
© Agostinho Franklim Pinto Marques Braga · competência física, técnica e psicológica...
93
Transcript of © Agostinho Franklim Pinto Marques Braga · competência física, técnica e psicológica...



© Agostinho Franklim Pinto Marques
Execução: Minhografe, Artes Gráficas, Lda. — BragaISBN: 978-989-20-7178-7Depósito Legal: 418449/16

ÍNDICE
Página
Prólogo 51. Introdução 92. Marcadores bioquímicos associados à função muscular 10
2.1. A alta competição e a função muscular 103. Dor muscular 194. Marcadores bioquímicos e fadiga 235. Marcadores bioquímicos associados ao stress oxidativo 31
5.1. Espécies oxidantes e radicais livres 315.1.1. Marcadores de lesão oxidativa 34
5.2. A Mecanismos de produção de radicais livres 355.2.1. Cadeia de transporte de eletrões (mitocôndria) 355.2.2. Xantina oxidase (metabolismo purínico) 365.2.3. Outras fontes 38
5.3. Capacidade antioxidante 395.4. Exercício físico e stress oxidativo.
Avaliação laboratorial 435.4.1. Futebol 445.4.2. Escalada indoor 505.4.3. Exercício em tapete rolante 545.4.4. Motocross 575.4.5. Canoagem 605.4.6. Alpinismo 62
5.5. Suplementação de antioxidantes 666. Conclusão 717. Referências bibliográficas 75Epílogo 89Discurso de presentación 91


5
PRÓLOGOEx.mo. Sr. Presidente
Ex.mos e Il.mos Sras e Srs AcadémicosSenhoras e Senhores
Caras amigas e caros amigos
Gostaria, antes de mais, de manifestar perante vós o meu contentamento por ter sido aceite, em nome individual, para fazer parte desta Ilustre Instituição que engloba e representa os melhores conhecimentos e saberes relativos ao farmacêutico e às Ciências Farmacêuticas.
É sempre um motivo de orgulho e de satisfação pessoal quando alguém reconhece o caminho que percorremos em defesa de algo ou orientados a um objetivo que acrescenta valor, seja de natureza científica, politico ou institucional, à sociedade e à cidadania.
E fico mais orgulhoso ainda quando uma ímpar Instituição, como a Academia de Farmácia da Galiza, me aceita como alguém capaz de caminhar lado a lado com Académicos de reconhecido mérito científico, político e profissional, nos distintos campos ou áreas de intervenção de cada um e de todos.
Sinto-me honrado por estar aqui, hoje, junto de vós, e permitam-me que nestas circunstâncias e antecipadamente, expresse o meu comprometimento ético, deontológico e livre perante esta camara, em defesa dos ideais, objetivos e dos valores que presidem a esta Academia.
As minhas relações pessoais, institucionais e de intercâmbio científico e de investigação com Espanha, de uma forma mais profunda e constante, são já de longa data, e reportam-se ao tempo dos meus estudos para a obtenção do grau de Doutor.
Na realidade, a parte experimental conducente à apresentação das minhas provas de doutoramento foi quase toda ela realizada em Madrid, na Faculdade de Medicina da Universidade Autónoma, sob orientação direta de uma dupla de relevantes e prolíficos cientistas e investigadores, o Prof. Doutor Antonio Sillero e a Doutora Maria

6
Antonia Sillero. O trabalho então realizado, e desenvolvido na área da neuroquímica, versou a purificação, identificação e caracterização de uma enzima presente no tecido cerebral, a 5´nucleotidase IMP-GMP especifica. Foi uma experiência intensa, sob o ponto de vista científico, e enriquecedora sob o ponto de vista pessoal e humano. Foi um tempo de aprendizagem e de crescimento.
Na sequência deste percurso, passei a fazer parte da Geração F2, da Saga Científica Española de Severo Ochoa, bioquímico espanhol, pioneiro dos estudos de síntese de ácidos nucleicos e Nobel de Fisiologia e Medicina, em 1959, pelos seus trabalhos em RNA, patrocinada pela Fundación Carmen y Severo Ochoa. Esta Geração F2 engloba os investigadores identificados pelos membros da designada Geração F1, a qual é constituída pelo conjunto de cientistas e de investigadores que trabalharam diretamente com Severo Ochoa, no seu laboratório nos Estados Unidos, enfim, seus discípulos diretos.
Parece que o ditado português ”de Espanha, nem bom vento nem bom casamento” não tem tido qualquer correspondência com as minhas relações pessoais, institucionais e fraternais com este país, em qualquer momento da minha vida.
Após terminar os meus estudos e obtido o grau de Doutor, a minha vida académica e científica sofreu alguns desenvolvimentos, nomeadamente na direção a seguir, no âmbito da investigação científica.
De facto, após alguns anos passados a realizar trabalho de investigação na área da neuroquímica, o foco de incidência da minha investigação passou a ser orientado para o estudo dos efeitos do exercício físico e a alta competição sobre o organismo humano, com enfase particular no stress oxidativo, e a sua avaliação e monitorização laboratorial.
A aplicabilidade prática dos resultados de investigação foi razão primeira desta opção. Mas, não posso omitir, nem deixar de ter também em consideração o facto de algumas Instituições de âmbito desportivo e competitivo, seguindo o parecer e a opinião de membros dos seus staff clínicos, terem solicitado a minha envolvência e o meu apoio laboratorial e científico ao trabalho desenvolvido e a desenvolver, no

7
âmbito da atividade, do exercício físico e do rendimento desportivo de atletas e desportistas de alta competição.
E ainda hoje mantenho esta linha de atuação primordial.Quando em Junho fui confrontado e presenteado com a agradável
notícia da vossa distinta resolução, e apercebendo-me da necessidade de perante vós apresentar um discurso de apresentação e de aceitação do cargo para o qual fui aceite, questionei-me sobre a temática a abordar.
Tenho sempre alguma dificuldade na livre escolha de temas a desenvolver, especialmente em situações como esta, em que a diversidade e amplitude de saberes cultural e científico de quem me escuta e a quem me dirijo são abrangentes. Não é fácil encontrar um tema inclusivo que englobe ou interesse à grande maioria dos ilustres académicos desta Instituição.
Quando parei um pouco para tentar encontrar um caminho, sentei-me ao computador e fui alertado por mensagem emanada do Research Gate para uma nova citação sobre um artigo publicado em 2010, na revista “European Journal of Applied Physiology”, pela equipa de investigação que integro, intitulado “Impact of Loughborough Intermittent Shuttle Test versus soccer match on physiological, bioche-mical and neuromuscular parameters”.
Num ano de competição europeia de futebol, surgir no sistema de alertas de citação de artigos, uma citação relacionada com futebol, não me pareceu nada de muito estranho, de muito inusitado. Nestas alturas os trabalhos nestas áreas de investigação e de estudo são mais lidos e mais procurados pela comunidade científica.
Mas foi extremamente curioso, o momento, a oportunidade, em especial num particular ano como este, em que o desporto de elite competitivo impera, com o Campeonato Europeu de Futebol (Euro 2016), em Paris, França, com os Jogos Olímpicos, no Rio de Janeiro, Brasil, e mais ainda quando o futebol português conquistou o seu primeiro grande troféu internacional, enquanto seleção de Campeão Europeu de Futebol
Pensei então em discorrer um pouco sobre a envolvência da minha investigação, mostrando alguns dos resultados obtidos por

8
mim e pela equipa que integro, publicados ou ainda não, em estudos efetuados no âmbito da investigação e monitorização laboratorial de atletas de alta competição, não só de futebol, mas também, de outras modalidades desportivas.
Espero, pois, conseguir transmitir-vos um pouco do meu trabalho, um pouco do percurso da minha vida como académico e como investigador, enfim, um pouco da minha vida participativa em prol do desporto, em defesa da saúde, da qualidade de vida e da cidadania.

9
1. INTRODUÇÃO
O sedentarismo constitui uma das principais fontes ou origem de muitos dos problemas de saúde que grassam nos dias de hoje. A prática de atividade ou exercício físico tem como fim primeiro o aperfeiçoamento da capacidade física, visando a melhoria e a promoção da saúde, do bem-estar e da qualidade de vida individual e coletiva. Variados são os estudos que comprovam as vantagens significativas em termos de benefícios na prevenção e melhoria da saúde da prática regular de exercício físico.
São hoje consensuais os benefícios que o exercício físico aporta consigo sobre distintas funções e processos fisiológicos (e.g. função imunitária, função muscular, desenvolvimento ósseo), bem como, sobre distintas patologias, entre as quais, pela sua dimensão e importância nos dias de hoje, as doenças do foro cardiovascular, a diabetes e a obesidade. De realçar ainda os benefícios que aporta a nível de bem-estar cognitivo e psicológico, nomeadamente na melhoria dos estados de depressão e ansiedade.
O conceito de desporto depreende uma prática de exercício físico que se desenvolve em ambiente regulamentado, tendo como fim, o alcançar de resultados concretos e em termos competitivos. Este conceito abarca em si mesmo as noções de capacidade e de competência física, técnica e psicológica individuais e/ou coletivos, vocacionados ao fim pretendido. As exigências são mais profundas e mais fortes que as requeridas na simples prática de atividade física, mais ainda quando o desporto se enquadra em ambientes de alta competição ou de elite. A alta competição exige que se vá cada vez mais além de si mesmo, que se ultrapassem os limites individuais de cada um até atingir um grau de excelência individual e coletiva.

10
2. MARCADORES BIOQUÍMICOS ASSOCIADOS À FUNÇÃO MUSCULAR
O tecido muscular é um dos tecidos do organismo mais suscetíveis a lesões ou traumas provocados pelo exercício físico. A prática de exercício físico com uma acentuada contração excêntrica, os exercícios de força, os realizados até à exaustão e/ou os inabituais são exemplos concretos de fatores causais desses processos de lesão.
A necessidade de prevenir ou de atempadamente avaliar e diagnosticar a existência de micro ou de qualquer outro tipo de lesões que possam de alguma maneira condicionar o desempenho desportivo ou limitar o tempo de inatividade associado particularmente em alta competição é de extrema importância.
2.1. A alta competição e a função muscularQualquer alteração ou modificação da função muscular condiciona
necessariamente o rendimento ou a performance desportiva de um jogador de alta competição.
A determinação laboratorial de marcadores bioquímicos associa-dos à função muscular constitui um dos parâmetros que podem ser utilizados na monitorização individual dos atletas de alta competição, permitindo uma melhor avaliação funcional e do desempenho despor-tivo dos mesmos. O conhecimento dos resultados destes marcadores não se esgota, contudo, neste âmbito, antes sim, podem constituir-se como um dado suplementar a ter consideração aquando do planea-mento e implementação dos processos de treino, nomeadamente no que à intensidade de esforço de um jogador ou de uma equipa de alta competição diz respeito.
No âmbito do acompanhamento e avaliação do desempenho e rendimento desportivo de uma equipa de futebol profissional de alta competição, procedeu-se ao estudo e monitorização laboratorial de variados parâmetros bioquímicos relacionados com a função muscular.
O futebol é uma modalidade ou desporto de alta competição com uma envolvência de atividade física de intensidade considerável, com potencial e significativo impacto no sistema muscular.

11
Uma das características mais identificativas desta modalidade desportiva consiste precisamente na notável variabilidade de inten-sidade, intermitência e de duração dos esforços efetuados em espaços de tempo muito curtos, que se verificam durante a realização dos jogos ou dos períodos de treino dos atletas/jogadores. As características de intermitência e variabilidade dos esforços físicos são causas potenciais dos níveis consideráveis de microlesões musculares associados a esta modalidade desportiva.
Participaram 27 atletas de uma equipa de futebol (25,7 ± 2,4 anos), que durante a realização do mesmo competiam regularmente a nível nacional, na 1ª Liga de Futebol português e, a nível internacional, na UEFA Champions League.
As colheitas de material biológico para determinação dos parâmetros analíticos selecionados, no âmbito do estudo em questão, foram obtidas em 3 tempos diferentes. A primeira colheita foi realizada no mês de agosto (pré-temporada), 7 dias após a realização dos primeiros treinos conducentes à preparação do início da temporada futebolística. As colheitas seguintes tiveram lugar nos meses de outubro (última semana) e de dezembro, 48 horas após realização dos jogos correspondentes à 1ª liga portuguesa e da Champions League, respetivamente. As colheitas foram efetuadas em período de jejum. Durante todo este período, para além dos jogos de competição, todos os atletas executaram treinos diários e, em alguns casos, bidiarios com a duração média de 2 horas.
Vários são, normalmente, os marcadores plasmáticos utilizados, sob o ponto de vista laboratorial, no estudo indireto de integridade e da lesão muscular, como a Creatina fosfocínase (CPK), o Aspartato aminotransférase (AST), a Aldolase (ALD), a Lactato desidrogenase (LDH), a Mioglobina (Mb).
Neste estudo, contudo, utilizaram-se como marcadores plasmáti-cos de integridade e de lesão muscular, a determinação das atividades plasmáticas das enzimas Creatina fosfocínase e Aspartato aminotrans-férase.
A CPK é uma enzima dimérica, constituído por 3 isoenzimas citosólicas, CPK-BB, CPK-MB e CPK-MM, sendo a forma MM a mais

12
abundante no músculo. Catalisa, reversivelmente, e à custa de ATP, a conversão de creatinina em creatinina-fosfato, a qual atua como reserva de energia em tecidos de grande e rápida utilização de ATP, como é o tecido muscular.
A CPK é considerada como o/um dos melhores marcadores plasmáticos de lesão muscular. Níveis plasmáticos aumentados desta enzima apontam para as lesões relacionadas com o exercício físico. Quanto maior for a intensidade do exercício ou o dano provocado, maior será o aumento dos níveis de CPK. Sendo o futebol uma atividade de grande envolvimento e sobrecarga muscular, pode este esforço ser causa de rutura das estruturas celulares, e de possível e subsequente fragilidade e diminuição da força muscular, libertando-se em consequência a CPK para a corrente sanguínea. Os níveis basais plasmáticos deste marcador em atletas de elite são superiores aos correspondentes de indivíduos sedentários ou de atletas menos treinados e evoluídos, sugerindo serem, estes valores, condicionados pelos processos de treino, nomeadamente pela sua intensidade e duração. O conhecimento dos valores plasmáticos de CPK pode assim constituir um valioso contributo a ser considerado aquando do planeamento, monitorização e modelação do esforço de treino de atletas. No entanto, a interpretação isolada deste parâmetro com este fim, deve ter sempre em consideração o facto de o comportamento da CPK apresentar grande variabilidade de resposta individual, face a um efeito causal idêntico ou de mesma intensidade.
No estudo efetuado pretendeu-se observar a evolução destes marcadores ao longo dos meses de monitorização, nomeadamente no início da temporada desportiva e dos treinos iniciais (agosto), e nos meses de outubro e de dezembro.
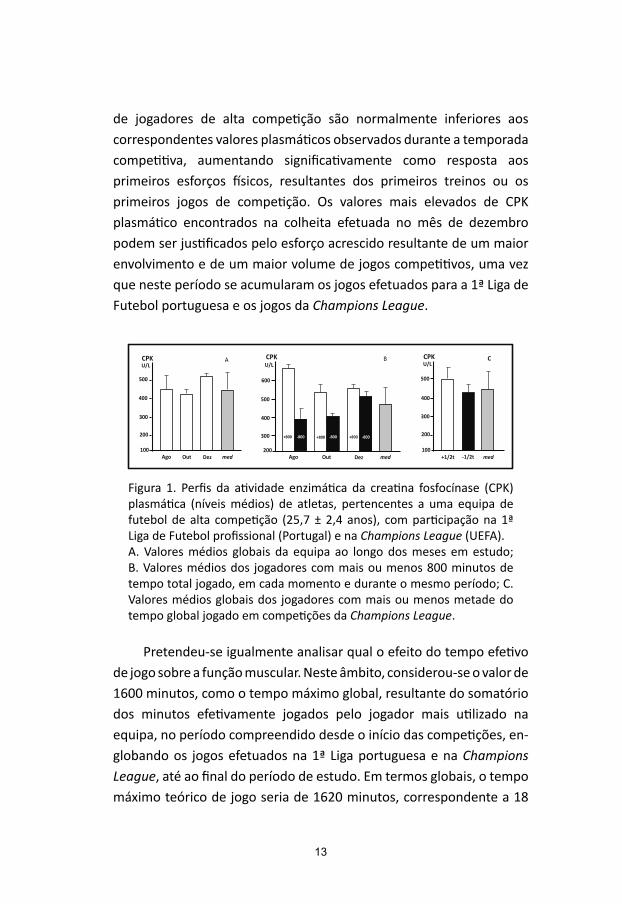
Os resultados obtidos (Figura 1A) mostram que os níveis plasmáticos de CPK relativos ao mês de agosto são mais elevados que os observados no mês de outubro e inferiores aos do mês de dezembro. Os valores médios globais de todos os jogadores da equipa em todo este período foram de 457 ± 231 UI/L.
Em períodos de pré-época ou de repouso, os valores de CPK
13
de jogadores de alta competição são normalmente inferiores aos correspondentes valores plasmáticos observados durante a temporada competitiva, aumentando significativamente como resposta aos primeiros esforços físicos, resultantes dos primeiros treinos ou os primeiros jogos de competição. Os valores mais elevados de CPK plasmático encontrados na colheita efetuada no mês de dezembro podem ser justificados pelo esforço acrescido resultante de um maior envolvimento e de um maior volume de jogos competitivos, uma vez que neste período se acumularam os jogos efetuados para a 1ª Liga de Futebol portuguesa e os jogos da Champions League.
Figura 1. Perfis da atividade enzimática da creatina fosfocínase (CPK) plasmática (níveis médios) de atletas, pertencentes a uma equipa de futebol de alta competição (25,7 ± 2,4 anos), com participação na 1ª Liga de Futebol profissional (Portugal) e na Champions League (UEFA). A. Valores médios globais da equipa ao longo dos meses em estudo; B. Valores médios dos jogadores com mais ou menos 800 minutos de tempo total jogado, em cada momento e durante o mesmo período; C. Valores médios globais dos jogadores com mais ou menos metade do tempo global jogado em competições da Champions League.
Pretendeu-se igualmente analisar qual o efeito do tempo efetivo de jogo sobre a função muscular. Neste âmbito, considerou-se o valor de 1600 minutos, como o tempo máximo global, resultante do somatório dos minutos efetivamente jogados pelo jogador mais utilizado na equipa, no período compreendido desde o início das competições, en-globando os jogos efetuados na 1ª Liga portuguesa e na Champions League, até ao final do período de estudo. Em termos globais, o tempo máximo teórico de jogo seria de 1620 minutos, correspondente a 18
Fig.1. Perfil da atividade enzimática da creatina fosfocínase (CPK) plasmática (níveis médios) de atletas, pertencentes a uma equipa de futebol de alta competição (25,7 ± 2,4 anos), com participação na 1ª Liga de Futebol profissional (Portugal) e na Champions League (UEFA). A. Valores médios globais da equipa ao longo dos meses em estudo; B. Valores médios dos jogadores com mais ou menos 800 minutos de tempo total jogado, em cada momento e durante o mesmo período; C. Valores médios globais dos jogadores com mais ou menos metade do tempo global jogado em competições da Champions League.
Fig 1
Ago med Out Dez
200
400
500
CPK U/L
300
A
+1/2t -1/2t med
200
400
500
CPK U/L
300
C CPK U/L
300
500
600
400
+800 +800 +800 -800 -800 -800
Ago med Out Dez
B
100 200 100

14
jogos competitivos efetivamente realizados. Tendo esse valor presente, adotou-se como valor de “cut-off” o tempo de 800 minutos de jogo efetivo, i.e. metade do tempo máximo de jogo efetivo, realizado pelo jogador mais utilizado. Quaisquer situações de lesões visíveis ou diagnos-ticadas durante ou após a realização dos jogos vivenciadas por qualquer jogador constituíram critério de exclusão deste perfil de análise.
Como se pode ver na Figura 1B, onde se descriminam os dados médios globais obtidos nestas condições, no primeiro mês verificou-se um mais acentuado aumento dos níveis plasmáticos de CPK nos jogadores com maior tempo global efetivo de jogo, os quais se foram atenuando e aproximando daqueles com menor tempo, ao longo desse período de estudo.
Uma explicação para esse perfil enzimático, prende-se com o facto de os jogadores mais efetivos na equipa, em termos de jogo jogado, terem desde logo, e possivelmente, iniciado os treinos com maior empenhamento e maior esforço muscular buscando a melhor forma possível, de que resultou um aumento mais significativo da libertação de CPK. À medida que a atividade prosseguia verificou-se a adaptação muscular ao esforço, resultando daí a diminuição progressiva dos valores de CPK até à estabilização dos seus níveis plasmáticos. Por outro lado, nos jogadores com menor tempo de jogo efetivo, observou-se, desde o início da temporada, um aumento gradual dos níveis plasmáticos de CPK, atingindo, no mês de dezembro, valores próximos dos encontrados nos jogadores mais utilizados. Estes resultados podem ser explicados por um mais lento e gradual envolvimento nos processos de treino que se traduz num menor grau de performance e de rendimento desportivo, quando comparados com os jogadores mais utilizados.
Do mesmo modo, considerando apenas os jogos realizados em partidas no âmbito da Champions League, e utilizando o mesmo critério de metade tempo de jogo efetivo (Figura 1C), verificou-se que os jogadores mais utilizados em termos médios apresentaram valores de CPK plasmáticos mais elevados. Uma fundamentação plausível para estes dados pode ser encontrada e justificada pelo maior esforço despendido aquando realização desses mesmos jogos.

15
Uma outra enzima utilizada igualmente como marcador da função muscular e que acompanha o perfil da CPK, é a Aspartato aminotransférase. A AST é uma enzima que faz parte do painel de diagnóstico laboratorial do perfil hepático juntamente com a Alanina aminotransférase (ALT) (bem como, como determinação complementar do perfil cardíaco), mas ao contrário desta, existe no citoplasma das células musculares em quantidade apreciável.
Como se pode observar na Figura 2A, o perfil dos níveis plasmáticos desta enzima segue o perfil da atividade de CPK anteriormente descrita, uma vez mais corroborando, por um lado, a adaptação do comportamento e da fisiologia muscular ao exercício e à atividade física e, por outro, a diferenciação entre os jogadores com maior ou menor carga temporal de jogos.
Verifica-se, de igual modo, uma diminuição dos níveis plasmáticos de AST ao longo do tempo de realização deste estudo, bem como, a diferenciação entre os diferentes grupos de jogadores, onde se observam níveis plasmáticos mais elevados deste marcador, naqueles que mais tempo foram utilizados, quer globalmente (Figura 2B) quer a nível da Champions League (Figura 2C).
Figura 2. Perfis da atividade enzimática da AST plasmática (níveis médios) de 27 atletas, pertencentes a uma equipa de futebol de alta competição (25,7 ± 2,4 anos), com participação na 1ª Liga de Futebol profissional (Portugal) e na Champions League (UEFA). A. Valores médios de AST plasmáticos da equipa ao longo dos meses em estudo; B. Valores médios de AST plasmáticos de jogadores com mais ou menos 800 minutos de tempo total jogado, durante o mesmo período; C. Valores médios de AST plasmáticos de jogadores com mais ou menos metade do tempo global jogado em competições da Champions League.
Fig.2. Perfil da atividade enzimática da Aspartato aminotransferase (AST) plasmática (níveis médios) de atletas, pertencentes a uma equipa de futebol de alta competição (25,7 ± 2,4 anos), com participação na 1ª Liga de Futebol profissional (Portugal) e na Champions League (UEFA). A. Valores médios globais da equipa ao longo dos meses em estudo; B. Valores médios dos jogadores com mais ou menos 800 minutos de tempo total jogado, em cada momento e durante o mesmo período; C. Valores médios globais dos jogadores com mais ou menos metade do tempo global jogado em competições da Champions League.
Fig 2
Ago med Out Dez
20
40
50
AST U/L
30
A
+1/2t -1/2t med
20
40
50
AST U/L
30
C AST U/L
30
50
60
40
+800 +800 +800 -800 -800 -800
Ago med Out Dez
B
10 10 10

16
De uma maneira geral, os atletas apresentam globalmente valores mais elevados de AST durante os períodos de competição, quando comparados com os sedentários (41,1 ± 15,0 U/L vs 19,9 ± 6,3 UI/L)).
Este estudo debruçou-se apenas sobre uma parte da temporada competitiva, durante a qual decorreu a participação em simultâneo de duas competições, uma a nível nacional e outra a nível europeu.
Num outro estudo, desenvolvido ao longo de toda uma temporada competitiva, procedeu-se à avaliação, entre outros, de uma série de parâmetros relacionados com possíveis alterações de parâmetros de natureza bioquímica, hormonal e de stress oxidativo (Silva e col, 2014). Catorze jogadores (idade, 25,7 ± 4,6 anos; altura, 178,1 ± 5,7 cm; peso, 76,5 ± 9,2 Kg) da liga secundária portuguesa de futebol profissional participaram neste estudo.
As colheitas de material biológico para a determinação labora-torial dos parâmetros a avaliar, foram efetuadas no início da pré-época (primeira, PE), a meio da época desportiva, no mês de janeiro (segunda, ME) e no final do campeonato (terceira, FE). As segundas e terceiras colheitas foram realizadas 72 h após a realização de um jogo de futebol. Uma quarta colheita foi realizada no período compreendido entre o fim da temporada desportiva e o início da pré-época (FIE).
Para além da CPK, utilizou-se a Mioglobina como um marcador laboratorial complementar de lesão muscular.
Os perfis comportamentais dos níveis plasmáticos destes 2 marca-dores de lesão muscular, CPK e Mioglobina, ao longo do tempo de estudo, são muito similares entre si (Figura 3). O aumento dos níveis plasmáticos destes marcadores, observado sensivelmente a meio da época desportiva (janeiro), embora dentro dos intervalos de referência para os futebolistas de alta competição, pode refletir um aumento do número de jogos e de volume de treinos efetuados, bem como, do envolvimento muscular associado.
O objetivo dos estudos anteriores consistiu na avaliação da evo-lução do comportamento fisiológico do organismo, efetuada através da análise dos níveis plasmáticos de biomarcadores da função muscular ao longo de um período de tempo considerável. Os resultados apre-

17
sentados reportam a resposta do organismo a essas condições, permitindo, através da sua análise, compreender, de alguma maneira, o efeito da adaptação fisiológica a esses estímulos de alta competição, ao longo de toda uma temporada desportiva.
Pouco nos diz, contudo, sobre a influência do exercício ou dos potenciais efeitos ou danos por ele causados, durante ou imediatamente após a sua realização (efeito agudo).
Figura 3. Perfis de marcadores de lesão muscular de jogadores de futebol profissional da Liga secundária portuguesa de futebol profissional. As colheitas foram efetuadas no início da pré-época (PE), a meio da época desportiva (ME), no final do campeonato (FE) e entre o fim da temporada desportiva e o início da pré-época (FIE). A. Valores médios de CPK plasmática dos participantes neste estudo, nos tempos indicados; B. Valores médios de Mioglobina plasmática de jogadores participantes neste estudo, nos tempos indicados.
Pouco nos diz também acerca do processo evolutivo de recupera-ção desse esforço muscular. O conhecimento mais pormenorizado do efeito da intensidade do esforço da prática de futebol sobre o sistema muscular pode ser melhor acompanhado se se avaliar o que se passa antes e nos momentos imediatamente seguintes à sua efetivação, e nos tempos iniciais da sua recuperação.
No sentido de analisar estes efeitos realizou-se um estudo onde se utilizaram como marcadores de lesão muscular a CPK e a Mioglobina (Ascensão e col, 2008).
Fig 3
Fig.3. Níveis médios plasmáticos de marcadores de lesão muscular, CPK e mioglobina (Mb), de jogadores de uma equipa da liga secundária de futebol portuguesa, em 4 momentos da época desportiva, pré-época (PE), meio da época (ME), final do campeonato (FE) e entre o fim da temporada desportiva e o início da pré-época (FIE).
150
350
450
CPK UI/L
PE PIE ME FE
15,0
25,0
30,0
Mb mg/L
20,0
PE PIE ME FE 10,0 50
250

18
A Mioglobina é uma proteína muscular, de estrutura globular, cuja função principal consiste no transporte (e reserva) do oxigénio molecular nesses tecidos. A Mioglobina está identificada como um marcador precoce de lesão do músculo-esquelético, sendo utilizada igualmente no estudo do perfil laboratorial de lesão cardíaca.
Os seus níveis plasmáticos aumentam 1 a 3 horas após lesão muscular, atingindo o seu pico entre as 6 e as 12 horas. O regresso aos valores plasmáticos normais ocorre a partir do momento do fim da lesão, essencialmente devido ao seu rápido metabolismo hepático e à sua clearance renal. A Mioglobina é nefrotóxica. A mioglobinúria é um parâmetro que, entre outros, é útil na avaliação clinica e laboratorial da rabdomiólise, situação em que ocorre perda da integridade celular, lise e morte da célula muscular, com libertação dos constituintes celulares para o plasma (Nayak e Jindal, 2015). O exercício muscular excessivo ou à exaustão constitui um, entre vários, dos fatores causais destas situações (George, 2014; Kima e col, 2016).
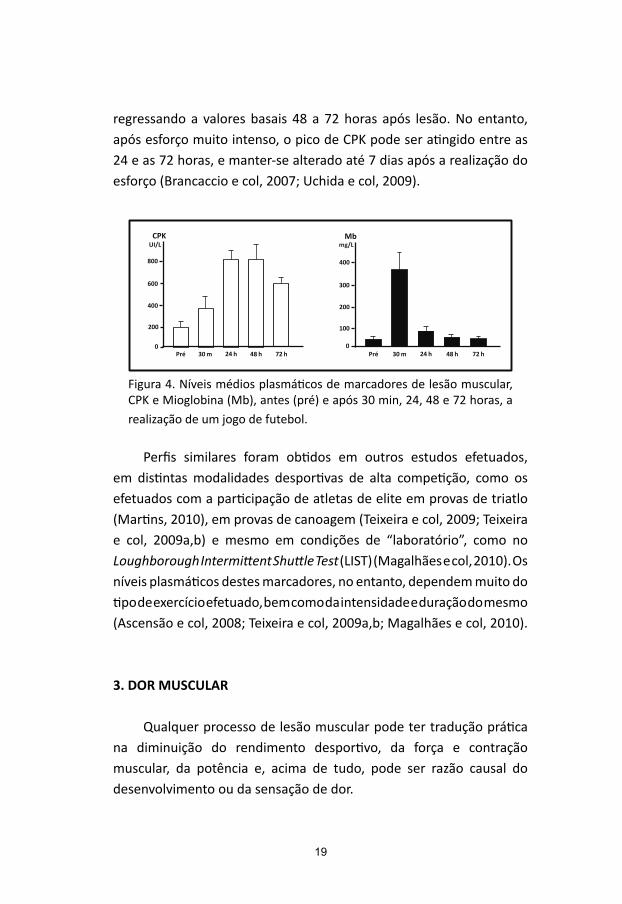
Os perfis dos níveis plasmáticos dos dois marcadores de lesão muscular em análise, CPK e Mioglobina, determinados imediatamente antes e após 30 minutos, 24, 48 e 72 horas da realização de um jogo de futebol amigável, no qual participaram 16 jogadores de futebol (idade, 25,7 ± 4,6 anos; altura, 178,1 ± 5,7 cm; peso, 76,5 ± 9,2 Kg) pertencentes a uma equipa do escalão secundário do futebol português, encontram-se plasmados na Figura 4.
Como se pode observar, verifica-se um aumento significativo dos níveis plasmáticos da Mioglobina 30 minutos após a realização do jogo, os quais atingem valores muito próximos dos níveis basais, ao fim de 24 horas. O aumento dos níveis plasmáticos de CPK ocorre mais lentamente que os da Mioglobina, como aliás seria expectável, atingindo os seus valores máximos entre as 24 e as 48 horas, com valores da ordem dos 800 UI/L, a partir das quais começam a diminuir, mantendo-se, contudo, em níveis significativamente aumentados, quando comparados com os basais, ao fim das 72 horas do período de recuperação, em estudo.
Os níveis plasmáticos de CPK começam a aumentar, normalmente, entre as 4 e as 6 horas, atingindo o seu pico entre as 18 e as 24 horas,
19
regressando a valores basais 48 a 72 horas após lesão. No entanto, após esforço muito intenso, o pico de CPK pode ser atingido entre as 24 e as 72 horas, e manter-se alterado até 7 dias após a realização do esforço (Brancaccio e col, 2007; Uchida e col, 2009).
Figura 4. Níveis médios plasmáticos de marcadores de lesão muscular, CPK e Mioglobina (Mb), antes (pré) e após 30 min, 24, 48 e 72 horas, a
realização de um jogo de futebol.
Perfis similares foram obtidos em outros estudos efetuados, em distintas modalidades desportivas de alta competição, como os efetuados com a participação de atletas de elite em provas de triatlo (Martins, 2010), em provas de canoagem (Teixeira e col, 2009; Teixeira e col, 2009a,b) e mesmo em condições de “laboratório”, como no Loughborough Intermittent Shuttle Test (LIST) (Magalhães e col, 2010). Os níveis plasmáticos destes marcadores, no entanto, dependem muito do tipo de exercício efetuado, bem como da intensidade e duração do mesmo (Ascensão e col, 2008; Teixeira e col, 2009a,b; Magalhães e col, 2010).
3. DOR MUSCULAR
Qualquer processo de lesão muscular pode ter tradução prática na diminuição do rendimento desportivo, da força e contração muscular, da potência e, acima de tudo, pode ser razão causal do desenvolvimento ou da sensação de dor.
Fig.4. Níveis médios plasmáticos de marcadores de lesão muscular, CPK e mioglobina (Mb), antes (pré) e após 30 min, 24, 48 e 72 horas, a realização de um jogo de futebol
Fig 4
CPK UI/L
Pré 48 h 30 m 24 h
200
600
800
400
72 h
100
300
400
Mb mg/L
200
Pré 48 h 30 m 24 h 72 h 0 0

20
A designada dor tardia, diferente da dor traumática, é um tipo de dor que aparece entre as 12 e as 24 horas após a realização de exercício físico, e que atinge o seu pico, normalmente, entre as 24 e as 72 horas. A partir de então, verifica-se uma diminuição progressiva da mesma que pode prolongar-se até 5 a 7 dias, após o término do exercício (Friden, 2013). A severidade associada a este tipo de dor depende da intensidade, duração e tipo de exercício (Cheung e col, 2003). Esta situação, apesar de transitória, pode, contudo, condicionar a atividade do atleta, e, em particular, limitar o seu rendimento ou desempenho desportivo.
Não se conhecem em rigor os mecanismos causadores de tal efeito, embora seja aceite que o exercício excêntrico é o/um dos primeiros fatores responsáveis pelo desencadear deste processo (Kanda e col, 2013; Zondi e col, 2015).
A lesão (ou micro lesões) do tecido muscular é uma das hipóteses explicativas desta situação de dor tardia. Na realidade, é de uma forma consensual aceite que a lesão muscular, causada pelo excesso de esforço despendido, pode ser responsável pelo aparecimento de dor muscular. Alguns estudos referem a existência de uma correlação significativa positiva entre níveis plasmáticos elevados de CPK e de Mioglobina e a dor tardia (Twist e col, 2005; Zondi e col, 2015). No entanto, pode ocorrer lesão muscular, a crer no aumento dos níveis plasmáticos de enzimas utilizadas como marcadores de lesão, e.g. CPK, sem o concomitante aparecimento de dor muscular. Na sequência destes quadros de (micro) lesões, ocorre o processo de cicatrização e de reconstrução do músculo lesado, do qual resulta um maior fortalecimento dos tecidos musculares e maiores fibras musculares (hipertrofia). Os processos de treino permitem uma maior adaptação e resistência muscular ao esforço desprendido, minorando ou compensando o estabelecimento deste tipo de dor.
Uma outra hipótese explicativa, e que pode ser cumulativa ou sobreponível, atribui este processo a uma ação inflamatória de reparação como resposta às (micro) lesões musculares ou do tecido conjuntivo. Esta resposta inflamatória será, pois, a responsável pelo

21
aparecimento da dor, do edema e da fixidez muscular (Kenney e col, 2016). O mecanismo teórico (modelo) hoje ainda mais aceite (Armstrong, 1984), inclui a existência de variados passos sequenciais que se inicia com o (i) aparecimento das micro lesões (micro ruturas) das fibras musculares durante os processos de contração muscular, prosseguindo (ii) pela necrose, resultante do dano muscular e (iii) pela libertação de compostos intracelulares (e.g. cininas, histamina) e a estimulação dos terminais nervosos associados à dor, por produtos
Fig. 5. Alterações leucocitárias, leucócitos, neutrófilos (escuro) e linfócitos (claro) com o exercício físico. A. Antes, 30 min, 24, 48 e 72 horas após a realização de um jogo de futebol; B. Antes, imediatamente após e 1 hora após a realização de uma prova competitiva de motocross; C. Antes, imediatamente após e 1 hora após a realização de uma prova de escalada. Imediatamente após a realização de exercício físico de elevada intensidade ocorre um aumento do número de leucócitos totais sanguíneos, devido principalmente ao aumento do número de neutrófilos e linfócitos (e de monócitos, em menor proporção). No entanto, o número de linfócitos, cerca de 30 minutos após a realização do exercício físico, diminui acentuadamente (linfopenia), especialmente à custa das células T e NK, mantendo-se contudo a neutrofilia (Pedersen e col, 1998). Esta linfopenia tem sido explicada, por alguns autores, por um mecanismo de apoptose mediada pelos níveis de cortisol e de catecolaminas aumentados pós exercício, os quais são fatores indutores deste processo (Cioca e col, 2000; Pederson e col, 2000; Morren e col, 2002; Krüger e Mooren, 2014). A diminuição destas células, ligada a uma perda da sua atividade funcional, em particular, devido à queda destas subpopulações linfocitárias (T e células NK) e à diminuição da IgA, que se verifica em consequência do exercício físico, parece ser responsável pela quebra de imunidade que tem sido associada à prática de exercício de alta intensidade e prolongado, e que explicaria o aumento da frequência de doenças do trato respiratório superior em atletas (MacKinnon e Hooper, 1994; Pedersen, 1998; Gleeson, 2007; Silva e Macedo, 2011; Trochimiak e Hübner-Woźniak, 2012; Sterczala, 2014)
Fig. 5. Alterações leucocitárias, leucócitos, neutrófilos (escuro) e linfócitos (claro) com o exercício físico. A. Antes, 30 min, 24, 48 e 72 horas após a realização de um jogo de futebol; B. Antes, imediatamente após e 1 hora após a realização de uma prova competitiva de motocross; C. Antes, imediatamente após e 1 hora após a realização de uma prova de escalada.
Imediatamente após a realização de exercício físico de elevada intensidade ocorre um aumento do número de leucócitos totais sanguíneos, devido principalmente ao aumento do número de neutrófilos e linfócitos (e de monócitos, em menor proporção). No entanto, o número de linfócitos, cerca de 30 minutos após a realização do exercício físico, diminui acentuadamente (linfopenia), especialmente à custa das células T e NK, mantendo-se contudo a neutrofilia (Pedersen e col, 1998). Esta linfopenia tem sido explicada, por alguns autores, por um mecanismo de apoptose mediada pelos níveis de cortisol e de catecolaminas aumentados pós exercício, os quais são fatores indutores deste processo (Cioca e col, 2000; Pederson e col, 2000; Morren e col, 2002; Krüger e Mooren, 2014). A diminuição destas células, associada a uma perda da sua atividade funcional, em particular, devido à queda destas subpopulações linfocitárias (T e células NK), e à diminuição da IgA que se verifica em consequência do exercício físico, parece ser responsável pela quebra de imunidade, que tem sido associada à prática de exercício de alta intensidade e prolongado, e que explicaria o aumento da frequência de doenças do trato respiratório superior em atletas (MacKinnon e Hooper, 1994; Pedersen, 1998; Gleeson, 2007; Silva e Macedo, 2011; Trochimiak e Hübner-Woźniak, 2012; Sterczala, 2014)
Fig 5
X109/L
Pré 48 h 30 m 24 h
A
3
9
12
6
72 h 0
X109/L
Pré 0H 1H
B
3
9
12
6
0
X109/L
Pré 0H 1H
c
3
9
12
6
0

22
da inflamação que se acumulam nos interstícios celulares (Leeder e col, 2014; Zondi e col, 2015). Após realização de exercício físico, a nível hematológico observa-se um aumento significativo de leucócitos aumento esse que é transitório (Figura 5). Os neutrófilos, que atingem o pico plasmático a partir da primeira hora após término do exercício físico e regressam aos valores basais, normalmente a partir das 6 horas após exercício físico, embora possam esses aumentos persistir até 5 dias, são as primeiras células a serem mobilizadas, após um processo de desmarginalização, para o local de lesão ou de inflamação por um processo de diapedese (Tidball, 2005; Magalhães e col, 2010). Estas células, por um processo de ativação, contribuem por mecanismos de fagocitose para a destruição de microrganismos, e para a ativação de outras células do sistema imunitário de defesa, através da libertação de citoquinas e quimiocinas. Por seu lado, os monócitos, após um aumento quantitativo dos seus níveis sanguíneos, como resposta ao exercício físico, diminuem nas 48 horas seguintes, uma vez que são convertidos em macrófagos após infiltração para os locais afetados (de necrose e de inflamação), onde contribuem para a sua limpeza, através da remoção dos tecidos lesados, por fagocitose (Gleeson e col, 1995; Tidball e Wehling-Henricks, 2007).
Mais ainda, podem colaborar nos processos de reparação do tecido muscular mediante a secreção de variados moléculas, e.g. fatores de crescimento semelhantes à insulina (IGF) e algumas citoquinas, como o fator de crescimento dos fibroblastos (FCF), que contribuem para a reabilitação dos tecidos (Butterfield e col, 2006).
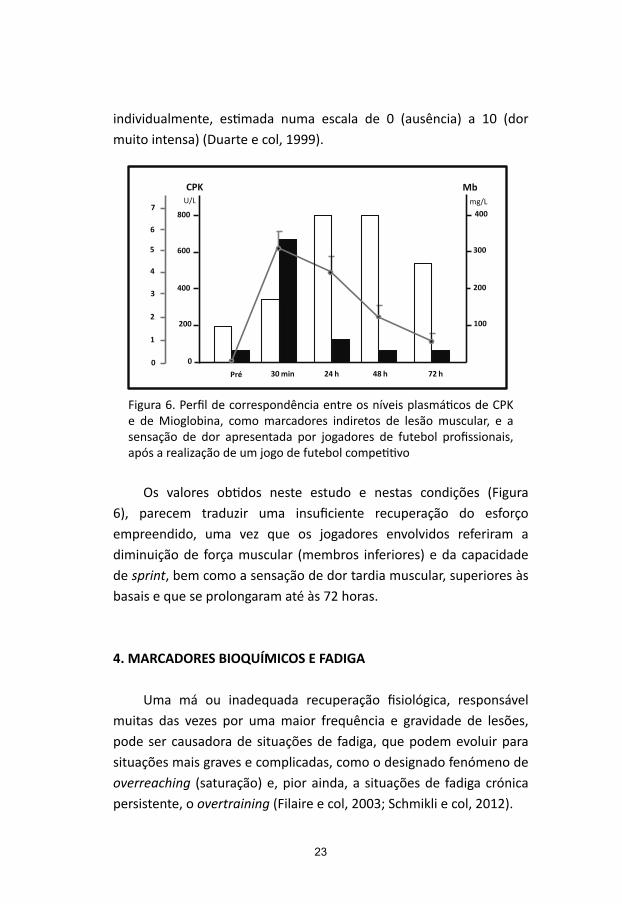
No sentido de procurar encontrar alguma tradução laboratorial entre dor muscular e lesão muscular, procedeu-se a um estudo visando encontrar correspondência entre os níveis plasmáticos de CPK e de Mioglobina, como marcadores indiretos de lesão muscular, e a sensação de dor apresentada por jogadores de futebol profissionais, após a realização de um jogo de futebol competitivo (Ascensão e col, 2008). A avaliação da sensação de dor foi efetuada mediante resposta a um inquérito distribuído a cada um dos participantes do estudo, onde se pedia que fosse manifestada a perceção de dor sentida
23
individualmente, estimada numa escala de 0 (ausência) a 10 (dor muito intensa) (Duarte e col, 1999).
Figura 6. Perfil de correspondência entre os níveis plasmáticos de CPK e de Mioglobina, como marcadores indiretos de lesão muscular, e a sensação de dor apresentada por jogadores de futebol profissionais, após a realização de um jogo de futebol competitivo
Os valores obtidos neste estudo e nestas condições (Figura 6), parecem traduzir uma insuficiente recuperação do esforço empreendido, uma vez que os jogadores envolvidos referiram a diminuição de força muscular (membros inferiores) e da capacidade de sprint, bem como a sensação de dor tardia muscular, superiores às basais e que se prolongaram até às 72 horas.
4. MARCADORES BIOQUÍMICOS E FADIGA
Uma má ou inadequada recuperação fisiológica, responsável muitas das vezes por uma maior frequência e gravidade de lesões, pode ser causadora de situações de fadiga, que podem evoluir para situações mais graves e complicadas, como o designado fenómeno de overreaching (saturação) e, pior ainda, a situações de fadiga crónica persistente, o overtraining (Filaire e col, 2003; Schmikli e col, 2012).
Fig 6
Fig.6. Perfil de correspondência entre os níveis plasmáticos de CPK e de mioglobina (Mb), como marcadores indiretos de lesão muscular, e a sensação de dor apresentada por jogadores de futebol profissionais, após a realização de um jogo de futebol competitivo
U/L mg/L
200
300
400
100
CPK Mb
Pré 30 min 24 h 48 h 72 h
1
3
5
7
200
400
600
800
0
2
4
6
0

24
O overtraining, de modo sucinto, é uma síndrome caracterizada por um conjunto de sintomas físicos, que englobam a perda de performance desportiva, diminuição da força e da velocidade, dificuldade ou incapacidade em treinar de modo consistente e eficaz, dificuldades de recuperação do esforço despendido, etc. A esta componente de natureza física, acresce ainda o envolvimento de outras componentes de natureza comportamental e de stress emocional, como o aparecimento de estados de medo, fobias, falta de confiança, insónias e a depressão, que complicam e agravam ainda mais este quadro fisiológico. Estas situações condicionam muito o atleta e a sua atividade e podem persistir durante períodos muito prolongados no tempo. São ocorrências de difícil diagnóstico, que necessitam de ser detetadas e prevenidas, de modo a evitar o sub-rendimento e a eficácia de um atleta de alta competição de qualquer modalidade desportiva, ou a sua eventual ausência forçada.
Sob o ponto de vista laboratorial não existem marcadores claros que permitam avaliar o estado de fadiga e/ou na sua forma mais evoluída e limite, de overtraining.
Foram vários os marcadores laboratoriais propostos para avaliar estes estados, onde se observa um desequilíbrio metabólico com predominância dos estados catabólicos sobre o anabolismo, mas nenhum é ainda hoje utilizado e aceite de forma clara e identificativa para esse fim. No entanto, a determinação dos níveis plasmáticos de algumas hormonas como a testosterona e o cortisol, as catecolaminas, bem como de alguns aminoácidos, nomeadamente, o triptofano e os aminoácidos ramificados, entre outros, tem sido utilizados como parte acrescida na avaliação global e clínica destas situações, em medicina desportiva.
O futebol constitui um bom exemplo de uma atividade que exige por parte do seu praticante uma preparação particular, dado o tipo e intensidade variável de esforço físico a esta modalidade associada. O tipo de esforço associado a esta modalidade desportiva é muito variado, englobando saltos, sprints ou corridas a alta velocidade, que alternam com situações de corrida em velocidade lenta e moderada, choques, quedas, etc. que exige uma boa recuperação através de

25
um período de descanso. Este período de descanso tem de ser bem doseado e adequado de modo a permitir a regeneração ou recuperação de possíveis microlesões das fibras musculares que ocorrem durante a atividade, bem como da reposição apropriada dos níveis energéticos nela consumidos, de modo a evitar os fenómenos de fadiga muscular que condicionam a perfomance e o rendimento desportivo.
No sentido de melhor compreender e monitorizar o estado de fadiga e o desempenho individual e coletivo de uma equipa de futebol de alta competição da 1ª Liga de Futebol profissional portuguesa, com participação na Champions League, procedeu-se à avaliação laboratorial dos níveis plasmáticos de testosterona e de cortisol.
Na Figura 7, encontram-se descriminados os dados médios glo-bais obtidos dos níveis plasmáticos de cortisol ao longo dos meses em estudo. Os valores de cortisol plasmático observados, em termos absolutos, são superiores nos dois primeiros momentos da colheita que no mês de dezembro.
Este perfil de resultados pode ser explicado pela maior carga e intensidade de exercício despendido durante o tempo competitivo, no sentido da obtenção da melhor performance e do melhor rendimento desportivo, relativamente ao período de repouso inicial. Os meses de agosto e de outubro correspondem ao período da temporada em estudo, onde, o stress causado pela retoma dos processos de treino e da forma física após período de férias e pelo volume de treino e de jogos competitivos da 1ª Liga de Futebol profissional e da Champions League sobre o organismo, têm um impacto catabólico mais significativo sobre os jogadores, diminuindo à medida que se atinge uma melhor adaptação dos atletas ao esforço realizado.
Por outro lado, os níveis plasmáticos médios globais de testos-terona, obtidos durante o mesmo período de tempo do estudo, apresentam valores mais elevados nos meses de agosto e dezembro, e mais baixos no mês de outubro. O facto do desenvolvimento de atividade física ser um fator indutor positivo da produção de testosterona, pode justificar essas variações plasmáticas globais encontradas. No entanto, não pode ser excluída a hipótese de o tempo escolhido para recolha

26
das amostras plasmáticas (48h após um jogo) poder de alguma maneira ter condicionado esses resultados. Durante a prática de exercício físico, e em particular um exercício de alta intensidade e duração, como é característico no futebol, os níveis plasmáticos de cortisol aumentam como resposta a essa situação stress (catabólico), mais ainda potenciados pela libertação de catecolaminas. Deste modo, este aumento da produção de cortisol, cujos níveis plasmáticos se mantêm durante algum tempo (primeiras horas) do período de recuperação, pode inibir o eixo hipotálamo-hipofisário, o que provoca um efeito contrário na produção e nos níveis de testosterona, e pode justificar as diferenças entre os níveis de testosterona observados.
Figura 7. Níveis plasmáticos médios globais de Cortisol e Testosterona de uma equipa de futebol de alta competição com participação na 1ª Liga de Futebol profissional (Portugal) e na Champions League (UEFA).
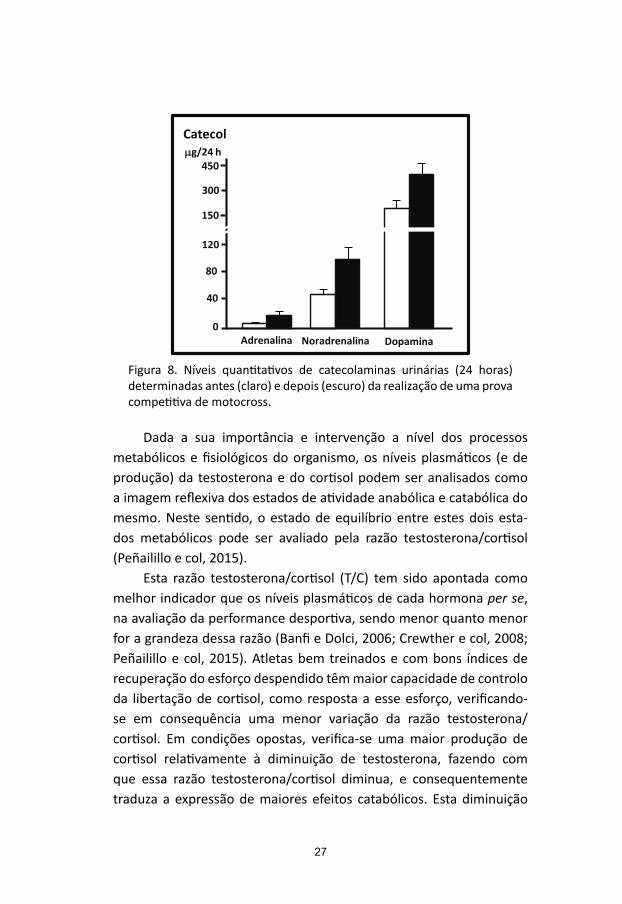
Não foram determinados os níveis plasmáticos de catecolaminas neste estudo. Mas, em outro estudo realizado (motocross), e unicamente a título de exemplo (Figura 8), pode avaliar-se bem o incremento da produção de catecolaminas, medido pelos níveis de excreção urinária (urina de 24 horas), em resposta ao stress provocado pela prática de uma modalidade desportiva altamente competitiva e de intensidade elevada (Ascensão e col, 2007).
Figura 7. Níveis plasmáticos médios globais de Cortisol e Testosterona de uma equipa de futebol de alta competição com participação na 1ª Liga de Futebol profissional (Portugal) e na Champions League (UEFA).
Fig. 7
Ago Out Dez
140
180
200
Cort ng/mL
160
Ago Out Dez
5,0
7,0
8.0
Test ng/mL
6,0
4,0 120
27
Figura 8. Níveis quantitativos de catecolaminas urinárias (24 horas) determinadas antes (claro) e depois (escuro) da realização de uma prova competitiva de motocross.
Dada a sua importância e intervenção a nível dos processos metabólicos e fisiológicos do organismo, os níveis plasmáticos (e de produção) da testosterona e do cortisol podem ser analisados como a imagem reflexiva dos estados de atividade anabólica e catabólica do mesmo. Neste sentido, o estado de equilíbrio entre estes dois esta-dos metabólicos pode ser avaliado pela razão testosterona/cortisol (Peñailillo e col, 2015).
Esta razão testosterona/cortisol (T/C) tem sido apontada como melhor indicador que os níveis plasmáticos de cada hormona per se, na avaliação da performance desportiva, sendo menor quanto menor for a grandeza dessa razão (Banfi e Dolci, 2006; Crewther e col, 2008; Peñailillo e col, 2015). Atletas bem treinados e com bons índices de recuperação do esforço despendido têm maior capacidade de controlo da libertação de cortisol, como resposta a esse esforço, verificando-se em consequência uma menor variação da razão testosterona/cortisol. Em condições opostas, verifica-se uma maior produção de cortisol relativamente à diminuição de testosterona, fazendo com que essa razão testosterona/cortisol diminua, e consequentemente traduza a expressão de maiores efeitos catabólicos. Esta diminuição
Fig.8. Níveis quantitativos de catecolaminas urinárias (24 horas) determinadas antes (claro) e depois (escuro) da realização de uma prova competitiva de motocross.
Ascenção e col. Br J Sports Med., 41(2007)101-5
Fig. 8
Adrenalina Noradrenalina 0
80
120
Catecol mg/24 h
40
150
300
450
Dopamina
28
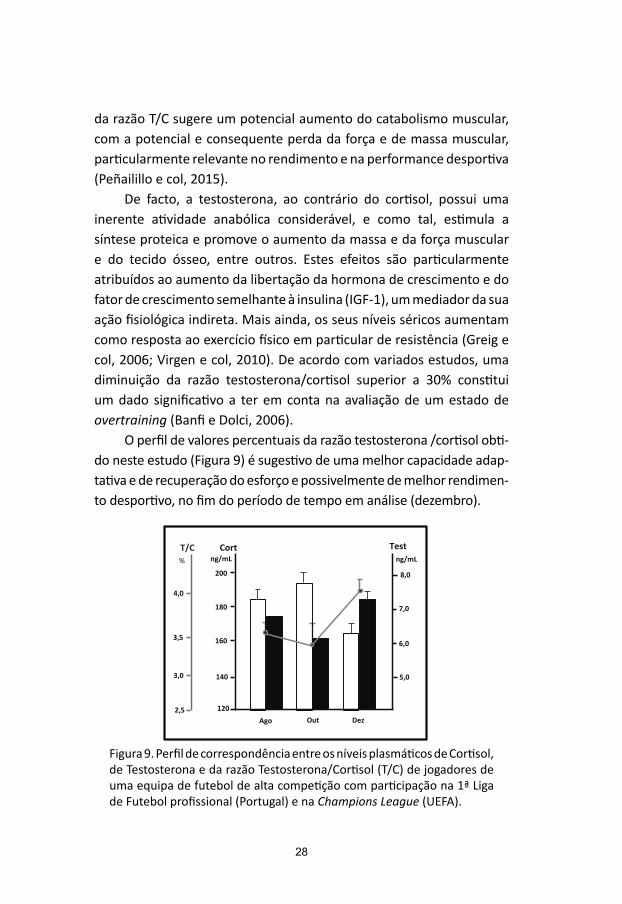
da razão T/C sugere um potencial aumento do catabolismo muscular, com a potencial e consequente perda da força e de massa muscular, particularmente relevante no rendimento e na performance desportiva (Peñailillo e col, 2015).
De facto, a testosterona, ao contrário do cortisol, possui uma inerente atividade anabólica considerável, e como tal, estimula a síntese proteica e promove o aumento da massa e da força muscular e do tecido ósseo, entre outros. Estes efeitos são particularmente atribuídos ao aumento da libertação da hormona de crescimento e do fator de crescimento semelhante à insulina (IGF-1), um mediador da sua ação fisiológica indireta. Mais ainda, os seus níveis séricos aumentam como resposta ao exercício físico em particular de resistência (Greig e col, 2006; Virgen e col, 2010). De acordo com variados estudos, uma diminuição da razão testosterona/cortisol superior a 30% constitui um dado significativo a ter em conta na avaliação de um estado de overtraining (Banfi e Dolci, 2006).
O perfil de valores percentuais da razão testosterona /cortisol obti-do neste estudo (Figura 9) é sugestivo de uma melhor capacidade adap-tativa e de recuperação do esforço e possivelmente de melhor rendimen-to desportivo, no fim do período de tempo em análise (dezembro).
Figura 9. Perfil de correspondência entre os níveis plasmáticos de Cortisol, de Testosterona e da razão Testosterona/Cortisol (T/C) de jogadores de uma equipa de futebol de alta competição com participação na 1ª Liga de Futebol profissional (Portugal) e na Champions League (UEFA).
Fig.9. Perfil de correspondência entre os níveis plasmáticos de Cortisol, de Testosterona e da razão Testosterona/Cortisol (T/C) de jogadores de uma equipa de futebol de alta competição com participação na 1ª Liga de Futebol profissional (Portugal) e na Champions League (UEFA)
Fig. 9 Ago Out Dez
6,0
7,0
8,0
5,0
Test Cort
Ago Out Dez
140
160
180
200
ng/mL ng/mL
3,0
3,5
T/C %
4,0
2,5 120

29
A acumulação dos efeitos resultantes de situações de stress provo-cadas pelo significativo volume de jogos competitivos e de treinos efetuados ao longo de toda uma época desportiva pode refletir-se em alterações fisiológicas e funcionais dos atletas.
Estas situações podem ter tradução a nível dos resultados oude biomarcadores específicos laboratoriais, como resultado da variabilidade de condições indutoras de alterações do “equilíbrio” entre os estados anabólicos e catabólicos do organismo, permitindo avaliar o impacto do exercício competitivo sobre distintas áreas, como a bioquímica, endocrinológica, hematologica, etc.. Mais ainda podem induzir alterações a nível físico e/ou psicológico cuja avaliação é de importância significativa, uma vez que podem afetar o desempenho e o rendimento desportivo.
Por outro lado, ao longo de toda uma época desportiva compe-titiva, ocorrem processos de adaptação fisiológica do organismo como resposta a estímulos de stress, os quais conferem condições de uma maior proteção face a situações potencialmente lesivas. A análise conjunta destes parâmetros constitui um valioso contributo adicional para os processos de recuperação do esforço e planeamento de uma época desportiva.
O estudo anterior foi limitado no tempo, abrangendo apenas os primeiros 4 meses da época competitiva desportiva. Por isso, um outro estudo, com participação de jogadores de futebol da Liga portuguesa de futebol profissional (14 jogadores), foi realizado durante toda uma temporada competitiva, monitorizada em 4 momentos, pré-época (PE), a meio da época desportiva (ME), no final do campeonato (FE) e no período compreendido entre o fim da temporada desportiva e o início da pré-época (FIE) (Silva e col, 2014).
Os resultados mostraram que ao longo da época desportiva (Figura 10) se verificou um aumento da razão testosterona/cortisol cujo valor máximo foi atingido no fim do período competitivo, regressando posteriormente a níveis sensivelmente basais ou de pré-época. Nos dois primeiros pontos de monitorização, os níveis de T/C e de cortisol sofreram pequenas variações.

30
Figura 10. Perfil de correspondência entre os níveis plasmáticos de testosterona, de cortisol e da razão Testosterona/Cortisol (T/C) de jogadores de uma equipa de futebol da Liga secundária de futebol profissional, em 4 momentos da época desportiva, pré-época (PE), meio da época (ME), final do campeonato (FE) e entre o fim da temporada desportiva e o início da pré-época (FIE).
Os níveis de cortisol refletem o estado catabólico associado a um aumento de stress provocado pelo exercício físico. Quanto mais intensa for a situação de stress, como acontece durante um jogo de futebol, em alta competição, maior é a resposta traduzida na produção de cortisol (Filaire, e col, 2003). Por outro lado, os níveis de T/C podem refletir uma melhor adaptação ao esforço, dado que o exercício físico induz o aumento da produção de testosterona, e o consequente melhor rendimento desportivo (Kraemer e col, 2004). Aliás, e neste sentido, a análise individual ou conjunta destes parâmetros tem sido sugerida como um contributo na avaliação do rendimento desportivo e em particular do estado de fadiga (repouso), e no limite, de overtraining. Distintos estudos efetuados na modalidade de futebol competitivo, contudo, manifestam algumas reservas sobre o contributo da razão T/C para a avaliação do rendimento desportivo ou do estado de fadiga. É possível, no entanto, que esta variabilidade de resultados seja explicada
Fig.10
Fig. 10. Perfil de correspondência entre os níveis plasmáticos de Cortisol (Cort), Testosterona (Test) e da razão Testosterona/Cortisol (T/C) de jogadores de uma equipa de futebol da Liga secundária de futebol profissional, em 4 momentos da época desportiva, pré-época (PE), meio da época (ME), final do campeonato (FE) e entre o fim da temporada desportiva e o início da pré-época (FIE).
6,5
7,0
7,5
6,0
Test Cort
PE ME FE
100
130
160
190
ng/mL ng/mL
2,0
4,0
T/C %
6,0
PIE
8,0
70

31
por distintos protocolos ou condições aplicadas, nomeadamente no que diz respeito a diferentes tempos de avaliação em função da época desportiva, durante esses estudos (Gorostiaga e col, 2004).
5. MARCADORES BIOQUÍMICOS ASSOCIADOS AO STRESS OXIDATIVO
A ação do stress oxidativo sobre o organismo humano induzido pelo exercício físico tem sido objeto de estudo há já vários anos. A manifestação destes efeitos incide de forma particular a nível muscular, sendo responsabilizado como fator causal ou indutor da lesão muscular e do estabelecimento de estado de fadiga, mas também na diminuição do rendimento e do desempenho desportivo e no desenvolvimento de quadros de overtraining (Powers e Jackson, 2008; Tanskanen e col, 2010; Kreher e Schwartz, 2012). Estes efeitos dependem muito da intensidade e da duração do exercício efetuado, sendo particularmente relevantes em situações de prática desportiva de alta competição. Se é verdade que distintos estudos se debruçam sobre os aspetos negativos do stress oxidativo associados à prática desportiva, outros há, contudo, que valorizam o facto de a sua implantação induzir uma resposta de adaptação fisiológica de defesa antioxidante, benéfica, por parte do organismo. De facto, a prática regular de exercício físico, resulta na diminuição quantitativa das espécies oxidantes formadas e no incremento da capacidade de defesa antioxidante, tornando o organismo mais bem preparado e mais resistente fisiologicamente para neutralizar ou diminuir os efeitos potencialmente deletérios do stress oxidativo no exercício seguinte (Knez e col, 2006; Nikolaidis e col, 2007; Radak e col, 2008; Morton e col, 2009; Majerczak e col, 2010; Wagner e col, 2011; Tong e col, 2012; Radak e col, 2013; Azizbeigia e col, 2014).
5.1. Espécies oxidantes e radicais livresEspécies reativas de oxigénio (ROS) são moléculas químicas muito
pequenas, presentes no organismo, com elevada capacidade oxidante,

32
que englobam em si nesta designação, os radicais livres de oxigénio, mas também outros compostos ou moléculas não radicalares, derivadas do oxigénio, e que podem gerar radicais livres. Podem ter uma origem endógena, geradas como subprodutos do metabolismo celular, ou serem de origem exógena ao organismo.
Sejam de origem fisiológica ou não, os radicais livres, subconjunto de moléculas ou compostos em que se subdividem as ROS, são moléculas de elevada reatividade, em particular pelo facto de os seus átomos possuírem um número ímpar de eletrões, e de terem vida muito curta (Viña e col, 2000).
Esta elevada reatividade apresenta algumas vantagens sob o pon-to de vista fisiológico de caracter “benigno”, podendo atuar como sina-lizadores ou mensageiros biológicos em inúmeras funções celulares, como, nos mecanismos de sinalização celular, nos processos biossin-téticos e de produção de energia, na regulação do crescimento celular, na defesa contra infeções mediadas do sistema imunológico, no relaxamento da musculatura lisa dos vasos e nos processos de inibição da adesão plaquetária, entre outros (Alderton e col, 2001; Dröge, 2002; Powers e col, 2011; Braakhuis e Hopkins, 2015).
Os radicais livres podem, por outro lado, exercer efeitos “per-versos” estando associados aos processos de envelhecimento, e como fator causal ou de agravamento de muitos quadros patológicos, como e.g. a aterosclerose, diabetes, cancro, doenças pulmonares, como enfisema, processos neurodegenerativos e do foro neurológico, como doença de Parkinson, Alzheimer, esclerose múltipla (Porto, 2001; Calvo e Lopez, 2011; Silva e col, 2011; Gospodaryov e Lushchak, 2012).
Entre as distintas espécies reativas de oxigénio, os principais radicais livres, porque formados nos processos celulares fisiológicos, são o superóxido (O
2•-), o hidroxilo (HO•) e o peroxilo (HOO•), sendo
o peróxido de hidrogénio, a mais representativa espécie reativa de oxigénio não radicalar.
De uma forma sucinta, o radical superóxido (anião) resulta daaceitação de um eletrão por parte do oxigénio no seu estado fundamental. Em meios aquosos possui pouca atividade reativa, tendo um tempo de

33
vida curta. É convertido em peróxido de hidrogénio por ação da enzima Superóxido dismutase (SOD), com um tempo de semivida mais longo. O peróxido de hidrogénio não é um radical livre. Existe nas células em maior quantidade que o radical superóxido e é pouco reativo. Porém, ao contrário do radical superóxido é capaz de atravessar as membranas celulares e, em locais distantes daquele onde foi formado, de reagir com iões metálicos, em particular, o Fe2+ (e Cu+), dando origem ao radical hidroxilo (reação de Fenton). Por este motivo, o peróxido de hidrogénio, ao potencialmente reagir com as membranas dos eritrócitos e com proteínas ligadas ao Ferro (Fe2+) faz com que apresente uma ainda mais elevada toxicidade para as células (Liochev, 2010).
O radical hidroxilo, também formado, como outros, pela expo-sição a radiações ionizantes (UV, raios X, radiação gama), é altamente reativo, o mais reativo e o mais deletério de todos radicais livres dos sistemas biológicos. De elevada inespecificidade, reage fácil e rapidamente com todas as biomoléculas ou outros radicais, da proximidade e mesmo do próprio local da sua formação. Este radical livre é o principal responsável pelo início do processo em cadeia da lipoperoxidação, e pelos processos oxidativos das proteínas e do DNA nuclear e mitocondrial.
Esta atividade oxidativa não pode unicamente ser atribuída aos radicais livres ou espécies não radicalares atrás referidos, mas também a outros como, e.g. os radicais peroxílo (ROO•) e o hidroperóxido (ROOH•) ou a espécie oxigénio singleto (1O2) e o ácido hipocloroso (HOCl). Acresce ainda, que esta atividade oxidativa não se limita às espécies reativas de oxigénio, podendo igualmente provir de outras espécie moleculares. De facto, existe outro tipo de espécies reativas, como os radicais livres de nitrogénio, dos quais se realçam o óxido nitrilo (NO•), o dióxido nitrilo (NO2
•) ou o nitroperóxido (ONOO•). Qualquer uma destas espécies, de elevada capacidade oxidante, se não forem devidamente controladas, podem facilmente exercer os seus efeitos deletérios, causando efeitos lesivos sobre as macromoléculas constituintes das distintas estruturas celular, podendo modificar a funcionalidade e condicionar por vezes a própria viabilidade celular (Halliwell, 2012).

34
5.1.1. Marcadores de lesão oxidativaDada a sua existência efémera, estes radicais são de difícil
quantificação e de deteção direta. No entanto, a sua produção pode ser avaliada de uma forma indireta recorrendo à determinação laboratorial de compostos designados como marcadores de lesão, que mais não são do que os produtos ou metabolitos resultantes dessa inespecífica atividade oxidativa e lesiva que se manifesta, nomeadamente, sobre (i) os lípidos (peroxidação lipídica), alterando a estrutura e a permeabilidade membrana celular, (ii) as proteínas (oxidação dos grupos sulfidrilo a pontes dissulfureto e oxidações de aminoácidos, ruturas, etc.) provocando a inativação e perda de funcionalidade biológica de proteínas da membrana ou de enzimas, e (iii) os ácidos nucleicos (oxidação da suas bases nucleotídeas) dando origem a mutações e interferindo com os processos de transmissão genética, entre outros (Barbosa e col, 2010; Wagner e col, 2011; Barrera, 2012; Teixeira e col, 2013; Silva e col, 2014; Kliszczewicz e col, 2015; Ribon e col, 2016; Wiecek e col, 2016).
Entre os variados marcadores utilizados laboratorialmente naavaliação da ação oxidativa sobre os lípidos, a determinação labo-ratorial do malondialdeído (MDA), produto secundário (aldeído) da lipo-peroxidação lipídica, e das substâncias que reagem com o ácido tiobarbitúrico (TBARS) têm sido os mais frequentemente utilizados. Outros marcadores, contudo, têm sido igualmente referenciados e usados para este fim, como os isoprostanos (PGF2-alfa-8-isoprostano) e os lipoperóxidos, entre outros. Da sua ação sobre deletéria os lípidos, nomeadamente sobre os ácidos gordos insaturados (peroxidação lipídica) das membranas celulares, resultam alterações na sua permeabi-lidade e estrutura e podem levar à inviabilidade e morte celular.
Do mesmo modo, os marcadores de lesão ou da ação oxidativa sobre as proteínas, e mais usuais sob o ponto de vista laboratorial, são os grupos tiol (SH) e dissulfureto (S-S), os grupos carbonilo, compostos (carbonilados ou dicarbonilados) que resultam da ação direta oxidativa dos radicais livres sobre as cadeias laterais dos aminoácidos, a nitrotirosina, e a 2-oxohistidina e a (2-oxo) orto-tirosina, entre outros.

35
A determinação do 8-hidroxi-2’-deoxiguanosina (8-OHdG) urinário, a 5-hidroximetil-2’-desoxiuridina), o 8-nitroguanina e a timidina glicol (TG) têm sido os marcadores mais utilizados laboratorialmente no estudo da ação oxidativa sobre os ácidos nucleicos.
5.2. Mecanismos de produção de radicais livresNa célula, as espécies reativas de oxigénio podem ser globalmente
formadas a nível da mitocôndria, do citoplasma ou da membrana celular (Castrogiovanni e Imbesi, 2012).
Entre os distintos mecanismos ou processos celulares descritos, como fonte ou responsáveis pela produção de radicais livres, a cadeia de transporte de eletrões na mitocôndria e a atividade da Xantina oxidase, através da sua intervenção no metabolismo das purinas, adquirem importância fundamental.
5.2.1. Cadeia de transporte de eletrões (mitocôndria)O exercício aeróbio está associado a um aumento do consumo
de oxigénio, aumento esse que será tanto maior quanto maior for a intensidade e a duração do mesmo.
Este consumo de oxigénio está por sua vez associado a uma for-mação continuada de ROS, estimando-se que em condições meta-bólicas basais, cerca de 1 a 5% da totalidade de oxigénio consumido seja responsável por esta produção (Fridovich, 1978; Turrens, 2003; Finaud e col, 2006; Halliwell e Gutteridge, 2007).
Ao longo da cadeia respiratória, localizada na membrana interna da mitocôndria, os eletrões vão sendo sucessivamente transferidos, entre os distintos complexos enzimáticos que a compõem, até ao seu aceitador final, o oxigeno, o qual é reduzido a água, de acordo com o aumento crescente de positividade do potencial redox. No entanto, ao longo desse percurso, alguns dos eletrões podem reagir diretamente com o oxigénio molecular dando origem de um modo mais particular, ao radical superóxido (O
2•–), a peróxido de hidrogénio (espécie de
oxigénio reativa, mas não radical livre) e o radical hidroxilo (HO•).

36
Esta produção de radicais livres (radical superóxido, o primeiro a ser formado), parece ocorrer preferencialmente ao nível do complexo I (complexo NADH desidrogenase, NADH-Ubiquinona oxidoredutase) e em particular do complexo III (complexo citocromo bc1, Ubiquinol-citocromo c oxidoredutase), com o envolvimento dos seus centros prostéticos FMN e Fe-S (Ji, 1999; Watson e col, 2005; Kudin e col, 2005; Gandra e col, 2006; Powers e col, 2011).
Assim, à medida que aumenta o fluxo de eletrões ao longo da cadeia respiratória da mitocôndria (i.e. quanto maior for a aumento do oxigénio consumido) maior é a quantidade de espécies de oxigénio reativas geradas.
Durante a realização exercício físico, e de acordo com as suas características, nomeadamente de intensidade e duração, a envolvente muscular é acentuada e exigente sob o ponto de vista de necessidades energéticas, tendo sido descritos aumentos de volume de oxigénio consumido por parte do tecido muscular, 100 a 200 vezes superiores aos valores apresentados em condições basais (Sem, 1995; Gandra e col, 2006). Estes acréscimos de volumes de oxigénio consumido explicam e corroboram a produção favorecida de radicais livres, potenciada ainda pelo aumento de volume de oxigénio consumido por parte do organismo, no exercício intenso, cujos valores descritos apontam para 10 a 20 vezes os valores normais (Halliwell e Gutteridge, 2007).
Este processo de formação de radicais livres, adquire assim im-portância mais fundamentada na prática de exercícios eminentemente aeróbios, típicos de atividades físicas contínuas e prolongadas.
5.2.2. Xantina oxidase (metabolismo purínico)Para além da assunção da mitocôndria como principal local de
produção fonte geradora deste tipo de espécies oxidativas e radicais livres durante a realização de exercício físico, a atividade da xantina oxidase tem merecido a maior relevância neste âmbito.
Durante a prática de exercício físico, ocorre consumo de ATP, que será tanto mais significativo quanto maior for a intensidade do esforço despendido, dando origem a quantidades significativas de hipoxantina,

37
como resultado do aumento do consumo de ATP necessário para cobrir as suas necessidades energéticas. Uma vez formada, a hipoxantina, é por sua vez convertida em xantina e esta em ácido úrico, produto final do metabolismo das purinas, numa reação catalisada pela enzima xantina oxidase.
Aquando da prática desportiva, ocorrem, adicionalmente, situa-ções de transitórias de hipoxia, mais intensas quanto mais agudo for o esforço despendido, devido à necessidade de aportes sanguíneos diferenciados entre os distintos tecidos envolvidos, tendo em vista o favorecimento da perfusão do tecido muscular. Estes “microambientes” de hipoxia podem ser ainda potenciados pelo facto do próprio tecido muscular poder ele mesmo estar envolvido nestas situações quando perante circunstâncias de elevada intensidade de exercício (Hellsten e col, 1997). Durante este estado e aquando a subsequente reperfusão (reoxigenação) destes tecidos observa-se a produção de ROS.
Um dos mecanismos proposto neste processo de isquémia-reperfusão e mais aceite como responsável pelo aumento da produção destas espécies oxidantes, parece ser mediado pela intervenção direta da xantina oxidase.
A Xantina desidrogenase, enzima que normalmente utiliza o NAD+ como cofactor enzimático, in vivo e em condições de homeostasia, nestas circunstâncias de hipoxia (isquémia-reperfusão) e/ou na presença de grandes quantidades de seu substrato, pode também sofrer alterações estruturais e funcionais de natureza proteolítica, sendo convertida de uma forma irreversível em Xantina Oxidase, passando a utilizar o oxigénio como recetor de eletrões (McCord, 1985; Chung e col, 1997; Vollaard e col, 2005; Ferreira e col, 2007). Desta atuação como oxidase, resulta a produção direta do radical superóxido, o qual pode dar origem subsequente a outros ROS (Hellesten, 2000; Mc Ardle e col, 2001; Chevion e col, 2003).
Esta conversão Xantina desidrogenase em Xantina oxidase (e concomitante aumento de produção de radicais livres) é potenciada em todos os casos de realização de exercícios físicos até à exaustão ou de elevada intensidade e intermitentes (curta duração), onde o consumo

38
de ATP, nestas condições de anaerobiose e por ativação da miocínase, é acentuado. São exemplos típicos de exercícios, a musculação, os sprints, treinos em altitude, alpinismo, entre outros.
5.2.3. Outras fontesPara além destes considerados como os principais mecanismos
ou processos geradores de espécies oxidativas, durante a realização de exercício físico tem sido propostas outras fontes de produção de ROS, como sejam, a auto oxidação de catecolaminas, auto-oxidação da mioglobina e hemoglobina, ativação de neutrófilos, reações envolvendo o ferro nos microssomas (cit. P450), lisossomas, células endoteliais, entre outras (Duarte e col, 1993; Magder, 2006; Moncada e col, 2006; Lambeth e col, 2014).
Em resposta ao exercício observa-se um aumento da produção de catecolaminas, em particular, a adrenalina e a noradrenalina, que será mais significativo quanto maior for a intensidade do mesmo. Quer os seus processos de auto-oxidação (processo não enzimático) quer os envolvidos na sua oxidação enzimática, mediada pela Monoami-noxidase (MAO) e a Catecol-O-metil transferase (COMT), são fontes potenciais de produção de espécies oxidantes reativas, como o peroxido de hidrogénio, que por sua vez pode dar origem ao radical hidroxilo (HO•) mais reativo e mais deletério para o organismo (Ishii e col, 1993; Duarte e col, 2004; Watson e col, 2005; Umbarkara e col, 2015).
Do mesmo modo, a hemoglobina e a mioglobina podem sofrer processos de auto-oxidação dando origem igualmente ao radical superóxido e peroxido de hidrogénio, (Gunther e col, 1999; Baron e Andersen, 2002; Reeder e col, 2004). A auto-oxidação da hemoglobina pode dar origem à metahemoglobina (Fe3+) (Cooper e col, 2002).
A ativação de neutrófilos e macrófagos é outra das hipóteses justificativas do aumento da produção de espécies reativas de oxigénio e dos efeitos de stress oxidativo. De um modo sucinto, a prática de exercício físico gera a ocorrência de microlesões a nível das fibras musculares, com o consequente desenvolvimento de uma resposta inflamatória, e a chamada dos leucócitos. A libertação subsequente de

39
fatores quimiotáticos (e.g. interleucinas, alfa-TNF) estimula a ativação das células circulantes.
Os leucócitos, em particular, os neutrófilos e os macrófagos têm uma ação interventiva nos processos defesa e de reparação dos tecidos lesados do organismo. Durante e como resposta ao exercicio verifica-se um aumento transitório do número de leucócitos (com predomínio dos neutrófilos) circulantes, atribuído a um processo de desmarginalização destas células. Para este processo, para além do quadro inflamatório, contribui igualmente o aumento dos níveis de catecolaminas e de cortisol, que ocorre em consequência do exercício que será tanto mais quanto maior for a duração e intensidade do mesmo (Smith e col, 1990; Robson e col, 1999; Shephard, 2003; Pyne, 2004; Butterfield e col, 2006; Schneider e Tiidus, 2007; Ascensao e col, 2008). Ao fim de algum tempo, 24-48 horas, dependendo da intensidade e duração do exercício, o número de neutrófilos (leucócitos) plasmáticos diminui, uma vez que, por efeitos de diapedese, invaginam para os locais de inflamação, passando dos capilares sanguíneos para o tecido conjuntivo, local de inflamação (Wolach e col, 2005; Magalhaes e col, 2007, 2010; Ascenção e col, 2008). Após estas células circulantes atingirem os locais de intervenção, libertam enzimas proteolíticas, lisozimas, que intervêm nos processos de limpeza e eliminação dos restos celulares e das proteínas danificadas. Estas ações são mediadas por mecanismos de fagocitose, e a destruição dos microrganismos mediadas pela intervenção de ROS por elas produzidas, em particular, pela ação das NADPH oxidases e mieloperoxidases (Leeuwenburgh e Heinecke, 2001; Cooper e col, 2002; Halliwell e Gutteridge, 2015).
De todas as formas, parece ser consensual, que o incremento de radicais livres observado durante o exercício físico é proveniente de variadas destas fontes de produção em simultâneo e não de apenas de uma, ou de cada uma delas, isoladamente.
5.3. Capacidade antioxidanteComo resposta a esta elevada carga de espécies ou radicais livres,
com capacidade oxidativa, o organismo desenvolve mecanismos de

40
adaptativos de defesa antioxidante, de natureza enzimática e não enzimática, no sentido de minorar ou no limite anular os efeitos lesivos por elas induzido.
Como componentes do sistema de defesa antioxidante de natureza enzimática, sob o ponto de vista fisiológico, são de realçar as enzimas: (i) Superóxido dismutase (SOD), que atua na prevenção do ião superóxido, intervindo cataliticamente na conversão do radical superóxido em peroxido de hidrogénio. Constituindo a primeira linha de defesa enzimática, existe em 2 formas, a Cu-Zn-SOD, presente no citoplasma, e a Mn-SOD mitocondrial; (ii) Glutationa peroxidase (GPx), dependente o selénio, e a Glutationa redutase (GR), ambas intervenientes no ciclo de oxidação/redução da glutationa. A GPx catalisa a reação de redução do peroxido de hidrogénio e de outros peróxidos, a água e aos álcoois, respetivamente. A GR intervém no processo enzimático de regeneração de glutationa reduzida (GSH) através da redução da GSSG (glutationa oxidada), utilizando como cofactor o NADPH; e (iii) Catalase, uma hemoproteína, que intervém na decomposição enzimática do peróxido de hidrogénio, presente na mitocôndria e peroxissomas (Halliwell, 2012).
No entanto, existem outros compostos de natureza não enzimática, endógenos ou exógenos, cujas capacidades antioxidantes contribuem igualmente para a fisiológica defesa adaptativa, antioxidante (Halliwell e Gutteridge, 2010; Nieman e col, 2010; Finley e col, 2011). Entre os vários compostos são de realçar, entre outros, glutationa, ácido úrico, albumina, bilirrubina, transferrina e a ceruloplasmina, o beta-caroteno (vitamina A), o ácido ascórbico (vitamina C), esta não sintetizada no organismo, apernas obtida de origem exógena, o alfa-tocoferol (vitamina E), compostos polifenóis, como os flavonoides, e alguns minerais, como o selénio.
Quando a quantidade de moléculas ou partículas com capacidade potencialmente oxidante e deletéria superarem a capacidade de defesa do organismo, os efeitos oxidativos prevalecem sobre os mecanismos de defesa e observam-se danos nas distintas moléculas e tecidos. Este desequilíbrio toma a designação de stress oxidativo. Deste modo, e de

41
um modo global, este quadro pode instalar-se como resultado de um aumento da produção de radicais livres, da diminuição da capacidade de defesa antioxidante ou da associação destes dois fatores. De realçar, contudo, que mesmo em condições normais fisiológicas observa-se um contínuo desequilíbrio destes sistemas a favor deste estado de stress oxidativo, o qual progride à medida que a idade avança.
A avaliação laboratorial destes mecanismos protetores contra os efeitos deletérios do stress oxidativo, é efetuada, normalmente, pela determinação da atividade das enzimas que constituem o sistema de defesa enzimático, SOD, GPx, GR e catalase, e/ou através do recurso à quantificação de qualquer um ou de vários dos compostos ou moléculas de natureza não enzimática, como a glutationa, o ácido úrico e as distintas vitaminas com capacidade antioxidante.
A glutationa (gama-glutamilcisteinilglicina) é um dos mais importantes compostos endógenos com atividade antioxidante, fa-zendo parte de um dos mais significativos sistemas de defesa dos radicais livres e dos xenobióticos. As suas propriedades como antioxi-dante residem no grupo sufidrilo (-SH) da molécula de cisteína, seu constituinte. Para além de atuar na neutralização do peroxido de hidrogénio e dos peróxidos lipídicos, intervém igualmente como “scavenger” de radicais livres ou de outras espécies reativas de oxigénio (e de nitrogénio).
Faz parte integrante do ciclo da glutationa onde pode ser encontrada quer na forma reduzida (GSH) quer na forma oxidada (GSSG). O equilíbrio quantitativo entre estas duas formas está condicionado pela proporção entre compostos oxidantes e antioxidantes. Em caso de stress oxidativo ou de inativação de qualquer composto oxidante ocorre um aumento de produção da forma GSSG com o concomitante incremento do consumo da forma GSH. Em situações normais fisiológicas do sistema redox, a regeneração enzimática da forma GSH ocorre no ciclo da glutationa, numa reação catalisada pela GR na presença de NADPH. A determinação de GSSG, de GSH e/ou da razão GSSH/GSH constitui pois bons indicadores do estado de stress oxidativo celular (Rover e col, 2004).

42
Do mesmo modo, a presença de compostos oxidantes como resultado de situação de stress oxidativo podem igualmente atuar sobre os grupos SH das proteínas, convertendo-os em pontes dissulfureto (S-S) com as consequentes a alterações estruturais e perda de função biológica. Níveis aumentados de GSSG, ao interferirem com o sistema redox intracelular, podem torná-lo mais oxidante e amplia e potencia esta situação. É por esta razão que a determinação laboratorial dos grupos SH e/ou dos grupos S-S têm sido utilizados, igualmente, como marcadores na avaliação do stress oxidativo.
O ácido úrico, produto final da ação da xantina oxidase e da xantina desidrogenase sobre a xantina (e hipoxantina), é considerado um bom indicador na avaliação do catabolismo purínico. Os seus ní-veis plasmáticos aumentam normalmente após a realização de exer-cício agudo de elevada intensidade. Possui uma relevante atividade antioxidante contribuindo deste modo para minorar os efeitos de stress oxidativo, nomeadamente a nível plasmático onde esse contri-buto é significativo (Maxwell e col, 1997; Hooper e col, 2000; Bailliee col, 2007). A sua capacidade antioxidante manifesta-se essencial-mente sobre os radicais hidroxilo e peroxinitrilo e sobre os grupos peróxidos, apresentando uma ação reduzida ou nula sobre os radicais superóxidos (Hellsten e col, 1997; Waring e col, 2003; Sautin e Johnson, 2008).
A avaliação da capacidade antioxidante total em fluidos biológicos pode ser ainda realizada utilizando-se algumas determinações labo-ratoriais como o TAS (capacidade ou “status” antioxidante total). A determinação, in vitro, deste parâmetro laboratorial, TAS, num dado fluido biológico (e.g. plasma, citoplasma celular), mede apenas a capacidade antioxidante total ou de inibição da produção e radicais livres nele presente, e dá-nos uma ideia do equilíbrio entre os sistemas antioxidantes e oxidantes aí existente. A grandeza ou o valor total dessa determinação está essencialmente condicionada pela contribuição das proteínas (10-28%), do ácido úrico (7-58%) e do ácido ascórbico (3-27%) (Wayner e col, 1987).

43
5.4. Exercício físico e stress oxidativo. Avaliação laboratorialDurante a prática de exercício físico ocorre um aumento da
produção radicais livres, como resultado das necessidades metabólicas do organismo, que se não for devidamente compensado, pode con-tribuir para a existência de danos ou lesões a nível celular e tecidual afetando o desempenho ou o rendimento desportivo do atleta (Powers e Jackson, 2008).
De um modo geral, a intensidade, a frequência e a duração do exercício físico são grandezas que podem modificar, de um modo significativo, este equilíbrio entre produção de radicais livres e sistemas de defesa antioxidante, induzindo quadros de stress oxidativo por vezes acentuados (Radak e col, 2008a).
Estudos demonstram que, quanto maior for a intensidade do exercício (VO
2max superiores a 70 % dos valores basais) maior é a formação de radicais livres e maior é a lesão oxidativa. Nestas situações, e de um modo geral, a resposta do sistema enzimático de proteção antioxidante celular é estimulada, traduzindo-se no aumento da atividade das enzimas GPx, SOD e Catalase. A diminuição de níveis de GSH e dos grupos tiol das proteínas e o aumento de GSSG e de ácido úrico constituem igualmente algumas alterações do sistema antioxidantenão enzimático, entre outras, que acompanham estas situações.
Quando perante situações de exercício físico de elevada intensidade e duração, os praticantes ou atletas podem estar sujeitos a estes desequilíbrios, que se poderão traduzir em lesões e danos oxidativos associados. Treinos de elevado volume e frequência podem igualmente ser causa de idênticos resultados (Palazzetti e col, 2003; Vollaard e col, 2005; Knez e col, 2006; König, 2007; Fisher-Wellman e Bloomer, 2009).
Estas alterações fisiológicas ocorrem igualmente como resposta ao exercício anaeróbio. No entanto, estudos apontam no sentido de, perante a mesma intensidade, a produção de radicais livres em aerobiose ser mais elevada no início do exercício, ao contrário do que ocorre em situações de anaerobiose, em que essa produção pode ser mais prolongada no tempo (Minyi e col, 2007).

44
Em ambos os casos, contudo, estes quadros de stress oxidativo apresentam-se mais moderados e mais reduzidos em atletas treinados, quando comparados com sedentários ou com atletas ou desportistas menos treinados (Vollaard e col, 2005; Patil e col, 2012; Scheffer e col, 2012).
Este dado sugere que prática continuada (crónica) de exercício pode induzir adaptações fisiológicas que reduzem os efeitos do stress oxidativo. Esta adaptação pode ser explicada pelo aumento das defesas antioxidantes ou da redução de produção de espécies oxidantes durante e após a realização do exercício físico. O incremento no mecanismos de upregulation endógena dos sistemas antioxidantes enzimático e não enzimático e/ou um mais eficaz fluxo de eletrões ao longo da cadeia de transporte de eletrões, diminuindo a “perda” de eletrões para o radical superóxido, foram mecanismos propostos para mediar essa adaptação (Ji, 1999; Bloomer e Goldfarb, 2004; Finaud e col, 2006; Shi e col, 2007; Wagner e col, 2011).
O desequilíbrio entre capacidade de defesa antioxidante e produção de radicais livres com capacidade deletéria oxidante, i.e. stress oxidativo, pode ser avaliado e estudado laboratorialmente com a determinação de variados marcadores de defesa antioxidante e/ou de marcadores de lesão oxidativa. A resposta do organismo perante a indução de quadros de stress oxidativo por parte da prática de exercício físico de elevada intensidade foi avaliada e analisada em distintos estudos efetuados com atletas de elite e de distintas modalidades desportivas alta de competição.
5.4.1. FutebolDurante realização de um jogo de futebol ocorrem alterações
significativas de natureza fisiológica, sendo o aumento dos batimentos e do fluxo sanguíneo cardíaco, o aumento da libertação das catecolaminas e a mobilização leucocitária, alguns dos exemplos mais expressivos. Dadas as suas especificidades, o futebol é um exemplo de modalidade desportiva de alta competição, predominantemente

45
aeróbico mas com uma carga anaeróbia considerável (a concentração de lactato durante uma partida de futebol competitiva pode atingir valores de 10 a 12 mmol/L), geradora de uma elevada produção de espécies reativas oxidantes e de radicais livres, com elevada capacidade oxidante e causa potencial de lesão oxidativa, fatores indutores ou contributivos para o desenvolvimento de quadros de stress oxidativo.
No sentido de avaliar o efeito do exercício físico e a prática desportiva de futebol competitivo como indutor de quadros de stress oxidativo, procedeu-se a um estudo cujo objetivo, entre outros, consistia na avaliação de uma série de parâmetros relacionados com possíveis alterações de parâmetros de natureza bioquímica, hormonal e de stress oxidativo, ao longo de toda uma temporada competitiva (Silva e col, 2014). Catorze jogadores (idade, 25,7 ± 4,6 anos; altura, 178,1 ± 5,7 cm; peso, 76,5 ± 9,2 Kg) da Liga secundária de Futebol profissional portuguesa participaram neste estudo. As colheitas de material biológico para a determinação laboratorial dos parâmetros a avaliar, foram efetuadas no início da pré-época (primeira, PE), a meio da época desportiva, no mês de Janeiro (segunda, ME), no final do campeonato (terceira, FE). As segundas e terceiras colheitas foram realizadas 72 h após a realização de um jogo de futebol. Uma quarta colheita foi realizada no período compreendido entre o fim da temporada desportiva e o início da pré-época (FIE).
Os resultados obtidos, que se podem observar nas Figura 11 e Figura 12, globalmente evidenciam um significativo quadro de stress oxidativo induzido pela prática futebolística, ao longo de toda a temporada profissional.
O perfil de atividade da enzima SOD, a primeira linha de defesa antioxidante contra o stress oxidativo (Finaud e col, 2006), que aumenta ao longo da época desportiva, parece sugerir um mecanismo adaptativo no sentido de contrariar a existência de uma atividade oxidativa continuada (atribuída ao radical superóxido) e inerente ao stress oxidativo.
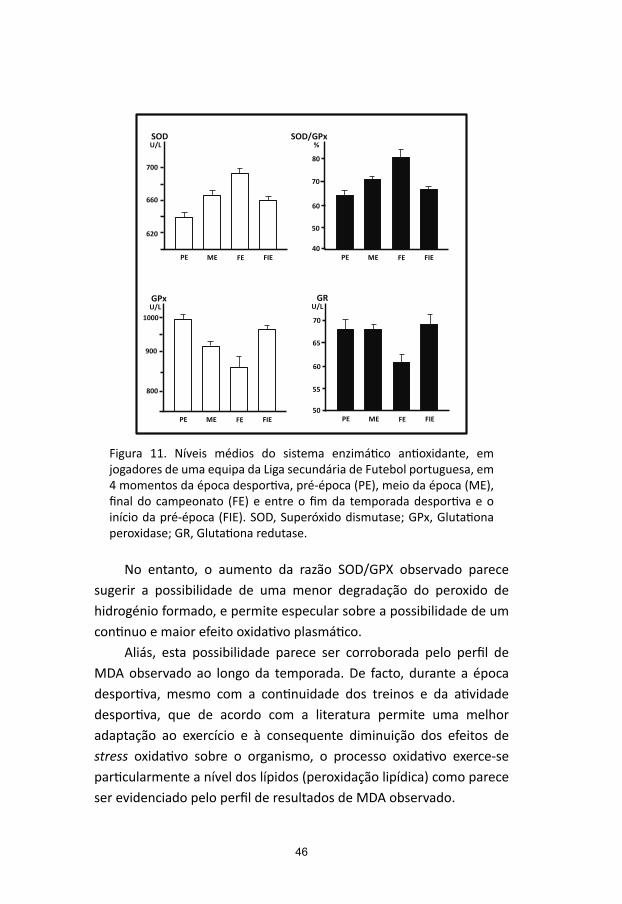
46
Figura 11. Níveis médios do sistema enzimático antioxidante, em jogadores de uma equipa da Liga secundária de Futebol portuguesa, em 4 momentos da época desportiva, pré-época (PE), meio da época (ME), final do campeonato (FE) e entre o fim da temporada desportiva e o início da pré-época (FIE). SOD, Superóxido dismutase; GPx, Glutationa peroxidase; GR, Glutationa redutase.
No entanto, o aumento da razão SOD/GPX observado parece sugerir a possibilidade de uma menor degradação do peroxido de hidrogénio formado, e permite especular sobre a possibilidade de um continuo e maior efeito oxidativo plasmático.
Aliás, esta possibilidade parece ser corroborada pelo perfil de MDA observado ao longo da temporada. De facto, durante a época desportiva, mesmo com a continuidade dos treinos e da atividade desportiva, que de acordo com a literatura permite uma melhor adaptação ao exercício e à consequente diminuição dos efeitos de stress oxidativo sobre o organismo, o processo oxidativo exerce-se particularmente a nível dos lípidos (peroxidação lipídica) como parece ser evidenciado pelo perfil de resultados de MDA observado.
Fig. 11. Níveis médios do sistema enzimático antioxidante, em jogadores de uma equipa da liga secundária de futebol portuguesa, em 4 momentos da época desportiva, pré-época (PE), meio da época (ME), final do campeonato (FE) e entre o fim da temporada desportiva e o início da pré-época (FIE). SOD, Superóxido dismutase; GPx, Glutationa peroxidase ; GR, Glutationa redutase. Fig.12
SOD/GPx
PE FIE ME FE
50
%
70
80
60
40
GR
PE FIE ME FE
55
U/L
65
70
60
50
PE FIE ME FE
620
SOD U/L
700
660
PE FIE ME FE
800
GPx U/L
1000
900

47
Figura 12. Níveis médios de marcadores de stress oxidativo, em jogadores de uma equipa da Liga secundária de Futebol portuguesa, em 4 momentos da época desportiva, pré-época (PE), temporada desportiva e o início da pré-época (FIE). TAS, Capacidade (“status”) antioxidante total; UA, Ácido úrico; MDA, Malonaldeído; SH, Grupos tiol.
O que se pode concluir, com base na análise global dos marcadores utilizados no estudo e avaliação do stress oxidativo, é que ao longo de uma época desportiva, os jogadores de futebol sofrem os efeitos oxidativos induzidos pela prática competitiva. Contudo, esses efeitos não se manifestam continuadamente, uma vez que são revertidos aos valores basais no período de repouso pós-temporada.
O que se pode concluir, com base na análise global dos marcadores utilizados no estudo e avaliação do stress oxidativo, é que ao longo de uma época desportiva, os jogadores de futebol sofrem os efeitos oxidativos induzidos pela prática competitiva. Contudo, esses efeitos não se manifestam continuadamente, uma vez que são revertidos aos valores basais no período de repouso pós-temporada.
Fig. 12. Níveis médios de marcadores de stress oxidativo, em jogadores de uma equipa da liga secundária de futebol portuguesa, em 4 momentos da época desportiva, pré-época (PE), meio da época (ME), final do campeonato (FE) e entre o fim da temporada desportiva e o início da pré-época (FIE). TAS, Capacidade (“status”) antioxidante total ; UA, Ácido úrico; MDA, Malonaldeído ; SH, Grupos tiol. Fig.11
PE FIE ME FE
4,00
MDA mmol/g
12,00
8,00
PE FIE ME FE
1,50
SH μmol/g
3,00
2,50
2,00
PE FIE ME FE
1,00
TAS mM
1,30
1,20
PE FIE ME FE
5,0
UA mg/dL
5,5
1,00 0,00
4,5 0,80

48
Perante estes resultados globais, é interessante observar as variações bioquímicas ocorridas nos momentos imediatamente posteriores à realização de um jogo de futebol sobre os níveis plasmáticos de stress oxidativo (Ascensão e col, 2008). Para isso, foram efetuadas colheitas de material biológico, para a determinação laboratorial de variados parâmetros utilizados laboratorialmente no estudo do efeito do exercício físico sobre o stress oxidativo, antes e após 30 minutos, 24, 48 e 72 horas da realização de um jogo amigável e competitivo entre 16 futebolistas (idade, 21,3 ± 1,1 anos; altura, 175,0 ± 6,0 cm; peso, 70,7 ± 6,3 Kg) da segunda Liga de Futebol profissional portuguesa. Na avaliação do stress oxidativo induzido pelo futebol, neste estudo, utilizaram-se como marcadores bioquímicos plasmáticos de defesa e lesão oxidativa, o MDA, os grupos SH, o TAS e o UA.
Como se pode observar na Figura 13, após a realização do jogo amigável de futebol (2x 45 minutos) e durante o período de recuperação, os níveis plasmáticos de MDA aumentaram significativamente até às 72 horas; os valores plasmáticos dos grupos sulfidrilo, no entanto, foram com exceção dos determinados às 72 horas, inferiores aos correspondentes valores basais. Os valores plasmáticos de TAS aumentaram significativamente aos 30 minutos após a realização do jogo. Os níveis de UA plasmático aumentaram significativamente durante o período de recuperação em análise. Estes valores demonstraram que a prática de um jogo de futebol amigável, mas competitivo, induzem alterações quantitativas a nível dos marcadores plasmáticos de lesão oxidativa, bem como, da capacidade antioxidante total. Este incremento da capacidade oxidativa teve uma incidência sobre os lípidos, como se pode inferir do aumento verificado de produtos da peroxidação lipídica, o malonaldeído (MDA).
Por outro lado, a ação oxidativa sobre as proteínas pode ser observada através da diminuição dos grupos sulfidrilo (SH), o que implica, desde logo, o aumento das ligações dissulfidrilo (S-S) presentes nas proteínas, bem como, na glutationa reduzida.
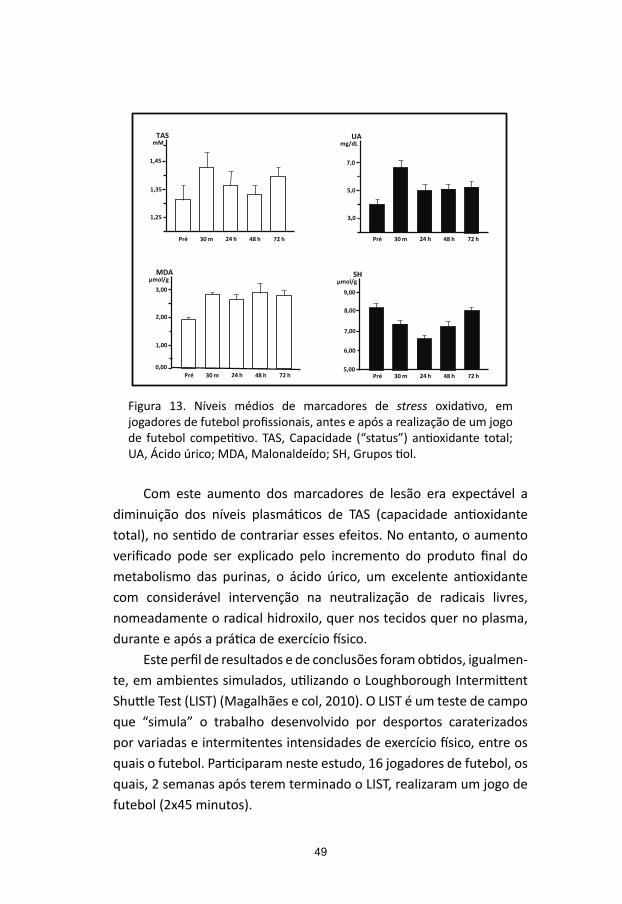
49
Figura 13. Níveis médios de marcadores de stress oxidativo, em jogadores de futebol profissionais, antes e após a realização de um jogo de futebol competitivo. TAS, Capacidade (“status”) antioxidante total; UA, Ácido úrico; MDA, Malonaldeído; SH, Grupos tiol.
Com este aumento dos marcadores de lesão era expectável a diminuição dos níveis plasmáticos de TAS (capacidade antioxidante total), no sentido de contrariar esses efeitos. No entanto, o aumento verificado pode ser explicado pelo incremento do produto final do metabolismo das purinas, o ácido úrico, um excelente antioxidante com considerável intervenção na neutralização de radicais livres, nomeadamente o radical hidroxilo, quer nos tecidos quer no plasma, durante e após a prática de exercício físico.
Este perfil de resultados e de conclusões foram obtidos, igualmen-te, em ambientes simulados, utilizando o Loughborough Intermittent Shuttle Test (LIST) (Magalhães e col, 2010). O LIST é um teste de campo que “simula” o trabalho desenvolvido por desportos caraterizados por variadas e intermitentes intensidades de exercício físico, entre os quais o futebol. Participaram neste estudo, 16 jogadores de futebol, os quais, 2 semanas após terem terminado o LIST, realizaram um jogo de futebol (2x45 minutos).
Fig. 13. Níveis médios de marcadores de stress oxidativo, em jogadores de futebol profissionais, antes e após a realização de um jogo de futebol competitivo. TAS, Capacidade (“status”) antioxidante total ; UA, Ácido úrico; MDA, Malonaldeído ; SH, Grupos tiol.
Fig.13
1,00
MDA μmol/g
3,00
2,00
1,25
TAS mM
1,45
1,35
SH μmol/g
3,0
UA mg/dL
7,0
5,0
Pré 48 h 30 m 24 h 72 h
Pré 48 h 30 m 24 h 72 h
Pré 48 h 30 m 24 h 72 h
Pré 48 h 30 m 24 h 72 h
6,00
9,00
8,00
7,00
5,00 0,00

50
O estudo foi objetivado no sentido de comparar os dois exercícios, teoricamente equivalentes em termos comparativos de intensidade e duração do esforço despendido, no que respeita, entre outos, ao efeito induzido no stress oxidativo e à lesão muscular associada. O que se pode concluir do estudo realizado é que o perfil dos valores em todos os parâmetros estudados, e em ambos os exercícios efetuados, foram similares entre si, variando apenas na sua grandeza. De facto, verificou-se, na pratica efetiva de um jogo de futebol, alterações mais significativas nos parâmetros de stress oxidativo, embora no mesmo sentido, que no LIST. No entanto, não se encontraram dife-renças significativas nos parâmetros de estudo muscular (CPK e Mioglobina).
Em resumo, o futebol, enquanto modalidade desportiva, é fonte considerável de desenvolvimento de stress oxidativo. Durante e após a sua prática, todas as condições e potenciais mecanismos de produção de radicais livres ou espécies oxidativas de oxigénio (e de nitrogénio) estão altamente favorecidas. O consumo aumentado de oxigénio, o incremento da atividade da xantina oxidase, bem como de situações transitórias de isquémia-reperfusão, os estados de inflamação associados à atividade ou lesões musculares, são alguns dos exemplos que permitem explicar e justificar este acréscimo de produção. No entanto, em resposta a esta potencial e desfavorável situação oxidativa e deletéria, o organismo é capaz de promover e desenvolver mecanismos protetores de defesa antioxidante, no sentido de obstar a estes efeitos de grande potencial deletério. A cessação desta atividade desportiva, como acontece em final dos períodos de competição, parece ter como consequência o retorno aos níveis fisiológicos basais de stress oxidativo, como pode ser comprovado pela recuperação dos níveis plasmáticos pré-esforço dos biomarcadores utilizados.
5.4.2. Escalada indoorA escalada indoor é uma modalidade desportiva cujo objetivo
consiste em atingir o topo de uma parede artificial, através de pontos

51
de apoio (agarras) previamente fixados de acordo com o grau de dificuldade e do percurso pretendido.
É uma modalidade desportiva muito exigente sobre o ponto de vista fisiológico e físico. De uma forma intensa e com imensos períodos de contração isométrica intermitente e sustentada, estão envolvidos os músculos do antebraço, flexores das mãos (e dedos), em particular, mas também os bíceps e os músculos das costas, necessários à propul-são ascendente. Cerca de 37% do tempo gasto durante a ascensão competitiva é passado em posições estáticas (Magalhães e col, 2007).
Assim, e considerando as características inerentes à prática desta modalidade, 14 atletas (idade, 25,9 ± 2,4 anos; peso, 63,7 ± 3,2 Kg; altura, 171,4 ± 5,2 cm) realizaram subidas e descidas numa parede de escalada indoor, de um modo continuo e até à exaustão, no sentido de avaliar o efeito da prática competitiva de escalada sobre os marcadores de stress oxidativo. A recolha de material biológico foi efetuada antes, imediatamente após (tempo zero) e 1 hora após a realização do exercício. Procedeu-se à determinação de TAS, ácido úrico, MDA e grupos SH, e Glutationa, nas suas distintas formas, como marcadores de stress oxidativo (Magalhães e col. 2007).
Na Figura 14, pode observar-se o perfil dos níveis plasmáticos de TAS, ácido úrico, e dos marcadores de lesão oxidativa sobre os lípidos (MDA) e sobre as proteínas (SH). Observa-se um incremento dos níveis de TAS e de ácido úrico, imediatamente após a realização do exercício, que diminuem para valores mais próximos dos basais, anteriores à realização do exercício, ao fim de 1 hora.
Este aumento dos valores de TAS, que, como referido anterior-mente, é um parâmetro plasmático que avalia da capacidade antioxi-dante total in vivo, pode ser explicado, pelo menos em parte, pelos valores dos níveis plasmáticos de ácido úrico cujo perfil plasmático acompanha. O contributo deste produto final do metabolismo das purinas, com comprovada atividade antioxidante, como “scavenger” de radicais livres (e.g. hidroxilo e peroxilo), pode ser significativo (7-58%).

52
Fig. 14. Níveis médios de marcadores de stress oxidativo, em praticantes de escalada indoor, determinados antes (Pré), imediatamente após (0H) e 1 hora depois (1H) da realização do exercício de um modo continuo e até à exaustão. TAS, Capacidade (“status”) antioxidante total; UA, Ácido úrico; MDA, Malonaldeído; SH, Grupos tiol.
Os níveis plasmáticos de glutationa total, bem como das suas distintas formas, reduzida (GSH) e oxidada (GSSG), têm sidos usados como bons marcadores do stress oxidativo.
Como se pode observar na Figura 15, verifica-se uma redução dos níveis glutationa total e de GSH e um aumento de GSSG, quer em valor absoluto quer percentual, após a realização exercício à exaustão (0H). Ao fim de 1 hora, esses níveis plasmáticos ainda se mantêm inferiores e superiores, respetivamente, aos níveis pré-escalada. O aumento dos valores de GSSG era expectável e pode ser explicado pelo efeito antioxidante da GSH, em resposta ao aumento da produção de radicais livres e consequente estímulo oxidativo celular provocado pelo exercício físico.
Fig. 14. Níveis médios de marcadores de stress oxidativo, em praticantes de escalada indoor, determinados antes (Pré), imediatamente após (0H) e 1 hora depois (1H) da realização do exercício de um modo continuo e até à exaustão. TAS, Capacidade (“status”) antioxidante total ; UA, Ácido úrico; MDA, Malonaldeído ; SH, Grupos tiol.
Fig.14
SH μmol/g
10,00
8,00
Pré 0H 1H
290
MDA μmol/g
330
310
Pré 0H 1H
UA mg/dL
Pré 0H 1H
4,30
4,90
4,50
4,70
1,20
TAS mM
1,50
1,30
Pré 0H 1H
1,40
4,10 1,10
270 6,00
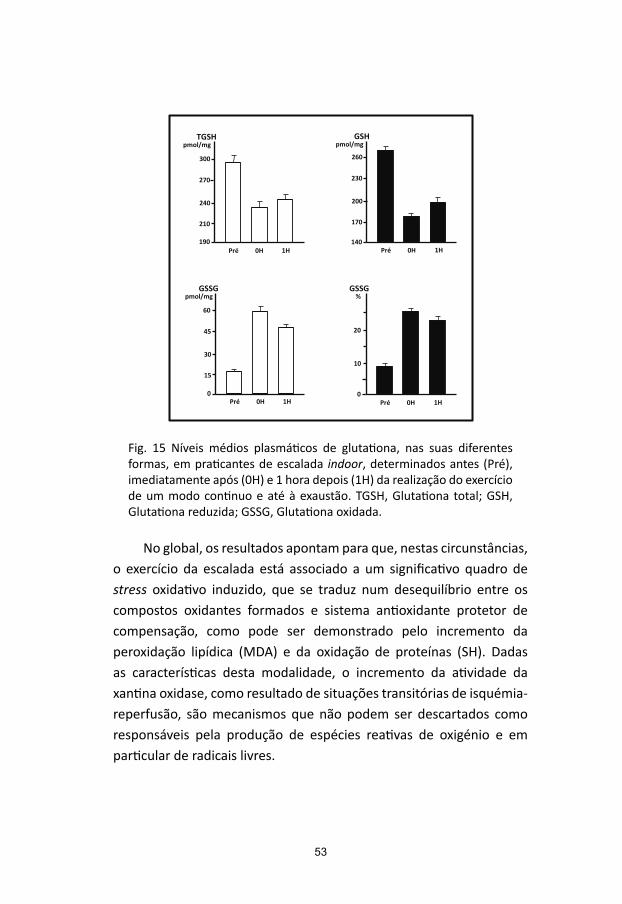
53
Fig. 15 Níveis médios plasmáticos de glutationa, nas suas diferentes formas, em praticantes de escalada indoor, determinados antes (Pré), imediatamente após (0H) e 1 hora depois (1H) da realização do exercício de um modo continuo e até à exaustão. TGSH, Glutationa total; GSH, Glutationa reduzida; GSSG, Glutationa oxidada.
No global, os resultados apontam para que, nestas circunstâncias, o exercício da escalada está associado a um significativo quadro de stress oxidativo induzido, que se traduz num desequilíbrio entre os compostos oxidantes formados e sistema antioxidante protetor de compensação, como pode ser demonstrado pelo incremento da peroxidação lipídica (MDA) e da oxidação de proteínas (SH). Dadas as características desta modalidade, o incremento da atividade da xantina oxidase, como resultado de situações transitórias de isquémia-reperfusão, são mecanismos que não podem ser descartados como responsáveis pela produção de espécies reativas de oxigénio e em particular de radicais livres.
Fig. 15 Níveis médios plasmáticos de glutationa, nas suas diferentes formas, em praticantes de escalada indoor, determinados antes (Pré), imediatamente após (0H) e 1 hora depois (1H) da realização do exercício de um modo continuo e até à exaustão. TGSH, Glutationa total ; GSH, Glutationa reduzida ; GSSG, Glutationa oxidada .
Fig.15
GSSG GSSG %
TGSH pmol/mg
GSH
170
pmol/mg
230
260
200
210
270
300
240
pmol/mg
0
20
10 15
45
60
30
Pré 0H 1H
Pré 0H 1H Pré 0H 1H
Pré 0H 1H 190 140
0

54
5.4.3. Exercício em tapete rolanteA prática de exercício físico é indutora de quadros de stress
oxidativo em atletas de elite, em diferentes modalidades desportivas de alta competição, como aliás estes e outros estudos comprovam (Powers e col, 2011). A intensidade é considerada um fator condicionante e da qual depende os efeitos induzidos pelo exercício físico. No entanto, o tipo de exercício físico ou a modalidade desportiva são igualmente preponderantes na avaliação do stress oxidativo em atletas ou desportistas.
Como na prática desportiva de escalada indoor ocorre o envolvi-mento muito particular de alguns músculos sobre outros, procedeu-se a um estudo comparativo, mas com a envolvência dinâmica e global de todo o corpo, recorrendo a um protocolo de corrida em tapete rolante (Magalhães e col, 2007). Para isso, os mesmos atletas que participaram na modalidade de escalada participaram, igualmente neste novo estudo seguindo um protocolo e uma diferente tipo de exercício físico, mas equivalente, em termos de em termos de intensidade de esforço (VO2 max) e de tempo (11,31 ± 3,22 min) ao exercício de escalada realizado. O protocolo de recolha de amostras biológicas e determinações laboratoriais foram idênticos às seguidas no exercício da escalada anterior.
Nas Figura 16 e Figura 17, estão representados os níveis médios plasmáticos de TAS, ácido úrico, e dos marcadores de lesão oxidativa, sobre os lípidos (MDA) e sobre as proteínas (SH), bem como, dos níveis plas-máticos da glutationa total e da sua distribuição quantitativapelas suas distintas formas, respetivamente, nos diferentesmomentos em estudo.
O aumento do MDA reflete o aumento dos processos de lipo-peroxidação, como resultado da ação dos radicais livres sobre os ácidos gordos polinsaturados da membrana. Do mesmo modo, essa capacidade oxidativa manifesta-se sobre as proteínas como se pode aduzir da diminuição dos grupos sulfidrilo (SH) plasmáticos.
Foram determinadas igualmente os níveis séricos de grupos carbonilo, um outro marcador da ação oxidativa sobre as proteínas. De acordo com o perfil obtido, os seus níveis aumentaram durante o exercício atingindo valores elevados imediatamente após exercício (0
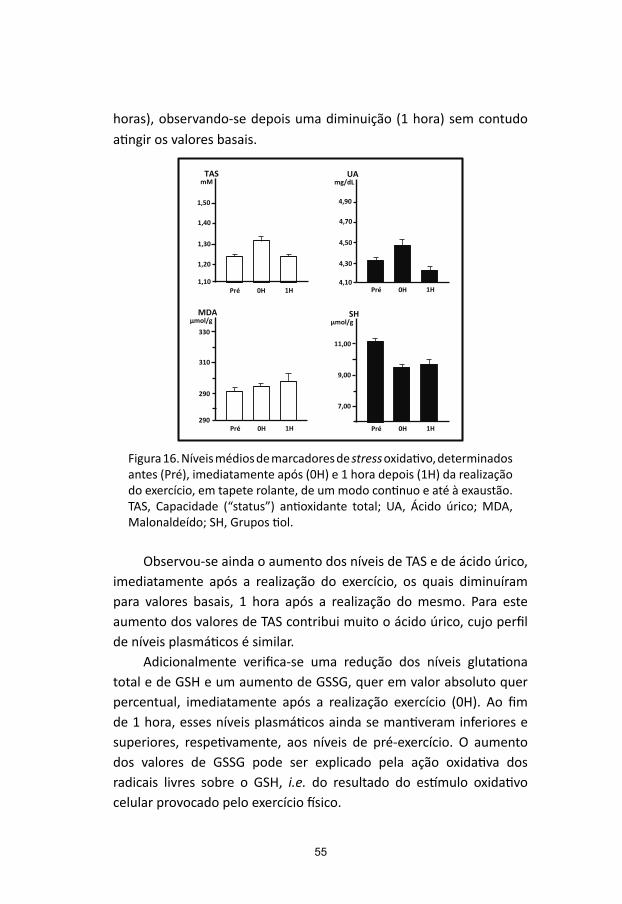
55
horas), observando-se depois uma diminuição (1 hora) sem contudo atingir os valores basais.
Figura 16. Níveis médios de marcadores de stress oxidativo, determinados antes (Pré), imediatamente após (0H) e 1 hora depois (1H) da realização do exercício, em tapete rolante, de um modo continuo e até à exaustão. TAS, Capacidade (“status”) antioxidante total; UA, Ácido úrico; MDA, Malonaldeído; SH, Grupos tiol.
Observou-se ainda o aumento dos níveis de TAS e de ácido úrico, imediatamente após a realização do exercício, os quais diminuíram para valores basais, 1 hora após a realização do mesmo. Para este aumento dos valores de TAS contribui muito o ácido úrico, cujo perfil de níveis plasmáticos é similar.
Adicionalmente verifica-se uma redução dos níveis glutationa total e de GSH e um aumento de GSSG, quer em valor absoluto quer percentual, imediatamente após a realização exercício (0H). Ao fim de 1 hora, esses níveis plasmáticos ainda se mantiveram inferiores e superiores, respetivamente, aos níveis de pré-exercício. O aumento dos valores de GSSG pode ser explicado pela ação oxidativa dos radicais livres sobre o GSH, i.e. do resultado do estímulo oxidativo celular provocado pelo exercício físico.
Fig. 16. Níveis médios de marcadores de stress oxidativo, determinados antes (Pré), imediatamente após (0H) e 1 hora depois (1H) da realização do exercício, em tapete rolante, de um modo continuo e até à exaustão. TAS, Capacidade (“status”) antioxidante total ; UA, Ácido úrico; MDA, Malonaldeído ; SH, Grupos tiol. Fig.16
SH μmol/g
11,00
9,00
7,00
Pré 0H 1H
290
MDA μmol/g
330
310
Pré 0H 1H
UA mg/dL
Pré 0H 1H
4,30
4,90
4,50
4,70
1,20
TAS mM
1,50
1,30
Pré 0H 1H
1,40
1,10 4,10
290
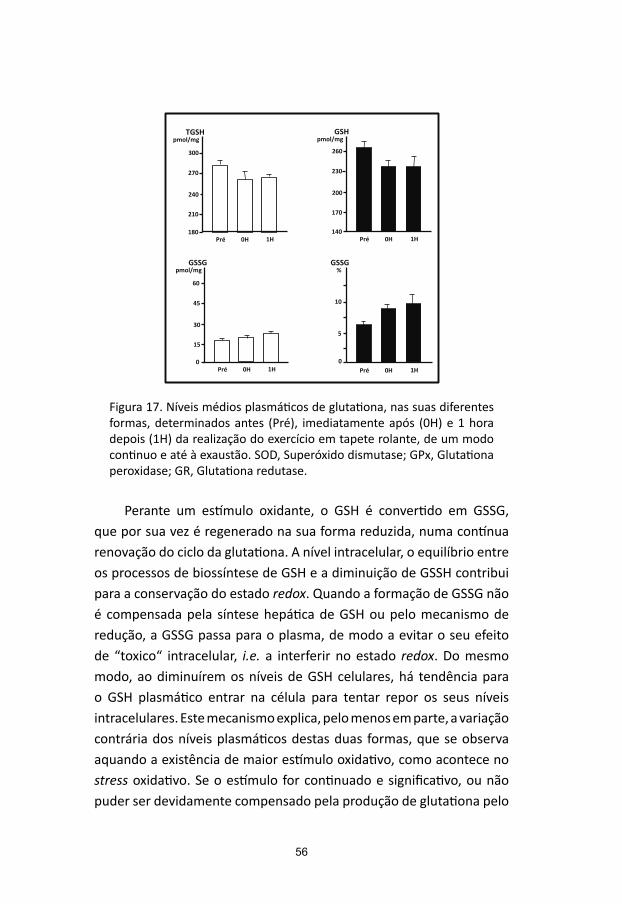
56
Figura 17. Níveis médios plasmáticos de glutationa, nas suas diferentes formas, determinados antes (Pré), imediatamente após (0H) e 1 hora depois (1H) da realização do exercício em tapete rolante, de um modo continuo e até à exaustão. SOD, Superóxido dismutase; GPx, Glutationa peroxidase; GR, Glutationa redutase.
Perante um estímulo oxidante, o GSH é convertido em GSSG, que por sua vez é regenerado na sua forma reduzida, numa contínua renovação do ciclo da glutationa. A nível intracelular, o equilíbrio entre os processos de biossíntese de GSH e a diminuição de GSSH contribui para a conservação do estado redox. Quando a formação de GSSG não é compensada pela síntese hepática de GSH ou pelo mecanismo de redução, a GSSG passa para o plasma, de modo a evitar o seu efeito de “toxico“ intracelular, i.e. a interferir no estado redox. Do mesmo modo, ao diminuírem os níveis de GSH celulares, há tendência para o GSH plasmático entrar na célula para tentar repor os seus níveis intracelulares. Este mecanismo explica, pelo menos em parte, a variação contrária dos níveis plasmáticos destas duas formas, que se observa aquando a existência de maior estímulo oxidativo, como acontece no stress oxidativo. Se o estímulo for continuado e significativo, ou não puder ser devidamente compensado pela produção de glutationa pelo
Fig. 17. Níveis médios plasmáticos de glutationa, nas suas diferentes formas, determinados antes (Pré), imediatamente após (0H) e 1 hora depois (1H) da realização do exercício em tapete rolante, de um modo continuo e até à exaustão. SOD, Superóxido dismutase; GPx, Glutationa peroxidase ; GR, Glutationa redutase.
Fig.17
TGSH pmol/mg
210
270
300
240
Pré 0H 1H
GSSG pmol/mg
15
45
60
30
Pré 0H 1H
GSSG %
5
10
Pré 0H 1H
GSH
170
pmol/mg
230
260
200
Pré 0H 1H 180 140
0 0

57
tecido hepático, ocorre a diminuição da Glutationa total com depleção dos seus pools intracelulares e maior dano oxidativo (Sies, 1985; Sies, 1999; Tapiero e Tew, 2003).
Deste modo, se se compararem os dois tipos de exercício realiz-ado, ambos até à exaustão e com níveis de intensidade idênticos, pode-se concluir que de acordo com a análise global dos parâmetros analisados, estes dois tipos de exercício, embora diferentes, estão associados a um desequilíbrio entre a capacidade oxidativa e a resposta antioxidante por eles induzida, e associada ao stress oxidativo. Mais ainda, se pode concluir que, tendo como base os valores dos níveis plasmáticos dos marcadores estudados, os níveis de stress oxidativo parecem ser menos acentuados na prática de exercício da modalidade de tapete rolante que na modalidade de escalada indoor.
5.4.4. Motocross O motocross é uma modalidade desportiva muito exigente, sob o
ponto de vista físico e fisiológico, para os pilotos. Dadas as condições particulares necessárias à prática da modalidade, terrenos irregulares, com lama, com curvas apertadas e saltos, muitas vezes de altura superior a 5 metros, o envolvimento do piloto, particularmente a nível muscular, é muito intenso, no sentido de fisicamente contrabalançar os choques com os joelhos, braços, pescoço e tronco. O esforço muscular dos membros superiores é muito acentuado dadas as inúmeras contrações isométricas e/ou excêntricas que sofrem para absorver esses impactos, durante a condução. Todas estas condições e características são sugestivas de envolvimento e de indução de stress oxidativo.
No estudo do stress oxidativo induzido por esta modalidade desportiva, 10 pilotos da elite de pilotos de motocross com experiencia significativa em provas internacionais (idade, 28,3 ± 7,9 anos; peso, 71,1 ± 7,0 Kg, altura, 169,8 ± 4,0 cm) realizaram uma prova competitiva num circuito de motocross de 1,5 Km. A recolha de material biológico foi efetuada antes, imediatamente após (tempo zero) e 1 hora após a realização da prova competitiva. Procedeu-se à determinação de TAS, ácido úrico, MDA e grupos SH, e glutationa, nas suas distintas formas,

58
como marcadores de stress oxidativo, assim como catecolaminas urinárias (urina 24 horas) horas) (Ascensão e col, 2007).
O valor percentual de GSSG (Figura 18), marcador sensível do stress oxidativo, aumenta no fim do período competitivo, em cerca de 25%, relativamente aos valores de basais.
Figura 18. Níveis médios da glutationa plasmática e suas distintas formas, de 10 participantes de uma prova competitiva de motocross, determinados antes (Pré), imediatamente após (0H) e 1 hora depois (1H) da sua realização. SOD, Superóxido dismutase; GPx, Glutationa peroxidase; GR, Glutationa redutase.
Estes valores elevados de GSSG podem ser explicados pelo facto de, durante a competição, a passagem de GSH do fígado para o plasma ter sido insuficiente para contrariar os níveis de espécies oxidantes plasmáticas, apesar de o GSH tecidular ter sido oxidado e compensada essa oxidação com a importação de mais GSH do plasma. No final, os níveis de GSH total diminuíram. Os níveis de GSSG plasmático podem assim ser o resultado aditivo da GSH que sofre um processo de oxidação no plasma, convertendo-se em GSSG, e da GSSG que foi exportada dos tecidos para o plasma.
Os perfis plasmáticos de MDA e de SH (Figura 19) e do grupo carbonilo (Figura 20) corroboram este efeito oxidativo exacerbado, resultante de um quadro de stress oxidativo induzido pelo exercício.
Figura 18. Níveis médios da glutationa plasmática e suas distintas formas, de 10 participantes de uma prova competitiva de motocross, determinados antes (Pré), imediatamente após (0H) e 1 hora depois (1H) da sua realização. SOD, Superóxido dismutase; GPx, Glutationa peroxidase; GR, Glutationa redutase.
Fig.18
TGSH pmol/mg
GSH
130
pmol/mg
190
220
160
150
210
240
180
Pré 0H 1H
GSSG pmol/mg
15
45
60
30
Pré 0H 1H
GSSG %
5
25
15
Pré 0H 1H
Pré 0H 1H 100 120
0 0
10
20

59
Do mesmo modo, pode observar-se, no final da competição, que os níveis de TAS plasmático foram superiores aos seus valores basais (Figura 19).
De facto, o aumento dos níveis plasmáticos de MDA traduz labo-ratorialmente o desenvolvimento do processo de lipoperoxidação, enquanto a diminuição dos grupos SH traduzem de maneira indireta a ação oxidativa sobre as proteínas.
Com o aumento de produção de radicais livres era de esperar que esses seus níveis diminuíssem, de modo a contrariar os inerentes efeitos oxidativos.
Figura 19. Níveis médios de marcadores de stress oxidativo, de 10 desportistas participantes de uma prova competitiva de motocross, determinados antes (Pré), imediatamente após (0H) e 1 hora depois (1H) da sua realização. TAS, Capacidade (“status”) antioxidante total; UA, Ácido úrico; MDA, Malonaldeído; SH, Grupos tiol.
No entanto, a justificação deste aumento pode ser encontrada no incremento do ácido úrico, cujo perfil de níveis plasmáticos o acompanha.
Para além do aumento do metabolismo das purinas, necessário para suprimir as exigentes necessidades energéticas associadas a esta modalidade desportiva, verificou-se ao longo da corrida de competição
Figura 19. Níveis médios de marcadores de stress oxidativo, de 10 participantes de uma prova competitiva de motocross, determinados antes (Pré), imediatamente após (0H) e 1 hora depois (1H) da sua realização. TAS, Capacidade (“status”) antioxidante total; UA, Ácido úrico; MDA, Malonaldeído ; SH, Grupos tiol.
Fig.19
SH μmol/g
6,00
4,00
2,00 Pré 0H 1H
300
MDA μmol/g
400
350
Pré 0H 1H
UA mg/dL
Pré 0H 1H
6,00
7,50
6,50
7,00
1,60
TAS mM
1,90
1,70
Pré 0H 1H
1,80
1,50 5,50
250
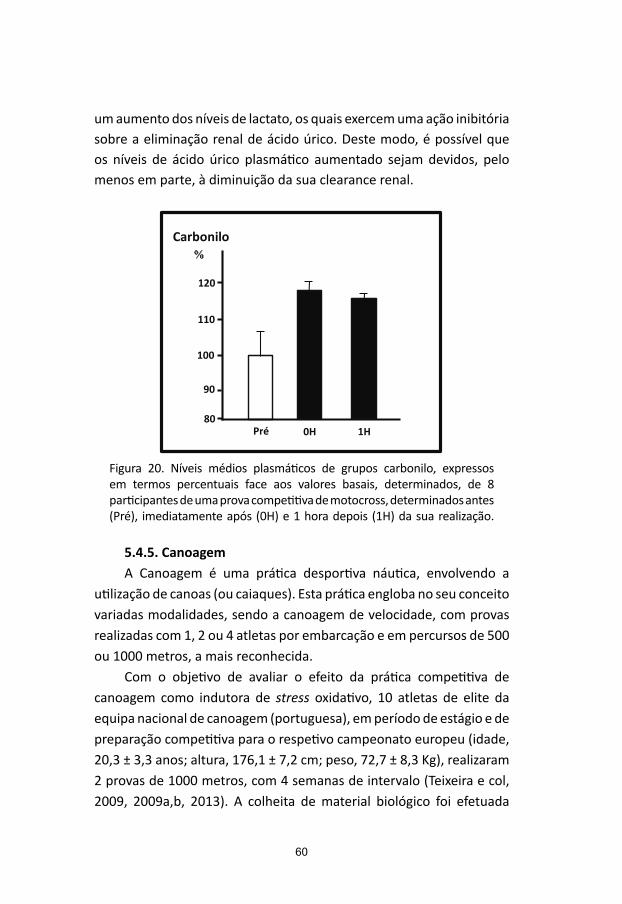
60
um aumento dos níveis de lactato, os quais exercem uma ação inibitória sobre a eliminação renal de ácido úrico. Deste modo, é possível que os níveis de ácido úrico plasmático aumentado sejam devidos, pelo menos em parte, à diminuição da sua clearance renal.
Figura 20. Níveis médios plasmáticos de grupos carbonilo, expressos em termos percentuais face aos valores basais, determinados, de 8 participantes de uma prova competitiva de motocross, determinados antes (Pré), imediatamente após (0H) e 1 hora depois (1H) da sua realização.
5.4.5. Canoagem A Canoagem é uma prática desportiva náutica, envolvendo a
utilização de canoas (ou caiaques). Esta prática engloba no seu conceito variadas modalidades, sendo a canoagem de velocidade, com provas realizadas com 1, 2 ou 4 atletas por embarcação e em percursos de 500 ou 1000 metros, a mais reconhecida.
Com o objetivo de avaliar o efeito da prática competitiva de canoagem como indutora de stress oxidativo, 10 atletas de elite da equipa nacional de canoagem (portuguesa), em período de estágio e de preparação competitiva para o respetivo campeonato europeu (idade, 20,3 ± 3,3 anos; altura, 176,1 ± 7,2 cm; peso, 72,7 ± 8,3 Kg), realizaram 2 provas de 1000 metros, com 4 semanas de intervalo (Teixeira e col, 2009, 2009a,b, 2013). A colheita de material biológico foi efetuada
Ascenção e col. Br J Sports Med., 41(2007)101-5
Figura. 20. Níveis médios plasmáticos de grupos carbonilo, expressos em termos percentuais face aos valores basais, determinados, de 8 participantes de uma prova competitiva de motocross, determinados antes (Pré), imediatamente após (0H) e 1 hora depois (1H) da sua realização.
Fig.20
Pré
90
110
120
Carbonilo %
100
0H 1H 80

61
antes e 15 minutos depois da realização de cada uma das duas provas executadas (P0, a primeira prova efetuada no início do período de estágio competitivo e P4, prova realizada 4 semanas depois).
Os resultados obtidos em cada uma das provas efetuadas, para cada um dos parâmetros utilizados (Figura 21), apresentam perfis muito semelhantes e refletem a instalação de um quadro de stress oxidativo induzido pelo exercício competitivo, traduzido no aumento dos valores de UA e TBARS e a diminuição de TAS.
Figura 21. Níveis médios plasmáticos de marcadores de stress oxidativo, determinados antes e 15 minutos depois da realização de duas provas de K1000, em período de estágio competitivo, a primeira realizada no início (P0) e a segunda (P4), realizada 4 semanas depois. TAS, Capacidade (“status”) antioxidante total; UA, Ácido úrico; TBARS, substâncias reativas ao ácido tiobarbitúrico.
O aumento dos TBARS traduz o efeito da ação oxidativa sobre os ácidos gordos insaturados (lipoperoxidação), como resultado do aumento da produção de radicais livres que não terá sido devidamente compensada pela quantidade de antioxidantes presentes no plasma, conforme se pode observar pela diminuição dos níveis de TAS. Os perfis dos níveis plasmáticos de ácido úrico traduzem o aumento do metabolismo das purinas para compensar as necessidades energéticas. Sendo o ácido úrico um dos compostos com propriedades antioxidantes englobadas na capacidade antioxidante total plasmática (TAS), a diminuição desta grandeza pode traduzir um aumento do consumo de outros dos seus constituintes antioxidantes, no sentido de contrariar a sobreprodução de radicais lives e de stress oxidativo.
Figura 21. Níveis médios plasmáticos de marcadores de stress oxidativo, determinados antes e 15 minutos depois da realização de duas provas de K1000, em período de estágio competitivo, a primeira realizada no inicio (P0) e a segunda (P4), realizada 4 semanas depois. TAS, Capacidade (“status”) antioxidante total ; UA, Ácido úrico; TBARS, substâncias reativas ao ácido tiobarbitúrico.
Fig.21
1,10
TAS mM
1,40
1,30
P0 P4
1,20 4,5
UA mg/dL
5,5
P0 P4
6,0
TBARS μM
10,0
8,0
P0 P4 1,00 3,5 4,0

62
5.4.6. AlpinismoO alpinismo é uma modalidade desportiva que exige por parte
dos seus praticantes uma elevada preparação em termos de condições físicas, psíquicas e fisiológicas.
As condições ambientais inerentes à altitude e, em particular, à alta altitude, são potencialmente adversas ao organismo humano, e têm como consequência uma grande variedade de respostas fisiológicas no sentido de obstar e contrariar essas mesmas adversidades. A redução da pressão de oxigénio, devido a condições hipobáricas (à medida que se sobe em altura, diminui a pressão atmosfera e a pressão parcial de oxigénio), reflete-se desde logo na diminuição da saturação arterial de oxigénio, originando situações de hipoxia celular e tecidular. Com a altitude verifica-se, assim, uma redução da disponibilidade da difusão de oxigénio do ar para o sangue através da membrana dos capilares alveolares, condição responsável pelo desenvolvimento de situações de hipoxia (Schoene, 2001; Askew, 2002). As consequências de altitude podem ser atribuídas, assim, às condições de insuficiência de oxigénio e à consequente diminuição de seus níveis no sangue arterial. Este tipo de hipoxia, resultante de uma baixa pressão, é designado de hipóxia hipobárica.
Paradoxalmente, apesar destas condições de deficiente disponibi-lidade de oxigénio, verifica-se, com a exposição à altitude, aguda ou crónica, um aumento da produção de espécies reativas de oxigénio e de azoto (ROS), de elevada capacidade deletéria sobre o organismo (Chao e col, 1999; Bailey e col, 2000; Moller e col, 2001; Magalhães e col 2004; Magalhães e col, 2004, 2005).
No sentido de estudar, pois, os efeitos do exercício físico em ambientes de hipoxia-hipobárica, 6 alpinistas foram acompanhados durante uma expedição de 4 semanas (1 semana de aclimatação, “trek” de 2800m a 5250m, e 3 semanas a altitude entre 5250m e pico) ao Pumori (7161m), nos Himalaias (Magalhães e col, 2005). A recolha de material biológico foi efetuada antes da partida e 1 dia após chegada da expedição (4 dias após ser atingido o cume do Pumori). Procedeu-se à determinação de SOD, GPx, GR eritrocitários e TBARS, TAS e grupos SH plasmáticos.
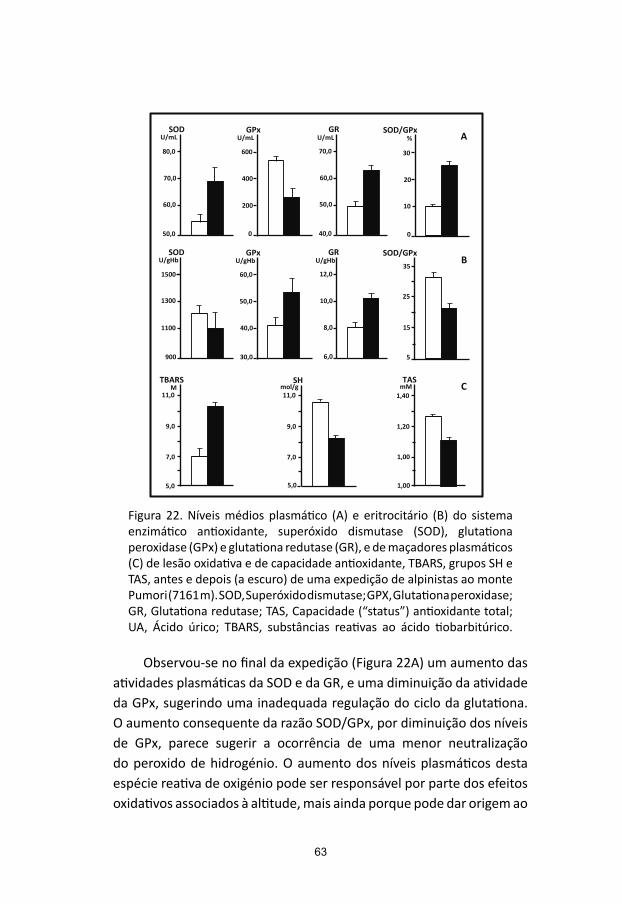
63
Figura 22. Níveis médios plasmático (A) e eritrocitário (B) do sistema enzimático antioxidante, superóxido dismutase (SOD), glutationa peroxidase (GPx) e glutationa redutase (GR), e de maçadores plasmáticos (C) de lesão oxidativa e de capacidade antioxidante, TBARS, grupos SH e TAS, antes e depois (a escuro) de uma expedição de alpinistas ao monte Pumori (7161 m). SOD, Superóxido dismutase; GPX, Glutationa peroxidase; GR, Glutationa redutase; TAS, Capacidade (“status”) antioxidante total; UA, Ácido úrico; TBARS, substâncias reativas ao ácido tiobarbitúrico.
Observou-se no final da expedição (Figura 22A) um aumento das atividades plasmáticas da SOD e da GR, e uma diminuição da atividade da GPx, sugerindo uma inadequada regulação do ciclo da glutationa. O aumento consequente da razão SOD/GPx, por diminuição dos níveis de GPx, parece sugerir a ocorrência de uma menor neutralização do peroxido de hidrogénio. O aumento dos níveis plasmáticos desta espécie reativa de oxigénio pode ser responsável por parte dos efeitos oxidativos associados à altitude, mais ainda porque pode dar origem ao
Fig.22
Figura 22. Níveis médios plasmático (A) e eritrocitário (B) do sistema enzimático antioxidante, superóxido dismutase (SOD), glutationa peroxidase (GPx) e glutationa redutase (GR), e de maçadores plasmáticos (C) de lesão oxidativa e de capacidade antioxidante, TBARS, grupos SH e TAS, antes e depois (a escuro) de uma expedição de alpinistas ao monte Pumori (7161 m). SOD, Superóxido dismutase; GPX, Glutationa peroxidase; GR, Glutationa redutase; TAS, Capacidade (“status”) antioxidante total; UA, Ácido úrico; TBARS, substâncias reativas ao ácido tiobarbitúrico.
SOD U/mL
GPx U/mL
80,0
50,0
70,0
60,0
600
0
400
200
GR U/mL
SOD/GPx
70,0
40,0
60,0
50,0
30
0
20
10
% A
SOD U/gHb
900
1300
1100
1500
GPx U/gHb
50,0
60,0
40,0
30,0
GR U/gHb
SOD/GPx
15
35
25
B
5
10,0
12,0
8,0
6,0
TBARS M
7,0
11,0
9,0
5,0
SH mol/g
TAS mM
1,00
1,40
1,20
C
7,0
11,0
9,0
5,0 1,00
CORRIGIDO

64
radical hidroxilo muito mais reativo e com maior capacidade deletéria que o peróxido de hidrogénio. O aumento dos níveis de atividade da SOD e da GR observado pode ser o resultado da ação oxidativa sobre as membranas provocando em consequência a lesão das mesmas e a sua passagem para o plasma, no que é corroborado pelo aumento dos níveis de TBARS verificado.
Por outro lado, a nível eritrocitário (Figura 22B), observa-se o aumento da atividade da GPx e da GR, i.e., das enzimas do ciclo da glutationa. Este mecanismo de regulação fisiológica contribui para a manutenção ou aumento da quantidade de glutationa reduzida intracelular disponível para compensar o aumento na produção de radicais livres. Este mecanismo constitui um sistema adaptativo e aumentado da capacidade de defesa antioxidante dessas células contra o stress oxidativo induzido pela altitude, conferindo e aumentando a capacidade de defesa antioxidante do glóbulo rubro (e do plasma), face a estas condições oxidativas adversas.
Acresce ainda que, a diminuição da razão SOD/GPx, por aumento de GPx, pode ser interpretado como mais um fator positivo na defesa do organismo face ao estímulo oxidativo.
Observou-se ainda (Figura 22C) um aumento nos níveis plasmáticos de substâncias reativas (TBARS) e diminuição dos grupos SH, indicando efeitos oxidativos sobre os lipídios das membranas (os ácidos gordos polinsaturados são sensíveis à oxidação pelas espécies de oxigénio reativas), e sobre em proteínas, com consequente aumento das ligações dissulfureto, respetivamente. A diminuição nos níveis plasmáticos de TAS corrobora a ação oxidativa sobre o organismo.
No entanto, e como dado suplementar, a altitude crónica parece induzir alterações significativas na composição qualitativa dos ácidos gordos constituintes da membrana dos eritrócitos. Os resultados por nós obtidos neste estudo apontam para um aumento dos ácidos gordos de polinsaturados, assim como, dos cis ácidos gordos. Mais ainda, observou-se uma diminuição quantitativa dos ácidos gordos trans, mantendo-se praticamente inalteráveis os níveis de ácidos gordos saturados. No seu conjunto, estes dados sugerem uma alteração fisiológica e adaptativa benéfica e de proteção face às condições

65
adversas associadas à altitude, em particular a hipóxia. De facto, esta modificação qualitativa, ao influenciar a fluidez da membrana eritrocitária (Subczynsky e Wisniewska, 2000; Halliwell e Gutteridge, 2015), poderá ser responsável por uma maior oxigenação dos tecidos, uma vez que contribuirá para uma melhor circulação dos eritrócitos pelos mais finos capilares, e a chegada do oxigénio às mais distintas partes do organismo.
Estes resultados observados em ambiente real foram igualmente encontrados em condições simuladas, corroborando a indução de situações de stress oxidativo pela altitude (Magalhães e col, 2004).
De facto, observou-se ao fim de uma exposição de 4 horas, a 5500m, em camara hipobárica, (Figura 23), a diminuição nos níveis plasmáticos de glutationa reduzida e o aumento significativo nos valores percentuais da glutationa oxidada relativamente a glutationa total, o aumento plasmático de TBARS e a diminuição dos grupos sulfidrilo (SH) e da concentração da capacidade total antioxidante (TAS).
Figura 23. Níveis médios plasmáticos de marcadores de stress oxidativo, imediatamente antes, durante (1H, 4H) e após o regresso a condições a nível do mar, de 6 voluntários, expostos durante 4 horas, a 5500m, em camara hipobárica. TBARS, substâncias reativas ao ácido tiobarbitúrico; SH, Grupos tiol; TAS, Capacidade (“status”) antioxidante total; GSH, Glutationa reduzida; GSSG, Glutationa oxidada.
Fig.23
Figura 23. Níveis médios plasmáticos de marcadores de stress oxidativo, imediatamente antes, durante (1H, 4H) e após o regresso a condições a nível do mar, de 6 voluntários (idade, 36,1 ± 2,0 anos; peso, 74,0 ± 2,3 kg; altura, 173,6 ± 1,6 cm), expostos , durante 4 horas, a 5500m, em camara hipobárica. ). TBARS, substâncias reativas ao ácido tiobarbitúrico; SH, Grupos tiol; TAS, Capacidade (“status”) antioxidante total; GSH, Glutationa reduzida; GSSG, Glutationa oxidada.
TBARS M
8,0
12,0
Antes 1H 4H Após
SH mol/g
7,0
10,0
9,0
Antes 1H 4H Após
TAS mM
1,05
1,15
1,10
Antes 1H 4H Após
GSH μmol/mg
120
200
160
Antes 1H 4H Após
GSSG %
15
45
30
Antes 1H 4H Após
6,0
10,0
6,0
8,0
1,00
1,20
0 80
CORRIGIDO

66
A análise global destes resultados permite concluir que a prática de exercício ou atividade física, não só na prática de alpinismo, mas também durante os processos de treino desportivo em altitude, quan-do realizada nestes ambientes de altitude e em particular a altitude ele-vada, está associada à indução de stress oxidativo, com todos os seus efeitos deletérios e de agressão orgânica sobre o organismo humano, in-dependentemente dos processos de aclimatação (Magalhães e col, 2004).
Em resposta às condições de baixos valores de pressão parcial de oxigénio inerentes à altitude, verificam-se variadas outras adaptações fisiológicas a diversos níveis, morfológico, hematológico e bioquímico, mas também um aumento “paradoxal” de espécies reativas de oxigénio, compostos que normalmente, são produzidos em ambientes ricos em oxigénio. O facto de se observar o estabelecimento de quadros de stress oxidativo nestes ambientes de hipoxia constitui o designado paradoxo da altitude.
Os mecanismos de resposta fisiológica a estas condições adversas, nomeadamente a nível da indução do sistema de defesa antioxidante, permitem minorar estes efeitos deletérios. Contudo, os efeitos oxidativos induzidos pela altitude com reflexos nesta modalidade desportiva devem ser analisados como de origem multifatorial, e não apenas atribuíveis às condições de hipoxia-hipobárica ou da prática do exercício físico, per se, uma vez que as condições ambientais inerentes às altas montanhas, como o frio, exposição às radiações UV solares, desidratação, etc., podem igualmente contribuir e potenciar os mais variados efeitos e ações oxidativas.
5.5. Suplementação de antioxidantesA prática de exercício físico é responsável pelo aumento da
produção de radicais livres, que pode determinar a instalação de quadros de stress oxidativo, se o sistema de compensação antioxidante do organismo não for suficiente para neutralizar ou minorar essa produção. Nestas situações, ocorre a manifestação potencial dos efeitos nefastos por ela causados, sendo de considerar danos celulares, de importância particular nos desportistas.

67
Esta produção ocorre durante e após a realização do exercício, sendo contudo mais acentuada durante a sua prática, e de acordo com a intensidade e duração do mesmo, devido a variados fatores, como o maior aumento do consumo de oxigénio, em particular, mas também, a uma maior atividade do metabolismo purínico, ao incremento das catecolaminas, aos processos inflamatórios como resposta à lesão muscular, entre outros.
A resposta do organismo a estes estímulos oxidantes traduz-se pelo aumento da sua capacidade de defesa antioxidante que nem é sempre suficiente, sob o ponto de vista quantitativo, para os compensar. Deste modo, a questão da suplementação com compostos antioxidantes, no sentido de melhor responder às alterações fisiológicas provocadas pela prática do exercício físico, coloca-se com toda a acuidade e premência, mais ainda e em particular, em situações de alta competição, onde o desempenho desportivo é fundamental.
Não existe consenso relativamente ao efeito do uso suplementar de antioxidantes, em particular de vitaminas, como forma de contrariar os efeitos menos positivos da prática do exercício físico ou como forma de favorecer a eficiência e o rendimento desportivo (Ji, 2002; Gross e Baum, 2015).
De facto, alguns autores encontraram efeitos protetores contra os efeitos deletérios causados aumento da produção de radicais livres, induzidos pelo exercício físico e pela prática desportiva competitiva, com a administração de suplementos vitamínicos ou de combinações de vitaminas (Mastaloudis e col, 2004; Williams, 2004; Zembron-Lacey e col, 2006; Davinson e col, 2007; Nazıroğlu e col, 2010; Bentley e col, 2015). Outros, porém, não encontraram quaisquer evidências ou vantagens na administração destes suplementos na redução dos efeitos causados destas situações potencialmente deletérias (Belko, 1987; Peake, 2003; Williams, 2004; Mastaloudis e col, 2006; Teixeira e col, 2009; Bailey e col, 2011; Theodorou e col, 2011). Na realidade, alguns estudos demonstram que a suplementação com antioxidantes pode mesmo ter efeitos contraproducentes, sugerindo que a suplementação com antioxidantes pode ser responsável por um efeito

68
pro-oxidativo (Nieman e col, 2004; Childs e col, 2001), bem como, por um atraso ou inadequação dos processos de adaptação fisiológica ao exercício físico, podendo interferir negativamente no desempenho e rendimento desportivo (Gomez-Cabrera e col, 2008; Peternelj e Coombes, 2011; Braakhuis, 2012; Nikolaidis e col, 2012; Makanae e col, 2013; Mendonça e col, 2014). Mais ainda, é possível que este impedimento ou dificuldade de adaptação do tecido muscular ao exercício seja o resultado, pelo menos em parte, da capacidade dos antioxidantes em bloquear os processos de sinalização, emanados pelos radicais livres, reguladores da expressão do sistema enzimático antioxidante (Gomez-Cabrera e col, 2006, 2008, 2008a).
Com o objetivo de avaliar o efeito da suplementação de antioxidantes na diminuição de stress oxidativo e da lesão muscular procedeu-se à realização de estudo (Teixeira e col, 2009, 2013) que envolveu um conjunto de atletas de elite pertencentes à equipa nacional de canoagem (portuguesa), em período de estágio e de preparação competitiva para o respetivo campeonato europeu.
Os atletas foram divididos em 2 grupos, cada um dos quais com 10 participantes. A um dos grupos (idade, 19,1 ± 3,8 anos; altura, 174,2 ± 9,7 cm; peso, 71,4 ± 9,1 Kg) foi administrado um suplemento de complexo de vitaminas antioxidantes e ao outro (idade, 20,3 ± 3,3 anos; altura, 176,1 ± 7,2 cm altura; peso, 72,7 ± 8,3 Kg) foi administrado placebo. A colheita de material biológico foi efetuada antes e 4 semanas depois da suplementação, em condições pré e pós-exercício físico realizado. Foram vários os parâmetros laboratoriais determinados neste estudo, entre eles, CPK (marcador de lesão muscular), TBARS (marcador da peroxidação lipídica), TAS, ácido úrico, IL-6 (inflamação) e cortisol.
Os resultados obtidos (Figura 24), permitiram concluir que, a administração de suplementos antioxidantes não parecem conferir proteção contra a peroxidação lipídica, nem contra a lesão muscular ou o processo inflamatório atribuído ao stress oxidativo induzido pelo exercício, pelo menos em atletas de elite altamente treinados.
De facto, em resposta ao exercício, observou-se um aumento significativo dos níveis de TBARS, ácido úrico e CPK em ambos os sub-
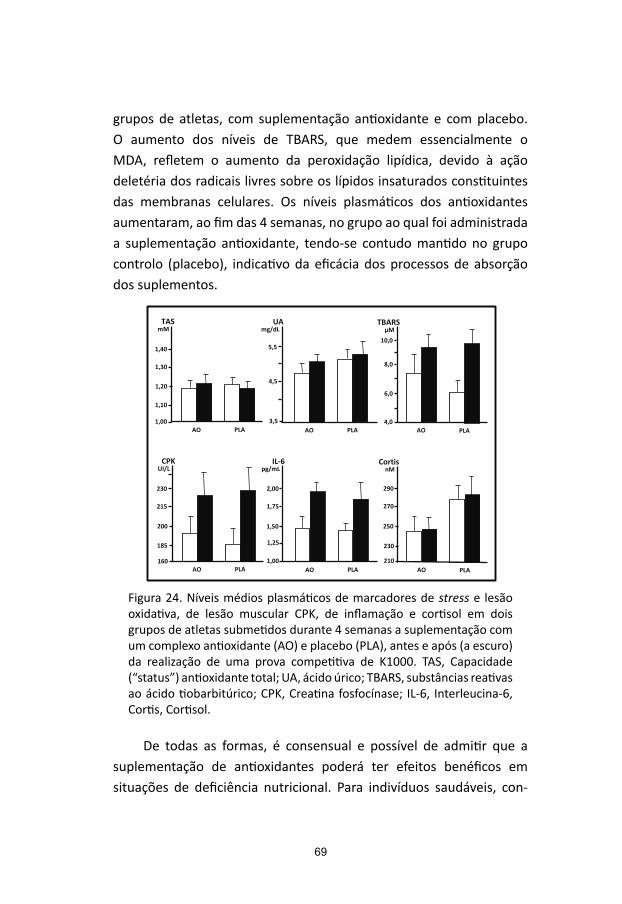
69
grupos de atletas, com suplementação antioxidante e com placebo. O aumento dos níveis de TBARS, que medem essencialmente o MDA, refletem o aumento da peroxidação lipídica, devido à ação deletéria dos radicais livres sobre os lípidos insaturados constituintes das membranas celulares. Os níveis plasmáticos dos antioxidantes aumentaram, ao fim das 4 semanas, no grupo ao qual foi administrada a suplementação antioxidante, tendo-se contudo mantido no grupo controlo (placebo), indicativo da eficácia dos processos de absorção dos suplementos.
Figura 24. Níveis médios plasmáticos de marcadores de stress e lesão oxidativa, de lesão muscular CPK, de inflamação e cortisol em dois grupos de atletas submetidos durante 4 semanas a suplementação com um complexo antioxidante (AO) e placebo (PLA), antes e após (a escuro) da realização de uma prova competitiva de K1000. TAS, Capacidade (“status”) antioxidante total; UA, ácido úrico; TBARS, substâncias reativas ao ácido tiobarbitúrico; CPK, Creatina fosfocínase; IL-6, Interleucina-6, Cortis, Cortisol.
De todas as formas, é consensual e possível de admitir que a suplementação de antioxidantes poderá ter efeitos benéficos em situações de deficiência nutricional. Para indivíduos saudáveis, con-
Fig.24
Figura 24. Níveis médios plasmáticos de marcadores de stress e lesão oxidativa , de lesão muscular CPK , de inflamação e cortisol em dois grupos de atletas submetidos durante 4 semanas a suplementação com um complexo antiopxidante (AO) e placebo (PLA), antes e após (a escuro) da realização de uma prova competitiva de K1000. TAS, Capacidade (“status”) antioxidante total; UA, ácido úrico; TBARS, substâncias reativas ao ácido tiobarbitúrico; CPK, Creatina fosfocínase; IL-6, Interleucina 6, Cortis, Cortisol.
1,10
TAS mM
1,40
1,30
AO PLA
1,20 4,5
UA mg/dL
5,5
AO PLA
6,0
TBARS μM
10,0
8,0
AO PLA
185
CPK UI/L
230
215
AO PLA
200
IL-6 pg/mL
AO PLA
Cortis nM
AO PLA
1,25
2,00
1,75
1,50
230
290
270
250
1,00 3,5 4,0
160 1,00 210
CORRIGIDO

tudo, a suplementação pode ser ineficaz ou mesmo constituir uma des-vantagem, uma vez que altas doses de algumas vitaminas podem apresentar risco de toxidade.
A análise global dos estudos de avaliação do stress oxidativo realizados em atletas de elite, de distintas modalidades desportivas, e de acordo com os resultados globais obtidos com a determinação laboratorial de marcadores de lesão e de defesa antioxidante, permite concluir que o exercício físico associado ao desporto de alta competição induz alterações significativas no estado redox, nomeadamente, por ser uma fonte considerável de produção de espécies oxidantes.
No entanto, através de alterações adaptativas de natureza fi-siológica, para as quais contribuem os processos continuados de treino e de prática desportiva, o organismo é capaz de contrabalançar esses efeitos, minorando os potenciais efeitos deletérios ao exercício físico associados.

71
6. CONCLUSÃO
Pode de uma maneira geral afirmar-se que todo o organismo é beneficiado pela prática de uma atividade ou de exercício físico, embora em certas circunstâncias esses benefícios possam ser reduzidos ou mesmo questionados.
De facto, qualquer prática de exercício extemporâneo ou de exercício agudo pode constituir um fator nocivo e potencial causador de muitos dos problemas de saúde física, pelas alterações fisiológicas abruptas por ele induzido. No entanto, é consensualmente aceite que o exercido praticado com regularidade, mesmo em condições de intensidade moderada ou leve, cria condições adaptativas fisiológicas muito benéficas para o organismo humano. Por cada esforço físico adicional, por cada aumento da intensidade ou duração do exercício físico realizado, maior é a resposta por parte do organismo. Perante um novo estímulo potencialmente deletério o organismo adquire uma maior capacidade de resposta no sentido de contrariar ou mesmo de anular esse estímulo.
A prática de exercício físico o desportiva é um indutor de quadros de stress oxidativo mais ou menos acentuados, que podem ser traduzidos pela identificação e avaliação de diversos marcadores relacionados, como sejam os marcadores de lesão oxidativa ou de defesa antioxidante. O incremento da atividade das enzimas que fazem parte do sistema de defesa enzimático antioxidante, pode por outro lado constituir um bom exemplo da adaptação fisiológica ao stress oxidativo induzido pelo exercício. Esta capacidade de defesa antioxidante faz com que, em atletas de alta competição e de elite sujeitos a situações de elevado potencial deletério, os efeitos lesivos expectáveis sejam minorados, ou mais prontamente compensados, minimizando os danos prolongados no tempo (Reichhold e col, 2009; Wagner e col, 2010; Wagner e col, 2011; Tong e col, 2012).
Do mesmo modo, é a regularidade da prática de exercício e do treino, de intensidade variável e adequada aos fins pretendidos, que faz com que praticantes ou desportistas treinados, mesmo atletas

72
de elite, estejam mais bem preparados sob o ponto de vista de flexibilidade e reforço muscular, no sentido de uma melhor resposta a um estímulo ou situação virtualmente lesiva. Os níveis plasmáticos basais mais elevados de marcadores laboratoriais utilizados na avaliação da função muscular, observados em atletas treinados por comparação com sedentários ou atletas menos treinados, sugerem a maior amplitude de adaptação muscular ao exercício físico e uma melhor defesa face ao desgaste por ele induzido. O facto de atletas de elite de provas de endurance (triatlo) apresentaram um menor aumento de valores plasmáticos de marcadores musculares quando comparados com outros atletas menos treinados (não de elite), após realização das respetivas provas competitivas, reforça o conceito de uma melhor adaptação ao esforço físico.
A natureza e a manifestação destes efeitos são independentes da idade do praticante, sendo perfeitamente estendíveis a todas as idades. Aliás, no âmbito da investigação científica relacionada com o desporto, o nosso grupo tem expandido o seu trabalho ao estudo dos efeitos da atividade ou da prática de exercício físico na terceira idade.
O envelhecimento populacional constitui um problema merecedor de um seguimento muito particular nos dias de hoje, pelas implicações sociais, económicas e de saúde a ele associadas. Os custos relacionados com o envelhecimento, e em particular os respeitantes aos gastos em saúde, são hoje consideráveis e tendem-se necessariamente a agravar nos próximos anos, bastando para tal considerar os dados sociodemográficos publicados em 2015 (The Ageing Report, 2015). Dada esta tomada de consciência, a União Europeia consagra, dentro das suas competências, um importante apoio à investigação dirigida ao envelhecimento.
A prática de exercício físico faz parte de uma forma de viver o envelhecimento saudável. A experiencia adquirida, a nível do acompanhamento com fins de estudo e de avaliação científica cientifico de uma população constituída por uma centena de homens e mulheres, com idades compreendidas entre os 65 anos e os 90 anos, mostra que a pratica regular de exercício físico melhora significativamente

73
a sociabilidade entre, previne ou atrasa o aparecimento e/ou o desenvolvimento de quadros patológicos, nomeadamente a nível das doenças cardiovasculares e do foro neurológico e psiquiátrico (Carvalho e col, 2010).
A citação latina Mens sana in corpore sano faz todo o sentido nos dias de hoje, onde o atropelar dos dias ocupados inibe ou dificulta, muitas vezes, o crescer e o viver em plenitude. A saúde mental condiciona a saúde física. Situações como o stress, a depressão a ansiedade constituem fatores condicionantes da saúde física que podem ser minorados com a prática regular do exercício físico. Vale a pena a prática de exercício físico.


75
7. REFERÊNCIAS BIBLIOGRÁFICAS
Alderton WK, Cooper CE, Knowles RG. Nitric oxide synthases: structure,
function and inhibition. Biochem J 357 (2001) 593-615
Armstrong RB. Mechanisms of exercise-induced delayed onset muscular
soreness: a brief review. Med Sci Sports Exerc 16 (1984)529-553)
Ascensão A, Ferreira R, Marques F, Oliveira E, Azevedo V, Soares J, Magalhaes
J. Effect of off-road competitive motocross race on plasma stress and
damage markers. Br J Sports Med 41 (2007) 101-105
Ascensão A, Rebelo A, Oliveira E, Marques F, Pereira L, Magalhães J.
Biochemical impact of a soccer match — analysis of oxidative stress and
muscle damage markers throughout recovery Clinical Biochemistry 41
(2008) 841–851
Askew EW. Work at high altitude and oxidative stress: antioxidant nutrients.
Toxicology 180 (2002) 107-119
Azizbeigia K, Stannardb SR, Atashakc S, Haghighid MM. Antioxidant enzymes
and oxidative stress adaptation to exercise training: Comparison of
endurance, resistance, and concurrent training in untrained males. J
Exerc Sci Fitness 12 (2014) 1-6
Bailey DM, Davies B, Baker J. Training in hypoxia: modulation of metabolic
and cardiovascular risk factors in men. Med Sci Sports Exerc 32 (2000)
1058-1066
Bailey DM, Williams C, Betts JA, Thompson D, Hurst TL. Oxidative stress,
inflammation and recovery of muscle function after damaging exercise:
effect of 6-week mixed antioxidant supplementation. Eur J Appl Physiol
111 (2001) 925-936
Baillie JK, Bates MGD, Thompson AAR, Waring WS, Partridge RW, Schnopp MF,
Simpson A, Gulliver-Sloan F, Maxwell SRJ. Endogenous Urate Production
Augments Plasma Antioxidant Capacity in Healthy Lowland Subjects
Exposed to High Altitude. Chest 131 (2007) 1473–1478
Bake JS, McCormick MC, Robergs RA. Interaction among Skeletal Muscle
Metabolic energy Systems during Intense Exercise. J Nutr Metabol 2010
(2010) 905612

76
Banfi G, Dolci A. Free testosterone/cortisol ratio in soccer: usefulness of a
categorization of values. J Sports Med Phys Fitness 46 (2006) 611-616
Barbosa KB, Costa NM, Alfenas RC, Paula DO, Minim VP, Bressan J. Estresse
oxidativo: conceito, implicações e fatores modulatórios Rev Nutr 23
(2010) 629-643
Baron CP, Andersen HJ. Myoglobin-induced lipid oxidation. A review. J Agric
Food Chem 50 (2002) 3887-3897
Barrera G. Oxidative Stress and Lipid Peroxidation Products in Cancer
Progression and Therapy. Oncology 2012 (2012) 1-21
Belko AZ. Vitamins and exercise - an update. Med Sci Sports Exerc 19 (1987)
S191-186
Bentley DJ, Ackerman J, Clifford T, Slattery KS. Acute and Chronic Effects of
Antioxidant Supplementation on Exercise Performance. In Antioxidants
in Sport Nutrition. Cap. 9. Ed. Lamprecht M. Boca Raton (FL): CRC Press/
Taylor & Francis. 2015
Bloomer RJ, Goldfarb AH. Anaerobic exercise and oxidative stress: a review.
Can J Appl Physiol 29 (2004) 245-63
Braakhuis G, Hopkins WG. Impact of dietary antioxidantes on sport
performance: a Review. Sports Med 45 (2015) 939-55
Braakhuis AJ. Effect of vitamin C supplements on physical performance. Curr
Sports Med Rep. 11 (2012) 180-184
Brancaccio P, Maffulli N, Limongelli, FM. Creatine Kinase monitoring in sport
medicine. Br. Med Bull 81 (2007) 209-230
Butterfield TA, Best TM, Merrick MA. The Dual Roles of Neutrophils and
Macrophages in Inflammation: A Critical Balance Between Tissue
Damage and Repair. J Athletic Training 41 (2006) 457-465.
Calvo G, Lopez G. Ejercicio físico y radicales libres, ¿es necesaria una
suplementación con antioxidantes? Rev Int Med Cienc Act Fis Deporte
12 (2011) 69-388
Carvalho J, Marques E, Ascensao A, Magalhaes J, Marques F, Mota J.
Multicomponent exercise program improves blood lipid profile and
antioxidant capacity in older women. Arch Gerontol and Geriatrics 51
(2010) 1-5
Castrogiovanni P, Imbesi R. Oxidative stress and skeletal muscle in exercise.
Ital J Anat Embryol 11 (2012) 107-117

77
Chevion S, Moran D, Heled Y, Shani Y, Regev G, Abbou B, Berenshtein E,
Stadtman E, Epstein Y. Plasma antioxidant status and cell injury after
severe physical exercise. Proc Natl Acad Sci USA 100 (2003) 5119-5123
Cheung K, Hume PA, Maxwell L. Delayed onset muscle soreness. Sports Med
33 (2003) 145-164
Childs A, Jacobs C, Kaminski C, Halliwell B, Leeuwenburg C. Supplementation
with vitamin C and N-acetyl-cystein increases oxidative stress in humans
after an acute muscle injury by eccentric exercise. Free Radic Biol Med
31 (2001) 745-753
Chung HY, Baek BS, Song SH, Kim MS, Huh JI, Shim KH, Kim KW. Xanthine
dehydrogenase/xanthine oxidase and oxidative stress. Age 20 (1997)
127–140
Cioca DP, Watanabe N, Isobe M. Apoptosis of peripherical blood lymphocytes
is induced by catecholamines. Jpn Heart J 41 (2000) 385-398
Cooper CE, Vollaard NB, Choeiri T, Wilsom MT. Exercise free radicals and
oxidative stress. Biochm Soc Trans 30 (2002) 280-285
Crewther B, Cronin J, Keogh J. Possible stimuli for strength and power
adaptation: acute metabolic responses. Sports Med 36 (2008) 65-78
Davinson G, Gleeson M, Philips S. Antioxidant supplementation and
immunoendocrine responses to prolonged exercise. Med Sci Sports
Exerc 39 (2007) 645-652
Dröge W. Free radicals in the physiological control of cell function. Physiol Rev
282 (2002) 47-95
Duarte JA, Appell HJ, Carvalho F, Bastos ML, Soares JM. Endothelium-derived
oxidative stress may contribute to exercise-induced muscle damage. Int
J Sports Med 14 (1993) 440-443
Duarte JA, Magalhaes JF, Monteiro L, Almeida-Dias A, Soares JM, Apell HJ.
Exercise-induced signs of muscle oversue in children. Int J Sports Med 2
(1999) 103-108
Duarte JA, Carvalho F, Remiao F, Bastos ML, Magalhaes J, Apell H.
D-Ampohetamine-induced hydrogen peroxide production in skeletal
muscles modulated by mono amine oxidase inhibition. Int J Sports Med
25 (2004) 446-449
Ferreira F, Ferreira R, Duarte JA. Stress oxidativo e dano oxidativo muscular
esquelético: influência do exercício agudo inabitual e do treino físico.
Rev Port Cien Desp 7 (2007) 257–275

78
Filaire E, Lac G, Pequignot JM. Biochemical, hormonal, and psychological
parameters in professional soccer players throughout a competitive
season. Percept Mot Skills 97 (2003) 1061-1072
Finaud J, Lac G, Filaire, E. Oxidative stress: relationship with Exercise end
training. Sports Med 36 (2006) 327-358
Finaud J, Lac G, Filaire E. Oxidative stress: relationship with exercise and
training. J Sports Sci 30 (2012) 327-358
Finley JW, Kong AN, Hintze KJ, Jeffery EH, Ji LL, Lei XG. Antioxidants in foods:
state of the science important to the food industry. J Agric Food Chem
59 (2011) 6837-6846
Fisher-Wellman K, Bloomer RJ. Acute exercise and oxidative stress: a 30 year
history. Dyn Med 8 (2009)1-25
Friden J. Delayed onset muscle soreness. In: Schmidt RF, Gebhart GF, editors.
Encyclopedia of pain. 2nd ed. New York: Springer, 2013; p. 874-877
Fridovich I. Superoxide radicals, superoxide dismutases and the aerobic
lifestyle. Photochem Photobiol 4-5 (1978) 733-41
Gandra PG, Macedo DV, Alves AA. Fontes de espécies reativas de oxigénio na
musculatura esquelética durante o exercício. Br Ensino Bioq Biol Mol 2
(2006) C1-C11
George Lee. Exercise-Induced Rhabdomyolysis. Rhode Island Med J Sports
Med 97 (2014) 22-24
Gleeson M. Immune function in sport and exercise. J Appl Physiol103 (2007)
693-699
Gleeson M, Almey J, Brooks S, Cave R, Lewis A, Griffiths H. Haematological and
acute-phase responses associated with delayed-onset muscle soreness
in humans. Eur J Appl Physiol Occup Physiol 71(1995) 137-142
Gomez-Cabrera MC, Martínez A, Santangelo G, Pallardó FV, Sastre J, Viña
J. Oxidative stress in marathon runners: interest of antioxidant
supplementation. Br J Nutr 96 (2006) S31-33
Gomez-Cabrera MC, Domenech E, Romagnoli M, Arduini A, Borras C, Pallardo
FV, Sastre J, Viña J. Oral administration of vitamin C decreases muscle
mitochondrial biogenesis and hampers training-induced adaptations in
endurance performance Am J Clin Nutr 87 (2008) 142-149
Gomez-Cabrera MC, Domenech E, Viña J. Moderate exercise is an antioxidant:
upregulation of antioxidant genes by training. Free Radic Biol Med 44
(2008a) 126-131

79
Gorostiaga EM, Izquierdo M, Ruesta M, Iribarren J, Gonzalez-Badillo JJ, Ibanez
J. Strengh training effects on physical performance and serum hormones
in young soccer players during a competition half-season. Eur J Appl
Physiol 91 (2004) 698-707
Gospodaryov D, Lushchak V. Oxidative Stress: Cause and Consequence of
Diseases, in Oxidative Stress and Diseases. Cap 2, pp 13-38. Ed. by Dr.
Volodymyr Lushchak. ISBN 978-953-51-0552-7 April, 2012
Greig MP, McNaughton LR, Lowell RJ. Physiological and mechanical response
to soccer-specific intermittent activity and steady-state activity. Res
Sports Med 14 (2006) 29-52
Gross M, Baum O. Supplemental Antioxidants and Adaptation to Physical
Training. Ch 7. In Antioxidants in Sport Nutrition. Lamprecht M, ed. Boca
Raton (FL): CRC Press/Taylor & Francis, 2015.
Gunther MR, Sampath V, Caughey MS. Potential role of myoglobin autoxidation
in myocardial isquemia-reperfusion injury. Free Radicals Biol Med 26
(1999) 1238-1295
Halliwell B. Free radicals and antioxidants: updating a personal view. Nutr Ver
70 (2012) 257-265
Halliwell B, Gutteridge JMC. Free Radical in Biology Medicine University Press,
4.ed. Oxford, Oxford, UK: Clarendon Press, 2007
Halliwell B, Gutteridge JMC. Antioxidants: molecules, medicines, and myths.
Biochem Biophys Res Commun 393 (2010) 561-564
Halliwell B, Gutteridge JMC. Free Radical in Biology Medicine University Press,
5.ed. Oxford, Oxford, UK: Clarendon Press, 2015
Hellsten Y, Tullson PC, Richter EA, Bangsbo J. Oxidation of urate in human
skeletal muscle during exercise. Free Radic Biol Med 22 (1997) 169-74
Hellesten Y. The role of xantina oxidase in exercise. In: Sen C, Packer L, &
Hanninen O (Eds.) Handbook of Oxidants and Antioxidants in Exercise.
Basel: Elsevier Science. pp. 153-176. 2000
Hooper DC, Scott GS, Zborek A, Mikheeva T, Kean RB, Koprowski H, Spitsin SV.
Uric acid, a peroxynitrite scavenger, inhibits CNS inflammation, blood-
CNS barrier permeability changes, and tissue damage in a mouse model
of multiple sclerosis. FASEB J 14 (2000) 691-698
Ishii H, Stanimirovic DB, Chang CJ, Mrsulja BB, Spatz M. Dopamine metabolism
and free-radical related mitochondrial injury during transient brain
ischemia in gerbils. Neurochem Res 18 (1993) 1193-1201

80
Ji LL. Antioxidants and oxidative stress in exercise. Proc. Soc. Exp Biol Med 222
(1999) 283-292
Ji LL. Exercise-induced modulation of antioxidante defense. Ann NY Acad Sci
959 (2002) 82-92
Kanda K, Sugama K, Hayashida H, et al. Eccentric exercise-induced delayed
on set muscle soreness and changes in markers of muscle damage and
inflammation. Exerc Immunol Rev 19 (2013) 72-78
Kima J, Leea J, Kimb S, Ryuc HY, Chac KS, Sungc DJ. Exercise-induced
rhabdomyolysis mechanisms and prevention: A literature review. J Sport
Health Sci 3 (2016) 324-333
Kliszczewicz B, Quindry CJ, Blessing LD, Oliver DG, Esco RM, Taylor JK. Acute
Exercise and Oxidative Stress: CrossFit(™) vs. Treadmill Bout. J Hum
Kinet 47 (2015) 81-90
Knez WL, Coombes JS, Jenkins DG. Ultra-endurance exercise and oxidative
damage: implications for cardiovascular health. Sports Med 36 (2006)
429-441
Kenney W, Larry, JW, Costill D. “Physiology of Sport and Exercise 5E: Delayed-
Onset Muscle Soreness.” Human-kinetics. N.p., 2015. Web. 18 Feb. 2016
König D, Neubauer O, Nics L. Biomarkers of exercise-induced myocardial stress
in relation to inflammatory and oxidative stress. Exerc Immunol Rev 13
(2007) 15-36
Kraemer WJ, French DN, Paxton NJ, Hakkinen K, Volek JS, Sebastienelli WJ.
Changes in exercise performance and hormonal concentrations over a
big ten soccer season in starters and nonstarters. J Strengh Cond Res 18
(2004) 121-128
Kreher JB, Schwartz JB. Overtraining syndrome: a practical guide. Sports
Health 4 (2012) 128-138
Krüger K, Mooren FC.Exercise-induced leukocyte apoptosis. Exerc Immunol
Rev 20 (2014)117-134
Kudin AP, Debska-Vielhaber G, Kunz WS. Characterization of superoxide
production sites in isolated rat brain and skeletal muscle mitochondria,
Biomed Pharmacoter 59 (2005) 163-168
Lambeth JD, Neish AS. “Regulation of Nox and Duox enzymatic activity and
expression”. Annual Review of Patholog: Mechanisms of Disease 9
(2014) 119
Leeder JD, van Someren KA, Gaze D, et al. Recovery and adaptation from

81
repeated intermittent-sprint exercise. Int J Sports Physiol Perform 9
(2014) 489-496
Leeuwenburgh C, Heinecke JW. Oxidative Stress and Antioxidants in Exercise.
Curr Med Chem 8 (2001) 829-838
Liochev SI. Reactive oxygen species and the free radicals theory of aging. Free
Radic Biol Med 60 (2010) 1-4
MacKinnon LT, Hooper S. Mucosal (secretory) immune system responses to
exercise of varying intensity and during overtraining. Int J Sports Med
15 (1994) S179–S183
Magalhaes J, Ascensao A, Viscor G, Soares J, Oliveira J, Marques F, Duarte J.
(2004a) Oxidative stress in humans during and after 4 hours of hypoxia
at simulated altitude of 5500m Aviat Space Environ Med 75 (2004) 16-22
Magalhaes J, Ascensao A, Marques F, Soares J, Ferreira R, Neuparth MJ,
Duarte J. (2005) Effect of a high-altitude expedition to a Himalayan peak
(Pumori, 7161m) on plasma and erythrocyte antioxidant profile. Eur J
Appl Physiol 93 (2005) 726-732
Magalhaes J, Ascensao A, Marques F, Soares J, Neuparth MJ, Ferreira R, Amado,
F, Duarte J. Skeletal muscle ultrastructural and plasma biochemical
signs of endothelium dysfunction induced by a high-altitude expedition
(Pumori, 7161m) Basic Appl Myol 15 (2005a) 1-7
Magalhães J, Ferreira R, Marques F, Oliveira E, Soares J, Ascensão A. Indoor clim-
bing elicits plasma oxidative stress. Med Sci Sports Exerc 39 (2007) 955-63
Magalhaes J, Rebelo A, Oliveira E, Silva JR, Marques F, Ascensao, A. Impact
of Loughborough Intermittent Shuttle Test versus soccer match on
physiological, biochemical and neuromuscular parameters. Eur J Appl
Physiol 108 (2010) 39-48
Magder S. Reactive oxygen species: toxic molecules or spark of life? Critical
Care 10 (2006) 208
Majerczak J, Rychlik B, Grzelak A, Grzmil P, Karasinski J, Pierzchalski P, Pulaski
L, Bartosz G, Zoladz JA. Effect of 5-week moderate intensity endurance
training on the oxidative stress, muscle specific uncoupling protein
(UCP3) and superoxide dismutase (SOD2) contents in vastus lateralis of
young, healthy men. J Physiol Pharmacol 61 (2010) 743-751
Makanae Y, Kawada S, Sasaki K, Nakazato K, Ishii N. Vitamin C administration
attenuates overload-induced skeletal muscle hypertrophy in rats. Acta
Physiol (Oxf) 208 (2013) 57-65

82
Martins, FSB. Alterações bioquímicas induzidas por diferentes tipos de provas
de triatlo. Dissertação de Doutoramento em Ciências do Desporto,
FADEUP. 2010
Mastaloudis A, Marrow JD, Hopkins DW, Devaraj S, Traber MG. Antioxidant
supplementation prevents exercise-induced lipid peroxidation but not
inflammation in ultramarathon runners. Free Radic Biol Med 36 (2004)
1329-1341
Mastaloudis A, Traber MG, Carstensen K, Widrick J. Antioxidants did not
prevent muscle damage in response to an ultramarathon runners. Med
Sci Sports Exerc 38 (2006) 72-80
Maxwell SRJ, Thomason H, Sandler D, Leguen C, Baxter MA, Thorpe GHG,
Jones AF, Barnett AH. “Antioxidant status in patients with uncomplicated
insulin-dependent and non-insulin-dependent diabetes mellitus”. Eur J
Clin Inv 27 (1997) 484-490
McArdle A, Pattwell A, Valsilaki A, Griffiths RD, Jackson MJ. Contractile activity-
induced oxidative stress: celular origin and adaptative response. Am J
Physiol Cell Physiol 280 (2001) C681-C687
McArdle WD, Katch FI, Katch VL. Fisiologia do exercício: Energia, nutrição e
desempenho humano. 6ª ed. Rio de Janeiro: Guanabara Koogan, 2008
McCord JM. Oxygen-derived free radicals in postischemic tissue injury. N Engl
J Med 312 (1985) 159-63
Mendonça RCL, Huertas MC, Hernandez GE, Zabala M, Escames G, Castroviejo
AD. Redox status and antioxidant response in professional cyclists during
training. Eur J Sport Sci 14 (2014) 830-838
Minyi SHI, Wang X, Yamanaka T, Ogita F, Nakatani K, Takeuchi T. Effects of
anaerobic exercise and aerobic exercise on biomarkers of oxidative
stress. Environ Health Prev Med 12 (2007) 202–208
Moller P, Loft S, Lundby C, Olsen NV. Acute hypoxia and hypoxia exercise
induced DNA strand breaks and oxidative DNA damage in humans.
FASEB J 15 (2001) 1181-1186
Moncada S, Higgs EA. The discovery of nitric oxide and its role in vascular
biology. British J Pharmacol 147 (2006) S193-201
Mooren FC, Bloming D, Lechtermann A, Lerch MM, Völker K. Lymphocyte
apoptosis after exhaustive and moderate exercise. J Appl Physiol 93
(2002) 147-153

83
Morton JP, Kayani AC, McArdle A, Drust B. The exercise-induced stress
response of skeletal muscle, with specific emphasis on humans. Sports
Med 39 (2009) 643-662
Nayak S, Jindal A. Myoglobinuria and Acute Kidney Injury. J Integr Nephrol
Androl 2 (2015) 50-54
Nazıroğlu M, Kılınç F, Uğuz AC, Çelik O, Bal R, Butterworth PJ, Baydar MF. Oral
vitamin C and E combination modulates blood lipid peroxidation and
antioxidant vitamin levels in maximal exercising basketball players. Cell
Biochem Function 28 (2010) 300-305
Nieman DC, Stear SJ, Castell LM, Burke LM. A-Z of nutritional supplements:
dietary supplements, sports nutrition foods and ergogenic aids for
health and performance: part 15. Br J Sports Med 44 (2010) 1202-1205
Nikolaidis MG, Paschalis V, Giakas G, Fatouros IG, Koutedakis Y, Kouretas D,
Jamurtas AZ. Decreased blood oxidative stress after repeated muscle-
damaging exercise. Med Sci Sports Exerc 39 (2007) 1080-1089
Nikolaidis MG, Kerksick CM, Lamprecht M, McAnulty SR. Does vitamin C and E
supplementation impair the favorable adaptations of regular exercise?
Oxid Med Cell Longev 2012 (2012) 707-741
Nieman DC, Henson DA, Mc Annulty SR. Vitamine E and immunity after the
Kona Triathlon World Championship. Med Sci Sports Exercise 36 (2004)
1328-1335
Palazzetti S, Richjard MJ, Favier A, Margaritis I. Overload training increases
exercise-induced oxidative stress and damage. Can J Appl Physiol 28
(2003) 588-604
Patil HR, O’keefe JH, Lavie CJ. Cardiovascular damage resulting from chronic
excessive endurance exercise. Mo Med 109 (2012) 312-21
Peake JM. Vitamin C: effects of exercise and requirements with training. Int J
Sport Nutr Exerc Metab 13 (2003) 125-151
Pedersen BK, Rohde T, Ostrowski K. Recovery of the immune system after
exercise. Acta Physiol Scand 162 (1998) 325-332
Peñailillo L, Maya L, Niño G, Torres H, Zbinden-Foncea H. Salivary hormones
and IgA in relation to physical performance in football. .J Sports Sci 33
(2015) 2080-2087
Peternelj TT, Coombes JS. Antioxidant supplementation during exercise
training: beneficial or detrimental (Review). Sports Med 41 (2011) 1043-
1069

84
Porto WG. Radicais Livres e Neurodegeneração. Entendimento Fisiológico:
Base para Nova Terapia? Rev Neurociências 9 (2001) 70-76
Powers SK, Jackson MJ. Exercise-induced oxidative stress: cellular mechanisms
and impact on muscle force production. Physiol Rev 88 (2008) 1243-
1276
Powers SK, Nelson WB, Hudson MB. Exercise-induced oxidative stress in
humans: cause and consequences. Free Radic Biol Med 51 (2011) 942-
950
Powers SK, Talbert EE, Adhihetty PJ. Reactive oxygen and nitrogen species as
intracellular signals in skeletal muscle. J Physiol 589 (2011) 2129-2138
Pyne DB. Regulation of Neutrophil Function During Exercise. Sports med 17
(2004) 245-258
Radak Z, Chung HY, Goto S. Systemic adaptation to oxidative challenge induced
by regular exercise. Free Radic Biol Med 15 (2008) 153-159
Radak Z, Chung HY, Koltai E, Taylor AW, Goto S. Exercise, oxidative stress and
hormesis. Ageing Res Rev. 7 (2008a) 34-42
Radak Z, Zhao Z, Koltai E, et al. Oxygen Consumption and Usage During Physical
Exercise: The Balance Between Oxidative Stress and ROS-Dependent
Adaptive Signaling. Antioxid Redox Signal 18 (2013) 1208-1246
Reeder BJ, Svistunenko DA, Cooper CE, Wilson MT. The radical and redox
chemistry of myoglobin and hemoglobin: from in vitro studies to human
pathology. Antioxid Redox Signal. 6 (2004) 954-966
Reichhold S, Neubauer O, Hoelzl C, et al. DNA damage in response to an
Ironman triathlon. Free Radic Res 43 (2009) 753-760
Ribon A, Pialoux V, Saugy JJ, Rupp T, Faiss R, Debevec T, Millet GP. Exposure
to hypobaric hypoxia results in higher oxidative stress compared to
normobaric hypoxia. Respir Physiol Neurobiol 223 (2016) 23-27
Robson PJ, Blannin AK, Walsh NP, Castell LM, Gleeson M (1999). Effects of
exercise intensity, duration and recovery on in vitro neutrophil function
in male athletes. Int J Sports Med 20 (1999) 128-135
Rover JL, Hoehr NF, Vellasco AP. Sistema antioxidante envolvendo o ciclo
metabólico da glutationa associado a métodos eletroanalíticos na
avaliação do estresse oxidativo. Química Nova 24 (2004) 112-119
Sautin Y, Johnson R. Uric Acid: The Oxidant-Antioxidant Paradox. Nucleosides,
Nucleotides and Nucleic Acids 27 (2008) 608-619

85
Scheffer DL, Silva LA, Tromm CB, da Rosa GL, Silveira PCL, de Souza CT, Latini A,
Pinho RA. Impact of different resistance training protocols on muscular
oxidative stress parameters. Appl Physiol Nutr Metabol 37 (2012) 1239-
1246
Schmikli SL, de Vries WR, Brink MS, Backx FJ. Monitoring performance,
pituitary-adrenal hormones and mood profiles: how to diagnose non-
functional over-reaching in male junior soccer players. Br J Sports Med
46 (2012) 1019-1023
Schneider BSP, Tiidus PM. Neutrophil Infiltration in Exercise-Injured Skeletal
Muscle. How Do We Resolve the Controversy? Sports Med 37 (2007)
837-856
Schoene RB. Limits of human lung function at high altitude. J Exp Biol 204
(2001) 3212-3217
Sen CK. Oxidants and antioxidants in exercise, J Appl. Physiol 79 (1995) 675-
686
Shephard RJ. Adhesion Molecules, Catecholamines and Leucocyte Redistribu-
tion During and Following Exercise. Sports Med 33 (2003) 261-284
Shi M, Wang X, Yamanaka T, Ogita F, Nakatani K, Takeuchi T. Effects of anaerobic
exercise and aerobic exercise on biomarkers of oxidative stress. Environ
Health Prev Med 12 (2007) 202-208
Sies H. Hydroperoxides and thiol oxidants in the study of oxidative stress
in intact cells and organs. In: Oxidative stress, H. Sies (Ed.) London:
Academic Press Inc., pp73-90, 1985
Sies H. Glutathione and its role in cellular functions. Free Rad Biol Med 27
(1999) 916-920
Silva FOC, Macedo DV. Exercício físico, processo inflamatório e adaptação:
uma visão geral Rev Bras Cineantropom Desempenho Hum 13 (2011)
320-328
Silva, DC, Cerchiaro, G, Honório, KM. Relações patofisiológicas entre estresse
oxi8dativo e ateriosclerose. Quim Nova 34 (2011) 300-305
Silva RS, Ascensao A, Marques F, Seabra A, Rebelo A, Magalhaes J.
Neuromuscular function, hormonal and redox status and muscle
damage of professional soccer players after a high-level competitive
match. Eur J Appl Physiol 113 (2013) 2193-2201
Silva JR, Rebelo A, Marques F, Pereira L, Seabra A, Ascensão A, Magalhães J.
Biochemical impact of soccer: an analysis of hormonal, muscle damage,

86
and redox markers during the season. Appl Physiol Nutr Metab 39 (2014) 432-438
Smith JA, Telford RD, Mason IB, and Weideman MJ. (1990). Exercise, training and neutrophil microbicial activity. In J Sports Med 11 (1990) 179-187
Soares J, Natal A. Fisiologia do treinamento no alto desempenho do atleta de futebol. Revista USP 99 (2013) 91-106
Sterczala AJ. “The Stress Response to an Acute Heavy Resistance Exercise Protocol“. 2014. Master’s Theses. Paper 586. http://digitalcommons.uconn.edu/gs_theses/ 586
Subczynsky WK, Wisniewska A. Physical properties of lipid bilayer membranes: relevance to membrane biological function. Acta Biochem Pol 3 (2000) 613-625
Tanskanen M, Atalay M, Uusitalo A. Altered oxidative stress in overtrained athletes. J Sports Sci 28 (2010) 309-317
Tapiero H, Tew KD. The importance of glutathione in human disease. Biomed Pharmacother 57 (2003) 145-55
Teixeira VH, Valente HF, Casal SI, Marques F, Moreira PA. Antioxidants do not prevent postexercise peroxidation and may delay muscle recovery. Med Sci Sports Exerc 41 (2009) 1752-1760
Teixeira V, Valente H, Casal S, Marques F, Moreira P. Antioxidant status, oxidative stress, and damage in elite kayakers after 1 year of training and competition in 2 seasons. Appl Physiol Nutr Metab 34 (2009a) 716-724
Teixeira V, Valente H, Casal S, Marques F, Moreira P. Antioxidant status, oxidative stress, and damage in elite trained kayakers and canoeists and sedentary controls. Int J Sport Nutr Exerc Metab 19 (2009b) 443-456
Teixeira VH, Valente HF, Casal SI, Marques F, Moreira PA. Blood antioxidant and oxidative stress biomarkers acute responses to a 1000-m kayak sprint in elite male kayakers. J Sports Med Phys Fitness 53 (2013) 71-79
The 2015 Ageing Report. Economic and budgetary projections for the 28 EU Member States (2013-2060). EUROPEAN ECONOMY 3. 2015. ISSN 1725-3217 (online)
Theodorou AA, Nikolaidis MG, Paschalis V, Koutsias S, Panayiotou G, Fatouros IG, Koutedakis Y, Jamurtas AZ. No effect of antioxidant supplementation on muscle performance and blood redox status adaptations to eccentric training. Am J Clin Nutr 93 (2011) 1373-1383
Tidball JG. Inflammatory processes in muscle injury and repair. Am J Physiol
Regul Integr Comp Physiol 288 (2005) R345-R353

87
Tidball JG, Wehling-Henricks M. Macrophages promote muscle membrane
repair and muscle fibre growth and regeneration during modified
muscle loading in mice. in vivo. J. Physiol 578 (2007) 327-36
Tong TK, Lin H, Lippi G, Nie J, Tian Y. Serum oxidant and antioxidant status
in adolescents undergoing professional endurance sports training. Oxid
Med Cell Longev 2012 (2012) 741239.
Trochimiak T, Hübner-Woźniak E. Effect of exercise on the level of
immunoglobulin A in saliva. Biol Sport 29 (2012) 255 -261
Turrens JF. Mitochondrial formation of reactive oxygen species. J Physiol 552
(2003) 335-344
Twist C, Eston R. The effects of exercise-induced muscle damage on maximal
intensity intermittent exercise performance. Eur J Appl Physiol 94 (2005)
652-658
Uchida MC, Nosaka, K. Ugrinowistsch CX, Yamashita A, Martins JR, Moriscot
AS, Aoki MS. Effect of bench press exercise intensity on muscle soreness
and inflammatory mediators. J Sports Sci 27 (2009) 499-507
Umbarkara P, Singha S, Arkata S, Bodhankarb SL, Lohidasanc S, Sitasawada
SL. Monoamine oxidase-A is an important source of oxidative stress
and promotes cardiac dysfunction, apoptosis, and fibrosis in diabetic
cardiomyopathy. Free Rad Biol Med 87 (2015) 263-273
UNESCO. Fisiologia do exercício. Brasília. Fundação Vale, UNESCO, 2013
Viña J, Gomez-Cabrera MC, Lloret A, Marquez R, Miñana JB, Pallardó FV, Sastre
J. Free radicals in exhaustive physical exercise: mechanism of production,
and protection by antioxidants. IUBMB Life 50 (2000) 271-277
Virgen JL, Kraemer WJ, Ratamess NA, Anderson JM, Volek JS, Maresh CM.
Testosterone physiology in resistance exercise and training: the up-
stream regulatory elements. Sports Med 40 (2010) 1037-1053.
Vollaard NB, Shearmen JP, Cooper CE. Exercise-induced oxidative stress myths,
realities and physiological relevance. Sports Med 35 (2005) 1045-1062
Wagner KH, Reichhold S, Hölzl C, Knasmüller S, Nics L, Meisel M, Neubauer
O. Well-trained, healthy triathletes experience no adverse health risks
regarding oxidative stress and DNA damage by participating in an ultra-
endurance event. Toxicology 278 (2010) 211-216
Wagner KH, Reichhold S, Neubauer O. Impact of endurance and ultraendurance
exercise on DNA damage. Ann N Y Acad Sci 1229 (2011) 115-123

88
Waring WS, Convery A, Mirsha V, Shenkin A, Webb DJ, Maxwell SR. Uric acid
reduces exercise-induced oxidative stress in health adults. Clin Sci 105
(2003) 425-430
Watson T, Lesley K, MacDonalds-Wicks, Gard M. Oxidative stress and oxidants
in athletes undertaking regular exercise training. Int J Sports Nutr Exerc
Metab 15 (2005) 131-146.
Wayner DD, Burton, GW, Ingold KU, Barclay LR, Locke SJ. The relative
contributions of vitamine E, urate, ascorbate and proteins to the total
peroxyl radical-trapping antioxidante activity of human blood plasma.
Biochem Biophys Acta 3 (1987) 408-419
Williams MH. Dietary Supplements and Sports Performance: Introduction and
Vitamins. J Int Soc Sports Nutr 1 (2004) 1-6
Wiecek M, Maciejczyk M, Szymura J, Szygula Z. Effect of maximal-intensity
exercise on systemic nitro-oxidative stress in men and women. Redox
Rep 14 (2016) 1-7
Wolach B, Gavrieli R, Ben-Dror SG, Zigel L, Eliakim A, Falk B. Transient decrease
of neutrophil chemotaxis following aerobic exercise. Med Sci Sports
Exerc 37 (2005) 949-955
Zembron-Lacey A, Szyszka K, Sobanska B, Pakula R. Prooxidant-antioxidant
equilibrium in rowers: effect of a single dose of vitamin E. J Sports Med
Phys Fitness 46 (2006) 257-264
Zondi PC, Rensburg DCJ, Grant CC, van Rensburg AJ. Delayed onset muscle
soreness: No pain, no gain? The truth behind this adage. South African
Family Practice 57 (2015) 29-33

89
EPÍLOGO
O caminho da ciência e da investigação científica exige cada vez mais a conjugação de distintos saberes para fazer face à constante evolução do pensamento e aproveitamento dos meios tecnológicos em contínuo desenvolvimento.
Na realidade, não existe ciência e investigação cientifica de um homem só. Tal como o homem, que é na essencia um animal social, também a ciencia apenas progride se permitir a flexibilidade e a liberdade de atuação e de objectivos dos seus “players” quando integrada em equipas multidisciplinares.
Ortega e Gasset dizia, na sua obra publicada, em 1937, La rebelión de las masas, que “nós somos nós e a nossa circunstância”. De facto, eu sou o fruto e a razão da minha circunstância. E nesta circunstância, que é a minha, eu sou o produto das relações de cooperação e de amizade que ao longo do tempo fui criando e acima de tudo mantendo, amizades e colaborações essas, desinteressadas e por isso mesmo profícuas.
Não posso deixar de destacar entre todos os elementos com quem mais frequentemente colaboro, José Soares, José Magalhães, António Ascenção, Vitor Hugo Teixeira, José Augusto Santos, todos docentes da Universidade do Porto, nomeadamente, nas Faculdades de Desporto e Ciencias da Nutrição e Alimentação, mas também, José Ramos, de Medicina Desportiva. Alguns destes foram meus estudantes de doutoramento; outros apareceram na minha vida cientifica e pessoal de uma maneira inusitada e inesperada, mas a quem agradeço o facto de nela terem permanecido. A todos, contudo, chamo de amigos.
Uma palavra especial aos meus colegas e amigos, que constituem a comunidade académica onde estou inserido e que dão vida ao Departamento de Ciências Biológicas da Faculdade de Farmácia da Universidade do Porto. Permitam-me, que entre todos destaque uma palavra particular à Prof. Natércia Teixeira, minha orientadora da

90
tese de doutoramento e atual Diretora do Departamento. São eles os motores e a essência da excelente investigação e da qualidade de ensino, que faz com que esta Instituição, a mais antiga das Faculdades de Farmácia em Portugal, seja entre estas, um caso impar.
Neste momento, sinto-me mais “rico”, mais completo, pois por um lado, tive a possibilidade de adquirir novos saberes e, por outro, de conhecer novas pessoas que certamente integrarão aventuras futuras.
Como dizia Fernando Pessoa, “Tudo o que é bom, dura o tempo necessário para ser inesquecível”. Seja qual for tempo destinado a este momento, ele já é para mim único e inesquecível.

100



















