ANATOMIA DO LENHO E DENDROCRONOLOGIA DE LIANAS DA FAMÍLIA...
104
Instituto de Pesquisas Jardim Botânico do Rio de Janeiro Escola Nacional de Botânica Tropical ANATOMIA DO LENHO E DENDROCRONOLOGIA DE LIANAS DA FAMÍLIA LEGUMINOSAE OCORRENTES NA MATA ATLÂNTICA ARNO FRITZ DAS NEVES BRANDES 2007
-
Upload
phungtuyen -
Category
Documents
-
view
245 -
download
0
Transcript of ANATOMIA DO LENHO E DENDROCRONOLOGIA DE LIANAS DA FAMÍLIA...

Instituto de Pesquisas Jardim Botânico do Rio de Janeiro Escola Nacional de Botânica Tropical
ANATOMIA DO LENHO E DENDROCRONOLOGIA DE
LIANAS DA FAMÍLIA LEGUMINOSAE OCORRENTES NA
MATA ATLÂNTICA
ARNO FRITZ DAS NEVES BRANDES
2007

II
Instituto de Pesquisas Jardim Botânico do Rio de Janeiro Escola Nacional de Botânica Tropical
ANATOMIA DO LENHO E DENDROCRONOLOGIA DE
LIANAS DA FAMÍLIA LEGUMINOSAE OCORRENTES NA
MATA ATLÂNTICA
ARNO FRITZ DAS NEVES BRANDES
Dissertação de Mestrado submetida ao Programa de Pós-Graduação em Botânica, Escola Nacional de Botânica Tropical, do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, como parte dos requisitos necessários para a obtenção do grau de Mestre em Botânica.
Orientadora: DRA. CLÁUDIA FRANCA BARROS
Rio de Janeiro 2007

III
ANATOMIA DO LENHO E DENDROCRONOLOGIA DE LIANAS DA
FAMÍLIA LEGUMINOSAE OCORRENTES NA MATA ATLÂNTICA.
ARNO FRITZ DAS NEVES BRANDES
Orientadora:
CLÁUDIA FRANCA BARROS
Dissertação de Mestrado submetida ao Programa de Pós-Graduação em Botânica, Escola
Nacional de Botânica Tropical, do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro,
como parte dos requisitos necessários para a obtenção do grau de Mestre em Botânica.
Aprovada por:
__________________________________
Prof.
__________________________________
Prof.
__________________________________
Prof.
Em ___/___/2007.
Rio de Janeiro
2007

IV
Brandes, Arno Fritz das Neves B817a Anatomia do lenho e dendrocronologia de lianas da família Leguminosae
ocorrentes na Mata Atlântica / Arno Fritz das Neves – Rio de Janeiro, 2007. X, 94 f. : il. Dissertação (mestrado) – Instituto de Pesquisas Jardim Botânico do Rio
de Janeiro/Escola Nacional de Botânica Tropical, 2005. Orientadora: Claudia Franca Barros. Bibliografia. 1. Anatomia da madeira. 2. Dendrocronologia. 3. Liana. 4. Leguminosae
5. Mata Atlântica. I. Título. II. Escola Nacional de Botânica Tropical.
CDD 575.46

V
A meus pais Arno (in memorium)
e Irene, e a minha esposa Monica.

VI
AGRADECIMENTOS
Agradeço a todos que de alguma forma contribuíram para a realização deste estudo e
em especial:
À Prof. Dra. Cláudia Franca Barros pela excelente orientação, pela grande amizade e
confiança.
À CAPES pela concessão da bolsa de estudo.
À Petrobrás, Fundação Botânica Margaret Mee, Programa Mata Atlântica pelo auxílio
financeiro e logístico na execução deste estudo.
À Escola Nacional de Botânica Tropical pelo ensino de qualidade.
Ao Instituto de Pesquisas Jardim Botânico do Rio de Janeiro - Laboratório de Botânica
Estrutural e aos técnicos responsáveis Rogério, Elaine e Monique.
À direção e funcionários do Parque Nacional do Itatiaia pela licença de execução deste
trabalho e pelo auxílio logístico.
Ao Instituto Nacional de Meteorologia - 6° DISME pela concessão dos dados
meteorológicos.
À Neusa Tamaio pela troca de informações sobre lianas e parceria no ensino da
anatomia da madeira.
A Dra. Cecília Gonçalves Costa pelo excelente convívio, pelas sugestões no
desenvolvimento deste trabalho e por todos os ensinamentos.
À Dra. Catia Callado por permitir a realização do estagio em docência na UERJ e
pelas valorosas conversas e informações sobre dendrocronologia.
À Maura da Cunha pela sugestões no manuscrito sobre a anatomia das camadas de
crescimento em lianas.
À Marli Pires Morim e Haroldo Cavalcante Lima pelos importantíssimos
ensinamentos a respeito das Leguminosas, e junto com Michel Barros e Robson Ribeiro pelo
auxilio na identificação do material coletado.
À Monica C. da Silva, pelo inestimável amor, companheirismo, pelas discussões
científicas, pelos ensinamentos da língua inglesa e ajuda em inúmeras outras situações.
À minha Mãe Irene Aparecida das Neves, pelo imenso amor, apoio e incentivo. E a
minha Avó Maria pelo grande amor e cuidados.
Aos meus sogros Lucas Alves Cardoso da Silva e Marialva Cardoso da Silva por toda
consideração, amizade e ajuda durantes estes anos.

VII
Aos grandes amigos parceiros de campo Izar Aximoff, Jaqueline Pires e Clarice pelo
auxilio e companhia durantes as excursões ao Itatiaia.
Aos grandes amigos escaladores e surfistas do JBRJ, Eduardo Saddi (Dudu), Izar
Aximoff, Juan Alarcon, Bruno Resende e Felipe Sobrinho, pelas confraternizações nos
momentos de laser.

VIII
RESUMO
As lianas representam uma parte considerável da riqueza e abundância nas florestas tropicais
e desempenham papel fundamental em diversos processos ecológicos. Para entender melhor
seu papel é necessário conhecer os vários aspectos da sua biologia, como a anatomia, o
crescimento e a idade destes organismos. O presente estudo descreveu e analisou a anatomia
do lenho de oito espécies de lianas da família Leguminosae, ocorrentes no Parque Nacional do
Itatiaia, localizado na região Sudeste do Brasil, inserido no bioma Mata Atlântica. Detalhes da
anatomia e da periodicidade de formação das camadas de crescimento foram verificados. As
espécies apresentaram características esperadas em lianas, como: elementos de vaso com
diâmetro elevado, com duas categorias de diâmetro; maior proporção de parênquima axial em
comparação às fibras; e variações cambiais. A anatomia do lenho das espécies estudadas é
muito semelhante por compartilharem o mesmo hábito e pertencerem à mesma família.
Apesar disso, as oito espécies estudadas puderam ser diferenciadas. A presença de camadas de
crescimento distintas foi observada nas quatro espécies que não apresentaram variação
cambial. As características que delimitam as camadas de crescimento nestas espécies são: anel
semi-poroso, parênquima marginal, zona fibrosa e achatamento radial das células do lenho
tardio. Nestas espécies foi detectado que as camadas de crescimento eram anuais. A idade
máxima observada nas lianas foi 27 anos e taxa média de crescimento radial de 2,3 mm por
ano. Através da análise das séries temporais, notou-se que ocorre grande semelhança entre os
raios do mesmo indivíduo, contudo pequena semelhança entre os indivíduos e entre as
espécies. A análise climatológica das séries temporais mostrou que poucos indivíduos
apresentam correlação significativa com precipitação, temperatura e insolação. Por meio do
incremento radial acumulado nota-se uma forte relação idade-diâmetro nas lianas, contudo o
desvio de um incremento acumulado médio para a espécie aumenta junto com a idade.
Palavras-chave: anatomia da madeira, lianas, Leguminosae, camadas de crescimento,
dendrocronologia, crescimento secundário, idade.

IX
ABSTRACT
The lianas represent a considerable part of the richness and abundance in tropical forests and
they play basic role in many ecological processes. To better understand its role, it is necessary
to know some aspects of its biology, as the anatomy, the growth and the age of these
organisms. In the present study it is described and analysed the wood anatomy of eight lianas
species of the Leguminosae family, from the Parque Nacional do Itatiaia, located in the
South-eastern region of Brazil, inserted in Atlantic Rain Forest biome. Details of anatomy and
formation periodicity of the growth rings were verified. The species show characteristics
waited for lianas, as: wide vessel, with two categories of diameter; bigger parenchyma axial
ratio in comparison to fibres; and cambial variants. The wood anatomy of the studied species
is very similar for sharing the same habit and belonging to the same family. Despite the
similarities all studied species could be differentiated. The presence of distinct growth rings
was observed in the four species that did not presented cambial variants. The features that
delimit the layers of growth in these species are: semi-ring porosity, marginal parenchyma,
fibrous zone and radially flattened latewood cells. In these species was detected that the
growth layers were annual. The observed maximum age of these lianas was 27 years and the
radial growth rate averaged was 2,3 mm per year. Through the analysis of the time series, it
was observed that great similarities occur between rays of the same individual, however small
similarity between individuals and species. The climatological analysis of the time series
showed that few individuals presented significant correlation with precipitation, temperature
and incident sunlight. By means of the accumulated radial increment, a strong relation age-
diameter was observed in lianas, however the deviation of an average accumulated increment
for the species increases together with the age.
Key-words: wood anatomy, lianas, Leguminosae, growth rings, dendrochronology,
secondary growth, age.

X
ÍNDICE
INTRODUÇÃO GERAL ........................................................................................................1
OBJETIVOS GERAIS ...........................................................................................................4
OBJETIVOS ESPECÍFICOS ..................................................................................................4
REVISÃO BIBLIOGRÁFICA ...............................................................................................5
ARTIGO 1 - ANATOMIA DO LENHO DE OITO ESPÉCIES DE LIANAS DA FAMÍLIA LEGUMINOSAE OCORRENTES NA FLORESTA ATLÂNTICA ..................................... 11
RESUMO ................................................................................................................. 12 ABSTRACT ............................................................................................................. 13 INTRODUÇÃO ........................................................................................................ 14 MATERIAL E MÉTODOS ....................................................................................... 15 RESULTADOS ........................................................................................................ 17 DISCUSSÃO ............................................................................................................ 28 AGRADECIMENTOS .............................................................................................. 33 REFERÊNCIAS BIBLIOGRÁFICAS ....................................................................... 33
ARTIGO 2 - GROWTH RINGS IN LIANAS OF LEGUMINOSAE FAMILY FROM ATLANTIC FOREST IN RIO DE JANEIRO, BRAZIL ....................................................... 42
ABSTRACT ............................................................................................................. 43 INTRODUCTION .................................................................................................... 44 MATERIALS AND METHODS ............................................................................... 46 RESULTS ................................................................................................................. 48 DISCUSSION ........................................................................................................... 51 ACKNOWLEDGEMENTS ...................................................................................... 55 REFERENCES ......................................................................................................... 55
ARTIGO 3 - CRESCIMENTO SECUNDÁRIO E IDADE EM LIANAS DA FAMÍLIA LEGUMINOSAE NA MATA ATLÂNTICA: UM ESTUDO COM CAMADAS DE CRESCIMENTO .................................................................................................................. 68
RESUMO ................................................................................................................. 69 INTRODUÇÃO ........................................................................................................ 70 MATERIAIS E MÉTODOS ...................................................................................... 72 RESULTADOS ........................................................................................................ 74 DISCUSSÃO ............................................................................................................ 76 REFERÊNCIAS BIBLIOGRÁFICAS ....................................................................... 79
CONSIDERAÇÕES GERAIS .............................................................................................. 87
REFERÊNCIAS BIBLIOGRÁFICAS GERAIS ................................................................... 89

INTRODUÇÃO GERAL
Nas florestas do mundo e particularmente nas regiões tropicais as lianas formam um
abundante e diverso grupo de plantas com características muito peculiares. Apesar de serem
muito importantes para os ecossistemas, a biologia das lianas tem sido relativamente pouco
estudada, em comparação com vegetais de outros hábitos. As lianas são um agrupamento
polifilético de vegetais que compartilham a mesma estratégia de desenvolvimento. O hábito
lianescente evoluiu independentemente em diferentes famílias de vegetais com flores e sem
flores (Ewers & Fisher, 1991; Gentry, 1991; Schnitzer & Bongers, 2002).
Os vegetais escaladores (trepadeiras) são organismos que dependem de outros vegetais
ou suportes para sustentação (Putz, 2006). Gentry (1985, 1991) classifica estes organismos
quanto à morfologia e ecologia, caracterizando as lianas por apresentarem caules lenhosos,
iniciarem sua vida no solo e terem a capacidade de desenvolver-se em florestas maduras,
diferindo dos outros tipos de trepadeiras que são herbáceas ou que em algum período de suas
vidas apresentam-se como epífitas.
Lianas são importantes componentes estruturais das florestas tropicais e representam
aproximadamente 25% da riqueza e da abundância nestes ambientes. Em certas regiões,
chegou-se a estimar que 44% das espécies lenhosas são lianas, como nas margens da bacia
amazônica. Sua abundância é controlada por vários fatores abióticos, incluindo precipitação
total, periodicidade das chuvas, fertilidade do solo e distúrbios. Outros estudos mostram a
importância das lianas com relação à biomassa, à produção de serrapilheira de folhas e à
interação com os animais. Podem contribuir com 5% da biomassa total acima do solo e suas
folhas constituem até 40% da área foliar da floresta. Para os animais, servem como alimento
e como componente estrutural do seu habitat, proporcionando conexões entre as copas
(Gentry, 1991; Putz, 2006; Schnitzer & Bongers, 2002). Nos processos em nível de
ecossistemas, as lianas têm fundamental importância na transpiração total da floresta e no
seqüestro de carbono (Schnitzer & Bongers, 2002).
A família Leguminosae, freqüentemente, é considerada como a maior família em
numero de espécies em levantamentos florísticos em florestas neotropicais. É também a
segunda mais importante família de lianas, sendo Bignoniaceae a mais importante em número
de espécies. No entanto, em algumas localidades no Peru e no Equador, a família
Leguminosae é observada como a de maior riqueza de espécies (Gentry, 1982,1991). No

2
Bioma Mata Atlântica, dois levantamentos mostram Leguminosae como uma das famílias
com maior riqueza de lianas (Rezende e Ranga, 2005). O hábito lianescente é observado nas 3
subfamílias, como observado nos gêneros: Bauhinia da subfamília Caesalpinoideae, Acacia
da subfamília Mimosoideae e Dalbergia e Machaerium da subfamília Papilionoideae
(Obaton, 1960; Caballé, 1993).
As lianas são referidas como importante componente da fisionomia dos remanescentes
de Floresta Atlântica do Parque Nacional do Itatiaia, desde o primeiro levantamento florístico
realizado na região por Brade, em 1956. Entretanto, poucos estudos posteriores levam em
consideração o hábito lianescente, como parâmetro de avaliação florística da região ou sua
importância na estrutura da floresta e nenhum estudo sobre a anatomia das lianas foi
realizado. Estudos realizados pelo Programa Mata Atlântica com as leguminosas arbóreas e
arbustivas, alertam para a necessidade de um aprofundamento no conhecimento das lianas da
família Leguminosae nessa região (Morim, 2002).
Na maioria dos vegetais que apresentam crescimento secundário, o câmbio vascular
produz xilema secundário internamente e floema secundário externamente, o que poderia ser
definido como tipo “comum” (Mauseth, 1988). As lianas apresentam vários tipos de
desenvolvimento ontogenético relacionados aos processos de desenvolvimento secundário,
diferindo do tipo “comum” (Carlquist, 1975). O desenvolvimento ontogenético variado deve-
se à atividade desigual do cambio vascular ou ao surgimento de outros câmbios vasculares e
foi chamado de “desenvolvimento secundário anômalo” (Dobbins, 1981) (Mauseth, 1988),
“desenvolvimento secundário incomum” (Fahn,1990), “variações no crescimento
secundário” (Fisher & Ewers, 1992), “câmbio anômalo” (Philipson, 1980) e “variações
cambiais” (Carlquist, 1988, 1991) (Rajput & Rao, 2003). Esta ultima denominação citada será
a utilizada neste trabalho.
Na história evolutiva de muitos vegetais, diferentes métodos de maximizar a condução
e a segurança no transporte da seiva foram desenvolvidos. Podem-se notar algumas tendências
ecológicas na anatomia do lenho que estariam relacionadas à resistência ao estabelecimento
de pressão negativa no xilema e também aos diferentes tipos ecológicos e diferentes hábitos
(árvores, arbustos e lianas). Aparentemente as lianas teriam sua última derivação de um
padrão de espécies lenhosas mésicas devido à área condutora, contudo tendem a formar um
grupo distinto destas espécies mésicas. As lianas tendem, excepcionalmente, a apresentar
vasos de grande diâmetro quando comparadas a outros grupos lenhosos, o que minimiza o
atrito no movimento da água, facilitando a condução (Carlquist, 1975).

3
Apesar das vantagens dos vasos grandes, Ewers (1985) questionou: “Por que todas as
plantas terrestres não apresentam vasos largos?” Vasos com diâmetro elevado são possíveis
apenas onde altas tensões negativas na coluna de água não existam, do contrário ocorreria a
cavitação dos vasos e o impedimento da condução. Assim, as lianas desenvolveram algumas
estratégias adaptativas para aumentar a segurança na condução. Entre elas está a presença de
dimorfismo dos vasos, ou seja, vasos com duas categorias de tamanho. Os vasos estreitos
continuam a conduzir mesmo quando ocorre embolismo nos vaso largos. Este autor também
observou que a pressão positiva gerada pelas raízes das lianas pode restaurar a funcionalidade
dos vasos cavitados.
O comprimento dos elementos de vasos das lianas não difere muito das plantas auto-
suportantes (Carlquist, 1991). Contudo o comprimento dos vasos, que é a série de elementos
de vasos interconectados verticalmente pelas placas de perfuração, é relativamente grande nas
lianas. Foi notado em análises que o diâmetro e comprimento dos vasos aparentam estar
correlacionados (Ewers & Fisher, 1987, 1989; Ewers et al., 1991).
A presença de tecido de sustentação é reduzida em favor do tecido condutivo e do
parênquima. As lianas apresentam abundância de parênquima o que confere maior
flexibilidade, auxilia na recuperação de injúrias, e no caso do parênquima axial, também pode
aumentar a segurança do sistema condutor (Carlquist , 1985).
Apesar de pouco relatadas, as lianas também podem apresentar camadas de
crescimento distintas (Baas & Schweingruber, 1987; Gasson e Dobbins, 1991; Carquist, 1995;
Araque, 2000). A aplicação dos métodos de dendrocronologia em lianas não é muito
conhecida (Ceccantini & Brandes, dados não publicados) (Kanngiesser, 1906 apud
Schweingruber & Poschlod, 2005), contudo é uma importante área a ser investigada que pode
prover importantes informações para o melhor entendimento deste hábito.
A Dendrocronologia pode ser definida como o estudo da seqüência cronológica de
formação de anéis de crescimento em espécies lenhosas (Stokes & Smiley, 1996). Até
recentemente regiões tropicais têm sido deixadas de lado pelos dendrocronologistas, sob a
alegação que os anéis de crescimento não são distintos ou quando distintos freqüentemente
não são anuais (Jacoby, 1989). Contudo, atualmente é bem aceito o conceito de que muitas
das espécies tropicais podem apresentar anéis de crescimento, não relacionados somente a
fatores climáticos como precipitação, mas também a vários outros fatores ambientais
sazonais, genéticos e endógenos, como por exemplo espécies que vivem nas margens dos
igarapés na Amazônia, algumas espécies de regiões inundáveis da Floresta Atlântica e
espécies decíduas (Callado et al., 2001a, 2001b; Worbes, 1988,1989).

4
OBJETIVOS GERAIS
� Conhecer a anatomia do lenho e a dendrocronologia de espécies de lianas da família
Leguminosae do Parque Nacional do Itatiaia.
OBJETIVOS ESPECÍFICOS
� Caracterizar anatomicamente o lenho das lianas da família Leguminosae do Parque
Nacional do Itatiaia, de forma quantitativa e qualitativa;
� Analisar a presença de camadas de crescimento nessas espécies e caracterizá-las
anatomicamente;
� Identificar a periodicidade de formação dos anéis de crescimento das espécies em
estudo que apresentarem esta característica anatômica;
� Caracterizar o crescimento radial dos caules das lianas estudadas e estimar a idade
através da análise dos anéis de crescimento.

5
REVISÃO BIBLIOGRÁFICA
Lianas
Historicamente a presença e a importância das lianas têm fascinado biólogos e
naturalistas. As primeiras publicações, que descrevem o hábito lianescente são do século XIX.
Mirbel (1828 apud Obaton, 1960) relatou a presença de feixes vasculares auxiliares surgindo
a partir do sistema vascular principal em Calycanthus; Gaudichaud (1833 apud Obaton, 1960)
estudou lianas brasileiras de cinco famílias; e Decaisne (1839 apud Obaton, 1960) descreveu
a aparição de zonas sucessivas de crescimento em Menispermaceae. Merecem destaque na
segunda metade do século XIX: o trabalho de Darwin (1867) intitulado “On the movement
and habits of climbing plants”, em que o autor investigou a estrutura e o comportamento dos
vegetais escaladores; Westermeier & Ambronn (1881 apud Carlquist, 1991) que descreveram
aspectos anatômicos do caule em que as plantas escaladoras diferem das auto-suportantes; e
os estudos clássicos de Schenck (1892, 1893), que focou suas pesquisas principalmente na
anatomia do caule das lianas, mas também incluiu investigações taxonômicas e geográficas e
realizou um excelente registro gráfico.
No inicio do século XX foram desenvolvidos vários trabalhos com lianas, a exemplo
de: Solereder (1908) que trata da anatomia comparada de dicotiledôneas e inclui as lianas em
suas análises; Shirley & Lambert (1918), que investigaram a anatomia de 53 espécies de
plantas escaladoras de 23 famílias com a intenção de determinar a família pelo exame da
estrutura interna; Dastur & Kapadia (1931) que estudaram a anatomia de vegetais escaladores
de 35 espécies. Estes trabalhos, citados neste parágrafo, enfatizaram as variações cambiais
encontradas.
Na segunda metade do século XX, ocorreu um aumento na investigação a respeito das
lianas e mais pesquisadores passaram a dar atenção ao hábito (Putz, 2006). Mas somente há
poucos anos, com o reconhecimento do importante papel das lianas nas muitas áreas da
ecologia vegetal, houve uma explosão dos relatos de pesquisas com lianas (Schnitzer &
Bongers, 2002). Mesmo assim, as lianas são menos estudadas quando comparadas a outros
hábitos, tendo sido muito negligenciadas por coletores e pesquisadores, possivelmente devido
às dificuldades de obtenção das amostras (Gentry, 1991). Bamber & Ter Welle (1994)
referem que os estudos anatômicos das lianas, assim como os da casca, foram negligenciados
devido a interesses econômicos. Os estudos concentravam-se no estudo anatômico do lenho
de árvores, que proveriam dados para identificação e utilização da madeira.

6
Na ultima década do século XX, um importante livro editado por Putz & Money
(1991) e intitulado “The Biology of Vines” reuniu informações a respeito das plantas
escaladoras. Esta obra trata em cinco grandes seções, da distribuição e evolução das plantas
escaladoras, da forma do caule e dos mecanismos usados para a ascensão, da fisiologia e
desenvolvimento, da ecologia de comunidades e da importância econômica das lianas.
Merecem destaque: “The distribuition and evolution of climbing plants” (Gentry, 1991), em
que o autor faz uma introdução a respeito das plantas escaladoras, define os tipos quanto à
morfologia e à ecologia, trata da importância e da distribuição geográfica, aborda aspectos
taxonômicos, evolutivos e de ecologia de comunidades; “Anatomy of vine and liana stems: a
review and syntesis” (Carlquist, 1991), que trata das variações cambiais, característica muito
comum nas plantas escaladoras, propõe uma classificação para essas variações e discorre
sobre os tecidos que compõem o xilema secundário relacionando-os com o hábito; “Structural
responses to stem injury in vines” (Fisher & Ewers, 1991), também trata dos aspectos
anatômicos, porém enfatiza o significado funcional, muitas vezes relacionado à recuperação
das injurias e à condução; “Water flux and xylem structure in vines” (Ewers et al., 1991), trata
de aspectos da condução, como a dimensão dos vasos; “Vine-host interactions” (Hegarty,
1991), traz uma importante classificação dos vegetais escaladores com relação aos métodos de
ascensão usados, discute cada um deles e aborda outros aspectos das interações com os
hospedeiros.
Os tecidos xilemáticos das lianas, comparados aos vegetais com outros hábitos,
mostram as seguintes tendências: maior diâmetro e/ou freqüência de vasos, ou seja, maior
área condutora (Araújo, 2005; Bamber & Ter Welle, 1994; Bass & Schweingruber, 1987;
Carlquist, 1985, 1991; Ewers, 1985; Ewers & Fisher, 1987,1989, 1991; Ewers et al., 1990;
Ewers et al. ,1991; Ewers et al.,1997; Fisher & Ewers, 1995; Gartner et al., 1990; Gasson &
Dobbins, 1991; Iqbal, 1994; Ter Welle, 1985; Zimmermann, 1983); longa série de elementos
de vasos ligados pelas placas de perfuração (Ewers, 1985; Ewers & Fisher, 1987; Ewers et al.,
1990; Ewers et al., 1991; Fisher & Ewers, 1995; Zimmermann, 1983); dimorfismo de vasos
(Araújo, 2005; Bass & Schweingruber, 1987; Carlquist, 1985, 1991; Gasson & Dobbins,
1991), maior abundância de parênquima axial (Carlquist, 1991, 1995; Dias-Leme, 1999;
Fisher & Ewers, 1989), maior largura e altura dos raios (Bamber & Ter Welle, 1994;
Carlquist, 1991, 1995, 1996; Fisher & Ewers, 1989; Gasson & Dobbins, 1991), placas de
perfuração simples (Araújo, 2005; Carlquist, 1991).

7
A notável largura dos elementos de vaso e dos raios nas lianas foi observada
primeiramente por Westermeier & Ambronn (1881 apud Carlquist, 1991), que relatam vasos
em Passiflora com mais de 500µm de diâmetro.
Dois dos pesquisadores que mais contribuíram para o conhecimento das tendências
relacionadas à condução de água e conseqüentemente à dimensões dos vasos, foram Ewers e
Fisher. Algumas das principais contribuições a este conhecimento são: a corroboração que as
lianas apresentam os mais largos e longos vasos do reino vegetal, que os vasos de lianas
podem permanecer funcionais por mais tempo que pode ser esperado (em comparação com as
árvores), que existe uma relação direta entre largura e comprimento dos vasos, e que não
ocorre somente pouco xilema por área foliar mas também pouco floema e tecido cortical
(Ewers, 1985; Ewers & Fisher, 1987, 1989, 1991; Ewers et al., 1989; Ewers et al., 1990;
Ewers et al., 1991; Ewers et al.,1997; Fisher & Ewers, 1995).
No Parque Nacional do Itatiaia são poucos os estudos que envolvem lianas. Brade
(1956) relata este hábito no Parque, apresentando-se abundante em regiões baixas e sofrendo
uma redução de acordo com as mudanças na altitude e vegetação. Acima dos 1900 m é
relatada a ocorrência de poucas espécies quando comparada a regiões baixas. Ele cita também
que lianas lenhosas são características do elemento florístico subtropical das matas higrófilas.
Outros estudos florísticos e fitossociológicos na Floresta Atlântica, como os realizados na
Reserva ecológica de Macaé de Cima, em Nova Friburgo, demonstram a importância das
trepadeiras como componente florístico, representando 18% das espécies (Lima & Guedes-
Bruni, 1994). Dois levantamentos florísticos desenvolvidos na região sudeste do Brasil,
mostraram Leguminosae como uma das famílias mais representativa em número de espécies
de lianas nas áreas estudadas (Resende & Ranga, 2005).
Os estudos da anatomia de lianas da Floreta Atlântica são escassos. Porém duas
pesquisas recentes podem ser citadas, uma com a espécie Serjania corrugata (Araújo, 2005) e
a outra com a família Menispermaceae (Tamaio, 2006). Estes estudos abordam vários
aspectos anatômicos do lenho incluindo o desenvolvimento e implantação das variações
cambiais.
A família Leguminosae
Na maioria das leguminosas algumas características anatômicas comuns ao lenho são:
porosidade difusa, vasos curtos, com placas de perfuração simples, pontoações intervasculares
alternas guarnecidas, pontoações raiovasculares semelhantes às intervasculares e cristais em
células subdivididas de parênquima axial. As variações na família são principalmente quanto:

8
ao tipo de parênquima axial e à presença de cristais em células subdivididas; ao diâmetro e à
freqüência dos vasos; às fibras que podem apresentar-se septadas ou não com paredes de
espessura variável; aos raios variando na largura, na altura e na composição celular; a
presença de estratificação (Barreta-Kuipers, 1981; Dias-Leme, 1999; Gasson, 1994). Alguns
dos autores tratam da anatomia de leguminosas e as discutiram num contexto taxonômico são:
Barreta-Kuipers (1981), Gasson (1994, 2000, 2003) e Höhn (1999). Merece atenção uma
recente e extensa pesquisa realizada por Evans et al. (2006), que trata detalhadamente da
anatomia da subfamília Mimosoideae, das tribos e dos grupos subordinados.
A família Leguminosae está presente em diversos estudos sobre anatomia de lianas,
possivelmente devido à representação da família em relação ao hábito, à riqueza de espécies e
à abundância. Os primeiros estudos relatados em lianas da família leguminosae datam do
século XIX. Sendo o primeiro a descrição de uma Acacia que apresentava 4 lobos (Netto,
1863 apud Obaton, 1960) e o segundo relato, a descrição da formação de arcos libero-
lenhosos (xilema e floema) terciários em Wistaria sinensis (Van Tieghem, 1891 apud Obaton,
1960). Dastur & Kapadia (1931) investigaram a anatomia de 7 espécies de leguminosas junto
com espécies de outras famílias. A análise da anatomia do lenho de Bauhinia radiata foi
realizada por Vieira (1990), que descreveu a variação cambial e constatou diferenças na
manifestação das estruturas anômalas em relação à luminosidade. Caballé (1993) investigou
muitas espécies de lianas tendo ilustrado em seu trabalho 11 espécies de leguminosas das 3
sub-famílias. As quais pertenciam aos gêneros Bauhinia, Acacia, Dalbergia e Machaerium. A
incidência de desenvolvimento anômalo foi estudada por Bamber & Ter Welle (1994) em 14
espécies de leguminosas, dentre um total de 117 espécies.
Vários trabalhos citam variações cambiais em gêneros da família Leguminosae. O
gênero mais citado, que se destaca por suas variações cambiais, é Bauhinia (Carlquist, 1988,
1991; Esau, 1985; Mauseth, 1988; Philipson, 1980; Vieira, 1990). O gênero Bauhinia
(Leguminosae) foi bem estudado pelos autores Fisher, Ewers e colaboradores pela sua
diversidade de hábitos. Outros gêneros da família leguminosae estudados por eles foram
Machaerium, Clitoria, Platypodium e Spatholobus (Ewers, 1985; Ewers & Fisher,
1987,1989,1991; Ewers et al., 1989; Ewers et al., 1990; Ewers et al., 1991; Ewers et al.,1997;
Fisher & Ewers, 1989, 1992).

9
Dendrocronologia
Pesquisas sobre anéis de crescimento em árvores tropicais, vêm sendo desenvolvidas e
publicadas desde o início do século XX, e um breve histórico e visão geral a respeito do tema
é apresentado por Worbes (2002).
Existem muitos trabalhos a respeito do tema, contudo, quatro eventos importantes,
com suas respectivas publicações, podem ser destacados com propulsores e compiladores de
pesquisas com camadas de crescimento em regiões tropicais: Iawa Committee (1989), Iawa
Committee (1995), Iawa Committee (1999) e Yale University: School of Forestry and
Environmental Studies (1981). Essas publicações tratam dos anéis e da periodicidade de
crescimento, discutem os métodos utilizados e destacam os problemas para o
desenvolvimento de pesquisas dessa natureza. Além disso, reforçam a necessidade de mais
estudos e relatam aplicações dos resultados. Muitos destes trabalhos demonstram resultados
positivos para contagem de anéis de crescimento e datação em espécies tropicais. Há poucos
anos, uma outra importante publicação, direciona o enfoque para a dendrocronologia na
América Latina, mostra o estado da arte nesta região e traz inúmeros exemplos bem sucedidos
(Roig, 2000).
Recentemente, Callado et al. (2001a, 2001b), estudando espécies arbóreas da Floresta
Atlântica detectaram a presença de anéis de crescimento distintos em 14 das 17 espécies
estudadas. Em algumas espécies, os autores relacionam a formação das zonas de crescimento
distintas à perda das folhas, ao fotoperíodo, a regimes de inundação e aos ritmos endógenos.
Além disso notam que a periodicidade de formação dos anéis de crescimento é anual.
Um importante livro a respeito dos anéis de crescimento que merece atenção, intitula-
se “An introduction to tree-ring dating” e foi escrito por Stokes & Smiley (1996). Eles
discutem os princípios da datação por anéis de crescimento, explicam métodos de campo e
técnicas de laboratório.
Os anéis de crescimento podem prover informações sobre condições ambientais no
presente ou no passado (Gray & Thompson, 1977; Sass-Klaassen et al., 2005; Savva et al.,
2003; Stokes & Smiley, 1996; Trouet et al., 2001). E também podem fornecer informações a
respeito do comportamento temporal, quanto ao crescimento radial (Boninsegna et al.,1989)
(Ferreira, 2002).
Poucos estudos relatam e tratam das camadas de crescimento em lianas. Ceccantini &
Brandes (dados não publicados) utilizaram métodos dendrocronológicos para comparar o
crescimento na espécie Dalbergia frutescens, que pode apresentar-se como liana ou como
auto-suportante. Além deste estudo, é conhecida apenas a citação de uma pesquisa muito

10
antiga que estimou a idade máxima de uma espécie de liana (Kanngiesser, 1906 apud
Schweingruber & Poschlod, 2005). Existem outros relatos da ocorrência de anéis de
crescimento neste hábito feitos por Baas & Schweingruber (1987), Gasson & Dobbins (1991),
Carlquist (1995) e Araque et al. (2000), mas os autores restringem-se relatar a distinção, não
entrando em detalhes. Recentemente Leon-Gómez & Monroy-Ata (2005) trataram da
periodicidade da atividade cambial em quatro espécies de lianas e foi revelado que em
algumas espécies o câmbio apresenta sazonalidade, em certos períodos apresenta grande
atividade e em outros não.

11
ARTIGO 1
Anatomia do lenho de oito espécies de lianas da família
Leguminosae ocorrentes na Floresta Atlântica
Arno F. N. Brandes1,2 e Cláudia F. Barros1
1) Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, Programa Mata Atlântica, Rua
Pacheco Leão 915, Jardim Botânico, CEP 22460-030 Rio de Janeiro, Rio de Janeiro, Brasil
2) Autor para correspondência: [email protected]
Artigo aceito para publicação na Acta Botanica Brasilica, provavelmente volume 21/2007.

12
RESUMO
(Anatomia do lenho de oito espécies de lianas da família Leguminosae ocorrentes na Floresta
Atlântica). Lianas são importantes componentes estruturais das florestas tropicais, apesar
disso a anatomia destes vegetais é pouco estudada. A família Leguminosae é relatada como a
segunda maior em número de espécies de lianas, contudo pouco se conhece da diversidade
anatômica das lianas desta família. O presente estudo descreveu e analisou a anatomia do
lenho de oito espécies de lianas da família Leguminosae, ocorrentes no Parque Nacional do
Itatiaia, localizado no Estado do Rio de Janeiro, região Sudeste do Brasil, local inserido no
bioma Floresta Atlântica. As espécies apresentaram características comuns ao hábito liana,
como: elementos de vaso com diâmetro elevado, com duas categorias de diâmetro; e maior
proporção de parênquima axial em comparação às fibras. Quatro espécies do gênero
Senegalia apresentaram variações cambiais produzidas por um único câmbio, normal em
produto mas anormal em conformação. A anatomia do lenho das espécies estudadas é muito
semelhante por compartilharem o mesmo hábito e pertencerem à mesma família. Apesar
disso, as oito espécies estudadas puderam ser diferenciadas. As espécies com variações
cambiais e Dalbergia frutescens são facilmente diferenciadas das demais. Contudo Senegalia
tenuifolia, Piptadenia micracantha e Piptadenia adiantoides necessitam ser examinadas com
muita cautela para identificá-las exclusivamente pela anatomia do lenho.
Palavras-chave: anatomia da madeira, lianas, Mata Atlântica, Leguminosae

13
ABSTRACT
(Wood anatomy of eight liana species of Leguminosae family from Atlantic Rain Forest).
Lianas are important structural component of tropical forests and even though the anatomy of
these plants is poorly studied. Leguminosae family is reported as the second larger family in
number of liana species, but little is know about the anatomical diversity of lianas from this
family. The present study described and analyzed the wood anatomy of eighth liana species of
Leguminosae family, from Parque Nacional do Itatiaia, located in Rio de Janeiro State,
Southeast region of Brazil and included in Atlantic Rain Forest Biome. The species show
common features with the liana habit, as: wide vessels, vessel dimorphism, and high
proportion of parenchyma when compared with fibers. Four species of Senegalia genus
showed cambial variants made by a single cambium, normal in products but abnormal in
conformation. The studied species have similar wood anatomy, many of which are because
they belong to the same family and share the same habit. Nevertheless, the eight species
studied could be distinguished. The species with cambial variants and Dalbergia frutescens
are easy distinguished from the others. However Senegalia tenuifolia, Piptadenia
micracantha and Piptadenia adiantoides require much wariness examination to identify the
species exclusively by wood anatomical features.
Key words: Wood anatomy, lianas, Atlantic Rain Forest, Leguminosae

14
INTRODUÇÃO
Lianas são trepadeiras lenhosas que iniciam seu desenvolvimento no solo e com o
crescimento perdem sua habilidade de se sustentar, necessitando de suporte para ascender à
copa (Gentry 1991; Gerwing et al. 2006). Nas florestas tropicais as lianas têm grande
representatividade na riqueza e abundância de espécies (Putz & Money 1991; Schnitzer &
Bongers 2002). Apesar de serem muito importantes nos ecossistemas tropicais sua anatomia e
biologia são pouco estudadas (Carlquist 1985; Ewers & Fisher 1991; Bamber & Ter Welle
1994).
A anatomia do caule das lianas é muito peculiar, geralmente exibe características
relacionadas ao hábito trepador como a presença de variações cambiais, elementos de vaso de
grande diâmetro, dimorfismo dos vasos, grande quantidade de parênquima e pequena
proporção de fibras (Carlquist 1991; Bamber & Ter Welle 1994). As lianas têm caules finos
em relação à área foliar, possivelmente devido à reduzida demanda mecânica, visto que utiliza
outros suportes para a sustentação (Ewers & Fisher 1991; Ewers et al. 1991). Em seus finos
caules estão presentes vasos largos que conferem grande eficiência na condução (Ewers et al.
1991; Bamber & Ter Welle 1994), e muitas vezes, simultaneamente, vasos pequenos que
conferem segurança na condução (Carlquist 1975).
A família Leguminosae é uma das mais representativas em número de espécies de
lianas na região tropical (Gentry 1991), fato comprovado em levantamentos realizados na
Floresta Atlântica (Resende & Ranga 2005). A literatura relaciona estudos sobre a anatomia
de lianas da família Leguminosae (Ewers & Fisher 1991; Vieira 1994), contudo o número de
trabalhos ainda é pequeno quando comparado ao número total de espécies de lianas.
Neste trabalho foi realizado o estudo e a caracterização anatômica de oito espécies de
lianas da família Leguminosae, num remanescente de Floresta Atlântica da região sudeste do
Brasil (Parque Nacional do Itatiaia). Algumas das questões que vamos elucidar são: As
características anatômicas do lenho destas espécies seguem as tendências anatômicas das
lianas? Correspondem às características atribuídas para a família Leguminosae? Pela
anatomia do lenho é possível diferenciar estas espécies?

15
MATERIAL E MÉTODOS
As coletas foram realizadas no Parque Nacional do Itatiaia, município de Itatiaia,
Estado do Rio de Janeiro, sudeste do Brasil. Localizado entre as coordenadas geográficas
22°19’S e 22°45’S, 44°15’W e 44°50’W, possui área de 30.000,00 hectares (Ibama 2006). A
vegetação é do tipo Floresta Atlântica stricto senso (Oliveira-Filho & Fontes 2000). Na região
ocorre o predomínio de relevo montanhoso com bem delimitados cinturões climáticos e de
vegetação relacionados com o gradiente altitudinal. As coletas foram realizadas entre 700 m e
1100 m de altitude, onde apresenta média anual de precipitação de 1699 mm e média anual de
temperatura de 18,2° C. O clima é temperado úmido, tipo Cfb pelo sistema de Köppen’s
(Segadas-Vianna & Dau 1965).
Oito espécies de lianas pertencentes à família Leguminosae e às subfamílias
Mimosoideae e Papilionoideae, foram coletadas (Tabela 1). Foram selecionadas lianas com
diâmetro do caule maior do que 2,0 cm a aproximadamente 1,3 m acima do solo, sem injúrias
e deformidades, região onde foi removida a amostra do lenho. De cada indivíduo amostrado,
foi coletado um testemunho com estruturas reprodutivas e/ou vegetativas, que foi depositado
na coleção do Herbário do Jardim Botânico do Rio de Janeiro (RB). Foram registradas as
coordenadas geográficas do local de coleta e a altitude com aparelho GPS Garmin.
Para a microscopia óptica, os corpos de prova foram amolecidos, seccionados no
micrótomo de deslize Spencer 860 com espessura entre 16 a 30 µm (Ceccantini 1997). Em
seguida, foram clarificados, corados com safranina e azul de astra (Bukatsch 1972),
desidratados (Johansen 1940; Sass 1958) e montados com resina sintética. Para a descrição e
mensuração os elementos celulares foram dissociados utilizando-se a solução de Franklin
(Jane 1956). Os macerados foram corados com Safranina aquosa 1% e montados em lâminas
semi-permanentes com glicerina 50% (Strasburger 1924). Cortes histológicos foram corados
com lugol para observação de amido (Johansen 1940).
As lâminas obtidas foram observadas ao microscópio Olympus BX50 e as imagens
capturadas com o software Image Pro Plus versão 3.0 para Windows, ligado ao microscópio
através de uma câmera de vídeo Media Cybernetics CoolSNAP-Pro, estrutura usada para
proceder às medidas e contagens. Os cristais de oxalato de cálcio foram observados com
microscópio de luz polarizada Zeiss com filtro vermelho �.

16
As amostras de madeira e as lâminas permanentes foram incorporadas ao acervo da
Xiloteca do Jardim Botânico do Rio de Janeiro (RBw).
A terminologia utilizada nas descrições, assim como as contagens e as mensurações
seguem recomendações do IAWA Committee (1989). As variações cambiais foram referidas
seguindo Carlquist (2001). Para o cálculo da porcentagem de vasos, parênquima axial,
parênquima radial e fibras em relação ao tecido xilemático, utilizou-se uma grade sobreposta
às imagens capturadas do plano transversal, onde foram considerados trinta pontos formados
pela interseção das linhas da grade que se distanciavam em 100 µm. Foram observadas dez
imagens por indivíduo e calculou-se a porcentagem de cada imagem observada.
Com os dados quantitativos, foram realizadas análises estatísticas descritivas para a
caracterização e comparação das espécies. Também foram realizadas análises de inferência
estatística como a análise de distribuição amostral e testes de normalidade de Kolmogorov-
Smirnov. Foi utilizada a análise de fatores pelo método de componentes principais (PCA) na
definição das características que mais influenciaram na diferenciação das espécies,
influenciando no modelo espacial produzido (Manly 1994). A análise de fatores foi realizada
com 14 variáveis de um total de 47 variáveis. Estas 14 variáveis são: camadas de crescimento
distintas, anel semi-poroso, fibras septadas, parênquima axial vasicêntrico, parênquima axial
aliforme, parênquima axial marginal, parênquima axial difuso, estratificação, variação
cambial presente, diâmetro tangencial dos vasos, freqüência de vasos por milímetro linear,
comprimento dos vasos, largura do raio em micrômetros, altura do raio em micrômetros. As
demais variáveis foram excluídas da análise por não apresentarem variação ou por
apresentarem o valor dos fatores muito baixo. Com os dados qualitativos, foi realizada a
análise de agrupamento através de distâncias de Manhattan (Manhattan distances, Unweighted
pair-group average) (Manly 1994). Foram realizadas analises de variância (ANOVA) para
testar se existem diferenças significativas nas médias de algumas características quantitativas
entre algumas espécies. Todas as análises foram realizadas no programa Statistic 6.0 para
Windows.

17
RESULTADOS
Senegalia grandistipula (Benth.) Seigler & Ebinger (Fig. 1 e 9)
Camadas de crescimento: distintas a indistintas, de ocorrência esporádica delimitadas por
anel semi-poroso, zona fibrosa, achatamento radial de células no lenho tardio, máculas e
canais traumáticos.
Elementos de vaso: porosidade difusa, mas pode apresentar anel semi-poroso
esporadicamente; solitários e múltiplos de 2 a 21 vasos em cadeias radiais e cachos (1-3-21);
placas de perfuração simples; pontoações intervasculares alternas guarnecidas pequenas a
médias, com 5,59-7,31-9,88 µm; pontoações raiovasculares com bordas distintas; vasos com
duas classes de diâmetro; diâmetro tangencial dos maiores 75,30-196,32-330,79 µm; diâmetro
tangencial dos menores 16,26-37,23-108,34 µm; freqüência de 6-14-44 /mm2; comprimento
147,41-409,94-727,02 µm; obstruções por tilos e substâncias que coram em vermelho e azul
com safranina e azul de astra nos maiores e menores; representam 20-32-57% do tecido
xilemático.
Fibras: pontoações simples; com espessamento helicoidal; septadas e não septadas; presença
de fibras gelatinosas; paredes finas a espessas, espessura da parede 3,20-5,11-7,50 µm;
comprimento 407,20-1021,82-1786,20 µm; representam 7-21-53% do tecido xilemático.
Parênquima axial: paratraqueal vasicêntrico, aliforme e confluente; com 2 a 4 células por
séries parenquimáticas; representam 17-35-53% do tecido xilemático.
Raios: uniseriados e multiseriados, largura de 1-2-6 células e 11,49-25,62-50,63 µm; altura de
3-16-54 células e 50,78-284,01-1068,33 µm; com freqüência de 3-6-9 /mm’; composto
exclusivamente por células procumbentes; representam 3-12-23% do tecido xilemático.
Máculas e canais traumáticos: presentes.
Variação cambial: do tipo caule lobado conferindo aspecto quadrangular.
Inclusões minerais: presença de cristais prismáticos de oxalato de cálcio em séries
parenquimáticas axiais com 1 cristal por câmara, chegando a mais de 30 na série.
Amido: presente em fibras e no parênquima axial.
Material examinado: RBw 8643, 8644.

18
Senegalia lacerans (Benth.) Seigler & Ebinger (Fig. 2 e 10)
Camadas de crescimento: distintas a indistintas, de ocorrência esporádica, delimitadas por
anel semi-poroso, parênquima marginal, zona fibrosa, achatamento radial de células no lenho
tardio, máculas e canais traumáticos.
Elementos de vaso: porosidade difusa, mas pode apresentar anel semi-poroso
esporadicamente; solitários e múltiplos de 2 a 74 vasos em cadeias radiais e cachos (1-5-74);
placas de perfuração simples; pontoações intervasculares alternas guarnecidas pequenas a
grandes, com 4,88-7,40-12,33 µm; pontoações raiovasculares com bordas distintas; vasos de
duas classes de diâmetro; diâmetro tangencial dos maiores 41,63-174,12-328,45 µm; diâmetro
tangencial dos menores 11,49-27,18-56,23 µm; freqüência de 6-25-98 /mm2; comprimento
172,29-389,62-539,77 µm; obstruções por tilos e substâncias que coram em vermelho e azul
com safranina e azul de astra nos maiores e menores; representam 3-29-47% do tecido
xilemático.
Fibras: pontoações simples; com espessamento helicoidal; septadas e não septadas; presença
de fibras gelatinosas; paredes finas; espessura da parede 2,78-5,31-8,67 µm; comprimento
443,27-1151,29-1930,55 µm; representam 12-26-46% do tecido xilemático.
Parênquima axial: paratraqueal aliforme e confluente formando faixas; células fusiformes e
formando séries com 2 a 5 células; representam 19-33-60% do tecido xilemático.
Raios: uniseriados e multiseriados, largura de 1-3-10 células e 9,20-34,79-103,68 µm; altura
de 2-23-94 células e 43,74-304,21-1045,81 µm; com freqüência de 2-5-9 /mm’; composto
exclusivamente por células procumbentes; representam 0-12-21% do tecido xilemático.
Máculas e canais traumáticos: presentes.
Variação cambial: xilema fissurado.
Inclusões minerais: presença de cristais prismáticos de oxalato de cálcio em séries
parenquimáticas axiais com 1 cristal por câmara, chegando a mais de 30 na série.
Amido: presente em fibras e no parênquima axial.
Material examinado: RBw 8607, 8608, 8624, 8625, 8631, 8641, 8642.

19
Senegalia martiusiana (Steud.) Seigler & Ebinger (Fig. 3 e 11)
Camadas de crescimento: distintas a indistintas, de ocorrência esporádica, delimitadas por
anel semi-poroso, zona fibrosa, máculas e canais traumáticos.
Elementos de vaso: porosidade difusa, mas pode apresentar anel semi-poroso
esporadicamente; solitários e múltiplos de 2 a 26 vasos em cadeias radiais e cachos (1-3-26);
placas de perfuração simples; pontoações intervasculares alternas guarnecidas pequenas a
grandes, com 4,89-7,67-11,48 µm; pontoações raiovasculares com bordas distintas; vasos de
duas classes de diâmetro; diâmetro tangencial dos maiores 80,46-235,93-440,96 µm; diâmetro
tangencial dos menores 11,72-32,23-96,91 µm; freqüência de 7-19-61 /mm2; comprimento de
178,93-418,43-673,88 µm; obstruções por tilos e substâncias que coram em vermelho e azul
com safranina e azul de astra nos maiores e menores; representam 23-50-70% do tecido
xilemático.
Fibras: pontoações simples; com espessamento helicoidal; septadas e não septadas; presença
de fibras gelatinosas; paredes finas a espessas; espessura da parede 2,23-5,48-10,72 µm;
comprimento 405,13-1009,63-1867,58 µm; representam 0-8-31% do tecido xilemático.
Parênquima axial: paratraqueal confluente; células fusiformes e formando séries com 2 a 4
células; representam 17-32-53% do tecido xilemático.
Raios: uniseriados e multiseriados, largura de 1-2-4 células e 7,57-24,01-48,77 µm; altura de
2-15-47 células e 43,68-202,78-540,99 µm; com freqüência de 2-5-10 /mm’; composto
exclusivamente por células procumbentes; representam 3-10-27% do tecido xilemático.
Máculas e canais traumáticos: presentes.
Variação cambial: caule lobado com lobos cilíndricos.
Inclusões minerais: presença de cristais prismáticos de oxalato de cálcio em séries
parenquimáticas axiais com 1 cristal por câmara, chegando a mais de 30 na série.
Amido: presente em fibras e no parênquima axial.
Material examinado: RBw 8653, 8655, 8656, 8657, 8660, 8661.

20
Senegalia pedicellata (Benth.) Seigler & Ebinger (Fig. 4 e 12)
Camadas de crescimento: distintas a indistintas, de ocorrência esporádica, delimitadas por
máculas, canais traumáticos, anel semi-poroso e zona fibrosa.
Elementos de vaso: porosidade difusa, mas pode apresentar anel semi-poroso
esporadicamente; solitários e múltiplos de 2 a 15 vasos em cadeias radiais e cachos (1-3-15);
placas de perfuração simples; pontoações intervasculares alternas guarnecidas pequenas a
médias, com 5,13-7,68-10,91 µm; pontoações raiovasculares com bordas distintas; vasos de
duas classes de diâmetro; diâmetro tangencial dos maiores 66,83-131,19-287,36 µm; diâmetro
tangencial dos menores 13,40-27,65-58,20 µm; freqüência de 17-31-70 /mm2; comprimento
de 178,81-393,97-605,30 µm; obstruções por tilos e substâncias que coram em vermelho e
azul com safranina e azul de astra nos maiores e menores; representam 17-38-57% do tecido
xilemático.
Fibras: pontoações simples; com espessamento helicoidal; septadas e não septadas; presença
de fibras gelatinosas; paredes finas a espessas; espessura da parede 2,05-4,61-8,07 µm;
comprimento 484,75-976,12-1739,57 µm; representam 0-11-27% do tecido xilemático.
Parênquima axial: paratraqueal confluente; células fusiformes e formando séries com 2 a 5
células; representam 20-43-60% do tecido xilemático.
Raios: uniseriados e multiseriados, largura de 1-2-3 células e 5,58-18,78-34,48 µm; altura 2-
11-31 células e 25,29-178,85-490,87 µm; com freqüência de 3-6-10 /mm’; composto
exclusivamente por células procumbentes; representam 0-8-17% do tecido xilemático.
Máculas e canais traumáticos: presentes.
Variação cambial: caule lobado com lobos achatados.
Inclusões minerais: presença de cristais prismáticos de oxalato de cálcio em séries
parenquimáticas axiais com 1 cristal por câmara, chegando a mais de 30 na série.
Amido: presente em fibras e no parênquima axial.
Material examinado: RBw 8609, 8615, 8662, 8663, 8664.

21
Senegalia tenuifolia (L.) Britton & Rose (Fig. 5)
Camadas de crescimento: distintas, delimitadas por anel semi-poroso, parênquima marginal,
zona fibrosa, achatamento radial de células no lenho tardio.
Elementos de vaso: porosidade em anel semi-poroso; solitários e múltiplos de 2 a 26 vasos
em cadeias radiais e cachos (1-4-26); placas de perfuração simples; pontoações
intervasculares alternas guarnecidas pequenas a médias 5,11-7,14-9,54 µm; pontoações
raiovasculares com bordas distintas; vasos de duas classes de diâmetro ; diâmetro tangencial
dos maiores 81,24-189,19-361,33 µm; diâmetro tangencial dos menores 11,49-33,84-74,77
µm; freqüência de 6-13-35 /mm2; comprimento de 225,47-405,32-615,90 µm; obstruções por
tilos e substâncias que coram em vermelho e azul com safranina e azul de astra ; representam
12-33-57% do tecido xilemático.
Fibras: pontoações simples; com espessamento helicoidal; septadas e não septadas; presença
de fibras gelatinosas; paredes finas a espessas; espessura da parede 3,02-5,07-6,91 µm;
comprimento 570,94-1193,24-2491,97µm; representam 3-21-39% do tecido xilemático.
Parênquima axial: paratraqueal vasicêntrico, aliforme e confluente formando faixas
marginais; células fusiformes e formando séries com 2 a 4 células; representam 20-32-41%
do tecido xilemático.
Raios: uniseriados e multiseriados, largura de 1-4-8 células e 9,48-45,24-73,60 µm; altura 4-
60-277células e 57,65-713,10-3258,75 µm; com freqüência de 3-5-9 /mm’; composto
exclusivamente por células procumbentes; representam 7-14-23% do tecido xilemático.
Máculas e canais traumáticos: presentes.
Inclusões minerais: presença de cristais prismáticos de oxalato de cálcio em séries
parenquimáticas axiais com 1 cristal por câmara, chegando a mais de 30 na série. Cristais
abundantes nos limites das camadas de crescimento.
Amido: presente no parênquima axial e raios.
Material examinado: RBw 8628, 8658.

22
Piptadenia adiantoides (Spreng.) J.F. Macbr. (Fig. 6)
Camadas de crescimento: distintas, delimitadas por anel semi-poroso, parênquima marginal,
zona fibrosa, achatamento radial de células no lenho tardio.
Elementos de vaso: porosidade em anel semi-poroso; solitários e múltiplos de 2 a 33 vasos
em cadeias radiais e cachos (1-4-33); placas de perfuração simples; pontoações
intervasculares alternas guarnecidas pequenas a grandes 5,81-8,34-12,38 µm; pontoações
raiovasculares com bordas distintas; vasos de duas classes de diâmetro; diâmetro tangencial
dos maiores 95,62-272,51-480,28 µm; diâmetro tangencial dos menores 13,00-48,78-144,75
µm; freqüência de 2-12-42 /mm2; comprimento de 240,34-416,89-653,86 µm; obstruções por
tilos e substâncias que coram em vermelho e azul com safranina e azul de astra nos maiores e
menores ; representam 10-38-57% do tecido xilemático.
Fibras: pontoações simples; com espessamento helicoidal; não septadas e ocasionalmente
septadas; presença de fibras gelatinosas; paredes finas a espessas; espessura da parede 2,36-
4,67-8,38 µm; comprimento 460,18-957,71-1433,00 µm; representam 5-22-53% do tecido
xilemático.
Parênquima axial: paratraqueal escasso, vasicêntrico, aliforme e confluente formando faixas
marginais; células fusiformes e formando séries com 2 a 4 células; representam 10-24-42%
do tecido xilemático.
Raios: uniseriados e multiseriados, largura de 1-3-7células e 13,79-44,44-95,26 µm; altura 2-
25-113 células e 67,30-419,10-1365,41 µm; com freqüência de 3-7-12 /mm’; composto
exclusivamente por células procumbentes; representam 7-17-26% do tecido xilemático.
Máculas e canais traumáticos: presentes.
Inclusões minerais: presença de cristais prismáticos de oxalato de cálcio em séries
parenquimáticas axiais com 1 cristal por câmara, chegando a mais de 30 na série. Cristais
abundantes nos limites das camadas de crescimento.
Amido: presente em fibras e no parênquima axial.
Material examinado: RBw 8635, 8636, 8646, 8648, 8652.

23
Piptadenia micracantha Benth. (Fig. 7)
Camadas de crescimento: distintas, delimitadas por anel semi-poroso, parênquima marginal,
zona fibrosa e algumas vezes por achatamento radial de células no lenho tardio.
Elementos de vaso: porosidade em anel semi-poroso; solitários e múltiplos de 2 a 47 vasos
em cadeias radiais e cachos (1-4-47); placas de perfuração simples; pontoações
intervasculares alternas guarnecidas pequenas a grandes, com 6,30-9,76-14,02 µm;
pontoações raiovasculares com bordas distintas; vasos de duas classes de diâmetro; diâmetro
tangencial dos maiores 69,16-300,36-543,43 µm; diâmetro tangencial dos menores 13,40-
57,37-154,30 µm; freqüência de 2-11-32 /mm2; comprimento de 154,04-435,16-881,95 µm;
obstruções por tilos e substâncias que coram em vermelho e azul com safranina e azul de astra
nos maiores e menores; representam 7-34-67% do tecido xilemático.
Fibras: pontoações simples; com espessamento helicoidal; não septadas e ocasionalmente
septadas; presença de fibras gelatinosas; paredes finas a espessas; espessura da parede 2,96-
5,64-9,38 µm; comprimento 535,57-1019,71-1517,20 µm; representam 7-25-47% do tecido
xilemático.
Parênquima axial: paratraqueal vasicêntrico, aliforme e confluente formando faixas
marginais; células fusiformes e formando séries com 2 a 5 células; representam 10-26-47%
do tecido xilemático.
Raios: uniseriados e multiseriados, largura de 1-4-10 células e 16,09-51,24-121,32 µm; altura
3-31-117 células e 48,28-461,41-1627,59 µm; com freqüência de 1-5-10 /mm’; composto
exclusivamente por células procumbentes; representam 7-15-24% do tecido xilemático.
Inclusões minerais: presença de cristais prismáticos de oxalato de cálcio em séries
parenquimáticas axiais com 1 cristal por câmara, chegando a mais de 30 na série. Cristais
abundantes nos limites das camadas de crescimento.
Amido: presente em fibras e no parênquima axial.
Material examinado: RBw 8605, 8632, 8637, 8645, 8651, 8654.

24
Dalbergia frutescens (Vell.) Britton var. frutescens (Fig. 8)
Camadas de crescimento: distintas, delimitadas por anel semi-poroso, parênquima marginal,
zona fibrosa, achatamento radial de células no lenho tardio.
Elementos de vaso: porosidade em anel semi-poroso; solitários e múltiplos de 2 a 12 vasos
em cadeias radiais e cachos (1-2-12); placas de perfuração simples; pontoações
intervasculares alternas guarnecidas pequenas a grandes, com 5,81-8,86-15,61 µm;
pontoações raiovasculares com bordas distintas; diâmetro tangencial 16,09-102,50-360,95
µm; freqüência de 3-10-21 /mm2; comprimento de 98,48-201,62-260,22 µm; obstruções por
tilos e substâncias que coram em vermelho e azul com safranina e azul de astra; representam
0-15-47% do tecido xilemático.
Fibras: pontoações simples; com espessamento helicoidal; não septadas; presença de fibras
gelatinosas; paredes finas a espessas; espessura da parede 2,62-4,56-6,65 µm; comprimento
452,38-927,79-1381,08 µm; representam 11-40-68% do tecido xilemático.
Parênquima axial: paratraqueal aliforme, confluente formando faixas marginais, difuso e
difuso em agregados, células fusiformes e formando séries com 2 células; representam 18-33-
57% do tecido xilemático.
Raios: uniseriados e multiseriados, largura de 1-2-3 células e 9,20-25,69-50,63 µm; altura 3-
9-20 células e 52,30-156,09-312,65 µm; com freqüência de 3-8-13 /mm’; composto
exclusivamente por células procumbentes; representam 0-12-21% do tecido xilemático.
Estratificação: de raios, parênquima axial e vasos.
Máculas e canais traumáticos: presentes.
Inclusões minerais: presença de cristais prismáticos de oxalato de cálcio em séries
parenquimáticas axiais com 1 cristal por câmara, chegando a mais de 30 na série. Cristais
abundantes nos limites das camadas de crescimento.
Amido: presente no parênquima axial e no parênquima radial.
Material examinado: RBw 8606, 8613, 8614, 8616, 8621, 8622, 8649.

25
Todas as espécies estudadas apresentaram: elementos de vaso solitários e múltiplos em
cadeias radiais e cachos; placas de perfuração simples; pontoações intervasculares alternas
guarnecidas; pontoações raiovasculares com bordas distintas; obstruções por tilos e
substâncias que coram em vermelho e azul com safranina e azul de astra. Fibras com
pontoações simples; de paredes finas a espessas; gelatinosas; com espessamento helicoidal; e
sem septos. Parênquima axial paratraqueal confluente; com células fusiformes e formando
séries. Raios uniseriados e multiseriados; compostos exclusivamente por células
procumbentes. Cristais prismáticos em séries parenquimáticas axiais com 1 cristal por
subdivisão. Abundância de amido.
Todas as espécies pertencentes à sub-família Mimosoideae, S. grandistipula, S.
lacerans, S. martiusiana, S. pedicellata, S. tenuifolia, P. adiantoides, P. micracantha,
apresentaram fibras septadas, dimorfismo de vasos e amido em fibras.
Senegalia tenuifolia, P. adiantoides, P. micracantha e D. frutescens apresentaram
camadas de crescimento distintas, parênquima marginal e anel semi-poroso. Somente D.
frutescens apresentou estratificação, parênquima axial difuso e não apresentou dimorfismo de
vasos. Piptadenia adiantoides apresentou parênquima axial paratraqueal escasso. Em P.
micracantha não foram observadas máculas e canais traumáticos.
Quatro espécies do gênero Senegalia apresentaram variações cambiais produzidas a
partir de um único câmbio normal em produto, mas anormal em conformação. Em S.
grandistipula, S. martiusiana e S. pedicellata a variação cambial é do tipo caule lobado,
contudo existem diferenças entre estas espécies (Fig. 9-12). Em S. grandistipula os lóbos
conferem um formato quadrangular para o caule (Fig. 9), em S. martiusiana os lóbos são
cilíndricos (Fig. 11), enquanto em S. pedicellata os lóbos são achatados (Fig. 12). S. lacerans
apresenta a variação cambial do tipo xilema fissurado (Fig. 10). Nessas espécies ocorrem
camadas de crescimento esporádicas e foi comum a presença de máculas e canais traumáticos.
A média e a amplitude do diâmetro dos vasos foi elevada em P. micracantha e P.
adiantoides, chegando a 543,43 µm em P. micracantha. Dalbergia frutescens, em
contraposição, tem a menor média e do diâmetro dos vasos. Piptadenia micracantha
apresentou as maiores pontoações intervasculares. As espécies S. martiusiana e S. lacerans
apresentaram a maior freqüência de vasos por milímetro quadrado. Em D. frutescens os vasos
são curtos, com menor média e variação, além disso, tem menor numero de vasos agrupados.
Foi realizada a análise de distribuição amostral para o diâmetro dos vasos e constatou-se que
todas as espécies apresentaram distribuição bimodal, exceto D. frutescens.

26
As fibras são longas em S. tenuifolia, com média de 1193,24 µm. Essa espécie
também se destaca pela elevada média e máximo de altura do raio em número de células e
micrometros. Em antagonismo, D. frutescens apresenta raios baixos com uma pequena média,
amplitude e variação. Senegalia tenuifolia, P. adiantoides, P. micracantha formam um grupo
que difere das outras espécies por apresentar raios largos e altos. Dalbergia frutescens foi a
espécie que apresentou maior freqüência de raios por milímetro linear. Essa espécie também
apresentou maior porcentagem média de fibras e menor porcentagem média de vasos, quando
comparadas com as outras espécies. Todas as espécies apresentaram alta porcentagem de
vasos e parênquima axial, destacando-se S. martiusiana pela elevada média de vasos
representando aproximadamente 50% do tecido xilemático e S. pedicellata com média de
43% de parênquima axial. Senegalia tenuifolia, P. adiantoides, P. micracantha não
apresentaram tão altas proporções de parênquima axial, mas apresentaram as mais elevadas de
parênquima radial. Senegalia martiusiana e S. pedicellata apresentaram a menor quantidade
relativa de fibras. Dalbergia frutescens apresentou um pequeno desvio padrão nas medidas de
comprimento de vasos, altura do raio em número de células e em comprimento. Isso expressa
numericamente a estratificação de raios, vasos e do parênquima axial que ocorre nesta
espécie. O sumário das características quantitativas com a estatística descritiva pode ser
observado na tabela 1.
Por meio da análise de fatores pelo método de componentes principais, foi constado
que as características que mais influenciaram na distribuição espacial bidimensional das
amostras foram: presença de variação cambial, freqüência de vasos, presença de fibras
septadas, comprimento dos vasos, camadas de crescimento distintas, anel semi-poroso,
parênquima axial marginal, parênquima axial vasicêntrico, parênquima axial aliforme,
diâmetro tangencial dos vasos, altura e largura dos raios em micrômetros, estratificação,
parênquima axial difuso (Fig. 13); explicando 74% da variação. A distribuição espacial
bidimensional permite a distinção de 3 grupos. Um com P. adiatoides, P. micracantha e S.
tenuifolia, um segundo grupo com S. grandistipula, S. lacerans, S. martuisiana e S.
pedicellata, e um grupo isolado somente com D. frutescens (Fig. 14).
Na análise de agrupamento as características qualitativas que se mostraram
importantes foram: presença de variação cambial, o tipo de variação cambial, presença de
fibras septadas, camadas de crescimento distintas, anel semi-poroso, parênquima axial
marginal, parênquima axial vasicêntrico, parênquima axial paratraqueal escasso, parênquima
axial aliforme, estratificação e parênquima axial difuso. Contudo, P. micracantha e S.
tenuifolia não apresentam características qualitativas que permitam a distinção, permanecendo

27
agrupadas (Fig. 15). Piptadenia adiantoides apresenta o parênquima axial paratraqueal
escasso que a diferencia dessas duas espécies, mas por não ser uma característica observada
em todas as amostras, não interfere no agrupamento. Observou-se pela análise da variância
que a média do diâmetro dos vasos têm diferenças significativas entre as espécies S.
tenuifolia, P. adiantoides e P. micracantha (p < 0,01).

28
DISCUSSÃO
As espécies analisadas apresentam algumas características anatômicas do lenho
atribuídas às lianas (Carlquist 1991; Bamber & Ter Welle 1994). Quase todas as espécies têm
vasos com dois diâmetros distintos (dimorfismo de vasos), todas as espécies têm grande
diâmetro de vasos e quatro espécies do gênero Senegalia apresentam variações cambiais.
A ocorrência de vasos com grande diâmetro é freqüente nas lianas (Carlquist 1985).
Carlquist (1975), comparando hábitos relata a média do diâmetro dos vasos de trepadeiras e
lianas de 157 µm, comprimento de elementos de vaso com 334 µm e grande área condutora
(ou área de vasos por milímetro quadrado). Neste estudo, P. micracantha apresentou vasos
com diâmetro médio de 300,36 µm e máximo de 543,43 µm. Senegalia pedicellata
apresentou a segunda menor média de diâmetro com 131,19 µm, valor muito próximo ao
citado por Carlquist (1975) para lianas, e considerado elevado.
O dimorfismo de vasos confere grande eficiência na condução, através dos vasos
largos, e grande segurança, através dos vasos pequenos (Carlquist 2001). Contudo outros
autores (Ewers et al. 1991) mencionam que a função dos vasos estreitos é incerta, mas pode
ser importante para reserva e transporte de água no sentido radial e tangencial, na condução
quando os vasos grandes estão embolizados e fazendo ligações hidráulicas entre partes da
planta. Neste estudo foi constatado que ocorrem vasos de dois diâmetros distintos na maioria
das espécies analisadas. Os vasos com diâmetro elevado são predominantemente solitários,
enquanto os vasos de menor diâmetro encontram-se principalmente agrupados. O dimorfismo
de vasos pode ser constatado através da análise de distribuição amostral. Carlquist (1985)
relata que em plantas escandentes ocorrem vasos largos e estreitos concomitantemente, com
relativamente poucos vasos com valores intermediários diferindo de uma distribuição normal.
O termo “dimorfismo” não implica necessariamente que a distribuição amostral forme uma
curva bimodal, mas sim qualquer derivação de uma curva de distribuição normal (Ewers &
Fisher 1989; Carquist 1991). A atenuação da curva de distribuição normal foi comum em
dados analisados por Fisher & Ewers (1989). Sete das espécies estudadas apresentaram
distribuição bimodal e somente D. frutescens não apresentou. Neste estudo, D. frutescens não
apresentou dimorfismo de vasos detectável pela análise de distribuição amostral. Contudo
ocasionalmente vasos pequenos podem ser observados próximo a vasos grandes o que poderia
levar outros autores a considerar como dimorfismo de vasos.

29
Na maioria das espécies em estudo os elementos de vaso são curtos com
aproximadamente 400 µm em média, exceto em D. frutescens onde foram observados
elementos de vaso muito curtos, com média de 201,62 µm. A espécie que apresentou os
elementos de vaso mais longos foi P. micracantha, com média de 435,16 µm. Estudos têm
demonstrado que o comprimento dos elementos de vaso está mais relacionado com gêneros e
espécies próximas do que com o hábito (Carlquist 1985), contudo podem ser verificadas
diferenças quando comparadas lianas e árvores da mesma espécie. As Leguminosas
apresentam elementos de vaso muito curtos a médios não ultrapassando 800 µm (Baretta-
Kuipers 1981; Wheeler & Baas 1992), o que foi corroborado neste estudo.
A ocorrência de variações cambiais em lianas é bem conhecida. Contudo variação
cambial na subfamília Mimosoideae é pouco relatada. Evans et al. (2006), que estudaram a
anatomia do lenho de 70% dos gêneros da subfamília, constatou somente no gênero Entada
tal característica. Neste estudo Senegalia grandistipula, S. martiusiana, S. pedicellata e S.
lacerans apresentam um cambio único, normal em produtos mas anormal na conformação,
com formato lobado e não fragmentado no corte transversal nas três primeiras espécies citadas
e com xilema fissurado em S. lacerans. A variação que confere o formato lobado já havia sido
observada por Obaton (1960) em Acacia pennata e em outra espécie de Acacia por Netto
(1863 apud Obaton 1960). Nestes tipos de variação o câmbio produz floema para o lado
externo e xilema para o lado interno, contudo em algumas regiões o câmbio produz menos
xilema que em outras (Carlquist 2001). No tipo caule lobado a variação é notada no lado
externo do caule sem necessitar a secção transversal, contudo na espécie que apresenta xilema
fissurado a variação é somente notada após ser seccionado transversalmente. O xilema
fissurado é decorrente de uma pequena porção do câmbio que produz mais floema e menos
xilema, tornando-se quase unidirecional (Carlquist 2001). Em todas as espécies com variação
cambial analisadas geralmente ocorre a redução da produção de xilema em 4 regiões do caule.
Ao parênquima axial paratraqueal nas lianas é atribuída a função de recuperação do
tecido vascular após injuria (Fisher & Ewers 1989), aumento da flexibilidade (Putz &
Holbrook 1991) ou ainda a função de segurança e manutenção do sistema condutor,
hidrolisando o amido a açúcar, movendo este para a água presente no vaso e gerando pressão
osmótica que eleve a água a esta porção do vaso, evitando ou eliminando ar dos vasos largos
(Carlquist 1985). Reserva de amido no parênquima adjacente aos vasos foi relatado por
Carlquist (1985) em três gêneros de lianas da família Leguminosae (Hardenbergia, Mucuna,
Wisteria). Nesse estudo foi observado que o amido não se concentrava apenas no parênquima

30
adjacente aos vasos, mas estava presente em quase todas as células de parênquima axial.
Observou-se também que a maioria das espécies não apresentava amido no parênquima radial.
Apenas D. frutescens apresentou amido nos raios.
Nas espécies estudadas, o parênquima e os vasos compõe a maior parte do xilema
secundário. Somente Senegalia martiusiana apresentou maior proporção de vasos do que
parênquima. Em todas as espécies as fibras representam uma pequena parte do xilema
secundário. Dalbergia frutescens apresentou mais parênquima (45%) do que os outros tipos
celulares, mas foi a única espécie a ser constituída por mais fibras (40%) do que vasos (15%).
A presença de grande quantidade de parênquima nas lianas e a grande área condutora, que
pode ser entendida como uma grande proporção de vasos, já havia sido citada por vários
autores (Carquist 1985; Bamber & Ter Welle 1994) e também foi evidenciada nesse estudo.
Todas as espécies desse estudo apresentaram fibras gelatinosas. As fibras gelatinosas
geralmente são observadas nos caules de espécies arbóreas no lenho de tensão (Carlquist
2001) e também podem ser observadas nos caules de lianas, onde foi sugerido que podem
aumentar a flexibilidade (Vieira 1994).
Além das características esperadas para o hábito muitas outras das características
observadas devem-se à história evolutiva da família Leguminosae, de suas subfamílias,
gêneros e espécies. Diversas características anatômicas observadas no lenho, comuns a todas
as espécies estudadas, correspondem às características atribuídas à família Leguminosae, que
são: vasos solitários a múltiplos, placas de perfuração simples, pontoações intervasculares
alternas guarnecidas; fibras com pontoações simples; raios homocelulares compostos por
células procumbentes; parênquima axial paratraqueal, presença de cristais em células de
parênquima subdivididas (Baretta-Kuipers 1981; Wheeler & Baas 1992).
A maioria das Mimosoideae apresentam parênquima axial aliforme com diferentes
graus de confluência. As células do parênquima axial podem ser fusiformes ou formar séries
com mais de 8 células, sendo mais comum a ocorrência de séries de 2 a 4 células. A família
Mimosoideae apresenta a anatomia do lenho muito homogênea e distinguível das outras duas
subfamílias de Leguminosae (Evans et al. 2006). A estratificação é uma característica de
pouca incidência na subfamília Mimosoideae e comum na subfamília Papilionoideae (Baretta-
Kuipers 1981). Dalbergia frutescens, uma Papilionoideae, foi a única espécie deste estudo a
apresentar essa característica. Todavia, fibras septadas são comuns na subfamília
Mimosoideae (Baretta-Kuipers 1981) e foram observadas em todas as espécies desta
subfamília neste estudo. Uma das funções atribuídas às fibras septadas, também chamadas de
fibras vivas, é de reserva de amido (Carlquist 2001). Neste estudo foram realizados testes

31
histoquímicos e foi confirmada a presença de amido no interior das fibras septadas. Em
algumas fibras não septadas também se notou a presença de amido. Apesar de não terem sido
quantificadas, constatou-se que as fibras septadas foram mais abundantes nas espécies do
gênero Senegalia, e menos abundantes no gênero Piptadenia. A presença de fibras septadas
também é referida como comum em lianas da família Bignoniaceae (Gasson & Dobbins
1991). As características observadas nas espécies de Piptadenia e Senegalia se enquadram nas
descrições realizadas por Evans et al. (2006) para o grupo Piptadenia da tribo Mimoseae e
para o grupo Acacia da tribo Acaciaeae, respectivamente.
Bamber & Ter Welle (1994) estudando a liana Acacia albizioides observaram
diâmetro de vasos com média de 265 µm, 39% de parênquima, 2-3 células de largura e 1 mm
de altura dos raios, maior porcentagem de parênquima do que de fibras. No presente estudo,
as espécies do gênero Senegalia, antes tratadas com Acacia, apresentaram menor diâmetro
dos vasos do que o observado pelos referidos autores e a largura dos raios na maioria das
espécies foi semelhante, exceto para S. tenuifolia que apresentou raios largos. Nenhuma
espécie do gênero Senegalia, apresentou raios com altura média maior que 1 mm. Em S.
tenuifolia os raios chegam a atingir 3,26 mm, mas a média é de 0,71 mm. A porcentagem de
parênquima foi maior que a observada pelos autores citados acima, com nenhuma espécie
possuindo menos de 42% de parênquima.
A presença de cristais de oxalato de cálcio é uma característica conspícua em
Leguminosae. No lenho de Papilionoideae e Mimosoideae os cristais são usualmente
solitários, em câmaras compartimentalizadas (Zindler-Frank 1987). Todas as espécies
estudadas apresentaram cristais prismáticos de oxalato de cálcio, em séries parenquimáticas
com um cristal por subdivisão. As espécies com camadas de crescimento distintas
apresentavam várias séries cristalíferas em seu limite, Segundo Evans et al. (2006), na família
Mimosoideae os cristais são freqüentes no parênquima axial e comuns em fibras
compartimentalizadas. Esses autores citam uma maior freqüência de cristais nos limites entre
parênquima axial e fibras o que dificulta a identificação do tipo de célula os encerram (Evans
et al. 2006). A mesma dificuldade foi encontrada no presente trabalho.
O parênquima axial pode auxiliar na distinção de gêneros (Baretta-Kuipers 1981), a
exemplo de Dalbergia dos demais gêneros deste estudo. No entanto o padrão do parênquima
axial não permitiu a diferenciação das espécies do gênero Senegalia e Piptadenia. O gênero
Dalbergia apresenta uma combinação de características anatômicas do lenho que permite uma
fácil distinção dos outros gêneros de Leguminosae (Wheeler & Baas 1992). Isto também foi
notado neste estudo, onde D. frutescens, diferiu das demais espécies por apresentar:

32
parênquima apotraqueal difuso, difuso em agregados, estratificação e ausência de fibras
septadas. Estas características correspondem às citadas em outros estudos (Baretta-Kuipers
1981; Gasson 2000).
Nas lianas da subfamília Mimosoideae ocorrentes na área de estudo, os caracteres
morfológicos de folhas, algumas vezes não permitem a identificação das espécies, porém a
anatomia do lenho pode auxiliar no reconhecimento das espécies. Evans et al. (2006) referem
que as espécies de Mimosoideae, de um mesmo gênero geralmente apresentam um alto grau
semelhança na anatomia do lenho. No presente trabalho tal aspecto foi observado em espécies
de um mesmo gênero e de gêneros distintos. Senegalia tenuifolia, P. adiantoides e P.
micracantha têm a anatomia do lenho muito semelhante. Piptadenia adiantoides apresenta
parênquima axial paratraqueal escasso que a diferencia da demais, contudo não é uma
característica evidente em todas as camadas de crescimento e amostras. As três espécies
podem ser diferenciadas pelo diâmetro dos vasos, como foi constatado na análise de variância.
Piptadenia micracantha apresenta os maiores vasos, enquanto P. adiantoides e S. tenuifolia
apresentam vasos menores respectivamente. Senegalia tenuifolia também apresenta as fibras
longas e raios altos quando comparada às outras duas espécies.
Foi constatado então que as espécies estudadas apresentam características anatômicas
do lenho concordantes com características atribuídas às lianas e à família Leguminosae.
Apesar de serem muito semelhantes, principalmente as espécies pertencentes à subfamília
Mimosoideae, elas podem ser diferenciadas pela anatomia do lenho. Algumas mais
facilmente, como as espécies com variação cambial e D. frutescens, e outras que necessitam
de muita cautela e uma boa comparação para serem notadas as sutilezas das diferenças, como
em S. tenuifolia, P. adiantoides e P. micracantha.

33
AGRADECIMENTOS
Agradecemos ao Instituto de Pesquisas Jardim Botânico do Rio de Janeiro – Programa
Mata Atlântica e a CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior)
pelo auxílio financeiro; ao Parque Nacional do Itatiaia e ao Programa Mata Atlântica pelo
apoio logístico; e a Dra. Cecília Gonçalves Costa pela atenta revisão e sugestões no
manuscrito.
REFERÊNCIAS BIBLIOGRÁFICAS
Bamber, R.K. & Ter Welle, B.J.H. 1994. Adaptative trends in the wood anatomy of lianas. In:
Iqbal, M. (ed.). 1994. Growth Patterns in Vascular Plants. Portland, Oregon, Dioscorides
Press.
Baretta-Kuipers, T. 1981. Wood Anatomy of Leguminosae: its relevance to taxonomy. Pp.
677-705. In: Polhill, R.M. & Raven, P.H. (eds.). Advances in legumes systematics 1. Kew,
Royal Botanic Gardens.
Bukatsch, F. 1972. Bemerkungen zur doppelfärbung astrablau-safranin. Mikrokosmos 61 (8):
33-36.
Carlquist, S. 1975. Ecological Strategies of Xylem Evolution. California, The University of
California Press.
Carlquist, S. 1985. Observations on functional wood histology of vines and lianas: vessel
dimorphism, tracheids, vasicentric tracheids, narrow vessels, and parenchyma. Aliso 11 (2):
139-157.
Carlquist, S. 1991. Anatomy of vine and liana stems: a review and synthesis. Pp. 53-72. In:
Putz, F.E. & Mooney, H.A. (eds.). The Biology of Vines. Cambridge, United Kingdom,
Cambridge University Press.
Carlquist, S. 2001. Comparative wood anatomy: systematic, ecological and evolutionary
aspects of dicotyledon wood. Berlin, Springer-Verlag, 2a ed..
Ceccantini, G. 1997. Cortes de madeira em micrótomo de deslize. Pp. 125-127. In.: Kraus, J.
E. & Arduin, M., Manual básico de métodos em morfologia vegetal. Rio de Janeiro, Edur,
Apêndice III.
Evans, J. A.; Gasson, P.E. & Gwilym, P.L. 2006. Wood Anatomy of the Mimosoideae
(Leguminosae). IAWA Journal Supplement 5: 0-118.

34
Ewers, F.W. & Fisher, J.B, 1989. Variation in vessel length and diameter in stems of six
tropical and subtropical lianas. American Journal of Botany 76 (10): 1452-1459.
Ewers, F.W. & Fisher, J.B, 1991. Why vines have narrow stems: histological trends in
Bauhinia (Fabaceae). Oecologia 88: 233-237.
Ewers, F.W.; Fisher, J.B. & Fichtner, K. 1991. Water flux and xylem structure in vines. Pp.
127-160. In: Putz, F.E. & Mooney, H.A. (eds.). The Biology of Vines. Cambridge, United
Kingdom, Cambridge University Press.
Fisher, J.B. & Ewers, F.W. 1989. Wound healing in stems of lianas after twisting and girdling
injuries. Botanical Gazette 150 (3): 251-265.
Gasson, P.2000. Does wood anatomy support tribal and generic classification in Papilionoid
Leguminosae. Pp. 201-215. In: Herendeen, P.S. & Bruneau, A. (eds.). Advances in Legume
Systematics 9. Kew, Royal Botanic Gardens.
Gasson, P. & Dobbins, D.R. 1991. Wood anatomy of the Bignoniaceae, with a comparison of
trees and lianas. IAWA Bulletin 12 (4): 389-417.
Gentry, A. H. 1991. The distribution and evolution of climbing plants. Pp. 3-50. In: Putz, F.E.
& Mooney, H.A. (eds.), The Biology of Vines. Cambridge, United Kingdom, Cambridge
University Press.
Gerwing, J.J; Schnitzer, S.A.; Burnham, R.J.; Bongers, F.; Chave, J.; Dewalt, S.J.; Ewango,
C.E.N.; Foster, R.; Kenfack, D.; Martínez-Ramos, M.; Parren, M.; Parthasarathy, N.; Pérez-
Salicrup, D.R.; Putz, F.E.; Thomas, D.W. 2006. A Standard Protocol for Liana Censuses.
Biotropica 38(2): 256-261.
Iawa Committee 1989. List of microscopic feature of hardwood identification. IAWA
Bulletin 10 (3): 219-332.
Ibama 2006. Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis.
Parque Nacional do Itatiaia. Publicado na internet.
http://www.ibama.gov.br/parna_itatiaia/index.php?id_menu=155.
Jane, F.W. 1956. The structure of wood. Londres, Adam & Black.
Johansen, D. A. 1940. Plant microtechnique. New York, McGraw-Hill Book Co.
Manly, B. F. J. 1994. Multivariate statistical methods. London, Chapman & Hall, 2a ed..
Obaton, M. 1960. Les lianes ligneuses à structure anormales des forêts dense d´Afrique
occidentale. Annales des Sciences Naturelles, (Botanique) 12 (1): 1-220.
Oliveira-Filho, A.T. & Fontes, M.A.L. 2000. Patterns of Floristic Differentiation among
Atlantic Forests in Southeastern Brazil and the influence of Climate. Biotropica 32(4b) : 793-
810.

35
Putz, F.E. & Mooney, H.A. 1991. The Biology of Vines. Cambridge, United Kingdom,
Cambridge University Press.
Putz, F.E. & Holbrook, N.M. 1991. Biomechanical studies of vines. Pp. 73-98. In: Putz, F.E.
& Mooney, H.A. (eds.). The Biology of Vines. Cambridge, United Kingdom, Cambridge
University Press.
Resende, A.A. & Ranga, N.T. 2005. Lianas da Estação Ecológica do Nordeste Paulista, São
José do Rio Preto / Mirassol, SP, Brasil. Acta Botanica Brasilica 19(2): 273-279.
Sass, J.E. 1958. Elements of botanical microtechnique. New York – London, Mc. Graw –
Hill Bood Co.
Schnitzer, S.A. & Bongers, F. 2002. The ecology of lianas and their role in forests. Trends in
Ecology and Evolution 17 (5): 223-230.
Segadas-Vianna, F. & Dau, L. 1965. Ecology of the Itatiaia Range, Southeastern Brazil.
Arquivos do Museu Nacional 53: 31-53.
Strasburger, E. 1924. Handbook of practical botany. New York, MacMillan Company.
Vieira, R.C. 1994. Estrutura do Caule de Bauhinia radiata Vell. em Diferentes Ambientes.
Revista Brasileira de Biologia 54(2): 293-310.
Zinder-Frank, E. 1987. Calcium oxalate crystals in legumes. Pp. 279-316. In: Stirton, C.H.
(ed.). Advances in Legume Systematics 3. Kew, Royal Botanic Gardens.
Wheeler, E. & Baas, P. 1992. Fossil wood of the Leguminosae: A case study in xylem
evolution and ecological anatomy. Pp. 281-301. In: Herendeen, P.S. & Dilcher, D.L. (eds.).
Advances in Legume Systematics 4. Kew, Royal Botanic Gardens.
36
Diâ
met
ro ta
ngen
cial
dos
vas
os
Diâ
met
ro ta
ngen
cial
dos
vas
os
men
ores
Freq
üênc
ia d
e va
sos
(/m
m2 )
Com
prim
ento
dos
vas
os
Agr
upam
ento
de
vaso
s
Diâ
met
ro d
a po
ntoa
ção
inte
rvas
cula
r
Com
prim
ento
das
fibr
as
Esp
essu
ra d
a pa
rede
das
fibr
as
Diâ
met
ro d
o lú
men
das
fibr
as
Lar
gura
dos
raio
s (n
o.cé
lula
s)
Lar
gura
dos
raio
s (�
m)
Altu
ra d
os ra
ios
(no.
cél
ulas
)
Altu
ra d
os ra
ios
(�m
)
Freq
üênc
ia d
e ra
ios
(/m
m li
near
)
% d
e va
sos
% d
e pa
rênq
uim
a ax
ial
% p
arên
quim
a ra
dial
% d
e fi
bras
mím 75,30 16,26 6 147,41 1 5,59 407,20 3,20 3,98 1 11,49 3 50,78 3 20 17 3 7
méd 196,32 37,23 14 409,94 3 7,31 1021,82 5,11 7,61 2 25,62 16 284,01 6 32 35 12 21
máx 330,79 108,34 44 727,02 21 9,88 1786,20 7,50 13,20 6 50,63 54 1068,33 9 57 53 23 53
d.p. 54,82 17,27 9 112,05 4 0,93 314,41 1,02 2,48 1 9,26 11 214,24 2 8 9 6 11
mím 41,63 11,49 6 172,29 1 4,88 443,27 2,78 2,51 1 9,20 2 43,74 2 3 19 0 12
méd 174,12 27,18 25 389,62 5 7,40 1151,29 5,31 6,47 3 34,79 23 304,21 5 29 33 12 26
máx 328,45 56,23 98 539,77 74 12,33 1930,55 8,67 14,11 10 103,68 94 1045,81 9 47 60 21 46
d.p. 60,80 9,71 18 79,02 10 1,37 275,70 1,13 2,31 2 18,00 17 206,17 1 8 9 5 9
mím 80,46 11,72 7 178,93 1 4,89 405,13 2,23 1,92 1 7,57 2 43,68 2 23 17 3 0
méd 235,93 32,23 19 418,43 3 7,67 1009,63 5,48 6,64 2 24,01 15 202,78 5 50 32 10 8
máx 440,96 96,91 61 673,88 26 11,48 1867,58 10,72 15,95 4 48,77 47 540,99 10 70 53 27 31
d.p. 66,17 13,03 11 103,51 5 1,40 368,13 1,61 2,74 1 7,57 9 94,62 2 10 8 4 6
mím 66,83 13,40 17 178,81 1 5,13 484,75 2,05 2,72 1 5,58 2 25,29 3 17 20 0 0
méd 131,19 27,65 31 393,97 3 7,68 976,12 4,61 7,18 2 18,78 11 178,85 6 38 43 8 11
máx 287,36 58,20 70 605,30 15 10,91 1739,57 8,07 15,95 3 34,48 31 490,87 10 57 60 17 27
d.p. 35,98 9,27 10 97,32 3 1,15 284,91 1,15 2,55 1 6,01 6 91,93 2 9 8 4 7
mím 81,24 11,49 6 225,47 1 5,11 570,94 3,02 2,72 1 9,48 4 57,65 3 12 20 7 3
méd 189,19 33,84 13 405,32 4 7,14 1193,24 5,07 5,33 4 45,24 60 713,10 5 33 32 14 21
máx 361,33 74,77 35 615,90 26 9,54 2491,97 6,91 9,79 8 73,60 277 3258,75 9 57 41 23 39
d.p. 67,17 14,46 7 110,54 6 0,97 376,76 1,06 1,59 2 14,64 62 627,66 2 15 6 4 11
mím 95,62 13,00 2 240,34 1 5,81 460,18 2,36 2,95 1 13,79 2 67,30 3 10 10 7 5
méd 272,51 48,78 12 416,89 4 8,34 957,71 4,67 8,72 3 44,44 25 419,10 7 38 24 17 22
máx 480,28 144,75 42 653,86 33 12,38 1433,00 8,38 16,89 7 95,26 113 1365,41 12 57 42 26 53
d.p. 78,89 30,40 9 85,45 6 1,06 224,01 1,16 3,21 1 17,10 19 253,73 2 12 9 4 12
mím 69,16 13,40 2 154,04 1 6,30 535,57 2,96 2,72 1 16,09 3 48,28 1 7 10 7 7
méd 300,36 57,37 11 435,16 4 9,76 1019,71 5,64 6,54 4 51,24 31 461,41 5 34 26 15 25
máx 543,43 154,30 32 881,95 47 14,02 1517,20 9,38 13,84 10 121,32 117 1627,59 10 67 47 24 47
d.p. 99,41 30,21 8 111,18 7 1,38 201,25 1,24 2,20 2 18,19 26 345,53 2 11 7 4 11
mím 16,09 3 98,48 1 5,81 452,38 2,62 3,36 1 9,20 3 52,30 3 0 18 0 11
méd 102,50 10 201,62 2 8,86 927,79 4,56 9,71 2 25,69 9 156,09 8 15 33 12 40
máx 360,95 21 260,22 12 15,61 1381,08 6,65 19,85 3 50,63 20 312,65 13 47 57 21 68
d.p. 72,56 5 25,60 2 1,51 178,38 0,87 3,12 0 8,54 3 42,43 2 9 9 4 13
Tabela 1. Dados quantitativos das espécies estudadas. Para cada característica é apresentadamínima (mín), média (méd), máxima (máx) e desvio padrão (d.p.).
Senegalia pedicellata (Benth.) Seigler & Ebinger
Senegalia martiusiana (Steud.) Seigler & Ebinger
Senegalia lacerans (Benth.) Seigler & Ebinger
Senegalia grandistipula (Benth.) Seigler & Ebinger
Dalbergia frutescens (Vell.) Britton var. frutescens
Piptadenia micracantha Benth.
Piptadenia adiantoides (Spreng.) J.F. Macbr.
Senegalia tenuifolia (L.) Britton & Rose

37
Figuras 1 a 4 - Visão microscópica dos planos: tranversal (A), radial (B) e tangencial (C).
Fig. 1 - Senegalia grandistipula (Benth.) Seigler & Ebinger. Fig. 2 - Senegalia lacerans
(Benth.) Seigler & Ebinger. Fig. 3 - Senegalia martiusiana (Steud.) Seigler & Ebinger. Fig. 4
- Senegalia pedicellata (Benth.) Seigler & Ebinger. Escala = 100 µm.
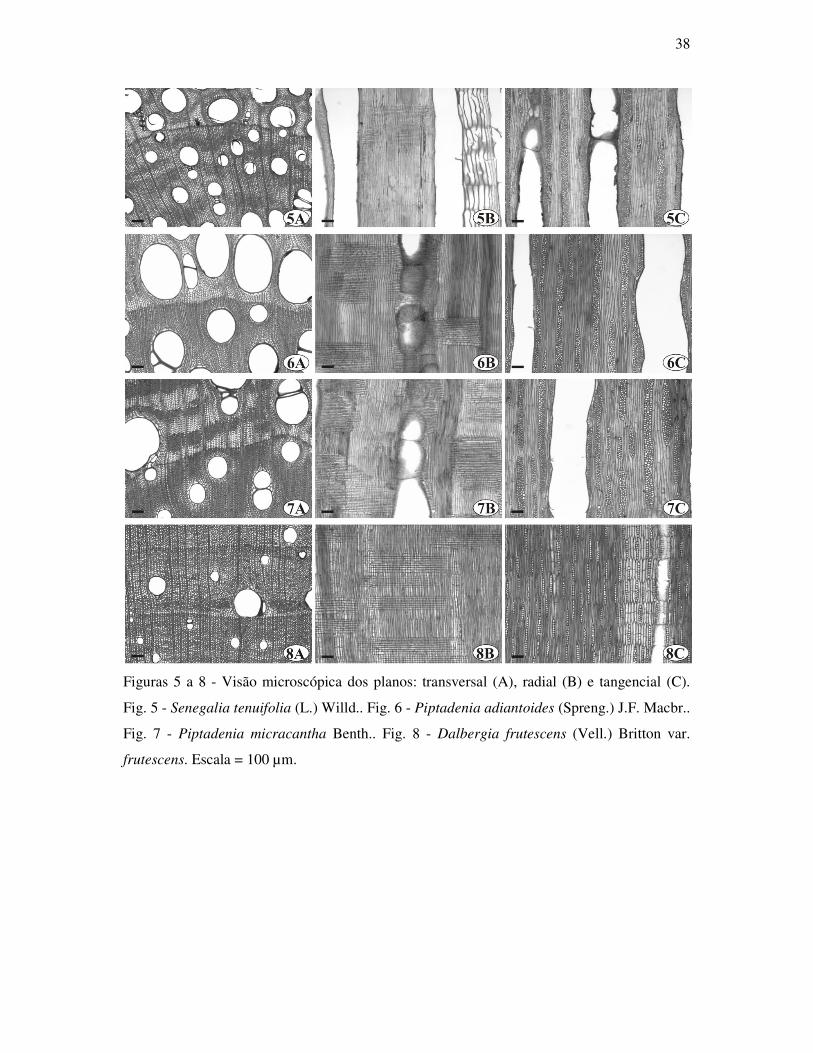
38
Figuras 5 a 8 - Visão microscópica dos planos: transversal (A), radial (B) e tangencial (C).
Fig. 5 - Senegalia tenuifolia (L.) Willd.. Fig. 6 - Piptadenia adiantoides (Spreng.) J.F. Macbr..
Fig. 7 - Piptadenia micracantha Benth.. Fig. 8 - Dalbergia frutescens (Vell.) Britton var.
frutescens. Escala = 100 µm.
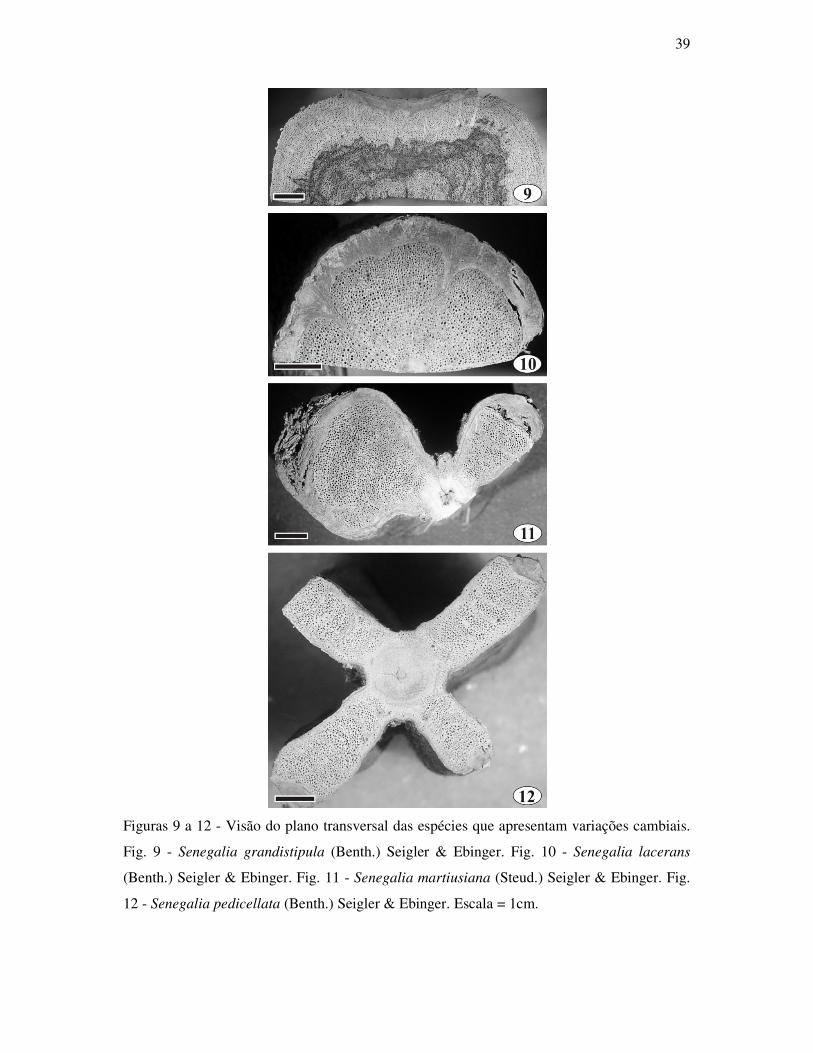
39
Figuras 9 a 12 - Visão do plano transversal das espécies que apresentam variações cambiais.
Fig. 9 - Senegalia grandistipula (Benth.) Seigler & Ebinger. Fig. 10 - Senegalia lacerans
(Benth.) Seigler & Ebinger. Fig. 11 - Senegalia martiusiana (Steud.) Seigler & Ebinger. Fig.
12 - Senegalia pedicellata (Benth.) Seigler & Ebinger. Escala = 1cm.
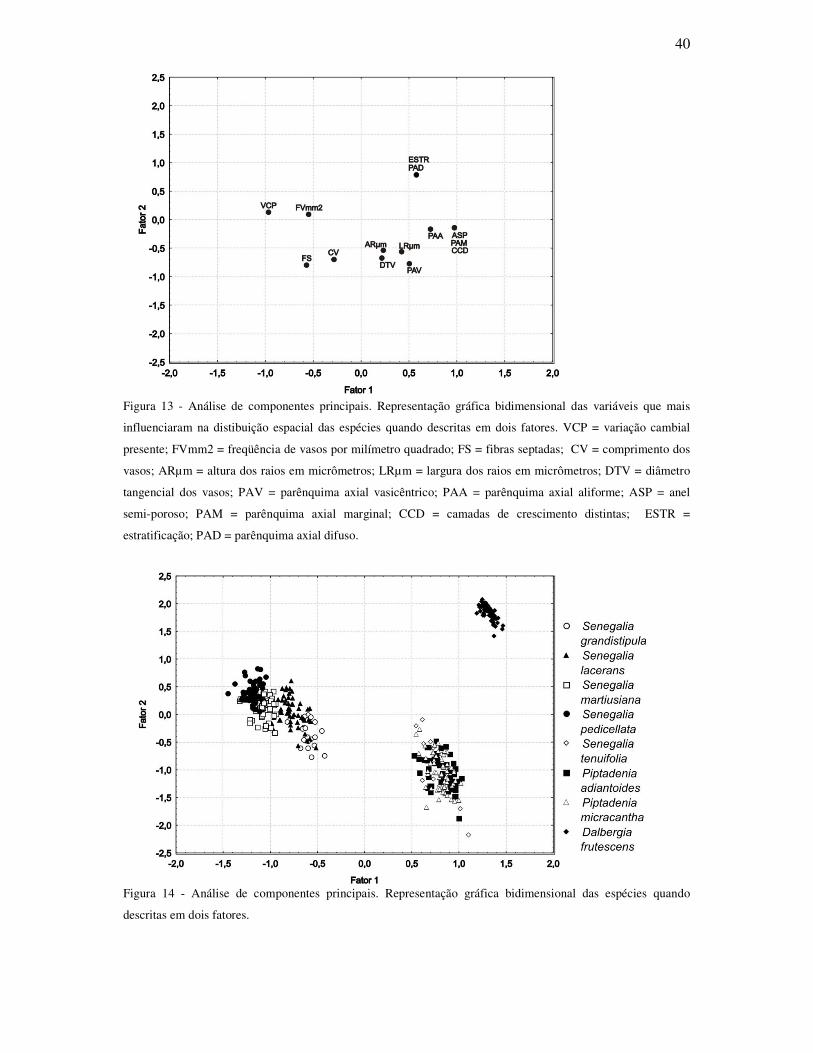
40
Figura 13 - Análise de componentes principais. Representação gráfica bidimensional das variáveis que mais
influenciaram na distibuição espacial das espécies quando descritas em dois fatores. VCP = variação cambial
presente; FVmm2 = freqüência de vasos por milímetro quadrado; FS = fibras septadas; CV = comprimento dos
vasos; ARµm = altura dos raios em micrômetros; LRµm = largura dos raios em micrômetros; DTV = diâmetro
tangencial dos vasos; PAV = parênquima axial vasicêntrico; PAA = parênquima axial aliforme; ASP = anel
semi-poroso; PAM = parênquima axial marginal; CCD = camadas de crescimento distintas; ESTR =
estratificação; PAD = parênquima axial difuso.
Figura 14 - Análise de componentes principais. Representação gráfica bidimensional das espécies quando
descritas em dois fatores.

41
Figura 15 - Análise de agrupamento. Dendrograma elaborado a partir de distâncias de
Manhattan entre as 8 espécies estudadas (Manhattan distances, unweighted pair-group
average). VCP = variação cambial presente; FS= fibras septadas; CCD = camadas de
crescimento distintas; ASP = anel semi-poroso; PAA = parênquima axial aliforme; PAM =
parênquima axial marginal; PAV = parênquima axial vasicêntrico; ESTR = estratificação;
PAD = parênquima axial difuso; PAE = parênquima axial paratraqueal escasso; CLQ = caule
lobado com aspecto quadrangular; CXL = caule com xilema fissurado; CLLC = caule lobado
com lobos cilíndricos; CLLA = caule lobado lobos achatados.

42
ARTIGO 2
Growth rings in lianas of Leguminosae family from
Atlantic forest in Rio de Janeiro, Brazil
Arno F. N. Brandes1* and Cláudia F. Barros1
1) Escola Nacional de Botânica Tropical, Instituto de Pesquisas Jardim Botânico do Rio de
Janeiro, Rua Pacheco Leão 915, Jardim Botânico, CEP 22460-030, Rio de Janeiro, Brazil.
Short running title: Growth rings in lianas
Artigo submetido ao “Botanical Journal of Linnean Society”

43
ABSTRACT
The anatomical features of growth rings of 8 lianas from Leguminosae family, occurring in
mountainous relief within Atlantic forest of the Rio de Janeiro State, south-eastern of Brazil,
were studied. Four species showed distinct growth rings. The others presented cambial
variants and growth rings of sporadic occurrence. The cambial variants observed were stem
with xylem furrowed and lobed stem. Semi-ring-porous, marginal parenchyma, fibrous zone
and radially flattened latewood cells were the main features that mark the growth rings. The
species with cambial variants have some boundaries made by pith flecks and traumatic canals.
Piptadenia adiantoides, Piptadenia micracantha, Senegalia tenuifolia and Dalbergia
frutescens show well-distinct growth rings in macroscopy and microscopy, that suggests the
utilization in futures growth rings researches.
ADDITIONAL KEYWORDS: cambial variants – Mimosoideae – Papilionoideae – secondary
xylem – tropical region – wood anatomy.

44
INTRODUCTION
In tropical forests, lianas are important component playing an essential ecological role in
many aspects. Over the past two decades has been an increasing interest and awareness for
this growth form, although it is still poorly understood in many aspects (Gentry, 1991;
Schnitzer & Bongers, 2002; Pérez-Salicrup, Schnitzer & Putz, 2004; Gerwing et al., 2006).
Little attention has been spent on anatomy of lianas (Carlquist, 1985; Bamber & Ter Welle,
1994) and almost none to growth rings in lianas, which few papers report this feature and it is
not focused on researches (Baas & Schweingruber, 1987; Gasson & Dobbins, 1991; Carlquist,
1995; Araque, De Pernía & León, 2000). Information about growth rings in trees is
accessible, but from others growth forms is scarce. Schweingruber & Poschlod (2005) widely
studied growth rings in herbs and shrubs, and they report distinct growth rings on these other
growth forms.
In the last decades, many dendrochronologic studies in tropical tree species did report
occurrence of seasonal growth rings, correlated with some climatic variations (Mariaux, 1981;
Villalba, Boninsegna & Holmes, 1985; Détienne, 1989; Worbes, 1989, 1995; Buckley et al.,
1995; Devall, Parresol & Wright, 1995; Gourlay, 1995a, 1995b; Heinrich & Banks, 2006;
Westbrook, Guilserson & Colinvaux, 2006). In Brazil, some studies were developed in the
Amazon forest (Botosso, 1984; Worbes, 1985; Vetter & Botosso, 1989) and in the Atlantic
forest biomes (Seitz & Kanninen, 1989; Callado et al., 2001a, 2001b, 2004; Marcati,
Angyalossy & Evert, 2006). The overview of tree-ring research in tropics is given by Worbes
(2002).
Lianas are woody climbers which development begins on the ground but their ability to self-
support is lost with the growth; they need a support to ascend to canopy and they are able to
grow in mature forests (Gentry, 1991; Gerwing et al., 2006). Most of lianas shows cambial
variants and a few parts have stem anatomy similar of those observed on trees and shrubs, in
which the cambium is single and cylindrical and produces internally secondary xylem and
externally secondary phloem, with alike amount in all portions of stem (Carlquist, 1991,
2001; Caballé, 1993). However, it follows anatomical tendencies correlated to habit, e.g.
higher conductive area and low proportion of supporting tissues (Carlquist, 1991).
Lianas are an important structural component of tropical forests and represent around of 25 %
in abundance and species richness (Gentry, 1991; Schnitzer & Bongers, 2002; Pérez-Salicrup
et al., 2004). Leguminosae is the third largest family of climbers in the New World and with
Bignoniaceae they are dominant climbing families in neotropical lowland forests (Gentry,

45
1991). Two liana censuses developed in Atlantic Forest biome show Leguminosae as the
second and the third most representative family in species number for studied ranges
(Resende & Ranga, 2005).
Brazilian Atlantic Forest has high species diversity and number of endemics species, but
retains only approximately 7,5 % of its original extension and it is one of the highest global
priorities for conservation (Morellato & Haddad, 2000; Myers et al., 2000). This biome
should be considered a mosaic comprising several types of vegetation, related with several
environments (Oliveira-Filho & Fontes, 2000; Scarano, 2002). The bulk of Brazilian Atlantic
Forest is on tropical region.
Growth rings analysis would contribute with temporal information that can be used in studies
of growth, wood production, age structure, population and forest dynamics, forest
management, conservation and restoration (Jacoby, 1989; Eickstein, Sass & Baas, 1995; Priya
& Bhat, 1999; Braker, 2002; Worbes, 2002; Grau, Easdale & Paolini, 2003; Worbes et al.,
2003; Brienen & Zuidema, 2006). Then, if some lianas have distinct growth rings, this might
contribute to the knowledge of the growth in lianas, the understanding of the tropical forest
dynamics and with sustainable management.
In this study, the anatomical features of eight liana species were analysed from a well-
preserved remnant of Atlantic Rain Forest, placed in mountain region (Serra da Mantiqueira –
Itatiaia Massif). This species represent almost all lianas of Leguminosae family recognized
from this place and registered in Programa Mata Atlântica data base. Some of the questions
that we try to answer are: Do lianas have growth rings? What are the anatomical features that
distinguish the growth rings? Species with cambial variants have growth rings? Some of
studied species can be used in future researches of dendrochronology?

46
MATERIALS AND METHODS
There were sampled fifty-seven lianas from eight species belonging to Leguminosae family,
subfamilies Mimosoideae and Papilionoideae (Table 1). The samples were collected at the
Parque Nacional do Itatiaia, municipality of Itatiaia, State of Rio de Janeiro, south-eastern
Brazil. Placed between geographical coordinates 22°15’ and 22°30’ S, 44°30’ and 44°45’ W,
within an area of 28,155.97 hectares (http://www.ibama.gov.br). The vegetation type is
Atlantic rain forest (Oliveira-Filho & Fontes, 2000). The Itatiaia range has a predominant
mountainous relief with several well-delimited climatic and vegetation belts related with
altitudinal gradient. The belt where the samples were taken is between 700 m and 1,100 m,
with annual mean precipitation of 1,699 mm, and annual mean temperature 18.2 °C. This
ambient is warm and rainy from December to February and cold and dry from June to August.
From December to February falls 50 % of the annual precipitation and January is the rainiest
month with an average of 289.2 mm. In these months the mean of temperature is 20.8°C.
From June to August falls only 5 % of the annual precipitation and the least rainy month is
June with average 31.5 mm. This cold and dry season has the mean temperature of 15.1 °C,
but was registered absolute minimum of 0 °C. The climate is temperate and humid, and by
Köppen’s system is the Cfb type (Segadas-Vianna & Dau 1965).
There were selected for this study lianas with diameter larger than 2.0 cm at approximately
1.3 m above the ground without wounds or deformities in this height. The number of samples
per species differed by the difficult to meet individuals with characteristics cited above and
different abundance of each species. From each sampled individual, a voucher was also
collected, with reproductive and others vegetative parts, which were deposited in herbarium
collection of the Jardim Botânico do Rio de Janeiro (RB).
Wood samples of stems were collected at approximately 1.3 m above the ground. There were
taken partial or complete cross-sections with approximately 2 cm length, including bark,
wood and some times the medulla. Blocks were polished for macroscopic observation with
hand lens and stereomicroscope Olympus SZ11. The images were taken with photographic
camera Olympus C5050 attached to the same stereomicroscope.
For light microscopy, boiling water and glycerine were used to soft part of the blocks.
Sections with 16 to 30 µm thick were made in transversal, tangential and radial plains with
Spencer 860 sliding microtome. After that they were cleaned, stained with safranin and astra
blue (Bukatsch, 1972), dehydrated and mounted on Permount resin in permanent slides.
Slides were observed at Olympus microscope BX50 and images captured with system Image

47
Pro Plus version 3.0 for Windows linked at that microscope through a Media Cybernetics
CoolSNAP-Pro video camera. The polarized light microscope Zeiss with red filter � was also
used for calcium oxalate crystals observation.
Wood samples and permanent slides were incorporated in the wood collection of the Jardim
Botânico do Rio de Janeiro (RBw). Terminology follows the IAWA Committee (1989).
Others considerations about growth rings and cambial variants follow classifications proposed
by Carlquist (2001).

48
RESULTS
GENUS SENEGALIA – MIMOSOIDEAE
Senegalia grandistipula (Benth.) Seigler & Ebinger shows cambial variant lobed stem given
to stem the appearance of square and growth rings boundaries of sporadic occurrence (Figure
1A, 2A). The distinctness in microscopic view is possible by fibrous zone and semi-ring-
porous, in some portions of stem. With less incidence radially flattened latewood cells and by
pith flecks and traumatic canals. The macroscopic and microscopic observations provide
distinction of growth rings boundaries, but macroscopic view facilities the detection of semi-
ring porosity.
Senegalia lacerans (Benth.) Seigler & Ebinger shows cambial variant xylem furrowed and
growth rings boundaries of sporadic occurrence, allowing the observation in some portions of
the samples (Figure 1C, 2B). The distinctness in microscopic view is possible by semi-ring-
porous; fibrous zone; marginal parenchyma. With less frequency, radially flattened latewood
cells, pith flecks and traumatic canals. The macroscopic and microscopic observations provide
distinction of growth rings boundaries.
Senegalia martiusiana (Steud.) Seigler & Ebinger shows cambial variant lobed stem, with
cylindrical lobes and growth rings boundaries of sporadic occurrence, allowing the
observation in some portions of samples (Figure 1D, 2C). The features that allow the
distinction in microscopic view are semi-ring-porous; fibrous zone; pith flecks and traumatic
canals. The macroscopic and microscopic observations provide distinction of growth rings
boundaries.
Senegalia pedicellata (Benth.) Seigler & Ebinger shows cambial variant lobed stem with
flattened lobes and growth rings boundaries of sporadic occurrence (Figure 1B, 2D). The
distinction in microscopic view is possible, in some portions of stem, mainly by pith flecks
and traumatic canals. But occasionally it is also possible by semi-ring-porous and fibrous
zone. The macroscopic and microscopic observations provide distinction of growth rings
boundaries.

49
Senegalia tenuifolia (L.) Britton & Rose (Figure 3D, 4A) in microscopic observation shows
distinct growth rings by semi-ring-porous; fibrous zone in latewood, radially flattened
latewood cells; and marginal parenchyma. Fibrous zone and radially flattened latewood cells
are not ever well-distinct on all samples and on all growth rings, but the absence of both
features in the same growth ring is rare. Abundance of calcium oxalate prismatic crystals was
observed in transition of growth rings. The macroscopic and microscopic observations
provide easy distinction of growth rings boundaries.
GENUS PIPTADENIA – MIMOSOIDEAE
Piptadenia adiantoides (Spreng.) J.F. Macbr. (Figure 3B, 4B) in microscopic observation
shows distinct growth rings boundaries marked by combination of: semi-ring-porous; fibrous
zone; radially flattened latewood cells; marginal parenchyma. Abundance of calcium oxalate
prismatic crystals in transition of growth rings was observed. Two specimens do not have
well-distinct semi-ring porosity, showing a thin layer of vessel with few diameters in
latewood. In the same two specimens not all growth ring boundaries have well distinct fibre
zone, because the pattern of decrease of confluence parenchyma bands toward the latewood
making fibre zones are not followed. The macroscopic observation provides easy distinction
of growth rings boundaries than microscopic observation, because the presence of axial
parenchyma lignified with thicker walls.
Piptadenia micracantha Benth. (Figure 3C, 4C) in microscopic observation shows distinct
growth rings boundaries marked by combination of: semi-ring-porous; fibrous zone; marginal
parenchyma. Few growth rings show radially flattened cells on latewood. Abundance of
calcium oxalate prismatic crystals in transition of growth rings was observed (Figure 5). The
macroscopic and microscopic observations provide easy distinction of growth rings
boundaries. Piptadenia micracantha display to be the species with the anatomical features
more constants delimiting growth rings.
GENUS DALBERGIA – PAPILIONOIDEAE
Dalbergia frutescens (Vell.) Britton var. frutescens (Figure 3A, 4D) in microscopic
observation has distinct growth rings boundaries marked by semi-ring porosity; marginal
parenchyma; fibrous zone in latewood; and radially flattened latewood cells. The features

50
fibrous zone and radially flattened latewood cells are not ever well-distinct on all samples and
on all growth rings, but the absence of both features in the same growth ring is unusual. The
macroscopic and microscopic observations provide easy distinction of growth rings
boundaries, however a broad view field of macroscopy facilitates the observation.

51
DISCUSSION
Distinct growth rings were observed in four of the eight studied species (Figure 3A-D, 4A-D).
The others presented cambial variants and growth rings boundaries of sporadic occurrence
(Figure 1A-D, 2A-D). The cambial variants observed were lobed stem but not fragmented and
stem with xylem furrowed, products of the single cambium but with unequal activity on some
regions, in studied species generally on four regions of the stem (Table 1).
The two species of Piptadenia genus show distinct growth rings boundaries. In four species of
the Senegalia genus, the occurrence of cambial variants and sporadic growth rings boundaries
were predominant. Only one studied species of Senegalia genus does not show cambial
variants and has distinct growth rings boundaries. The unique studied species of Dalbergia
genus shows distinct growth ring boundaries.
IAWA Committee (1989) states that distinctness of the ring boundaries should be judged with
a light microscope and when growth rings are nonperiodical and of sporadic occurrence
should be recorded as rings absent or indistinct. Therefore, S. grandistipula, S. lacerans, S.
martiusiana, and S. pedicellata should be classified as indistinct growth rings because of the
sporadic occurrence of ring boundaries. Many of these sporadic and nonperiodical growth
rings are linked with traumatic events.
The other four species have distinct growth rings and were classified within the types 1B, 1D,
8 and 11, descript by Carlquist (2001). The type 1B was observed in P. micracantha and D.
frutescens, and related to the reduction of vessel diameter from earlywood to latewood and the
other cell types unaffected. The type 1D was observed in P. adiantoides, S. tenuifolia and D.
frutescens, it is related to reduction of vessel diameter and imperforate traqueary elements
from earlywood to latewood, this may be associated with the feature radially flattened
latewood cells. The type 8 may be applicable to P. micracantha and P. adiantoides, which
some samples have much wider vessels in earlywood than in latewood. The type 11, related to
marginal parenchyma and vessel elements change from wide to narrow, can be associated to
four species with distinct growth rings, because all of them show marginal parenchyma and
present semi-ring porosity. Categorize the subtypes of the type 11 was not possible, because
the period of formation of marginal parenchyma was not investigated.
The more evident anatomical features of growth rings for species with distinct growth ring
are: semi-ring porosity, fibrous zone, marginal parenchyma and radially flattened cell in
latewood. The species with cambial variant also have traumatic canals and pith flecks
delimiting some growth boundaries, this caused by natural wounding. Some features are not

52
ever present in all samples from each species and in all growth rings from each sample. The
macroscopic observation provides the distinction by the same features of microscopic
observation; with exception of radially flattened latewood cells that is only possible to detect
in microscopic view. Generally, the macroscopic view facilitates the detection of growth rings
boundaries because of a broad field view.
The features that distinguish the growth ring boundaries observed in this study are frequently
associated to a great number of the tropical tree species (Boninsegna et al. 1989; Détienne,
1989; Worbes, 1989; Heinrich et al., 2006; Marcati et al., 2006), and may be viewed in
several combinations within Leguminosae family, e.g.: marginal parenchyma bands
(Boninsegna et al., 1989; Détienne, 1989; Worbes, 1989; Gourlay, 1995a, 1995b; Eshete &
Stahl, 1999; Roig et al., 2005); radially flattened and thick-walled latewood fibres (Barros &
Callado, 1997; Barros et al., 2001); marginal parenchyma bands and thick-walled fibres in the
latewood (Callado et al., 2001a); marginal parenchyma, radially flattened and thick-walled
latewood fibres (Vetter & Botosso, 1989, Tomazello Filho et al., 2004); marginal parenchyma
bands, and pattern of concurring parenchyma and fibre tissue (fibrous zone) (Worbes, 2002),
change in vessel diameter and marginal parenchyma (Villalba & Boninsegna, 1989; Richter &
Dallwitz, 2000)
Wood anatomy studies related to Leguminosae systematic show that many species present
diffuse porosity and only few species have ring-porous or semi-ring-porous (Wheeler & Baas,
1992; Gasson, 1994, 2000; Höhn, 1999). Some African Caesalpinioideae and Mimosoideae
species when grouped by wood anatomy produce eighteen groups all having diffuse porous,
among them, seven show distinct growth rings and have marginal parenchyma (Höhn, 1999).
The occurrence of mainly diffuse porous was not verified in this study, which one half of
species shows diffuse porosity and other half shows semi-ring-porous.
Studies with African Acacia species show that they have growth rings distinguished mainly
by marginal parenchyma, many times represent one year (Gourlay, 1995a, 1995b; Eshete &
Stahl, 1999) and abundant prismatic crystals of calcium oxalate are related to marginal
parenchyma (Gourlay, 1995a, 1995b). In present study S. tenuifolia and the other three
species with distinct growth rings have abundant calcium oxalate prismatic crystals related to
marginal parenchyma. The others Senegalia species with growth rings of sporadic occurrence
also show calcium oxalate prismatic crystals related to marginal parenchyma in distinct
boundaries. The Senegalia species in the present study before belonged to Acacia genus.
Recently the New World taxa from Acacia subgenus Aculeiferum Vassal were relocated to
Senegalia genus (Seigler et al., 2006).

53
In the last decades the knowledge about growth rings in tropics improved, but its occurrence
remains with low scientific acceptance (Worbes, 2002). This laid on concept that tropical
region lack marked seasons, which produce distinct growth rings. But the contrary had been
demonstrated in many studies, in which approximately half of species from these regions had
distinct growth rings. The same was realized in this study. Alves & Angyalossy-Alfonso
(2000) observed ecological trends in some Brazilian tree species, which detect that 48 % of
the studied species have growth rings correlated positively with mild and medium
mesothermic climates and negatively related to super-humid environments. In this study 136
species of Leguminosae family were sampled and 71 species presented distinct growth rings.
Among them, four species are of the genus Piptadenia and five of genus Dalbergia, including
D. frutescens. The interactive key for commercial timbers, from Delta Databases (Richter &
Dallwitz, 2000), indicate eleven species with distinct growth ring of 24 Leguminosae species
from tropical South America. Seven with marginal parenchyma and among them two have
semi-ring-porosity. Dalbergia decipularis Matt. & Rizz. and Piptadenia macrocarpa Benth.
are examples of tree species with distinct growth ring from tropical South America. Roig et
al. (2005) report that 42 % of the studied Leguminosae species in Yucatan Peninsula had
visible rings. These previous studies and the present study show that growth rings in
Leguminosae species, trees and lianas, from tropical region is a frequent feature.
Growth rings in lianas were detected in other studies. Gasson & Dobbins (1991) report
distinct growth rings in Bignoniaceae family, genus Capsis from tribe Tecomeae, in genus
Schlegelia from tribe Schlegelieae and in nine genera from tribe Bignonieae. Among diverse
plant habits from Europe, Baas & Schweingruber (1987) observed that climbers show porous-
ring, semi-ring-porous. But they do not sampled Leguminosae species and the samples
analysed were from temperate climate. In other study within lianas in Venezuela, it was
observed growth rings in Anomospermun schomburgkii Miers (Menispermaceae) and
Strycnos brachistantha Standley (Loganiaceae), marked by marginal parenchyma (Araque et
al., 2000). These two species showed diffuse porosity. Semi-ring porous and marginal
parenchyma were important features observed in lianas analysed on this study, because they
are present in all species with distinct growth ring. The growth rings are made of variation in
cambium activity and its products, and each ring may represent a period of one year (Fahn,
1982). A cambium activity study detected in four lianas from a Mexican tropical rainforest
that cambium is active along all year. Although, three species showed strong cambial
seasonality, differing by number of cambial layers, positively associated with rainfall. Two

54
species of Machaerium (Leguminosae - Papilionoideae) was studied and presented cambial
seasonality (Leon-Gómez & Monroy-Ata, 2005).
The changes in anatomical structure, which produces growth rings, are a dynamic process
affected by many ecological factors (Wimmer, 2002). Some factors that make cyclic
responses in tree growth are: water supply; temperature; photoperiod; light intensity; and
phenological behaviors, which control or are controlled by hormones (Amobi, 1974; Mariaux,
1981; Tomlinson & Longman, 1981; Carlquist, 2001). Borchert (1999) detect that the
phenology and cambial activity is affected by rainfall seasonality in tree species growing in
dry sites, and most of them have annual growth rings. In brevideciduous and evergreen
species the consecutive reduction in correlation of phenology and cambium activity with
climatic seasonality was noticed. These factors may act in lianas and produce growth
responses; making distinct growth rings or producing growth rings of sporadic occurrence, as
observed in this study.
The results obtained by this study demonstrate that P. micracantha, P. adiantoides, S.
tenuifolia and D. frutescens might be used for further dendrochronological studies. Because
the very clear rings visible under microscopy and macroscopy, which are fundamental to
enable a precise count and width measurement. Several species of the same genus or the same
species are used or indicated in literature for growth rings researches, e.g. Piptadenia excelsa
(Grau et al., 2003), Dalbergia ecastaphyllum (Amobi, 1974), D. frutescens and four
Piptadenia species (Alves & Angyalossy-Alfonso, 2000), Acacia species (Gourlay, 1995a,
1995b; Eshete & Stahl, 1999). Senegalia grandistipula, S. lacerans, S. martiusiana and S.
pedicellata from Itatiaia are species with cambial variants and they should not be used in
dendrochronologic studies, because the rings are of sporadic occurrence. However, these
species taken from places with more drastic environmental change around year can make
distinct growth rings with frequent occurrence, showing that more studies with this species
from other ranges are required.

55
ACKNOWLEDGEMENTS
We thank the Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, Programa Mata
Atlântica and CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) for
financial support; the Laboratório de Botânica Estrutural do Instituto de Pesquisas Jardim
Botânico do Rio de Janeiro for logistic support; the Dra. Maura da Cunha by suggestions on
manuscript.
REFERENCES
Alves ES, Angyalossy-Alfonso V. 2000. Ecological trends in the wood anatomy of some
Brazilian species. 1. Growth rings and vessels. International Association of Wood Anatomists
Journal 21: 3-30.
Amobi CC. 1974. Periodicity of wood formation in twigs of some tropical trees in Nigeria.
Annals of Botany 38: 931-936.
Araque OZ, De Pernía NE, León WJ. 2000. Estudio anatómico del leño de seis especies de
lianas. Revista Forestal Venezolana 44: 39-48.
Baas P, Schweingruber FH. 1987. Ecological trends in the wood anatomy of trees, shrubs
and climbers from Europe. International Association of Wood Anatomists Bulletin 8: 245-
274.
Bamber RK, Ter Welle BJH. 1994. Adaptative trends in the wood anatomy of lianas. In:
Iqbal M, ed. Growth Patterns in Vascular Plants. Portland: Dioscorides Press, 272-287.
Barros CF, Callado CH. 1997. Madeiras da Mata Atlântica – Anatomia do lenho de
espécies ocorrentes nos remanescentes florestais do Estado do Rio de Janeiro. Vol. I. Brazil:
Jardim Botânico do Rio de Janeiro.
Barros CF, Callado CH, Marcon ML, Costa CG, Cunha M, Lima HRP, Marquete O.
2001. Madeiras da Mata Atlântica – Anatomia do lenho de espécies ocorrentes nos
remanescentes florestais do Estado do Rio de Janeiro. Vol. II. Brazil: Jardim Botânico do Rio
de Janeiro.
Boninsegna JA, Villalba R, Amarilla L, Ocampo J. 1989. Studies on tree rings, growth
rates and age-size relationships of tropical tree species in Missiones, Argentina. International
Association of Wood Anatomists Bulletin n.s. 10: 161-169.
Borchert R. 1999. Climatic periodicity, phenology and cambium activity in tropical dry
forest trees. International Association of Wood Anatomists Journal 20: 239-247.

56
Botosso PC. 1984. Some anatomical wood characteristics as source of cyclic structural
change (regular or irregular) of growth periodicity for 20 Amazonian species. International
Association of Wood Anatomists Bulletin n.s. 5: 545-546.
Bräker OU. 2002. Measuring and data processing in tree-ring research – a methodological
introduction. Dendrochronologia 20: 203-216.
Brienen RJW, Zuidema PA. 2006. The use of tree rings in tropical forest management:
Projecting timber yields of four Bolivian tree species. Forest Ecology and Management 226:
256-267.
Buckley BM, Barbetti M, Watanasak M, D’arrigo DR, Boonchirdchoo S, Sarutanon S.
1995. Dendrochronological investigations in Thailand. International Association of Wood
Anatomists Journal 16: 393-409.
Bukatsch F. 1972. Bemerkungen zur doppelfärbung astrablau-safranin. Mikrokosmos 61(8):
33-36.
Caballé G. 1993. Liana structure, function and selection: a comparative study of xylem
cylinders of tropical rainforest species in Africa and America. Botanical Journal of the
Linnean Society 113: 41-60.
Callado CH, Neto SJS, Scarano FR, Barros CF, Costa CG. 2001a. Anatomical features of
growth rings in flood-prone trees of the Atlantic Rain Forest in Rio de Janeiro, Brazil.
International Association of Wood Anatomists Journal 22: 29-42.
Callado CH, Neto SJS, Scarano FR, Costa CG. 2001b. Periodicity of growth rings in some
flood-prone trees of the Atlantic Rain Forest in Rio de Janeiro, Brazil. Trees 15: 492-497.
Callado CH, Neto SJS, Scarano FR, Costa CG. 2004. Radial growth dynamics of Tabebuia
umbellata (Bignoniaceae), a flood-tolerant trees from the Atlantic Forest swamps in Brazil.
International Association of Wood Anatomists Journal 25:175-183.
Carlquist S. 1991. Anatomy of vine and liana stems: a review and synthesis. In: Putz FE,
Mooney HA, eds. The Biology of Vines. Cambridge: Cambridge University Press, 53-72.
Carlquist S. 1985. Observations on functional wood histology of vines and lianas: vessel
dimorphism, tracheids, vasicentric tracheids, narrow vessels, and parenchyma. Aliso 11: 139-
157.
Carlquist S. 1995. Wood and Bark anatomy of Ranunculaceae (including Hydrastis) and
Glaucidiaceae. Aliso 14: 65-84.
Carlquist S. 2001. Comparative wood anatomy: systematic, ecological and evolutionary
aspects of dicotyledon wood. Berlin: Springer-Verlag, 2a ed.

57
Détienne P. 1989. Appearance and periodicity of growth ring in some tropical woods.
International Association of Wood Anatomists Bulletin 10: 123-132.
Devall MS, Parresol BR, Wright SJ. 1995. Dendroecological analysis of Cordia alliodora,
Pseudobombax septenatum and Annona spraguei in central Panama. International
Association of Wood Anatomists Journal 16: 411-424.
Eickstein D, Sass U, Baas P. 1995. Growth periodicity in tropical trees – Preface.
International Association of Wood Anatomists Journal 16: 325.
Eshete G, Stahl G. 1999. Tree rings as indicators of growth periodicity of acacias in the Rift
Valley of Ethiopia. Forest Ecology and Management 116: 107-117.
Ewers FW, Fisher JB. 1991. Why vines have narrow stems: histological trends in Bauhinia
(Fabaceae). Oecologia 88: 233-237.
Fahn A. 1982. El xilema secundario. In: Anatomia Vegetal. Madri: Ediciones Pirámide S.A.,
350-391.
Fahn A, Burley J, Longman KA, Mariaux A, Tomlinson PB. 1981. Possible contributions
of wood anatomy to the determination of the age of tropical trees. Yale University: School of
Forestry and Environmental Studies Bulletin 94: 31-54.
Gasson P. 1994. Wood anatomy of the Tribe Sophoreae and related Caesalpinoideae and
Papilionoideae. Advances in Legume Systematics 6: 165-203.
Gasson P. 2000. Does wood anatomy support tribal and generic classification in Papilionoid
Leguminosae. Advances in Legume Systematics 9: 201-215.
Gasson P, Dobbins DR. 1991. Wood anatomy of the Bignoniaceae, with a comparison of
trees and lianas. International Association of Wood Anatomists Bulletin n.s. 12(4): 389-417.
Gentry AH. 1991. The distribution and evolution of climbing plants. In: Putz FE, Mooney
HA, eds. The Biology of Vines. Cambridge: Cambridge University Press, 3-50
Gerwing JJ, Schnitzer SA, Burnham RJ, Bongers F, Chave J, Dewalt SJ, Ewango CEN,
Foster R, Kenfack D, Martínez-Ramos M, Parren M, Parthasarathy N, Pérez-Salicrup
DR, Putz FE, Thomas DW. 2006. A Standard Protocol for Liana Censuses. Biotropica 38:
256-261.
Gourlay ID. 1995a. Growth ring characteristics of some African Acacia species. Journal of
Tropical Ecology 11: 121-140.
Gourlay ID. 1995b. The definition of seasonal growth zones in some African Acacia species
– A review. International Association of Wood Anatomists Journal 16: 353-359.

58
Grau HR, Easdale AT, Paolini L. 2003. Subtropical dendroecology – dating disturbances
and forest dynamics in northwestern Argentina montane ecosystems. Forest Ecology and
Management 177: 131-143.
Heinrich I, Banks JCG. 2006. Tree-ring anomalies in Toona ciliata. International
Association of Wood Anatomists Journal 27: 213-231.
Höhn A. 1999. Wood anatomy of selected west african species of Caesalpinioideae and
Mimosoideae (Leguminosae): A comparative study. International Association of Wood
Anatomists Journal 20: 115-146.
Iawa Committee. 1989. List of microscopic feature of hardwood identification. International
Association of Wood Anatomists Bulletin n.s. 10: 219-332.
Jacoby GC. 1989. Overview of tree-ring analysis in tropical regions. International
Association of Wood Anatomists Bulletin n.s. 10: 99-108.
Leon-Gómez C, Monroy-Ata A. 2005. Seasonality in cambial activity of four lianas from a
Mexican lowland tropical rainforest. International Association of Wood Anatomists Journal
26: 111-120.
Marcati CR, Angyalossy V, Evert RF. 2006. Seasonal variation in wood formation of
Cedrela fissilis (Meliaceae). International Association of Wood Anatomists Journal 27: 199-
211.
Mariaux A. 1981. Past efforts in measuring age and annual growth in tropical trees. Yale
University: School of Forestry and Environmental Studies Bulletin 94: 20-30.
Morellato LPC, Haddad CFB. 2000. Introduction: The Brazilian Atlantic Forest. Biotropica
32: 786-792.
Myers N, Mittermeier RA, Mittermeier CG, Fonseca GAB, Kent J. 2000. Biodiversity
hotspots for conservation priorities. Nature 403: 853-858.
Oliveira-Filho AT, Fontes MAL. 2000. Patterns of floristic differentiation among Atlantic
Forests in Southeastern Brazil and the influence of climate. Biotropica 32: 793-810.
Pérez-Salicrup DR, Schnitzer S, Putz FE. 2004. Community ecology and management of
lianas. Forest Ecology and Management 190: 1-2.
Priya PB, Bhat KM. 1999. Influence of rainfall, irrigation and age on the growth periodicity
and wood structure in Teak (Tectona grandis). International Association of Wood Anatomists
Journal 20: 181-192.
Resende AA, Ranga NT. 2005. Lianas da Estação Ecológica do Nordeste Paulista, São José
do Rio Preto / Mirassol, SP, Brasil. Acta Botanica Brasilica 19: 273-279.

59
Richter HG, Dallwitz MJ. 2000 onwards. Commercial timbers: descriptions, illustrations,
identification, and information retrieval. In English, French, German, Portuguese, and
Spanish. Version: 16th April 2006. http://delta-intkey.com.
Roig FA, Osornio JJJ, Diaz JV, Luckman B, Tiessen H, Medina A, Noellemeyer EJ.
2005. Anatomy of growth rings at the Yucatán Peninsula. Dendrochronologia 22: 187-193.
Sass U, Eckstein D. 1995. The variability of vessel size in beech (Fagus sylvatica L.) and its
ecophysical interpretation. Trees 9: 247-252.
Scarano FR. 2002. Structure, Function and Floristic Relationship of Plant Communities in
Stressful Habitats Marginal to the Brazilian Atlantic Rainforest. Annals of Botany 90: 517-
524.
Schnitzer SA, Bongers F. 2002. The ecology of lianas and their role in forests. Trends in
Ecology and Evolution 17: 223-230.
Schweingruber FH, Poschlod P. 2005. Growth rings in herbs and shrubs: life span, age
determination and stem anatomy. Forest Snow and Landscape Research 79: 195-415.
Segadas-Vianna F, Dau L. 1965. Ecology of the Itatiaia Range, Southeastern Brazil. II.
Climates and climatic altitudinal zonation. Arquivos do Museu Nacional 53: 31-53.
Seigler DS, Ebinger JE , Miller JT. 2006. The genus Senegalia (Fabaceae: Mimosoideae)
from the New World. Phytologia 88(1): 38-94.
Seitz RS, Kanninen M. 1989. Tree ring analysis of Araucaria angustifolia in southern
Brazil: Preliminary results. International Association of Wood Anatomists Bulletin n.s. 10:
170-174.
Tomazello Filho M, Lisi CS, Hansen N, Cury G. 2004. Anatomical features of increment
zones in different tree species in the State of São Paulo, Brazil. Scientia Forestalis 66: 46-55.
Tomlinson PB, Longman KA. 1981. Growth Phenology of Tropical Trees in Relation to
Cambial Activity. Yale University: School of Forestry and Environmental Studies Bulletin 94:
7-19.
Vetter RE, Botosso PC. 1989. Remarks on age and growth rate determination of Amazonian
trees. International Association of Wood Anatomists Bulletin n.s. 10: 133-145.
Villalba R, Boninsegna JA. 1989. Dendrochronological studies on Prosopis flexuosa DC.
International Association of Wood Anatomists Bulletin n.s. 10: 155-160.
Villalba R, Boninsegna JA, Holmes RL. 1985. Cedrela angustifolia and Juglans australis:
Two new tropical species useful in dendrochronology. Tree-ring Bulletin 45: 25-35.
Westbrook JA, Guilserson ThP, Colinvaux PA. 2006. Annual growth rings in a sample of
Hymenaea courbaril. International Association of Wood Anatomists Journal 27: 193-197.

60
Wheeler E, Baas P. 1992. Fossil wood of the Leguminosae: A case study in xylem evolution
and ecological anatomy. Advances in Legume Systematics 4: 281-301.
Wimmer R. 2002. Wood anatomical features in tree-rings as indicators of environmental
change. Dendrochronologia 20: 21-36.
Worbes M. 1985. Structural and other adaptation to long-term flooding by trees in Central
Amazonia. Amazoniana 9: 459-484.
Worbes M. 1989. Growth rings, increment and age of trees in inundation forests, savannas
and a mountain forest in the neotropics. International Association of Wood Anatomists
Bulletin n.s. 10: 109-122.
Worbes M. 1995. How to measure growth dynamics in tropical trees – A review.
International Association of Wood Anatomists Journal 16: 337-351.
Worbes M. 2002. One hundred year of tree-ring research in the tropics – a brief history and
an outlook to future challenges. Dendrochronologia 20: 217-231.
Worbes M, Staschel R, Roloff A, Junk WJ. 2003. Tree ring analysis reveals age structure,
dynamics and wood production of a natural forest stand in Cameroon. Forest ecology and
management 173: 105-123.
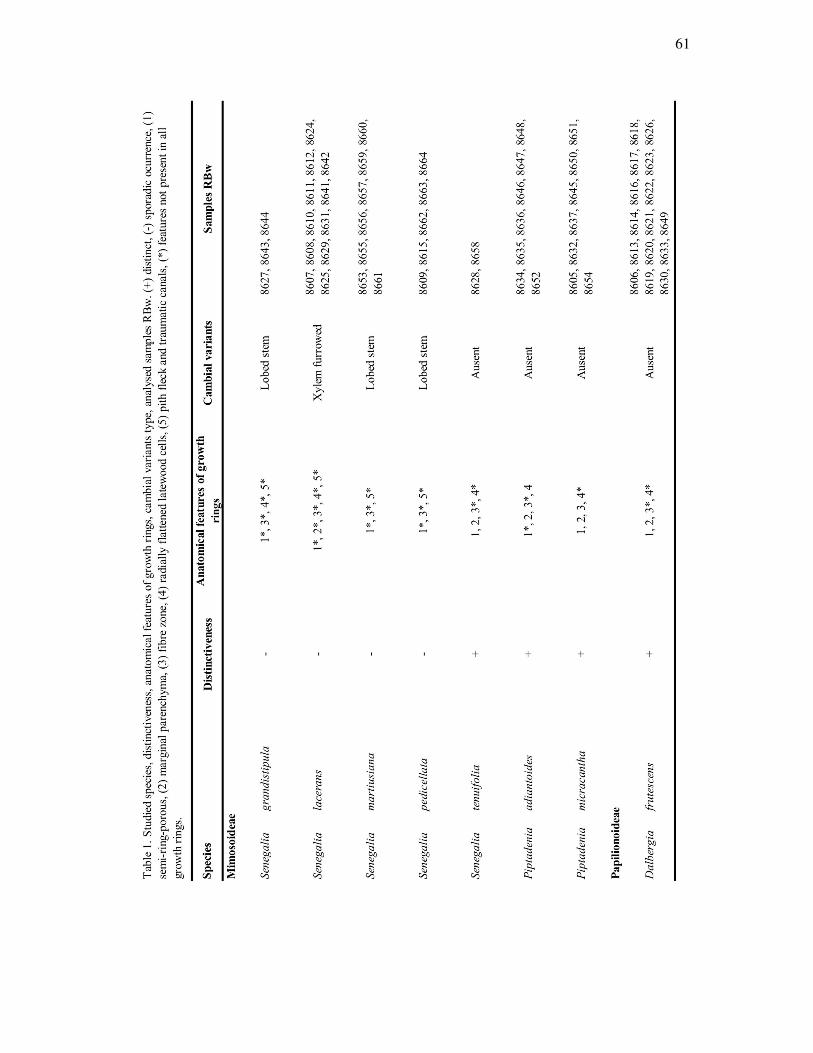
61

62
Figure 1: Transverse sections in light microscope. A: Senegalia grandistipula. B: S.
pedicellata. C: S. lacerans. D: S. martiusiana. Arrows indicate the growth ring boundaries.
Scale bar = 100µm.

63
64
Figure 2: Transverse sections in stereomicroscope. A: Senegalia grandistipula. B: S. lacerans.
C: S. martiusiana. D: S. pedicellata. Arrows indicate the growth ring boundaries. Scale bar =
1 mm.

65
Figure 3: Transverse sections in light microscope. A: Dalbergia frutescens. B: Piptadenia
adiantoides. C: P. micracantha. D: Senegalia tenuifolia. Arrows indicate the growth ring
boundaries. Scale bar = 100 µm.

66

67
Figure 4: Transverse sections in stereomicroscope. A: Senegalia tenuifolia. B: Piptadenia
adiantoides. C: P. micracantha. D: Dalbergia frutescens. Arrows indicate the growth ring
boundaries. Scale bar = 1mm.
Figure 5: Prismatic crystals of calcium oxalate on polarized light microscopy in Piptadenia
micracantha. Scale bar = 100 µm.

68
ARTIGO 3
Crescimento secundário e idade em lianas da família
Leguminosae na Mata Atlântica: um estudo com camadas
de crescimento
Arno Fritz das Neves Brandes e Cláudia Franca Barros
Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, Programa Mata Atlântica, Rua
Pacheco Leão 915, Jardim Botânico, CEP 22460-030, Rio de Janeiro, Rio de Janeiro, Brasil.
Artigo a ser traduzido para o inglês e submetido a “Trees: Structure and Functions”.

69
RESUMO
O crescimento e a idade das plantas são informações importantes para o entendimento dos
complexos processos florestais. As lianas desempenham um papel muito importante nas
florestas tropicais e pouco se sabe sobre idade e crescimento deste hábito. No presente estudo
foram analisadas as camadas de crescimento de quatro espécies de lianas da família
Leguminosae, que se desenvolvem no Parque Nacional do Itatiaia, localizado no Sudeste do
Brasil, região compreendida no Bioma Mata Atlântica e que apresenta um relevo montanhoso
com uma estação seca bem definida no ano. Foi detectada periodicidade anual de formação
das camadas de crescimento nas quatro espécies. Através da análise das séries temporais
notou-se que ocorre grande semelhança entre os raios do mesmo indivíduo, contudo pequena
semelhança entre os indivíduos e entre as espécies. A análise climatológica das séries
temporais mostrou que poucos indivíduos apresentam correlação significativa com
precipitação e temperatura. Foi detectada a idade máxima de 27 anos e taxa média de
crescimento radial entre as espécies de 2,3 mm por ano. Por meio do incremento radial
acumulado nota-se uma forte relação idade-diâmetro nas lianas, contudo o desvio do
incremento acumulado médio para a espécie aumenta junto com a idade. A análise das
camadas de crescimento, neste estudo, mostrou-se um método útil na estimativa de
parâmetros, como o crescimento e idade, que podem ser utilizados em futuros estudos de
ecologia florestal.

70
INTRODUÇÃO
Desde o final do século 19 são conhecidos trabalhos na região tropical, que
comprovam a periodicidade anual de crescimento secundário e aplicam métodos
dendrocronológicos em espécies arbóreas (Worbes, 2002). Na Mata Atlântica as pesquisas são
mais recentes, mas também foram observadas espécies que expressam a periodicidade anual
de crescimento e ressaltam o potencial para se desenvolver este tipo de pesquisa neste bioma
(Callado et al., 2001).
A análise das camadas de crescimento tem-se mostrado um dos melhores métodos
para estimar a idade e o crescimento de espécies arbóreas (Worbes & Junk, 1999). Através da
largura das camadas de crescimento é possível extrair informações sobre o crescimento, idade
e relações com as condições ambientais (Worbes, 2002; Brienem e Zuidema, 2006).
As lianas representam aproximadamente 25% da riqueza e da abundância nas florestas
tropicais (Schnitzer & Bongers, 2002). De forma discrepante podem contribuir com 5% da
biomassa total acima do solo e com até 40% da área foliar da floresta (Putz, 2006). Elas
competem com as espécies arbóreas por nutrientes, luz e água (Gentry, 1991). Devido a seus
caules finos e o lenho de pouca densidade, as lianas seqüestram menos carbono que as
espécies arbóreas. Infestações de lianas inibem a regeneração da floresta por árvores o que
reduz a quantidade de carbono que seria assimilada na biomassa (Schnitzer & Bongers, 2002).
Conhecem-se dois estudos que aplicam a análise das camadas de crescimento em
lianas. Um realizado no início do século 20, que estima a idade de Clematis vitalba L.
(Kanngiesser, 1906 apud Schweingruber & Poschlod, 2005) e outro recente que compara o
crescimento de uma espécie que se apresenta como lianas e auto-suportantes (Ceccantini &
Brandes, dados não publicados). Um estudo desenvolvido por Brandes & Barros (dados não
publicados), detalha a anatomia das camadas de crescimento de quatro espécies de lianas da
família Leguminosae, Senegalia tenuifolia (L.) Britton & Rose, Piptadenia adiantoides
(Spreng.) J.F. Macbr., Piptadenia micracantha Benth. e Dalbergia frutescens (Vell.) Britton
var. frutescens, que serão as lianas tratadas no presente estudo. Alguns outros relatos da
ocorrência de camadas de crescimento distintas em lianas são reportados em Baas &
Schweingruber (1987), Gasson & Dobbins (1991), Carlquist (1995), Araque et al. (2000) e
demonstram que a dendrocronologia neste hábito é um campo de pesquisas a ser explorado.
Em plantas herbáceas existem relatos antigos do uso da análise das camadas de crescimento,
mas foram negligenciados até recentemente serem redescobertos e mostrarem as diversas
aplicações nos estudos de ecologia e de biologia da conservação (Schweingruber & Poschlod,

71
2005). Partindo do princípio que as angiospermas com seus diversos hábitos, lianas, árvores,
arbustos e ervas, compartilham os mesmos mecanismos de crescimento secundário, o mesmo
pode-se esperar no estudo dendrocronológico das lianas.
Este estudo provem informações a respeito da idade e crescimento de lianas e algumas
das perguntas a serem respondidas são: Existe periodicidade de crescimento em lianas da
Mata Atlântica? Qual a taxa de crescimento das espécies analisadas? Há concordância entre as
séries temporais de um mesmo indivíduo, entre indivíduos de uma mesma espécie e entre
espécies? Ocorrem correlações entre as séries temporais e os dados climáticos?

72
MATERIAIS E MÉTODOS
O estudo foi realizado no Parque Nacional do Itatiaia, município de Itatiaia, no estado
do Rio de Janeiro, sudeste do Brasil (22°15’S - 22°30’ S, 44°30’W - 44°45’ W), com área de
28.155,97 hectares (http://www.ibama.gov.br). O tipo da vegetação é Floresta Atlântica senso
stricto (Oliveira-Filho & Fontes, 2000). O Parque Nacional do Itatiaia apresenta a
predominância de relevo montanhoso com bem delimitados cinturões climáticos e de
vegetação relacionados ao gradiente altitudinal (Segadas-Vianna & Dau 1965). As amostras
foram coletadas no cinturão compreendido entre 700 m e 1100 m. Esta área apresenta
precipitação anual média de 1699 mm e temperatura anual média de 18,2 °C, com uma
estação fria e seca de Junho a Agosto, onde chove somente 5 % da precipitação anual. O
clima é temperado úmido e pelo sistema Köppen do tipo Cfb (Segadas-Vianna & Dau 1965)
(Fig. 1).
Foram selecionadas para o estudo 31 lianas de quatro espécies da família Leguminosae
(Tabela 1). Os indivíduos selecionados para o estudo não apresentavam anomalias ou
deformidades aparentes na região da coleta e o diâmetro era maior que 2 cm a 1,3 m da base.
Foi utilizada a técnica de lesão no câmbio vascular, chamada “Janela de Mariaux”
(Mariaux, 1967), para detectar as características do xilema produzido durante o ano e a
presença de camadas de crescimento anuais. Foram realizadas quatro marcações ao longo do
ano e após um ano da primeira lesão foi coletada uma secção parcial do lenho para análise.
Durante o procedimento foi removido o câmbio de uma área de 0,5 cm de largura por
aproximadamente 5 cm de altura, onde se aplicou calda bordalesa para evitar a ação de
microorganismos patogênicos. As amostras utilizadas para a contagem e medida das camadas
de crescimento foram retiradas 5 cm acima da região onde foi realizada a marca no câmbio.
Todas as amostras deste estudo foram polidas com o auxílio de navalha, observadas
com lupa de mão (10x) e ao microscópio estereoscópico Olympus SZ11. Foram capturadas
imagens com uma câmera fotográfica Olympus C5050 acoplada ao microscópio citado. As
imagens foram processadas e a largura das camadas de crescimento foi medida com o
software Image-pro plus com precisão de 0,01 mm.
Quatro raios (séries temporais) foram medidos por amostra e estes valores combinados
constituíram uma curva média para o indivíduo. Antes das curvas serem combinadas, foi
realizada a datação cruzada (cross-dating) das séries temporais, o que permite a identificação
de anéis descontínuos (Lorimer et al. 1999). As séries também foram padronizadas
(standardized), ou seja, convertidas a índices pelo traçado da curva spline e pelo modelo

73
autoregressivo, removendo tendências de baixa freqüência. Para o tratamento e análise dos
dados foram seguidos passos metodológicos recomendados por Bräker (2002). As medidas
foram submetidas ao programa Cofecha (Holmes, 1983), para testar a qualidade das medidas
e detectar possíveis falhas na datação cruzada.
A média de incremento radial anual (taxa de incremento) para cada liana, foi calculada
através da média da largura dos anéis ao longo de quatro raios. O incremento médio anual em
diâmetro foi obtido multiplicando este valor por dois. Para cada liana foram estabelecidas as
relações idade-diâmetro ao longo de sua vida, através do incremento radial acumulado.
Todo o processamento das amostras foi realizado no Laboratório de Botânica
Estrutural do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro e as amostras foram
depositadas na Xiloteca do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro (RBw).
Os dados climáticos utilizados neste estudo foram coletados e cedidos pelo Instituto Nacional
de Meteorologia - 6° Distrito de Meteorologia na estação de Resende (22° 27' S, 44° 26' W).

74
RESULTADOS
As quatro espécies estudadas apresentam periodicidade no crescimento radial com a
formação de uma camada de crescimento por ano. Ao início da estação chuvosa, em
setembro, o câmbio produz vasos grandes, que vão gradativamente diminuindo a medida que
a estação seca se aproxima (Fig. 2 a 5). Não foi possível detectar o tipo de parênquima
marginal. Em nenhum indivíduo ocorreu a formação de mais de uma camada por ano.
A idade das lianas coletadas variou de 5 a 27 anos. Os indivíduos que apresentaram
maior idade são da espécie D. frutescens, chegando à 27 anos. Em P. micracantha o indivíduo
mais velho apresentou 19 anos. A média de crescimento radial por ano (taxa de crescimento)
foi maior nas espécies S. tenuifolia (2,76 mm/ano) e P. adiantoides (2,44 mm/ano), e um
pouco menor em P. micracantha (2,07 mm/ano) e D. frutescens (1,91 mm/ano) (Tabela 1,
Fig. 8). Entre todas as espécies estudadas, a média de crescimento foi 2,30 mm por ano. A
taxa de crescimento máxima observada entre indivíduos de S. tenuifolia foi 2,89 mm/ano, em
P. adiantoides 3,15 mm/ano, em P. micracantha 2,85 mm/ano e em D. frutescens 3,26
mm/ano.
Foi estimada a idade utilizando a média de crescimento radial de cada espécie para
caules com 5 cm de diâmetro (25 mm de raio) e observaram-se os seguintes valores: S.
tenuifolia com 9 anos, P adiantoides com 10 anos, P. micracantha com 12 anos e D.
frutescens com 13 anos. A mesma estimativa, quando realizada com a taxa de crescimento
máxima de cada espécie, obtém-se S. tenuifolia com 8 anos, P adiantoides com 7 anos, P.
micracantha com 8 anos e D. frutescens com 7 anos. Contudo a idade obtida através da média
do incremento acumulado de cada espécie, que reflete as medidas realizadas das camadas de
crescimento dos indivíduos, foi 10 anos em S. tenuifolia, 8 anos em P. adiantoides, 12 anos
em P. micracantha e 13 anos em D. frutescens (Fig. 8). Pôde-se também constatar que o
caule, com 2,5 cm de diâmetro (12,5 mm de raio), apresentou em média 6 anos nas espécies S.
tenuifolia, P. micracantha e D. frutescens e 5 anos em P. adiantoides. (Fig. 8).
A ocorrência de anéis descontínuos e de anéis muito estreitos foi notada em algumas
regiões do caule nas lianas deste estudo e foram mais freqüentes em D. frutescens, onde
somente um indivíduo não apresentou camadas descontínuas.
Foi observada alta intercorrelação entre os raios medidos em cada indivíduo e os
valores mais altos foram de P. adiantoides e D. frutescens (Fig. 6). Foram excluídas das
análises de correlação subseqüentes as séries temporais de indivíduos que apresentaram baixa
correlação entre os raios (coeficiente de intercorrelação < 0,4).

75
A intercorrelação das séries temporais entre indivíduos da mesma espécie foi baixa. A
espécie cujos indivíduos apresentaram a maior intercorrelação foi P. adiantoides.
Com valores de correlação baixos, não é recomendável traçar uma curva média de
cada espécie e testar a sua relação com fatores climáticos. Então, foi realizada a análise de
correlação entre os indivíduos e os fatores climáticos. Poucos indivíduos mostraram
correlação positiva dos índices com fatores climáticos. Senegalia tenuifolia e P. micracantha
não apresentaram nenhuma correlação significativa. Em P. adiantoides um indivíduo
apresentou correlação com precipitação total anual e dois com temperatura média anual. Em
D. frutescens foi observada correlação significativa somente em dois indivíduos com
temperatura (Fig. 7).
A correlação entre o crescimento radial acumulado e idade foi significativa para todas
as espécies. A média de incremento radial acumulado para cada espécie foi calculada (Fig. 8).
Foram observadas diferenças nas trajetórias de crescimento das espécies e entres os
indivíduos de uma mesma espécie (Fig. 8 e 9). As maiores variações na trajetória de
crescimento foram observadas em D. frutescens e P. micracantha (Fig. 9). O desvio padrão
do incremento radial acumulado médio de cada espécie aumenta com o aumento da idade
(Fig. 9).

76
DISCUSSÃO
Foi observado neste estudo a ocorrência de camadas de crescimento anuais. Tal
constatação é reforçada pelo regime anual de precipitação na área de estudo, com uma estação
seca com duração de três meses, onde a média mensal é menor que 30 mm. Segundo Worbes
(1995), regiões com um período seco de três meses ou mais no ano e precipitação abaixo de
60 mm mensais neste período, favorecem a ocorrência de espécies que formam camadas de
crescimento anuais. A ocorrência de camadas de crescimento anuais também foram relatadas
em espécies arbóreas da Mata Atlântica (Callado et al., 2001) e em diversas espécies do
gênero Acacia na região tropical na África com um período seco anual (Gourlay, 1995). No
presente estudo, tratando-se de outro hábito que não o arbóreo, foi utilizada a técnica de
lesões no câmbio para demonstrar a periodicidade do crescimento secundário além das
evidências climatológicas e anatômicas.
O crescimento em diâmetro observado nas lianas no presente trabalho foi bem superior
aos observados em outros estudos (Nabe-Nielsen, 2002; Putz,1990; Gerwing, 2004; Ewers et
al., 1991). Observou-se um incremento médio de diâmetro de 3,82 mm a 5,51 mm por ano, ou
1,91 mm a 2,76 mm de incremento radial. Em lianas da espécie Machaerium cuspidatum
(Leguminosae), estudadas por Nabe-Nielsen (2002), foi observada a taxa de crescimento no
diâmetro de 1,4 mm por ano em indivíduos com caule de 31 mm a 50 mm de diâmetro, e em
indivíduos com diâmetro de 51 mm a 100 mm foi 0,7 mm por ano. As maiores taxas de
crescimento observadas por ele foram em lianas que se desenvolviam sob grande quantidade
de luz. Entretanto poucas cresceram rapidamente quando estavam totalmente expostas ao sol.
O valor médio de 1,4 mm por ano de crescimento também foi observado por Putz (1990),
estudando 15 espécies de lianas no Panamá, as que apresentavam maior taxa de crescimento
pertenciam à família Leguminosae. Segundo o autor, Acacia hayessi (Britt. & Rose) Standl.
apresentou incremento médio anual no diâmetro de 2,58 mm e Entada monstachya DC. de
5,82 mm.
Gerwing (2004) registrou o crescimento anual variando de 0,3 mm a 2,2 mm por ano
em seis espécies de lianas da Amazônia brasileira. Algumas das espécies por ele estudadas
que pertenciam a família Leguminosae: Bauhinia guianensis, com incremento de 1,1 mm por
ano, Bauhinia cupreonitans com 1,8 mm por ano e Acacia multipinnata com 1,9 mm por ano.
Segundo o autor, as espécies com desenvolvimento mais lento estavam associadas a florestas
em estagio sucessional tardio ou maduras e as que se desenvolveram mais rapidamente,
associadas à abertura de clareiras e a florestas em recente estágio sucessional. Ewers et al.

77
(1991) refere dados de Lieberman et al., coletados na Costa Rica, que demonstram que o
aumento anual no diâmetro das espécies varia de 0,0 a 5,2 mm, contudo algumas espécies
foram pouco representadas. Machaerium seemannii apresentou crescimento de com 5,2 mm
por ano e Bauhinia sp. de 4,3 mm por ano, valores próximos aos obtidos no presente trabalho.
Os resultados destes estudos apresentados acima somados aos obtidos no presente,
demonstram que as lianas da família Leguminosae têm o crescimento rápido quando
comparado a lianas de outras famílias.
Foi observado nos indivíduos estudados que existe relação entre diâmetro e idade.
Notam-se variações na taxa de crescimento entre os indivíduos, mas geralmente apresentam
uma relação próxima a linear, como também observou Bonisegna et al. (1989) em Cedrella
fissilis na Argentina e Worbes (1999) em espécies arbóreas tropicais, que em situações
naturais tendem a apresentar um crescimento contínuo. Esta relação mais ou menos linear
permite uma simples estimação da idade através do diâmetro. A estimativa da idade deve ser
assumida com muita cautela visto o desvio de uma tendência linear de crescimento. Gerwing
(2004) estimou a idade de lianas através da taxa de crescimento médio e máximo de algumas
espécies de lianas e encontrou valores discrepantes para lianas com 5 cm de diâmetro. Em
Cronton ascendens estimou 25 anos através da média e 7 anos através do máximo. Em
Memora schomburgkii estimou 169 anos através da média e 30 anos através do máximo. Nas
espécies estudadas no presente trabalho, quando estimada a idade de caules com 5 cm de
diâmetro pela média de crescimento, S. tenuifolia apresentou 9 anos, P adiantoides 10 anos,
P. micracantha 12 anos e D. frutescens 13 anos. A mesma estimativa, quando realizada com o
crescimento máximo foi 8 anos em S. tenuifolia e P. micracantha e 7 anos em P adiantoides e
D. frutescens. Os valores estimados pela taxa de crescimento médio foram bem próximos aos
valores observados através da contagem e mensuração. Contudo quando estimado pelo
crescimento máximo estes valores foram subestimados. Isso demonstra que o crescimento
médio seria o melhor parâmetro para estimação da idade em lianas.
Poucos estudos estimaram a idade máxima de algumas lianas. Kanngiesser (1906 apud
Schweingruber & Poschlod, 2005) estimou 41 anos em Clematis vitalba L. e utilizou as
camadas de crescimento para este fim. Burkill (1944 apud Schweingruber & Poschlod, 2005)
estima a idade máxima de Tamus communis L. em 20 anos, porém o método de estimação por
ele utilizado é desconhecido. A idade máxima observada em lianas deste estudo foi 27 anos
em D. frutescens. É importante salientar que as idades obtidas são subestimadas devido ao
tempo que o indivíduo levou para desenvolver-se de plântula até a altura de onde foi removida
a amostra.

78
No presente trabalho, quando analisadas as séries temporais (raios) de cada indivíduo,
foram observados altos valores de intercorrelação, demonstrando que em diferentes regiões do
caule o câmbio responde proporcionalmente da mesma maneira aos fatores que influenciam
no crescimento. Contudo a baixa correlação entre as séries temporais dos indivíduos de uma
mesma espécie sugere que as lianas podem estar respondendo mais a distúrbios locais. As
análises climatológicas das séries temporais reforça tal suposição, pois poucos indivíduos
apresentaram correlações significativas com precipitação e temperatura.
Anéis ocultos e descontínuos são um problema conhecido e pode levar a subestimação
da idade e superestimação do crescimento (Lorimer et al. 1999). Neste estudo foram
detectadas camadas de crescimento descontínuas em todas as espécies através da datação
cruzada das séries temporais do mesmo indivíduo. A presença de anéis descontínuos é
relatada em diversas espécies arbóreas neotropicais, e se observa que são mais comuns em
árvores pequenas do sub-bosque do que em árvores do dossel ou emergentes. Nota-se que os
indivíduos que mais apresentam anéis descontínuos estão sob pequena quantidade de luz e
desenvolvem-se sob competição (Worbes, 2002; Lorimer et al. 1999). A presença de anéis
descontínuos possivelmente deve-se a diferenças no suprimento local de carboidratos, água,
elementos minerais e fitohormônios, geradas por mudanças na saturação de luz decorrentes de
modificações da pressão de competição (Dünisch et al., 1999 apud Worbes, 2002). Estudos
da ecologia das lianas relatam a forte pressão de competição com árvores por nutrientes, luz e
por água em ambientes onde ocorre uma estação seca (Schnitzer & Bongers, 2002). Os
resultados obtidos neste estudo, quanto à presença de camadas descontínuas, reforçam a idéia
de que as lianas estão sujeitas a processos de competição que as levam a alterações da
atividade cambial em determinadas regiões do caule.
As lianas da família Leguminosae em Itatiaia apresentam: camadas de crescimento
anuais; têm a taxa de crescimento maior que as observadas em outros estudos; mostram
grande correlação das séries temporais de um mesmo indivíduo e pequena entre indivíduos e
espécies; têm poucos indivíduos com correlação com fatores climáticos; e possuem relação
idade diâmetro, mas o desvio do incremento radial acumulado médio aumenta junto com o a
idade. A análise das camadas de crescimento das lianas mostrou-se um método útil na
estimação de parâmetros, como o crescimento e idade, que podem ser utilizados em futuros
estudos de ecologia florestal. Contudo são necessárias mais investigações, com mais espécies,
para se ter um panorama mais preciso sobre idade e crescimento deste hábito.

79
REFERÊNCIAS BIBLIOGRÁFICAS
Araque OZ, De Pernía NE, León WJ. 2000. Estudio anatómico del leño de seis especies de
lianas. Revista Forestal Venezolana 44: 39-48.
Baas P, Schweingruber FH. 1987. Ecological trends in the wood anatomy of trees, shrubs and
climbers from Europe. International Association of Wood Anatomists Bulletin 8: 245-
274.
Boninsegna, J.A., Villalba, R., Amarilla, L., e Ocampo, J., 1989. Studies on tree rings, growth
rates and age-size relationships of tropical tree species in Missiones, Argentina. IAWA
Bulletin n.s., vol. 10 (2) : 161-169.
Bräker, O.U. 2002. Measuring and data processing in tree-ring research – a methodological
introduction. Dendrochronologia 20 / 1-2 : 203-216.
Brienen, R.J.W. & Zuidema, P.A. 2006. Lifetime growth patterns and ages of Bolivian rain
forest trees obtained by tree ring analysis. Journal of Ecology 94: 481-493.
Callado, C.H., Neto, S.J.S., Scarano, F.R. & Costa, C.G. 2001. Periodicity of growth rings in
some flood-prone trees of the Atlantic Rain Forest in Rio de Janeiro, Brazil. Trees , vol
15:492–497.
Carlquist S. 1995. Wood and Bark anatomy of Ranunculaceae (including Hydrastis) and
Glaucidiaceae. Aliso 14: 65-84.
Ewers, F.W., Fisher, J.B. & Fichtner, K. 1991. Water flux and xylem structure in vines. In:
Putz, F.E., Mooney, H.A. (Eds), The Biology of Vines. Cambridge University Press,
Cambridge, United Kingdom, pp. 127-160.
Gasson, P. & Dobbins, D.R. 1991. Wood anatomy of the Bignoniaceae, with a comparison of
trees and lianas. IAWA Bulletin 12 (4): 389-417.
Gentry, A.H. 1991. The distribution and evolution of climbing plants. In: Putz, F.E., Mooney,
H.A. (Eds), The Biology of Vines. Cambridge University Press, Cambridge, United
Kingdom, pp. 3-50.
Gerwing, J.J 2004. Life history diversity among six species of canopy lians in an old-growth
forest of the eastern Brazilian Amazon. Forest Ecology and Management 190 : 57-72.
Gourlay, I.D. 1995. Growth ring characteristics of some African Acacia species. Journal of
Tropical Ecology, 11 : 121-140.
Holmes, R.L. 1983. Computer-assisted quality control in tree-ring dating and measurement.
Tree-Ring Bulletin 43: 69-78.

80
Lorimer, C.G., Dahir, S.E. & Singer, M.T. 1999. Frequency of partial and missing rings in
Acer saccharum in relation to canopy position and growth rate. Plant ecology 143 :
189-202.
Mariaux, A. 1967. Les cernes dans les bois tropicaux africains, nature et periodicité. Rev.
Bois For. Trop., 113: 3-14/114 : 23-37.
Nabe-Nielsen, L. 2002. Growth and Mortality Rates of the Liana Machaerium cuspidatum in
Relation to Light and Topographic Position. Biotropica 34(2) : 319 – 322.
Oliveira-Filho, A.T. & Fontes, M.A.L. 2000. Patterns of Floristic Differentiation among
Atlantic Forests in Southeastern Brazil and the influence of Climate. Biotropica 32(4b)
: 793-810
Putz, F.E. 1990. Liana stem diameter growth and mortality rates on Barro Colorado Island,
Panama. Biotropica 22 : 103 – 105.
Putz, F.E. 2006. Vine ecology. ECOLOGY.INFO #24.
Schnitzer, S.A. & Bongers, F. 2002. The ecology of lianas and their role in forests. Trends in
Ecology and Evolution 17 (5): 223-230.
Schweingruber, F.H., Poschlod, P. 2005. Growth rings in herbs and shrubs: life span, age
determination and stem anatomy. Forest Snow and Landscape Research 79: 195-415.
Segadas-Vianna, F. & Dau, L. 1965. Ecology of the Itatiaia Range, Southeastern Brazil.
Arquivos do Museu Nacional, vol.LIII : 31-53.
Worbes, M., 1995. How to measure growth dynamics in tropical trees – A review. IAWA
Journal, Vol. 16 (4) : 337-351
Worbes, M., 1999. Annual growth rings, rainfall-dependent growth and long-term growth
patterns of tropical trees from the Caparo Forest Reserve in Venezuela. Journal of
Ecology, 87: 391-403.
Worbes, M., 2002. One hundred year of tree-ring research in the tropics – a brief history and
an outlook to future challenges. Dendrochronologia, 20/1-2 : 217-231.
Worbes, M. & Junk, W.J. 1999. How old are tropical trees? The persistence of a myth. IAWA
Journal 20 (3) : 255-260.
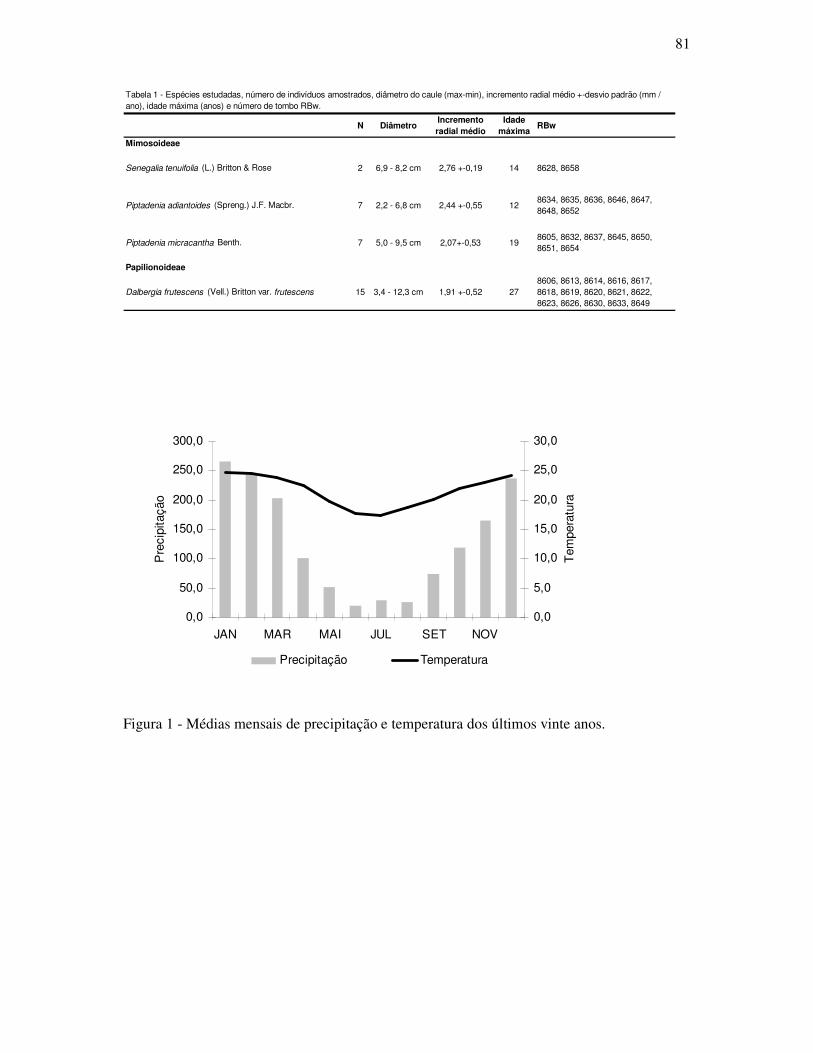
81
Figura 1 - Médias mensais de precipitação e temperatura dos últimos vinte anos.
0,0
50,0
100,0
150,0
200,0
250,0
300,0
JAN MAR MAI JUL SET NOV
Pre
cipi
taçã
o
0,0
5,0
10,0
15,0
20,0
25,0
30,0
Tem
pera
tura
Precipitação Temperatura
N DiâmetroIncremento radial médio
Idade máxima
RBw
Mimosoideae
Senegalia tenuifolia (L.) Britton & Rose 2 6,9 - 8,2 cm 2,76 +-0,19 14 8628, 8658
Piptadenia adiantoides (Spreng.) J.F. Macbr. 7 2,2 - 6,8 cm 2,44 +-0,55 128634, 8635, 8636, 8646, 8647, 8648, 8652
Piptadenia micracantha Benth. 7 5,0 - 9,5 cm 2,07+-0,53 198605, 8632, 8637, 8645, 8650, 8651, 8654
Papilionoideae
Dalbergia frutescens (Vell.) Britton var. frutescens 15 3,4 - 12,3 cm 1,91 +-0,52 278606, 8613, 8614, 8616, 8617, 8618, 8619, 8620, 8621, 8622, 8623, 8626, 8630, 8633, 8649
Tabela 1 - Espécies estudadas, número de indivíduos amostrados, diâmetro do caule (max-min), incremento radial médio +-desvio padrão (mm / ano), idade máxima (anos) e número de tombo RBw.
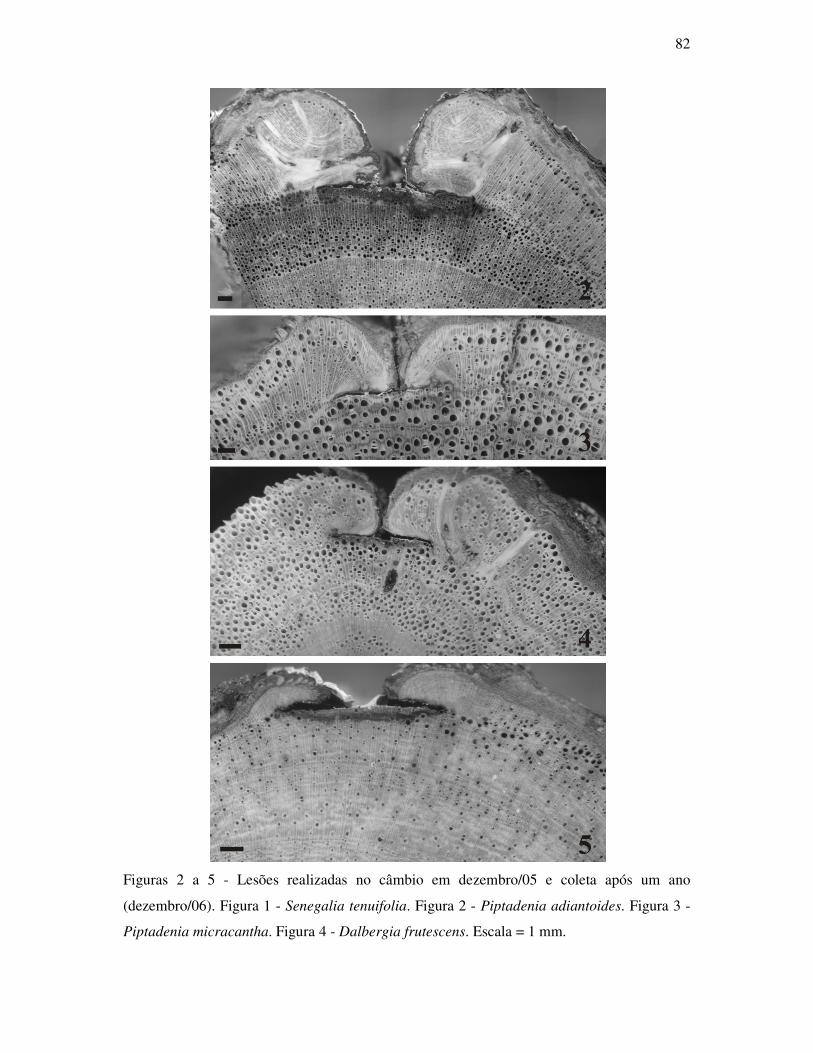
82
Figuras 2 a 5 - Lesões realizadas no câmbio em dezembro/05 e coleta após um ano
(dezembro/06). Figura 1 - Senegalia tenuifolia. Figura 2 - Piptadenia adiantoides. Figura 3 -
Piptadenia micracantha. Figura 4 - Dalbergia frutescens. Escala = 1 mm.
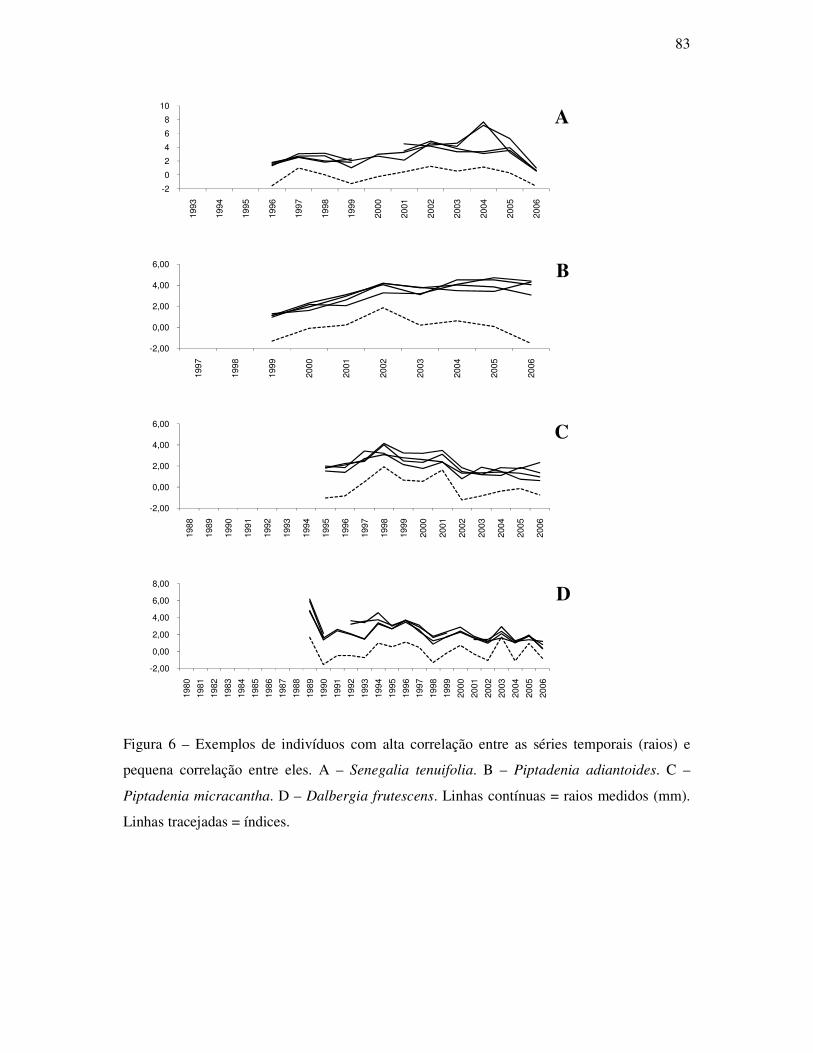
83
Figura 6 – Exemplos de indivíduos com alta correlação entre as séries temporais (raios) e
pequena correlação entre eles. A – Senegalia tenuifolia. B – Piptadenia adiantoides. C –
Piptadenia micracantha. D – Dalbergia frutescens. Linhas contínuas = raios medidos (mm).
Linhas tracejadas = índices.
-2,00
0,00
2,00
4,00
6,00
8,00
1980
1981
1982
1983
1984
1985
1986
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
D
-2
0
2
4
6
8
10
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
A
-2,00
0,00
2,00
4,00
6,00
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
B
-2,00
0,00
2,00
4,00
6,00
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
C

84
Figura 7 – Gráficos dos dados climáticos e índices. São representados apenas os indivíduos
que apresentaram alta correlação dos índices com os dados climáticos. A - Piptadenia
adiantoides e precipitação total anual (mm). B - Piptadenia adiantoides e temperatura média
anual (°C). C - Dalbergia frutescens. e temperatura média anual (°C). Linhas cinzas = dados
climáticos. Linhas pretas = índices.
A
B
-2,00
-1,00
0,00
1,00
2,00
1986
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
Índi
ce
1000,00
1250,00
1500,00
1750,00
2000,00
Pre
cipi
taçã
o
C
-2,00
-1,00
0,00
1,00
2,00
1986
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
Índi
ce
18,00
20,00
22,00
24,00
Tem
pera
tura
-2,00
-1,00
0,00
1,00
2,00
3,00
1986
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
2003
2004
2005
2006
Índi
ce
18,00
20,00
22,00
24,00
Tem
pera
tura
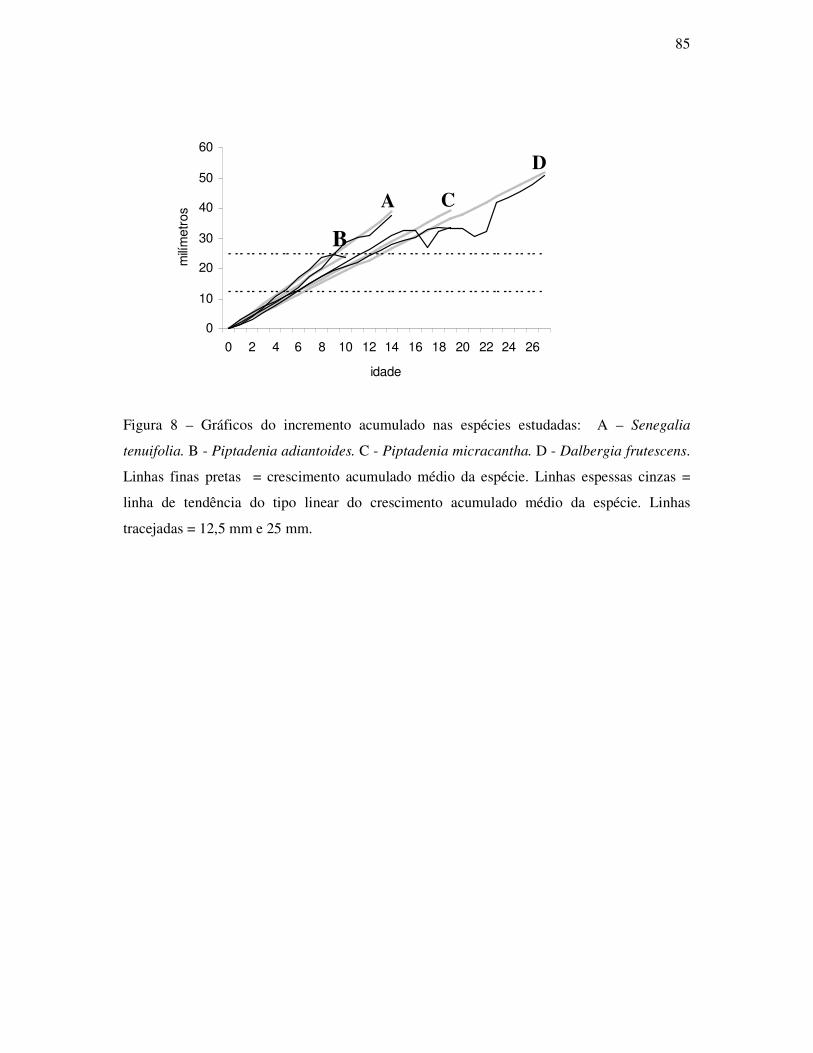
85
Figura 8 – Gráficos do incremento acumulado nas espécies estudadas: A – Senegalia
tenuifolia. B - Piptadenia adiantoides. C - Piptadenia micracantha. D - Dalbergia frutescens.
Linhas finas pretas = crescimento acumulado médio da espécie. Linhas espessas cinzas =
linha de tendência do tipo linear do crescimento acumulado médio da espécie. Linhas
tracejadas = 12,5 mm e 25 mm.
0
10
20
30
40
50
60
0 2 4 6 8 10 12 14 16 18 20 22 24 26
idade
milí
met
ros A
B
C
D
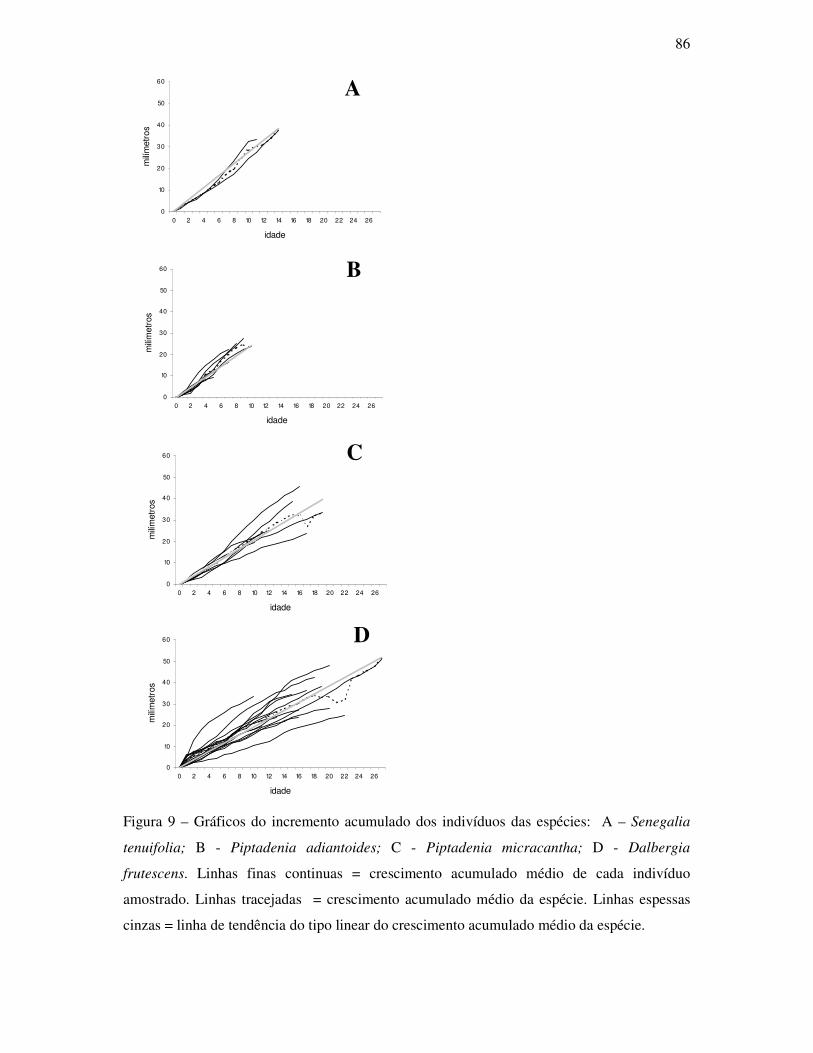
86
Figura 9 – Gráficos do incremento acumulado dos indivíduos das espécies: A – Senegalia
tenuifolia; B - Piptadenia adiantoides; C - Piptadenia micracantha; D - Dalbergia
frutescens. Linhas finas continuas = crescimento acumulado médio de cada indivíduo
amostrado. Linhas tracejadas = crescimento acumulado médio da espécie. Linhas espessas
cinzas = linha de tendência do tipo linear do crescimento acumulado médio da espécie.
0
10
20
30
40
50
60
0 2 4 6 8 10 12 14 16 18 20 22 24 26
idade
milí
met
ros
0
10
20
30
40
50
60
0 2 4 6 8 10 12 14 16 18 20 22 24 26
idade
milí
met
ros
0
10
20
30
40
50
60
0 2 4 6 8 10 12 14 16 18 20 22 24 26
idade
milí
met
ros
0
10
20
30
40
50
60
0 2 4 6 8 10 12 14 16 18 20 22 24 26
idade
milí
met
ros
A
B
C
D

87
CONSIDERAÇÕES GERAIS
Neste estudo foram produzidas descrições anatômicas das espécies de lianas da família
Leguminosae ocorrentes no Parque Nacional do Itatiaia, Senegalia grandistipula, Senegalia
lacerans, Senegalia martiusiana e Senegalia pedicellata, Senegalia tenuifolia, Piptadenia
adiantoides, Piptadenia micracantha e Dalbergia frutescens. Tendo sido constatadas
características esperadas para o hábito e para a família. Algumas características importantes
nestas espécies foram: variações cambiais; camadas de crescimento; diâmetro dos vasos,
altura e largura dos raios, fibras septadas e o tipo de parênquima axial. Através destas
características é possível distinguir as espécies da área de estudo.
As camadas de crescimento foram investigadas com mais detalhes, quanto a sua
anatomia. E constatou-se que poucos trabalhos já haviam relatado tal característica em lianas.
As espécies com variações cambiais, Senegalia grandistipula, Senegalia lacerans, Senegalia
martiusiana e Senegalia pedicellata, apresentaram camadas de crescimento de ocorrência
esporádica. As sem variações cambiais, Senegalia tenuifolia, Piptadenia adiantoides,
Piptadenia micracantha e Dalbergia frutescens, apresentaram camadas de crescimento
distintas bem delimitadas. Esta constatação mostrou a possibilidade de utilizá-las para análise
temporal.
Pelo método de lesão no câmbio, foi observado que as camadas de crescimento são
anuais. O comportamento temporal do crescimento radial e a idade dos indivíduos foram
obtidas através da análise das camadas de crescimento. Nas lianas deste estudo o incremento
médio do caule foi 2,30 mm, valor maior que o relatado em outros estudos. As lianas
estudadas respondem aos fatores que controlam o crescimento de maneira muito similar nas
diferentes regiões da circunferência, devido à correlação dos raios do mesmo indivíduo.
Contudo, parece que fatores locais são mais expressados no crescimento do que fatores de
larga escala, como fatores climáticos. Isto devido a não ter sido encontrada similaridade na
análise das séries temporais entre os diversos indivíduos e espécies, além de poucos
indivíduos apresentarem correlações com precipitação e temperatura.
Com este estudo mostra-se que a anatomia da madeira é uma ferramenta muito útil na
identificação das espécies de lianas que ocorrem na área de estudo. Através anatomia da
madeira de lianas é possível conhecer melhor características e estruturas pouco relatadas que
permitem a produção de novas hipóteses, como exemplo as camadas de crescimento em
lianas.

88
A dendrocronologia, por sua vez, tem-se mostrado muito eficiente em estudos com
espécies arbustivas e herbáceas, bem como em espécies arbóreas na floresta tropical. Com o
presente estudo acredita-se que também seja aplicável nas lianas, apesar das limitações e
ressalvas. Permitindo assim conhecer parâmetros úteis no manejo florestal e conservação,
como o crescimento secundário do caule e a idade.
Por fim, nota-se a importância de mais estudos com outros hábitos, especialmente com
lianas, visto a importância destes componentes florestais e a escassez de estudos a seu
respeito. Sejam estes estudos de natureza anatômica, ecológica, fisiológica, entre outras.

89
REFERÊNCIAS BIBLIOGRÁFICAS GERAIS
Araque, O.Z., De Pernía, N.E. & León, W.J. 2000. Estudio anatómico del leño de seis
especies de lianas. Revista Forestal Venezolana 44 (2): 39-48.
Araújo, G.U.C. 2005. Aspectos anatômicos do desenvolvimento do caule de Serjania
corrugata Radlk. (Sapindaceae). Dissertação de Mestrado – Instituto de Pesquisas
Jardim Botânico do Rio de Janeiro/Escola Nacional de Botânica Tropical.
Baas, P. & Schweingruber, F. H. 1987. Ecological trends in the wood anatomy of trees,
shrubs and climbers from Europe. IAWA Bulletin, Vol. 8(3):245-274.
Bamber, R.K. & Ter Welle, B.J.H. 1994. Adaptative trends in the wood anatomy of lianas. In:
Iqbal, M. (ed). 1994. Growth Patterns in Vascular Plants. Dioscorides Press, Portland,
Oregon.
Barreta-Kuipers, T. 1981. Wood Anatomy of Leguminosae: its relevance to taxonomy. In:
Advances in legumes systematics. Part 1. Royal Botanic Gardens, Kew.
Boninsegna, J.A., Villalba, R., Amarilla, L., & Ocampo, J. 1989. Studies on tree rings, growth
rates and age-size relationships of tropical tree species in Missiones, Argentina. IAWA
Bulletin n.s., vol. 10 (2) : 161-169.
Brade, A. C.1956. A flora do Parque Nacional do Itatiaia. Boletim Parque Nacional Itatiaia 5:
1-92.
Caballé, G. 1993. Liana structure, function and selection: a comparative study of xilem
cylinders of tropical rainforest species in Africa and America. Botanical Journal of the
Linnean Society, 113 : 41-60.
Callado, C.H., Neto, S.J.S., Scarano, F.R., Barros, C.F. & Costa, C.G. 2001a. Anatomical
features of growth rings in flood-prone trees of the Atlantic Rain Forest in Rio de
Janeiro, Brazil. IAWA Journal, vol. 22(1) 29-42.
Callado, C.H., Neto, S.J.S., Scarano, F.R. & Costa , C.G. 2001b. Periodicity of growth rings
in some flood-prone trees of the Atlantic Rain Forest in Rio de Janeiro, Brazil. Trees ,
vol 15:492–497.
Carlquist, S. 1975. Specialization in Dicotyledonous Wood. Ecological Strategies of Xylem
Evolution. California: The University of California Press.
Carlquist, S. 1985. Observations on functional wood histology of vines and lianas: vessel
dimorphism, tracheids, vasicentric tracheids, narrow vessels, and parenchyma. Aliso 11
(2): 139-157.

90
Carlquist, S. 1988. Comparative wood anatomy: Systematic, ecological and evolutionary
aspects of dicotyledon wood. Springer-Verlag, Berlin.
Carlquist, S. 1991. Anatomy of vine and liana stems: a review and synthesis. In: Putz, F.E.,
Mooney, H.A. (Eds), The Biology of Vines. Cambridge University Press, Cambridge,
United Kingdom, pp. 53-72.
Carlquist, S. 1995. Wood and Bark anatomy of Ranunculaceae (including Hydrastis) and
Glaucidiaceae. Aliso 14 (2) : 65-84.
Carlquist, S. 1996. Wood and stem anatomy of Menispermaceae. Aliso 14 (3) : 155-170.
Darwin, C. 1867. On the movements and habits of climbing plants. Journal of the Linnean
Society (Botany) 9: 1-118.
Dastur, R.H. & Kapadia, G.A. 1931. The anatomy of climbing plants. Journal of the Indian
Botanical Society 10 (2): 110-121.
Dias-Leme, C.L. 1999. Árvores e lianas de Leguminosae Papilionoideae: anatomia do caule.
Tese de Doutorado. Universidade de São Paulo. São Paulo.
Dobbins, D.R. 1981. Anomalous secundary growth in lianas of the Bignoniaceae is correlated
with the vascular pattern. Amer. J. Bot. 68(1) :142-144.
Esau, K. 1985. Anatomia Vegetal. 3ed. Omega Barcelona.
Evans, J. A., Gasson, P.E. e Gwilym, P.L. 2006. Wood Anatomy of the Mimosoideae
(Leguminosae). IAWA Journal, suplemento 5.
Ewers, F.W. 1985. Xylem structure and water conduction in conifer trees, dicot trees, and
lianas. IAWA Bulletin 6 (4): 309-371.
Ewers, F.W. & Fisher, J.B. 1987. Water transport in lianas. Fairchild Tropical Garden
Bulletin 42 (4): 28-29.
Ewers, F.W. & Fisher, J.B. 1989. Variation in vessel length and diameter in stems of six
tropical and subtropical lianas. American Journal of Botany 76 (10): 1452-1459.
Ewers, F.W. & Fisher, J.B. 1991. Why vines have narrow stems: histological trends in
Bauhinia (Fabaceae). Oecologia 88: 233-237.
Ewers, F.W., Fisher, J.B. & Chiu, S.-T. 1989. Water transport in the liana Bauhinia
fassoglensis. Plant Physiology, 91: 1625-31.
Ewers, F.W., Fisher, J.B. & Chiu, S.-T. 1990. A survey if vessel dimensions in stem of
tropical lianas and others growth forms. Oecologia 84: 544-552.
Ewers, F.W., Fisher, J.B. & Fichtner, K. 1991. Water flux and xylem structure in vines. In:
Putz, F.E., Mooney, H.A. (Eds), The Biology of Vines. Cambridge University Press,
Cambridge, United Kingdom, pp. 127-160.

91
Ewers, F.W., Carlton, M.R., Fisher, J.B., Kolb, K.J. & Tyree, M.T. 1997. Vessel diameters in
roots versus stems of tropical lianas and other growth forms. IAWA Journal 18 (3):
261-279.
Fahn, A. 1990. Plant Anatomy. 4 ed. Pergamon Press, Oxford.
Ferreira, L. 2002. Periodicidade do crescimento e formação da madeira de algumas espécies
arbóreas de florestas estacionais semidecíduas da região sudeste do estado de São
Paulo. São Paulo, 103p. Dissertação de mestrado, Escola Superior de Agricultura “Luiz
de Queiroz”, USP.
Fisher, J.B. & Ewers, F.W. 1989. Wound healing in stems of lianas after twisting and girdling
injuries. Botanical Gazette 150 (3): 251-265.
Fisher, J.B. & Ewers, F.W. 1991. Structural responses to stem injury in vines. In: Putz, F.E.,
Mooney, H.A. (Eds), The Biology of Vines. Cambridge, U.K., Cambridge University
Press, pp. 99-124.
Fisher, J. B. & Ewers, F.W. 1992. Xilem pathways in liana stems with variant secundary
growth. Botanical Journal of the Linnean Society, 108 : 181-202.
Fisher, J.B. & Ewers, F.W. 1995. Vessel dimension in liana and tree species of Gnetum
(Gnetales). American Journal of Botany 82(11) : 1350-1357.
Gartner, B.L., Bullock, S.H. Mooney, H.A., Brown, V.B. & Whitbeck, J.L. 1990. Water
transport properties of vine and tree stems in a tropical deciduous forest. AM.J.Bot. 77 :
742-749.
Gasson, P. 1994. Wood anatomy of the Tribe Sophoreae and related Caesalpinoidae and
Papilionoidae. In: Ferguson, I.K. & Tucker, S. (eds.). Advances in Legume Systematics
6, Structural Botany, 165-203.
Gasson, P. 2000. Does wood anatomy support tribal and generic classification in Papilionoid
Leguminosae. In: Herendeen, P.S. & Bruneau, A. (eds.). Advances in Legume
Systematics 9, 201-215. Royal Botanic Gardens, Kew.
Gasson, P. 2003. Wood anatomy of Caesalpinioideae. In: Klitgaard, B.B. & & Bruneau, A.
(eds.). Advances in Legume Systematics, part 10, 63-93. Royal Botanic Gardens, Kew.
Gasson, P. & Dobbins, D.R. 1991. Wood anatomy of the Bignoniaceae, with a comparison of
trees and lianas. IAWA Bulletin 12 (4): 389-417.
Gentry, A.H. 1982. Patterns os neotropical plant species diversity. Evolutionary Ecology 15:
1-84.

92
Gentry, A.H. 1985. An ecotaxonomic survey of Panamanian lianas. In: Historia Natural de
Panama, ed. D´arcy, W.G. & Correa, M., pp. 29-42. Missouri Botannical Garden, St
Louis.
Gentry, A. H. 1991. The distribution and evolution of climbing plants. In: Putz, F. E. &
Mooney, H. A. (eds). The Biology of Vines. Cambridge University Press.
Gray, J. & Thompson, P. 1977. Climatic information from 18O/16O analysis of cellulose,
lignin and whole wood from tree rings. NATURE 270 : 708 – 709.
Hegarty, E.E. 1991. Vine-host interactions. In: Putz, F.E., Mooney, H.A. (Eds), The
Biology of Vines. Cambridge University Press, Cambridge, United Kingdom, pp. 357-376.
Höhn, A. 1999. Wood anatomy of selected West African species of Caesalpinioideae and
Mimosoideae (Leguminosae): A comparative study. IAWA Journal Vol. 20 : 115.
Iawa Committee 1989. Growth Rings in Tropical Trees. IAWA Bulletin n.s. vol. 10 (2): 95-
174.
Iawa Committee 1995. Growth Periodicity in Tropical Trees. IAWA Journal 16 (4): 323-442.
Iawa Committee 1999. Dendrochronology in Monsoon Asia. IAWA Journal vol. 20 (3): 223-
350.
Iqbal, M. 1994. Growth patterns in vascular plantas. Dioscorides Press. Theodore R. Dudley,
PHD., General Editor Portland, Oregon.
Jacoby, G.C. 1989. Overview of tree-ring analysis in tropical regions. IAWA Bulletin, vol.10
(2) : 99-108.
Leon-Gómez, C. & Monroy-Ata, A. 2005. Seasonality in cambial activity of four lianas from
a Mexican lowland tropical rainforest. IAWA Journal 26(1) : 111-120.
Lima, M. P. M. & Guedes-Bruni, R.R. 1994. Reserva Ecológica de Macaé de Cima, Nova
Friburgo, RJ: Aspectos florísticos das espécies vasculares, v.1. Jardim Botânico do Rio
de Janeiro. RJ.
Mariaux, A. 1981. Past efforts in measuring age and annual growth in tropical trees. Yale
University: School of forestry and environmental studies, bulletin no.94, p. 20-30.
Mauseth, J.D. 1988. Plant Anatomy. Menlo Park: Benjamin/Cummings Publishing Company
Inc.
Morim, M.P. 2002. Leguminosae arbustivas e arbóreas do Parque Nacional do Itatiaia:
abordagem florístico-taxonômica. Tese (doutorado) – UFRJ/Museu Nacional, 2002.
Obaton, M. 1960. Les lianes ligneuses à structure anormales des forêts dense d´Afrique
occidentale. Annales des Sciences Naturelles, (Botanique), Sér. 12 -1: 1-220.

93
Philipson, W.R. 1980. Anomalous Cambia. In: IQBAL, M. The Vascular Cambium. P. 201-
212.
Putz, F.E. 2006. Vine ecology. ECOLOGY.INFO 24.
Putz, F.E. & Mooney, H.A. 1991. The Biology of Vines. Cambridge University Press.
Rajput, K.S. & Rao, K.S. 2003. Cambial variant and xylem structure in the stem of Cocculus
hirsutus (MENISPERMACEAE). IAWA Journal 24(4) : 411-420.
Resende, A.A. & Ranga, N.T. 2005. Lianas da Estação Ecológica do Nordeste Paulista, São
José do Rio Preto / Mirassol, SP, Brasil. Acta bot. bras. 19(2) : 273-279.
Roig, F. A.. 2000. Dendrocronologia en América Latina. Mendonza: EDIUNC.
Sass-Klaassen, U., Poole, I. Wils, T., Helle, G., Schleser, G.H. & Van Bergen, P.F.
2005. Carbon and oxygen isotope dendrochronology in sub-fossil bog oak tree rings – a
preliminary study. IAWA Journal, vol. 26(1) : 121-136.
Savva, Y.V., Schweingruber, F.H., Vaganov, E.A. & Milyutin, L.I. 2003. Influence of
Climate Changes on Tree-ring Characteristics of Scots Pine Provenances in Southern
Siberia (Forest-Steppe). IAWA Journal, vol. 24(4) : 371-383.
Schenck, H. 1892. Beiträge zur Biologie und Anatomie der Lianen, im Besonderon der in
Brasilien einheimischen Arten. 1. Beiträge zur biologie der lianen. In: Schimper, A.F.
(ed.). Botanische Mittheilungen aus den Tropen 4. Gustav Fischer, Jena.
Schenck, H. 1893. Beiträge zur Biologie und Anatomie der Lianen, im Besonderon der in
Brasilien einheimischen Arten. 2. Beiträge zur anatomie der lianen. In: Schimper, A.F.
(ed.). Botanische Mittheilungen aus den Tropen 5. Gustav Fischer, Jena.
Schnitzer, S.A. & Bongers, F. 2002. The ecology of lianas and their role in forests. Trends in
Ecology and Evolution 17 (5): 223-230.
Schweingruber, F.H., Poschlod, P. 2005. Growth rings in herbs and shrubs: life span, age
determination and stem anatomy. Forest Snow and Landscape Research 79: 195-415.
Shirley, J. & Lambert, C.A. 1918. The stems of climbing plants. Proceedings of the Linnean
Society of NSW 43: 600-609.
Solereder, H. 1908. Systematic anatomy of the Dicotyledons. Vol. 1 e 2, Oxford, Clarendon
Press.
Stokes, M. A. & Smiley, T. L. 1996. An Introduction to Tree-Ring Dating. Arizona: The
University of Arizona Press.
Tamaio, N. 2006. Anatomia do caule e da raiz em Menispermaceae. Tese de doutorado.
Instituto de Biociências da Universidade de São Paulo.
Ter Welle, B.J.H. 1985. Differences in wood anatomy of lianas and tree. IAWA Bull. 6:70.

94
Trouet, V., Haneca, K., Coppin, P. & Beeckman, H. 2001. Tree ring analysis of Brachystegia
spiciformis and Isoberlinia tomentosa: evaluation of the ENSO-Signal in the Miombo
Woodland of Eastern Africa. IAWA Journal, vol. 22(4) : 385-399.
Vieira, C.R. 1990. Contribuição ao conhecimento da anatomia ecológica de Bauhinia radiata
Vell.. Dissertação de mestrado em Botânica da Universidade Federal do Rio de Janeiro.
Worbes, M. 1988. Dendrochronological Studies on Tropical Trees. Plant Research and
Development, vol. 32: 86-98.
Worbes, M. 1989. Growth rings, increment and age of trees in imundation forests, savanas
and a mountain forest in the neotropics. IAWA Bulletin, Vol. 10 (2) : 109-122.
Worbes, M. 2002. One hundred year of tree-ring research in the tropics – a brief history and
an outlook to future challenges. Dendrochronologia, 20/1-2 : 217-231.
Yale University 1981. Age and Growth Rate of Tropical Trees: new directions for research.
Yale University: School of forestry and environmental studies, bulletin no.94.
Zimmermann, M.H. 1983. Xylem structure and the ascent of sap. Springer-Verlag, Berlin.