus e Thamnophilus caerulescens) coincidiram
entre as pesquisas e nenhum tempo de anilha apresentado por aqueles autores foi maior
do que o encontrado neste trabalho.
Os resultados obtidos para a composição da listagem de aves para o RECD
demonstram semelhança a alguns trabalhos desenvolvidos nas matas pernambucanas, a
exemplo de AZEVEDO-JÚNIOR (1990) que apresentou uma listagem com 181 espécies
para a Estação Ecológica do Tapacurá e LYRA-NEVES et al. (2000), com o registro de
187 espécies para a Área de Proteção Ambiental de Guadalupe, ambas com áreas
superiores a 60 ha. Estes resultados diferem daquele encontrado para a Reserva Estadual
de Gurjaú, com 220 espécies de aves que foram registradas em 1077,10 ha de mata
dividida em três regiões, apresentando um resultado alto se comparado aos
levantamentos realizados nos trabalhos descritos (LYRA-NEVES et al. 2004). Essa

34
aproximação do número de espécies encontradas no pequeno fragmento do RECD com
as demais áreas citadas pode estar relacionada à presença de florestas circunvizinhas, a
exemplo da Usina São José (distando aproximadamente 4 km da área estudada), o que
certamente contribui com a existência de muitas espécies que sobrevivem em pequenos
fragmentos interligados a plantações, pastagens e outros fragmentos, promovendo a
ampliação do uso de recurso por parte desses animais (ALEIXO & GALETTI 1997;
MALDONADO-COELHO & MARINI 2000; TELINO-JÚNIOR et al. 2000).
A partir de outubro o número de capturas aumentou e isto pode corresponder a
maiores padrões de atividades das aves no verão. ALMEIDA et al. (1999) considera o mês
de setembro de grande atividade das aves, período a ser considerado quanto à realização
de levantamentos avifaunísticos, quando é possível amostrar um maior número de
espécies.
Independente da elevação das capturas em períodos quentes do ano, resultados de
baixa FO são esperados. Isto se deve, juntamente ao elevado índice de registros únicos
de algumas espécies, à metodologia aplicada, que restringe a captura a determinadas
espécies pelo fato de ocuparem diferentes estratos da floresta que não contemplam o
intervalo de altura das redes quando desenvolvem as suas atividades como alimentação e
nidificação. Concomitante a isso se faz necessário observar padrões fitogeográficos da
área, uma vez que o fragmento encontra-se em fase de regeneração, o que pode limitar
sua diversidade.
A FO demonstrou que apenas três famílias já representavam mais de 50% das
espécies capturadas (Pipridae, seguida de Emberizidae e Tyrannidae). Este advento é
explicado quando se observam que as espécies mais numerosas são aquelas
caracterizadas pela vasta distribuição e habitarem, sobretudo, a mata e capoeira.

35
TERBORGH (1992) afirma ser altamente incerto o futuro de muitas espécies nas pequenas
“sobras” de habitats fragmentados, embora esses fragmentos possam prover
perfeitamente bem determinadas espécies, sendo insuficientes para outras. Manacus
manacus, Neopelma pallescens e Arremon taciturnus, espécies frugívoras/insetívoras e
onívora, provavelmente tiveram uma maior representatividade devido ao efeito de borda
que esta área está sofrendo, pois possui estreita faixa de vegetação natural. A maior
incidência de luz neste ambiente proporciona maior produção de frutos e de plantas
invasoras produtoras de sementes, que são a base alimentar destas espécies (DÁRIO et al.
2002).
Informações obtidas acerca da bionomia dos indivíduos capturados neste
fragmento, oferecem subsídios para um maior conhecimento sobre a ornitofauna,
principalmente no que diz respeito à região nordestina do Brasil.
Alguns estudos prévios (DUNNING 1987, BELTON 1994, SICK 1997) compilaram
dados sobre a massa corpórea de aves do mundo. Outras pesquisas fizeram registros
mais localizados, com aves da Mata Atlântica, a exemplo de ONIKI (1981), ONIKI &
WILLIS (1993), REINERT et al. (1996). CLARK JÚNIOR (1979), em revisão sobre o
potencial uso de pesos das aves, comprova a relevância para várias áreas de
conhecimento de uma coletânea com dados sobre massa corpórea, onde tais informações
podem ser utilizadas como padrões para determinar o tamanho da ave, avaliar condições
fisiológicas, uso nas análises de comunidades ecológicas e na sistemática. Por outro
lado, REINERT et al. (1996) ressaltam a importância dessas informações principalmente
para aves endêmicas da Mata Atlântica, que ainda são pobremente conhecidas. Os
achados que comprovam diferenças entre pesos de machos e fêmeas, a exemplo da
espécie Arremon taciturnus, corroboram com a literatura pesquisada. BALDWIN &

36
KENDEIGH (1938) e CLARK JÚNIOR (1979) registraram essas desigualdades e comprovam
que a relação não está distribuída de maneira constante. Este registro foi reforçado uma
vez que duas das três espécies analisadas não apresentaram o mesmo padrão, não sendo
significativa a variação de peso entre sexos. Quanto às comparações entre massa
corpórea e estações do ano, embora não tenham existido valores significativos, variações
interespecíficas são comuns, além das alterações individuais ao longo do tempo
(BALDWIN & KENDEIGH 1938). Ou seja, as mudanças de peso ocorrem tanto dentro da
mesma espécie quanto no indivíduo, variando ao longo do dia e das estações do ano e
está relacionada a outras variáveis, tais como: diferenças na temperatura do ar,
quantidade de alimentação, migração e outras atividades, não sendo possível estabelecer
um padrão geral. A maneira e extensão de flutuação de peso observada evidenciam que
as aves em seu ajustamento fisiológico são altamente sensíveis a influências ambientais,
e que esta inter-relação entre função e ambiente afeta extremamente esse comportamento
(BALDWIN & KENDEIGH op cit.).
Por outro lado, novas correlações devem ser testadas, além de análises de massa
corpórea e diferenças entre sexos e podem contribuir para responder a presença de
fatores biológicos ocultos. Isto é, diferenças observadas entre medidas de alguns
caracteres, como comprimento de asa ou tarso entre sexos, podem sugerir um novo
método para determinar dimorfismo sexual em espécies que não apresentam essa
distinção fenotípica aparente.
Para as observações de mudas, o presente estudo confirma aquelas informações
encontradas na literatura quanto à simultaneidade entre mudas de rêmiges e retrizes
(POULIN et al. 1992, SICK 1997, MARINI & DURÃES 2001). As penas de contorno não
seguem esse padrão, fato que corrobora com MARINI & DURÃES op cit. em estudos sobre

37
a bionomia de aves da Mata Atlântica. O grande percentual de mudas de contorno
encontrado em comparação com as mudas de vôo faz crer que o critério empregado,
utilizando apenas três penas em crescimento, em quaisquer regiões do corpo (cabeça,
dorso e/ou ventre) para afirmar a existência de muda pós-nupcial, pode mascarar a
análise em questão. Entretanto, MARINI & DURÃES (2001) aumentaram esse critério para
observações de no mínimo dez penas em crescimento em uma ou mais regiões do corpo,
encontrando mais de 15% de aves repondo-as durante os meses de estação chuvosa ou
seca. Com isso concluíram que este tipo de muda ocorre ao longo de todo o ano,
propondo que mudas de contorno não estejam necessariamente relacionadas com a muda
pós-nupcial. Apesar das diferenças metodológicas, fato semelhante foi encontrado no
presente estudo. Sugere-se, portanto, o emprego apenas das penas primárias para avaliar
o ciclo de muda pós-nupcial conforme recomendado por MILLER (1961) e POULIN et al.
(1992).
No ciclo anual das aves, reprodução e atividade de mudas são eventos que
demandam grande energia, levando vários autores a apontar a ausência de
simultaneidade entre mudas primárias e condições reprodutivas em regiões temperadas
(MILLER 1961, PAYNE 1969, POULIN et al. 1992). Para aves tropicais ao menos,
sobreposição de mudas e período reprodutivo é mais comum do que geralmente suposto,
devido ao prolongado período reprodutivo nos trópicos (FOSTER 1975), e ocorre
geralmente em uma fase de alta oferta de alimento (CRUZ & ANDREWS 1989, POULIN et
al. 1992). Neste trabalho não foi possível comprovar estatisticamente a sobreposição das
mudas, possivelmente devido à reduzida quantidade de espécimes capturados. Todavia,
uma correlação negativa entre retriz e placa de incubação e rêmige e placa de incubação
foi observada, o que sugere estar de acordo com os achados dos autores acima citados.

38
Em regiões tropicais POULIN et al. (1992) e MARINI & DURÃES (2001) afirmam
que o ciclo do ano com grande disponibilidade de alimento é no fim da estação seca e na
estação chuvosa. POULIN et al. op cit. demonstraram, em florestas tropicais da
Venezuela, que durante a estação reprodutiva das aves, houve um aumento no consumo
de artrópodes em detrimento a ingestão de frutos, o que sugere que a abundância desses
invertebrados é um fator crucial para governar a velocidade das atividades reprodutivas,
nivelando as espécies que normalmente incluem alta proporção de néctar e frutas em sua
dieta. Igualmente a este padrão, tanto as mudas quanto as placas de incubação estiveram
mais freqüentes no final da estação seca, início da estação chuvosa para este trabalho.
MARINI & DURÃES (2001) afirmam que padrões climáticos regionais no sudeste
do Brasil, em lugar de variações locais, são mais prováveis para determinar os períodos
reprodutivos e mudas em pássaros florestais. Algumas variações ocorreram em 2004
quanto aos índices pluviométricos, mas atividades de muda e reprodução, seguiram o
padrão regional.
Quanto a hábitos alimentares SICK (1997) classifica os piprídeos (Manacus
manacus e Neopelma pallescens) como frugívoros/insetívoros e o Turdus leucomelas
como onívoro, o que os torna bastante generalistas, fato que, aliado a outros hábitos e
características, amplia sua representatividade no ambiente. A estrutura da comunidade
de aves é drasticamente alterada em fragmentos menores quando se observa, por
exemplo, que a representatividade de insetívoros especializados cuja captura em rede de
neblina é significativa para áreas contínuas, reduz dramaticamente (BIERREGAARD et al.
1992). Com o passar do tempo a maioria dessas espécies insetívoras presentes nesses
fragmentos, que forrageiam em conjunto com outras espécies, desaparece em fragmentos
menores de 10 hectares (POWELL 1985, BIERREGAARD & LOVEJOY 1989).

39
A presença constante e fortemente indicativa de aumento das capturas no período
de maior oferta alimentar pela espécie Basileuterus flaveolus sugere explicação para a
alta relação observada frente aos dados abióticos. Esta espécie insetívora procura seu
alimento de preferência no solo ou muito baixo na ramagem, estando de acordo com o
observado por SICK (1997). Neste período, percebeu-se também uma quantidade maior
de insetos, possivelmente aqueles que necessitam de água para completarem o seu ciclo
de vida, fato similar aos achados de POULIN et al. (1992), que apresentam resultados em
relação a maior abundância de artrópodes na estação chuvosa, onde as aves seguem o
mesmo padrão, independente da guilda, mantendo predominantemente seus períodos de
muda e reprodução nas estações úmidas, estando em sincronia com a alta abundância de
recursos alimentares. Isto pode explicar a relação existente entre esta espécie e os
padrões de precipitação e umidade, embora LEFEBVRE et al. (1992) afirmem que, para
regiões tropicais, a disponibilidade de insetos é relativamente constante ao longo do ano.
Por outro lado, a espécie da família Pipridae (Manacus manacus) apresentou
maior relação com a temperatura. Isso pode ser explicado quando se consideram
períodos mais quentes que antecedem a época reprodutiva. Estas aves utilizam
comumente poleiros de exibição para atrair as fêmeas, ou seja, complexos
comportamentos de exibição pré-nupcial, assim como observado por SICK (1997) e
SILVA & RÊGO (2004), facilitando a sua visualização e o seu deslocamento, aumentando
a probabilidade de captura. Este fator também pode estar relacionado com a frutificação
que ocorre no período após as chuvas, a exemplo das melastomatáceas que se
apresentam abundantes nas bordas da mata e são muito utilizadas por pequenos
frugívoros, como os Pipridae (GALETTI & STOTZ 1996).

40
São escassas as informações acerca das espécies Touit surda e Ornithion inerme
no Estado de Pernambuco. O único registro de pele do T. surda data de dezembro de
1944, quando BERLA (1946 e 1954) apud TELINO JÚNIOR et al. (2000) coletou três
exemplares da espécie, nos arredores do Recife. Entre setembro de 1996 a agosto de
1997, estudos realizados no RECD acrescentaram informações acerca da distribuição
geográfica, biologia e história natural do Touit surda no Estado através de observação,
gerando sugestões para estratégias de conservação da espécie (TELINO JÚNIOR et al. op
cit.). Naquela ocasião foram observados bandos de até 40 indivíduos sobrevoando a área
à procura de alimento e abrigo, quase sempre identificados pela vocalização. Estas
informações se assemelham com as observações contidas nesta pesquisa quanto à
identificação por vocalização, entretanto bandos menores com aproximadamente 12
indivíduos foram avistados sobrevoando a área, demonstrando utilizá-la naquele
momento, ao menos, como ambiente de transição.
Quanto aos hábitos alimentares do T. surda, embora a oferta máxima do sapoti
tenha ocorrido apenas durante a primeira captura, o fruto verde parece atrair essas aves,
fato observado por TELINO JÚNIOR et al. (2000), quando apresentam o incremento de
itens alimentares para a espécie no Estado e citam frutos verdes e maduros como um
atrativo para a espécie. Os primeiros, ao serem capturados juntos demonstraram a
formação de casal, fato confirmado quando realizado a taxidermia. Isto sugere que tal
espécie, apesar de conviver em bando e mais freqüentemente na copa das árvores
(TELINO JÚNIOR et al. op cit.), pode ainda apresentar hábitos de forrageamento mais
restrito a pequenos grupos, chegando a atingir estratos inferiores da floresta em busca de
alimento. SICK (1997) relata esse procedimento para várias espécies da família dos
Psittacidae, que eventualmente chegam até ao solo atrás de cocos ou sementes caídos.

41
Para Pernambuco o primeiro registro do Ornithion inerme data de 2002. José
Fernando Pacheco e Ricardo Parrini relataram a observação de um único exemplar desta
espécie, identificado pela vocalização característica, em pleno forrageamento, na
Reserva Ecológica de Dois Irmãos, Recife, ampliando a listagem de aves para o Parque
(AZEVEDO-JÚNIOR et al. 1998, FARIAS et al. 2002). Até então, era assinalado apenas para
regiões fronteiriças com Alagoas e Paraíba (FARIAS et al. op cit.).
MORELLATO & HADDAD (2000) sugerem que as pesquisas na Floresta Tropical e
especialmente na Floresta Atlântica Brasileira, devem requerer maior coordenação e
direção no sentido de conduzir efetivamente a estudos sobre conservação, manejo
racional e políticas públicas, do contrário a degradação causará transformações
irreversíveis para a estrutura das comunidades em geral.
O efeito de borda, por exemplo, promove modificações na estrutura da floresta
causando alterações na composição de marsupiais, roedores, répteis e anfíbios
(LAURANCE 1991), exercendo uma pressão de predação sobre ovos e filhotes de muitas
aves o que pode causar a diminuição do sucesso reprodutivo das mesmas nestes locais
(TABARELLI & MANTOVANI 1997), conforme observado para o Trogon curucui no
ambiente estudado. A importância desta redução está diretamente relacionada ao
tamanho do fragmento e à área ocupada pela borda (TABARELLI & MANTOVANI op cit.).
A floresta que compõe o fragmento alvo do presente estudo, mantém-se como
um remanescente de floresta em decorrência da ação depredadora do homem no
passado. Entretanto, foi possível observar sua busca pela recomposição, mesmo com as
áreas vizinhas sofrendo pressão antrópica para servir a agropecuária. Esta recuperação se
deve às medidas conservacionistas desenvolvidas na região (LYRA-NEVES et al. 2000).
Áreas de floresta atlântica circunvizinhas ao RECD, como exemplo da Usina São José,

42
com estimados 323 ha considerados como Reserva Ecológica associados a outros 5.000
ha de mata fragmentada (TELINO-JÚNIOR et al. 2000), certamente contribuem com a
existência de muitas espécies que sobrevivem em pequenos fragmentos interligados a
plantações, pastagens e a outras pequenas áreas florestais, promovendo a ampliação do
uso de recurso por parte desses animais (ALEIXO & GALETTI 1997; TELINO-JÚNIOR et al.
op cit.), servindo de corredores ecológicos.
TERBORGH (1992) sugere como forma de amenizar os efeitos da fragmentação,
um adequado uso da terra com prioridades de planejamento. O desenvolvimento de
atividades que visem melhorar a qualidade de ambientes degradados deve ser empregado
para cada área atingida de forma específica, onde se torna necessário uma análise da
problemática em particular da região para que seja possível viabilizar medidas a serem
aplicadas para a condução de ações efetivas de preservação e conservação dos recursos
naturais locais.
Todos os fatores observados indicam que o emprego do método de captura em
rede para acrescentar informações biológicas sobre as aves, apresenta algumas respostas,
sendo suficiente para pesquisas pontuais. Comprova, através dos dados obtidos, a
importância do RECD para a manutenção das aves presentes nesse ecossistema.
A ampliação do montante de dados referentes à composição da avifauna e sua
manutenção em fragmentos florestais contribuem para o planejamento de estratégias de
conservação dessas áreas que são, em muitos casos, os últimos redutos para a fauna do
local.

43
6. CONSIDERAÇÕES FINAIS
► Embora as capturas realizadas tenham sido efetuadas por amostragem, a presença de
espécies ameaçadas de extinção, bem como a ocorrência de espécies até o momento com
registro em apenas uma área em todo o Estado de Pernambuco ou ainda a presença de
espécies da família Psittacidae, que apresentam dieta mais especializada e, portanto,
maior necessidade de áreas extensas de vegetação cujo suprimento de frutos esteja
disponível ao longo do ano, comprova a importância do fragmento na conservação da
diversidade biológica da área;
► Apesar de não ter sido comprovada a sobreposição entre mudas e placa de incubação
no presente estudo, possivelmente devido ao baixo número de capturas (considerando
esta análise), a correlação negativa entre retriz vs placa de incubação e rêmige vs placa
de incubação sugerem que devido ao período reprodutivo prolongado nos trópicos e a
presença das placas de incubação no final da estação seca e início da estação chuvosa,
haja uma possível sobreposição destas atividades, embora essa proporção seja
provavelmente considerada baixa;
► Pesquisas em ambientes alterados, principalmente aqueles de tamanho reduzido, são
de extrema importância para comprovar a sustentabilidade das espécies, gerando a
necessidade de maior coordenação e direção no sentido de conduzir efetivamente a
conservação e o manejo racional através do desenvolvimento de políticas públicas que
visem estabelecer planos de manejo que gerenciem essas áreas e regiões afins, evitando

44
que tamanha degradação cause transformações irreversíveis para a estrutura das
comunidades em geral.

45
7. REFERÊNCIAS BIBLIOGRÁFICAS
ALEIXO, A. & J. M. E. VIELLIARD. 1995. Composição e dinâmica da avifauna da mata de
Santa Genebra, Campinas, São Paulo, Brasil. Revista Brasileira de Zoologia, São
Paulo, 12(3):493-511.
ALEIXO, A. & M. GALETTI. 1997. The conservation of the avifauna in a lowland Atlantic
forest in south-east Brazil. Bird Conservation International, Cambridge, 7:235-
261.
ALMEIDA, M. E. DE C.; J. M. E. VIELLIARD; & M. M. DIAS. 1999. Composição da
avifauna em duas matas ciliares na bacia do rio Jacaré-Pepira, São Paulo, Brasil.
Revista Brasileira de Zoologia, São Paulo, 16(4):1087-1098.
ANCIÃES, M. & M. Â. MARINI. 2000a. Assimetria flutuante em passeriformes da mata
Atlântica. pp. 187-204. Em: Maria Alice dos Santos Alves, José Maria Cardoso da
Silva, Monique Van Sluys, Helena de Godoy Bergallo & Carlos Frederico Duarte da
Rocha (Orgs.) A ornitologia no Brasil: pesquisa atual e perspectivas. Rio de Janeiro:
EdUERJ.
ANCIÃES, M. & M. Â. MARINI. 2000b. The effects of fragmentation on fluctuating
asymmetry in passerine birds of Brazilian tropical forests. Journal of Applied
Ecology, Oxford, 37:1013-1028.
ANDRADE-LIMA, D. DE. 1960. Estudos fitogeográficos de Pernambuco. Arquivos do
Instituto de Pesquisas Agronômicas, Recife, 5:305-341.
ANDRADE-LIMA, D. DE. 1961. Tipos de floresta de Pernambuco. In XII Anais da
Associação dos Geógrafos Brasileiros. São Paulo. 69-85.

46
AVERY, M. L. 1985. Annual molt pattern in a Malaysian population of fantail warblers
(Cisticola juncidis). The Condor, Los Angeles, 87:346-349.
AZEVEDO-JÚNIOR, S. M. de. 1990. A Estação Ecológica do Tapacurá e suas aves. Anais
do Encontro Nacional de Anilhadores de Aves, Recife, 4:92-99.
AZEVEDO-JÚNIOR, S. M. DE; A. G. M. COELHO; M. E. DE LARRAZÁBAL; R. M. DE LIRA
NEVES & W. R. TELINO JÚNIOR. 1998. Conservação e diversidade das aves da
Reserva Ecológica de Dois Irmãos. pp. 241-250. Em: Isabel Cristina Machado,
Ariadna Valentina Lopes & Kátia Cavalcanti Pôrto (Orgs.) Reserva Ecológica de
Dois Irmãos: estudos em um remanescente de Mata Atlântica em área urbana
(Recife, Pernambuco, Brasil). Recife: Secretaria de Ciência, Tecnologia e Meio
Ambiente.
AZEVEDO-JÚNIOR, S. M. DE.; M. M. DIAS; M. E. DE; W. R. TELINO-JÚNIOR, R. M. DE
LYRA-NEVES & C. J. G. FERNANDES. 2001. Recapturas e recuperações de aves
migratórias no litoral de Pernambuco, Brasil. Ararajuba, Rio de Janeiro, 9(1):23-42.
BALDWIN, P. & C. KENDEIGH. 1938. Variations in the weight of birds. Auk, Lawrence,
55:416-467.
BELTON, W. 1994. Aves do Rio Grande do Sul: distribuição e biologia. São Leopoldo:
Ed. UNISINOS, 584p.
BIERREGAARD JR., R. O. & LOVEJOY, T. E. 1989. Effects of forest fragmentation on
Amazonia understory bird communities. Acta Amazonica, Manaus, 19:215-241.
BIERREGAARD JR., R. O.; T. E. LOVEJOY; V. KAPOS; A. A. DOS SANTOS & R. W.
HUTCHINGS. 1992. The biological dynamics of tropical rainforest fragments: a
prospective comparison of fragments and continuous forest. BioScience,
Washington, 42(11):859-866.

47
BROWN, K. S. JR. & G. G. BROWN. 1992. Habitat alteration and species loss in Brazilian
forests, p. 119-142. In: T. C. Whitmore and J. A. Sayer (Eds.). Tropical
deforestation and species extinction. Chapman and Hall, London, England.
CENTRO DE PESQUISAS PARA CONSERVAÇÃO DAS AVES SILVESTRES.
1994. Manual de anilhamento de aves silvestres. 2 ed., Brasília: IBAMA, 146p.
CHIARELLO, A.G. 1999. Effects of fragmentation of the Atlantic forest on mammal
communities in south-eastern Brazil. Biol. Cons, Essex, 89: 71-82.
CLARK JÚNIOR, G. A. 1979. Body weights of birds: a review. The Condor, Los Angeles,
81:193-202.
CRUZ, A. & R. W. ANDREWS. 1989. Observations on the breeding of passerines in a
seasonally flooded savanna in Venezuela. Wilson Bull, Lawrence, 101(1):62-76.
DÁRIO, F. R.; M. C. V. DE VICENZO, & A. F. DE ALMEIDA. 2002. Avifauna em fragmentos
da Mata Atlântica. Ciência Rural, Santa Maria, 32(6):989-996.
DUNNING, J. S. 1987. South America birds: a photographic aid to identification.
Pennsylvania, Harrowood Books, 364p.
ELDER, W. H. & D. ZIMMERMAN. 1983. A comparison of recapture versus resighting data
in a 15-year study of survivorship of the black-capped chickadee. J. Field Ornithol,
54(2): 138-145.
FARIAS, G. B. DE.; M. T. DE BRITO & G. L. PACHECO. 2002. Registros ornitológicos de
Pernambuco. Observadores de Aves de Pernambuco, Recife, 1-67.
FIDEM, FUNDAÇÃO PARA O DESENVOLVIMENTO DA REGIÃO METROPOLITANA DO
RECIFE. 1993. Monitoramento das reservas ecológicas da região metropolitana
do Recife. Governo do Estado de Pernambuco, Secretaria de Planejamento do

48
Estado de Pernambuco, Fundação de Desenvolvimento da Região Metropolitana do
Recife, Recife, 55p.
FONSECA, G. A. B. DA. 1985. The vanishing Brazilian Atlantic Forest. Biological
Conservation, Essex, 34: 17-34.
FOSTER, M. S. 1975. The overlap of molting and breeding in some tropical birds. The
Condor, Los Angeles, 77:304-314.
GALETTI, M. & D. STOTZ. 1996. Miconia hypoleuca (Melastomataceae) como espécie-
chave para aves frugívoras no sudoeste do Brasil. Revista Brasileira de Zoologia,
São Paulo, 56(2):435-439.
GONZAGA DE CAMPOS. 1912. Mappe florestal. Rio de Janeiro, Serviço Geológico de
Mineralogia do Brasil.
IBAMA. 2003. Ecossistemas brasileiros [on line] URL: http://www.ibama.org.br
INMET - INSTITUTO NACIONAL DE METEOROLOGIA – 2004. Seção de Observação e
Meteorologia Aplicada – SEOMA.
LAURANCE, W. F. 1991. Edges effects in tropical forest fragments: application of a
model for the design of nature reserves. Biological Conservation, Essex, 57:205-
219.
LEFEBVRE, G.; B. POULIN & R. MCNEIL. 1992. Settlement period and function of long-
term territory in tropical mangrove passerines. The Condor, Los Angeles, 94:83-92.
LIMA, M. L. F. DA C. 1998. A reserva da Biosfera da Mata Atlântica em Pernambuco -
situação atual, ações e perspectivas. Caderno n. 12. Instituto Florestal, São Paulo,
43p.
LOPES, O. DE S.; L. DE A. SACCHETTA & E. DENTE. 1980. Longevity of wild birds
obtained during a banding program in SP, Brasil. J. Field Ornithol. 51(2):144-148.

49
LYRA-NEVES, R. M. 1998. Estudo de ectoparasitos de aves silvestres (Passeriformes:
Emberizidae) de Mata Atlântica, Igarassu, Pernambuco. Dissertação em Biologia
Animal do Centro de Ciências Biológicas. Universidade Federal de Pernambuco:
Recife. 70p.
LYRA-NEVES, R. M.; Ã. M. I. DE FARIAS; W. R. TELINO-JÚNIOR; M. ARZUA; M. C. N.
BOTELHO & M. DA C. ABREU E LIMA. 2000. Ectoparasitismo em aves silvestres
(Passeriformes – Emberizidae) de Mata Atlântica, Igarassu, Pernambuco.
Melopsittacus, Belo Horizonte, 3(2):64-71.
LYRA-NEVES, R. M. DE; M. M. DIAS; S. M. DE AZEVEDO-JÚNIOR; W. R. TELINO-JÚNIOR
& M. E. L. DE LARRAZÁBAL. 2004. Comunidade de aves da Reserva de Gurjaú,
Pernambuco, Brasil. Revista Brasileira de Zoologia, Londrina, 21(3):581-592.
MALDONADO-COELHO, M. & M. Â. MARINI. 2000. Effects of forest fragment size and
successional stage on mixed-species bird flocks in southeastern Brazil. The Condor,
Los Angeles, 102:585-594.
MARINI, M. A. 1998. Efeitos da fragmentação florestal sobre as aves em Minas Gerais.
Em: Maria Alice dos Santos Alves, José Maria Cardoso da Silva, Monique Van
Sluys, Helena de Godoy Bergallo & Carlos Frederico Duarte da Rocha (Orgs.) A
Ornitologia no Brasil: pesquisa atual e perspectivas. EdUERJ, Rio de Janeiro,
352p.
MARINI, M. Â & R. DURÃES. 2001. Annual patterns of molt and reproductive activity of
passerines in south-central Brazil. The Condor, Los Angeles, 103:767-775.
MCNEIL, R. 1971. Lean-season fat in a South American population of Black-necked
stilts. Condor, Los Angeles, 73:472-475.

50
MILLER, A. H. 1961. Molt cycles in Equatorial Andean sparrows. The Condor, Los
Angeles, 63:143-161.
MORELLATO, L. P. C. & C. F. B. HADDAD. 2000. Introduction: the Brazilian Atlantic
forest. Biotropica, Washington, 32(4b):786-792.
MYERS, N.; R. A. MITTERMEIER; C. G. MITTERMEIER; G. A. B. FONSECA & J. KENT.
2000. Biodiversity hotspots for conservation priorities. Nature, London, 403:853-
858.
ONIKI, Y. 1981. Weights, cloacal temperatures, plumage and molt condition of birds in
the state of São Paulo. Rev. Brasil. Biol., Rio de Janeiro, 41(2):451-460.
ONIKI, Y. & E. O. WILLIS. 1993. Pesos, medidas, mudas, temperaturas cloacais e
ectoparasitos de aves da Reserva Ecológica do Panga, Minas Gerais, Brasil. Bol.
CEO. 9:2-10.
PARKER III, T. A.; D. E. STOTZ & J. W. FITZPATRICK. 1996. Ecological and distributional
databases, p. 113-436. In: D. F. STOTZ; J. W. FITZPATRICK; T. A. PARKER III & D. K.
MOSKOVITS (Eds.). Neotropical birds: ecology and consertation. Chicago,
University of Chicago Press, XI+700p.
PAYNE, R. B. 1969. Overlap of breeding and molting schedules in a collection of African
birds. The Condor, Los Angeles, 71:140-145.
PAYNE, R. B. & L. L. PAYNE. 1990. Survival estimates of indigo buntings: comparison of
banding recoveries and local observations. The Condor, Los Angeles, 92:938-946.
POULIN, B.; G. LEFEBVRE & R. MCNEIL. 1992. Tropical avian phenology in relation to
abundance and exploitation of food resources. Ecology, Durham, 73(6):2295-2309.
POWELL, G. V. N. 1985. Sociobiology and adaptive significance of interspecific foraging
flock in the neotropics. p.713-732. In: P. A. Buckley, M. S. Foster, E. S. Morton, R.

51
S. Ridgely, and F. G. Buckley (Eds.). Neotropical Ornithology. American
Ornithologists Union, Washington, DC.
RANTA, P.; T. BLOM; J. NIEMELÃ; E. JOENSUU & M. SIITONEN. 1998. The fragmented
Atlantic rain forest of Brazil: size, shape and distribution of forest fragments.
Biodiversity and Conservation, London, 7:385-403.
REINERT, B. L., J. C. PINTO, BORNSCHEIN, M. R., PICHORIM & MARINI, M. Â. 1996. Body
masses and measurements of birds from Southern Atlantic Forest, Brazil. Revista
Brasileira de Zoologia, São Paulo, 13(4):815-824.
RIDGELY, R. S. & G. TUDOR. 1994a. The birds of South America: the oscines
passerines. 2a ed., Austin, Univ. Texas Press, 516p.
RIDGELY, R. S. & G. TUDOR. 1994b. The birds of South America: the suboscines
passerines. Austin, Univ. Texas Press, 814p.
SICK, H. 1997. Ornitologia Brasileira. Edição revista e ampliada por José Fernando
Pacheco. Rio de Janeiro, Editora Nova Fronteira, II+862p.
SILVA, W. A. DE G. & P. S. DO RÊGO. 2004. Conservação do soldadinho-do-araripe
Antilophia bokermanni (Aves: Pipridae): subsídios para a elaboração do plano de
manejo. Observadores de Aves de Pernambuco, Curitiba, Fundação O Boticário
de Proteção à Natureza, 32p.
SILVA, J. M. C. & M. TABARELLI. 2000. Tree species impoverishment and the future flora
of the Atlantic Forest of Northeast Brazil. Nature, London, 404: 72-74.
SNOW, D. W. & A. LILL. 1974. Longevity records for some neotropical land birds. The
Condor, Los Angeles, 76:262-267.
SOUZA, D. G. S. 2002. All the birds of Brazil: an identification guide. Bahia, Editora
DALL, 356p.

52
TABARELLI, M. 2000. Dois Irmãos: o desafio da conservação biológica em um
fragmento de floresta tropical. In: Isabel Cristina Machado, Ariadna Valentina
Lopes & Kátia Cavalcanti Pôrto (Orgs.) Reserva Ecológica de Dois Irmãos: Estudos
em um Remanescente de Mata Atlântica em Área Urbana (Recife – Pernambuco –
Brasil). Editora Universitária, SECTMA. pp.311-323. Recife, PE.
TABARELLI, M. & W. MANTOVANI. 1997. Predação de ovos e remoção de propágulos em
um fragmento de floresta atlântica, Espírito Santo, Brasil. Rev. Brasil. 57(4):699-
707.
TELINO-JÚNIOR, W. R.; R. M. DE LYRA-NEVES & R. S. CARNEIRO. 2000. Observações de
Touit surda (Psittacidae) em fragmentos florestais de Pernambuco, Brasil.
Melopsittacus, Belo Horizonte, 3(4):159-165.
TERBORGH, J. 1992. Maintenance of the diversity in tropical forests. Biotropica,
Washington, 24:283-292.
TERBORGH, J.; S. K. ROBINSON;. T. A. PARKER III; C. A. MUNN & N. PIERPONT. 1990.
Structure and organization of an Amazonian forest bird community. Ecological
Monographs, Washington, 60 (2): 213-238.
VIANA, V. M.; A. A. J. TABANEZ & J. L. F. BATISTA. 1997. Dynamics and restoration of
forest fragments in the Brazilian Atlantic Moist Forest. In: Laurance, W. F. and
Bierregaard, R. O. (Eds.). Tropical forest remnants: ecology, management and
conservation of fragmented communities. University of Chicago Press, Chicago.
WILLIS, E. O. 1979. The composition of avian communities in remanescent woodlots in
southern Brazil. Papéis Avulsos Zool., São Paulo, 33(1):1-25.
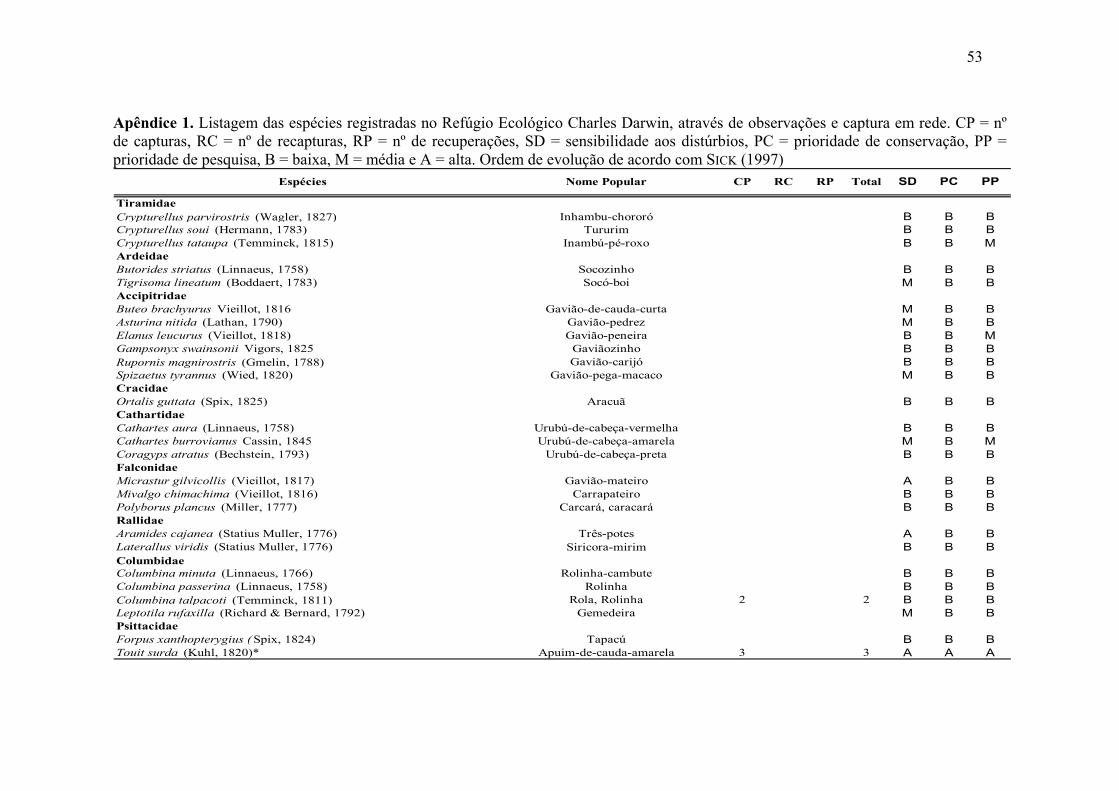
53
Apêndice 1. Listagem das espécies registradas no Refúgio Ecológico Charles Darwin, através de observações e captura em rede. CP = nº de capturas, RC = nº de recapturas, RP = nº de recuperações, SD = sensibilidade aos distúrbios, PC = prioridade de conservação, PP = prioridade de pesquisa, B = baixa, M = média e A = alta. Ordem de evolução de acordo com SICK (1997)
TiramidaeCrypturellus parvirostris (Wagler, 1827) Inhambu-chororó B B BCrypturellus soui (Hermann, 1783) Tururim B B BCrypturellus tataupa (Temminck, 1815) Inambú-pé-roxo B B MArdeidaeButorides striatus (Linnaeus, 1758) Socozinho B B BTigrisoma lineatum (Boddaert, 1783) Socó-boi M B BAccipitridaeButeo brachyurus Vieillot, 1816 Gavião-de-cauda-curta M B BAsturina nitida (Lathan, 1790) Gavião-pedrez M B BElanus leucurus (Vieillot, 1818) Gavião-peneira B B MGampsonyx swainsonii Vigors, 1825 Gaviãozinho B B BRupornis magnirostris (Gmelin, 1788) Gavião-carijó B B BSpizaetus tyrannus (Wied, 1820) Gavião-pega-macaco M B BCracidaeOrtalis guttata (Spix, 1825) Aracuã B B BCathartidaeCathartes aura (Linnaeus, 1758) Urubú-de-cabeça-vermelha B B BCathartes burrovianus Cassin, 1845 Urubú-de-cabeça-amarela M B MCoragyps atratus (Bechstein, 1793) Urubú-de-cabeça-preta B B BFalconidaeMicrastur gilvicollis (Vieillot, 1817) Gavião-mateiro A B BMivalgo chimachima (Vieillot, 1816) Carrapateiro B B BPolyborus plancus (Miller, 1777) Carcará, caracará B B BRallidaeAramides cajanea (Statius Muller, 1776) Três-potes A B BLaterallus viridis (Statius Muller, 1776) Siricora-mirim B B BColumbidaeColumbina minuta (Linnaeus, 1766) Rolinha-cambute B B BColumbina passerina (Linnaeus, 1758) Rolinha B B BColumbina talpacoti (Temminck, 1811) Rola, Rolinha 2 2 B B BLeptotila rufaxilla (Richard & Bernard, 1792) Gemedeira M B BPsittacidaeForpus xanthopterygius ( Spix, 1824) Tapacú B B BTouit surda (Kuhl, 1820)* Apuim-de-cauda-amarela 3 3 A A A
SD PC PPEspécies Nome Popular CP RC RP Total
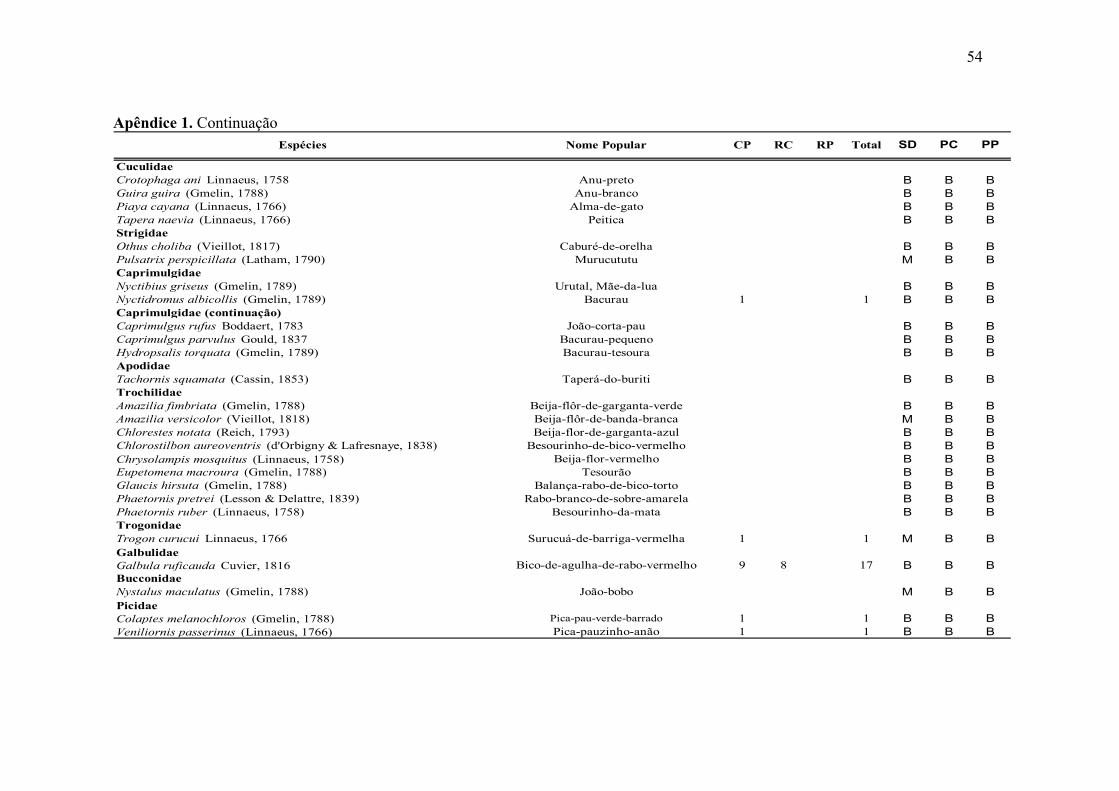
54
Apêndice 1. Continuação
CuculidaeCrotophaga ani Linnaeus, 1758 Anu-preto B B BGuira guira (Gmelin, 1788) Anu-branco B B BPiaya cayana (Linnaeus, 1766) Alma-de-gato B B BTapera naevia (Linnaeus, 1766) Peitica B B BStrigidaeOthus choliba (Vieillot, 1817) Caburé-de-orelha B B BPulsatrix perspicillata (Latham, 1790) Murucututu M B BCaprimulgidaeNyctibius griseus (Gmelin, 1789) Urutal, Mãe-da-lua B B BNyctidromus albicollis (Gmelin, 1789) Bacurau 1 1 B B BCaprimulgidae (continuação)Caprimulgus rufus Boddaert, 1783 João-corta-pau B B BCaprimulgus parvulus Gould, 1837 Bacurau-pequeno B B BHydropsalis torquata (Gmelin, 1789) Bacurau-tesoura B B BApodidaeTachornis squamata (Cassin, 1853) Taperá-do-buriti B B BTrochilidaeAmazilia fimbriata (Gmelin, 1788) Beija-flôr-de-garganta-verde B B BAmazilia versicolor (Vieillot, 1818) Beija-flôr-de-banda-branca M B BChlorestes notata (Reich, 1793) Beija-flor-de-garganta-azul B B BChlorostilbon aureoventris (d'Orbigny & Lafresnaye, 1838) Besourinho-de-bico-vermelho B B BChrysolampis mosquitus (Linnaeus, 1758) Beija-flor-vermelho B B BEupetomena macroura (Gmelin, 1788) Tesourão B B BGlaucis hirsuta (Gmelin, 1788) Balança-rabo-de-bico-torto B B BPhaetornis pretrei (Lesson & Delattre, 1839) Rabo-branco-de-sobre-amarela B B BPhaetornis ruber (Linnaeus, 1758) Besourinho-da-mata B B BTrogonidaeTrogon curucui Linnaeus, 1766 Surucuá-de-barriga-vermelha 1 1 M B BGalbulidaeGalbula ruficauda Cuvier, 1816 Bico-de-agulha-de-rabo-vermelho 9 8 17 B B BBucconidaeNystalus maculatus (Gmelin, 1788) João-bobo M B BPicidaeColaptes melanochloros (Gmelin, 1788) Pica-pau-verde-barrado 1 1 B B BVeniliornis passerinus (Linnaeus, 1766) Pica-pauzinho-anão 1 1 B B B
Nome Popular CP RC RPEspécies Total SD PC PP
55
Apêndice 1. Continuação
Picidae (continuação)Picumnus cirratus Temminck, 1825 Pica-pau-anão-barrado B B MPicumnus exilis (Lichtenstein, 1823)** Pica-pau-anão-de-pintas-amarelas M B BThamnophilidaeDysithamnus mentalis (Temminck, 1823) Choquinha-lisa M B BFormicivora grisea (Boddaert, 1783) Papa-formigas-pardo 8 1 9 B B BHerpsilochmus atricapillus Pelzeln, 1868 Chorozinho-de-chapéu-preto 5 1 6 M B BHerpsilochmus rufomarginatus (Temminck, 1822) Chorozinho-de-asa-vermelha M B BMyrmotherula axillaris (Vieillot, 1817) Choquinha-de-flancos-brancos M B BTaraba major (Vieillot, 1816) Choró-boi B B BThamnophilus caerulescens Vieillot, 1816** Choca-da-mata 7 1 1 9 B B MThamnophilus paliatus (Lichtenstein, 1823) Choca-listrada B B MConopophagidaeConopophaga lineata (Wied, 1831)** Chupa-dente 1 1 2 M A MFurnariidaeCerthiaxis cinnamomeus (Gmelin, 1788) Corruíra-do-brejo M B BPhacellodomus rufifrons (Wied, 1821) João-graveto M B BSynallaxis frontalis Pelzeln,1859 Tio-antônio B B BSynallaxis scutata Sclater, 1859 Viu-vi M M MXenops minutus (Sparrman, 1788)** Bico-virado-miúdo 7 3 10 M B BXenops rutilans Temminck, 1821 Bico-virado-carijó M B BDendrocolaptidaeSittasomus griseicapillus (Vieillot, 1818) Arapaçu-verde 6 4 10 M B MXiphorhynchus picus (Gmelin, 1788) Arapaçu-de-bico-branco 5 1 1 7 B B BTyrannidaeCamptostoma obsoletum (Temminck, 1824) Risadinha 2 2 B B MCapsiemps flaveola ( Lichtenstein, 1823) Marianinha-amarela 2 2 B B BElaenia cristata Pelzeln, 1868 Guaracava-de-topete-uniforme 1 1 M B BElaenia flavogaster (Thunberg, 1822) Guaracava-de-barriga-amarela 11 1 12 B B BLeptopogon amaurocephalus Tschudi, 1846 Cabeçudo 9 14 23 M B BMionectes oleagineus (Lichtenstein, 1823) Abre-asas 5 5 M B BMyiozetetes similis (Spix, 1825) Bentevizinho-penacho-vermelho 3 3 B B BOrnithion inerme Hartlaub, 1853 Poaieiro-de-sobrancelha 2 2 M B BPachyramphus polychopterus (Vieillot, 1818) Caneleiro-preto 1 1 B B BPachyramphus viridis (Vieillot, 1816) Caneleiro-verde 2 2 M B MPhyllomyias fasciatus (Thunberg, 1822) Piolhinho M B M
Espécies Nome Popular CP RC RP Total SD PC PP

56
Apêndice 1. Continuação
Tyrannidae (continuação)Pitangus sulphuratus (Linnaeus, 1766) Bentevi 1 1 B B BTolmomyias flaviventris (Wied, 1831) Bico-chato-amarelo 12 4 16 B B BElaenia mesoleuca (Deppe, 1830) Tuque B B BElaenia spectabilis Pelzeln, 1868 Guaracava-grande B B BFluvicola nengeta (Linnaeus, 1766) Lavadeira-mascarada B B BHemitriccus margaritaceiventer (d'Orbigny & Lafresnaye, 1837) Sebinho-de-olho-de-ouro M B BHemitriccus zosterops (Pelzeln, 1868) Maria-de-olho-branco A B MLathrotriccus euleri (Cabanis, 1868) Enferrujado B B BLegatus leucophaius (Vieillot, 1818) Bentevi-pirata B B BMachetornis rixosus (Vieillot, 1819) Bentevi-do-gado B B BMegarynchus pitangua (Linnaeus, 1766) Bentevi-de-bico-chato B B BMyiarchus ferox (Gmelin, 1789) Maria-cavaleira B B BMyiarchus swaisoni Cabanis & Heine, 1859 Irrê B B BMyiarchus tuberculifer (d'Orbigny & Lafresnaye, 1837) Maria-cavaleira-pequena B B BMyiarchus tyrannulus (Statius Muller, 1776) Maria-caveleira-de-rabo-enferrujado B B BMyiodynastes maculatus (Statius Muller, 1776) Bentevi-rajado B B BMyiopagis gaimardii (d'Orbigny, 1839) Maria-pechim M B BMyiopagis viridicata (Vieillot, 1817) Guaracava-de-olheiras M B BMyiophobus fasciatus (Statius Muller, 1776) Filipe B B BPhaeomyias murina (Spix, 1825) Bagageiro B B BPoecilotriccus fumifrons (Hartlaub, 1853) Ferreirinho-de-testa-parda B B BTodirostrum cinereum (Linnaeus, 1766) Ferreirinho, relógio B B BTolmomyias poliocephalus (Taczanowski, 1884) Bico-chato-de-cabeça-fina M B BTyrannus melancholicus Vieillot, 1819 Suiriri B B BZimmerius gracilipes (Sclater & Salvin, 1868) Poiaeiro-de-pata-fina M B BPipridaeChiroxiphia pareola (Linnaeus, 1766) Tangará-falso 16 1 17 A B BManacus manacus (Linnaeus, 1766) Rendeira 26 4 6 36 B B BNeopelma pallescens (Lafresnaye, 1853) Fruxu-do-cerradão 28 17 3 48 M B BPipra rubrocapilla Temminck, 1821 Cabeça-encarnada 17 1 18 A B BHirundinidaeProgne chalybea (Gmelin, 1789) Andorinha-doméstica-grande B B BStelgidopteryx ruficollis (Vieillot, 1817) Andorinha-serrador 4 4 B B BCotingidaeIodopleura pipra (Lesson, 1831)* Anambé-de-crista M A A
Espécies Nome Popular CP RC RP Total SD PC PP

57
Apêndice 1. Continuação
Troglodytidae (continuação)Thryothorus genibarbis Swainson, 1838 Pai-avô, Pia-vovó B B BTroglodytes musculus Naumann, 1823 Corruíra, Rouxinol 3 3 B B MMuscicapidaePolioptila plumbea (Gmelin, 1788) Balança-rabo B B BRamphocaenus melanurus Vieillot, 1819 Bico-assovelado 2 2 B B BTurdus amaurochalinus Cabanis, 1850 Sabiá-bico-de-osso B B BTurdus leucomelas Vieillot, 1818 Sabiá-barranco 24 4 28 B B BTurdus rufiventris Vieillot, 1818 Sabiá-laranjeira B B BVireonidaeCyclarhis gujanensis (Gmelin, 1789) Pitiguari 1 1 B B BVireo olivaceus (Linnaeus, 1766) Juruviara-norte-americano 3 3 B B MEmberizidaeArremon taciturnus ( Hermann, 1783) Tico-tico-do-mato-de-bico-preto 16 10 6 32 M B BBasileuterus flaveolus ( Baird, 1865) Canário-do-mato 18 12 30 M B BCoereba flaveola (Linnaeus, 1758) Cambacica 10 1 11 B B BCyanerpes cyaneus (Linnaeus, 1766) Saíra-beija-flor 15 1 1 17 B B BDacnis cayana (Linnaeus, 1766) Saí-azul 3 1 4 B B BEuphonia chlorotica (Linnaeus, 1766) Vem-vem B B BEuphonia violacea (Linnaeus, 1758) Gaturamo-verdadeiro 11 2 2 15 B B BIcterus cayanensis (Linnaeus, 1766) Encontro 1 1 B B BMolothrus bonariensis (Gmelin, 1789) Chopim 1 1 B B MPasserina brissonii (Lichtenstein, 1823) Azulão M B BRamphocelus bresilius (Linnaeus, 1766)* Tiê-sangue 1 1 1 3 B B MSaltator maximus (Müller, 1776) Tempera-viola 2 2 B B BSchistochlamys melanopis (Latham, 1790) Sanhaço-de-coleira B B BSicalis flaveola (Linnaues, 1766) Canário-da-terra B B BSporophila angolensis (Linnaeus, 1766) Curió B B MSporophila bouvreuil (Statius Muller, 1776) Caboclinho M M MSporophila leucoptera (Vielillot, 1817) Patativa-chorona B B MSporophila nigricollis (Vieillot, 1823) Baiano 1 1 B B BTachyphonus cristatus (Linnaeus, 1766) Tiê-galo 1 1 M B BTachyphonus rufus (Boddaert, 1783) Pipira-preta 10 3 13 B B BTangara cayana (Linnaeus, 1766) Saíra-amarelo 5 1 6 M B BTersina viridis (Illiger, 1811) Saí-andorinha B B MThlypopsis sordida (D'Orbigny & Lafresnaye, 1837) Canário-sapé 1 1 B B B
Espécies Nome Popular CP RC RP Total SD PC PP
58
Apêndice 1. Continuação
Emberizidae (Continuação)Thraupis palmarum (Wied, 1821) Sanhaço-de-coqueiro 7 7 B B BThraupis sayaca (Linnaeus, 1766) Sanhaço-cinzento 2 2 B B BVolatina jacarina (Linnaeus, 1766) Tiziu B B BPasseridaePasser domesticus (Linnaeus, 1758) Pardal B B MEstrildidaeEstrilda astrild (Linnaeus, 1758) Bico-de-lacre * * *
335 98 23 456
Espécies Nome Popular CP RC RP Total SD PC PP
* Espécies endêmicas; ** Espécies ameaçadas de extinção.
59
Apêndice 2. Planilha utilizada para registro dos dados coletados
LOCAL: Data: Data: Data:
1. Área 1 (0 m) Temperatura: Temperatura: Temperatura:2. Àrea 1 (200 m)3. Área 2 (0 m) MÊS / ANO: Hora: Hora: Hora:4. Área 2 (200 m)
Tempo: Tempo: Tempo:Obs: As anilhas devem ser colocadas no tarso direito, salvo exceções descritas na observação.
PLANILHA DE CAMPO
AMBIENTE
CONTORNO
C = CabeçaD = DorsoV = Ventre
No DA ANILHA (metal)
AMBIENTE
4. 4º segmentoN = NinhegoI = Indeterminado
MALHA
ESPÉCIEDIA
STATUS
MUDA
REPOSIÇÃO
HORA
MALHA
IDADE
PESO (g)
PESO COM
SACO (g)
OBS.
PESO DO
SACO (g)
RÊMIGES
CONTORNO
RETRIZES
NARINA/ PONTA
MEDIDAS (mm)
ASA
DIREITA
CAUDA CÚLMEN TOTAL
TARSO DIREITO
TARSO ESQUER.
∅ TARSO
ESQUER.
ASA
ESQUERDA
∅ TARSO
DIREITO
PLACA
4. Anilha destruída
IDADE
A = Adulto J = Jovem
STATUS
1. Ave nova2. Recaptura3. Recuperação 3. 3º segmento
SEXO
M = Macho F = FêmeaI = Indeterminado
1. 1º segmento2. 2º segmento
SEXO
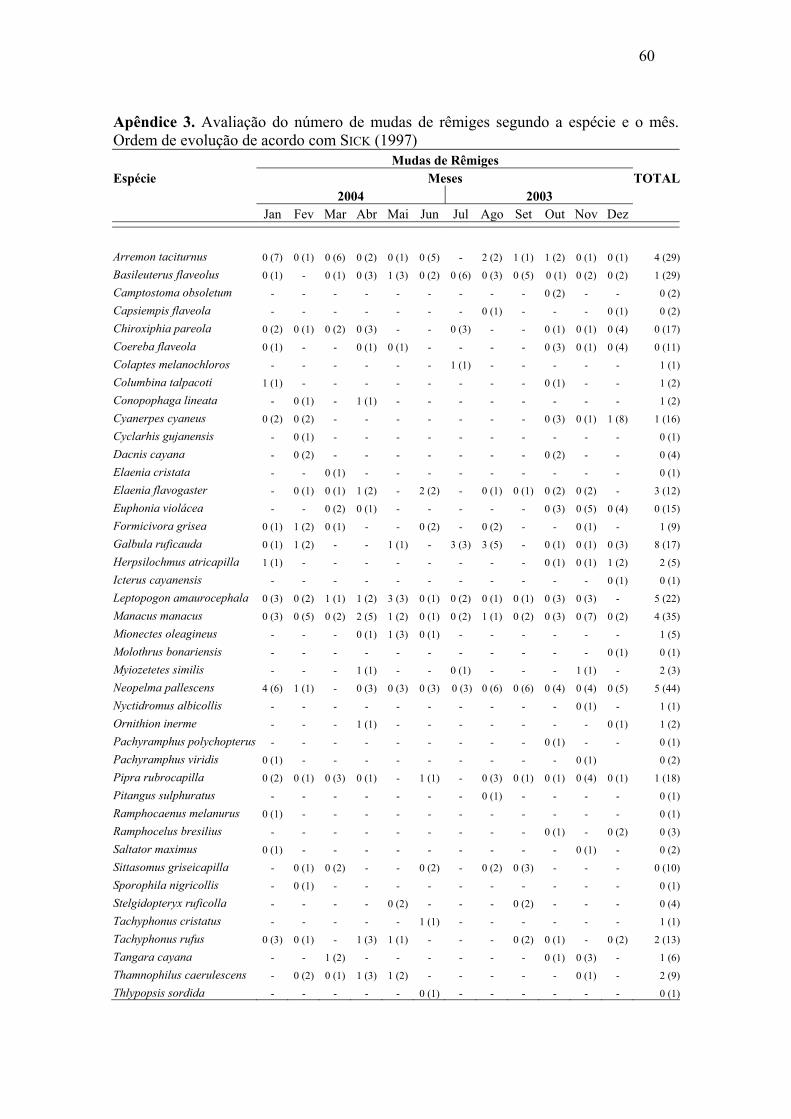
60
Apêndice 3. Avaliação do número de mudas de rêmiges segundo a espécie e o mês. Ordem de evolução de acordo com SICK (1997) Mudas de Rêmiges Espécie Meses TOTAL 2004 2003 Jan Fev Mar Abr Mai Jun Jul Ago Set Out Nov Dez
Arremon taciturnus 0 (7) 0 (1) 0 (6) 0 (2) 0 (1) 0 (5) - 2 (2) 1 (1) 1 (2) 0 (1) 0 (1) 4 (29)
Basileuterus flaveolus 0 (1) - 0 (1) 0 (3) 1 (3) 0 (2) 0 (6) 0 (3) 0 (5) 0 (1) 0 (2) 0 (2) 1 (29)
Camptostoma obsoletum - - - - - - - - - 0 (2) - - 0 (2)
Capsiempis flaveola - - - - - - - 0 (1) - - - 0 (1) 0 (2)
Chiroxiphia pareola 0 (2) 0 (1) 0 (2) 0 (3) - - 0 (3) - - 0 (1) 0 (1) 0 (4) 0 (17)
Coereba flaveola 0 (1) - - 0 (1) 0 (1) - - - - 0 (3) 0 (1) 0 (4) 0 (11)
Colaptes melanochloros - - - - - - 1 (1) - - - - - 1 (1)
Columbina talpacoti 1 (1) - - - - - - - - 0 (1) - - 1 (2)
Conopophaga lineata - 0 (1) - 1 (1) - - - - - - - - 1 (2)
Cyanerpes cyaneus 0 (2) 0 (2) - - - - - - - 0 (3) 0 (1) 1 (8) 1 (16)
Cyclarhis gujanensis - 0 (1) - - - - - - - - - - 0 (1)
Dacnis cayana - 0 (2) - - - - - - - 0 (2) - - 0 (4)
Elaenia cristata - - 0 (1) - - - - - - - - - 0 (1)
Elaenia flavogaster - 0 (1) 0 (1) 1 (2) - 2 (2) - 0 (1) 0 (1) 0 (2) 0 (2) - 3 (12)
Euphonia violácea - - 0 (2) 0 (1) - - - - - 0 (3) 0 (5) 0 (4) 0 (15)
Formicivora grisea 0 (1) 1 (2) 0 (1) - - 0 (2) - 0 (2) - - 0 (1) - 1 (9)
Galbula ruficauda 0 (1) 1 (2) - - 1 (1) - 3 (3) 3 (5) - 0 (1) 0 (1) 0 (3) 8 (17)
Herpsilochmus atricapilla 1 (1) - - - - - - - - 0 (1) 0 (1) 1 (2) 2 (5)
Icterus cayanensis - - - - - - - - - - - 0 (1) 0 (1)
Leptopogon amaurocephala 0 (3) 0 (2) 1 (1) 1 (2) 3 (3) 0 (1) 0 (2) 0 (1) 0 (1) 0 (3) 0 (3) - 5 (22)
Manacus manacus 0 (3) 0 (5) 0 (2) 2 (5) 1 (2) 0 (1) 0 (2) 1 (1) 0 (2) 0 (3) 0 (7) 0 (2) 4 (35)
Mionectes oleagineus - - - 0 (1) 1 (3) 0 (1) - - - - - - 1 (5)
Molothrus bonariensis - - - - - - - - - - - 0 (1) 0 (1)
Myiozetetes similis - - - 1 (1) - - 0 (1) - - - 1 (1) - 2 (3)
Neopelma pallescens 4 (6) 1 (1) - 0 (3) 0 (3) 0 (3) 0 (3) 0 (6) 0 (6) 0 (4) 0 (4) 0 (5) 5 (44)
Nyctidromus albicollis - - - - - - - - - - 0 (1) - 1 (1)
Ornithion inerme - - - 1 (1) - - - - - - - 0 (1) 1 (2)
Pachyramphus polychopterus - - - - - - - - - 0 (1) - - 0 (1)
Pachyramphus viridis 0 (1) - - - - - - - - - 0 (1) 0 (2)
Pipra rubrocapilla 0 (2) 0 (1) 0 (3) 0 (1) - 1 (1) - 0 (3) 0 (1) 0 (1) 0 (4) 0 (1) 1 (18)
Pitangus sulphuratus - - - - - - - 0 (1) - - - - 0 (1)
Ramphocaenus melanurus 0 (1) - - - - - - - - - - - 0 (1)
Ramphocelus bresilius - - - - - - - - - 0 (1) - 0 (2) 0 (3)
Saltator maximus 0 (1) - - - - - - - - - 0 (1) - 0 (2)
Sittasomus griseicapilla - 0 (1) 0 (2) - - 0 (2) - 0 (2) 0 (3) - - - 0 (10)
Sporophila nigricollis - 0 (1) - - - - - - - - - - 0 (1)
Stelgidopteryx ruficolla - - - - 0 (2) - - - 0 (2) - - - 0 (4)
Tachyphonus cristatus - - - - - 1 (1) - - - - - - 1 (1)
Tachyphonus rufus 0 (3) 0 (1) - 1 (3) 1 (1) - - - 0 (2) 0 (1) - 0 (2) 2 (13)
Tangara cayana - - 1 (2) - - - - - - 0 (1) 0 (3) - 1 (6)
Thamnophilus caerulescens - 0 (2) 0 (1) 1 (3) 1 (2) - - - - - 0 (1) - 2 (9)
Thlypopsis sordida - - - - - 0 (1) - - - - - - 0 (1)
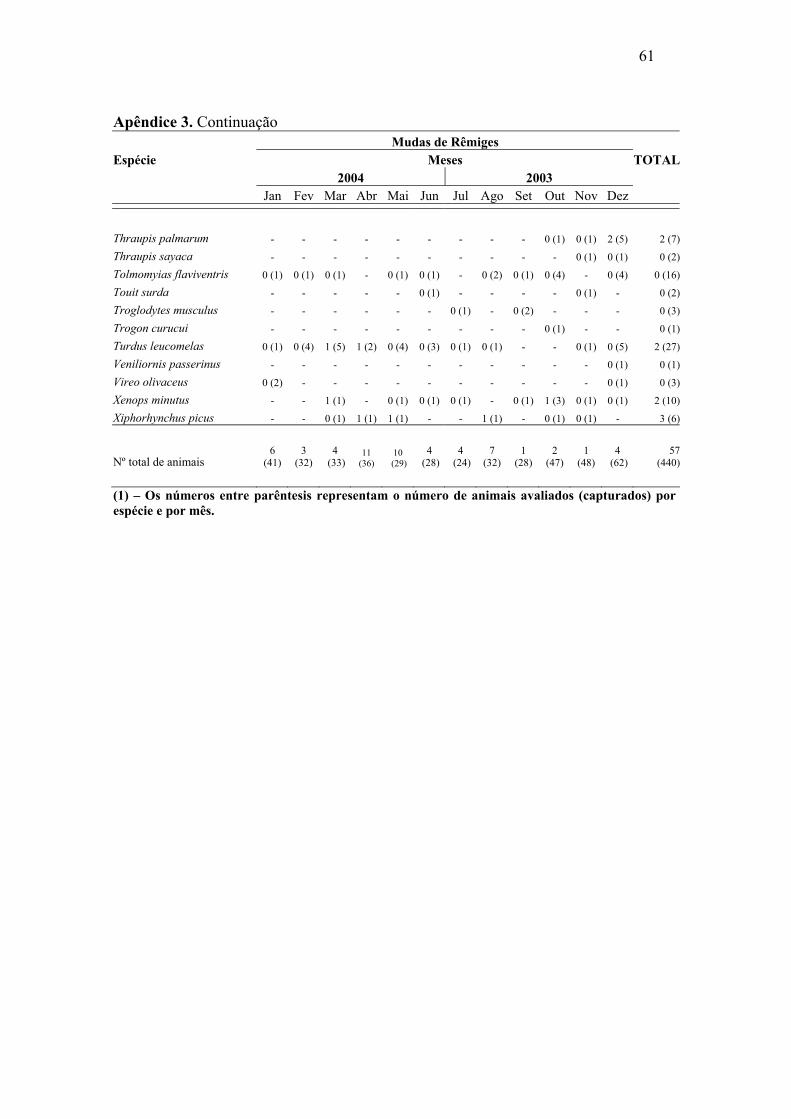
61
Apêndice 3. Continuação Mudas de Rêmiges Espécie Meses TOTAL 2004 2003 Jan Fev Mar Abr Mai Jun Jul Ago Set Out Nov Dez
Thraupis palmarum - - - - - - - - - 0 (1) 0 (1) 2 (5) 2 (7)
Thraupis sayaca - - - - - - - - - - 0 (1) 0 (1) 0 (2)
Tolmomyias flaviventris 0 (1) 0 (1) 0 (1) - 0 (1) 0 (1) - 0 (2) 0 (1) 0 (4) - 0 (4) 0 (16)
Touit surda - - - - - 0 (1) - - - - 0 (1) - 0 (2)
Troglodytes musculus - - - - - - 0 (1) - 0 (2) - - - 0 (3)
Trogon curucui - - - - - - - - - 0 (1) - - 0 (1)
Turdus leucomelas 0 (1) 0 (4) 1 (5) 1 (2) 0 (4) 0 (3) 0 (1) 0 (1) - - 0 (1) 0 (5) 2 (27)
Veniliornis passerinus - - - - - - - - - - - 0 (1) 0 (1)
Vireo olivaceus 0 (2) - - - - - - - - - - 0 (1) 0 (3)
Xenops minutus - - 1 (1) - 0 (1) 0 (1) 0 (1) - 0 (1) 1 (3) 0 (1) 0 (1) 2 (10)
Xiphorhynchus picus - - 0 (1) 1 (1) 1 (1) - - 1 (1) - 0 (1) 0 (1) - 3 (6)
Nº total de animais 6
(41) 3
(32) 4
(33) 11
(36) 10
(29) 4
(28) 4
(24) 7
(32) 1
(28) 2
(47) 1
(48) 4
(62) 57
(440)
(1) – Os números entre parêntesis representam o número de animais avaliados (capturados) por espécie e por mês.
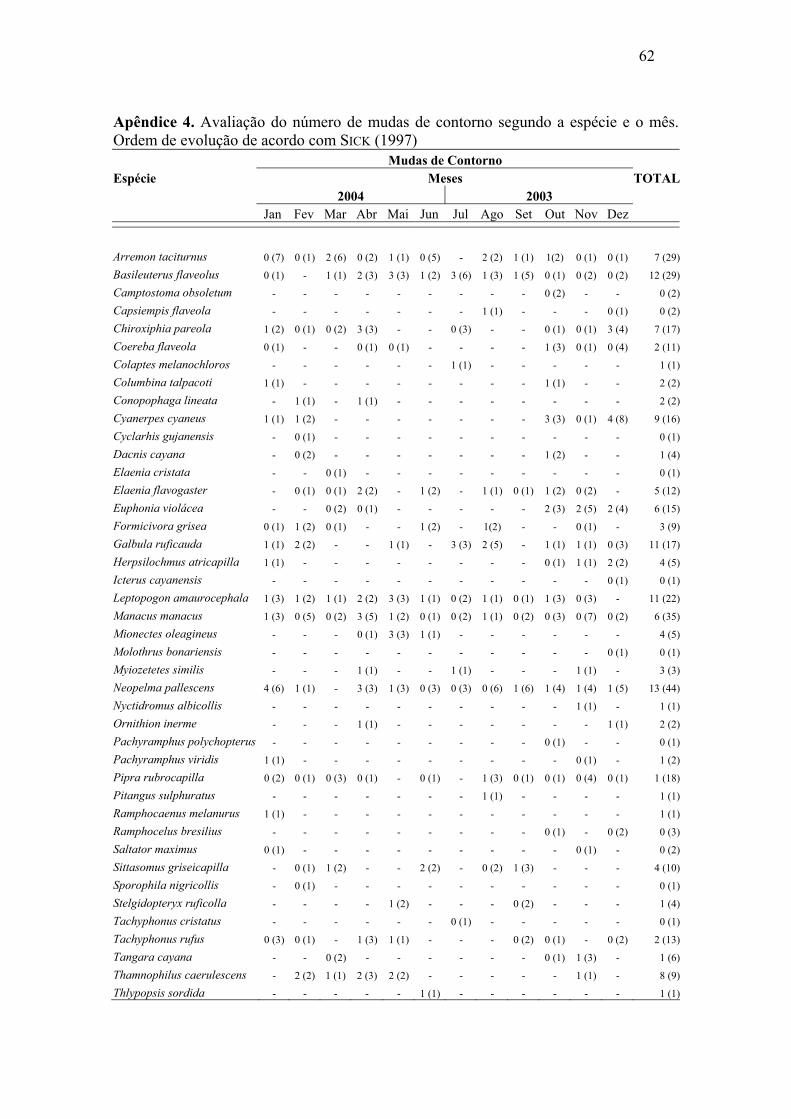
62
Apêndice 4. Avaliação do número de mudas de contorno segundo a espécie e o mês. Ordem de evolução de acordo com SICK (1997) Mudas de Contorno Espécie Meses TOTAL 2004 2003 Jan Fev Mar Abr Mai Jun Jul Ago Set Out Nov Dez
Arremon taciturnus 0 (7) 0 (1) 2 (6) 0 (2) 1 (1) 0 (5) - 2 (2) 1 (1) 1(2) 0 (1) 0 (1) 7 (29)
Basileuterus flaveolus 0 (1) - 1 (1) 2 (3) 3 (3) 1 (2) 3 (6) 1 (3) 1 (5) 0 (1) 0 (2) 0 (2) 12 (29)
Camptostoma obsoletum - - - - - - - - - 0 (2) - - 0 (2)
Capsiempis flaveola - - - - - - - 1 (1) - - - 0 (1) 0 (2)
Chiroxiphia pareola 1 (2) 0 (1) 0 (2) 3 (3) - - 0 (3) - - 0 (1) 0 (1) 3 (4) 7 (17)
Coereba flaveola 0 (1) - - 0 (1) 0 (1) - - - - 1 (3) 0 (1) 0 (4) 2 (11)
Colaptes melanochloros - - - - - - 1 (1) - - - - - 1 (1)
Columbina talpacoti 1 (1) - - - - - - - - 1 (1) - - 2 (2)
Conopophaga lineata - 1 (1) - 1 (1) - - - - - - - - 2 (2)
Cyanerpes cyaneus 1 (1) 1 (2) - - - - - - - 3 (3) 0 (1) 4 (8) 9 (16)
Cyclarhis gujanensis - 0 (1) - - - - - - - - - - 0 (1)
Dacnis cayana - 0 (2) - - - - - - - 1 (2) - - 1 (4)
Elaenia cristata - - 0 (1) - - - - - - - - - 0 (1)
Elaenia flavogaster - 0 (1) 0 (1) 2 (2) - 1 (2) - 1 (1) 0 (1) 1 (2) 0 (2) - 5 (12)
Euphonia violácea - - 0 (2) 0 (1) - - - - - 2 (3) 2 (5) 2 (4) 6 (15)
Formicivora grisea 0 (1) 1 (2) 0 (1) - - 1 (2) - 1(2) - - 0 (1) - 3 (9)
Galbula ruficauda 1 (1) 2 (2) - - 1 (1) - 3 (3) 2 (5) - 1 (1) 1 (1) 0 (3) 11 (17)
Herpsilochmus atricapilla 1 (1) - - - - - - - - 0 (1) 1 (1) 2 (2) 4 (5)
Icterus cayanensis - - - - - - - - - - - 0 (1) 0 (1)
Leptopogon amaurocephala 1 (3) 1 (2) 1 (1) 2 (2) 3 (3) 1 (1) 0 (2) 1 (1) 0 (1) 1 (3) 0 (3) - 11 (22)
Manacus manacus 1 (3) 0 (5) 0 (2) 3 (5) 1 (2) 0 (1) 0 (2) 1 (1) 0 (2) 0 (3) 0 (7) 0 (2) 6 (35)
Mionectes oleagineus - - - 0 (1) 3 (3) 1 (1) - - - - - - 4 (5)
Molothrus bonariensis - - - - - - - - - - - 0 (1) 0 (1)
Myiozetetes similis - - - 1 (1) - - 1 (1) - - - 1 (1) - 3 (3)
Neopelma pallescens 4 (6) 1 (1) - 3 (3) 1 (3) 0 (3) 0 (3) 0 (6) 1 (6) 1 (4) 1 (4) 1 (5) 13 (44)
Nyctidromus albicollis - - - - - - - - - - 1 (1) - 1 (1)
Ornithion inerme - - - 1 (1) - - - - - - - 1 (1) 2 (2)
Pachyramphus polychopterus - - - - - - - - - 0 (1) - - 0 (1)
Pachyramphus viridis 1 (1) - - - - - - - - - 0 (1) - 1 (2)
Pipra rubrocapilla 0 (2) 0 (1) 0 (3) 0 (1) - 0 (1) - 1 (3) 0 (1) 0 (1) 0 (4) 0 (1) 1 (18)
Pitangus sulphuratus - - - - - - - 1 (1) - - - - 1 (1)
Ramphocaenus melanurus 1 (1) - - - - - - - - - - - 1 (1)
Ramphocelus bresilius - - - - - - - - - 0 (1) - 0 (2) 0 (3)
Saltator maximus 0 (1) - - - - - - - - - 0 (1) - 0 (2)
Sittasomus griseicapilla - 0 (1) 1 (2) - - 2 (2) - 0 (2) 1 (3) - - - 4 (10)
Sporophila nigricollis - 0 (1) - - - - - - - - - - 0 (1)
Stelgidopteryx ruficolla - - - - 1 (2) - - - 0 (2) - - - 1 (4)
Tachyphonus cristatus - - - - - - 0 (1) - - - - - 0 (1)
Tachyphonus rufus 0 (3) 0 (1) - 1 (3) 1 (1) - - - 0 (2) 0 (1) - 0 (2) 2 (13)
Tangara cayana - - 0 (2) - - - - - - 0 (1) 1 (3) - 1 (6)
Thamnophilus caerulescens - 2 (2) 1 (1) 2 (3) 2 (2) - - - - - 1 (1) - 8 (9)
Thlypopsis sordida - - - - - 1 (1) - - - - - - 1 (1)
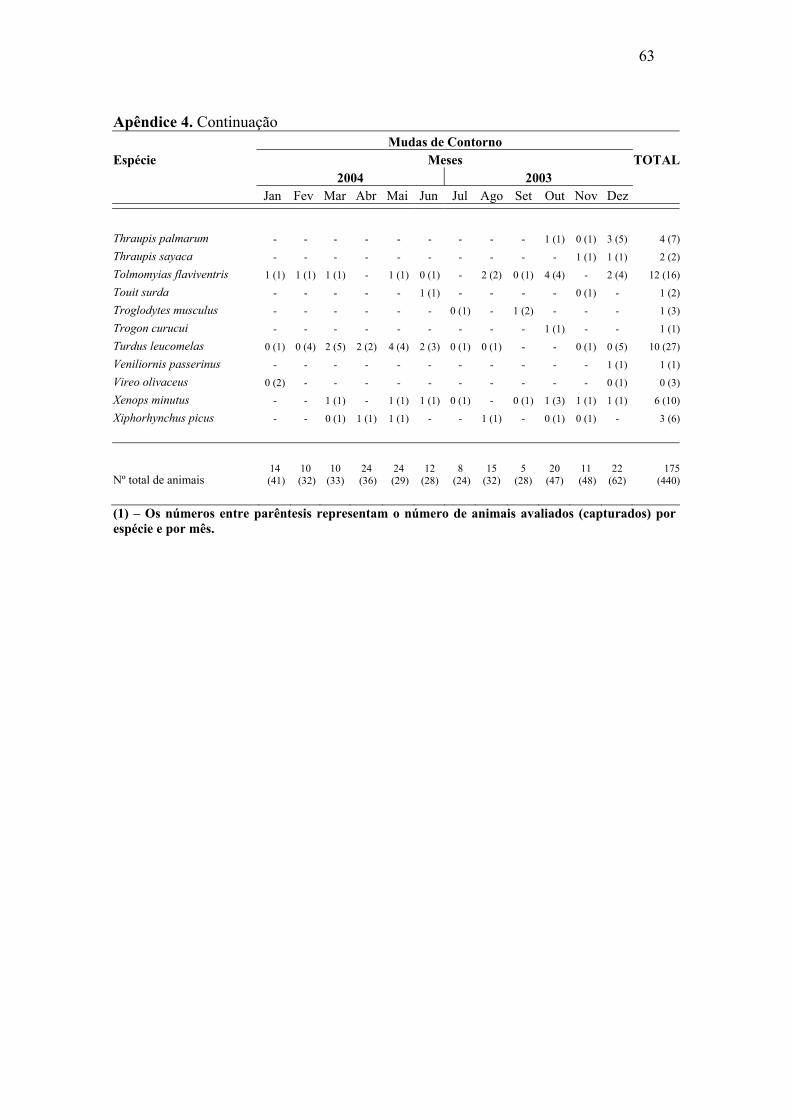
63
Apêndice 4. Continuação Mudas de Contorno Espécie Meses TOTAL 2004 2003 Jan Fev Mar Abr Mai Jun Jul Ago Set Out Nov Dez
Thraupis palmarum - - - - - - - - - 1 (1) 0 (1) 3 (5) 4 (7)
Thraupis sayaca - - - - - - - - - - 1 (1) 1 (1) 2 (2)
Tolmomyias flaviventris 1 (1) 1 (1) 1 (1) - 1 (1) 0 (1) - 2 (2) 0 (1) 4 (4) - 2 (4) 12 (16)
Touit surda - - - - - 1 (1) - - - - 0 (1) - 1 (2)
Troglodytes musculus - - - - - - 0 (1) - 1 (2) - - - 1 (3)
Trogon curucui - - - - - - - - - 1 (1) - - 1 (1)
Turdus leucomelas 0 (1) 0 (4) 2 (5) 2 (2) 4 (4) 2 (3) 0 (1) 0 (1) - - 0 (1) 0 (5) 10 (27)
Veniliornis passerinus - - - - - - - - - - - 1 (1) 1 (1)
Vireo olivaceus 0 (2) - - - - - - - - - - 0 (1) 0 (3)
Xenops minutus - - 1 (1) - 1 (1) 1 (1) 0 (1) - 0 (1) 1 (3) 1 (1) 1 (1) 6 (10)
Xiphorhynchus picus - - 0 (1) 1 (1) 1 (1) - - 1 (1) - 0 (1) 0 (1) - 3 (6)
Nº total de animais 14
(41) 10
(32) 10
(33) 24
(36) 24
(29) 12
(28) 8
(24)15
(32) 5
(28) 20
(47) 11
(48) 22
(62) 175
(440)
(1) – Os números entre parêntesis representam o número de animais avaliados (capturados) por espécie e por mês.
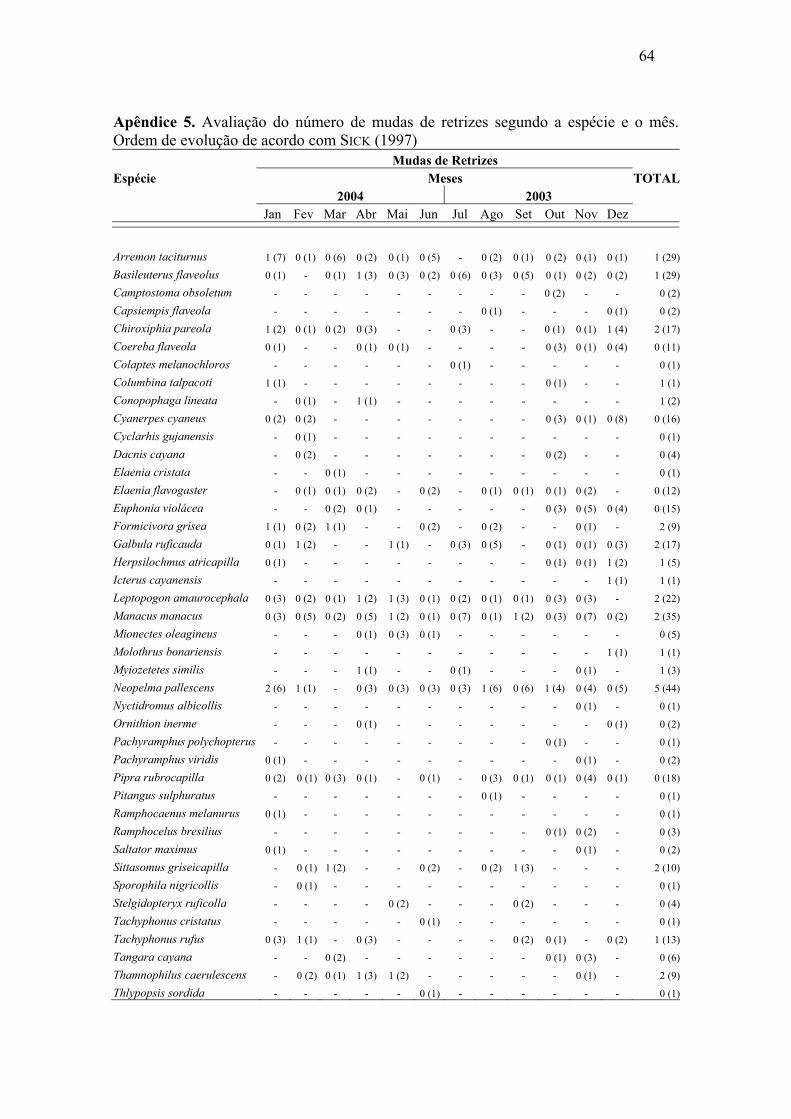
64
Apêndice 5. Avaliação do número de mudas de retrizes segundo a espécie e o mês. Ordem de evolução de acordo com SICK (1997) Mudas de Retrizes Espécie Meses TOTAL 2004 2003 Jan Fev Mar Abr Mai Jun Jul Ago Set Out Nov Dez
Arremon taciturnus 1 (7) 0 (1) 0 (6) 0 (2) 0 (1) 0 (5) - 0 (2) 0 (1) 0 (2) 0 (1) 0 (1) 1 (29)
Basileuterus flaveolus 0 (1) - 0 (1) 1 (3) 0 (3) 0 (2) 0 (6) 0 (3) 0 (5) 0 (1) 0 (2) 0 (2) 1 (29)
Camptostoma obsoletum - - - - - - - - - 0 (2) - - 0 (2)
Capsiempis flaveola - - - - - - - 0 (1) - - - 0 (1) 0 (2)
Chiroxiphia pareola 1 (2) 0 (1) 0 (2) 0 (3) - - 0 (3) - - 0 (1) 0 (1) 1 (4) 2 (17)
Coereba flaveola 0 (1) - - 0 (1) 0 (1) - - - - 0 (3) 0 (1) 0 (4) 0 (11)
Colaptes melanochloros - - - - - - 0 (1) - - - - - 0 (1)
Columbina talpacoti 1 (1) - - - - - - - - 0 (1) - - 1 (1)
Conopophaga lineata - 0 (1) - 1 (1) - - - - - - - - 1 (2)
Cyanerpes cyaneus 0 (2) 0 (2) - - - - - - - 0 (3) 0 (1) 0 (8) 0 (16)
Cyclarhis gujanensis - 0 (1) - - - - - - - - - - 0 (1)
Dacnis cayana - 0 (2) - - - - - - - 0 (2) - - 0 (4)
Elaenia cristata - - 0 (1) - - - - - - - - - 0 (1)
Elaenia flavogaster - 0 (1) 0 (1) 0 (2) - 0 (2) - 0 (1) 0 (1) 0 (1) 0 (2) - 0 (12)
Euphonia violácea - - 0 (2) 0 (1) - - - - - 0 (3) 0 (5) 0 (4) 0 (15)
Formicivora grisea 1 (1) 0 (2) 1 (1) - - 0 (2) - 0 (2) - - 0 (1) - 2 (9)
Galbula ruficauda 0 (1) 1 (2) - - 1 (1) - 0 (3) 0 (5) - 0 (1) 0 (1) 0 (3) 2 (17)
Herpsilochmus atricapilla 0 (1) - - - - - - - - 0 (1) 0 (1) 1 (2) 1 (5)
Icterus cayanensis - - - - - - - - - - - 1 (1) 1 (1)
Leptopogon amaurocephala 0 (3) 0 (2) 0 (1) 1 (2) 1 (3) 0 (1) 0 (2) 0 (1) 0 (1) 0 (3) 0 (3) - 2 (22)
Manacus manacus 0 (3) 0 (5) 0 (2) 0 (5) 1 (2) 0 (1) 0 (7) 0 (1) 1 (2) 0 (3) 0 (7) 0 (2) 2 (35)
Mionectes oleagineus - - - 0 (1) 0 (3) 0 (1) - - - - - - 0 (5)
Molothrus bonariensis - - - - - - - - - - - 1 (1) 1 (1)
Myiozetetes similis - - - 1 (1) - - 0 (1) - - - 0 (1) - 1 (3)
Neopelma pallescens 2 (6) 1 (1) - 0 (3) 0 (3) 0 (3) 0 (3) 1 (6) 0 (6) 1 (4) 0 (4) 0 (5) 5 (44)
Nyctidromus albicollis - - - - - - - - - - 0 (1) - 0 (1)
Ornithion inerme - - - 0 (1) - - - - - - - 0 (1) 0 (2)
Pachyramphus polychopterus - - - - - - - - - 0 (1) - - 0 (1)
Pachyramphus viridis 0 (1) - - - - - - - - - 0 (1) - 0 (2)
Pipra rubrocapilla 0 (2) 0 (1) 0 (3) 0 (1) - 0 (1) - 0 (3) 0 (1) 0 (1) 0 (4) 0 (1) 0 (18)
Pitangus sulphuratus - - - - - - - 0 (1) - - - - 0 (1)
Ramphocaenus melanurus 0 (1) - - - - - - - - - - - 0 (1)
Ramphocelus bresilius - - - - - - - - - 0 (1) 0 (2) - 0 (3)
Saltator maximus 0 (1) - - - - - - - - - 0 (1) - 0 (2)
Sittasomus griseicapilla - 0 (1) 1 (2) - - 0 (2) - 0 (2) 1 (3) - - - 2 (10)
Sporophila nigricollis - 0 (1) - - - - - - - - - - 0 (1)
Stelgidopteryx ruficolla - - - - 0 (2) - - - 0 (2) - - - 0 (4)
Tachyphonus cristatus - - - - - 0 (1) - - - - - - 0 (1)
Tachyphonus rufus 0 (3) 1 (1) - 0 (3) - - - - 0 (2) 0 (1) - 0 (2) 1 (13)
Tangara cayana - - 0 (2) - - - - - - 0 (1) 0 (3) - 0 (6)
Thamnophilus caerulescens - 0 (2) 0 (1) 1 (3) 1 (2) - - - - - 0 (1) - 2 (9)
Thlypopsis sordida - - - - - 0 (1) - - - - - - 0 (1)
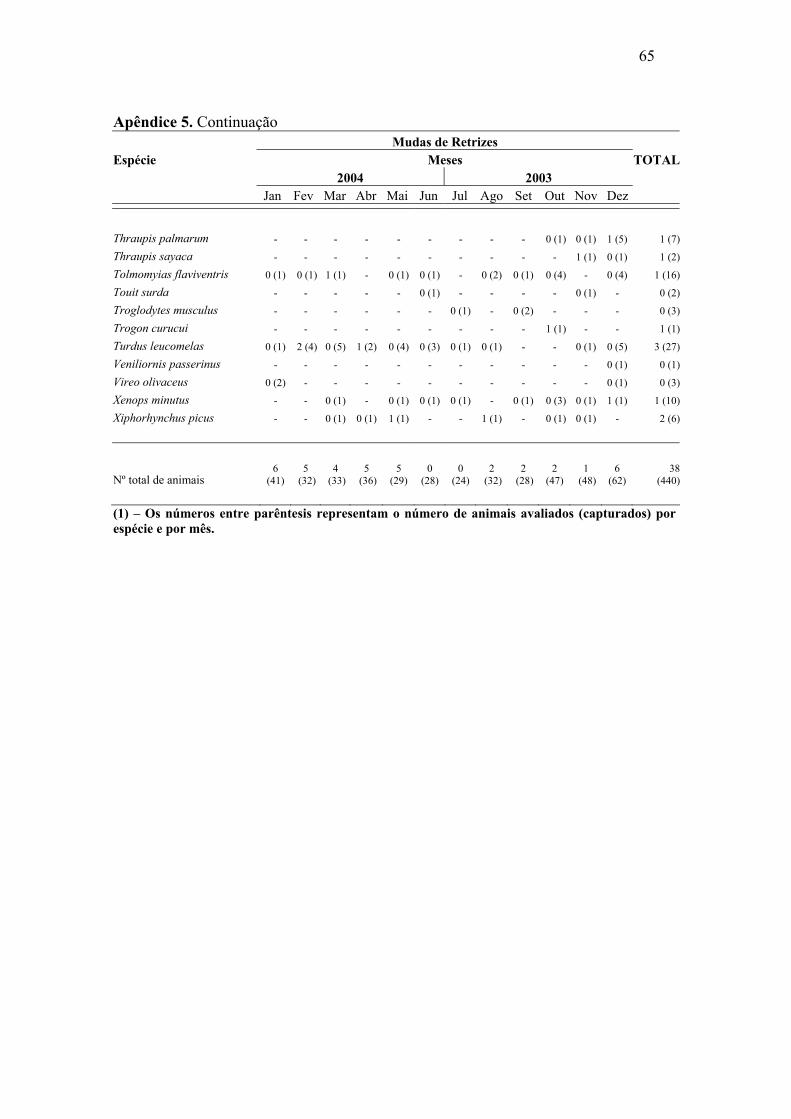
65
Apêndice 5. Continuação Mudas de Retrizes Espécie Meses TOTAL 2004 2003 Jan Fev Mar Abr Mai Jun Jul Ago Set Out Nov Dez
Thraupis palmarum - - - - - - - - - 0 (1) 0 (1) 1 (5) 1 (7)
Thraupis sayaca - - - - - - - - - - 1 (1) 0 (1) 1 (2)
Tolmomyias flaviventris 0 (1) 0 (1) 1 (1) - 0 (1) 0 (1) - 0 (2) 0 (1) 0 (4) - 0 (4) 1 (16)
Touit surda - - - - - 0 (1) - - - - 0 (1) - 0 (2)
Troglodytes musculus - - - - - - 0 (1) - 0 (2) - - - 0 (3)
Trogon curucui - - - - - - - - - 1 (1) - - 1 (1)
Turdus leucomelas 0 (1) 2 (4) 0 (5) 1 (2) 0 (4) 0 (3) 0 (1) 0 (1) - - 0 (1) 0 (5) 3 (27)
Veniliornis passerinus - - - - - - - - - - - 0 (1) 0 (1)
Vireo olivaceus 0 (2) - - - - - - - - - - 0 (1) 0 (3)
Xenops minutus - - 0 (1) - 0 (1) 0 (1) 0 (1) - 0 (1) 0 (3) 0 (1) 1 (1) 1 (10)
Xiphorhynchus picus - - 0 (1) 0 (1) 1 (1) - - 1 (1) - 0 (1) 0 (1) - 2 (6)
Nº total de animais 6
(41) 5
(32) 4
(33)5
(36) 5
(29) 0
(28) 0
(24) 2
(32) 2
(28) 2
(47) 1
(48) 6
(62) 38
(440)
(1) – Os números entre parêntesis representam o número de animais avaliados (capturados) por espécie e por mês.
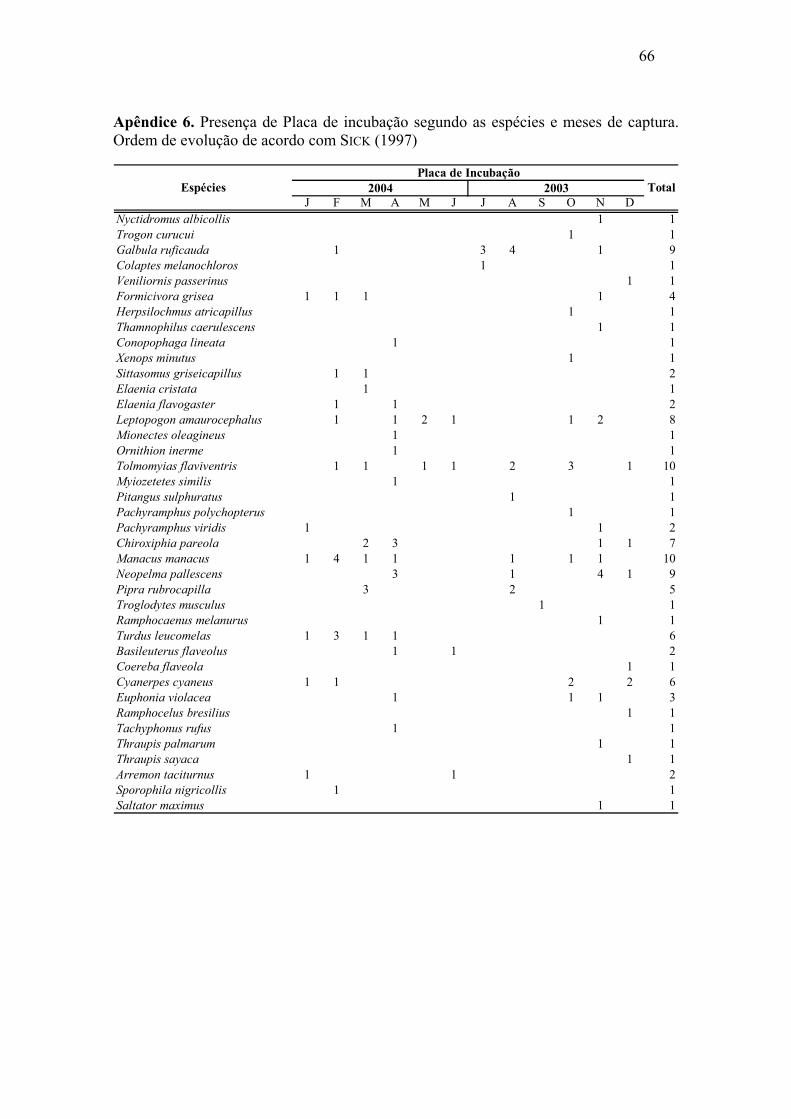
66
Apêndice 6. Presença de Placa de incubação segundo as espécies e meses de captura. Ordem de evolução de acordo com SICK (1997)
J F M A M J J A S O N DNyctidromus albicollis 1 1Trogon curucui 1 1Galbula ruficauda 1 3 4 1 9Colaptes melanochloros 1 1Veniliornis passerinus 1 1Formicivora grisea 1 1 1 1 4Herpsilochmus atricapillus 1 1Thamnophilus caerulescens 1 1Conopophaga lineata 1 1Xenops minutus 1 1Sittasomus griseicapillus 1 1 2Elaenia cristata 1 1Elaenia flavogaster 1 1 2Leptopogon amaurocephalus 1 1 2 1 1 2 8Mionectes oleagineus 1 1Ornithion inerme 1 1Tolmomyias flaviventris 1 1 1 1 2 3 1 10Myiozetetes similis 1 1Pitangus sulphuratus 1 1Pachyramphus polychopterus 1 1Pachyramphus viridis 1 1 2Chiroxiphia pareola 2 3 1 1 7Manacus manacus 1 4 1 1 1 1 1 10Neopelma pallescens 3 1 4 1 9Pipra rubrocapilla 3 2 5Troglodytes musculus 1 1Ramphocaenus melanurus 1 1Turdus leucomelas 1 3 1 1 6Basileuterus flaveolus 1 1 2Coereba flaveola 1 1Cyanerpes cyaneus 1 1 2 2 6Euphonia violacea 1 1 1 3Ramphocelus bresilius 1 1Tachyphonus rufus 1 1Thraupis palmarum 1 1Thraupis sayaca 1 1Arremon taciturnus 1 1 2Sporophila nigricollis 1 1Saltator maximus 1 1
Placa de IncubaçãoEspécies Total2004 2003


















