
A comunidade de moluscos terrestres retrata a alteração...
77
A comunidade de moluscos terrestres retrata a alteração ambiental da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro Gleisse Kelly Meneses Nunes Rio de Janeiro Setembro - 2009 UNIVERSIDADE DO ESTADO DO RIO DE JANEIRO INSTITUTO DE BIOLOGIA ROBERTO ALCANTARA GOMES DEPARTAMENTO DE ZOOLOGIA
Transcript of A comunidade de moluscos terrestres retrata a alteração...

A comunidade de moluscos terrestres retrata a
alteração ambiental da Vila Dois Rios, Ilha Grande,
Angra dos Reis, Rio de Janeiro
Gleisse Kelly Meneses Nunes
Rio de Janeiro
Setembro - 2009
UNIVERSIDADE DO ESTADO DO RIO DE JANEIRO
INSTITUTO DE BIOLOGIA ROBERTO ALCANTARA GOMES
DEPARTAMENTO DE ZOOLOGIA

ii
Gleisse Kelly Meneses Nunes
A comunidade de moluscos terrestres retrata a
alteração ambiental da Vila Dois Rios, Ilha Grande,
Angra dos Reis, Rio de Janeiro
Orientadora: Profa. Dra. Sonia Barbosa dos Santos
Monografia apresentada ao Instituto de Biologia
Roberto Alcantara Gomes da Universidade do Estado do
Rio de Janeiro, como parte dos requisitos necessários à
obtenção do grau de Bacharel em Ciências Biológicas.
Rio de Janeiro
Setembro - 2009

iii
FICHA CATALOGRÁFICA:

iv
Gleisse Kelly Meneses Nunes
A comunidade de moluscos terrestres retrata a
alteração ambiental da Vila Dois Rios, Ilha Grande,
Angra dos Reis, Rio de Janeiro
Resultado: _________________________com grau _______
Monografia de Bacharelado submetida como requisito parcial para
obtenção do grau de Bacharel em Ciências Biológicas, pela Universidade do
Estado do Rio de Janeiro, avaliada pela comissão formada pelos professores:
Orientadora: _____________________________________________ Dra. Sonia Barbosa dos Santos (UERJ) 1° Examinador ____________________________________________ Dr. José Ricardo Miras Mermudes (UERJ) 2° Examinador _____________________________________________ Msc. Amilcar Brum Barbosa (UERJ) 3° Examinador _____________________________________________ Msc. Claudia Leal Rodrigues (UERJ) Suplentes: ____________________________________________ Dr. Sandra Aparecida Magalhães-Fraga (Fiocruz) _____________________________________________ Dr. Oscar Rocha Barbosa (UERJ)
Rio de Janeiro, 21de setembro de 2009.

v
À minha família por todo
apoio e carinho.

vi
AGRADECIMENTOS
À minha orientadora, Professora Dra. Sonia Barbosa dos Santos.
À minha família, Neusa, Hudson, Gleidson, Gleivid e Luiz Eduardo pelo
conforto, carinho e apoio ao longo da minha jornada. Aos meus familiares pelo
incentivo.
A toda equipe do Laboratório de Malacologia da UERJ: Amilcar Brum
Barbosa, Claudia Leal Rodrigues, Francielle Cardoso Fonseca, Igor Christo Miyahira,
Isabela Cristina B. Gonçalves, Luciane Guilhermino, Jaqueline Lopes de Oliveira,
Luiz Eduardo Macedo de Lacerda, Patrícia do Socorro da Silva, Renata Ximenes e
Tiago Abreu Viana.
A todos os meus professores que foram sempre grandes educadores e
incentivadores.
A todos os funcionários do Centro de Estudos Ambientais e Desenvolvimento
Sustentável (CEADS) pelo grande apoio prestado durante os trabalhos de campo.
Aos funcionários do Departamento de Zoologia e da Secretaria do Instituto de
Biologia Roberto Alcantara Gomes.
Ao Instituto Estadual do Ambiente pela concessão da licença de coleta.
Ao Sr. Getúlio Cantuária por sua contribuição sobre a história de ocupação da
região da Vila Dois Rios.
Aos bibliotecários das Bibliotecas da Universidade do Estado do Rio de
Janeiro, de Museu da Universidade de São Paulo e do Museu Nacional do Rio de
Janeiro.

vii
A todos que contribuíram de forma direta ou indireta para a realização deste
trabalho.
À Universidade do Estado do Rio de Janeiro e ao Instituto de Biologia Roberto
Alcantara Gomes pela infra-estrutura.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
pela bolsa de Iniciação Científica concedida.

viii
SUMÁRIO
página
LISTA DE TABELAS........................................................................................................ x
LISTA DE FIGURAS........................................................................................................ xi
RESUMO........................................................................................................................ xiii
ABSTRACT..................................................................................................................... xv
INTRODUÇÃO................................................................................................................ 01
OBJETIVOS.................................................................................................................... 04
MATERIAL E MÉTODOS................................................................................................. 05
Caracterização da área de trabalho.................................................................... 05
Ilha Grande........................................................................................ 05
Vila Dois Rios.................................................................................... 06
Coleta dos moluscos terrestres.......................................................................... 07
Curadoria........................................................................................................... 11
Coleta de dados ambientais............................................................................... 11
Identificação dos moluscos terrestres................................................................ 12
RESULTADOS E DISCUSSÃO........................................................................................... 13
Características das espécies coletadas................................................................ 14
Alcadia Gray, 1840............................................................................. 14
Neocyclotus prominulus (d’Orbigny, 1835)....................................... 14
Succinea meridionalis d’Orbigny, 1837............................................ 15
Gastrocopta servilis (Gould, 1843) ................................................... 16
Bulimulus tenuissimus (d’Orbigny, 1835)........................................... 17
Beckianum beckianum (Pfeiffer, 1846)................................................ 18
Subulina octona (Bruguière, 1789)....................................................... 19
Lamellaxis gracilis (Hutton, 1834)........................................................22
Leptinaria unilamellata (d’Orbigny, 1835)...........................................23
Obeliscus Beck 1837..............................................................................25
Miradiscops Baker, 1925.......................................................................26

ix
página
Happiella Baker, 1925......................................................................... 26
Tamayoa Baker, 1925.......................................................................... 26
Ptychodon Ancey, 1888....................................................................... 26
Ptychodon schuppi (Suter, 1900)......................................................... 27
Stephanoda Albers, 1860..................................................................... 28
Habroconus semenlini (Moricand, 1846)............................................ 28
Bradybaena similaris (Férussac, 1821)............................................... 29
Área urbana da Vila Dois Rios........................................................................... 31
Encosta da Vila Dois Rios a 350 metros de altitude........................................... 33
Encosta da Vila Dois Rios a 450 metros de altitude........................................... 35
Dados ambientais................................................................................................ 38
CONCLUSÕES................................................................................................................. 47
REFERÊNCIAS BIBLIOGRÁFICAS..................................................................................... 48

x
L ISTA DE TABELAS
Página
Tabela I. Famílias e espécies coletadas na Vila Dois Rios, Ilha Grande, Angra dos
Reis, Rio de Janeiro......................................................................................................
13
Tabela II: Análise de Variância com os dados ambientais da Vila Dois Rios, Ilha
Grande, Angra dos Reis, Rio de Janeiro e teste Tukey................................................
44
Tabela III. Sugestão para diagnóstico de ambientes de Floresta Atlântica
degradados/ recuperados, utilizando a proporção de espécies de moluscos
sinantrópicas e nativas, para cada estágio de recuperação...........................................
46

xi
L ISTA DE FIGURAS
Página
Figura 1. Localização da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de
Janeiro...........................................................................................................................
8
Figura 2. Rua da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro.......... 8
Figura 3. Canteiros gramados na parte central da Vila Dois Rio, Ilha Grande, Angra
dos Reis, Rio de Janeiro e encosta ao fundo.................................................................
9
Figura 4. Encosta da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro.... 9
Figura 5. Vista aérea da Vila Dois Rios, Ilha Grande, Angra dos Reia, Rio de
Janeiro com localização dos pontos de coleta..............................................................
10
Figura 6. Neocyclotus prominulus (d’Orbigny, 1835).................................................. 15
Figura 7. Succinea meridionalis d´Orbigny, 1837....................................................... 16
Figura 8. Gastrocopta servilis (Gould, 1843)............................................................... 17
Figura 9. Bulimulus tenuissimus (d´Orbigny, 1835)..................................................... 18
Figura 10. Beckianum beckianum. (Pfeiffer, 1846)...................................................... 19
Figura 11. Subulina octona (Bruguière, 1789)............................................................. 21
Figura 12. Lamellaxis gracilis (Hutton, 1834)............................................................. 23
Figura 13. Leptinaria unilamellata (d’Orbigny, 1835). .............................................. 25
Figura 14. Ptychodon schuppi (Suter, 1900). .............................................................. 28
Figura 15. Habroconus semenlini (Moricand, 1846). ................................................. 29
Figura 16. Bradybaena similaris (Férussac, 1821). ..................................................... 31
Figura 17. Representação das famílias de moluscos terrestres coletadas na área
urbana da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro....................
32
Figura 18. Abundância dos moluscos terrestres coletados na Vila Dois Rios, Ilha
Grande, Angra dos Reis, Rio de Janeiro. .....................................................................
33
Figura 19. Representação das famílias de moluscos terrestres coletadas na encosta
da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro, a 350 metros de
altitude. ........................................................................................................................
34
Figura 20. Abundância das espécies coletadas na encosta da Vila Dois Rios, Ilha
Grande, Angra dos Reis, Rio de Janeiro, a 350 m de altitude. ....................................
35

xii
Figura 21. Representação das famílias de moluscos terrestres coletadas na encosta
da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro, a 450 metros de
altitude. ........................................................................................................................
36
Figura 22. Abundância das espécies coletadas na encosta da Vila Dois Rios, Ilha
Grande, Angra dos Reis, Rio de Janeiro, a 450 m de altitude. ...................................
36
Figura 23. Percentual de espécies coletas na Vila Dois Rios, Ilha Grande, Angra dos
Reis, Rio de Janeiro. 0m: área urbana; 350m: encosta a 350m de altitude; 450m:
encosta a 450m de altitude............................................................................................
37
Figura 24. Temperatura ambiente e do solo (ºC) da Vila Dois Rios, Ilha Grande,
Angra dos Reis, Rio de Janeiro....................................................................................
39
Figura 25. Percentual de umidade ambiente da Vila Dois Rios, Ilha Grande, Angra
dos Reis, Rio de Janeiro. .............................................................................................
40
Figura 26. Luminosidade (Lux) da Vila Dois Rios, Ilha Grande, Angra dos Reis,
Rio de Janeiro...............................................................................................................
41
Figura 27. Profundidade da serapilheira (cm) da Vila Dois Rios, Ilha Grande, Angra
dos Reis, Rio de Janeiro...............................................................................................
42
Figura 28. Percentual de umidade da serapilheira da Vila Dois Rios, Ilha Grande,
Angra dos Reis, Rio de Janeiro....................................................................................
43

xiii
RESUMO
Este trabalho teve como objetivo comparar a composição da comunidade de
moluscos terrestres em três diferentes locais da Vila Dois Rios (VDR), que se
apresentam segundo um gradiente, no qual a luminosidade e a temperatura do
ambiente e do solo diminuem e a umidade do ambiente, a profundidade e a umidade
da serapilheira aumentam, conforme passamos da área urbana para a floresta
secundária e depois, para a floresta climáxica, para construir índices utilizando a
proporção de espécies sinantrópicas de moluscos presentes nas comunidades, como
indicadores de alterações ambientais. Os locais de coleta foram: 1) área urbana (AU)
onde existe influência antrópica; 2) encosta da VDR a 350 m de altitude (VDR 350),
onde cessou a influência antrópica em 1975; 3) encosta da VDR a 450 m de altitude
(VDR 450), onde não houve influência antrópica. Em cada local foram utilizados dez
quadrats de 25x75 cm, de forma aleatória, para recolher a serapilheira, que foi
transportada até o laboratório, onde foi triada, buscando conchas e animais vivos.
Dados ambientais como profundidade da serapilheira, temperatura ambiente e do solo,
umidade relativa do ar e luminosidade foram aferidos em cada quadrat. No total,
foram coletadas 20 espécies, em 17 gêneros e 10 famílias. Na AU todas as espécies
são sinantrópicas, sete foram exclusivas. A família Subulinidade (91,5 %) foi
dominante, em especial a espécie Beckianum beckianum (Pfeiffer, 1846) (67 %). Na
VDR 350 quatro espécies foram exclusivas, a família Systrophiidae (61%) foi
dominante. Na VDR 450 quatro espécies foram exclusivas, a família Systrophiidae
(53%) foi dominante. Houve redução da proporção da família Subulinidae da AU para
VDR 350, até sua ausência na VDR 450. Concluímos que a comunidade de moluscos
terrestres respondeu ao gradiente ambiental através de sua composição. A partir destes

xiv
dados, sugerimos a utilização da proporção de espécies de moluscos sinantrópicas para
fazer o diagnóstico de ambientes de Floresta Atlântica degradados/recuperados, da
seguinte forma: ambiente com grande influência antrópica terá de 90 a 100% de
espécies sinantrópicas; ambiente que se recupera a pelo menos 35 terá de 50 a 60% de
espécies sinantrópicas; já uma floresta a pelo menos 50 anos sem influência antrópica
terá de 0 a 10% de espécies sinantrópicas. Este trabalho permitiu estabelecer uma
metodologia rápida, de pouco impacto e principalmente de baixo custo para classificar
ou diagnosticar o estado de conservação ou modificação de fragmentos florestais na
Ilha Grande.

xv
ABSTRACT
The aim of this study was to compare the community composition of land
snails in three different localities in the Vila Dois Rios (VDR), to build indexes using
the proportion of synanthropic species of molluscs ocurring in communities, as
indicators of environmental change. The localities appear as a gradient in which the
luminosity, atmospheric and soil temperature decreases; in the other hand, relative air
humidity, depth and moisture content of litter increases from the urban area to the
secondary forest and then to the climax forest. The sampling sites were: 1) urban area
(UA) where there is human influence; 2) VDR’s mountain slope at 350 m als (VDR
350), where human influence ceased in 1975; 3) VDR’s mountain slope at 450 m als
(VDR 450), where there were no human influence. At each area we used ten quadrats
of 25x75 cm, to collect the litter. The samples, collected at random, were transported
to the laboratory where they were searched for shells and live animals. Environmental
data such as litter depth, atmospheric and soil temperature, relative air humidity and
luminosity were measured in each quadrat. In UA all species were synanthropic being
seven exclusive. Family Subulinidae (91.5%) was dominant, especially the species
Beckianum beckianum (Pfeiffer, 1846) (67%). In VDR 350 four species were
exclusive and Systrophiidae (61%) was dominant. In VDR 450 four species were
exclusive and, one more time, Systrophiidae (53%) was dominant. There was a
reduction in the Subulinidae proportion from UA to VDR 350 and total disappearance
in VDR 450. We concluded that the composition of terrestrial molluscs responds to
environmental gradients. From these data, we suggest the use of the proportion of
synanthropic mollusc species as a index to the diagnosis of Atlantic Forest
environments as follows: environments with large human influence will have 90 to

xvi
100% of synanthropic species; environments that are recovering for at least 35 years
will have 50 to 60% of synanthropic species; and a forest that is recovering for at least
50 years with no human influence will have 0 to 10% of synanthropic species. The aim
of this work was to establish a rapid method of low impact and low cost to classify or
evaluate the stage of conservation of forest fragments in the Ilha Grande.

1
INTRODUÇÃO
A distribuição das espécies nos ecossistemas é influenciada por diversos fatores
bióticos e abióticos, os quais operam como filtros para a dispersão. Os antigos processos de
colonização e migração humana e a recente tendência de globalização são os principais
responsáveis pela transposição das barreiras geográficas e introdução de espécies em regiões
fora de sua distribuição original (ESPINOLA & JUNIOR 2007).
A introdução de espécies exóticas tem sido amplamente discutida na comunidade
científica, devido aos problemas que causam. Representam uma ameaça à biodiversidade
local, por ter forte influência sobre as espécies nativas, via predação ou competição, e também
destruição ou modificação do hábitat, podendo inclusive levar à extinção de outras espécies
(COWIE 2001, PRIMACK & RODRIGUES 2001, AUBRY et al. 2005). As espécies introduzidas
podem ainda se tornar pestes na agricultura e ecossistemas naturais (BARKER & MAYHILL
1999).
Diversos trabalhos relatam a introdução de espécies de moluscos terrestres em várias
partes do mundo; geralmente, a distribuição dessas espécies está relacionada à ocupação
humana e sua consequente modificação de hábitat (COWIE 1998, CAMERON 2002). Algumas,
cuja origem não pode ser identificada, são denominadas espécies sinantrópicas, possuindo
ampla distribuição geográfica, ocorrendo em diversos países. Muitas das introduções recentes
foram provavelmente acidentais através de plantas de horticultura e ornamentação (COWIE
2001), sendo que a destruição ou modificação do hábitat facilita o estabelecimento de
espécies não nativas (COWIE 1998). Os impactos de espécies exóticas são uma das maiores
ameaças a muitos ecossistemas, especialmente ilhas (COWIE 2000b).
Um modo de detectar e monitorar os padrões de mudança na biodiversidade
provocados por ações humanas é utilizar espécies, ou grupo de espécies, que funcionam como

2
bioindicadoras de degradação ambiental (SANTOS et al. 2006). SCHILTHUIZEN & RUTJES
(2001) defendem que os moluscos têm várias características que permitem ter sucesso como
indicadores para avaliação da diversidade. Segundo os autores, a malacofauna pode ser
coletada facilmente de forma não destrutiva, por coleta de conchas vazias da serapilheira;
além disso, qualquer grupo de molusco terrestre pode ser coletado na serapilheira, mesmo os
arborícolas, porque suas conchas caem no chão.
Enquanto o declínio ou desaparecimento de vertebrados chama a atenção do público, a
homogeneização global de plantas e invertebrados é menos divulgada, embora, mais
preocupante, já que constitui uma maior perda de biodiversidade (COWIE 2001). Por isso é
importante estudar a malacofauna terrestre tanto em ambientes florestados como em
ambientes antrópicos, para conhecer a composição de gastrópodes terrestres nesses diferentes
ambientes e verificar se a introdução de espécies está sendo restrita aos ambientes urbanos, ou
se há risco de introdução em ambientes florestados, o que potencialmente traria diversos
prejuízos para as espécies nativas.
Em outras partes do mundo, existe a preocupação de relatar a ocorrência de espécies
exóticas ou sinantrópicas, por exemplo, República Checa (JUŘIČKOVÁ 2006), Israel (M IENIS
1977), Nova Zelândia (BARKER 1982), Austrália (HOLLAND et al. 2007), São Tomé e Príncipe
(GASCOIGNE 1994), Ilhas de Tristão da Cunha (PREECE 2001), México (CORREA 1999), Havaí
(COWIE 1998, 2000a,b, 2001), Argentina (MIQUEL & PARENT 1996), Uruguai (OLAZARRI
1986), Colômbia (HAUSDORF 2002) e Peru (RAMÍREZ et al. 2001).
Devido à relevância do tema, no Brasil podemos apontar algumas iniciativas do
governo brasileiro, desde 2001, com a realização em Brasília da “Reunião de Trabalho sobre
Espécies Exóticas Invasoras”, por meio de parceria entre o Ministério do Meio Ambiente -
MMA e a Empresa Brasileira de Pesquisa Agropecuária – Embrapa, onde se discutiu as linhas
gerais do problema e as principais estratégias para amenizá-lo, uma delas foi a produção de

3
um Informe Nacional sobre Espécies Exóticas Invasoras no país, visando sistematizar e
divulgar a informação já existente sobre o tema (MMA 2006).
Todavia, apesar dos esforços, ainda não existe uma listagem completa para espécies de
moluscos terrestres em ambientes antrópicos no território nacional, mas alguns trabalhos,
mesmo com resultados parciais ou incompletos, já foram realizados no Rio Grande do Sul (C.
OLIVEIRA 2002, SILVA 2007), no Paraná (CAMPOS et al. 2005) e na Ilha Grande (HAAS 1953,
NUNES et al. 2003, NUNES & SANTOS 2006, FONSECA et al. 2007, FONSECA et al. 2009).
O problema é ainda maior, porque parte considerável da produção científica recente
sobre florestas tropicais tem sido descritiva e fragmentada. Desse modo, apesar do crescente
interesse pela questão ambiental, pode-se observar que a importância da presença e da
influência humanas sobre o meio natural não é geralmente considerada em toda sua extensão
(HADLEY 1991 apud OLIVEIRA & NETTO 2006).

4
OBJETIVOS
O objetivo deste trabalho foi comparar a composição da comunidade de moluscos
terrestres em três diferentes locais da Vila Dois Rios, que constituem um gradiente ambiental,
visando construir índices utilizando a proporção de espécies sinantrópicas de moluscos
terrestres presentes nas comunidades malacológicas como indicadoras de alterações
ambientais.

5
MATERIAL E MÉTODOS
Caracterização da área de trabalho
ILHA GRANDE
A Ilha Grande está localizada ao sul do Estado do Rio de Janeiro, no município de
Angra dos Reis, entre a latitude 23º05' e 23º15' S e a longitude 44º05' e 44º23' O. Por ser um
importante remanescente de Floresta Atlântica do Estado, faz parte da Área de Proteção
Ambiental dos Tamoios. Além disso, abriga o Parque Estadual da Ilha Grande, a Reserva
Biológica da Praia do Sul e o Parque Estadual Marinho da Praia do Aventureiro. Situada no
domínio da Floresta Atlântica, predominando floresta ombrófila densa de encosta com
variados graus de influência antrópica (ALHO et al. 2002, R. OLIVEIRA 2002, ALVES et al.
2005), em diversas etapas de recomposição, além de ambientes como mangues, restingas,
lagoas, rios, etc. (SANTOS & MONTEIRO 2001).
Os sinais de ocupação da Ilha Grande datam de três mil anos atrás, com uma
população de pescadores primitivos e coletores de mariscos que provavelmente praticavam
canibalismo, é possível que a atuação nas florestas se limitasse à caça e à derrubada de
grandes árvores com machado (com preferência por certas espécies) para a construção de
canoas de madeira (TENÓRIO 1999 apud WUNDER 2006). Inicialmente a Ilha Grande foi
habitada por índios goianás. Após a fase de ocupação indígena, durante todo o século XIX, a
Ilha Grande foi um significativo centro agrícola. A demanda por terra multiplicou e grandes
posses de terras agrícolas (fazendas) foram estabelecidas em diversos locais, como o Abraão,
Dois Rios e Parnaioca (WUNDER 2006), de forma que, na primeira metade do século XIX, o
café era produzido em abundância, tornando-se o elemento principal da sua lavoura (MELLO
1987).
Mais tarde, as fazendas de Dois Rios e do Abraão tornaram-se pontos de desembarque
de escravos vindos da costa da África (MACIEL et al. 1984). Às vezes, o desmatamento

6
causava uma degradação permanente da terra cultivada, mas esses impactos se restringiam a
terrenos próximos à costa que não eram inclinados demais para plantações (OLIVEIRA &
COELHO NETTO 1996 apud WUNDER 2006). Na passagem para o século XX, as plantações
haviam decaído e as florestas estavam se recuperando, uma tendência que foi reforçada ao
longo desse século. A antiga fazenda de Dois Rios foi convertida em prisão no período de
1893 a 1903, quando a Ilha Grande teve sua história associada a instituições carcerárias, com
a criação da Colônia Correcional de Dois Rios, que perdurou até 1994, quando houve a
implosão do Instituto Penal Cândido Mendes e, desde então, o local e seu entorno passaram
por várias mudanças.
O impacto resultante dessa sequencia histórica de intervenções na Ilha Grande é que
atualmente, nas regiões onde cessou a atividade agrícola, as florestas se recuperam
amplamente, graças à predominância de ciclos neutros durante quase um século (WUNDER
2006). O que não ocorreu nas regiões da baixada da Vila Dois Rios, que durante as atividades
do presídio, continuaram sendo utilizadas para plantação de diversos produtos agrícolas, que
serviam para a população carcerária, funcionários do presídio e seus familiares. Dentre os
principais produtos, podemos destacar banana, aipim, milho, capim, hortaliças, etc., além da
retirada de madeira para servir de lenha aos fornos.
VILA DOIS RIOS
Atualmente as áreas ocupadas pelas fazendas apresentam mata secundária em
processo de sucessão ecológica. A vegetação da área urbana da Vila Dois Rios (23º 05’ – 23º
15’S; 44º 05’ – 44º 23’ O) é constituída por grandes jardins e gramados, algumas árvores
introduzidas de grande porte e arbustos de médio porte. Ao redor temos floresta secundária
em regeneração há cerca de 20 anos. Em geral, os topos dos morros se mantiveram

7
razoavelmente preservados, ocorrendo apenas cortes seletivos de árvores de grande porte. A
Vila Dois Rios faz parte do Parque Estadual da Ilha Grande.
A equipe do Laboratório de Malacologia da UERJ vem realizando coletas na Vila
Dois Rios e região de entorno desde 1995, quando a Vila Dois Rios passou a ser administrada
pela UERJ, que se tornou um campus avançado. Atualmente, na referida vila, está instalado o
Centro de Estudos Ambientais e Desenvolvimento Sustentável (CEADS), que é utilizado por
pesquisadores e alunos que contam com uma boa infra-estrutura para desenvolver suas
atividades de campo.
Buscando comparar locais com diferentes fases de regeneração da mata, foram
realizadas coletas em três localidades da Vila Dois Rios que representam um gradiente
ambiental:
1) Área urbana da Vila Dois Rios, onde atualmente moram aproximadamente 115
moradores. Esta vila foi estabelecida junto com a implantação do Complexo Penal, em 1894,
antes disso, o local fazia parte da fazenda de Dois Rios. Nível do mar (23º 11.106’ S; 44º
11.547’ O) (Fig. 2 e 3).
2) Encosta voltada para a Vila Dois Rios a 350 metros de altitude (23º 10.410’ S; 44º
11.746’ O), onde ocorreu cultivo de capim para alimentação de bois até o ano de 1975 (Fig.
4).
3) Encosta voltada para a Vila Dois Rios a 450 metros de altitude (23º 10.339’ S; 44º
11.812’ O), onde não ocorreu cultivo Fig. 4).
Coleta dos moluscos terrestres
As coletas foram realizadas em 24/10/2003 (região de encosta) e 01/04/2004 (área
urbana) na Vila Dois Rios, Ilha Grande, no município de Angra dos Reis, ao sul do estado do
Rio de Janeiro (Fig. 1).

8
Figura 1. Localização da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro.
Figura 2. Rua da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro.

9
Figura 3. Canteiros gramados na parte central da Vila Dois Rio, Ilha Grande, Angra dos Reis, Rio de Janeiro e
encosta ao fundo.
Figura 4. Encosta da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro. Foto: Luiz Eduardo M. de
Lacerda.

10
Em cada local foram utilizados dez quadrats de 25 x 75 cm (Fig. 5), de forma
aleatória, para coletar a serapilheira, que foi recolhida com ancinhos e as mãos até que o solo
ficasse exposto e, acondicionada em sacos plásticos identificados, para transporte até o
laboratório (NUNES 2007).
Figura 5. Vista aérea da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro com localização dos pontos
de coleta marcados com asteriscos vermelhos.
No Laboratório do CEADS, a serapilheira foi triada, minuciosamente, a olho nu, com
o auxílio de bandejas e pincéis. Os moluscos encontrados em cada ponto de coleta foram
mantidos em placas de Petri, separados, identificados, contados e tabulados para posterior
identificação ao menor nível taxonômico possível.
**
**
**
** **
**
** **
**
**

11
Curadoria
Para limpar as conchas vazias coletadas, elas foram separadas e identificadas,
colocadas em placas de Petri, imersas em água por aproximadamente 48 horas, e escovadas
com pincéis macios para retirar o excesso de sujeira. Conchas maiores ou mais sujas
demoraram mais tempo de molho. Quando as conchas estavam limpas, elas foram colocadas
para secar sobre papel absorvente em placa de Petri por aproximadamente 96 horas, ou até
que ficassem completamente secas.
Os animais encontrados vivos foram mantidos imersos em água previamente fervida e
resfriada, até o relaxamento total (THOMÉ & LOPES 1973, THOMÉ 1975), depois foram fixados
com álcool a 96° GL, para futuros estudos taxonômicos e moleculares.
Tanto as conchas como os animais coletados com a parte mole foram armazenados em
frascos plásticos de diversos tamanhos, potes de vidro ou ependorfes, receberam um número
de tombo, uma etiqueta contendo os dados de coleta e foram depositados na Coleção de
Moluscos da Universidade do Estado do Rio de Janeiro (Col. Mol. UERJ).
Coleta de dados ambientais
Em cada ponto de coleta (quadrat) foram coletadas as seguintes variáveis ambientais,
de acordo com propostas da literatura (MENEZ 2002):
a) Profundidade da serapilheira: foi medida em seis pontos dentro do quadrat de 25 X
75 cm, afundando um palito graduado em centímetros na serapilheira até que fosse percebida
a resistência oferecida pelo solo. Utilizou-se a média de cada ponto de coleta nas análises
estatísticas.
b) Temperatura ambiente (máxima, média e mínima): foi medida com uso de
higrotermômetro digital TFA da marca Biocristal que ficou em repouso no quadrat durante a
coleta da serapilheira.

12
c) Temperatura do solo: foi medida com um termômetro Incotherm (graduação -35 a
55 °C) que foi parcialmente enterrado no solo, imediatamente ao lado do quadrat, por alguns
minutos durante a coleta da serapilheira.
d) Umidade relativa do ar (máxima, média e mínima): foi medida com uso de
higrotermômetro digital TFA da marca Biocristal que ficou em repouso ao lado do quadrat
durante a coleta da serapilheira.
e) Luminosidade: foi medida através de luxímetro digital LDR- 208 da marca
Instrutherm posicionado cerca de 50 cm acima do quadrat.
No momento da triagem, a serapilheira foi separada em folhas e galhos, e estes foram
acondicionados em sacos de papel com peso conhecido, e pesados com a intenção de obter o
peso úmido. Depois foram colocados em estufa a 55ºC até a secagem total do material, que
foi novamente pesado, para obter o peso seco. O processo foi repetido para cada amostra até
que fosse verificada a estabilização do peso. Calculando a diferença entre o peso úmido e o
peso seco, é possível calcular o percentual de umidade da serapilheira por regra de três
simples.
Foi verificada a normalidade para cada variável ambiental antes da realização da Análise
de Variância através do programa Systat 11.0, com o objetivo de verificar se existia diferença
estatisticamente significativa entre as médias das áreas (UNDERWOOD 1997), posteriormente,
foi realizado teste de Tukey para verificar entre quais áreas há diferença estatisticamente
significativa.
Identificação dos moluscos terrestres
Ainda no CEADS, todos os animais foram observados sob microscópio estereoscópio
da marca Leica modelo MZ6, separados por morfotipos e tabulados. Depois, no Laboratório
de Malacologia da UERJ, foram novamente analisados e determinados até o menor nível
taxonômico possível. Um representante de cada espécie ou morfotipo foi selecionado e
desenhado em câmara clara acoplada ao microscópio Olympus, modelo SZH10, para facilitar
a observação de caracteres de importância taxonômica.
13
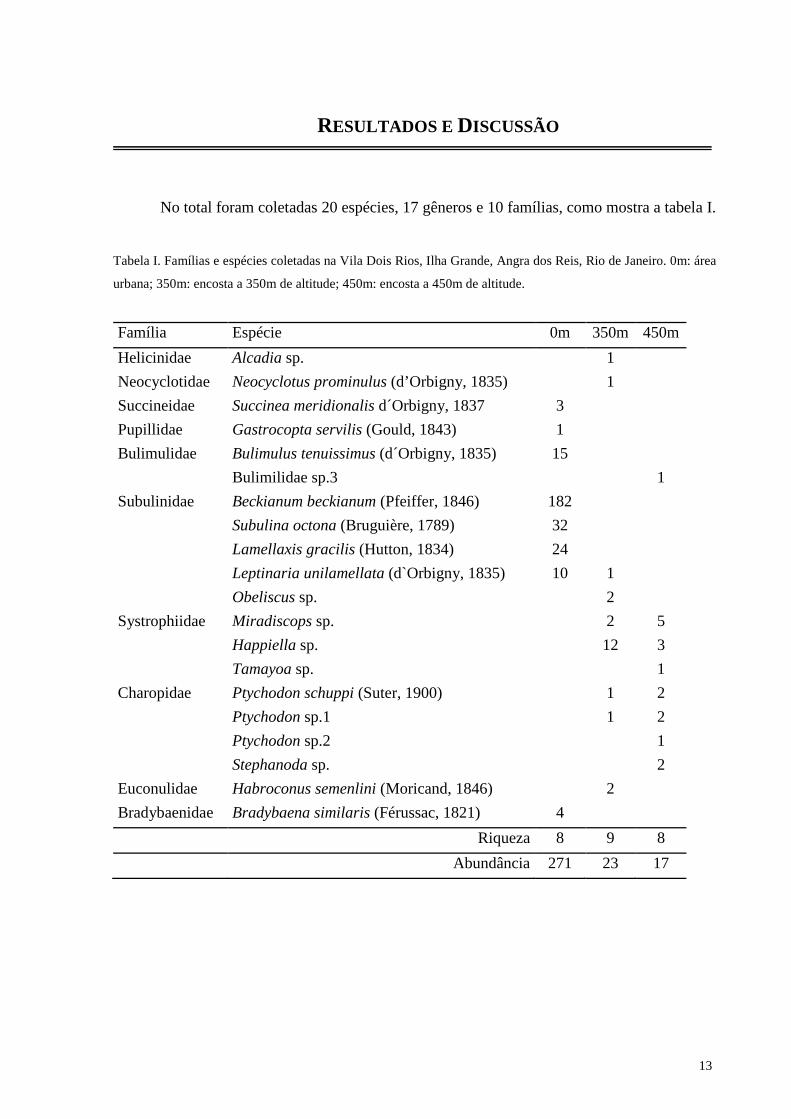
RESULTADOS E DISCUSSÃO
No total foram coletadas 20 espécies, 17 gêneros e 10 famílias, como mostra a tabela I.
Tabela I. Famílias e espécies coletadas na Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro. 0m: área
urbana; 350m: encosta a 350m de altitude; 450m: encosta a 450m de altitude.
Família Espécie 0m 350m 450m
Helicinidae Alcadia sp. 1
Neocyclotidae Neocyclotus prominulus (d’Orbigny, 1835) 1
Succineidae Succinea meridionalis d´Orbigny, 1837 3
Pupillidae Gastrocopta servilis (Gould, 1843) 1
Bulimulidae Bulimulus tenuissimus (d´Orbigny, 1835) 15
Bulimilidae sp.3 1
Subulinidae Beckianum beckianum (Pfeiffer, 1846) 182
Subulina octona (Bruguière, 1789) 32
Lamellaxis gracilis (Hutton, 1834) 24
Leptinaria unilamellata (d`Orbigny, 1835) 10 1
Obeliscus sp. 2
Systrophiidae Miradiscops sp. 2 5
Happiella sp. 12 3
Tamayoa sp. 1
Charopidae Ptychodon schuppi (Suter, 1900) 1 2
Ptychodon sp.1 1 2
Ptychodon sp.2 1
Stephanoda sp. 2
Euconulidae Habroconus semenlini (Moricand, 1846) 2
Bradybaenidae Bradybaena similaris (Férussac, 1821) 4
Riqueza 8 9 8
Abundância 271 23 17

14
Características das espécies coletadas
Uma breve busca bibliográfica foi realizada, procurando a localidade tipo, as
características das espécies ou gênero (para aqueles que não foram identificadas até a
categoria específica), a distribuição geográfica e os trabalhos que foram realizados no país
abordando as espécies encontradas neste trabalho.
Família Helicinidae
Alcadia Gray, 1840
Concha heliciforme, enrolada, subglobosa ou ligeiramente deprimida, apresenta
“pêlos”, com calo perto da columela, que é achatada, quase retilínea. Perióstraco muitas vezes
coberto com linhas de “pêlos”. Abertura semioval, quase triangular. Perístoma mais ou menos
expandido, separado da columela, na maior parte curvo; opérculo sólido, semioval (GRAY
1857).
Família Neocyclotidae
Neocyclotus prominulus (d’Orbigny, 1835) (Fig. 6)
Localidade-tipo: Rio de Janeiro (D’ORBIGNY 1837). Espécie nativa, encontrada em
ambientes florestados.
Concha dextrógira, helicoidal, deprimida, de cor marrom, apresenta estrias
unilamelares regulares entre as quais existem microestrias. Abertura é circular e o opérculo
também. Espira apresenta-se cônica com ápice obtuso e convexo; linha de sutura profunda.
Protoconcha é elevada, lisa e possui, em média, 1 ½ voltas sem estrias ou pontuações.
MONTEIRO (2005) comparou a morfologia da concha desta espécie em três diferentes
regiões da Ilha Grande e concluiu que a concha dessa espécie não sofre interferência das
alterações ambientais, nas áreas estudadas, o que poderia ser explicada pela presença do

15
opérculo, que auxiliaria na manutenção das condições hidroeletrolíticas, minimizando a
influência do ambiente sobre o animal.
Ocorrência: Minas Gerais, Rio de Janeiro, São Paulo e Paraná (MORRETES 1949). Ilha
Grande, Angra dos Reis Rio de Janeiro (HAAS 1953), nas trilhas da Parnaioca, Caxadaço e
Jararaca (MONTEIRO 2005), na trilha da Jararaca de 100 a 400 m de altitude e na trilha do Pico
do Papagaio de 200 a 400 m de altitude (NUNES 2007).
Família Succineidae
Succinea meridionalis d´Orbigny, 1837 (Fig. 7)
A localidade-tipo desta espécie não foi encontrada no material pesquisado.
Concha de forma oval, brilhante, geralmente translúcida e muito frágil; existem
conchas opacas e relativamente fortes. O número de voltas varia de 2 1/6 a 3 1/6 e as conchas
atingem o comprimento de 13,1 mm. As voltas são convexas. As linhas de sutura ligeiramente
inclinadas, são bem marcadas e profundas. As linhas de crescimento são relativamente
acentuadas. Abertura ovalada, vertical e bem arredondada na base. Columela arqueada,
suboblíqua. Perístoma delgado e sem sinuosidade (LANZIERI 1965). Espécie sinantrópica.
A espécie é muito higrófila e pode abrigar parasitas. LANZIERI (1965) relatou o
encontro de dois exemplares com esporocisto de Trematoda no tentáculo.
Distribuição: Rio de Janeiro, Curitiba e Paraná (MORRETES 1949). Maceió, Rio de
Janeiro, Paraná, Rio Grande do Sul, Uruguai, Argentina, Peru e Chile (LANZIERI 1965). Ilha
Figura 6. Neocyclotus prominulus (d’Orbigny, 1835). Escala 5 mm.
Foto: Amilcar B. Barbosa.

16
Grande, Angra dos Reis, Rio de Janeiro (HAAS 1953). Do Rio de Janeiro à Patagônia (SIMONE
2006).
Figura 7. Succinea meridionalis d´Orbigny, 1837. Desenho copiado de LANZIERI (1965).
Família Pupillidae
Gastrocopta servilis (Gould, 1843) (Fig. 8)
Localidade-tipo: Matanzas, Cuba (SOLEM 1961), México (BIELER & SLAPCINSKY
2000), espécie neotropical (CHRISTENSEN & KIRCH 1986, COWIE 1998). Espécie sinantrópica.
Concha alongada; ápice agudo, cor castanho pálido; cinco voltas, muito delicada,
sutura bem definida. Abertura semioval, perto do eixo da concha, a porção transversa
ligeiramente oblíqua. Um dente lamelar perto da junção do lábio externo, um grande dente
columelar cônico, na base um dente pequeno, e próximo, um outro dente, colocado de modo a
formar um tripé regular com os outros dois, e acima deste em quinto dente inconspícuo. Lábio
ligeiramente evertido, não achatado, às vezes um pouco flexionado à direita; umbílico
pequeno (GOULD 1843 apud PISBRY 1916-1918). O número e a forma dos dentes da abertura
são característicos da espécie (PILSBRY 1926).
Distribuição: do México à Argentina (PILSBRY 1926). Levada para o Havaí desde o
século XIX através do comércio (CHRISTENSEN & KIRCH 1986). Ceará (MORRETES 1949). Ilha
Grande, Angra dos Reis, Rio de Janeiro (HAAS 1953).

17
Família Bulimulidae
Bulimulus tenuissimus (d´Orbigny, 1835) (Fig. 9)
Localidade-tipo: Rio de Janeiro (PILSBRY 1897-1898). Espécie nativa, sinantrópica.
Concha umbilicada, oval-cônica, muito fina, transparente, pouco brilhante, decussada
pelo crescimento de estrias e linhas espirais delicadas; pálida córnea, algumas vezes fulvo.
Espira conóide, ápice bastante agudo, sutura simples. Seis voltas, ligeiramente convexas,
aumentando com velocidade moderada; a última volta convexa, não descendente na frente,
ligeiramente atenuada na base. Columela suboblíqua, algumas vezes quase vertical. Abertura
oval, perístoma simples, não expandido, agudo; margem columelar refletida na parte superior
(PILSBRY 1897-1898).
ARAÚJO et al. (1960) caracterizam a concha com comprimento total de 12 a 20 mm,
com 5 a 6 voltas da espira. Concha embrionária ornamentada em zig-zag na primeira volta da
espira e até a metade da segunda volta. O resto da concha apresenta apenas estrias de
crescimento mais visíveis nas suturas.
DUTRA (1988) observou que os indivíduos desta espécie ocorrem verticalmente nos
ramos e talos de ervas; eventualmente foram encontrados no solo, em locomoção sobre folhas
caídas; possuem excepcional habilidade de adaptação às condições ambientais.
SILVA et al. (2008) ao estudarem aspectos do desenvolvimento e reprodução em
laboratório, relataram que este molusco tropical, que é amplamente distribuído no Brasil, é a
Figura 8. Gastrocopta servilis (Gould, 1843). Imagem disponível em:
www.jaxshells.org/gast20.jpg; Imagem do Dr. Wayne Van Devender do
Departamento de Biologia, Appalachian State University, EUA.

18
única espécie de sua família com importância parasitológica. MEIRELES et al. (2008)
estudaram a influência da dieta e do isolamento no crescimento dos indivíduos desta espécie
em condições de laboratório, verificaram a influência da densidade populacional no
crescimento e que a dieta oferecida exerceu grande influência na biologia dos animais.
Distribuição: Natal, Maranhão e Pará (BAKER 1913). Rio de Janeiro, Bahia,
Pernambuco e Mato Grosso (MORRETES 1949). Pernambuco (DUTRA 1988). Espírito Santo e
São Paulo (SIMONE 2006).
Família Subulinidae
Beckianum beckianum (Pfeiffer, 1846) (Fig. 10)
A localidade-tipo desta espécie não foi encontrada, no material pesquisado. PILSBRY
(1906) relata que esta espécie está presente em várias partes do mundo. Segundo o autor,
algumas formas de Opeas (atualmente Beckianum e Lamellaxis), nenhuma delas bem
conhecidas, foram descritas a partir da Índia Ocidental, em geral são espécies bem
distribuídas, como: O. beckianum, O. gracilis, O. micra e O. goodalli. Por outro lado, COWIE
Figura 9. Bulimulus tenuissimus (d´Orbigny, 1835). Desenho
copiado de ARAÚJO et al. (1960).

19
(1998) afirma que a região de origem desta espécie é América Central e do Sul. Espécie
sinantrópica.
Concha umbilicada, oval-oblonga, ápice aguçado, fina, translúcida, hialina esverdeada,
densamente costelada e estriada. Nove voltas, convexa, a última sobre um quarto final do
comprimento total, base arredondada, columela curta. Abertura ampla, semioval; perístoma
simples, agudo, margem columelar estendida. (PFEIFFER 1846 apud PILSBRY 1906). Mede
cerca de 8 mm, tem 8 ½ voltas fortemente convexas, apresenta estriação (PILSBRY 1926).
Distribuição: Índia Ocidental PILSBRY (1906). Do México ao Brasil (PILSBRY 1926).
Pará, Rio de Janeiro e São Paulo (MORRETES 1949). Do México ao Rio de Janeiro (TILLIER
1980). Rio Grande do Norte e Roraima (SIMONE 2006). Ilha Grande, Angra dos Reis, Rio de
Janeiro (HAAS 1953), nas trilhas do Caxadaço e da Jararaca (SANTOS & MONTEIRO 2001), nas
trilhas do Pico do Papagaio e da Jararaca apenas em 100m de altitude (NUNES 2007).
Subulina octona (Bruguière, 1789) (Fig. 11)
Localidade-tipo: Guadalupe, Índias Ocidentais (SOLEM 1961). Antilhas (BIELER &
SLAPCINSKY 2000). THOMÉ et al. (2006) consideram a espécie nativa no Brasil tropical,
embora afirme que a localidade tipo e holótipo são desconhecidos. TILLIER (1980) afirma que
a espécie é introduzida em toda a região intertropical, sendo impossível determinar com
Figura 10. Beckianum beckianum. (Pfeiffer, 1846). Foto retirada de
SIMONE (2006), exemplar com 6 mm.

20
precisão seu local de origem. Por outro lado, COWIE (1998) relata o Neotrópico como região
de origem. SIMONE (2006) destaca esta espécie como introduzida e relata sua ocorrência na
África, mas com um ponto de interrogação. BESSA et al. (2000) afirmam que esta espécie é
encontrada em várias partes do mundo, sempre associadas a ambientes antrópicos. Pelo
exposto, é possível perceber que a origem desta espécie é muito controversa, embora a
maioria dos autores pesquisados indique a região Neotropical. Espécie sinantrópica.
ARAÚJO & BESSA (1993) caracterizam a concha como pequena, medindo entre 13 x
4mm, com espira muito longa, graças ao número de voltas que apresenta (oito). A amplitude
das voltas aumenta progressivamente até a volta corporal. Não apresenta nenhuma
ornamentação característica, evidenciando de forma não muito marcada, as linhas de
crescimento. A abertura é de forma ovalada sem dentes ou lamelas e com perístoma cortante.
O bordo columelar apresenta uma leve projeção no sentido do eixo, mais alargada em sua
metade. As suturas são evidentes, porém muito profundas. THOMÉ et al. (2006) caracterizam a
espécie da seguinte forma: concha alongada, amarelada, lisa, levemente estriada e com
protoconcha lisa.
É hospedeira intermediária de parasitos. BESSA et al. (2000) relataram a infecção por
Angiontrongylus vasorum (Baillet, 1866). Em laboratório, já foi verificada a influência do
substrato na reprodução (D’ÁVILA & BESSA 2005a, BESSA & ARAÚJO 1995b) e no crescimento
(D’ÁVILA & BESSA 2005b) desta espécie.
DUTRA (1988) relatou o encontro de indivíduos desta espécie no solo e também abaixo
do entrelaçado de ramos e raízes sobre o solo, segundo o autor, a espécie tem excepcional
habilidade de adaptação às condições ambientais. D’ÁVILA et al. (2006) verificaram a
presença de comportamento agregativo.
Segundo BESSA & ARAÚJO (1995a), são capazes de fazer autofecundação, o que
aumentaria seu sucesso em colonizar novos ambientes (SANTOS & MONTEIRO 2001).

21
D’ÁVILA et al. (2004) estudaram a resistência à dessecação de S. octona e verificaram
que, depois de 36 horas em câmara climatizada à 35ºC e 80% de umidade relativa do ar, todos
os indivíduos morreram, contudo, os indivíduos desta espécie podem se enterrar no substrato
para superar períodos desfavoráveis. Esse comportamento é muito frequente para esta espécie,
observado mesmo em períodos favoráveis, ele reduz o risco de dessecação já que o substrato
fornece a umidade necessária à sobrevivência destes moluscos e pode funcionar como uma
barreira física à evaporação de água corporal.
Figura 11. Subulina octona (Bruguière, 1789). Desenho
copiado de ARAÚJO & BESSA (1993).
Distribuição: Ceará e Pará (BAKER 1913). Do México ao
Brasil e Equador (PILSBRY 1926). Pernambuco, Rio de Janeiro
e Paraná (MORRETES 1949). Ilha Grande, Angra dos Reis, Rio
de Janeiro (HAAS 1953). Amapá, Amazonas, Pará, Ceará,
Bahia, Rio de Janeiro, Minas Gerais, São Paulo, Paraná, Rio
Grande do Sul e Rondônia (ARAÚJO & BESSA 1993). Rio
Grande do Sul (THOMÉ et al. 2006). Amplamente distribuída
em regiões tropicais (BIELER & SLAPCINSKY 2000). Sua
presença é assinalada em várias partes do mundo, vivendo em
jardins próximos aos domicílios humanos (BESSA et al.
2000).

22
Lamellaxis gracilis (Hutton, 1834) (Fig. 12)
Localidade-tipo: Mirzapur, Ceylon, Índia, China, Formosa e Japão (PILSBRY 1906).
Embora ROTH (1997) relate que a origem desta espécie é incerta, COWIE (1998) relata sua
origem como desconhecida, mas sugere que possa ser Neotropical. Espécie sinantrópica.
Concha dextrógira, heliciforme, nos indivíduos jovens e cônica-alongada com ápice
obtuso nos adultos, branca, translúcida; com duas a sete voltas moderadamente convexas; a
protoconcha é lisa com 1 ½ voltas e apresenta crenulações junto à sutura, observáveis apenas
ao Microscópio Eletrônico de Varredura; sutura profunda; a abertura é oval alongada, com
margem columelar lisa, reta e refletida junto à base e margem parietal lisa; o umbílico é
parcialmente encoberto pela margem columelar (C. OLIVEIRA 2002). A altura da abertura
ocupa cerca de um quarto da altura total da concha; espira elevada, lábio simples, perióstraco
translúcido, com menos brilho que os outros Subulinídeos (OLAZARRI 1986).
São capazes de fazer autofecundação (OLIVEIRA et al. 1971). São herbívoros, se
protegem embaixo de talos e folhas em épocas secas (OLAZARRI 1986). São encontrados em
ambientes muito variados, embaixo de pedras, enterrados entre raízes e em solos arenosos
(M IQUEL & PARENT 1996). Habitam a serapilheira de muitos lugares (ROTH 1997).
Distribuição: Regiões temperadas dos dois hemisférios, provavelmente introduzida
pelo comércio (OLAZARRI 1986, ROTH 1997, BIELER & SLAPCINSKY 2000). Na América tem
vasta distribuição: nas Antilhas, na América Central e na América do Sul. Pará e São Paulo
(MORRETES 1949). Regiões tropicais de ambos os hemisférios (PILSBRY 1926, BERG 1994).
Distribuição mundial (MIQUEL & PARENT 1996). CHRISTENSEN & KIRCH (1986) relatam sua
ocorrência no Havaí em período pré-histórico. Na Ilha Grande (HAAS 1953), ocorre na trilha
do Pico do Papagaio (400 e 500 m de altitude) e na trilha da Jararaca, em altitudes (100, 400 e
500m de altitude) (NUNES 2007).

23
Leptinaria unilamellata (d`Orbigny, 1835) (Fig. 13)
A localidade-tipo desta espécie não foi encontrada no material pesquisado. Segundo
PILSBRY (1906), Leptinaria é um gênero da América tropical amplamente distribuído,
principalmente na América do Sul. Espécie encontrada em ambientes antrópicos e também em
ambientes florestados, mas com densidade baixa.
Segundo ARAÚJO & KELLER (1993) a concha é pequena, com 4,5 mm e cerca de 6,5
voltas de espira, com a superfície mostrando as linhas de crescimento não muito marcadas,
suturas bem evidentes, mas não muito profundas. Volta corporal com comprimento
equivalente a pouco mais que as demais. Abertura ovalada com perístoma cortante, bordo não
refletido e desprovido de dentes ou lamelas no bordo externo. Bordo columelar com uma
lamela lisa, triangular, baixa e projetada para a abertura. Bordo parietal com uma lamela
transversal à abertura e situada a meia distância entre o término da lamela columelar e a
inserção da última volta.
Apresenta peristômio simples, lamela parietal e columela truncada desde o embrião
(PILSBRY 1906), não dando assim indicativo conquiliológico externo do seu grau de
desenvolvimento (DUTRA 1988).
Figura 12. Lamellaxis gracilis (Hutton, 1834). Escala 1 mm.
Foto: Amilcar B. Barbosa.

24
Considerada espécie cosmopolita, tem sua distribuição ampliada principalmente por
ação antrópica (DUTRA 1988). ALMEIDA & BESSA (2001b) estudaram o crescimento e a
reprodução desta espécie em laboratório e verificaram que ela pode fazer autofecundação.
ARAÚJO (1982) apud (DUTRA 1988) cita uma constante simpatria dessa espécie com
espécies de Opeas (Lamellaxis) e Subulina octona.
DUTRA (1988) estudou aspectos da ecologia e reprodução e observou que essa espécie
ocorre junto com Bulimulus tenuissimus e Sarassinula sp., mas raramente com Subulina
octona. Essa espécie habita o solo, sobre e sob talos e folhas caídas, abaixo do entrelaçado de
ramos e raízes; apresentou grande densidade populacional, superando as outras espécies de
moluscos terrestres associadas.
Estudos fisiológicos foram realizados por ALMEIDA & BESSA (2000) e verificaram que
a densidade populacional influenciou o crescimento em L. unilamellata.
D’ÁVILA et al. (2004) estudaram a resistência à dessecação de L. unilamellata e
verificaram que, depois de 36 horas em câmara climatizada à 35ºC e 80% de umidade relativa
do ar, todos os indivíduos morreram, contudo, os indivíduos desta espécie podem se enterrar
para superar períodos desfavoráveis.
L. unilamellata é hospedeiro intermediário de Postharmostomum gallinum Witenberg,
1923 (DUARTE 1980 apud ALMEIDA & BESSA 2000) e de Paratanaisia bragai Santos, 1934
(ARAÚJO & KELLER 1993).
Distribuição: Peru (HAAS 1949). Pernambuco (DUTRA 1988). Bahia, Curitiba, Paraná
(MORRETES 1949). Mato Grosso, Amazonas, Roraima, Minas Gerais, Bahia, Rio de Janeiro,
São Paulo e Paraná ARAÚJO (1982) apud DUTRA (1988). Amazonas, Pará, Pernambuco,
Bahia, Rio de Janeiro, Minas Gerais, São Paulo, Mato Grosso, Rondônia (ARAÚJO & KELLER
1993). Ilha Grande, Angra dos Reis, Rio de Janeiro (HAAS 1953), na trilha do Pico do

25
Papagaio nas altitudes de 100, 300 e 500m, e trilha da Jararaca a 200m de altitude (NUNES
2007).
Obeliscus Beck, 1837
Concha sem umbílico, subcilíndrica ou afunilada, usualmente grande, sólida e opaca;
de nove a 18 voltas, um pouco convexa, não carenada. Voltas embrionárias suaves ápice
obtuso. Abertura oval, lábio simples e curto, columela côncava ou vertical, truncada na base, a
protoconcha é globosa ou oblonga, com cerca de três voltas, e muito pequena se comparada
com um exemplar adulto; são animais vivíparos (PILSBRY 1906-1907).
Distribuição: América do Sul e Antilhas (PILSBRY 1906-1907).
Os três gêneros a seguir são da família Systrophiidae, que é endêmica da Região
Neotropical (RAMÍREZ 1993).
Figura 13. Leptinaria unilamellata (d’Orbigny, 1835).
Desenho retirado de ARAÚJO & KELLER (1993).

26
Família Systrophiidae
Miradiscops Baker, 1925
Concha subdiscoidal, pequena, hialina, umbílico aberto, linhas de crescimento
fracamente marcadas, espira minimamente levantada, voltas com incremento lento, a última
volta não é maior que a anterior; sem dentes; a forma da última volta é arredondada,
perístoma afiado; dente radular em forma de garra, dente central vestigial ou ausente, dente
com cúspides longas e delgadas (RAMÍREZ 1993).
Distribuição: Bolívia, Brasil, Peru, Venezuela, Costa Rica, Guatemala, Panamá,
México, Porto Rico, Providence e Trinidade (RAMÍREZ 1993).
Happiella Baker, 1925
Concha com espira muito baixa e umbílico reduzido a uma pequena perfuração
(BAKER 1925). Concha sem carena no umbílico, dentes central e laterais fimbriados (RAMÍREZ
1993).
Distribuição: da Colômbia até as Guianas, Brasil, Paraguai e Trinidad (RAMÍREZ
1993).
Tamayoa Baker, 1925
Concha com carena no umbílico, estendendo até o ápice; pequeno dente central
radular, tricúspide (RAMÍREZ 1993).
Distribuição: Peru, Brasil, Venezuela e Trinidade (RAMÍREZ 1993).
Família Charopidae
Ptychodon Ancey, 1888
Concha pequena, aproximadamente três milímetros de diâmetro máximo e altura de
dois milímetros, deprimida, discoidal, com espira às vezes praticamente oculta pela última

27
volta, de cor ambarina, parede muito fina e opaca ou semitransparente, coberta por perióstraco
geralmente fino e brilhoso. Apresenta até cinco voltas e meia, convexas, de crescimento lento,
regular e levemente comprimidas. A protoconcha é lisa. As voltas pós-embrionárias são
dotadas de costelas radiais ripiformes, finas, convexas e baixas de traçado perfeitamente
sigmóide. Os espaços intercostelares apresentam linhas de crescimento, podendo ocorrer
também estrias. A abertura é reniforme, comprimida palatalmente, com perístomo de borda
simples e cortante. O umbílico tem cerca de 1/3 a 1/4 do diâmetro máximo da concha, é amplo
e perspectivo. Apresenta barreiras peristomais (FONSECA & THOMÉ 1993).
Ptychodon schuppi (Suter, 1900) (Fig. 14)
Localidade-tipo: São Leopoldo, Rio Grande do Sul (SUTER 1900). Espécie nativa,
encontrada em ambientes florestados.
Concha exígua, discoidea, umbilicada, estriada, não lustrosa, fulva; espira quase plana,
de 4 voltas, aumentando paulatina e regularmente, convexas, com costelas finas, direitas
equidistantes, em número mais ou menos de 14 por milímetro; entre estas existem linhas
microscópicas de incremento, cruzadas por linhas espirais. A protoconcha é lisa de 1 ½ voltas.
A sutura é muito profunda, a abertura subvertical, estreita e redondamente lunar com margens
simples, e extremidades distantes, convergentes. O lábio columelar é curto, vertical, não
reflexo, o umbigo bem largo, perspectivo, mostrando todas as voltas. A base é convexa
(SUTER 1900).
Distribuição: Rio Grande do Sul (SUTER 1900). Ilha Grande, Angra dos Reis, Rio de
Janeiro, nas trilhas da Jararaca e do Caxadaço (SANTOS & MONTEIRO 2001), na trilha da
Jararaca nos 300 e 500 m de altitude e na trilha do Pico do Papagaio de 200 a 300 m de
altitude (NUNES 2007).

28
Stephanoda Albers, 1860
Concha deprimida, de paredes finas, espira baixa e plana, até sete voltas convexas de
sutura profunda e não escavada, abertura reniforme e obliqua de perístomo simples, umbílico
muito amplo (ALBERS 1860 apud FONSECA & THOMÉ 1993).
Família Euconulidae
Habroconus semenlini (Moricand, 1846) (Fig. 15)
Localidade-tipo: Bahia (MORICAND 1846). Espécie nativa encontrada em ambientes
antrópicos e florestados.
Micromolusco de concha dextrógira, discóide, frágil; amarela, translúcida; número de
voltas variando de 2 ½ a 4 ½; sutura pouco profunda; a protoconcha com 1 ½ voltas
apresenta linhas finas, espirais e contínuas, que se estendem por toda a superfície da concha e
aumentam o espaçamento entre as mesmas à medida que a concha cresce; em vista dorsal e
ventral são em número de 19 a 20 linhas na volta do corpo; abertura em formato de meia lua;
perístomo fino; umbílico oval encoberto parcialmente ou totalmente (C. OLIVEIRA 2002). A
concha vista pelo microscópio é claramente decussada. A protoconcha apresenta miúdas
estrias radiais, tendo linhas espirais visíveis apenas nas proximidades da sutura. A perfuração
umbilical é muito estreita e quase completamente coberta pela dobra columelar (SUTER 1900).
Figura 14. Ptychodon schuppi (Suter, 1900). Foto: Carla
W. Gabriel.

29
Distribuição: Uruguai, Argentina, Paraguai, São Paulo, Paraná, Rio Grande do Sul (C.
OLIVEIRA 2002). São Paulo (SUTER 1900). Bahia (MORICAND 1846). Na Ilha Grande, Angra
dos Reis, Rio de Janeiro (HAAS 1953), na trilha da Jararaca de 100 a 500 m de altitude e na
trilha do Pico do Papagaio de 200 a 400 m de altitude (NUNES 2007).
Família Bradybaenidae
Bradybaena similaris (Férussac, 1821) (Fig. 16)
Localidade-tipo: Timor (Indonésia) (SOLEM 1961). Várias localidades (BIELER &
SLAPCINSKY 2000). COWIE (1998) relata sua origem na Ásia. THOMÉ et al. (2006) afirmam
que esta espécie possui localidade tipo e holótipo desconhecidos. BURCH (1962) e BERG
(1994) são mais precisos, afirmando que é nativa da China, a partir de onde se expandiu pelo
comércio a todas as partes do mundo onde houve cultivo de café, podendo ser uma praga
grave para a floricultura e horticultura (BOFFI 1979, THOMÉ et al. 2006). Espécie sinantrópica,
encontrada exclusivamente em ambientes modificados, esta espécie não é encontrada em
ambientes florestados.
A concha mede de 9 a 13 mm de diâmetro, sub-heliciforme, com perfil das voltas
convexo, base arredondada, translúcida e lisa. Cor castanho clara, com ou sem uma faixa
espiral castanha na altura da sutura e no seu prolongamento imaginário pela volta do corpo;
lábio e interior da abertura brancos e brilhantes, região parietal castanha. Abertura luniforme,
lábio refletido e umbílico estreito. Escultura de estrias espirais; linhas de crescimento finas e
oblíquas (BOFFI 1979).
Figura 15. Habroconus semenlini (Moricand, 1846). Escala 1
mm. Foto: Antônio C. Freitas.

30
De acordo com ARAÚJO (1989), a concha tem aspecto geral heliciforme, com parede
espessa e forte, constituída de cinco a seis voltas da espira. Mostra duas suturas evidentes
porém não muito profundas, acompanhadas no seu limite superior por uma linha de coloração
castanha que se estende na volta corporal até o bordo da abertura e é visível também pela face
interna da concha. As linhas de crescimento são mais pronunciadas na volta corporal, dando
um aspecto levemente rugoso à superfície da concha. O umbílico é aberto, com os bordos se
abrindo amplamente sem, contudo, ser muito profundo. A abertura é semilunar, com os
bordos refletidos largamente, sem lamelas ou dentes.
THOMÉ et al. (2006) caracterizam a espécie como pequeno caracol herbívoro, com
concha arredondada, castanho clara, em geral, apresenta uma banda espiral avermelhada.
Espécie invasora, provavelmente asiática, comum em hortas e jardins.
ALMEIDA & BESSA (2001a) estudaram o crescimento e a reprodução desta espécie em
laboratório, verificando que é capaz de fazer autofecundação. Pode ser infectada por
Postharmostum gallinum Witenberg, 1923 e Eurytrema coelomaticum (Giard & Biliet, 1892),
ambos com ciclos de vida associados a animais domésticos (AMATO & BEZERRA 1992).
D’ÁVILA et al. (2004) relataram a formação de epifragma em alguns indivíduos e uma
grande resistência à dessecação, o que poderia ser explicado por seu tropismo positivo por
superfícies verticais sendo, então, menos dependente do substrato e possivelmente tem menor
exposição à luz e ao calor.
No Brasil, alguns trabalhos já foram realizados com a espécie, abordando reprodução,
fisiologia e parasitologia, por exemplo: AMATO & BEZERRA (1992), PASCHOAL & AMATO
(1996), ALMEIDA & BESSA (2000), LIRA et al. (2000), BRANDOLINI & AMATO (2001),
PINHEIRO et al. (2001), MOREIRA et al. (2003), FURTADO et al. (2004), ARÉVALO et al. (2006),
NASCIMENTO et al. (2006), GARCIA & PINHEIRO (2007), LUSTRINO et al. (2008) e DE OLIVEIRA
et al. (2008).

31
Distribuição: Um dos moluscos mais amplamente distribuídos pelo mundo (SOLEM
1961). HAAS (1955) relatou que esta espécie oriental estava rapidamente se espalhando no
Peru. Curitiba, Paraná, São Paulo, Rio de Janeiro (MORRETES 1949). Cincuntropical,
introduzida nas Américas (BOFFI 1979). É encontrada em quase todos os estados do Brasil
(ARAÚJO 1989). Rio Grande do Sul (THOMÉ et al. 2006). Encontra-se distribuído
principalmente nas regiões tropicais, disseminado pelo comércio de plantas; ocorre na
América do Norte, Central e do Sul; no Brasil, é encontrado desde o Amapá até o Rio Grande
do Sul (ALMEIDA & BESSA 2001a).
Área urbana da Vila Dois Rios
Na área urbana foram coletados 271 indivíduos, distribuídos em cinco famílias e oito
espécies (Tab. I) (Col. Mol. UERJ Nº 7691-7723). Destas, uma espécie é nativa, cinco
espécies são exóticas, uma espécie não conseguimos localizar a origem, mas todas estão
sempre associadas a ambientes antrópicos.
A família dominante foi Subulinidae com 91,50 % do total coletado, seguido por
Bulimulidae (5,54%), Fruticolicidae (1,48%), Succineidae (1,11%) e Pupillidae (0,37%),
como mostra a figura 17.
Figura 16. Bradybaena similaris (Férussac,
1821). Desenho retirado de ARAÚJO (1989).

32
Famílias
Subulinidae
Bulimulidae
Fruticicolidae
Succineidae
Pupillidae
Figura 17. Representação das famílias de moluscos terrestres coletadas na área urbana da Vila Dois Rios, Ilha
Grande, Angra dos Reis, Rio de Janeiro.
Podemos observar que nesta comunidade existe a dominância da espécie Beckianum
beckianum (Pfeiffer, 1846), com 67% de exemplares (Fig. 18). Embora a origem geográfica
desta espécie seja desconhecida, vários autores relatam a ocorrência dela em diversas partes
do mundo, sempre associadas a ambientes modificados pelo homem, seja pela agricultura, por
canteiros de hortas ou jardins. Esta espécie apresenta elevada abundância em ambientes
antrópicos, mas em ambientes onde a influência humana foi muito tênue, ela não é
encontrada.

33
0
20
40
60
80
100
120
140
160
180
200
Abu
ndân
cia
B. bec
kianu
m
S. octo
na
L. gr
acilis
B. ten
uissim
us
L. un
ilam
ellat
a
B. sim
ilaris
S. mer
idion
alis
G. ser
vilis
Figura 18. Abundância dos moluscos terrestres coletados na Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de
Janeiro.
Encosta da Vila Dois Rios a 350 metros de altitude
Na encosta da Vila Dois Rios, a 350 m de altitude, foram coletados 23 indivíduos, de
nove espécies diferentes, compreendidos em seis famílias (Tab. I) (Col. Mol. UERJ Nº 7677-
7690, 7724).
A família Systrophiidae (61%), endêmica do neotrópico (RAMÍREZ 1993), foi
dominante, seguida respectivamente pelas famílias Subulinidae (13%), Euconulidae e
Charopidae (8, 7% cada) e Helicinidae e Neocyclotidae (4,3% cada) (Fig. 19).

34
Famílias
Helicinidae
Neocyclotidae
Subulinidae
Systrophiidae
Charopidae
Euconulidae
Figura 19. Representação das famílias de moluscos terrestres coletadas na encosta da Vila Dois Rios, Ilha
Grande, Angra dos Reis, Rio de Janeiro, a 350 metros de altitude.
Observando a figura 20, percebemos que a espécie dominante foi Happiella sp,
seguida por Obeliscus sp., Miradiscops sp. e H. semenlini. As espécies Alcadia sp., N.
prominulus, L. unilamellata, Ptychodon sp.1 e P. schuppi foram coletados com apenas um
indivíduo cada, representam espécies singletons que refletem a raridade e/ou a pequena
abundância da maioria das espécies dentro de uma comunidade, padrão comumente
observado, principalmente em regiões tropicais (WHITTAKER 1975, MAGURRAN 1988, ODUM
1988, KAGEYAMA & L EPSCH-CUNHA 2001, RICKLEFS 2003).
Das espécies coletadas, quatro foram identificadas até a categoria específica, três das
quais possuem localidade tipo no país e uma com localidade tipo descrita para a América
Tropical.
As espécies Obeliscus sp., H. semenlini, Alcadia sp. e N. prominulus foram exclusivas
desta localidade.
Famílias

35
Figura 20. Abundância das espécies coletadas na encosta da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de
Janeiro, a 350 m de altitude.
3) Encosta da Vila Dois Rios a 450 metros de altitude
Na coleta quantitativa da encosta da Vila Dois Rios, a 450 m de altitude, foram
coletados 17 indivíduos, divididos em oito espécies diferentes, e três famílias (Tab. I) (Col.
Mol. UERJ Nº 7662-7669, 7671-7674).
A família Systrophiidae foi dominante, com 53% do total de indivíduos coletados,
seguido pela família Charopidae, que representou 41% do total de indivíduos.

36
Famílias
Bulimulidae
Systrophiidae
Charopidae
Figura 21. Representação das famílias de moluscos terrestres coletadas na encosta da Vila Dois Rios, Ilha
Grande, Angra dos Reis, Rio de Janeiro, a 450 metros de altitude.
Observando a figura 22, percebemos que a espécie dominante foi Miradiscops sp.,
seguida por Happiella sp. Das espécies coletadas, apenas uma foi identificada até a categoria
específica, sendo ela nativa.
As espécies Bulimulidae sp.3, Tamayoa sp., Ptychodon sp.2 e Stephanoda sp. foram
exclusivas desta localidade.
Figura 22. Abundância das espécies coletadas na encosta da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de
Janeiro, a 450 m de altitude.

37
Na figura 23 é possível observar que nas amostras coletadas existem espécies
exclusivas da área urbana da Vila Dois Rios: Beckianum beckianum, Subulina octona,
Lamellaxis gracilis, Bulimulus tenuissimus, Bradybaena similaris, Succinea meridionalis,
Gastrocopta servilis. A espécie Leptinaria unilamellata, também foi coletada na área de
floresta a 350 m de altitude, apenas um indivíduo.
0%
20%
40%
60%
80%
100%
B. bec
kianu
m
S. octo
na
L. gracil
is
B. te
nuiss
imus
L. unil
amell
ata
B. si
mila
ris
S. m
eridi
onali
s
G. servil
is
Obeliscu
s sp.
H. se
men
lini
Alca
dia sp
N. prominu
lus
Happ
iella
sp.
P. sc
hupp
i
Ptyc
hodo
n sp
.1
Mira
disco
ps sp.
Bulim
ilidae
sp. 3
Tamay
oa sp
.
Ptycho
don sp
.2
Step
hano
da sp
.
450m
350m
0m
Figura 23. Percentual de espécies coletas na Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro. 0m:
área urbana; 350m: encosta a 350m de altitude; 450m: encosta a 450m de altitude.
De acordo com a nossa experiência na região (NUNES et al. 2003) observamos que
algumas espécies são encontradas somente em ambientes não florestados, fortemente
influenciados pelo homem, tais como gramados, jardins e hortas: Bulimulus tenuissimus,
Bradybaena similaris, Succinea meridionalis e Gastrocopta servilis, estas espécies não são
encontradas onde a vegetação está se estabelecendo, apresentando um dossel razoavelmente
fechado, constituído por árvores e arbustos de várias alturas. Contudo, outras espécies
conseguem viver tanto no ambiente antrópico, bem alterado, e também em ambientes onde a
influencia antrópica cessou e o ambiente está se recuperando, são elas B. beckianum, S.
octona, L. gracilis e L. unilamellata. A espécie B. beckianum, em ambientes florestados em

38
recuperação, também apresenta abundância e dominância com valores bastante elevados
(NUNES et al. 2003).
Nas regiões florestadas da Ilha Grande, onde a influência antrópica se restringiu a
entrada do homem para realizar cortes de madeira, ou realizar caça, por exemplo,
eventualmente encontramos um ou dois exemplares de Lamellaxis gracilis e Leptinaria
unilamellata.
As espécies do ambiente florestado, provavelmente são mais sensíveis a variações dos
fatores ambientais, por isso não são encontradas em ambientes antrópicos, onde existe uma
grande variação de condições ambientais ao longo do dia.
Dados ambientais
Podemos considerar que existe um gradiente ambiental, que vai desde a região mais
alterada, a área urbana da Vila Dois Rios, para a região menos alterada, a encosta da Vila
Dois Rios a 450 m de altitude, passando pela região intermediária, a encosta da Vila Dois
Rios a 350 m de altitude. Visualmente é fácil perceber este gradiente e os dados ambientais
coletados corroboram a existência deste gradiente.

39
Tmax Tmed Tmin Tsolo
1 2 3
Área
16
18
20
22
24
26
28
30
32T
empe
ratu
ra (
ºC)
Figura 24. Temperatura ambiente e do solo (ºC) da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro.
1: área urbana; 2: encosta a 350m de altitude; 3: encosta a 450m de altitude.
A área urbana apresentou temperaturas mais elevadas que as outras áreas, o que é
justificado pelo fato desta área ser aberta e, portanto, receber forte incidência luminosa do sol.
A região florestada a 450 m de altitude possui temperaturas mais baixas devido ao seu dossel
que é mais fechado quando comparada com a região de 350 m de altitude. A ausência de
cobertura vegetal favorece a elevação da temperatura na área urbana da Vila Dois Rios (Fig.
24). A diminuição de temperatura de acordo com o aumento da altitude é um evento bem
conhecido (RICKLEFS 2003, BEGON et al. 2006).

40
Umax Umed Umin
1 2 3
Área
40
45
50
55
60
65
70
75
80
85
Um
idad
e am
bien
te (
%)
Figura 25. Percentual de umidade ambiente da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro. 1:
área urbana; 2: encosta a 350m de altitude; 3: encosta a 450m de altitude.
A umidade foi menor na área urbana da Vila Dois Rios, devido principalmente a sua
elevada temperatura, que associada à estrutura da vegetação, constituída por algumas árvores
de grande porte e muitas plantas herbáceas, principalmente gramíneas, o que não contribui
para reter a umidade no local. Na região florestada, a umidade média aumentou conforme o
gradiente ambiental e altitudinal, mas a umidade mínima e máxima reduziu com a elevação da
altitude (Fig. 25). A diminuição de umidade de acordo com o aumento da altitude já foi relata
na literatura (RICKLEFS 2003, BEGON et al. 2006).

41
1 2 3
Área
-4000
-2000
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
20000
Lum
inos
idad
e (L
ux)
Figura 26. Luminosidade (Lux) da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro. 1: área urbana;
2: encosta a 350m de altitude; 3: encosta a 450m de altitude.
A luminosidade na área urbana da Vila Dois Rios teve uma enorme variação, e
apresentou valores muito elevados, pelos mesmos motivos apresentados anteriormente. A
localidade de encosta a 350m de altitude apresentou valores de luminosidade mais baixos e
mais homogêneos, embora o esperado fosse que a área de 450 m de altitude apresentasse
valores comparativamente mais baixos que as outras localidades (Fig. 26). Mas se tratando de
uma região íngreme, a vegetação apresenta várias alturas de dossel, que associado ao ângulo
do sol, poderia ocasionar um aumento de possibilidades de entrada de luz solar, aumentando
assim a luminosidade no local.

42
1 2 3
Área
0
1
2
3
4
5
6
7
Pro
fund
idad
e da
ser
apilh
eira
(cm
)
Figura 27. Profundidade da serapilheira (cm) da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de Janeiro. 1:
área urbana; 2: encosta a 350m de altitude; 3: encosta a 450m de altitude.
A profundidade da serapilheira aumentou conforme o gradiente ambiental (Fig. 27). O
mesmo ocorreu para o percentual de umidade de folhas da serapilheira (Fig. 28). Quanto
maior for a profundidade do folhiço, maior são as suas chances de reter umidade nas folhas da
serapilheira. Por outro lado, o percentual de umidade de galhos da serapilheira não apresentou
valores que diferenciem as localidades de maneira significativa (p = 0,057) (Tab. II).

43
Folha Galho
1 2 3
Área
0
10
20
30
40
50
60
70
80
90
Um
idad
e da
ser
apilh
eira
(%
)
Figura 28. Percentual de umidade da serapilheira. da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de
Janeiro. 1: área urbana; 2: encosta a 350m de altitude; 3: encosta a 450m de altitude.
Na análise de variância, com exceção do percentual de umidade de galhos presentes na
serapilheira, todos os outros dados ambientais quantificados diferiram estatisticamente entre
as três localidades (Tab. II). Em relação ao teste Tukey, para os dados ambientais que na
análise de variância diferiram significativamente, a área urbana da Vila Dois Rios difere
estatisticamente das outras duas localidades florestadas da encosta. Em contrapartida, essas
áreas florestadas de encosta são mais semelhantes entre si, já que quase todos os dados
ambientais não foram estatisticamente diferentes para essas localidades, apenas a temperatura
do solo e umidade relativa mínima do ar diferiram significativamente essas duas localidades.
De forma simplificada, os dados ambientais indicam que as duas localidades de
encosta da Vila Dois Rios (350 e 450 m de altitude) são mais semelhantes entre si e
completamente diferentes da área urbana da Vila Dois Rios.

44
Tabela II: Análise de Variância com os dados ambientais da Vila Dois Rios, Ilha Grande, Angra dos Reis, Rio de
Janeiro e teste Tukey. 1: área urbana; 2: encosta a 350m de altitude; 3: encosta a 450m de altitude.Valores em
negrito indicam p < 0,05; ou seja, diferença significativa entre as localidades consideradas.
Dados ambientais Anova teste Tukey
1 X 2 1 X 3 2 X 3
Temperatura máxima do ambiente 0,000 0,000 0,000 0,056
Temperatura média do ambiente 0,000 0,000 0,000 0,079
Temperatura mínima do ambiente 0,000 0,000 0,000 0,061
Temperatura do solo 0,000 0,000 0,000 0,000
Umidade relativa máxima do ar 0,000 0,000 0,000 0,065
Umidade relativa média do ar 0,000 0,000 0,000 0,130
Umidade relativa mínima do ar 0,000 0,000 0,000 0,001
Luminosidade 0,001 0,000 0,023 0,266
Profundidade da serapilheira 0,000 0,000 0,000 0,268
Umidade de folhas da serapilheira 0,001 0,009 0,002 0,589
Umidade de galhos da serapilheira 0,507 0,706 0,915 0,517
A composição encontrada na comunidade da área urbana é totalmente diferente da
comunidade dos ambientes florestados de encosta, o que também ocorre para as condições
ambientais que são totalmente diferentes. A Vila Dois Rios apresenta elevada intensidade
luminosa, temperatura ambiente e do solo; baixa umidade do ar e pouca serapilheira,
condições completamente opostas às encontradas no ambiente florestado. Como as condições
ambientais se apresentam segundo um gradiente, no qual a luminosidade e a temperatura do
ambiente e do solo diminuem; e a umidade do ambiente, a profundidade e a umidade da
serapilheira aumenta, conforme passamos de uma área completamente alterada, para a floresta
secundária e depois para a floresta primária, a comunidade de moluscos terrestres responde a
este gradiente através de sua composição (Fig. 23).
A fauna urbana atual da Vila Dois Rios é constituída apenas por espécies
sinantrópicas, as quais em geral, possuem ampla distribuição geográfica, mas estão associadas

45
a ambientes modificados pela ação do homem. A introdução dessas espécies provavelmente
se deu pela ocupação humana e pela modificação do hábitat, principalmente pela agricultura.
A introdução de plantas ornamentais e para o cultivo através de mudas, permitiu a entrada na
Ilha Grande de ovos e indivíduos de moluscos presentes na terra das mudas de plantas, HAAS
(1953) já tinha citado a ocorrência destas espécies na Ilha Grande, elas devem estar nesta ilha
desde a época das lavouras de café, na primeira metade do século XIX.
É interessante observar que muitas destas espécies sinantrópicas ficam restritas ao
ambiente antrópico, como jardins, hortas e gramados. Enquanto que algumas outras espécies
sinantrópicas conseguem estabelecer-se também em um ambiente que está iniciando seu
processo de recuperação.
Observamos uma tendência de que o ambiente uma vez modificado, a paisagem
consegue se recuperar, mas durante este processo, a malacofauna modifica sua estrutura, com
perda de espécies nativas e introdução de espécies sinantrópicas. Com o passar do tempo e o
fim da influência humana no ambiente, a malacofauna nativa consegue se restabelecer, e as
espécies sinantrópicas vão aos poucos desaparecendo, por isso a abundância delas é reduzida
em ambientes intermediários.
Tendo os moluscos terrestres características que os tornam potencialmente excelentes
bioindicadores (SCHILTHUIZEN & RUTJES 2001) e tendo como base tudo que aqui foi exposto,
queremos propor um índice de diagnóstico de alteração/degradação ambiental.

46
Tabela III. Sugestão para diagnóstico de ambientes de Floresta Atlântica degradados/ recuperados, utilizando a
proporção de espécies de moluscos sinantrópicas e nativas, para cada estágio de recuperação.
Ambiente
urbano
Floresta
intermediária
Floresta climáxica
(OLIVEIRA 2002)
Tempo de recuperação (anos) 0 anos 35 anos Mais de 50 anos sem
interferência antrópica
Riqueza e abundância das
espécies sinantrópicas
90 a 100 % 50 a 60 % 0 a 10 %
Riqueza e abundância das
espécies nativas
0 a 10 % 40 a 50 % 90 a 100 %
Os moluscos são animais extremamente sensíveis a mudanças no micro-habitat e
podem ser bons indicadores de alteração ambiental, por isso, o estudo comparativo de
comunidades sinantrópicas e nativas, juntamente com suas tolerâncias ambientais foi
necessário e permitiu estabelecer uma metodologia rápida, de pouco impacto e principalmente
de baixo custo para classificar ou diagnosticar o estado de conservação ou modificação de
fragmentos florestais na Ilha Grande. Estes dados poderão ser utilizados para estudos de
conservação e manejo da vida silvestre. Contudo, mais estudos utilizando gradientes
ambientais em outras localidades devem ser realizados com a finalidade de estabelecer
proporções de espécies sinantrópicas/nativas para cada fase de regeneração florestal em
diferentes ambientes.

47
CONCLUSÕES
A comunidade de moluscos terrestres retrata a alteração ambiental da Vila Dois Rios,
Ilha Grande, Angra dos Reis, Rio de Janeiro.
Com a reestruturação da floresta, a composição da malacofauna vai se modificando,
com a redução do númeor de espécies sinantrópicas e aumento do número de espécies nativas.
A realização de mais estudos deste tipo é necessária e permitirá estabelecer intervalos
mais precisos para diagnósticos de fragmentos de Floresta Atlântica usando uma metodologia
rápida e barata.
Esta metodologia pode auxiliar estudos de escolha de áreas prioritárias para
conservação de ambientes de Floresta Atlântica.

48
REFERÊNCIAS BIBLIOGRÁFICAS
ALBERS, J.C. 1860. Die Heliceen nach natürlicher Verwandtschaft systematisch
gerodnet. 2. Ed. Leipzig: W. Engelmann. p.80-98.
ALHO, C.J.R.; M. SCHNEIDER & L.A. VASCONCELLOS. 2002. Degree of threat to the biological
diversity in the Ilha Grande State Park (RJ) and guidelines for conservation. Brazilian
Journal of Biology 62 (3): 375-385.
ALMEIDA , M.N. & E.C.A. BESSA. 2000. Efeito da densidade populacional sobre o crescimento
e a reprodução de Bradybaena similaris (Férussac, 1821) (Mollusca, Xanthonychidae) e
Leptinaria unilamellata (Mollusca, Subulinidae). Revista Brasileira de Zoociências 2
(2): 97-104.
ALMEIDA , M.N. & E.C.A. BESSA. 2001a. Estudo do crescimento e da reprodução de
Bradybaena similaris (Férussac) (Mollusca, Xanthonychidae) em laboratório. Revista
Brasileira de Zoologia 18 (4): 1115-1122.
ALMEIDA , M.N. & E.C.A. BESSA. 2001b. Estudo do crescimento e da reprodução de
Leptinaria unilamellata (d’Orbigny) (Mollusca, Subulinidae) em laboratório. Revista
Brasileira de Zoologia 18 (4): 1107-1113.
ALVES, S.L.; A.S. ZÁU, R.R. OLIVEIRA , D.F. LIMA & C.J.R. MOURA. 2005. Sucessão florestal e
grupos ecológicos em Floresta Atlântica de encosta, Ilha Grande, Angra dos Reis/ RJ.
Revista Universidade Rural, Série Ciências da Vida 25 (1): 26-32.
AMATO, S.B. & J.C.B. BEZERRA. 1992. Concurrent infection of Postharmostum gallinum
(Digenea, Brachylaimidae) and Eurytrema coelomaticum (Digenea, Dicrocoeliidae in
Bradybaena similaris (Stylommatophora, Xanthonichidae). Memórias do Instituto
Oswaldo Cruz 87 (2): 309-311.
ARAÚJO, J.L.B. 1982. Alguns moluscos terrestres como hospedeiros intermediários de
parasitas de animais domésticos, no Brasil: estudos sobre a anatomia, sistemática e

49
participação em helmintoses. Itaguaí, 103 pp. Tese de Doutorado, Universidade Federal
Rural do Rio de Janeiro.
ARAÚJO, J.L.B. 1989. Moluscos de importância no Brasil. I. Xanthonychidae: Bradybaena
similaris (Férussac, 1821). (Mollusca, Gastropoda, Pulmonata, Stylommatophora).
Revista Brasileira de Zoologia 6 (4): 583- 592.
ARAÚJO, J.L.B. & D.G. KELLER. 1993. Moluscos de importância econômica no Brasil. III.
Subulinidae, Leptinaria unilamellata (Orbigny) (Mollusca, Gastropoda, Pulmonata,
Stylommatophora). Revista Brasileira de Zoologia 10 (3): 499-507.
ARAÚJO, J.L.B. & E.C.A. BESSA. 1993. Moluscos de importância econômica no Brasil. II.
Subulinidae, Subulina octona (Bruguiére) (Mollusca, Gastropoda, Pulmonata,
Stylommatophora). Revista Brasileira de Zoologia 10 (3): 489-497.
ARAÚJO, J.L.B.; H.E.B. RESENDE & P.A.F. RODRIGUES. 1960. Sobre “Bulimulus tenuissimus”
(Orbigny, 1835) (Gastropoda, Pulmonata). Revista Brasileira de Biologia 20 (1): 33- 42.
ARÉVALO, E.G; F.O. JUNQUEIRA; G.L.G. SOARES & E.C.A. BESSA. 2006. Atividade
fagoinibidora do ácido salicílico sobre Bradybaena similaris (Férussac, 1821) (Mollusca,
Bradybaenidae) em condições de laboratório. Revista Brasileira de Zoociências 8 (2):
111- 115.
AUBRY, S.; F. MAGNIN; V. BONNET & R. C. PREECE. 2005. Multi-scale altitudinal patterns in
species richness of land snail communities in south-eastern France. Journal of
Biogeography 32: 985-998.
BAKER, F. 1913. The land and fresh-water mollusks of the Stanford Expedition to Brazil.
Proceedings of the Academy of Natural Sciences of Philadelphia 65: 618-675.
BAKER, H.B. 1925. Agnathomorphous Aulacopoda. Nautilus 38: 86-89.
BARKER, G.M. 1982. Notes on the introduced terrestrial Pulmonata (Gastropoda: Mollusca) of
New Zealand. Journal of Molluscan Studies 48: 174- 181.

50
BARKER, G.M. & P.C. MAYHILL . 1999. Patterns of diversity and habitat relationships in
terrestrial mollusc communities of the Pukeamaru Ecological District, northeastern New
Zealand. Journal of Biogeography 26: 215-238.
BEGON, M.; C.R. TOWNSEND & J.L. HARPER. 2006. Ecology: from individuals to
ecosystems. Oxford, Blackwell Publishing. XII+738 p.
BERG, G.H. 1994. Caracoles y babosas de importância cuarentenaria, agrícola y medica
para America Latina y el Caribe. San Salvador, Organismo Internacional Regional de
Sanidad Agropecuaria, 132p.
BESSA, E.C.A. & J.L.B. ARAÚJO. 1995a. Ocorrência de autofecundação em Subulina octona
(Bruguiére) (Pulmonata, Subulinidae) sob condições de laboratório. Revista Brasileira de
Zoologia 12 (3): 719-723.
BESSA, E.C.A. & J.L.B. ARAÚJO. 1995b. Oviposição, tamanho de ovos e medida do
comprimento da concha em diferentes fases do desenvolvimento de Subulina octona
(Bruguiére) (Pulmonata, Subulinidae) em condições de laboratório. Revista Brasileira de
Zoologia 12 (3): 647-654.
BESSA, E.C.A.; W.S. LIMA ; E. DAEMON; M.C. CURY & J.L.B. ARAÚJO. 2000.
Desenvolvimento biológico de Angiostrongylus vasorum (Baillet) Kamensnky
(Nematoda, Angiostrongylidae) em Subulina octona (Bruguière) (Molusca, Subulinidae)
em condições de laboratório. Revista Brasileira de Zoologia 17 (1): 29-41.
BIELER, R. & J. SLAPCINSKY. 2000. A case study for the development of an Island fauna:
recent terrestrial mollusks of Bermuda. Nemouria 44: 2-99.
BOFFI, A.V. (1979). Moluscos de brasileiros de interesse médico e econômico. São Paulo.
HCITEC. 182p.

51
BRANDOLINI, S.V.P.B. & S.B. AMATO. 2001. Desenvolvimento de Eurytrema coelomaticum
(Giard & Billet) (Digenea, Dicrocoeliidae) em Bradybaena similaris (Férussac)
(Gastropoda, Xanthonychidae). Revista Brasileira de Zoologia 18 (2): 499- 510.
BURCH, J.B. 1962. How to Know the Eastern Land Snails. Iowa, WM. C, Brown Company
Publishers. 215p.
CAMERON, R.A.D. 2002. The land molluscs of North Ronaldsay, Orkney: human intervention
and island faunal diversity. Journal of Conchology 37 (5): 445-453.
CAMPOS, J.B.; M.G.O. TOSSULINO & C.R.C. MÜLLER (org.). 2005. Unidades de Conservação
– Ações para valorização da biodiversidade. Paraná, Instituto Ambiental do Paraná.
348p.
CHRISTENSEN, C.C. & P.V. KIRCH. 1986. Nonmarine mollusks and ecological change at
Barbers Point, O’ahu, Hawai´i. Bishop Museum Occasional Papers 26: 52-80.
CORREA, A.S. 1999. Zoogeografía de los gastrópodos terrestres de la region oriental de San
Luis Potosí, México. Revista de Biologia Tropical 47: 493-502.
COWIE, R.H. 1998. Patterns of introduction of non-indigenous non-marine snails and slugs in
the Hawaiian Islands. Biodiversity and Conservation 7: 349-368.
COWIE, R.H. 2000a. New records of alien land snails and slugs in the Hawaian Islands.
Bishop Museum Occasional Papers 64: 51-53.
COWIE, R.H. 2000b. Non-indigenous land and freshwater mollusks in the islands of the
Pacific: conservation impacts and threats, p. 143-172. In: G. SHERLEY (ed.). Invasive
species in the Pacific: a technical review and draft regional strategy. South Pacific
Regional Environment Programme, Apia. 190p.
COWIE, R.H. 2001. Invertebrate invasions on Pacific Islands and the replacement of unique
native faunas: a synthesis of the land and freshwater snails. Biological Invasions 3: 119-
136.

52
D’ÁVILA S. & E.C.A. BESSA. 2005a. Influência do substrato sobre a reprodução de Subulina
octona (Brugüière) (Mollusca, Subulinidae), sob condições de laboratório. Revista
Brasileira de Zoologia 22 (1): 197–204.
D’ÁVILA S. & E.C.A. BESSA. 2005b. Influência do substrato sobre o crescimento de Subulina
octona (Brugüière) (Mollusca, Subulinidae), sob condições de laboratório. Revista
Brasileira de Zoologia 22 (1): 205–211.
D’ÁVILA S.; R.J.P. DIAS & E.C.A. BESSA. 2006. Comportamento agregativo em Subulina
octona (Brugüière) (Mollusca, Subulinidae). Revista Brasileira de Zoologia 23 (2): 357–
363.
D’ÁVILA S.; R.J.P. DIAS; E.C.A. BESSA & E. DAEMON. 2004. Resistência à dessecação em três
espécies de moluscos terrestres: aspectos adaptativos e significado para o controle de
helmintos. Revista Brasileira de Zoociências 6 (1): 115-127.
DE OLIVEIRA , C.S.; M.C. VASCONCELLOS & J. PINHEIRO. 2008. The population density effects
on the reproductive biology of the snail Bradybaena similaris (Férussac, 1821) (Mollusca,
Gastropoda). Brazilian Journal of Biology 68 (2): 367-371.
D’ORBIGNY, A. 1837. Voyage dans l’Amerique Meridionale... Mollusques. P. Bertarnd,
Paris V. Levrault, Strasbourg. 5 (3): 185-376.
DUARTE, M.J.F. 1980. O ciclo evolutivo de Postharmostomum gallinum Witenberg, 1923, no
estado do Rio de Janeiro, Brasil (Trematoda, Brachylaemidae). Revista Brasileira de
Biologia 40 (4): 793-809.
DUTRA, A.V.C. 1988. Aspectos da ecologia e da reprodução de Leptinaria unilamellata
(Orbigny, 1835) (Gastropoda, Subulinidae). Revista Brasileira de Zoologia 5 (4): 581-
591.
ESPINOLA, L.A.E & H.F.J. JUNIOR. 2007. Espécies invasoras: conceitos, modelos e atributos.
Interciência 32 (9): 580-585.

53
FONSECA, A.L.M. & J.W. THOMÉ. 1993. Classificação das espécies neotropicais de
micromoluscos dos gêneros Stephanoda Albers, 1860, Stephadiscus Scott, 1981 e
Ptychodon Ancey, 1888 (Charopidae: Endodontoidea: Stylommatophora: Gastropoda).
Acta Biologica Leopoldensia 15 (2): 65-76.
FONSECA, F.C., G.K.M. NUNES & S.B. SANTOS. 2007. Lista preliminar da malacofauna
“urbana” da Vila Dois Rios, Ilha Grande, Brasil. In: XX Encontro Brasileiro de
Malacologia, 2007, Rio de Janeiro Resumos... Rio de Janeiro: UERJ. p. 231.
FONSECA, F.C., G.K.M. NUNES & S.B. SANTOS. 2009. Malacofauna urbana da Vila Dois Rios
e da Vila do Abraão, Ilha Grande, Angra dos reis, Rio de Janeiro. In: XXI Encontro
Brasileiro de Malacologia, 2009, Rio de Janeiro Cd de Resumos... Rio de Janeiro: UERJ.
p. 250.
FURTADO, M.C.V.; E.C.A. BESSA & M.C.M.N. CASTANÕN. 2004. Ovoteste de Bradybaena
similaris (Férussac, 1821) (Molusca, Xanthonychidae): histologia e produção de gametas.
Revista Brasileira de Zoociências 6 (1): 7-17.
GARCIA, T.A. & J. PINHEIRO. 2007. The photoperiod may modulate the carbohydrate
metabolismo of Bradybaena similaris (Férussac, 1821) (Molusca, Bradybaenidae).
Revista Brasileira de Zoociências 9 (1): 7-12.
GASCOIGNE, A. 1994. The dispersal of terrestrial gastropod species in the Gulf of Guinea.
Journal of Conchology 35 (1): 1-7.
GOULD, A.A. 1843. Monograph of the species of Pupa found in the United States, with
figures. Boston Journal of Natural History 4:350-360.
GRAY, J. E. 1857. Guide to the systematic distribution of Mollusca in the British Museum
– Part I. London, British Museum. 230p.
HAAS, F. 1949. Land and fresh-wather Mollusks from Peru. Fieldiana: Zoology 31 (28):
235-250.

54
HAAS, F. 1953. Mollusks from Ilha Grande, Rio de Janeiro, Brazil. Fieldiana: Zoology 34
(20): 203-209.
HAAS, F. 1955. On some small collections of inland shell from South America – Land and
fresh-wather Mollusks from Peru. Fieldiana: Zoology 34 (35): 361-387.
HADLEY , M. 1991. Rain Forest regeneration and manegement: strengthening the Research
Management Connection. In: GOMES-POMBA, A.; HADLEY , M. & WHITMOR, T. C. (Orgs.).
Rain Forest regeneration and manegement. Man and Biosfere Series. Londres: Unesco &
The Parthenon, Publ. Group.
HAUSDORF, B. 2002. Introduced land snails and slugs in Colombia. Journal of Molluscan
Studies 68: 127-131.
HOLLAND , K.D.; M.J. MCDONNELL & N.S.G. WILLIAMS . 2007. Abundance, species richness
and feeding preferences of introduced mollusks in native grasslands of Victoria, Australia.
Austral Ecology 32: 626-634.
JUŘIČKOVÁ , L. 2006. Subulina octona (Bruguière, 1798) – a new greenhouse species for the
Czech Republic (Mollusca: Gastropoda: Subulinidae). Malacologica Bohemoslovaca 5:
1–2.
KAGEYAMA , P. & N.M. LEPSCH-CUNHA. 2001. Singularidade da biodiversidade nos trópicos,
p. 199-214. In: I. GARAY & B. DIAS. Conservação da biodiversidade em ecossistemas
tropicais: avanços conceituais e revisão de novas metodologias de avaliação e
monitoramento. Petrópolis, Editora Vozes. 430 p.
LANZIERI, P.D. 1965. Sobre a cópula e constituição do aparelho genital de Succinea
meridionalis Orbigny, 1846 (Gastropoda, Pulmonata, Succineidae). Memórias do
Instituto Oswaldo Cruz 63: 207-228.

55
LIRA, C.R.S.; E. M. GOMES; G.M. CHAGAS & J. PINHEIRO. 2000. Influência do jejum severo
sobre o conteúdo de proteínas totais e de amônio na hemolinfa de Bradybaena similaris
(Férussac). Revista Brasileira de Zoologia 17 (4): 907- 913.
LUSTRINO, D.; V.M. TUNHOLI-ALVES; V.M. TUNHOLI; E.C.A. BESSA & J. PINHEIRO. 2008.
Allamanda catartica L. (Apocynacea) seeds induces changes on carbohydrates deposits of
Bradybaena similaris (Férussac, 1821) (Mollusca, Bradybaenidae). Revista Brasileira de
Zoociências 10 (1): 23-27.
MACIEL, N.C.; D.S.D. ARAUJO & A. MAGNANINI . 1984. Reserva Biológica da Praia do Sul
(Ilha Grande, Angra dos Reis, RJ): Contribuição para conhecimento da fauna e da flora.
Boletim da Fundação Brasileira para a Conservação da Natureza 19: 126-148.
MAGURRAN, A.E. 1988. Ecologycal diversity and its measurement. Cambridge, University
Press. 179p.
MEIRELES, L.M.O.; L.C. SILVA ; F.O. JUNQUEIRA & E.C.A. BESSA. 2008. The influence of diet
and isolation on growth and survivalin the land snail Bulimulus tenuissimus (Mollusca:
Bulimulidae) in laboratory. Revista Brasileira de Zoologia 25 (2): 224–227.
MELLO, C.E.H.V. 1987. Apontamentos para servir à história fluminense (Ilha Grande)
Angra dos Reis. Angra dos Reis, Conselho Municipal de Cultura de Angra dos Reis.
115p.
MENEZ, A. 2002. The standardization of abiotic variable data collection in land mollusc
research. Journal of Conchology 37 (5): 581- 583.
MIENIS, H.K. 1977. North American land snails in Israel. The Nautilus 91 (1): 30-31.
MIQUEL, S.E. & H. PARENT. 1996. Moluscos gasteropodos de la Provincia de Santa Fe,
Argentina. Malacological Review 29: 107- 112.
MINISTÉRIO DO MEIO AMBIENTE - MMA. 2006. Espécies exóticas invasoras: Situação
Brasileira/ Ministério do Meio Ambiente, Secretaria de Biodiversidade e Florestas. 24p.

56
MONTEIRO, D. P. 2005. Morfologia da concha de Neocyclotus prominulus (d’Orbigny, 1835)
de três áreas de floresta na Ilha Grande, Rio de Janeiro. Dissertação de Mestrado,
Instituto de Biologia Roberto Alcantara Gomes, Universidade do Estado do Rio de
Janeiro. 97 p.
MOREIRA, C.S.D.R.; E.M.GOMES; G.M. CHAGAS & J. PINHEIRO. 2003. Calcium changes in
Bradybaena similaris (Férussac, 1821) (Mollusca, Xanthonychidae) under starvation.
Revista Brasileira de Zoociências 5 (1): 45-54.
MORICAND, S. 1846. Troisième supplément au mémoire sur les coquilles terrestres e
fluviatiles de la province de Bahia, envoyeés par M. Blanchet. Memóire de la Societé
Physique et d’Histoire Naturelle de Genève 11: 147-160.
MORRETES, F.L. 1949. Ensaio de Catálogo dos Moluscos do Brasil. Arquivos do Museu
Paranaense 7: 5- 216.
NASCIMENTO, C.A.A.; E. ARÉVALO; I.S. AFONSO-NETO; E.C.A. BESSA & G.L.G. SOARES.
2006. Efeito do extrato aquoso de folhas de Allamanda cathartica L. (Apocynaceae) sobre
Bradybaena similaris (Férussac, 1821) (Mollusca, Bradybaenidae) em condições de
laboratório. Revista Brasileira de Zoociências 8 (1):77-82.
NUNES, G.K.M. 2007. Comparação da diversidade da malacofauna terrestre em duas
vertentes, oceânica e continental, da Ilha Grande, Angra dos reis, Rio de Janeiro, Brasil.
Dissertação de Mestrado, Instituto de Biologia Roberto Alcantara Gomes, Universidade
do Estado do Rio de Janeiro. 148 p.
NUNES, G.K.M.; M.A. EZEQUIEL & S.B. SANTOS. 2003 Composição da malacofauna em dois
sub-ambientes em uma área de mata secundária na Ilha Grande, Rio de Janeiro. In: XVIII
Encontro Brasileiro de Malacologia, 2003, Rio de Janeiro. Resumos... Rio de Janeiro:
UERJ. p. 196.

57
NUNES, G. K. M & S. B. SANTOS. 2006. Levantamento prévio da malacofauna urbana da Vila
Dois Rios, Ilha Grande, Angra dos Reis, Brasil. In: XXVI Congresso Brasileiro de
Zoologia, 2006, Londrina. CD de Resumos... Londrina: UEL.
ODUM, E.P. 1988. Ecologia. Rio de Janeiro, Guanabara Koogan S. A., 434p.
OLAZARRI, J. 1986. Lamellaxis gracilis (Hutton, 1834) (Gastropoda: Subulinidae) en el
Uruguay. Comunicaciones de la Sociedad Malacologica del Uruguay VI (50): 361-364.
OLIVEIRA , C.R.P. 2002. Estudo bionômico de uma espécie de Gastrocopta (Gastropoda;
Vertiginidae; Gastrocoptinae) e caracterização da malacofauna acompanhante do Parque
Moinhos de Vento, Porto Alegre e do Sistema Arroio-Lagoa Bolaxa, Rio Grande, Rio
Grande do Sul. Dissertação de Mestrado, Instituto de Biociências, Universidade Federal
do Rio Grande do Sul. 127 p.
OLIVEIRA , M.O.; I. VIEIRA & M.H.R. OLIVEIRA . 1971. Sobre Bradybaena similaris Férussac
(Gastropoda, Pulmonata, Stylommatophora, Fruticicolidae) copula y funcionamiento del
oviducto durante el período de fecundacion y formacion del huevo. Comunicaciones de
la Sociedad Malacologica del Uruguay III (21): 155-161.
OLIVEIRA , R.R. 2002. Ação antrópica e resultantes sobre a estrutura e composição da Mata
Atlântica na Ilha Grande, RJ. Rodriguésia 53 (82): 33-58.
OLIVEIRA , R.R. & A. L. COELHO NETTO. 1996. O rastro do homem na floresta. A construção
da paisagem da Reserva Biológica Estadual da Praia do Sul (Ilha Grande, Rio de Janeiro)
a partir das intervenções antrópicas. Alberoa 4 (10): 110-118.
OLIVEIRA , R.R. & NETTO, A.L.C. 2006. O rastro do homem na floresta: a construção da
paisagem da Reserva Biológica Estadual da Praia do Sul (Ilha Grande, Rio de
Janeiro) a partir das intervenções antrópicas: 39 – 58. In: PRADO, R.M. (Org.). 2006.
Ilha Grande: do sambaqui ao turismo. Garamond, EDUERJ, 288p.

58
PASCHOAL, S.V. & S.B. AMATO. 1996. Eurytrema coelomaticum (Giard & Billet) (Digenea,
Dicrocoeliidae) em Bradybaena similaris (Férussac) (Gastropoda, Xanthonychidae):
Alterações nos depósitos de cálcio e na função reprodutiva do primeiro hospedeiro
intermediário. Revista Brasileira de Zoologia 13: 411-418.
PFEIFFER, L. 1846. Symbolae ad historiam heliceorum. Casselis: 1-100.
PILSBRY, H.A. 1897-1898. American Bulimulidae: Bulimulus, Neopetraeus, Oxychona,
and South American Drymaeus. In TRYON, G.B. Manual of Conchology, Second
Series: Pulmonata 11. Philadelphia, The Academy of Natural Sciences of Philadelphia,
339p + 51 pls.
PILSBRY, H.A. 1906-1907. Achatinidae: Stenogyrinae and Coeliaxinae. In TRYON, G.B.
Manual of Conchology, Second Series: Pulmonata 18. Philadelphia, The Academy of
Natural Sciences of Philadelphia, 357p + 51 pls.
PILSBRY, H.A. 1916 -1918. Pupillidae (Gastrocoptinae). Manual of Conchology, Second
Series: Pulmonata 24. Philadelphia, The Academy of Natural Sciences of Philadelphia,
380p + 49 pls.
PILSBRY, H.A. 1926. The land mollusks of the republic of Panama and the Canal Zone.
Proceedings of the Academy of Natural Sciences of Philadelphia 78: 57-127.
PINHEIRO, J.; E.M. GOMES & G.M. CHAGAS. 2001. Aminotransferases activity in the
hemolymph of Bradybaena similaris (Gastropoda, Xanthonychidae) under starvation.
Memórias do Instituto Oswaldo Cruz 96 (8): 1161-1164.
PREECE, R.C. 2001. Introduced land mollusks on the Island of the Tristan da Cunha – Gough
Group (South Atlantic). Journal of Conchology 37 (3): 253-259.
PRIMACK, R.B. & E. RODRIGUES. 2001. Biologia da Conservação. Londrina, Editora Vida,
328p.

59
RAMIREZ, R. L. 1993. A generic analysis of the family Systrophiidae (Mollusca: gastropoda):
taxonomy, phylogeny and biogeography. Dissertação de Mestrado, Faculty of the
Graduate School, University of Kansas, 218 p.
RAMIREZ, R.L.; S. CÓRDOVA & K. CARO. 2001. Diversity of land mollusks in the bamboo-
dominated Forest of the Lower Urubamba Region, Peru, p. 49-53. In: A. ALONSO; F.
DALMEIR & P. CAMPBELL (eds). Urubamba: the biodiversity of a Peruvian rainforest.
Smithsonian Institution, SI/MAB Biodiversity Program, 216pp.
RICKLEFS, R.E. 2003. A economia da natureza. Rio de Janeiro, Guanabara Koogan, 5ª.ed.,
503p.
ROTH, B. 1997. “Aclis” californica Bartsch, 1997: a land snail misintrepeted (Gastropoda:
Pulmonata: Subulinidae). The Veliger 40 (4): 364-366.
SANTOS, S.B. & D.P. MONTEIRO. 2001. Composição de gastrópodes terrestres em duas áreas
do Centro de Estudos Ambientais e Desenvolvimento Sustentado (CEADS), Vila Dois
Rios, Ilha Grande, Brasil – um estudo piloto. Revista Brasileira de Zoologia 18 (supl.1):
181-190.
SANTOS, M. S.; J. N. C. LOUZADA; N. DIAS; R. ZANETTI; J. H. C. DELABIE & I. C.
NASCIMENTO. 2006. Riqueza de formigas (Hymenoptera, Formicidae) da serapilheira em
fragmentos de floresta semidecídua da Mata Atlântica na região do Alto do Rio Grande,
Minas Gerais, Brasil. Iheringia, Série Zoologia 96 (1): 95-101.
SCHILTHUIZEN, M. & H. A. RUTJES. 2001. Land snail diversity in a square kilometre of
tropical rainforest in Sabah, Malaysian Borneo. Journal of Molluscan Studies 67: 417-
423.
SILVA , A.L. 2007. Habroconus (Pseudoguppya) semenlini (Moricand, 1845) (Gastropoda,
Stylommatophora, Euconulidae) e malacofauna acompanhante de um fragmento de
floresta nativa alterada, no Campus do Vale da Universidade Federal do Rio Grande do

60
Sul, Porto Alegre, Brasil. Dissertação de Mestrado, Instituto de Biociências,
Universidade Federal do Rio Grande do Sul. 118 p.
SILVA L.C.; L.M.O. MEIRELES; F.O. JUNQUEIRA & E.C.A. BESSA. 2008. Development and
reproduction in Bulimulus tenuissimus (Mollusca: Bulimulidae) in laboratory. Revista
Brasileira de Zoologia 25 (2): 220–223.
SIMONE, L. R. L. 2006. Land and freshwater molluscs of Brazil. São Paulo, EGB, Fapesp,
390p.
SOLEM, A. 1961. New Caledonian land and fresh-water snails an annotated check list.
Fieldiana Zoology 41 (3): 412-501.
SUTER, H. 1900. Observações sobre alguns caracoes terrestres do Brazil. Revista do Museu
Paulista 4 (3): 329-337.
TENÓRIO, M.C. 1999. Os fabricantes de machado da Ilha Grande. In: TENÓRIO, M.C. (Org.).
Pré-história da Terra brasilis. Rio de Janeiro: Editora da UFRJ, 380 p.
THOMÉ, J.W. 1975. Distensão de moluscos terrestres para fixação, com comentários sobre
coleta e transporte. Nota prévia. Arquivos do Museu Nacional do Rio de Janeiro 55:
153-154.
THOMÉ, J.W.; S.R. GOMES & J.B. PICANÇO. 2006. Guia ilustrado: Os caracóis e as lesmas
dos nossos bosques e jardins. Pelotas, USEB. 123p.
THOMÉ, J.W. & V.L.R. LOPES. 1973. Aulas práticas de zoologia, I. Dissecação de um molusco
gastrópode desprovido de concha. Iheringia 3: 34- 45.
TILLIER , S. 1980. Gastéropodes terrestres et fluviatiles de Guyane Française. Mémoires du
Muséum National d’histoire naturelle Série A, Zoologie 118: 1- 189.
UNDERWOOD, A. J. 1997. Experiments in ecology. Cambridge, Cambridge University Press.
504p.

61
WHITTAKER, R. H. 1975. Communities and ecossistems. New York, Macmilian Publishing
CO., Inc., 385p.
WUNDER, S. 2006. A história do uso do solo e da cobertura florestal. 105-130. In: PRADO,
R.M. (Org.). Ilha Grande: do sambaqui ao turismo. Garamond,










![Filo Molusca [Modo de Compatibilidade] · São celomados. Sistema digestivo completo. Respiração Pode ser de três tipos; Pulmonar : nos moluscos terrestres. ... Os moluscos apresentam](https://static.fdocumentos.tips/doc/165x107/5bb3a0f209d3f2d3728bc422/filo-molusca-modo-de-compatibilidade-sao-celomados-sistema-digestivo-completo.jpg)






![[I] Moluscos terrestres e a malacologia paranaense: histórico e … · 2019. 10. 10. · Os moluscos terrestres são gastrópodos que formam um grupo bastante diversificado, com](https://static.fdocumentos.tips/doc/165x107/60bbc316fdc3e37dec7bfab1/i-moluscos-terrestres-e-a-malacologia-paranaense-histrico-e-2019-10-10.jpg)
