
37. CURVAS DE PROGRESSO DA DOENÇA - dbbe.fcen.uba.ar · A curva de progresso da doença, ......
21
405 Capítulo 37 37. CURVAS DE PROGRESSO DA DOENÇA Armando Bergamin Filho A curva de progresso da doença, usualmente expressa pela plotagem da proporção de doença versus tempo, é a melhor representação de uma epidemia. Por meio dela, por exemplo, interações entre patógeno, hospedeiro e ambiente podem ser caracterizadas, estratégias de controle avaliadas, níveis futuros de doença previstos e simuladores verificados. A análise de epidemias por meio da curva de progresso da doença constitui-se apenas em um segmento de uma especialidade mais ampla, conhecida dos ecologistas por análise da curva de crescimento. Crescimento é definido como uma mudança de magnitude de qualquer característica mensurável, como peso, número, comprimento ou, neste contexto, proporção de doença. A análise da curva de crescimento tem uma história muito mais antiga que a análise da curva de progresso da doença. Enquanto que esta tem sua origem no trabalho pioneiro de Vanderplank (1963), aquela pode ser seguida desde o famoso trabalho de Malthus, no longínquo 1798, sobre o crescimento humano, passando por Verhulst e o crescimento logístico, de 1838, até os biomatemáticos modernos (ver, por exemplo, Newby, 1980). Modelos matemáticos de crescimento são capazes de resumir, na forma de expressões matemáticas relativamente simples, a relação existente entre doença e tempo. Estas expressões facilitam a análise dos dados de progresso da doença, dados obtidos, por exemplo, em experimentos onde produtos químicos ou variedades com diferentes níveis de resistência são testados. A capacidade de permitir comparações já se constitui numa justificativa suficiente para o estudo das curvas de progresso de doenças e para o desenvolvimento de expressões matemáticas (modelos) que as descrevem. Além desse interesse prático, modelos matemáticos de curvas de progresso da doença também podem contribuir para um melhor entendimento do processo epidêmico. Curvas de progresso da doença podem ser construídas para qualquer patossistema: o hospedeiro, por exemplo, pode ser anual, perene ou semi-perene, de origem tropical ou temperada; o patógeno, por sua vez, pode ser um fungo, um vírus, uma bactéria ou qualquer outro agente causal; a epidemia pode ser de curta, média ou longa duração; a área na qual a doença está ocorrendo pode ser desde uma pequena parcela experimental até um continente inteiro. Independentemente da situação considerada, os parâmetros importantes da curva de progresso da doença – como a época de início da epidemia, a quantidade de inóculo inicial (x 0 ), a taxa de aumento da doença (r), a forma e a área sob a curva de progresso da doença, as quantidades máxima (x max ) e final (x f ) de doença e a duração da epidemia – podem ser caracterizados.
Transcript of 37. CURVAS DE PROGRESSO DA DOENÇA - dbbe.fcen.uba.ar · A curva de progresso da doença, ......

Importância das Doenças de Plantas
405
Capítulo 37
37. CURVAS DE PROGRESSO DA DOENÇAArmando Bergamin Filho
A curva de progresso da doença, usualmente expressa pela plotagem da proporção de doença versus tempo, é a melhor representação de uma epidemia. Por meio dela, por exemplo, interações entre patógeno, hospedeiro e ambiente podem ser caracterizadas, estratégias de controle avaliadas, níveis futuros de doença previstos e simuladores verifi cados.
A análise de epidemias por meio da curva de progresso da doença constitui-se apenas em um segmento de uma especialidade mais ampla, conhecida dos ecologistas por análise da curva de crescimento. Crescimento é defi nido como uma mudança de magnitude de qualquer característica mensurável, como peso, número, comprimento ou, neste contexto, proporção de doença. A análise da curva de crescimento tem uma história muito mais antiga que a análise da curva de progresso da doença. Enquanto que esta tem sua origem no trabalho pioneiro de Vanderplank (1963), aquela pode ser seguida desde o famoso trabalho de Malthus, no longínquo 1798, sobre o crescimento humano, passando por Verhulst e o crescimento logístico, de 1838, até os biomatemáticos modernos (ver, por exemplo, Newby, 1980).
Modelos matemáticos de crescimento são capazes de resumir, na forma de expressões matemáticas relativamente simples, a relação existente entre doença e tempo. Estas expressões facilitam a análise dos dados de progresso da doença, dados obtidos, por exemplo, em experimentos onde produtos químicos ou variedades com diferentes níveis de resistência são testados. A capacidade de permitir comparações já se constitui numa justifi cativa sufi ciente para o estudo das curvas de progresso de doenças e para o desenvolvimento de expressões matemáticas (modelos) que as descrevem. Além desse interesse prático, modelos matemáticos de curvas de progresso da doença também podem contribuir para um melhor entendimento do processo epidêmico.
Curvas de progresso da doença podem ser construídas para qualquer patossistema: o hospedeiro, por exemplo, pode ser anual, perene ou semi-perene, de origem tropical ou temperada; o patógeno, por sua vez, pode ser um fungo, um vírus, uma bactéria ou qualquer outro agente causal; a epidemia pode ser de curta, média ou longa duração; a área na qual a doença está ocorrendo pode ser desde uma pequena parcela experimental até um continente inteiro. Independentemente da situação considerada, os parâmetros importantes da curva de progresso da doença – como a época de início da epidemia, a quantidade de inóculo inicial (x0), a taxa de aumento da doença (r), a forma e a área sob a curva de progresso da doença, as quantidades máxima (xmax) e fi nal (xf) de doença e a duração da epidemia – podem ser caracterizados.

Manual de Fitopatologia
406
37.1 CLASSIFICAÇÃO EPIDEMIOLÓGICA DE DOENÇA37.1.1 Taxas de juros e capitalVanderplank (1963), no clássico livro “Plant diseases: epidemics and control”,
baseou toda sua teoria epidemiológica na analogia entre crescimento de capital (dinheiro) e crescimento de doença. Dois tipos de crescimento de capital foram considerados: a juros simples e a juros compostos. Um exemplo será útil para recordar esses conceitos: capital guardado na gaveta do criado-mudo não cresce. Em termos matemáticos, o capital Ct no tempo t será o mesmo que o capital C0 no tempo t0, C Ct = 0 (37.1)
Isto é equivalente a dizer que o capital tem uma taxa de crescimento nula dC dt/ = 0 (37.2)Se, porém, este mesmo capital for depositado num banco, a uma taxa de juros
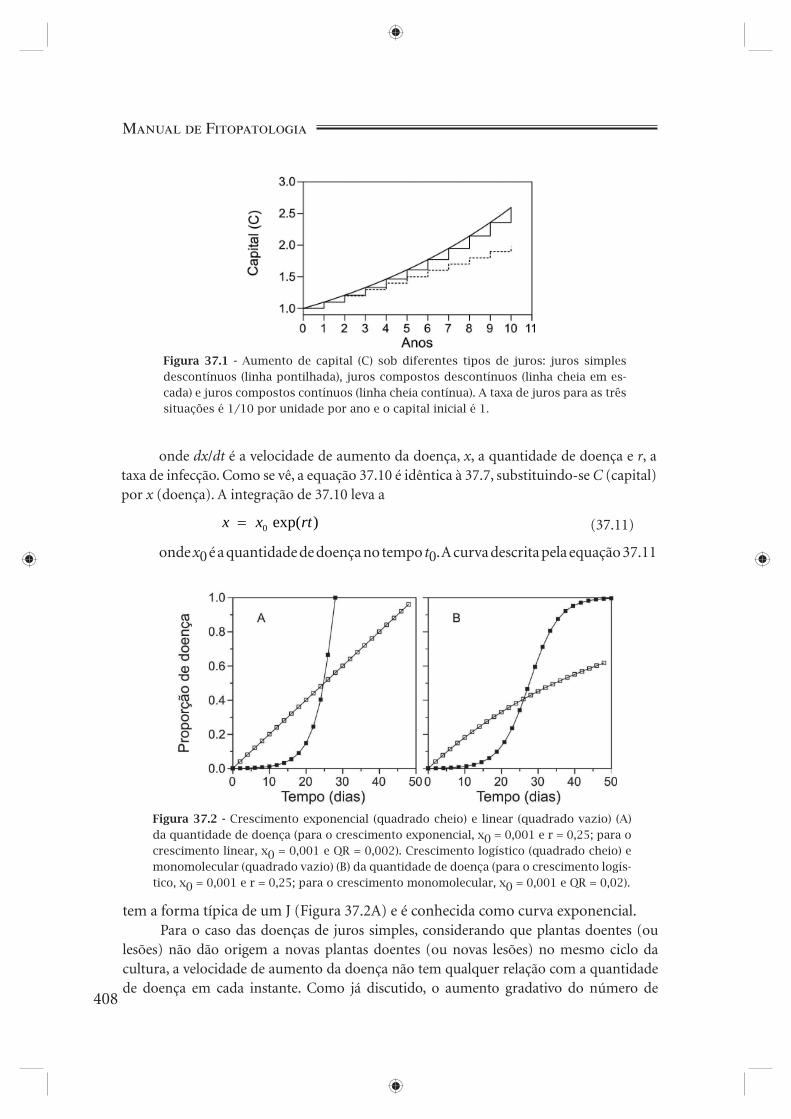
de 10% ao ano, e se os juros ganhos ao fi nal de cada ano forem guardados no já citado criado-mudo, o capital total acumulado (banco + criado-mudo) crescerá a uma taxa constante. Esta situação caracteriza um crescimento descontínuo a juros simples (Figura 37.1 - linha pontilhada). Após t anos, o capital inicial C0 modifi cou-se para
C C rtt = +0 1( ) (37.3)onde r é a taxa de juros, no exemplo igual a 0,1 por ano. A correspondente taxa
média de crescimento é expressa por dC dt r/ = (37.4)
Considere agora uma situação diferente: os juros ganhos anualmente, ao invés de serem guardados no criado-mudo, são deixados no banco para serem adicionados ao capital inicial. Assim, os juros ganhos no ano anterior irão, eles mesmos, render juros nos anos subsequentes. A consequência disso, não é difícil imaginar, é que os ganhos são crescentes a cada ano. Essa situação caracteriza um crescimento também descontínuo, mas agora, a juros compostos (Figura 37.1 - linha cheia em escada). Após t anos, o capital inicial C0 modifi cou-se para
C C rt
t= +0 1( ) (37.5)
A unidade de tempo que tem sido empregada até aqui é o ano, mas a equação 37.5 mantém sua validade ainda que meses, dias ou qualquer outra unidade seja escolhida. O cálculo mostra que a equação 37.5 transforma-se em
C C rtt = 0 exp( ) (37.6)
caso a unidade de tempo seja infi nitesimamente pequena. Nesta equação, exp representa e (base dos logaritmos naturais, com valor aproximado de 2,7182) elevado a uma potência específi ca. Esta situação caracteriza um crescimento contínuo a juros compostos (Figura 37.1 - linha contínua). A correspondente taxa média de crescimento é

Importância das Doenças de Plantas
407
expressa por:
dC dt rC/ = (37.7)Usando logaritmo, a equação 37.6 pode ser escrita como
ln lnC rt C= + 0 (37.8)que é o mesmo que
r t C C= ( / ) ln( / )1 0 (37.9)A equação 37.8 torna claro que a curva de crescimento de capital é uma linha reta
caso utilize-se uma escala logarítmica (ordenada)-linear (abscissa).
37.1.2 Taxas de infecção e doençaNeste item, taxas de juros tornam-se taxas de infecção e capital torna-se doença.
Como no item anterior, dois grupos são conceituados: doenças de juros compostos e doenças de juros simples (Vanderplank, 1963). No primeiro grupo, plantas infectadas durante o ciclo da cultura servirão de fonte de inóculo para novas infecções durante o mesmo ciclo. É o caso típico da ferrugem do trigo, por exemplo, cujo agente causal (Puccinia graminis f. sp. tritici), em condições favoráveis, pode produzir uma geração a cada 10 dias. Esta situação é análoga ao crescimento de capital a juros compostos, onde os juros ganhos rendem novos juros; no caso de doenças de juros compostos, plantas doentes rendem novas plantas doentes durante o ciclo da cultura. Para que isto ocorra, está implícita uma movimentação do patógeno a partir de plantas doentes em direção a novos sítios de infecção. No segundo grupo, plantas infectadas durante o ciclo da cultura não servirão de fonte de inóculo para novas infecções durante o mesmo ciclo. É o caso típico da murcha do algodoeiro, por exemplo, cujo agente causal (Fusarium oxysporum f. sp. vasinfectum) coloniza principalmente o interior do xilema das plantas infectadas. Esta situação é análoga ao crescimento de capital a juros simples, onde os juros ganhos não rendem novos juros; no caso de doenças de juros simples, o aumento gradativo do número de plantas doentes durante o ciclo da cultura não é devido, primariamente, à movimentação do patógeno a partir de plantas doentes em direção a novos sítios de infecção e, sim, ao inóculo original, neste caso clamidósporos, previamente existente no solo.
Uma vez conceituados os dois grupos epidemiológicos de Vanderplank (1963), é tempo de voltar a atenção para o tema deste capítulo: curvas de progresso da doença. Como seria a curva típica de cada um deles?
Para o caso das doenças de juros compostos, considerando que plantas doentes (ou lesões) dão origem a novas plantas doentes (ou novas lesões) no mesmo ciclo da cultura, a velocidade de aumento da doença é proporcional à própria quantidade de doença em cada instante. Assim, se uma lesão der origem a 10 lesões, 10 lesões darão origem a 100, 100 a 1000, 1000 a 10.000 e assim por diante. Essa cinética de crescimento é expressa
matematicamente através da equação diferencial
dx dt rx/ = (37.10)
Manual de Fitopatologia
408
onde dx/dt é a velocidade de aumento da doença, x, a quantidade de doença e r, a taxa de infecção. Como se vê, a equação 37.10 é idêntica à 37.7, substituindo-se C (capital) por x (doença). A integração de 37.10 leva a
x x rt= 0 exp( )
onde x0 é a quantidade de doença no tempo t0. A curva descrita pela equação 37.11
Figura 37.1 - Aumento de capital (C) sob diferentes tipos de juros: juros simples descontínuos (linha pontilhada), juros compostos descontínuos (linha cheia em es-cada) e juros compostos contínuos (linha cheia contínua). A taxa de juros para as três situações é 1/10 por unidade por ano e o capital inicial é 1.
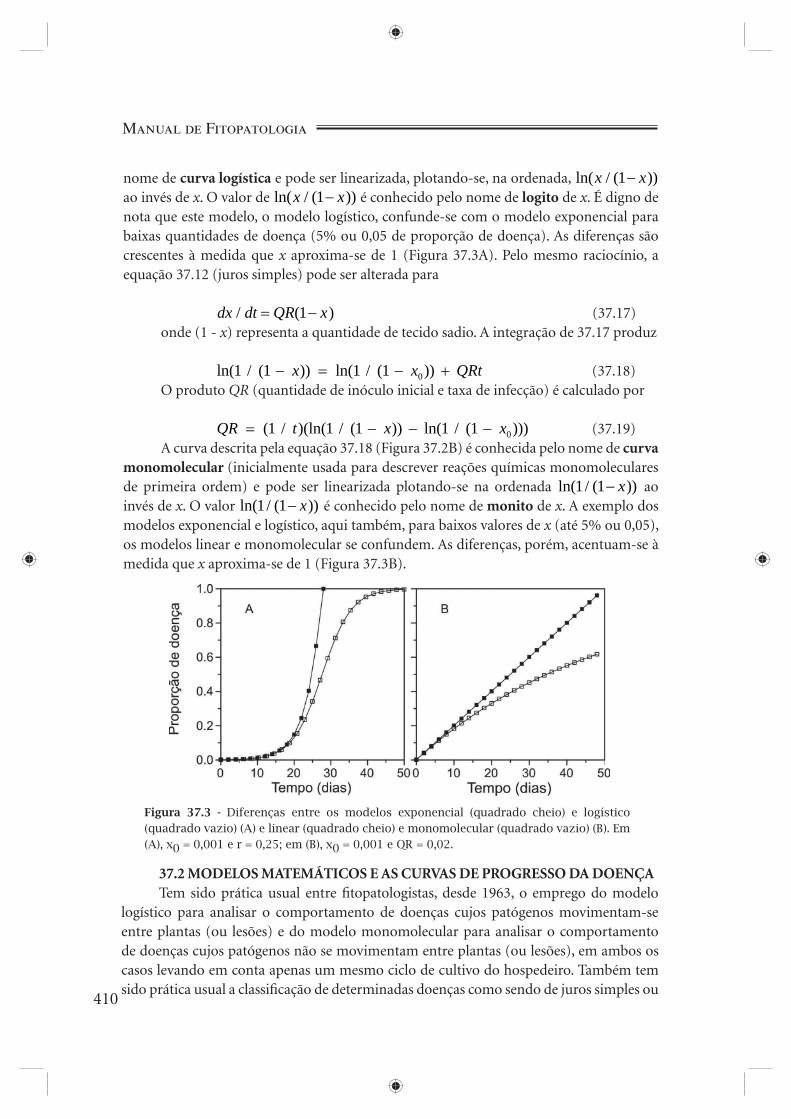
tem a forma típica de um J (Figura 37.2A) e é conhecida como curva exponencial.Para o caso das doenças de juros simples, considerando que plantas doentes (ou
lesões) não dão origem a novas plantas doentes (ou novas lesões) no mesmo ciclo da cultura, a velocidade de aumento da doença não tem qualquer relação com a quantidade de doença em cada instante. Como já discutido, o aumento gradativo do número de
Figura 37.2 - Crescimento exponencial (quadrado cheio) e linear (quadrado vazio) (A) da quantidade de doença (para o crescimento exponencial, x0 = 0,001 e r = 0,25; para o crescimento linear, x0 = 0,001 e QR = 0,002). Crescimento logístico (quadrado cheio) e monomolecular (quadrado vazio) (B) da quantidade de doença (para o crescimento logís-tico, x0 = 0,001 e r = 0,25; para o crescimento monomolecular, x0 = 0,001 e QR = 0,02).
(37.11)

Importância das Doenças de Plantas
409
plantas doentes durante o ciclo da cultura é função do inóculo original previamente existente. A quantidade de inóculo existente é, na maioria dos casos, desconhecida, mas por conveniência, considerada constante durante cada período de vegetação. A fração de plantas que se torna doente (x) depende da frequência de contatos efetivos entre hospedeiro e patógeno (inóculo original). Contato efetivo é defi nido como aquele contato que leva à doença. Assim,
dx dt QR/ = (37.12)
sendo Q a quantidade de inóculo previamente existente e R, a taxa de infecção. O produto QR representa o número de contatos efetivos. Como se vê, a equação 37.12 é semelhante à 37.4, substituindo-se C (capital) por x (doença) e r (taxa de juros) pelo produto QR (inóculo pré-existente e taxa de infecção). Tanto r quanto QR são considerados constantes. A integração de 37.12 resulta em
x x QRt= +0 (37.13)onde x0 é a quantidade de doença no tempo t0. A curva descrita pela equação
37.13 é uma linha reta (Figura 37.2A).Seriam os modelos exponencial (equação 37.11, Figura 37.2A) e linear (equação
37.13, Figura 37.2A) espelhos fi éis da realidade? Simulariam eles com razoável precisão o crescimento da doença em condições naturais? As epidemias reais, para vários patossistemas, mostram um acordo parcial entre esses modelos e a realidade: aparentemente, com quantidades pequenas de doença, os modelos fi cam próximos da realidade; à medida que a quantidade de doença aumenta, aumenta também o divórcio entre realidade e modelo. E, pensando bem, as coisas não poderiam ser diferentes: tanto o modelo exponencial quanto o linear permitem à quantidade de doença crescer até o infi nito. Nenhum processo biológico comporta-se desta maneira: leveduras ou bactérias cultivadas em meio de cultura não crescem ao infi nito, pois o meio, entre outras causas, esgota-se; a quantidade de doença, do mesmo modo, não pode tender ao infi nito, pois seu crescimento é limitado, entre outras causas, pela disponibilidade de tecido sadio. Um fator de correção obviamente faz-se necessário, fator este que reduza a velocidade de crescimento da doença proporcionalmente à diminuição da oferta de tecido sadio. A equação 37.10 (juros compostos), assim, pode ser alterada para
dx dt rx x/ ( )= −1 (37.14)onde (1 - x) representa a quantidade de tecido sadio (x, neste contexto, é sempre
expresso em proporção de doença). A integração de 37.14 produz ln( / ( )) ln( / ( ))x x x x rt1 10 0− = − + (37.15)Em consequência, o valor da taxa r (chamada de taxa aparente de infecção por
Vanderplank, 1963) é calculado por
r t x x x x= − − −( / )(ln( / ( )) ln( / ( )))1 1 10 0 (37.16)A curva descrita pela equação 37.15 tem a forma de S (Figura 37.2B), é conhecida pelo
Manual de Fitopatologia
410
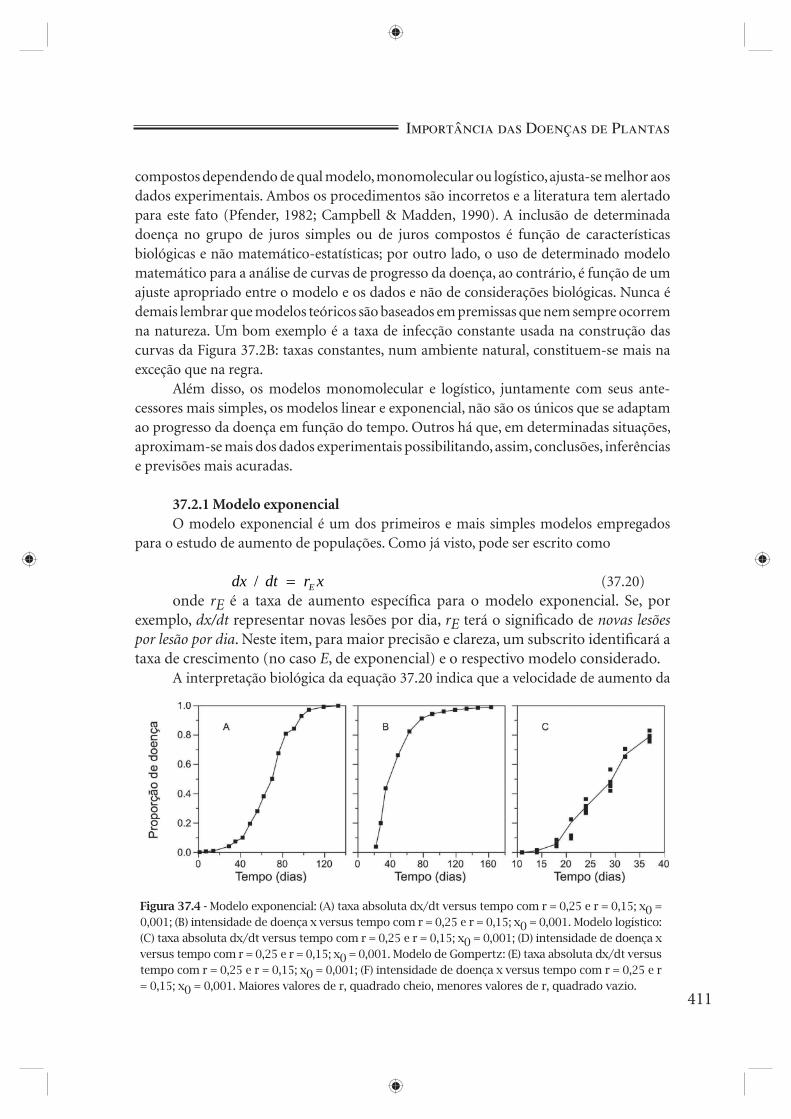
nome de curva logística e pode ser linearizada, plotando-se, na ordenada, ln( / ( ))x x1− ao invés de x. O valor de ln( / ( ))x x1− é conhecido pelo nome de logito de x. É digno de nota que este modelo, o modelo logístico, confunde-se com o modelo exponencial para baixas quantidades de doença (5% ou 0,05 de proporção de doença). As diferenças são crescentes à medida que x aproxima-se de 1 (Figura 37.3A). Pelo mesmo raciocínio, a equação 37.12 (juros simples) pode ser alterada para
dx dt QR x/ ( )= −1 (37.17)onde (1 - x) representa a quantidade de tecido sadio. A integração de 37.17 produz ln( / ( )) ln( / ( ))1 1 1 1 0− = − +x x QRt (37.18)O produto QR (quantidade de inóculo inicial e taxa de infecção) é calculado por QR t x x= − − −( / )(ln( / ( )) ln( / ( )))1 1 1 1 1 0 (37.19)A curva descrita pela equação 37.18 (Figura 37.2B) é conhecida pelo nome de curva
monomolecular (inicialmente usada para descrever reações químicas monomoleculares de primeira ordem) e pode ser linearizada plotando-se na ordenada ln( / ( ))1 1− x ao invés de x. O valor ln( / ( ))1 1− x é conhecido pelo nome de monito de x. A exemplo dos modelos exponencial e logístico, aqui também, para baixos valores de x (até 5% ou 0,05), os modelos linear e monomolecular se confundem. As diferenças, porém, acentuam-se à medida que x aproxima-se de 1 (Figura 37.3B).
Figura 37.3 - Diferenças entre os modelos exponencial (quadrado cheio) e logístico (quadrado vazio) (A) e linear (quadrado cheio) e monomolecular (quadrado vazio) (B). Em (A), x0 = 0,001 e r = 0,25; em (B), x0 = 0,001 e QR = 0,02.
37.2 MODELOS MATEMÁTICOS E AS CURVAS DE PROGRESSO DA DOENÇATem sido prática usual entre fi topatologistas, desde 1963, o emprego do modelo
logístico para analisar o comportamento de doenças cujos patógenos movimentam-se entre plantas (ou lesões) e do modelo monomolecular para analisar o comportamento de doenças cujos patógenos não se movimentam entre plantas (ou lesões), em ambos os casos levando em conta apenas um mesmo ciclo de cultivo do hospedeiro. Também tem sido prática usual a classifi cação de determinadas doenças como sendo de juros simples ou
Importância das Doenças de Plantas
411
compostos dependendo de qual modelo, monomolecular ou logístico, ajusta-se melhor aos dados experimentais. Ambos os procedimentos são incorretos e a literatura tem alertado para este fato (Pfender, 1982; Campbell & Madden, 1990). A inclusão de determinada doença no grupo de juros simples ou de juros compostos é função de características biológicas e não matemático-estatísticas; por outro lado, o uso de determinado modelo matemático para a análise de curvas de progresso da doença, ao contrário, é função de um ajuste apropriado entre o modelo e os dados e não de considerações biológicas. Nunca é demais lembrar que modelos teóricos são baseados em premissas que nem sempre ocorrem na natureza. Um bom exemplo é a taxa de infecção constante usada na construção das curvas da Figura 37.2B: taxas constantes, num ambiente natural, constituem-se mais na exceção que na regra.
Além disso, os modelos monomolecular e logístico, juntamente com seus ante-cessores mais simples, os modelos linear e exponencial, não são os únicos que se adaptam ao progresso da doença em função do tempo. Outros há que, em determinadas situações, aproximam-se mais dos dados experimentais possibilitando, assim, conclusões, inferências e previsões mais acuradas.
37.2.1 Modelo exponencialO modelo exponencial é um dos primeiros e mais simples modelos empregados
para o estudo de aumento de populações. Como já visto, pode ser escrito como dx dt r xE/ = (37.20)onde rE é a taxa de aumento específi ca para o modelo exponencial. Se, por
exemplo, dx/dt representar novas lesões por dia, rE terá o signifi cado de novas lesões por lesão por dia. Neste item, para maior precisão e clareza, um subscrito identifi cará a taxa de crescimento (no caso E, de exponencial) e o respectivo modelo considerado.
A interpretação biológica da equação 37.20 indica que a velocidade de aumento da
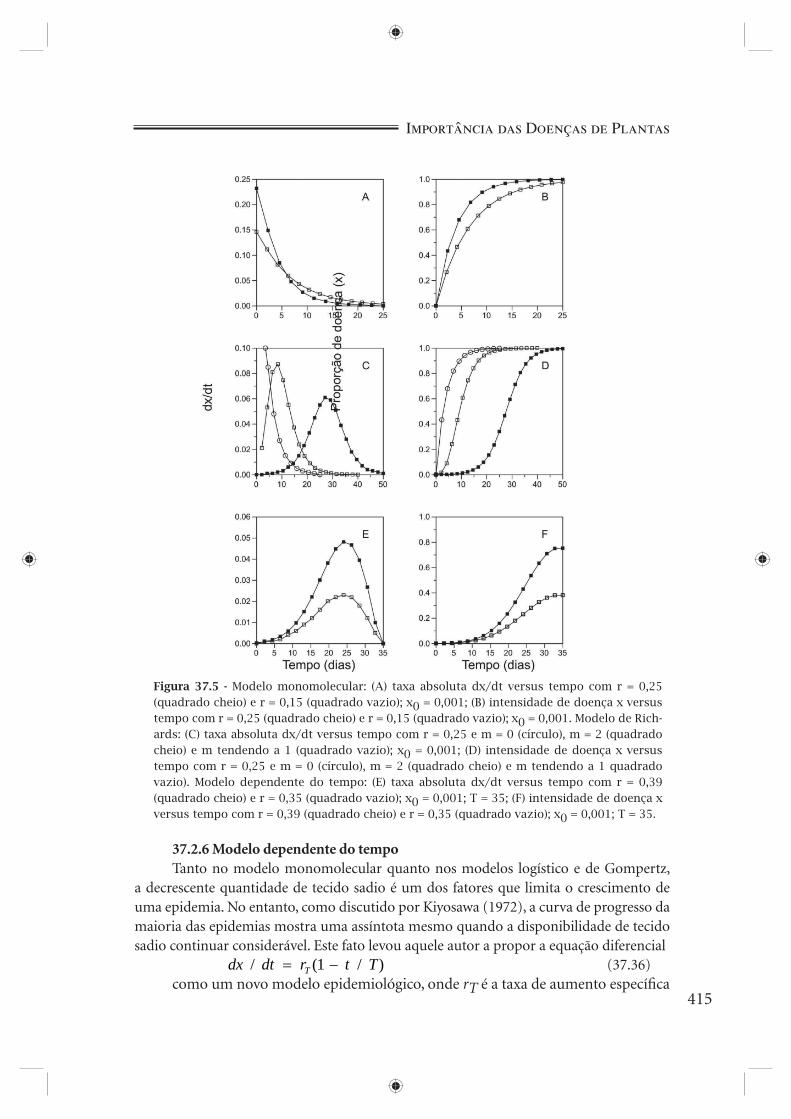
Figura 37.4 - Modelo exponencial: (A) taxa absoluta dx/dt versus tempo com r = 0,25 e r = 0,15; x0 = 0,001; (B) intensidade de doença x versus tempo com r = 0,25 e r = 0,15; x0 = 0,001. Modelo logístico: (C) taxa absoluta dx/dt versus tempo com r = 0,25 e r = 0,15; x0 = 0,001; (D) intensidade de doença x versus tempo com r = 0,25 e r = 0,15; x0 = 0,001. Modelo de Gompertz: (E) taxa absoluta dx/dt versus tempo com r = 0,25 e r = 0,15; x0 = 0,001; (F) intensidade de doença x versus tempo com r = 0,25 e r = 0,15; x0 = 0,001. Maiores valores de r, quadrado cheio, menores valores de r, quadrado vazio.

Manual de Fitopatologia
412
doença, dx/dt, também chamada de taxa absoluta de aumento da doença, é proporcional à própria quantidade de doença. Isto é o mesmo que dizer que níveis maiores de doença levarão sempre a incrementos maiores de doença, fato claramente visível no gráfi co de dx/dt e tempo (Figura 37.4A). Taxas absolutas de aumento da doença são muito úteis para se ter uma idéia de como cresce a população. Na prática, entretanto, o que se observa é o nível de doença (x) em diferentes tempos (t). A equação que descreve a variação de x em função de t é obtida pela integração de 37.20 (Figura 37.4B)
x x r tE= 0 exp( ) (37.21)O modelo exponencial, apesar de demasiado simplista, pode ser empregado
para as fases iniciais da epidemia, enquanto a quantidade de doença não exceder 5%. Experimentalmente, o valor de rE pode ser determinado conhecendo-se dois valores de quantidade de doença em tempos diferentes (x e x0) através da fórmula
r t x xE = ( / ) ln( / )1 0 (37.22)ou, quando vários valores de x forem conhecidos, por meio da regressão linear entre
ln(x) e tempo.
37.2.2 Modelo logísticoO modelo logístico, originalmente proposto por Verhulst (1838), tem sido o modelo
mais empregado para descrever o progresso de epidemias desde que Vanderplank (1963) redescobriu-o em seu clássico livro. A equação diferencial do modelo logístico pode ser escrita como
dx dt r x xL/ ( )= −1 (37.23)onde rL é a taxa de aumento específi ca para o modelo logístico e 1, a quantidade
máxima de doença. O fator de correção ( )1− x representa a quantidade de tecido sadio. Vanderplank (1963) chamou a taxa rL de taxa aparente de infecção uma vez que, nas avaliações de doença no campo, o que realmente se determina é o tecido com aparência doente, ou seja, aquele tecido infectado que já passou pelo período de incubação. A taxa rL tem o signifi cado, caso dx/dt represente novas lesões por dia, de novas lesões por lesão por dia.
A interpretação biológica da equação 37.23 indica que a velocidade de aumento da doença, dx/dt, é proporcional à própria quantidade de doença x e à quantidade de tecido sadio disponível ( )1− x . Para quantidades pequenas de doença (abaixo de 5%), o modelo logístico confunde-se com o modelo exponencial (Figura 37.3A). Quando dx/dt é plotado contra o tempo, incrementos crescentes ocorrem no início, com uma taxa absoluta máxima ou ponto de infl exão quando x = 50% (0,5), seguidos por incrementos decrescentes que tendem a zero. Como se pode ver na Figura 37.4C, a curva é simétrica em torno de seu ponto de infl exão. A integração de 37.23 produz
x x r tL= + − −1 1 1 10/ ( (( / ) ) exp( )) (37.24)A plotagem de x contra o tempo origina uma curva em forma de S, conhecida

Importância das Doenças de Plantas
413
como curva logística, simétrica em torno de x = 0,5 (Figura 37.4D). A equação 37.24 pode ser linearizada (ver equação 37.15) e rL pode ser avaliado (ver equação 37.16). Quando várias estimativas de x para diferentes tempos forem conhecidas, a taxa aparente de infecção rL pode ser calculada por meio de regressão linear, utilizando-se o logito de x (ln(x/(1-x))) contra o tempo.
37.2.3 Modelo de GompertzApesar do modelo de Gompertz ter sido introduzido na epidemiologia vegetal
algum tempo depois do modelo logístico, sua origem é mais antiga (Gompertz, 1825). Hoje em dia, no domínio das doenças de plantas, ambos os modelos são bastante empregados (Berger, 1981; Waggoner, 1986; Campbell & Madden, 1990). A equação diferencial para o modelo de Gompertz pode ser escrita como
dx dt r x xG/ (ln( ) ln( ))= −1 (37.25)que é o mesmo que
dx dt r x xG/ ( ln( ))= − (37.26)onde rG é a taxa específi ca para este modelo. A plotagem de dx/dt contra o tempo,
a exemplo do que acontece com o modelo logístico, mostra incrementos crescentes até que o ponto de infl exão seja atingido, seguindo-se incrementos decrescentes que tendem a zero. O ponto de infl exão, diferentemente do modelo logístico, ocorre no ponto x = 0,37 (1/e), fazendo com que a curva de dx/dt seja assimétrica, inclinada para a esquerda (Figura 37.4E). A integração de 37.25 ou 37.26 produz
x x r tG= − − −exp( ( ln( )) exp( ))0 (37.27)A plotagem de x contra o tempo origina uma curva também em forma de S que
difere, porém, do S logístico por apresentar um crescimento mais acentuado em seu início (Figura 37.4F). A equação 37.27 pode ser linearizada, transformando-se em
− − = − − +ln( ln( )) ln( ln ))x x r tG0 (37.28)A taxa rG pode ser determinada, quando se dispõe das estimativas de x e x0,
por r t x xG = − − − − −( / )( ln( ln( ))) ( ln( ln( )))1 0 (37.29)Quando várias estimativas de x para diferentes tempos forem conhecidas a
taxa de infecção rG pode ser calculada por meio de regressão linear, utilizando-se o gompito de x ( ln( ln( )))− − x contra o tempo.
37.2.4 Modelo monomolecularOs três modelos que foram examinados neste item até o momento (exponencial,
logístico e Gompertz) têm em comum que, neles, a velocidade de aumento da doença (dx/dt) é proporcional à própria quantidade de doença (x). Este, porém, não é o caso do modelo monomolecular. Aqui, a velocidade de aumento da doença é proporcional ao

Manual de Fitopatologia
414
inóculo inicial e a uma taxa, que em 37.1.2 foram chamados de Q e R, respectivamente, ambos supostos constantes. A equação 37.17, já discutida, pode ser reescrita, apenas para fi ns de uniformização, como
dx dt r xM/ ( )= −1 (37.30)onde rM é a taxa de aumento específi ca para o modelo monomolecular (rM
= QR) e ( )1− x , como sempre, representa tecido sadio. A plotagem de dx/dt contra o tempo mostra que os incrementos são sempre decrescentes, e aproximam-se de zero à medida que se esgota o tecido sadio (Figura 37.5A). A integração de 37.30 leva a
x x r tM= − − −1 1 0( ) exp( ) (37.31)que, plotado contra o tempo, produz uma curva côncava em relação à abscissa,
aproximando-se assintoticamente da quantidade máxima de doença (Figura 37.5B). A equação 37.31 pode ser linearizada fazendo-se
ln( / ( )) ln( / ( ))1 1 1 1 0− = − +x x r tM (37.32)e rM pode ser avaliado, desde que x e x0 sejam conhecidos, através de
r t x xM = − − −( / )(ln( / ( )) ln( / ))1 1 1 1 1 0 (37.33)Quando várias estimativas de x para diferentes tempos forem conhecidas,
rM pode ser calculado por meio de regressão linear, utilizando-se o monito de x (ln( / ( )))1 1− x contra o tempo.
37.2.5 Modelo de RichardsO modelo de Richards (Richards, 1959) difere dos demais aqui discutidos por ser o
único fl exível, ou seja, o único que pode tomar várias formas. Sua equação diferencial é dx dt r x x mR
m/ ( ( )) / ( )= − −
−1 1
1 (37.34)
onde rR é a taxa de aumento e m, o parâmetro de forma (m tem uma gama de valores de 0 a infi nito). Quando m= 0 , a equação 37.34 reduz-se à diferencial do modelo monomolecular (equação 37.30); quando m= 2 , à do modelo logístico (equação 37.23); quando m tende a 1, à do modelo de Gompertz (equação 37.25 ou 37.26); com outros valores de m, um número ilimitado de curvas dx/dt versus tempo pode ser gerado (Figura 37.5C). A integração de 37.34 produz
x x r tm
Rm
= + − −− −( (( ) ) exp( ))( ) / ( )1 10
1 1 1 (37.35)
Aqui também, para m= 0 a equação 37.35 transforma-se na função monomolecular (equação 37.31); para m= 2 , na logística (37.24); para m tendendo a 1, na de Gompertz (37.27) (Figura 37.5D). É importante notar que valores de rR não podem ser comparados diretamente caso tenham sido estimados com diferentes valores de m.
Importância das Doenças de Plantas
415
37.2.6 Modelo dependente do tempoTanto no modelo monomolecular quanto nos modelos logístico e de Gompertz,
a decrescente quantidade de tecido sadio é um dos fatores que limita o crescimento de uma epidemia. No entanto, como discutido por Kiyosawa (1972), a curva de progresso da maioria das epidemias mostra uma assíntota mesmo quando a disponibilidade de tecido sadio continuar considerável. Este fato levou aquele autor a propor a equação diferencial
dx dt r t TT/ ( / )= −1 (37.36)como um novo modelo epidemiológico, onde rT é a taxa de aumento específi ca
Figura 37.5 - Modelo monomolecular: (A) taxa absoluta dx/dt versus tempo com r = 0,25 (quadrado cheio) e r = 0,15 (quadrado vazio); x0 = 0,001; (B) intensidade de doença x versus tempo com r = 0,25 (quadrado cheio) e r = 0,15 (quadrado vazio); x0 = 0,001. Modelo de Rich-ards: (C) taxa absoluta dx/dt versus tempo com r = 0,25 e m = 0 (círculo), m = 2 (quadrado cheio) e m tendendo a 1 (quadrado vazio); x0 = 0,001; (D) intensidade de doença x versus tempo com r = 0,25 e m = 0 (círculo), m = 2 (quadrado cheio) e m tendendo a 1 quadrado vazio). Modelo dependente do tempo: (E) taxa absoluta dx/dt versus tempo com r = 0,39 (quadrado cheio) e r = 0,35 (quadrado vazio); x0 = 0,001; T = 35; (F) intensidade de doença x versus tempo com r = 0,39 (quadrado cheio) e r = 0,35 (quadrado vazio); x0 = 0,001; T = 35.

Manual de Fitopatologia
416
para este modelo e T, o valor máximo de t (tempo), no qual rT = 0. A estimativa de T como o valor máximo a ser considerado é muito importante, pois após este período, os incrementos tornam-se negativos e x decresce. A plotagem de dx/dt contra o tempo produz uma curva assimétrica, com ponto de infl exão deslocado para a direita (Figura 37.5E), ao contrário do que acontece com o modelo de Gompertz (Figura 37.4E). A integral de 37.36 tem a forma
x x r t t TT= −0 1 2exp( ( / )) (37.37)O gráfi co de x versus tempo também produz uma curva em forma de S (Figura
37.5F), curva que pode se adaptar àquelas doenças que têm seu crescimento limitado mais pela diminuição da suscetibilidade do hospedeiro ou pela desfavorabilidade do clima do que pela falta de tecido sadio disponível.
37.3 EXEMPLOS E APLICAÇÕES37.3.1 Como escolher o melhor modelo?A estatística dispõe de diversos critérios para que se possa escolher o modelo
que melhor se ajuste a um determinado conjunto de dados. No entanto, o coefi ciente de determinação (R2), obtido da regressão linear entre os valores transformados da proporção de doença (variável dependente) e o tempo (variável independente), apesar de largamente empregado na literatura, não pode servir como critério (Jeger, 1986). Em compensação, o coefi ciente de determinação (R*2), obtido da regressão linear entre os valores previstos (variável dependente) e observados (variável independente), ambos sem transformação, é uma boa escolha. Além deste critério, a forma da curva da derivada (dx/dt), as estimativas do desvio padrão dos parâmetros r* e x0 e, mais importante, a plotagem do resíduo padrão (x observado menos x previsto) em função da variável independente são procedimentos estatisticamente aconselháveis (Campbell & Madden, 1990).
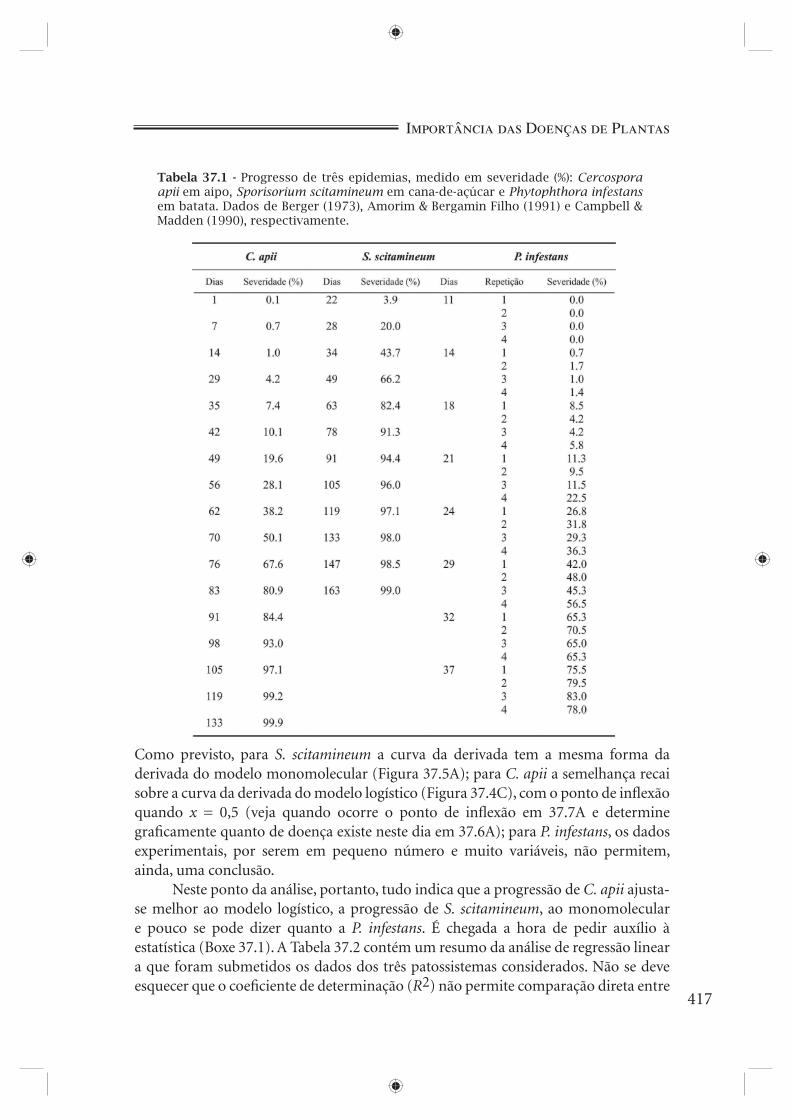
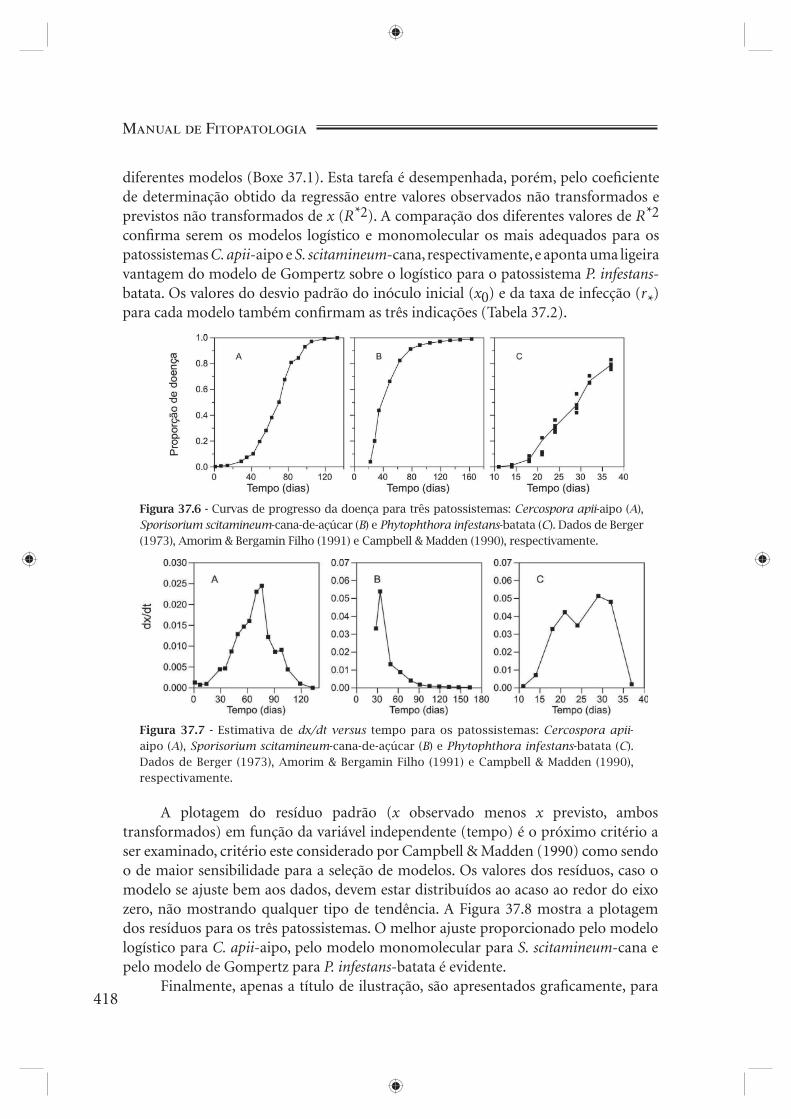
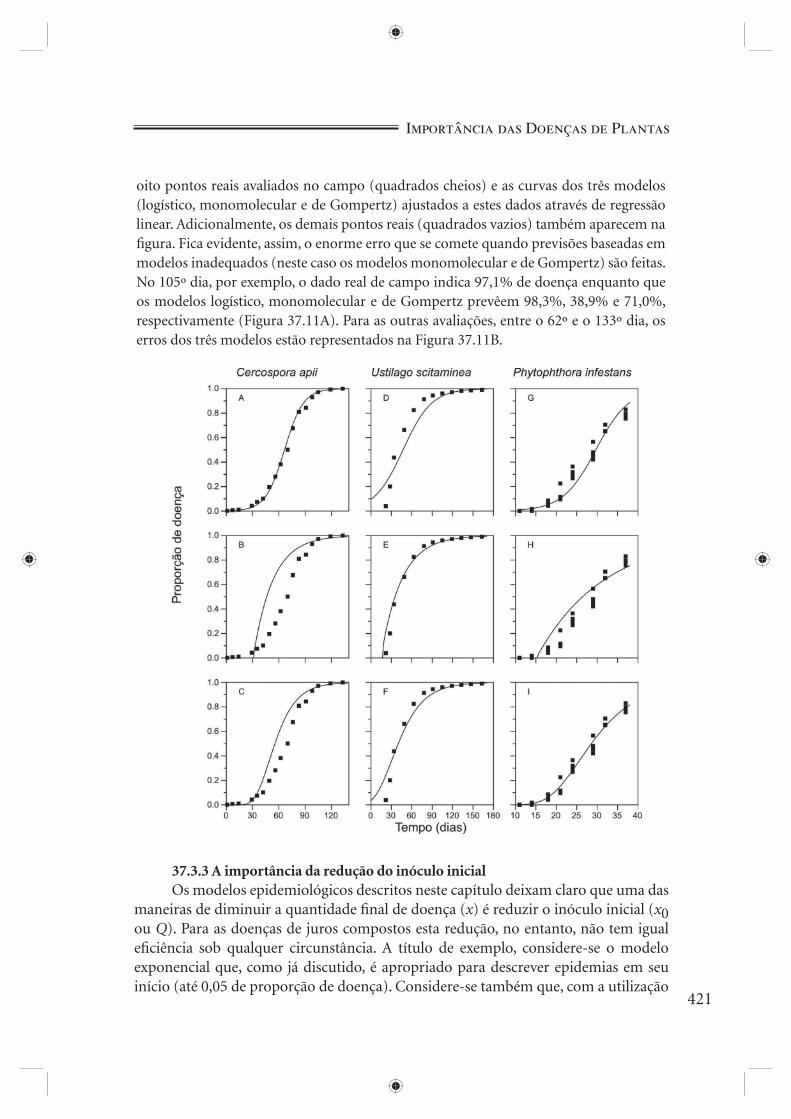
Experimentos reais servirão de exemplo. A Tabela 37.1 e a Figura 37.6 apresen-tam o progresso de três diferentes epidemias: Cercospora apii em aipo (Berger, 1973), Sporisorium scitamineum (Ustilago scitaminea) em cana-de-açúcar (Amorim & Bergamin Filho, 1991) e Phytophthora infestans em batata (Campbell & Madden, 1990). Qual seria o modelo mais apropriado para cada caso? Didaticamente, vamos nos ater aos modelos exponencial, logístico, de Gompertz e monomolecular. Antes da estatística, uma atenta observação visual dos dados constitui-se numa prática útil. As duas primeiras curvas (Figura 37.6A e 37.6B) mostram claramente uma assíntota, assíntota esta menos evidente, mas mesmo assim, perceptível na terceira curva (Figura 37.6C). A presença de assíntota é incompatível com o modelo exponencial (Figura 37.4B). A primeira e última curvas em nada se parecem com a curva típica do modelo monomolecular (Figura 37.5B): no entanto, dizer que uma ou outra está mais próxima do modelo logístico (Figura 37.4D) que do modelo de Gompertz (Figura 37.4F), ou vice-versa, seria temerário a esta altura. A segunda curva, ao contrário, fortemente se parece com a curva monomolecular (Figura 37.5B).
E que informação nos traz a plotagem de dx/dt contra o tempo (Figura 37.7)?
Importância das Doenças de Plantas
417
Como previsto, para S. scitamineum a curva da derivada tem a mesma forma da derivada do modelo monomolecular (Figura 37.5A); para C. apii a semelhança recai sobre a curva da derivada do modelo logístico (Figura 37.4C), com o ponto de infl exão quando x = 0,5 (veja quando ocorre o ponto de infl exão em 37.7A e determine grafi camente quanto de doença existe neste dia em 37.6A); para P. infestans, os dados experimentais, por serem em pequeno número e muito variáveis, não permitem, ainda, uma conclusão.
Neste ponto da análise, portanto, tudo indica que a progressão de C. apii ajusta-se melhor ao modelo logístico, a progressão de S. scitamineum, ao monomolecular e pouco se pode dizer quanto a P. infestans. É chegada a hora de pedir auxílio à estatística (Boxe 37.1). A Tabela 37.2 contém um resumo da análise de regressão linear a que foram submetidos os dados dos três patossistemas considerados. Não se deve esquecer que o coefi ciente de determinação (R2) não permite comparação direta entre
Tabela 37.1 - Progresso de três epidemias, medido em severidade (%): Cercospora apii em aipo, Sporisorium scitamineum em cana-de-açúcar e Phytophthora infestans em batata. Dados de Berger (1973), Amorim & Bergamin Filho (1991) e Campbell & Madden (1990), respectivamente.
Manual de Fitopatologia
418
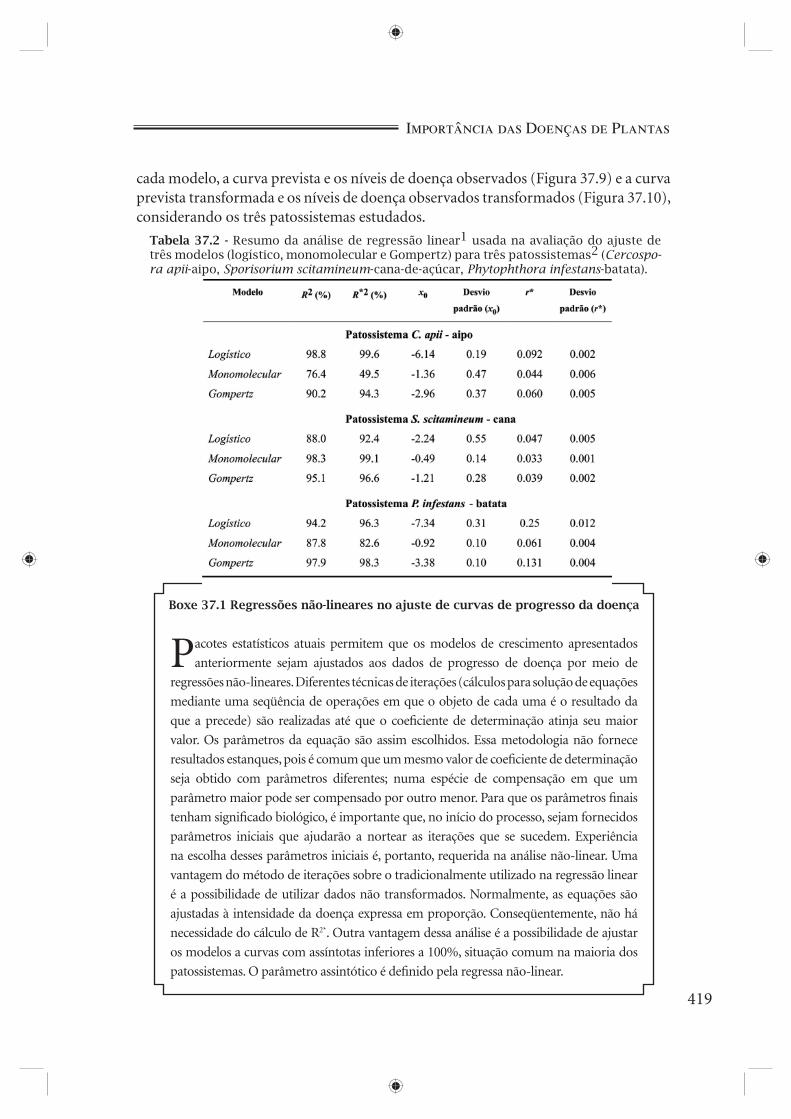
diferentes modelos (Boxe 37.1). Esta tarefa é desempenhada, porém, pelo coefi ciente de determinação obtido da regressão entre valores observados não transformados e previstos não transformados de x (R*2). A comparação dos diferentes valores de R*2 confi rma serem os modelos logístico e monomolecular os mais adequados para os patossistemas C. apii-aipo e S. scitamineum-cana, respectivamente, e aponta uma ligeira vantagem do modelo de Gompertz sobre o logístico para o patossistema P. infestans-batata. Os valores do desvio padrão do inóculo inicial (x0) e da taxa de infecção (r*) para cada modelo também confi rmam as três indicações (Tabela 37.2).
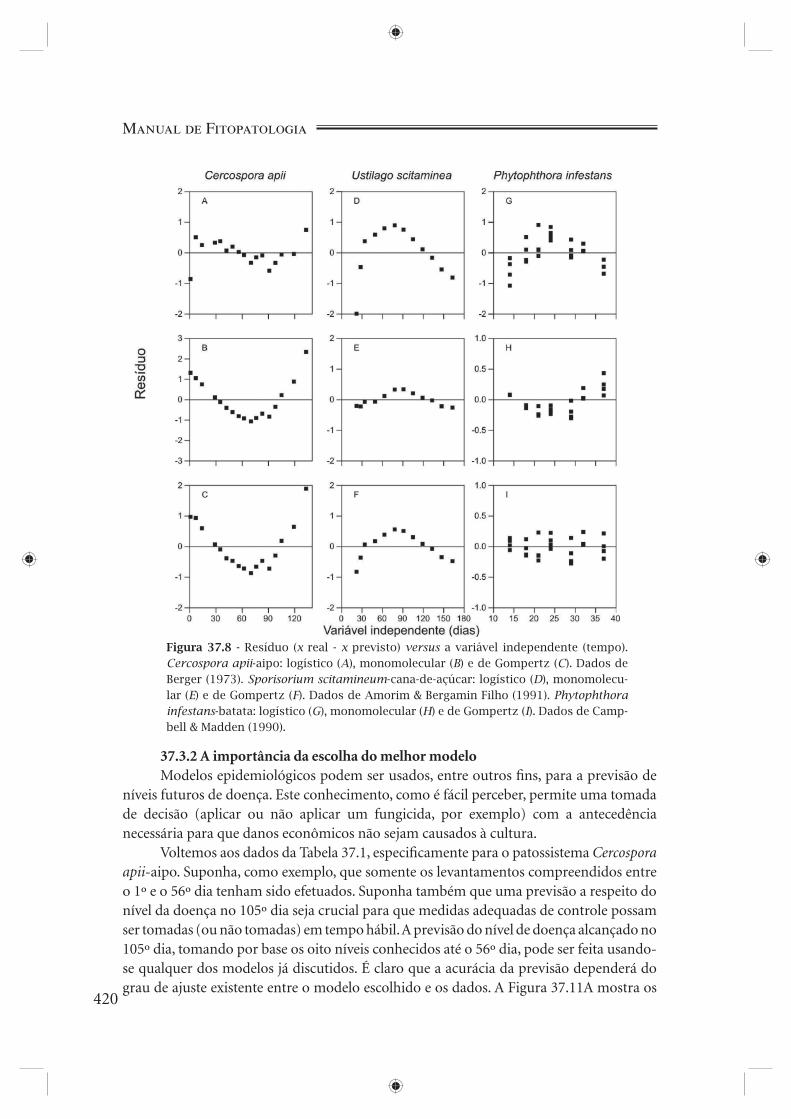
A plotagem do resíduo padrão (x observado menos x previsto, ambos transformados) em função da variável independente (tempo) é o próximo critério a ser examinado, critério este considerado por Campbell & Madden (1990) como sendo o de maior sensibilidade para a seleção de modelos. Os valores dos resíduos, caso o modelo se ajuste bem aos dados, devem estar distribuídos ao acaso ao redor do eixo zero, não mostrando qualquer tipo de tendência. A Figura 37.8 mostra a plotagem dos resíduos para os três patossistemas. O melhor ajuste proporcionado pelo modelo logístico para C. apii-aipo, pelo modelo monomolecular para S. scitamineum-cana e pelo modelo de Gompertz para P. infestans-batata é evidente.
Finalmente, apenas a título de ilustração, são apresentados grafi camente, para
Figura 37.6 - Curvas de progresso da doença para três patossistemas: Cercospora apii-aipo (A), Sporisorium scitamineum-cana-de-açúcar (B) e Phytophthora infestans-batata (C). Dados de Berger (1973), Amorim & Bergamin Filho (1991) e Campbell & Madden (1990), respectivamente.
Figura 37.7 - Estimativa de dx/dt versus tempo para os patossistemas: Cercospora apii-aipo (A), Sporisorium scitamineum-cana-de-açúcar (B) e Phytophthora infestans-batata (C). Dados de Berger (1973), Amorim & Bergamin Filho (1991) e Campbell & Madden (1990), respectivamente.
Importância das Doenças de Plantas
419
Tabela 37.2 - Resumo da análise de regressão linear1 usada na avaliação do ajuste de três modelos (logístico, monomolecular e Gompertz) para três patossistemas2 (Cercospo-ra apii-aipo, Sporisorium scitamineum-cana-de-açúcar, Phytophthora infestans-batata).
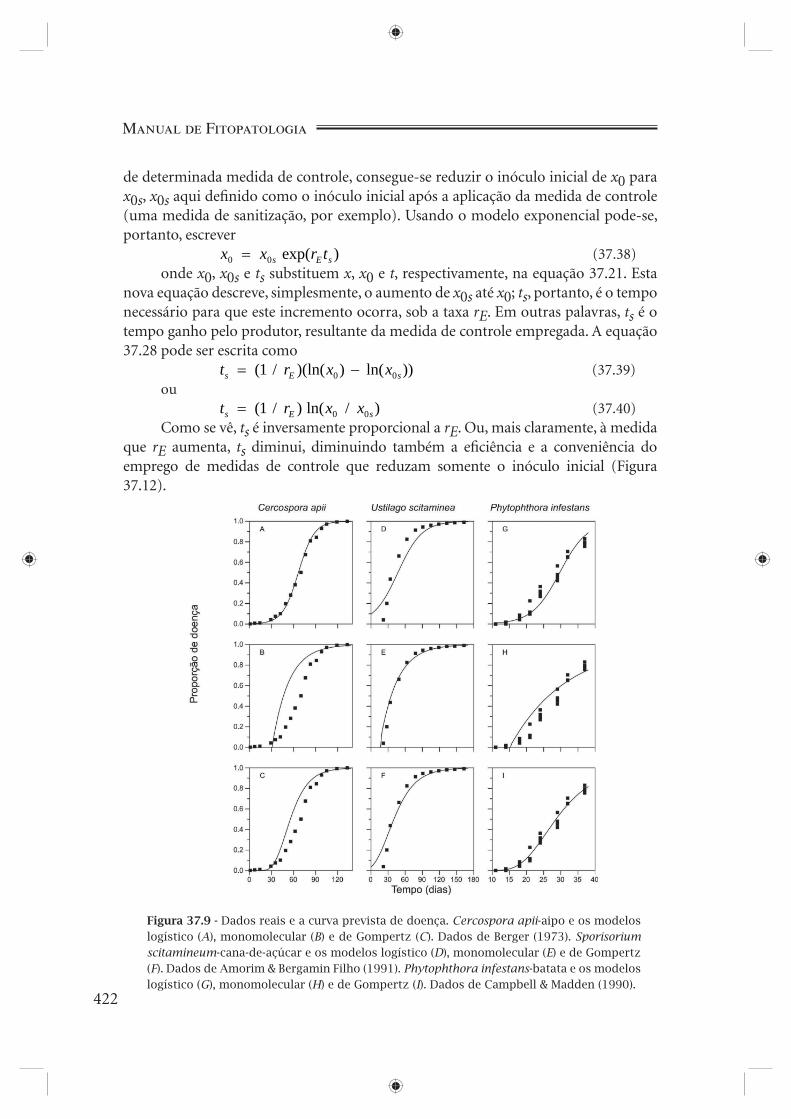
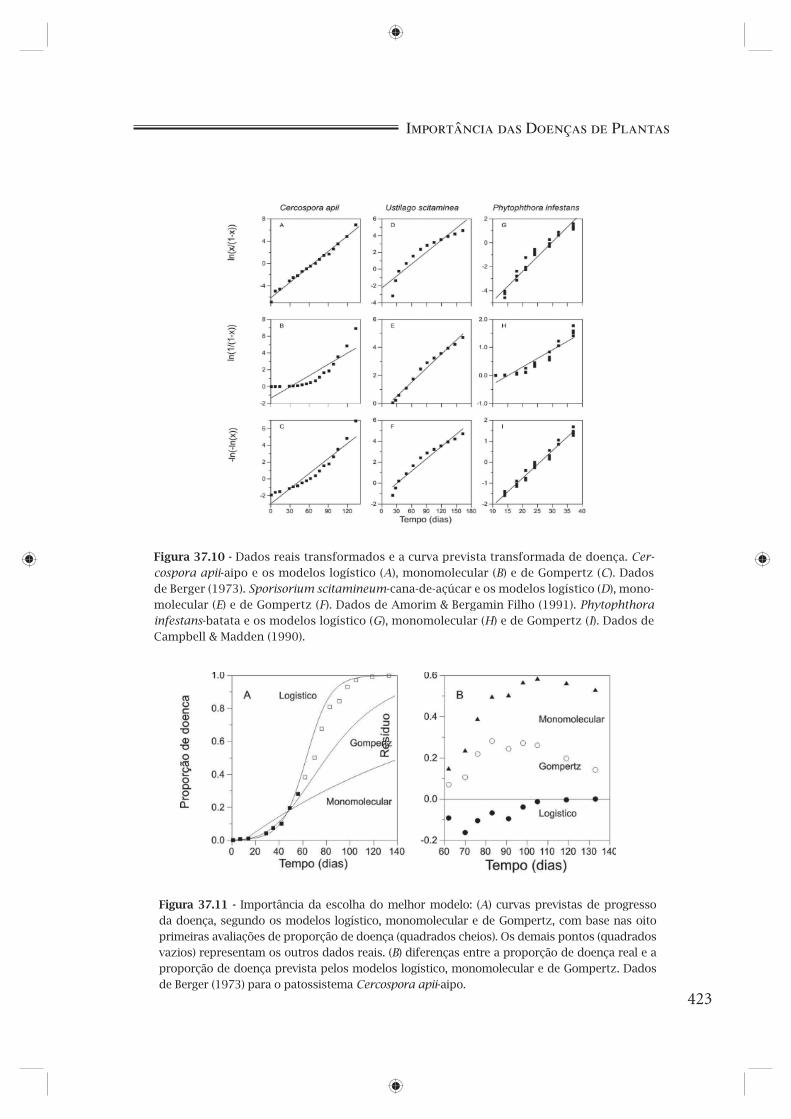
cada modelo, a curva prevista e os níveis de doença observados (Figura 37.9) e a curva prevista transformada e os níveis de doença observados transformados (Figura 37.10), considerando os três patossistemas estudados.
Boxe 37.1 Regressões não-lineares no ajuste de curvas de progresso da doença
Pacotes estatísticos atuais permitem que os modelos de crescimento apresentados
anteriormente sejam ajustados aos dados de progresso de doença por meio de
regressões não-lineares. Diferentes técnicas de iterações (cálculos para solução de equações
mediante uma seqüência de operações em que o objeto de cada uma é o resultado da
que a precede) são realizadas até que o coefi ciente de determinação atinja seu maior
valor. Os parâmetros da equação são assim escolhidos. Essa metodologia não fornece
resultados estanques, pois é comum que um mesmo valor de coefi ciente de determinação
seja obtido com parâmetros diferentes; numa espécie de compensação em que um
parâmetro maior pode ser compensado por outro menor. Para que os parâmetros fi nais
tenham signifi cado biológico, é importante que, no início do processo, sejam fornecidos
parâmetros iniciais que ajudarão a nortear as iterações que se sucedem. Experiência
na escolha desses parâmetros iniciais é, portanto, requerida na análise não-linear. Uma
vantagem do método de iterações sobre o tradicionalmente utilizado na regressão linear
é a possibilidade de utilizar dados não transformados. Normalmente, as equações são
ajustadas à intensidade da doença expressa em proporção. Conseqüentemente, não há
necessidade do cálculo de R2*. Outra vantagem dessa análise é a possibilidade de ajustar
os modelos a curvas com assíntotas inferiores a 100%, situação comum na maioria dos
patossistemas. O parâmetro assintótico é defi nido pela regressa não-linear.
Manual de Fitopatologia
420
37.3.2 A importância da escolha do melhor modeloModelos epidemiológicos podem ser usados, entre outros fi ns, para a previsão de
níveis futuros de doença. Este conhecimento, como é fácil perceber, permite uma tomada de decisão (aplicar ou não aplicar um fungicida, por exemplo) com a antecedência necessária para que danos econômicos não sejam causados à cultura.
Voltemos aos dados da Tabela 37.1, especifi camente para o patossistema Cercospora apii-aipo. Suponha, como exemplo, que somente os levantamentos compreendidos entre o 1º e o 56º dia tenham sido efetuados. Suponha também que uma previsão a respeito do nível da doença no 105º dia seja crucial para que medidas adequadas de controle possam ser tomadas (ou não tomadas) em tempo hábil. A previsão do nível de doença alcançado no 105º dia, tomando por base os oito níveis conhecidos até o 56º dia, pode ser feita usando-se qualquer dos modelos já discutidos. É claro que a acurácia da previsão dependerá do grau de ajuste existente entre o modelo escolhido e os dados. A Figura 37.11A mostra os
Figura 37.8 - Resíduo (x real - x previsto) versus a variável independente (tempo). Cercospora apii-aipo: logístico (A), monomolecular (B) e de Gompertz (C). Dados de Berger (1973). Sporisorium scitamineum-cana-de-açúcar: logístico (D), monomolecu-lar (E) e de Gompertz (F). Dados de Amorim & Bergamin Filho (1991). Phytophthora infestans-batata: logístico (G), monomolecular (H) e de Gompertz (I). Dados de Camp-bell & Madden (1990).
Importância das Doenças de Plantas
421
oito pontos reais avaliados no campo (quadrados cheios) e as curvas dos três modelos (logístico, monomolecular e de Gompertz) ajustados a estes dados através de regressão linear. Adicionalmente, os demais pontos reais (quadrados vazios) também aparecem na fi gura. Fica evidente, assim, o enorme erro que se comete quando previsões baseadas em modelos inadequados (neste caso os modelos monomolecular e de Gompertz) são feitas. No 105º dia, por exemplo, o dado real de campo indica 97,1% de doença enquanto que os modelos logístico, monomolecular e de Gompertz prevêem 98,3%, 38,9% e 71,0%, respectivamente (Figura 37.11A). Para as outras avaliações, entre o 62º e o 133º dia, os erros dos três modelos estão representados na Figura 37.11B.
37.3.3 A importância da redução do inóculo inicialOs modelos epidemiológicos descritos neste capítulo deixam claro que uma das
maneiras de diminuir a quantidade fi nal de doença (x) é reduzir o inóculo inicial (x0 ou Q). Para as doenças de juros compostos esta redução, no entanto, não tem igual efi ciência sob qualquer circunstância. A título de exemplo, considere-se o modelo exponencial que, como já discutido, é apropriado para descrever epidemias em seu início (até 0,05 de proporção de doença). Considere-se também que, com a utilização
Manual de Fitopatologia
422
de determinada medida de controle, consegue-se reduzir o inóculo inicial de x0 para x0s, x0s aqui defi nido como o inóculo inicial após a aplicação da medida de controle (uma medida de sanitização, por exemplo). Usando o modelo exponencial pode-se, portanto, escrever
x x r ts E s0 0= exp( ) (37.38)onde x0, x0s e ts substituem x, x0 e t, respectivamente, na equação 37.21. Esta
nova equação descreve, simplesmente, o aumento de x0s até x0; ts, portanto, é o tempo necessário para que este incremento ocorra, sob a taxa rE. Em outras palavras, ts é o tempo ganho pelo produtor, resultante da medida de controle empregada. A equação 37.28 pode ser escrita como t r x xs E s= −( / )(ln( ) ln( ))1 0 0 (37.39)
ou t r x xs E s= ( / ) ln( / )1 0 0 (37.40)
Como se vê, ts é inversamente proporcional a rE. Ou, mais claramente, à medida que rE aumenta, ts diminui, diminuindo também a efi ciência e a conveniência do emprego de medidas de controle que reduzam somente o inóculo inicial (Figura 37.12).
Figura 37.9 - Dados reais e a curva prevista de doença. Cercospora apii-aipo e os modelos logístico (A), monomolecular (B) e de Gompertz (C). Dados de Berger (1973). Sporisorium scitamineum-cana-de-açúcar e os modelos logístico (D), monomolecular (E) e de Gompertz (F). Dados de Amorim & Bergamin Filho (1991). Phytophthora infestans-batata e os modelos logístico (G), monomolecular (H) e de Gompertz (I). Dados de Campbell & Madden (1990).
Importância das Doenças de Plantas
423
Figura 37.10 - Dados reais transformados e a curva prevista transformada de doença. Cer-cospora apii-aipo e os modelos logístico (A), monomolecular (B) e de Gompertz (C). Dados de Berger (1973). Sporisorium scitamineum-cana-de-açúcar e os modelos logístico (D), mono-molecular (E) e de Gompertz (F). Dados de Amorim & Bergamin Filho (1991). Phytophthora infestans-batata e os modelos logístico (G), monomolecular (H) e de Gompertz (I). Dados de Campbell & Madden (1990).
Figura 37.11 - Importância da escolha do melhor modelo: (A) curvas previstas de progresso da doença, segundo os modelos logístico, monomolecular e de Gompertz, com base nas oito primeiras avaliações de proporção de doença (quadrados cheios). Os demais pontos (quadrados vazios) representam os outros dados reais. (B) diferenças entre a proporção de doença real e a proporção de doença prevista pelos modelos logístico, monomolecular e de Gompertz. Dados de Berger (1973) para o patossistema Cercospora apii-aipo.

Manual de Fitopatologia
424
Figura 37.12 - Relação entre o atraso na epidemia (ts) em função de diferentes relações de sanitização (10, 102, 103, 104, 105 e 106 - as diferentes curvas de baixo para cima) e diferentes taxas exponenciais de infecção (rE).
A equação 37.40 deixa claro que os valores reais de x0 e x0s não infl uenciam o atraso (ts) conseguido na epidemia. O que importa é a relação (x0/x0s), também chamada de relação de sanitização (Vanderplank, 1963). Assim, consegue-se mesmos valores de ts com diferentes combinações de x0 e x0s como, por exemplo, 0,01 e 0,001 ou 0,001 e 0,0001: em ambos os casos a relação de sanitização é 10.
Boxe 37.2 - Seria rE dependente de x
o?
No item 37.3.3 foi defi nida a relação de sanitização como sendo
t r x xs E s= ( / ) ln( / )1 0 0 (37.40)
para os estádios iniciais da epidemia. Esta relação pressupõe ser a
taxa rE independente da quantidade de inóculo inicial x
o. Seria esta suposição
verdadeira? Berger (1988) garante que não. Usando seus próprios dados (Plaut &
Berger, 1981) para os patossistemas Cercospora arachidicola-amendoim, Botrytis
cinerea-begônia e Uromyces appendiculatus-feijoeiro, aquele autor mostrou que
epidemias que começaram com inóculos menores progrediram a taxas maiores
(três níveis de inóculo foram usados). Estes resultados, como enfatiza Berger
(1988), são de grande importância na análise de epidemias e no controle de
doenças. Caso este fenômeno seja generalizado, medidas de controle que atuem
unicamente no inóculo inicial, como o uso de sementes tratadas ou sadias,
podem ter efi ciência menor do que a imaginada até aqui.
Outros patossistemas que exibem o mesmo fenômeno são milho-Cochliobolus
heterostrophus (Helminthosporium maydis, Gregory et al., 1981) e trigo-Erysiphe
graminis (Rouse et al., 1981).

Importância das Doenças de Plantas
425
37.4 BIBLIOGRAFIA CONSULTADAAmorim, L. & Bergamin Filho, A. Sugarcane smut development models: I. Annual curves of disease progress.
Zeitschrift fur Pfl anzenkrankheiten und Pfl anzenschutz 98: 605-611, 1991.
Berger, R.D. Infection rates of Cercospora apii in mixed populations of susceptible and tolerant celery.
Phytopathology 63: 535-537, 1973.
Berger, R.D. Comparison of the Gompertz and logistic equations to describe plant disease progress.
Phtytopathology 71: 716-719, 1981.
Berger, R.D. The analysis of effects of control measures on the development of epidemics. In Kranz, J. & Rotem, J.
(ed.). Experimental Techniques in Plant Disease Epidemiology. Berlin, Springer, 1988. p. 137-151.
Campbell, C.L. & Madden, L.V. Introduction to Plant Disease Epidemiology. New York, John Wiley, 1990.
Gompertz, B. On the nature of the function expressive of the law of human mortality. Philos. Trans. R. Soc.
London 36: 513-585, 1825.
Gregory L.V.; Ayers, J.E.; Nelson, R.R. Reliability of apparent infection rates in epidemiological research. Journal
of Phytopathology 100: 135-142, 1981.
Jeger, M.J. The potential of analytic compared with simulation approaches to modeling in plant disease
epidemiology. In: Leonard, K.J. & Fry, W.E. (ed.). Plant Disease Epidemiology. Population Dynamics and
Management. New York, Macmillan, 1986. p.255-281.
Kiyosawa, S. Mathematical studies on the curve of disease increase - a technique for forecasting epidemic
development. Annals of the Phytopathological Society of Japan 38: 30-40, 1972.
Newby, J.C. Mathematics for the Biological Sciences. Oxford, Clarendon, 1980.
Pfender, W.F. Monocyclic and policyclic root diseases: distinguishing between the nature of the disease cycle and
the shape of the disease progress curve. Phytopathology 72: 31-32, 1982.
Plaut, J.L. & Berger, R.D. Infection rates in three pathosystem epidemics initiated with reduced disease severities.
Phytopathology 71: 917-921, 1981.
Richards, F.J. A fl exible growth function for empirical use. J. Exp. Bot. 10: 290-300, 1959.
Rouse, D.I.; MacKenzie, D.R.; Nelson, R.R. A relationship between initial inoculum and apparent infection rate
in a set of disease progress data for powdery mildew on wheat. Journal of Phytopathology 100: 143-149,
1981.
Vanderplank, J.E. Plant Diseases: Epidemics and Control. New York, Academic Press, 1963.
Verhulst, P.F. Notice sur la loi que la population suit dans son accroissement. Coord. Math. Phys. 10: 112-121,
1838.
Waggoner, P.E. Progress curves of foliar diseases: their interpretation and use. In Leonard, K.J. & Fry, W.E. (ed.).
Plant Disease Epidemiology. Population Dynamics and Management. New York, Macmillan, 1986. p.3-27.


















