
Aula09BioqII Metabolismo de Aminoácidos · nitrogenados: heme, aminas biologicamente ativas,...
44
Aula de Bioquímica II Tema: Metabolismo de aminoácidos Prof. Dr. Júlio César Borges Depto. de Química e Física Molecular – DQFM Instituto de Química de São Carlos – IQSC Universidade de São Paulo – USP E-mail: [email protected]
Transcript of Aula09BioqII Metabolismo de Aminoácidos · nitrogenados: heme, aminas biologicamente ativas,...

Aula de Bioquímica II
Tema:
Metabolismo de aminoácidos
Prof. Dr. Júlio César BorgesDepto. de Química e Física Molecular – DQFM
Instituto de Química de São Carlos – IQSC
Universidade de São Paulo – USP
E-mail: [email protected]

Metabolismo do Nitrogênio
���� Nitrogênio: para ser utilizado pelos animais, ele precisa ser fixado- Grande custo energético
� Reduzido de N2 para NH3 (amônia)- Somente algumas poucas bactérias de solo ou que vivem associadas a raízes de plantas
podem converter N2 a NH3- ���� Fixação de nitrogênio
���� As rotas de biossíntese de aminoácido e nucleotídeos necessitam de nitrogênio na forma solúvel e reacional
� Amônia, aminoácidos e nucleotídeos são utilizados de forma econômica pela maioria dos organismos
- Reciclagem ���� menor custo energético- Excedente é eliminado na naturezaPrincipais formas de excreção de N

Degradação oxidativa de Aminoácidos
Energia metabólica gerada nos tecidos
1. Metabolismo dos carboidratos e lipídeos (~90%)
2. Metabolismo dos aminoácidos (~10%)- Derivados de proteínas endógenas e exógenas
���� Carnívoros – podem obter até 90% de energia proveniente de aminoácidos���� Herbívoros – obtém apenas uma pequena fração de suas necessidades energéticas a
partir dessa via���� Microrganismos – retiram aminoácidos do ambiente e podem utilizá-lo como fonte de
energia���� Plantas – utilizado para biossíntese de proteínas. Raramente ou nunca oxidam
aminoácidos
Aminoácidos podem sofrer degradação oxidativa:
1. Dieta rica em proteínas – excedente de aminoácidos podem ser catabolizados e virar gordura ou glicogênio
2. Durante o jejum ou diabetes mellitus – proteínas são utilizadas como combustível
3. Protein turnover – Durante a dinâmica normal de síntese e degradação das proteínas no organismo
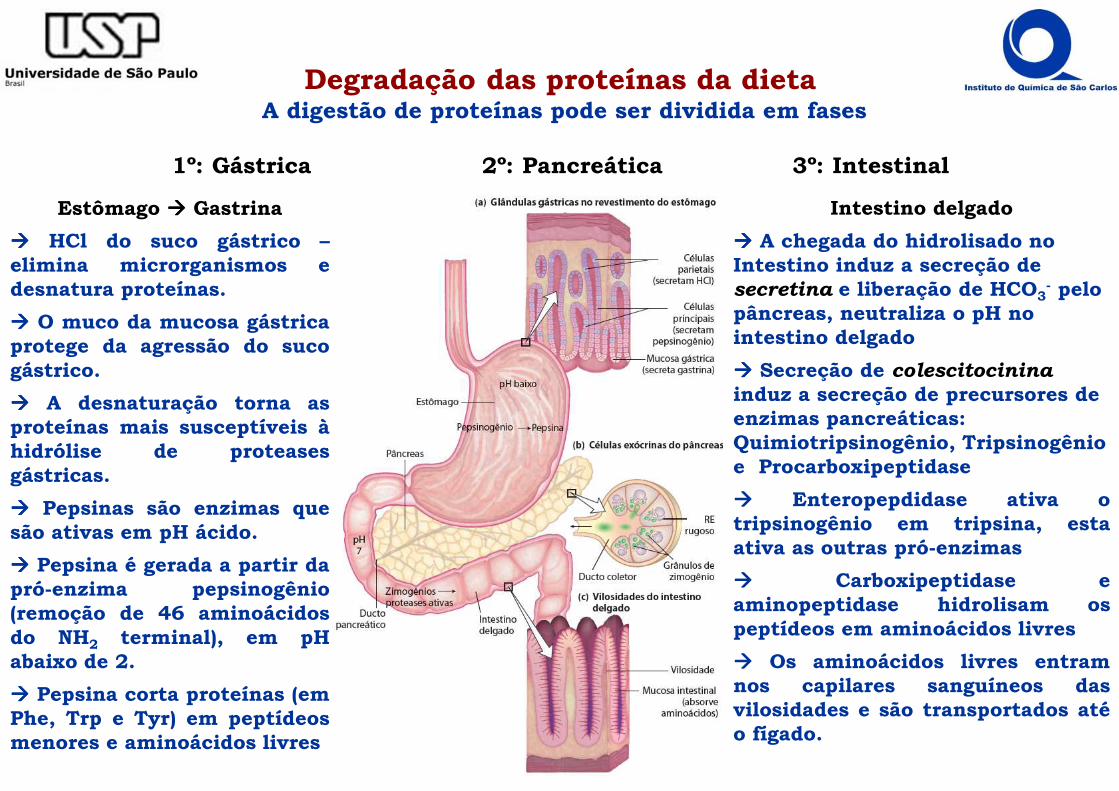
Degradação das proteínas da dietaA digestão de proteínas pode ser dividida em fases
1º: Gástrica 2º: Pancreática 3º: Intestinal
Estômago ���� Gastrina
���� HCl do suco gástrico –elimina microrganismos edesnatura proteínas.
���� O muco da mucosa gástricaprotege da agressão do sucogástrico.
���� A desnaturação torna asproteínas mais susceptíveis àhidrólise de proteasesgástricas.
���� Pepsinas são enzimas quesão ativas em pH ácido.
���� Pepsina é gerada a partir dapró-enzima pepsinogênio(remoção de 46 aminoácidosdo NH2 terminal), em pHabaixo de 2.
���� Pepsina corta proteínas (emPhe, Trp e Tyr) em peptídeosmenores e aminoácidos livres
Intestino delgado
���� A chegada do hidrolisado no Intestino induz a secreção de secretina e liberação de HCO3
- pelo pâncreas, neutraliza o pH no intestino delgado
���� Secreção de colescitocininainduz a secreção de precursores de enzimas pancreáticas: Quimiotripsinogênio, Tripsinogênio e Procarboxipeptidase
���� Enteropepdidase ativa otripsinogênio em tripsina, estaativa as outras pró-enzimas
���� Carboxipeptidase eaminopeptidase hidrolisam ospeptídeos em aminoácidos livres
���� Os aminoácidos livres entramnos capilares sanguíneos dasvilosidades e são transportados atéo fígado.

Renovação de proteínas
� As proteínas estão em constante processo de degradação e síntese.
- A concentração proteica geral mantém-se constante no indivíduo adulto e saudável.
- Entretanto há grande variação na velocidade de degradação para cada proteína.
- Os aminoácidos excedentes são oxidados e o N é excretado ���� NH3 é tóxico.
Renovação
���� Renovação de cerca de 400 g de proteínas/dia em um adulto com dieta adequada
���� Eliminação de N correspondente a 100 g de proteína/dia
� Como 400 g são renovados, os 100 g eliminados devem ser repostos pela alimentação
���� Aminoácidos em excesso não são armazenados para utilização futura e tampouco excretados
São convertidos em intermediários metabólicos, como piruvato, oxalacetato, acetil-coenzima
(Acetil-CoA) e αααα-cetoglutarato
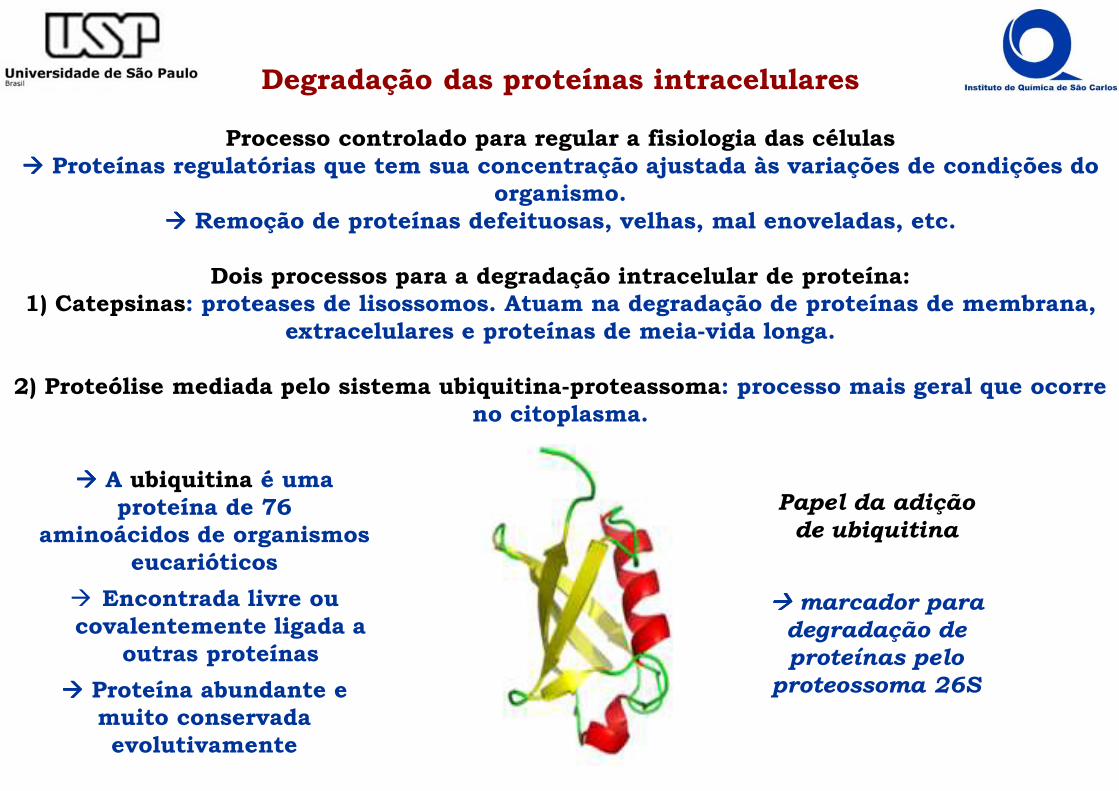
Degradação das proteínas intracelulares
Processo controlado para regular a fisiologia das células���� Proteínas regulatórias que tem sua concentração ajustada às variações de condições do
organismo.���� Remoção de proteínas defeituosas, velhas, mal enoveladas, etc.
Dois processos para a degradação intracelular de proteína:1) Catepsinas: proteases de lisossomos. Atuam na degradação de proteínas de membrana,
extracelulares e proteínas de meia-vida longa.
2) Proteólise mediada pelo sistema ubiquitina-proteassoma: processo mais geral que ocorre no citoplasma.
���� A ubiquitina é uma proteína de 76
aminoácidos de organismos eucarióticos
� Encontrada livre ou covalentemente ligada a
outras proteínas
���� Proteína abundante e muito conservada evolutivamente
Papel da adição de ubiquitina
���� marcador para degradação de proteínas pelo
proteossoma 26S
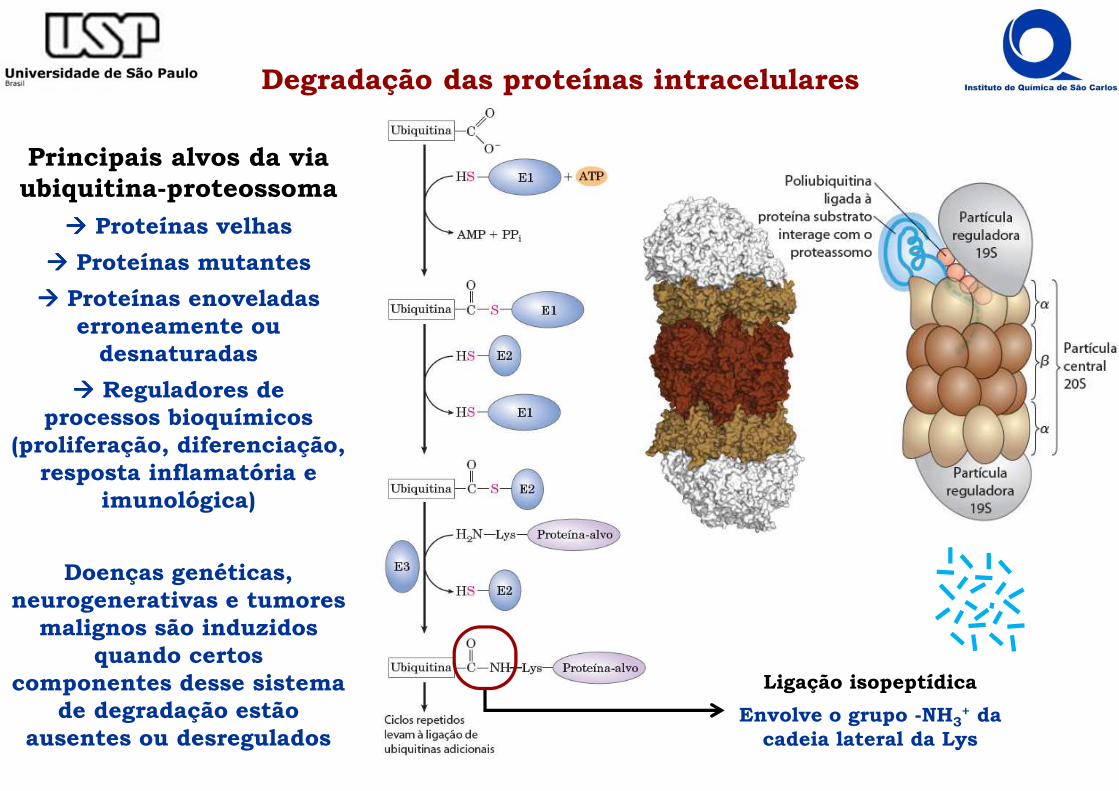
Degradação das proteínas intracelulares
Principais alvos da via ubiquitina-proteossoma
���� Proteínas velhas
���� Proteínas mutantes
���� Proteínas enoveladas erroneamente ou
desnaturadas
���� Reguladores de processos bioquímicos
(proliferação, diferenciação, resposta inflamatória e
imunológica)
Doenças genéticas, neurogenerativas e tumores
malignos são induzidos quando certos
componentes desse sistema de degradação estão
ausentes ou desregulados
Ligação isopeptídica
Envolve o grupo -NH3+ da
cadeia lateral da Lys
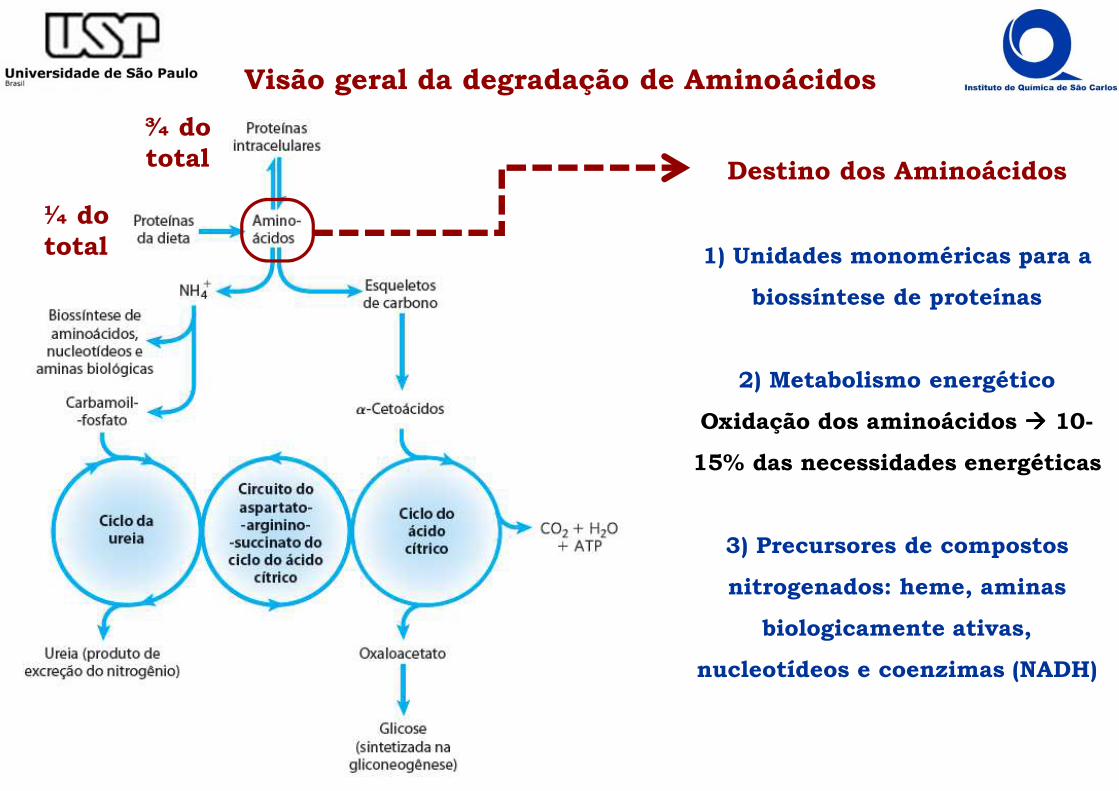
Visão geral da degradação de Aminoácidos
¼ do total
¾ do total Destino dos Aminoácidos
1) Unidades monoméricas para a
biossíntese de proteínas
2) Metabolismo energético
Oxidação dos aminoácidos ���� 10-
15% das necessidades energéticas
3) Precursores de compostos
nitrogenados: heme, aminas
biologicamente ativas,
nucleotídeos e coenzimas (NADH)
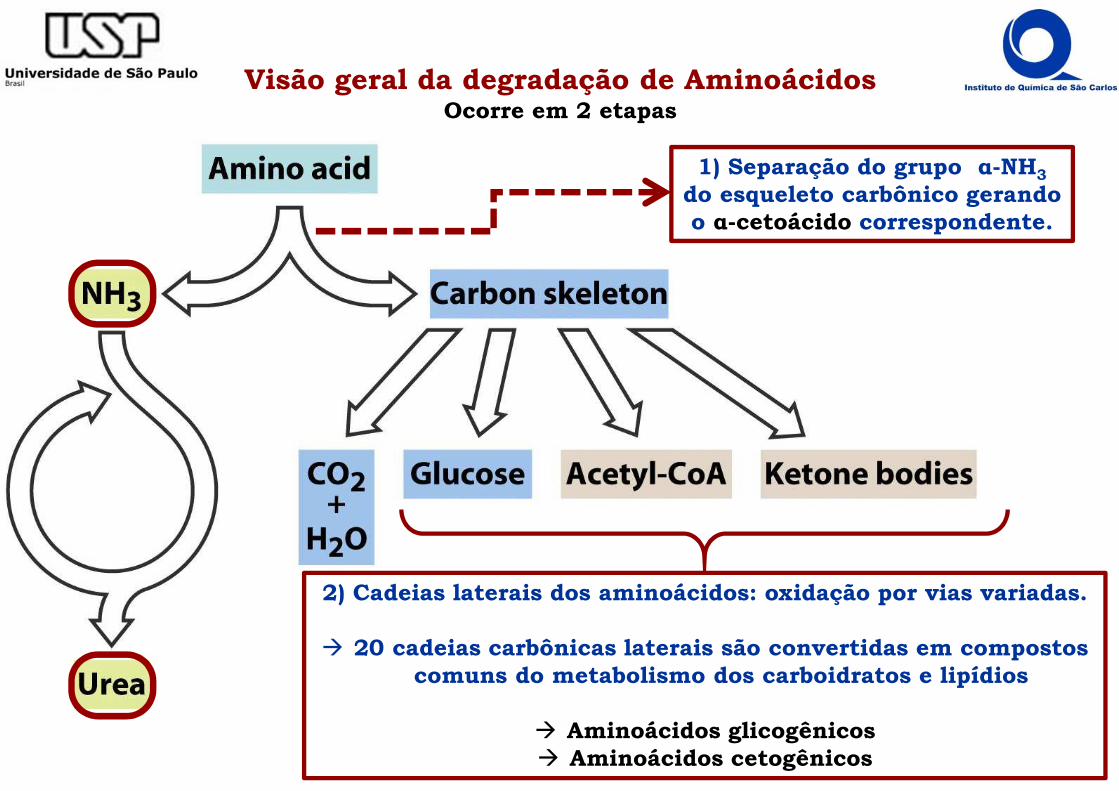
Visão geral da degradação de AminoácidosOcorre em 2 etapas
2) Cadeias laterais dos aminoácidos: oxidação por vias variadas.
� 20 cadeias carbônicas laterais são convertidas em compostos comuns do metabolismo dos carboidratos e lipídios
� Aminoácidos glicogênicos� Aminoácidos cetogênicos
1) Separação do grupo α-NH3
do esqueleto carbônico gerando o α-cetoácido correspondente.
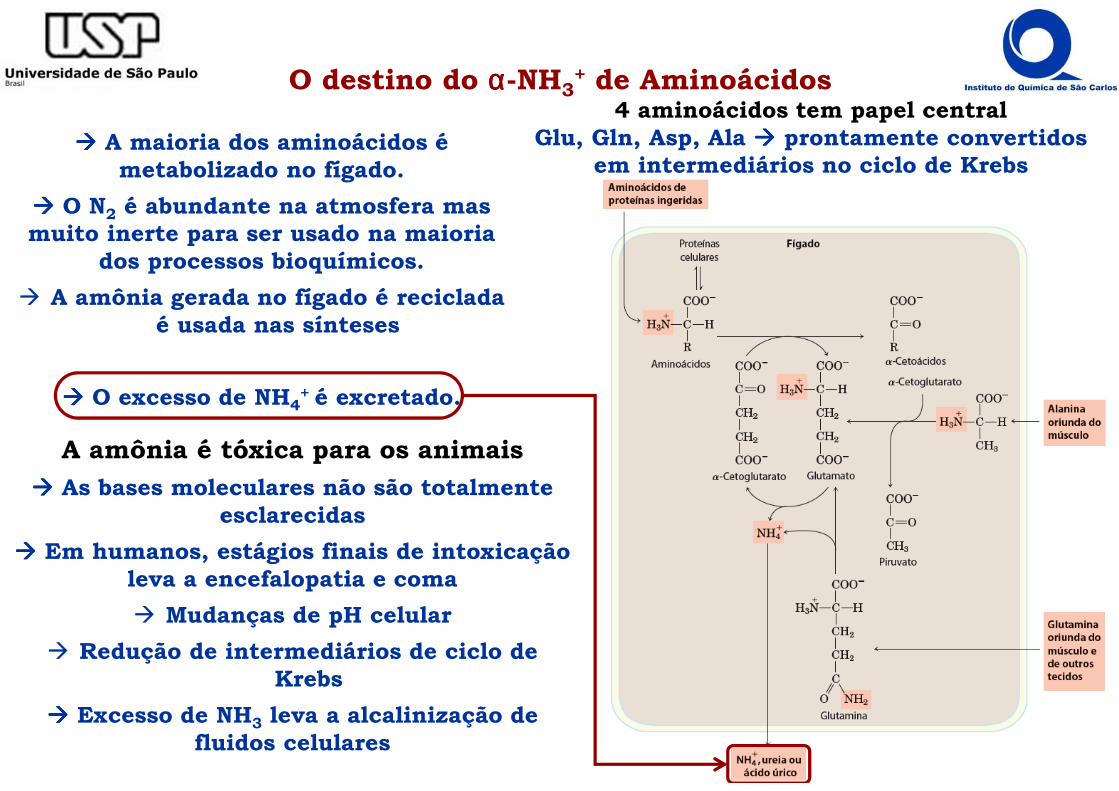
O destino do αααα-NH3+ de Aminoácidos
A amônia é tóxica para os animais
���� As bases moleculares não são totalmente esclarecidas
���� Em humanos, estágios finais de intoxicação leva a encefalopatia e coma
� Mudanças de pH celular
� Redução de intermediários de ciclo de Krebs
���� Excesso de NH3 leva a alcalinização de fluidos celulares
���� A maioria dos aminoácidos é metabolizado no fígado.
���� O N2 é abundante na atmosfera mas muito inerte para ser usado na maioria
dos processos bioquímicos.
� A amônia gerada no fígado é reciclada é usada nas sínteses
���� O excesso de NH4+ é excretado.
4 aminoácidos tem papel centralGlu, Gln, Asp, Ala ���� prontamente convertidos
em intermediários no ciclo de Krebs
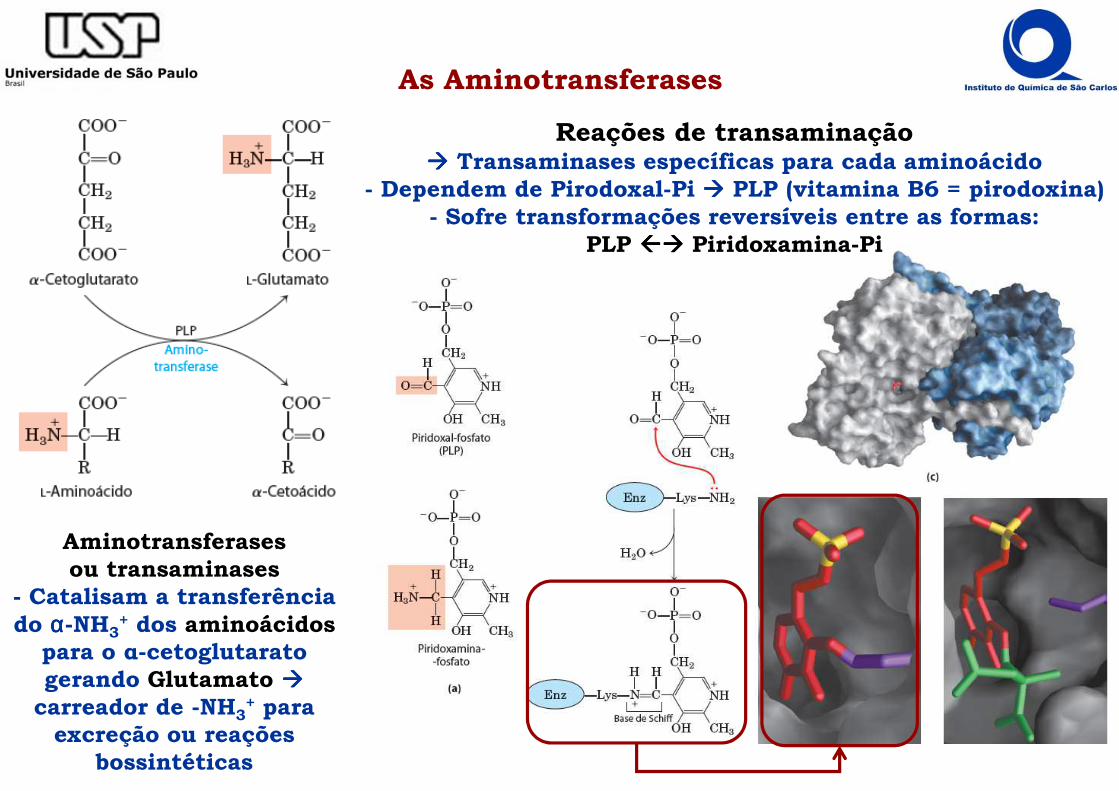
As Aminotransferases
Aminotransferasesou transaminases
- Catalisam a transferência do αααα-NH3
+ dos aminoácidospara o α-cetoglutaratogerando Glutamato ����
carreador de -NH3+ para
excreção ou reações bossintéticas
Reações de transaminação���� Transaminases específicas para cada aminoácido
- Dependem de Pirodoxal-Pi ���� PLP (vitamina B6 = pirodoxina)- Sofre transformações reversíveis entre as formas:
PLP ���� Piridoxamina-Pi
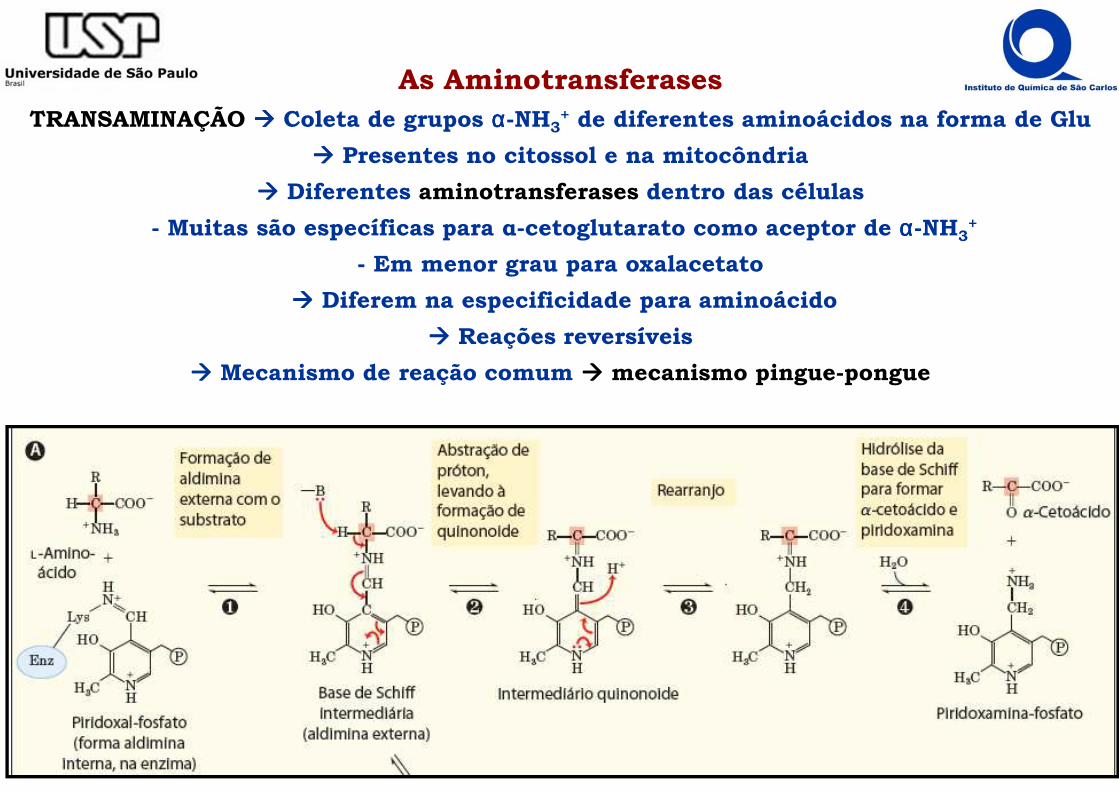
As AminotransferasesTRANSAMINAÇÃO ���� Coleta de grupos αααα-NH3
+ de diferentes aminoácidos na forma de Glu
���� Presentes no citossol e na mitocôndria
���� Diferentes aminotransferases dentro das células
- Muitas são específicas para α-cetoglutarato como aceptor de αααα-NH3+
- Em menor grau para oxalacetato
���� Diferem na especificidade para aminoácido
���� Reações reversíveis
���� Mecanismo de reação comum ���� mecanismo pingue-pongue
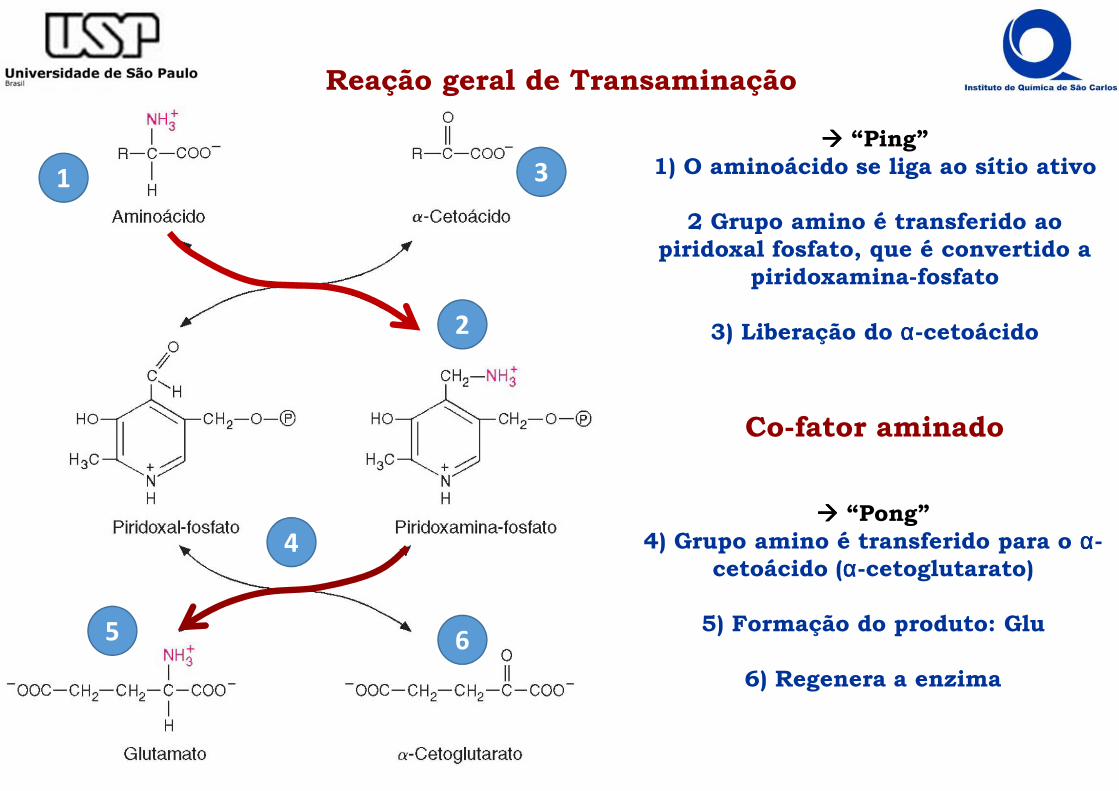
Reação geral de Transaminação
���� “Ping”1) O aminoácido se liga ao sítio ativo
2 Grupo amino é transferido ao piridoxal fosfato, que é convertido a
piridoxamina-fosfato
3) Liberação do αααα-cetoácido
���� “Pong”4) Grupo amino é transferido para o αααα-
cetoácido (αααα-cetoglutarato)
5) Formação do produto: Glu
6) Regenera a enzima
Co-fator aminado
1
2
3
4
5 6
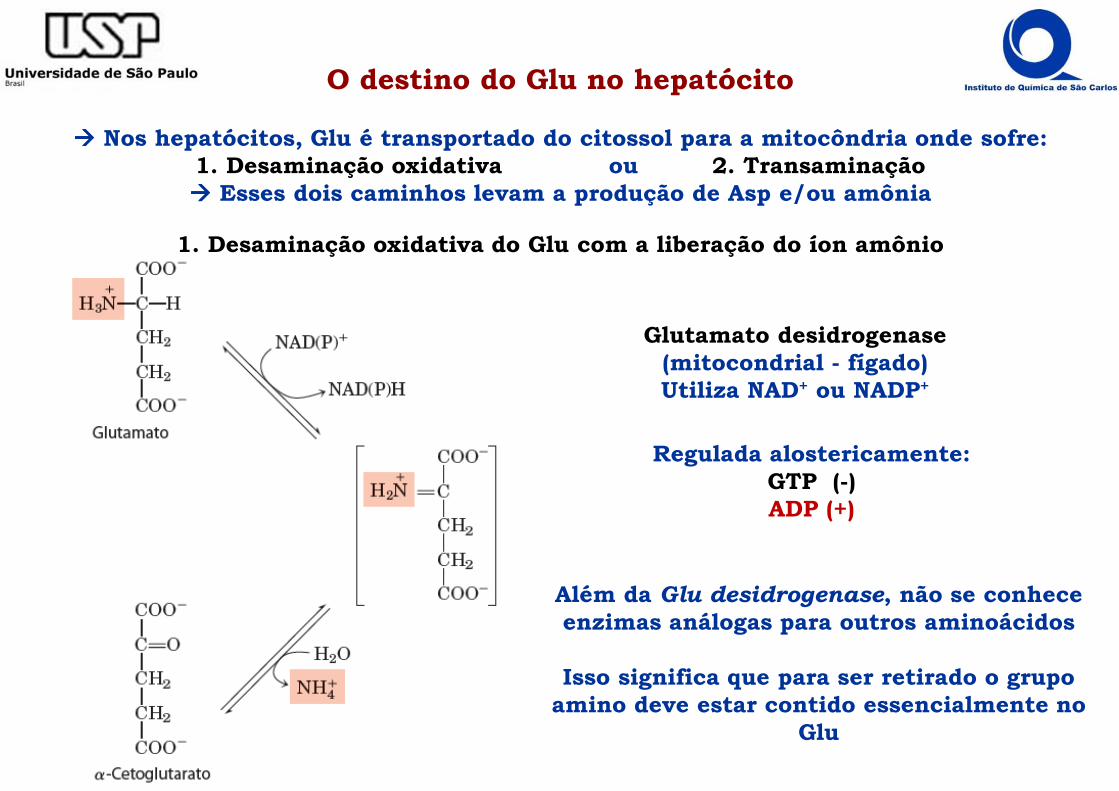
O destino do Glu no hepatócito
���� Nos hepatócitos, Glu é transportado do citossol para a mitocôndria onde sofre:1. Desaminação oxidativa ou 2. Transaminação���� Esses dois caminhos levam a produção de Asp e/ou amônia
1. Desaminação oxidativa do Glu com a liberação do íon amônio
Glutamato desidrogenase(mitocondrial - fígado)Utiliza NAD+ ou NADP+
Além da Glu desidrogenase, não se conhece enzimas análogas para outros aminoácidos
Isso significa que para ser retirado o grupo amino deve estar contido essencialmente no
Glu
Regulada alostericamente:GTP (-)ADP (+)
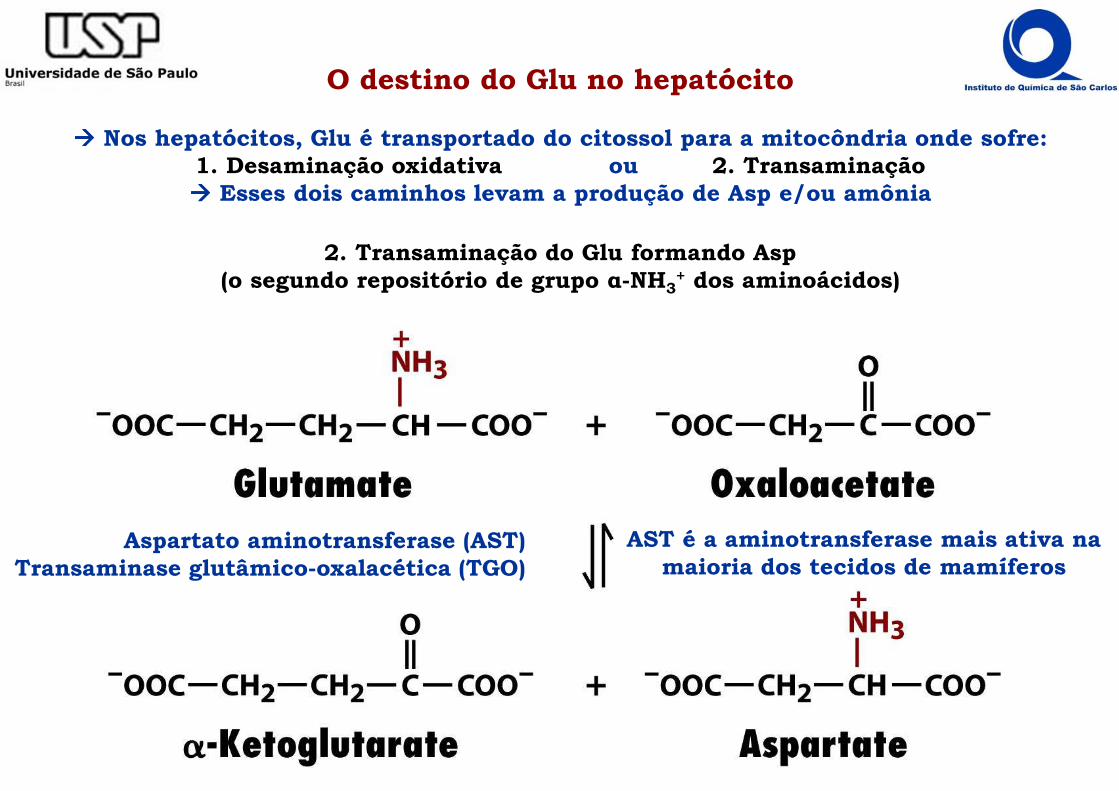
O destino do Glu no hepatócito
���� Nos hepatócitos, Glu é transportado do citossol para a mitocôndria onde sofre:1. Desaminação oxidativa ou 2. Transaminação���� Esses dois caminhos levam a produção de Asp e/ou amônia
2. Transaminação do Glu formando Asp(o segundo repositório de grupo α-NH3
+ dos aminoácidos)
Aspartato aminotransferase (AST)Transaminase glutâmico-oxalacética (TGO)
AST é a aminotransferase mais ativa na maioria dos tecidos de mamíferos
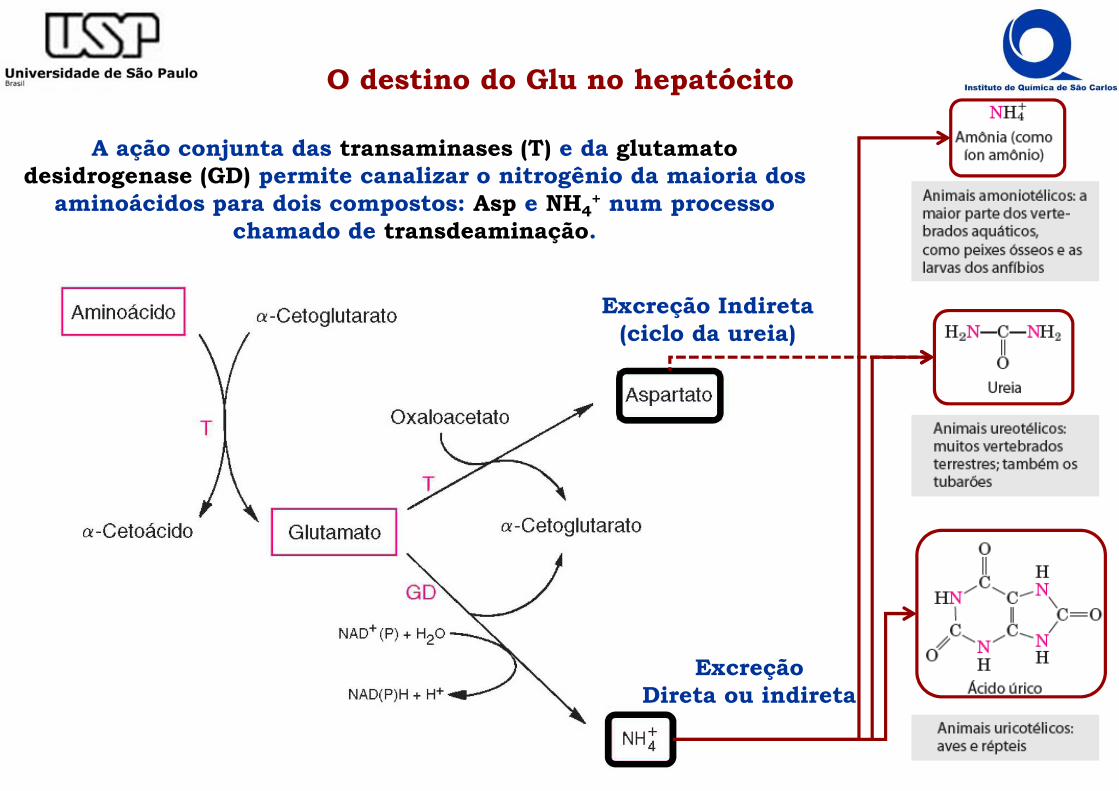
O destino do Glu no hepatócito
A ação conjunta das transaminases (T) e da glutamato desidrogenase (GD) permite canalizar o nitrogênio da maioria dos
aminoácidos para dois compostos: Asp e NH4+ num processo
chamado de transdeaminação.
ExcreçãoDireta ou indireta
Excreção Indireta(ciclo da ureia)
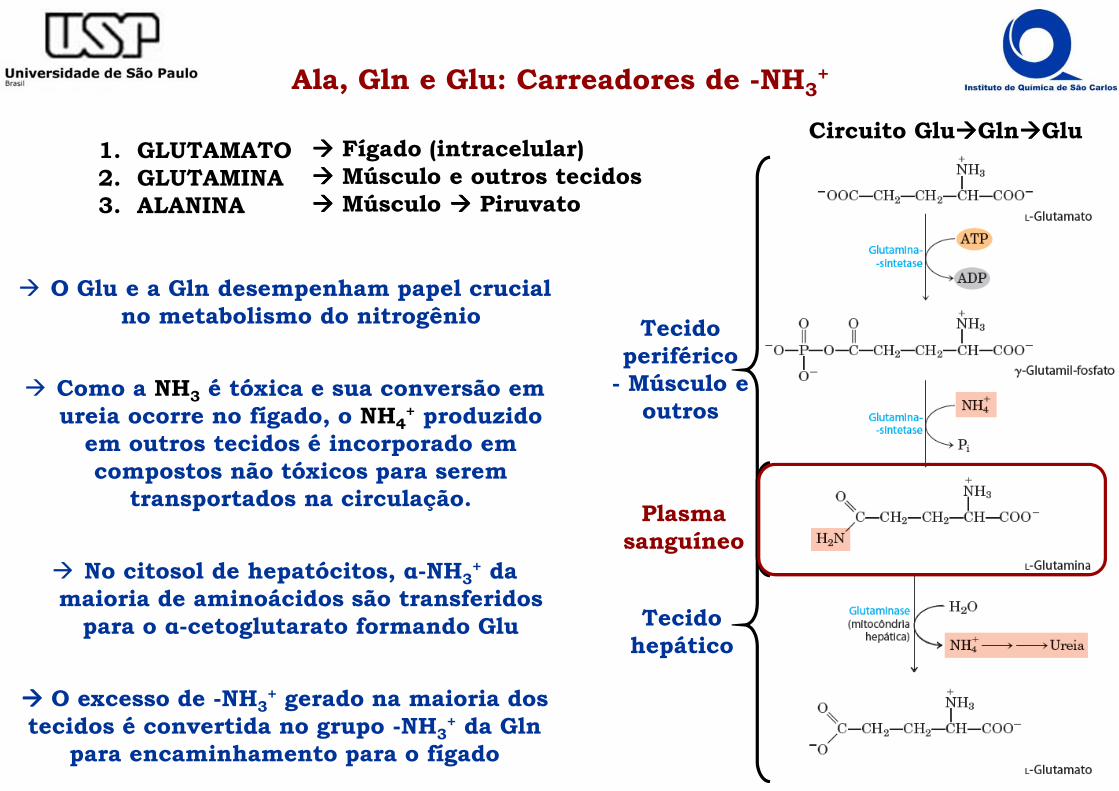
Ala, Gln e Glu: Carreadores de -NH3+
Plasma sanguíneo
Tecido hepático
Circuito Glu����Gln����Glu
Tecido periférico
- Músculo e outros
� O Glu e a Gln desempenham papel crucial no metabolismo do nitrogênio
� Como a NH3 é tóxica e sua conversão em ureia ocorre no fígado, o NH4
+ produzido em outros tecidos é incorporado em compostos não tóxicos para serem
transportados na circulação.
� No citosol de hepatócitos, α-NH3+ da
maioria de aminoácidos são transferidos para o α-cetoglutarato formando Glu
���� O excesso de -NH3+ gerado na maioria dos
tecidos é convertida no grupo -NH3+ da Gln
para encaminhamento para o fígado
1. GLUTAMATO2. GLUTAMINA3. ALANINA
���� Fígado (intracelular)���� Músculo e outros tecidos���� Músculo ���� Piruvato
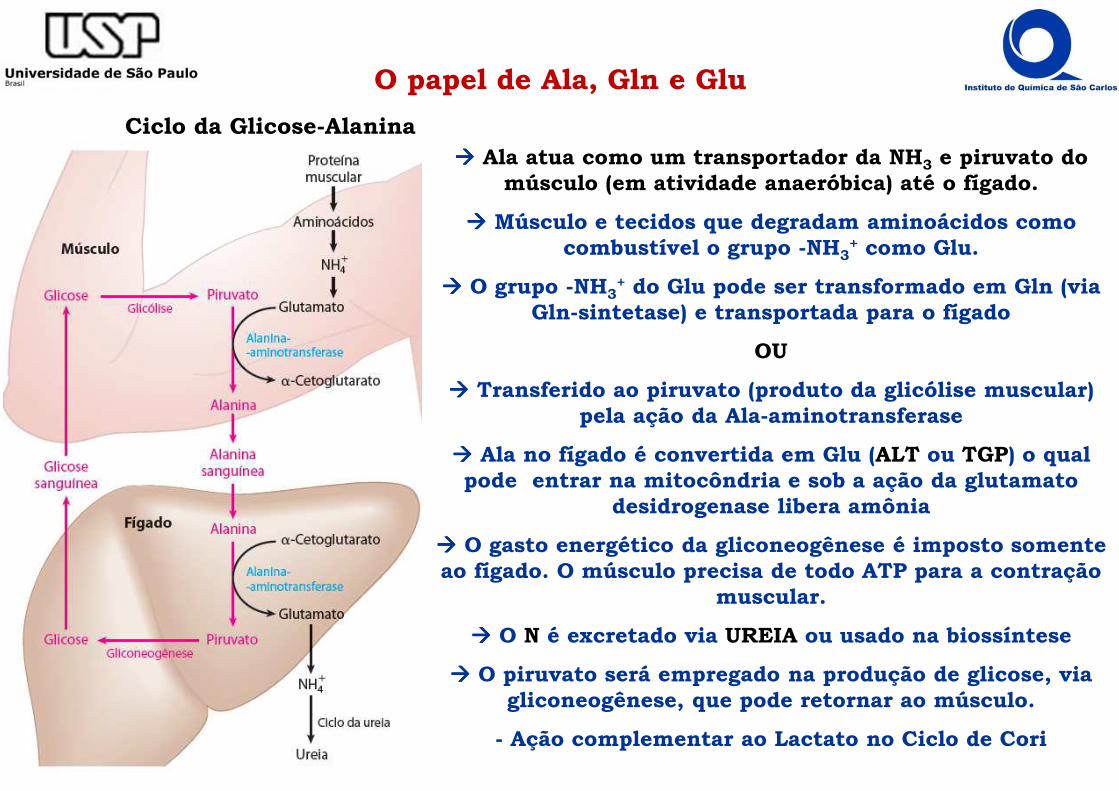
O papel de Ala, Gln e Glu
Ciclo da Glicose-Alanina���� Ala atua como um transportador da NH3 e piruvato do
músculo (em atividade anaeróbica) até o fígado.
���� Músculo e tecidos que degradam aminoácidos como combustível o grupo -NH3
+ como Glu.
���� O grupo -NH3+ do Glu pode ser transformado em Gln (via
Gln-sintetase) e transportada para o fígado
OU
���� Transferido ao piruvato (produto da glicólise muscular) pela ação da Ala-aminotransferase
���� Ala no fígado é convertida em Glu (ALT ou TGP) o qual pode entrar na mitocôndria e sob a ação da glutamato
desidrogenase libera amônia
���� O gasto energético da gliconeogênese é imposto somente ao fígado. O músculo precisa de todo ATP para a contração
muscular.
���� O N é excretado via UREIA ou usado na biossíntese
���� O piruvato será empregado na produção de glicose, via gliconeogênese, que pode retornar ao músculo.
- Ação complementar ao Lactato no Ciclo de Cori

O destino final do NH3 em ureotélicos:
Excreção na forma de ureia
���� Animais terrestres necessitam de vias metabólicas para a excreção de N e diminuição da toxicidade da NH3 e diminuição da perda de água
���� Plantas reciclam essencialmente todos os grupos NH3 e a excreção de N ocorrem em condições específicas
���� Nos animais ureotélicos a NH3 atinge a mitocôndria dos hepatócitos será convertida em UREIA
���� Ciclo da UREIA ou ciclo Krebs-Henseleit (1932; 1° ciclo metabólico a ser descrito!!)
���� No ciclo da UREIA um dos átomos de N é derivado do Asp, o outro é derivado do próprio NH4
+ livre.
���� O átomo de C vem do bicarbonato (HCO3-) oriundo do metabolismo oxidativo.
���� A UREIA, que tem alta solubilidade (~10-11 mol/L), atinge a circulação e chega aos rins onde é excretada na urina
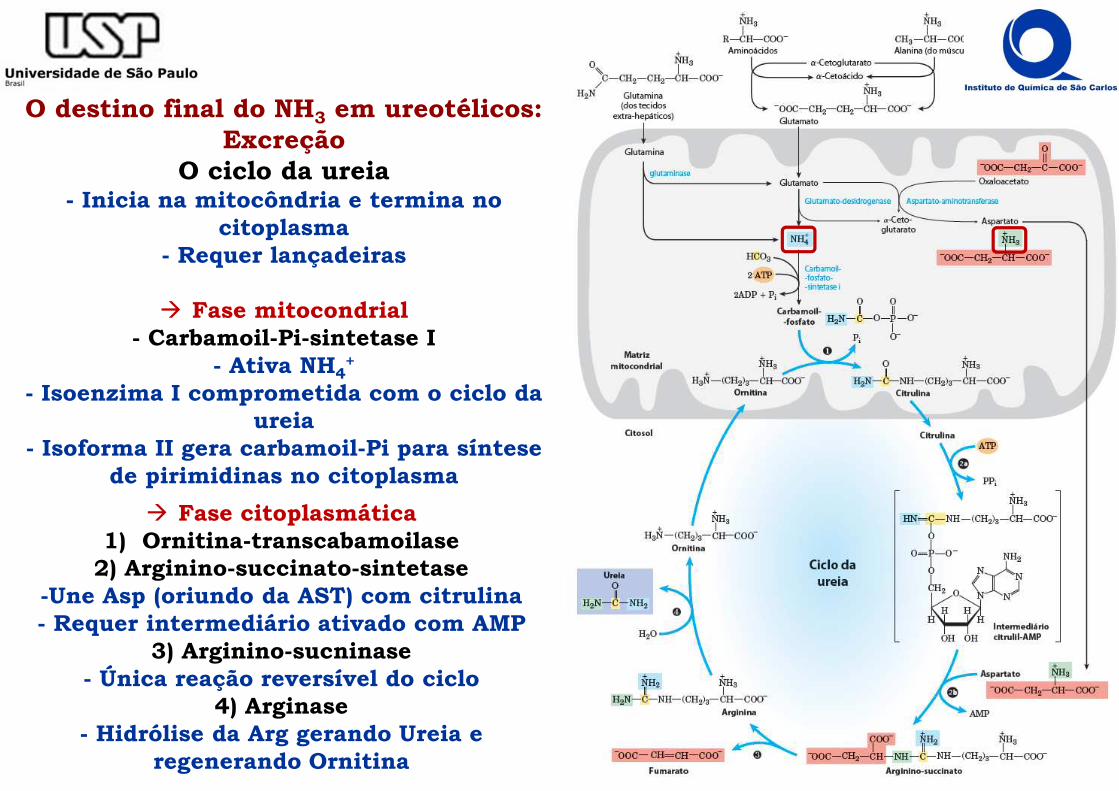
O destino final do NH3 em ureotélicos: Excreção
O ciclo da ureia- Inicia na mitocôndria e termina no
citoplasma- Requer lançadeiras
� Fase mitocondrial- Carbamoil-Pi-sintetase I
- Ativa NH4+
- Isoenzima I comprometida com o ciclo da ureia
- Isoforma II gera carbamoil-Pi para síntese de pirimidinas no citoplasma
� Fase citoplasmática1) Ornitina-transcabamoilase
2) Arginino-succinato-sintetase-Une Asp (oriundo da AST) com citrulina- Requer intermediário ativado com AMP
3) Arginino-sucninase- Única reação reversível do ciclo
4) Arginase- Hidrólise da Arg gerando Ureia e
regenerando Ornitina
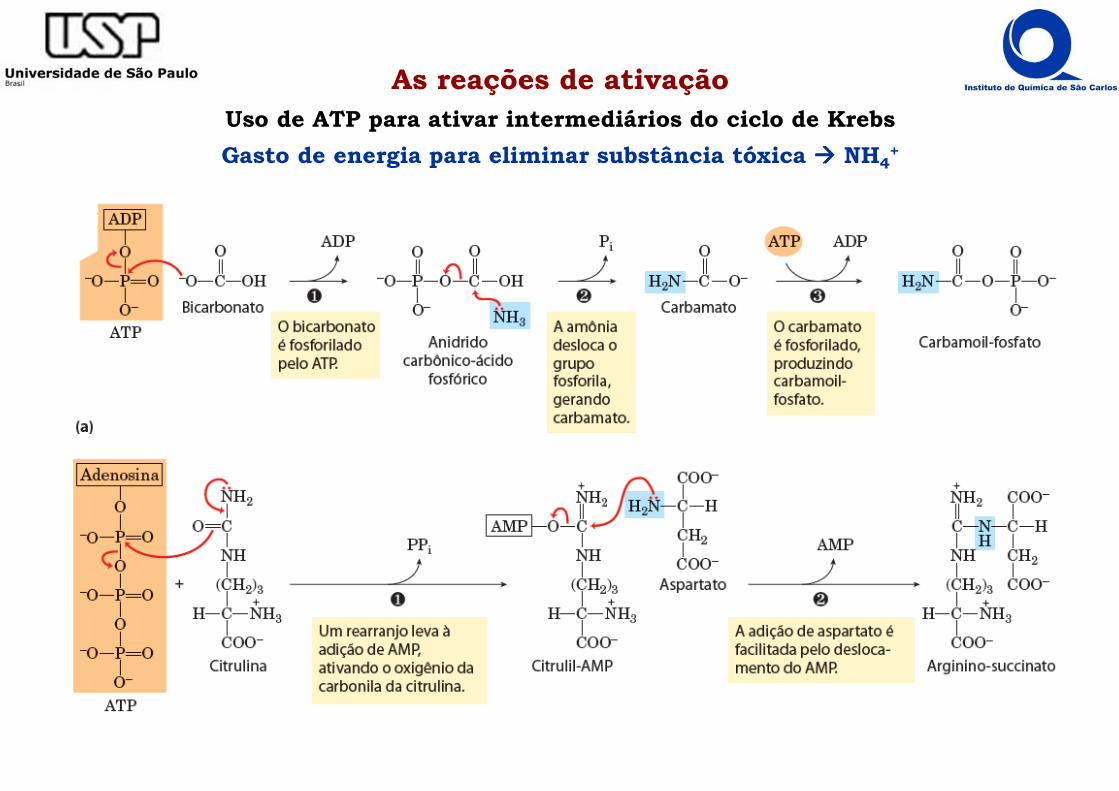
As reações de ativaçãoUso de ATP para ativar intermediários do ciclo de Krebs
Gasto de energia para eliminar substância tóxica ���� NH4+
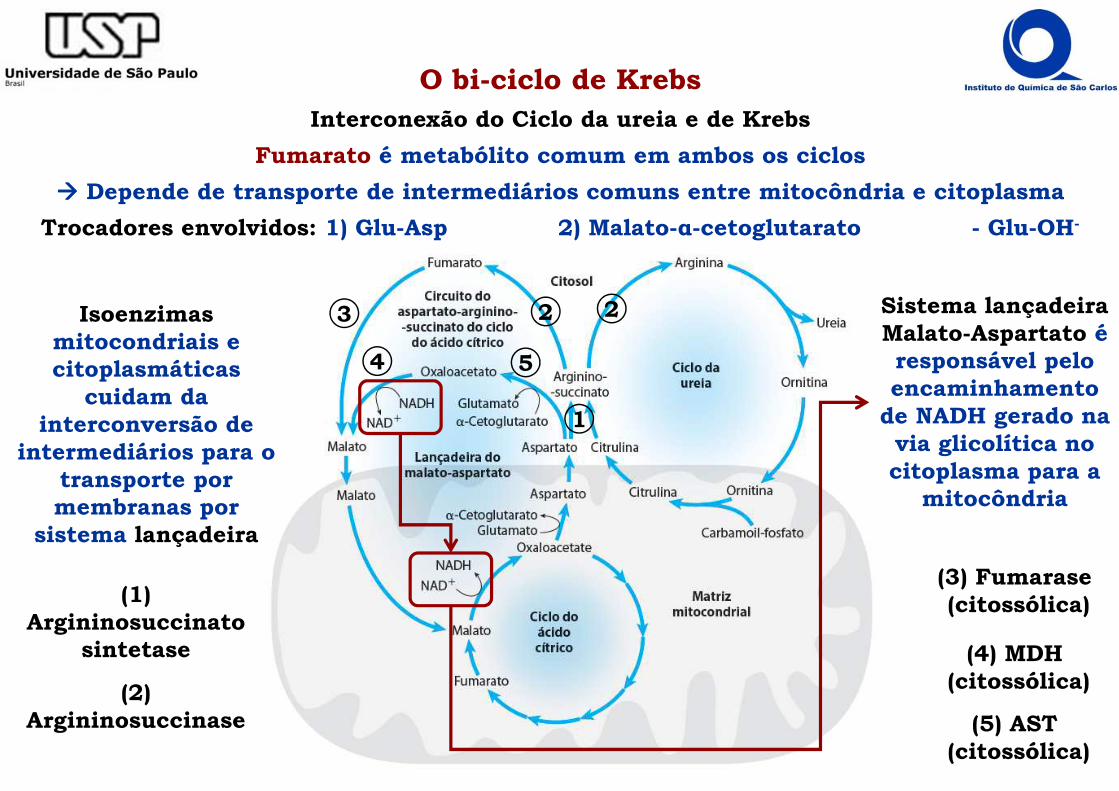
O bi-ciclo de KrebsInterconexão do Ciclo da ureia e de Krebs
Fumarato é metabólito comum em ambos os ciclos
���� Depende de transporte de intermediários comuns entre mitocôndria e citoplasma
Trocadores envolvidos: 1) Glu-Asp 2) Malato-α-cetoglutarato - Glu-OH-
Isoenzimasmitocondriais e citoplasmáticas
cuidam da interconversão de
intermediários para o transporte por membranas por
sistema lançadeira
Sistema lançadeiraMalato-Aspartato é responsável pelo encaminhamento
de NADH gerado na via glicolítica no citoplasma para a
mitocôndria
(1) Argininosuccinato
sintetase
(2)Argininosuccinase
1
22
(3) Fumarase(citossólica)
3
(4) MDH(citossólica)
4 5
(5) AST(citossólica)

Regulação do ciclo da ureia
Regulação de curto prazo���� Regulação alostérica da carbamoil fosfato
sintetase I- Estimulo por N-acetilglutamato (produzido a partir de condensação de glutamato e acetil-
CoA pela N-acetilglutamato sintase).
Regulação a longo prazo
1) A quantidade de ureia excretada em humanos é dependente da dieta
- ↑ em condições alta ingestão proteica –produção de ácidos graxos a partir dos α-
cetoácidos correspondentes dos aminoácidos ingeridos.
- ↑ em condições de jejum prolongado onde as cadeias carbônicas dos aminoácidos são
utilizadas como precursores gliconeogênicos.
���� Nessas condições há ↑ da síntese das enzimas do ciclo da ureia e da carbamoil
fosfato sintetase I (elevação de cerca de 10 a 20 vezes).
2) Diminui em condições de dieta pobre em proteínas e rica em carboidratos e
lipídios
Redução da concentração das enzimas envolvidas no ciclo
- A N-acetilglutamatosintase é ativada por arginina, que
sinaliza acúmulo de intermediários do ciclo da ureia e
excesso de NH4+.
- A Arg adequa a velocidade de
formação de NH4+ a
sua conversão em ureia.

Balanço energéticoA síntese de uma molécula de ureia consome 4 ligações Pi
- 2 ATP para a síntese de carbamoil-Pi
- 2 equivalentes ATP: ativação citrulina em citruil-AMP libera PPi
Pirofosfatase alcalina hidroliza PPi em 2 Pi
2 NH4+ + 1 HCO3
- + 3 ATP4- + 1 H2O → 1 ureia + 2 ADP3- + 4 Pi2- + 1 AMP2- + 2 H+
O custo é substancialmente reduzindo pelo acoplamento do transporte de equivalentes redutor na forma de NADH para a mitocôndria pela lançadeira Malato-aspartato
1 NADH = 2,5 ATP na fosforilação oxidativa
Hiperamonemia� Depleção de intermediários do ciclo de Krebs e diminuição da taxa de oxidação da
glicose ���� vital para o tecido cerebral.- Alto consumo de α-cetoglutarato para a síntese de Glu.
� Comprometimento da transmissão do impulso nervoso e encefalopatia via ação exacerbada da transmissão GABAérgica ���� alta [Glu].
� Falhas genéticas ocasionam hiperamonemia ou aumento dos intermediários- Intolerância a dietas proteicas
- Reações irreversíveis causam aumento dos intermediários do ciclo- Dieta deve ser suprida com os aminoácidos essenciais
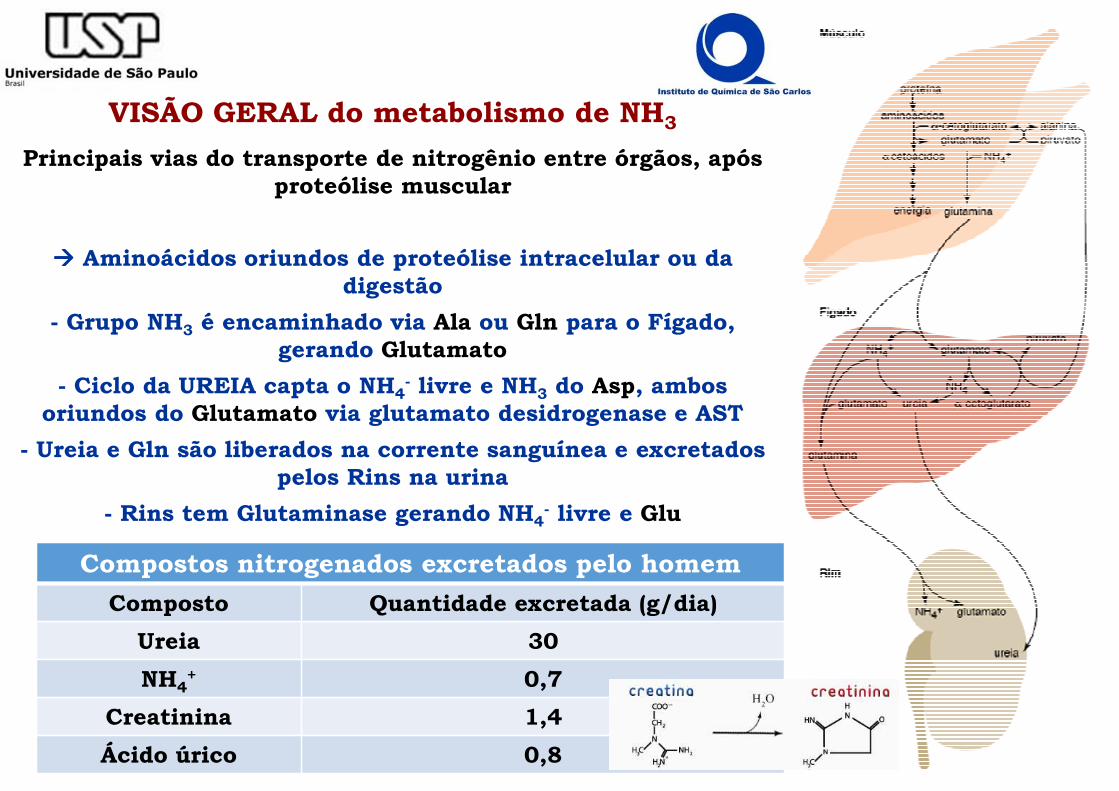
VISÃO GERAL do metabolismo de NH3
Principais vias do transporte de nitrogênio entre órgãos, após proteólise muscular
���� Aminoácidos oriundos de proteólise intracelular ou da digestão
- Grupo NH3 é encaminhado via Ala ou Gln para o Fígado, gerando Glutamato
- Ciclo da UREIA capta o NH4- livre e NH3 do Asp, ambos
oriundos do Glutamato via glutamato desidrogenase e AST
- Ureia e Gln são liberados na corrente sanguínea e excretados pelos Rins na urina
- Rins tem Glutaminase gerando NH4- livre e Glu
Compostos nitrogenados excretados pelo homem
Composto Quantidade excretada (g/dia)
Ureia 30
NH4+ 0,7
Creatinina 1,4
Ácido úrico 0,8

Degradação da cadeia carbônica dos aminoácidos
���� Rende 10-15% da energia extraída pelo organismo
���� Removido o grupo NH3, resta o esqueleto carbônico do aminoácido na forma de α-cetoácido.
���� As 20 cadeias carbônicas são oxidadas por vias próprias, porém convergem para 6
componentes do metabolismo: piruvato, acetil CoA, oxaloacetato, fumarato, α-cetoglutarato e succinato (4 últimos são intermediários do ciclo de Krebs).
� A partir desse ponto o metabolismo da cadeia carbônica dos aminoácidos confunde-se com o dos carboidratos ou ácidos graxos.
� Um mesmo aminoácido pode contribuir para diferentes componentes do metabolismo.
���� O destino final do α-cetoácido depende do tecido e do estado fisiológico podendo ser:
- Oxidado pelo ciclo de Krebs
- Utilizado na Gliconeogênese
- Conversão em triacilglicerol
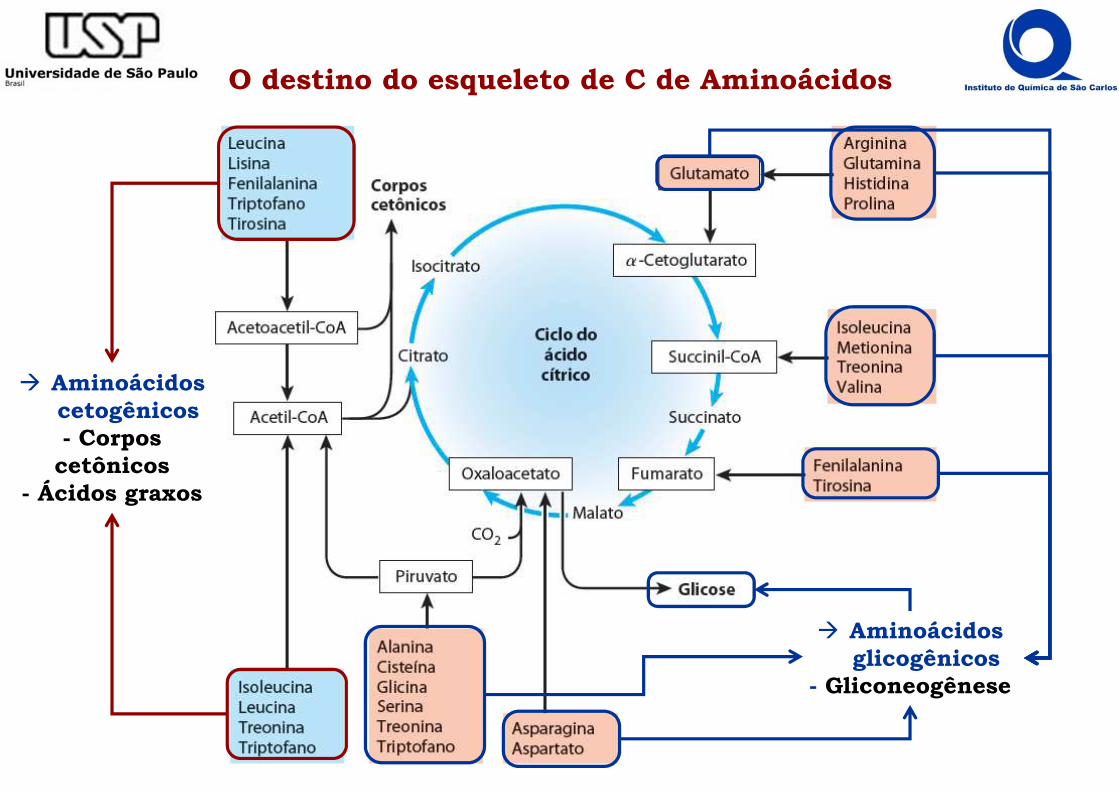
O destino do esqueleto de C de Aminoácidos
� Aminoácidos cetogênicos- Corpos
cetônicos- Ácidos graxos
� Aminoácidos glicogênicos
- Gliconeogênese
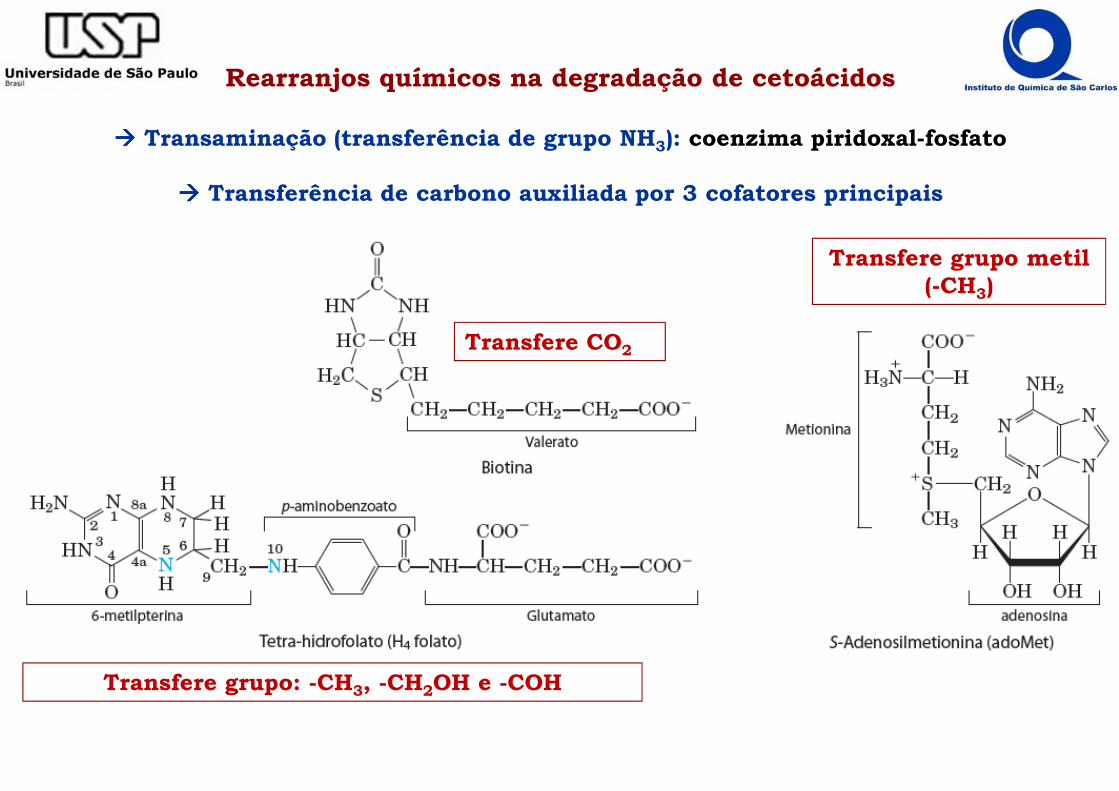
Rearranjos químicos na degradação de cetoácidos
���� Transaminação (transferência de grupo NH3): coenzima piridoxal-fosfato
���� Transferência de carbono auxiliada por 3 cofatores principais
Transfere CO2
Transfere grupo metil (-CH3)
Transfere grupo: -CH3, -CH2OH e -COH
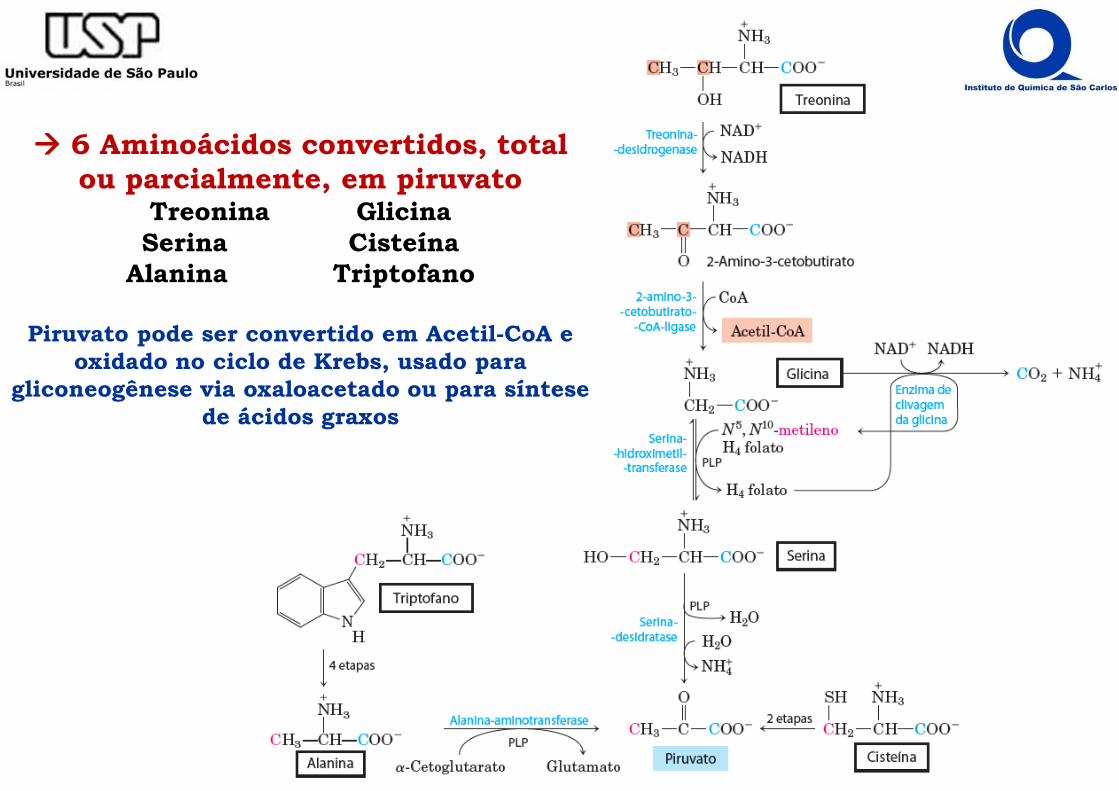
���� 6 Aminoácidos convertidos, total ou parcialmente, em piruvato
Treonina GlicinaSerina Cisteína
Alanina Triptofano
Piruvato pode ser convertido em Acetil-CoA e oxidado no ciclo de Krebs, usado para
gliconeogênese via oxaloacetado ou para síntese de ácidos graxos
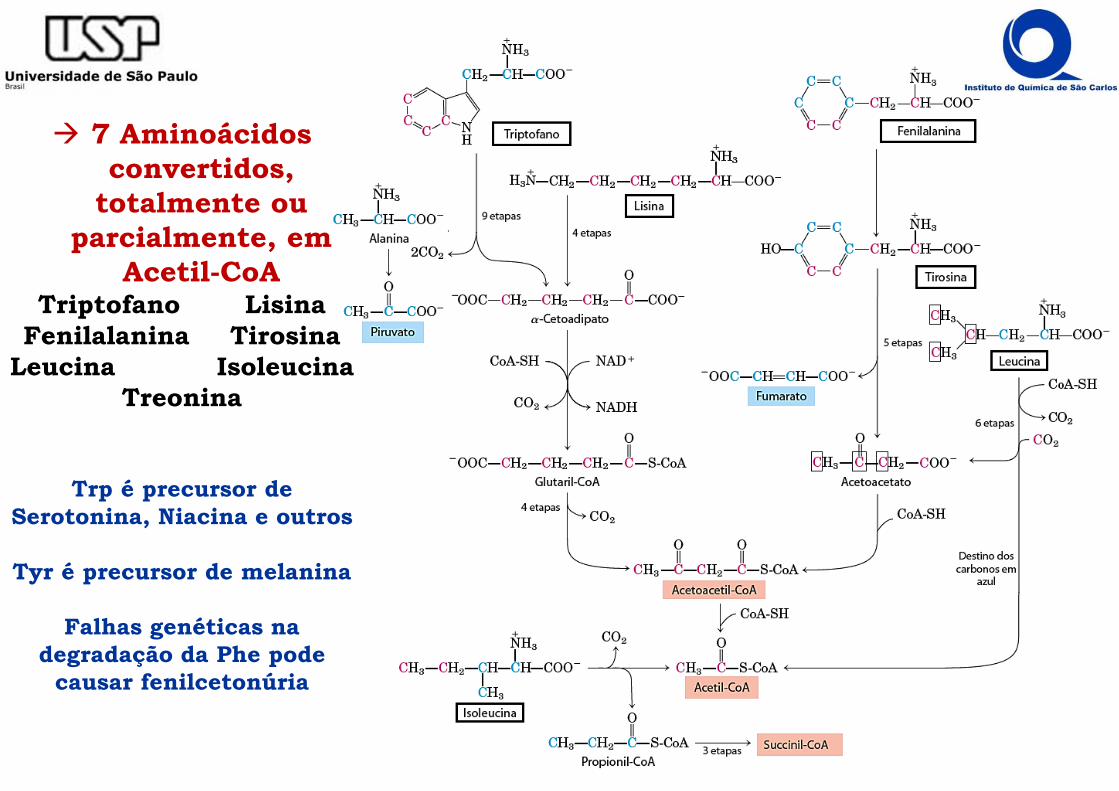
� 7 Aminoácidos convertidos,
totalmente ou parcialmente, em
Acetil-CoATriptofano Lisina
Fenilalanina TirosinaLeucina Isoleucina
Treonina
Trp é precursor de Serotonina, Niacina e outros
Tyr é precursor de melanina
Falhas genéticas na degradação da Phe pode
causar fenilcetonúria
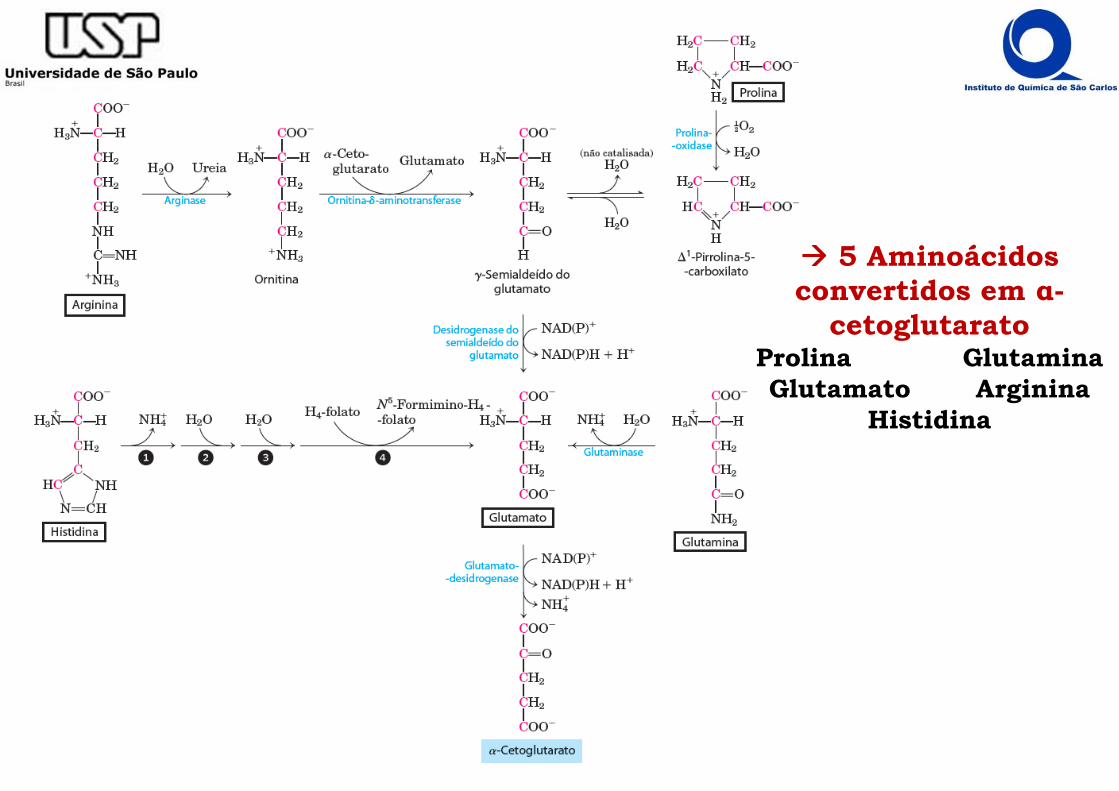
���� 5 Aminoácidos convertidos em α-
cetoglutaratoProlina GlutaminaGlutamato Arginina
Histidina
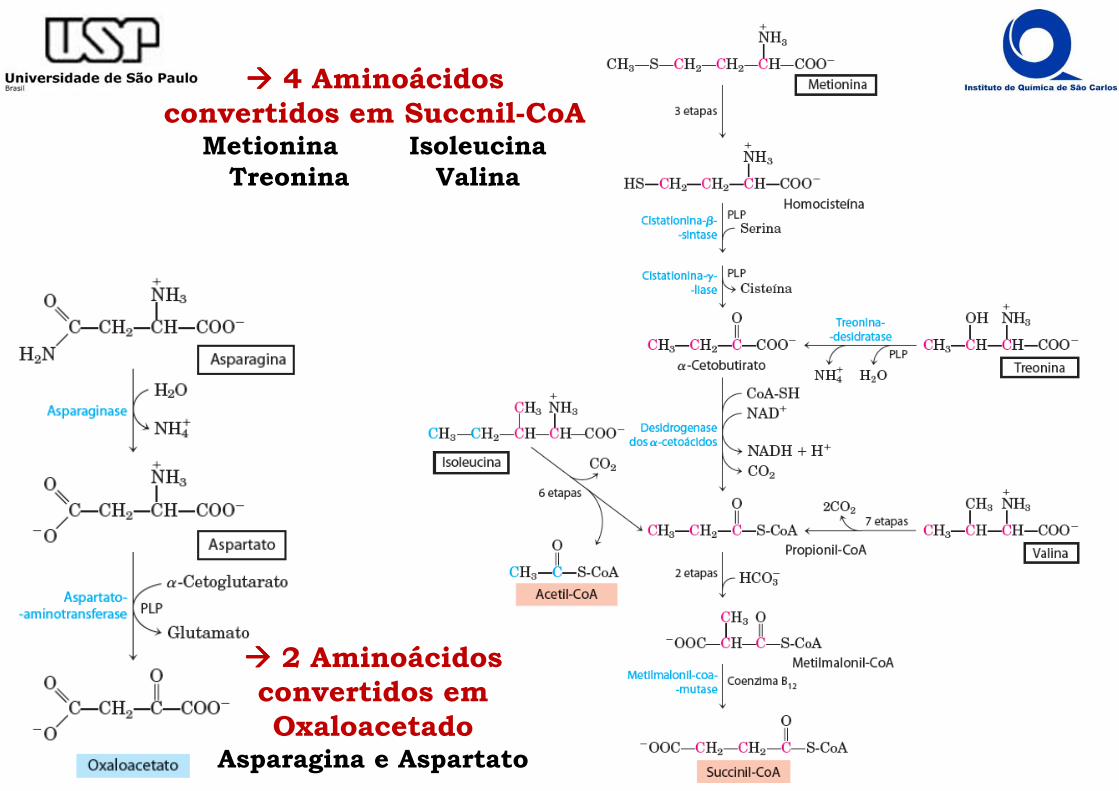
���� 4 Aminoácidos convertidos em Succnil-CoA
Metionina IsoleucinaTreonina Valina
���� 2 Aminoácidos convertidos em Oxaloacetado
Asparagina e Aspartato
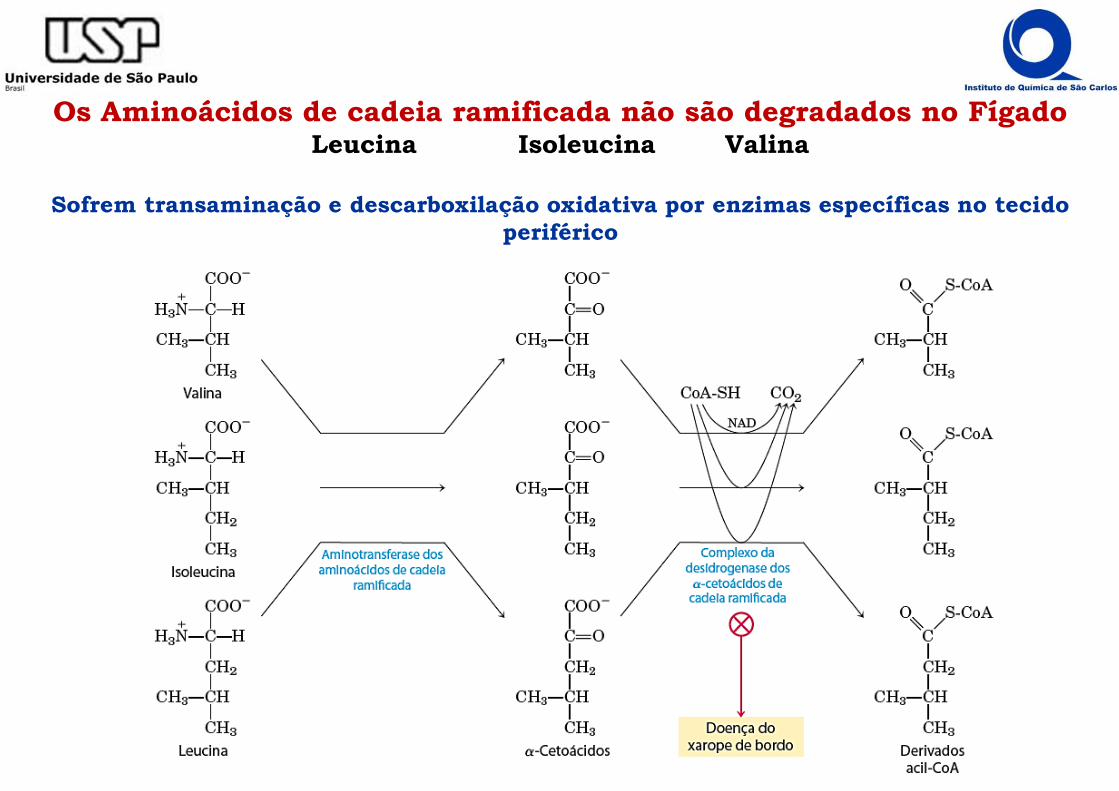
Os Aminoácidos de cadeia ramificada não são degradados no FígadoLeucina Isoleucina Valina
Sofrem transaminação e descarboxilação oxidativa por enzimas específicas no tecido periférico

A origem do Nitrogênio das moléculas orgânicas
���� Os organismos apresentam dependência variada do meio ambiente com relação ao suprimento de aminoácidos
���� A principal fonte de nitrogênio para os seres vivos é o N2 atmosférico- Diferentemente do O2, o N2 é um gás pouco reativo
���� Para ser assimilado pelas células, o N2 precisa ser convertido em NH3
- Processo chamado de Fixação de nitrogênio
� 30-40% são fixados por:- Processo naturais, não biológicos como descargas elétricas (raios) e radiação UV (10-15%)
- Processos industriais (fertilizantes) (25%) ���� processo Haber-Bosch: ~10% da energia industrial mundial

A origem do Nitrogênio das moléculas orgânicas
Fixação de nitrogênio� 60-70% do nitrogênio disponível para os seres vivos são fixados por bactérias
- bactérias fixadoras de N2 compreendem um grande número de espécies que habitam os mais diversos nichos ecológicos (solo, oceano, rios, etc.)
���� Vegetais e muitas bactérias conseguem sintetizar todos os
aminoácidos sendo o grupo NH3+ obtido
a partir de NH3 e a cadeia carbônica a partir de carboidratos
As bactérias, produzem um excesso de NH3 que é liberado para o solo.
� Bactérias da família Rhizobiaceae invadem as raízes de leguninosas, induzindo a formação de nódulos complexos, responsáveis pela fixação do nitrogênio.
���� Bactérias endofíticas como as do gênero Burkholderia ganham acesso ao sistema vascular de não-leguminosas como gramíneas (milho, arroz, gramas, etc)
����Bactérias de vida livre como Azobacter
���� A fixação por simbiose é mais eficiente que a realizada por bactérias de vida livre.
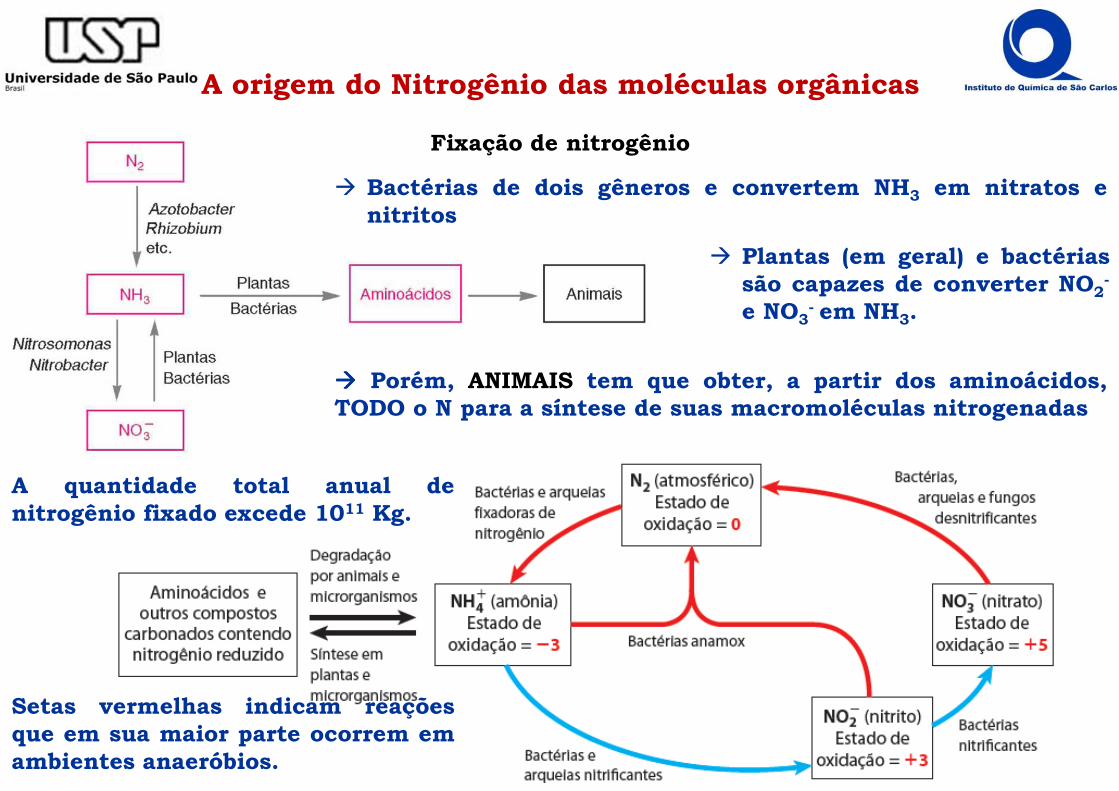
A origem do Nitrogênio das moléculas orgânicas
Fixação de nitrogênio
� Bactérias de dois gêneros e convertem NH3 em nitratos enitritos
���� Porém, ANIMAIS tem que obter, a partir dos aminoácidos,TODO o N para a síntese de suas macromoléculas nitrogenadas
� Plantas (em geral) e bactériassão capazes de converter NO2
-
e NO3- em NH3.
A quantidade total anual denitrogênio fixado excede 1011 Kg.
Setas vermelhas indicam reaçõesque em sua maior parte ocorrem emambientes anaeróbios.
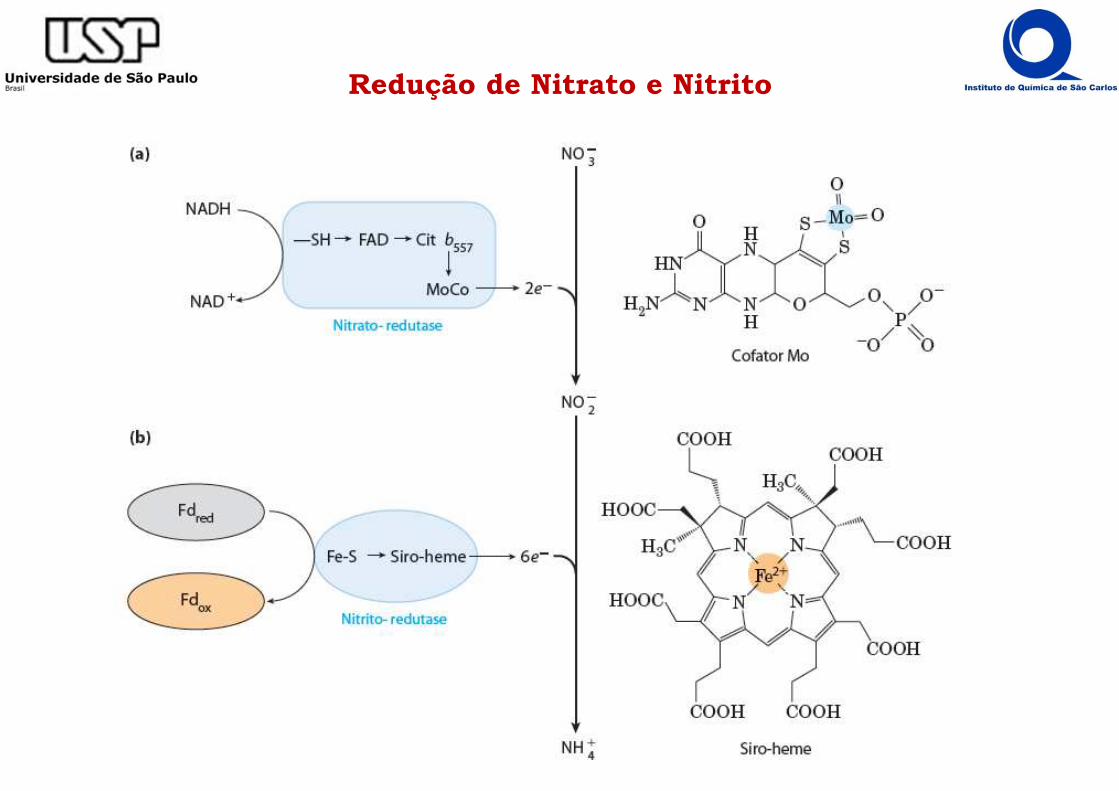
Redução de Nitrato e Nitrito
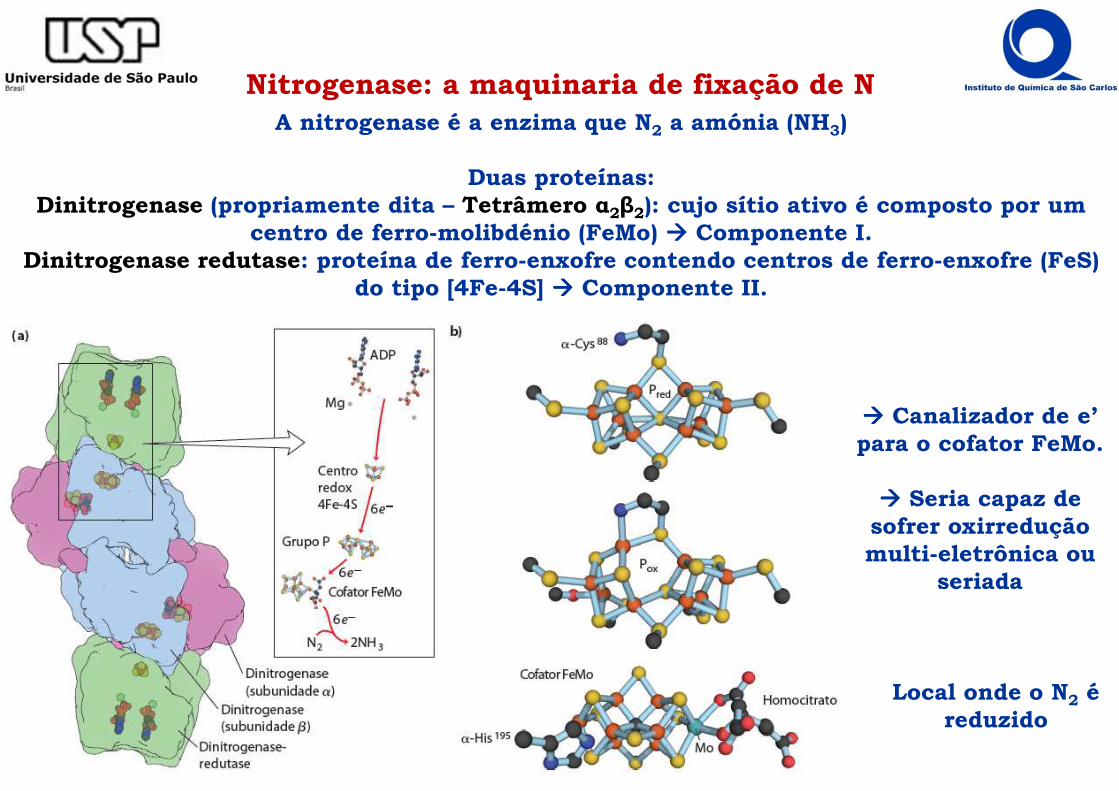
Nitrogenase: a maquinaria de fixação de NA nitrogenase é a enzima que N2 a amónia (NH3)
Duas proteínas: Dinitrogenase (propriamente dita – Tetrâmero α2β2): cujo sítio ativo é composto por um
centro de ferro-molibdénio (FeMo) ���� Componente I.Dinitrogenase redutase: proteína de ferro-enxofre contendo centros de ferro-enxofre (FeS)
do tipo [4Fe-4S] ���� Componente II.
Local onde o N2 é reduzido
���� Canalizador de e’ para o cofator FeMo.
���� Seria capaz de sofrer oxirredução multi-eletrônica ou
seriada
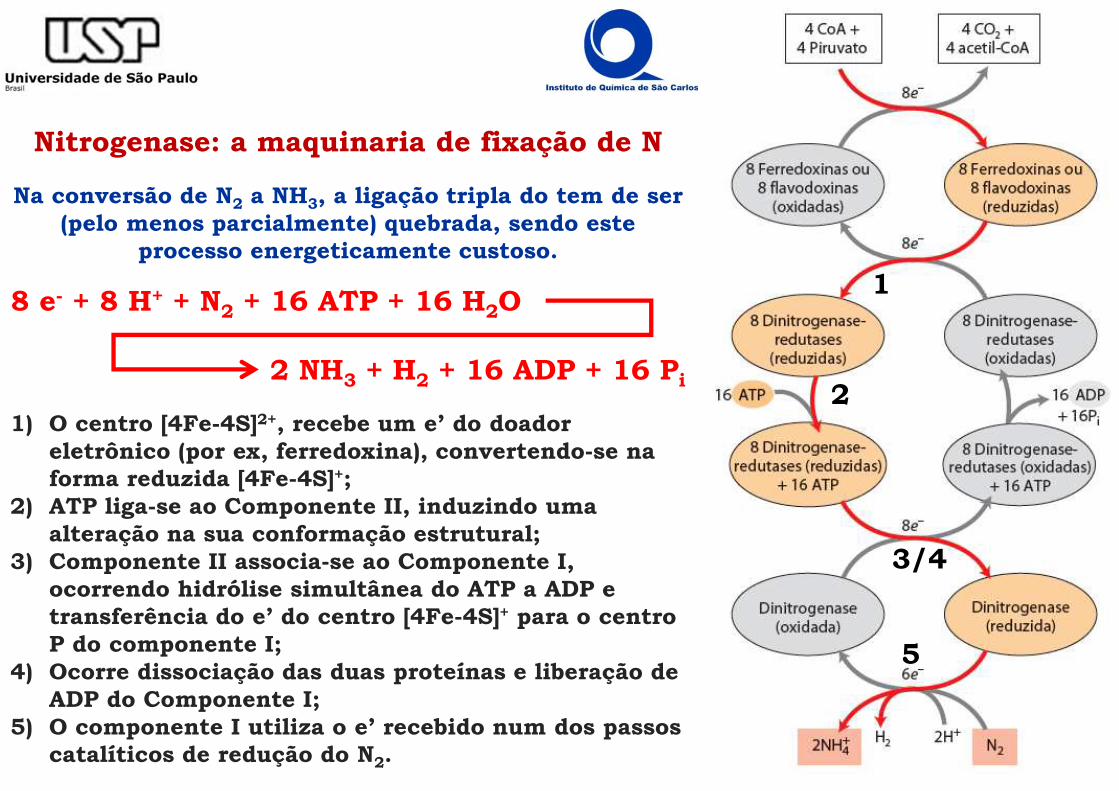
Nitrogenase: a maquinaria de fixação de N
Na conversão de N2 a NH3, a ligação tripla do tem de ser (pelo menos parcialmente) quebrada, sendo este
processo energeticamente custoso.
8 e- + 8 H+ + N2 + 16 ATP + 16 H2O
1) O centro [4Fe-4S]2+, recebe um e’ do doador eletrônico (por ex, ferredoxina), convertendo-se na forma reduzida [4Fe-4S]+;
2) ATP liga-se ao Componente II, induzindo uma alteração na sua conformação estrutural;
3) Componente II associa-se ao Componente I, ocorrendo hidrólise simultânea do ATP a ADP e transferência do e’ do centro [4Fe-4S]+ para o centro P do componente I;
4) Ocorre dissociação das duas proteínas e liberação de ADP do Componente I;
5) O componente I utiliza o e’ recebido num dos passos catalíticos de redução do N2.
1
2
3/4
5
2 NH3 + H2 + 16 ADP + 16 Pi
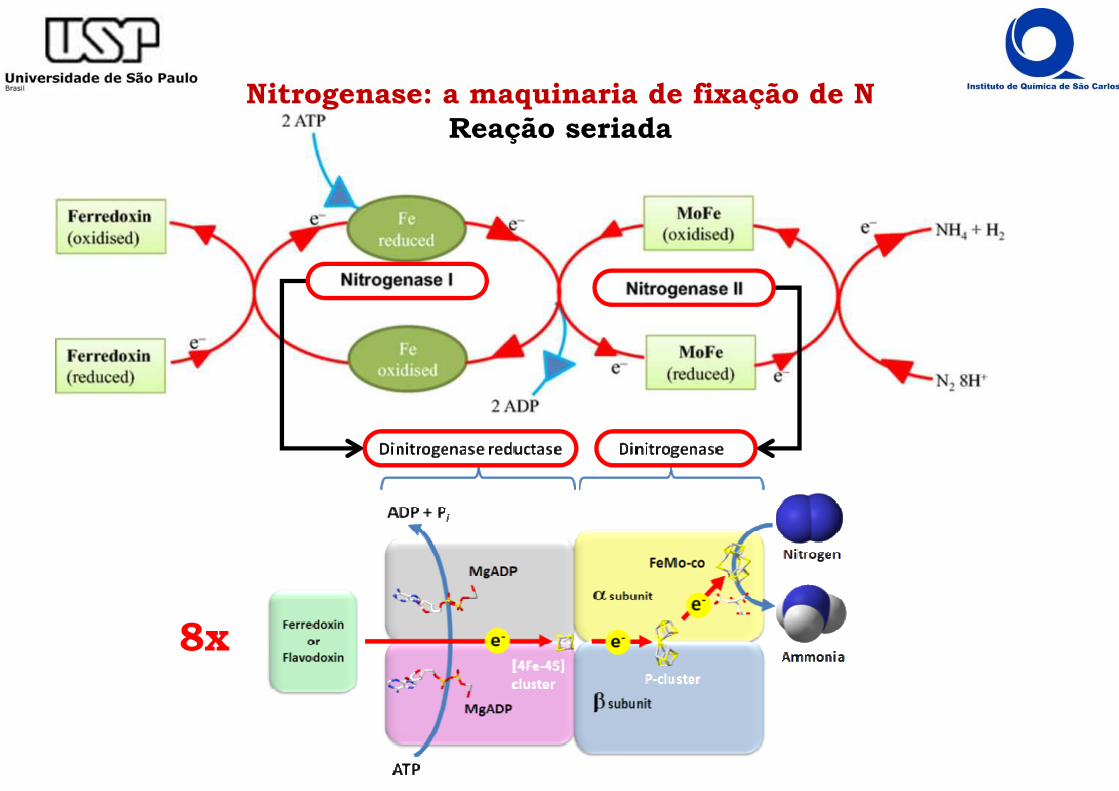
Nitrogenase: a maquinaria de fixação de NReação seriada
8x
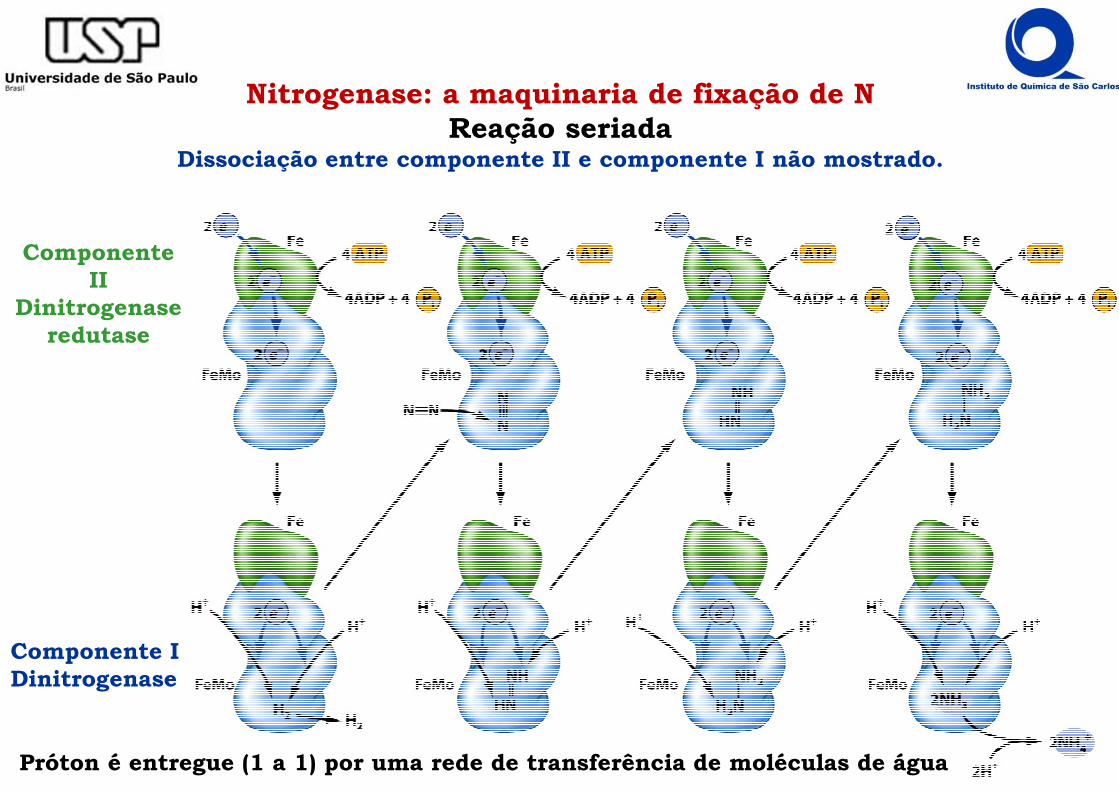
Nitrogenase: a maquinaria de fixação de NReação seriada
Dissociação entre componente II e componente I não mostrado.
Componente IDinitrogenase
Componente II
Dinitrogenaseredutase
Próton é entregue (1 a 1) por uma rede de transferência de moléculas de água

A amônia é incorporada em Glu e Gln
���� A amônia é, primeiramente, incorporada em Glu e Gln- os coletores de grupos amino, sendo, então, utilizados para a síntese dos demais
aminoácidos.
- Bactérias como E. coli sintetizam os 20 aminoácidos
- Humanos sintetizam apenas 11 dos 20 aminoácidos
- Os demais 9 aminoácidos são denominados AMINOÁCIDOS ESSENCIAIS
que devem ser obtidos da dieta.
Tyr e Cys, apesar de serem considerados não essenciais, são sintetizados a partir de aminoácidos essenciais obtidos da dieta.
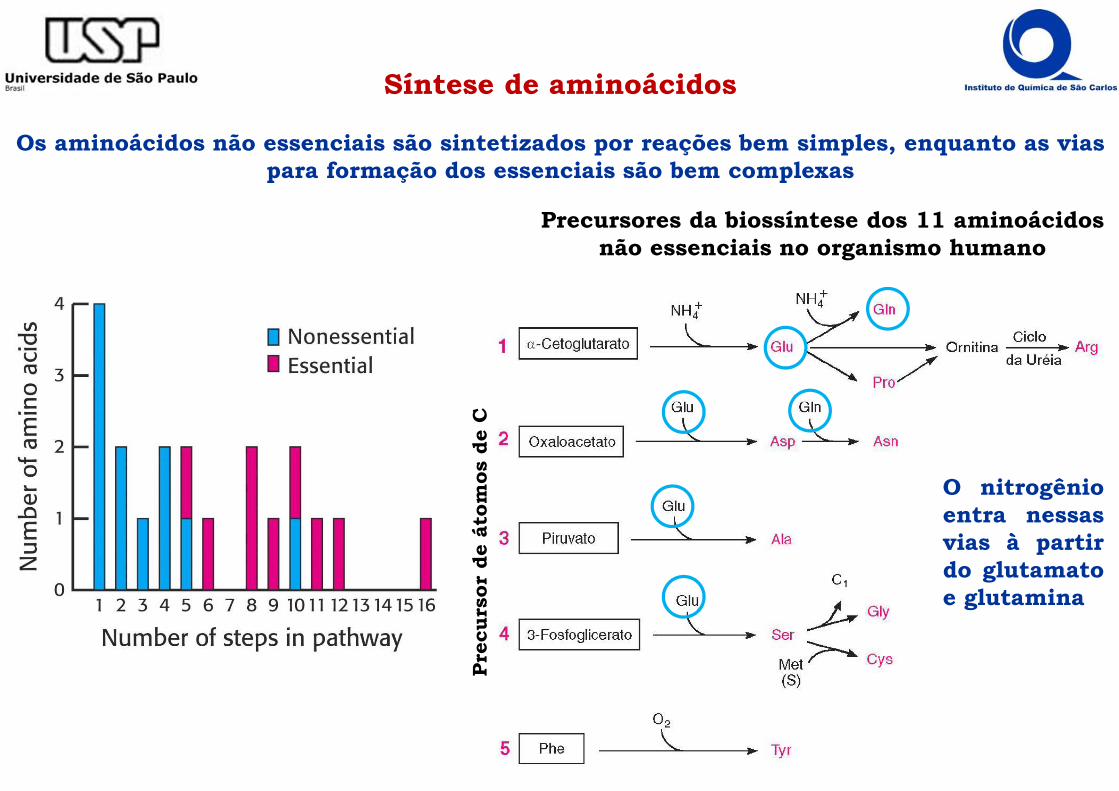
Síntese de aminoácidos
Os aminoácidos não essenciais são sintetizados por reações bem simples, enquanto as vias para formação dos essenciais são bem complexas
Precursores da biossíntese dos 11 aminoácidos não essenciais no organismo humano
O nitrogênioentra nessasvias à partirdo glutamatoe glutamina
Pre
curs
or
de á
tom
os
de C
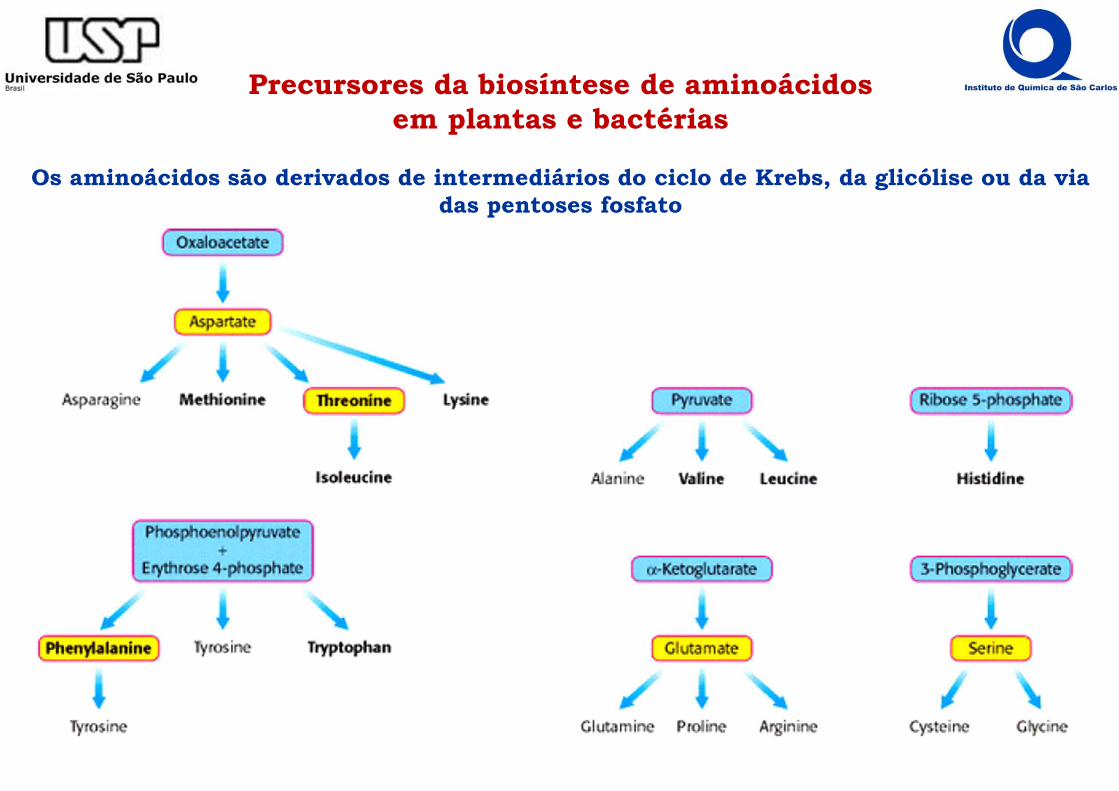
Precursores da biosíntese de aminoácidos em plantas e bactérias
Os aminoácidos são derivados de intermediários do ciclo de Krebs, da glicólise ou da via das pentoses fosfato


















